

Halorotetin A: A Novel Terpenoid Compound Isolated from Ascidian Halocynthia rotetzi Exhibits the Inhibition Activity on Tumor Cell Proliferation


Abstract
:1. Introduction
2. Results
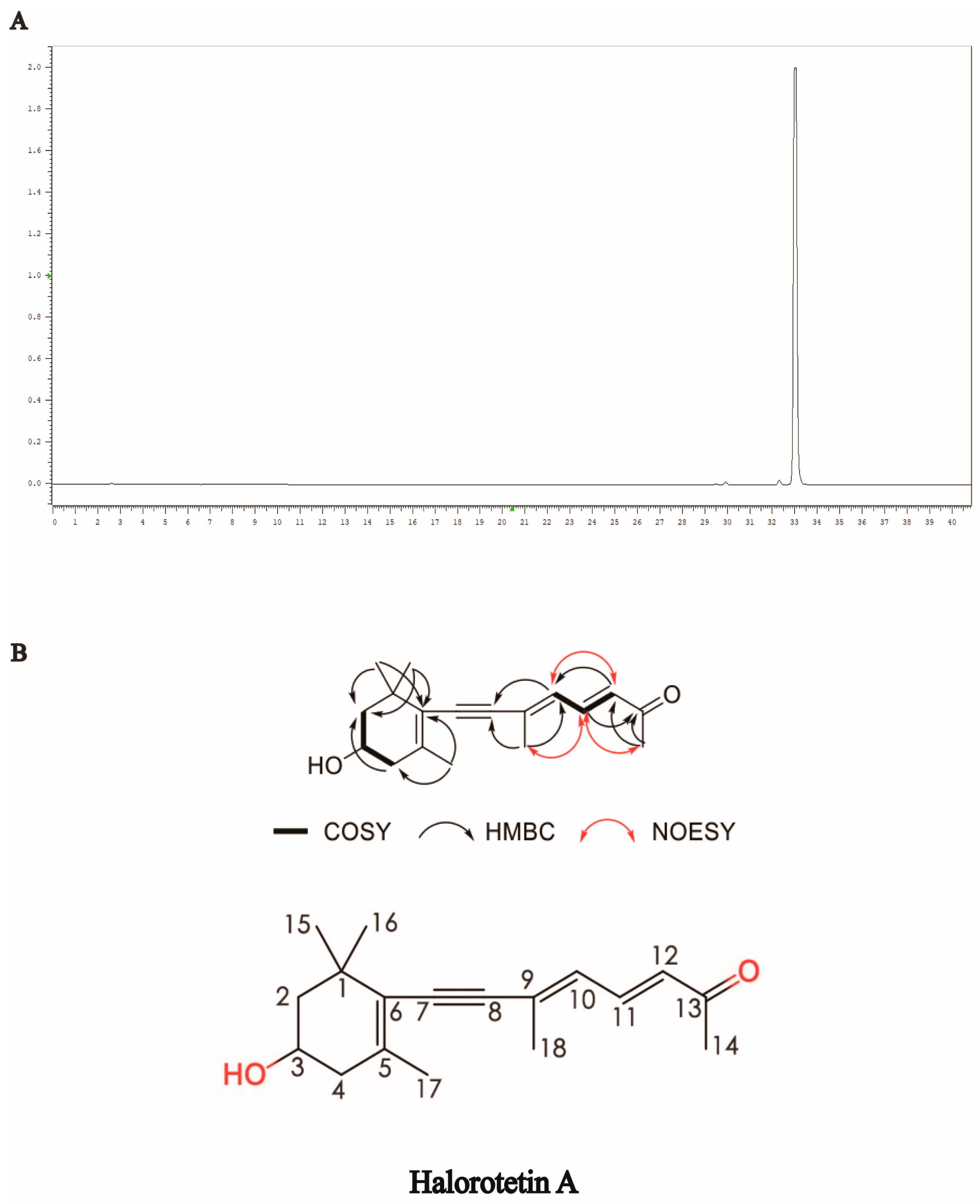
2.1. Isolation and Purification of Halorotetin A
2.2. Structural Identification of Halorotetin A
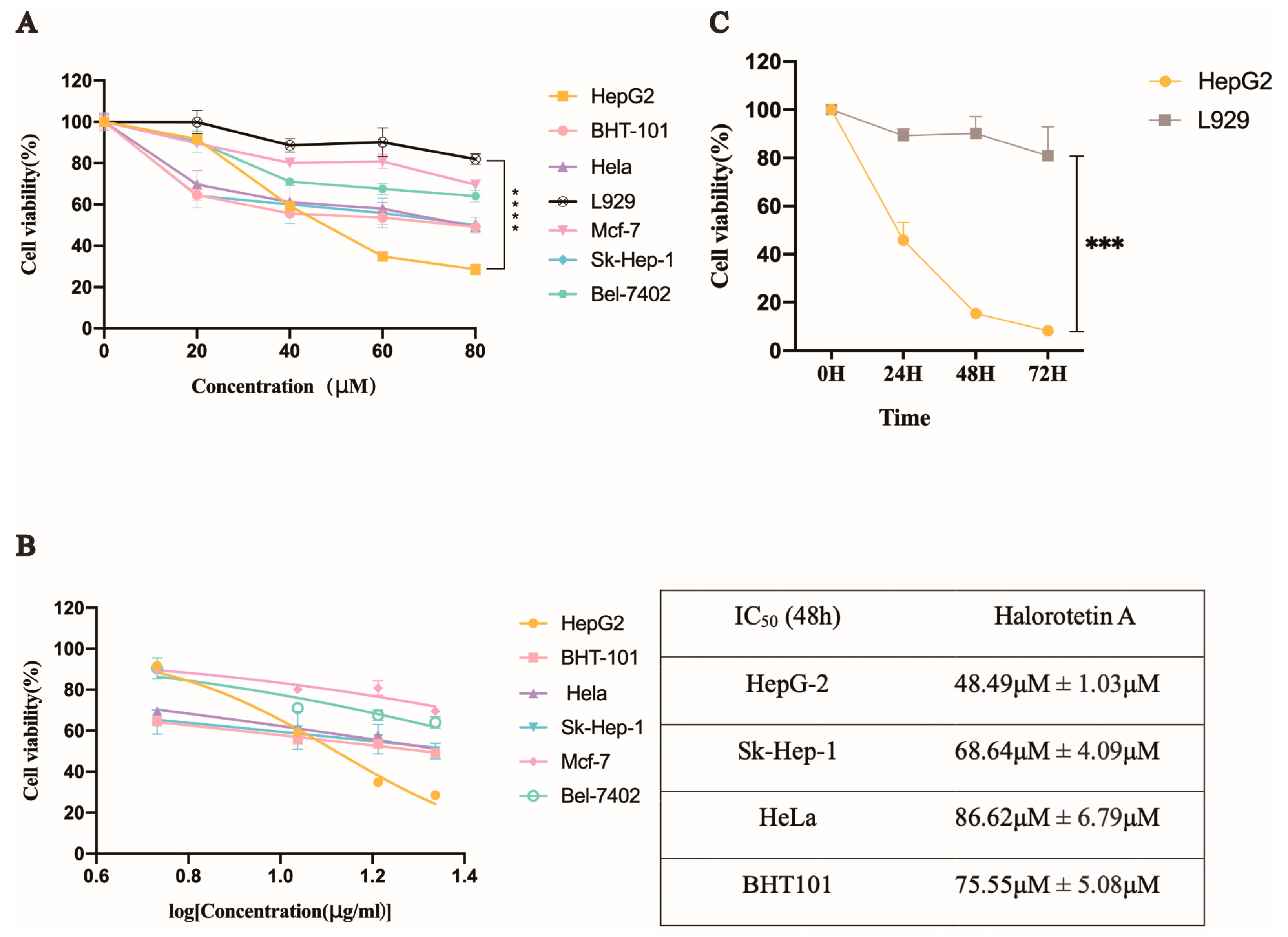
2.3. Halorotetin A Inhibits the Proliferation of a Series of Tumor Cells
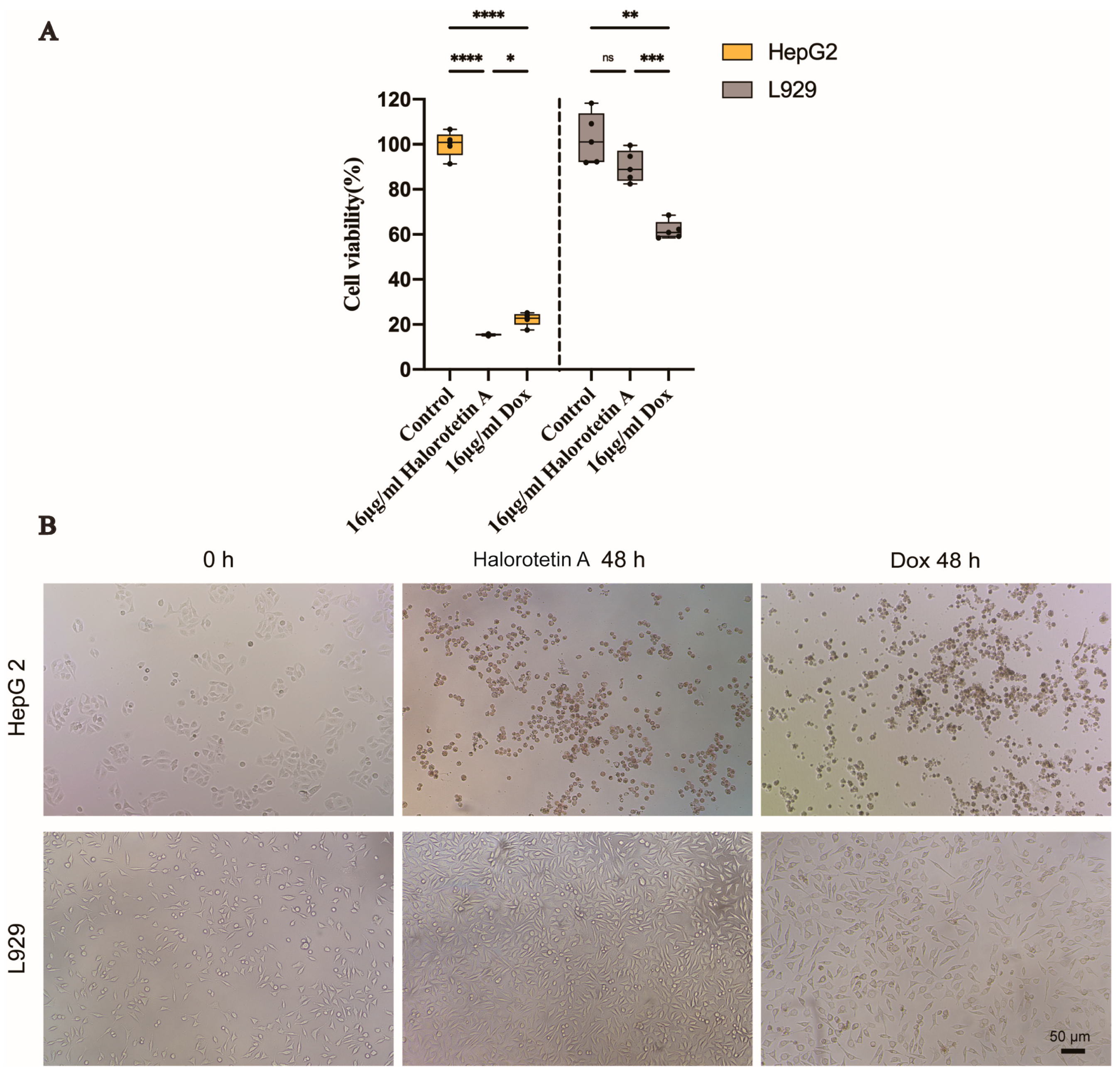
2.4. Halorotetin A Shows the Cytotoxicity to HepG-2 Cells but Much Less to L929 Cells
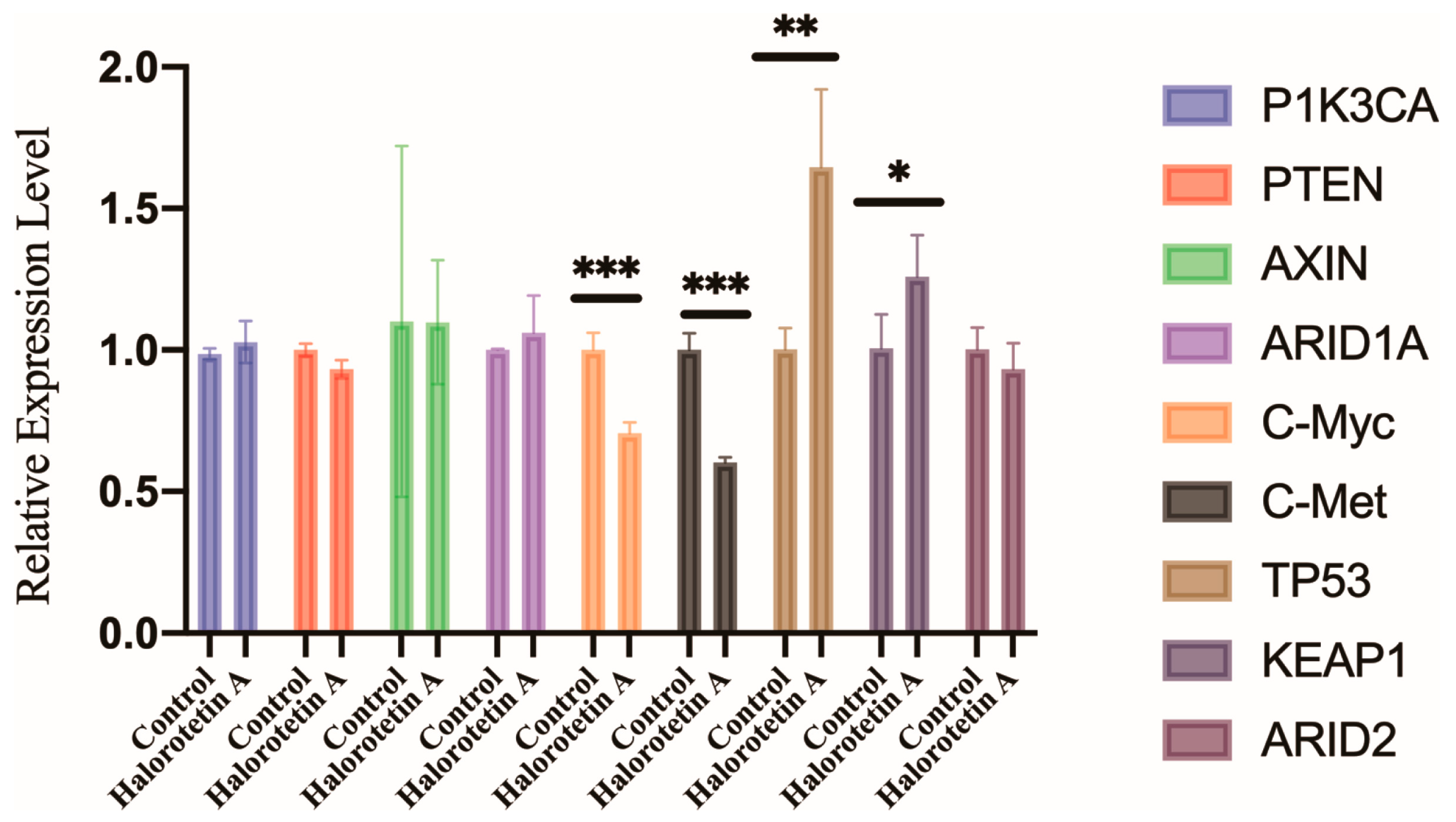
2.5. Halorotetin A Inhibits c-myc and c-met but Promotes the TP53 Transcription Level in HepG-2 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of SGC-II by Silica Gel Column Chromatography
4.3. Preparation of Halorotetin A by PLC and HPLC
4.4. Structural Identification of Halorotetin A
4.5. Cell Culture
4.6. MTT Assay for Cell Viability
4.7. RNA Extraction from HepG-2 Cells
4.8. Reverse Transcription
4.9. Detecting of Gene Expression
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Gerwick, W.H.; Fenner, A.M. Drug Discovery from Marine Microbes. Microb. Ecol. 2013, 65, 800–806. [Google Scholar] [CrossRef]
- Kowalski, R.J.; Giannakakou, P.; Gunasekera, S.P.; Longley, R.E.; Day, B.W.; Hamel, E. The Microtubule-Stabilizing Agent Discodermolide Competitively Inhibits the Binding of Paclitaxel (Taxol) to Tubulin Polymers, Enhances Tubulin Nucleation Reactions More Potently than Paclitaxel, and Inhibits the Growth of Paclitaxel-Resistant Cells. Mol. Pharmacol. 1997, 52, 613–622. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Shin, H.J. Natural Products from Marine Fungi. Mar. Drugs 2020, 18, 230. [Google Scholar] [CrossRef]
- Sang, V.T.; Dat, T.T.H.; Vinh, L.B.; Cuong, L.C.V.; Oanh, P.T.T.; Ha, H.; Kim, Y.H.; Anh, H.L.T.; Yang, S.Y. Coral and Coral-Associated Microorganisms: A Prolific source of Potential Bioactive Natural Products. Mar. Drugs 2019, 17, 468. [Google Scholar] [CrossRef] [Green Version]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [Green Version]
- Dou, X.; Dong, B. Origins and Bioactivities of Natural Compounds Derived from Marine Ascidians and Their Symbionts. Mar. Drugs 2019, 17, 670. [Google Scholar] [CrossRef] [Green Version]
- Sakai, R.; Rinehart, K.L.; Guan, Y.; Wang, J. Additional antitumor ecteinascidins from a Caribbean tunicate: Crystal structures and activities in vivo. Proc. Nati.Acad. Sci. USA 1992, 89, 5. [Google Scholar] [CrossRef]
- Hendriks, H.R.; Fiebig, H.H.; Giavazzi, R.; Langdon, S.P.; Jimeno, J.M.; Faircloth, G.T. High antitumour activity of ET743 against human tumour xenografts from melanoma, non-small-cell lung and ovarian cancer. Ann. Oncol. 1999, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- D’Incalci, M.; Galmarini, C.M. A Review of Trabectedin (ET-743): A Unique Mechanism of Action. Mol. Cancer Ther. 2010, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yovine, A.; Riofrio, M.; Blay, J.-Y.; Brain, E.; Alexandre, J.; Kahatt, C.; Taamma, A.; Jimeno, J.; Martin, C.; Salhi, Y.; et al. Phase II Study of Ecteinascidin-743 in Advanced Pretreated Soft Tissue Sarcoma Patients. J. Clin. Oncol. 2004, 22, 890–899. [Google Scholar] [CrossRef]
- Zhao, J.; Wei, J.; Liu, M.; Xiao, L.; Wu, N.; Liu, G.; Huang, H.; Zhang, Y.; Zheng, L.; Lin, X. Cloning, characterization and expression of a cDNA encoding a granulin-like polypeptide in Ciona savignyi. Biochimie 2013, 95, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Hirose, E.; Ohtake, S.-I.; Azumi, K. Morphological characterization of the tunic in the edible ascidian, Halocynthia roretzi (Drasche), with remarks on ‘soft tunic syndrome’ in aquaculture. J. Fish Dis. 2009, 32, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Mikami, N.; Miyashita, K. Effects of sea squirt (Halocynthia roretzi) lipids on white adipose tissue weight and blood glucose in diabetic/obese KK-Ay mice. Mol. Med. Rep. 2010, 3, 449–453. [Google Scholar] [CrossRef]
- Watanabe, H.; Okawara, M.; Matahira, Y.; Mano, T.; Wada, T.; Suzuki, N.; Takara, T. The Impact of Ascidian (Halocynthia roretzi)-derived Plasmalogen on Cognitive Function in Healthy Humans: A Randomized, Double-blind, Placebo-controlled Trial. J. Oleo Sci. 2020, 69, 1597–1607. [Google Scholar] [CrossRef]
- Wei, J.; Gao, H.; Yang, Y.; Liu, H.; Yu, H.; Chen, Z.; Dong, B. Seasonal dynamics and starvation impact on the gut microbiome of urochordate ascidian Halocynthia roretzi. Anim. Microbiome 2020, 2, 30. [Google Scholar] [CrossRef]
- Zhu, Y.; Han, S.; Li, J.; Gao, H.; Dong, B. Aqueous Extract of Sea Squirt (Halocynthia roretzi) with Potent Activity against Human Cancer Cells Acts Synergistically with Doxorubicin. Mar. Drugs 2022, 20, 284. [Google Scholar] [CrossRef]
- Oh, Y.; Shim, K.-B.; Ahn, C.-B.; Kim, S.S.; Je, J.-Y. Sea Squirt (Halocynthia roretzi) Hydrolysates Induce Apoptosis in Human Colon Cancer HT-29 Cells through Activation of Reactive Oxygen Species. Nutr. Cancer 2019, 71, 118–127. [Google Scholar] [CrossRef]
- Kawasaki, M. Effects of dietary sea squirt (Halocynthia roretzi) on lipid metabolism in rats. Biofactors 2004, 22, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Andersson, T.; Borhan, B.; Berova, N.; Nakanishi, K.; Haugan, J.A.; Liaaen-Jensen, S. Absolute configurational assignment of 3-hydroxycarotenoids. J. Chem. Soc. Perkin Trans. 2000, 5. [Google Scholar] [CrossRef]
- Matsuno, T.; Ookubo, M.; Nishizawa, T.; Shimizu, I. Carotenoids of Sea Squirts. I. New Marine Carotenoids, Halocynthiaxanthin and Mytiloxanthinone from Halocynthia roretzi. Chem. Pharm. Bull. 1984, 32, 4309–4315. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, S.; Marais, R.; Zhu, A.X. The role of signaling pathways in the development and treatment of hepatocellular carcinoma. Oncogene 2010, 29, 4989–5005. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, J.J.; Huang, M.Q.; Bao, J.L.; Chen, X.P.; Wang, Y.T. Terpenoids: Natural products for cancer therapy. Expert Opin. Investig. Drugs 2012, 21, 1801–1818. [Google Scholar] [CrossRef]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibasami, H.; Iwase, H.; Yoshioka, K.; Takahashi, H. Glycyrrhizin induces apoptosis in human stomach cancer KATO III and human promyelotic leukemia HL-60 cells. Int. J. Mol. Med. 2005, 16, 233–236. [Google Scholar] [CrossRef]
- Gnanasekar, M.; Thirugnanam, S.; Xu, L.; Ramaswamy, K. Glycyrrhizin induces apoptosis in prostate cancer cell lines DU-145 and LNCaP. Oncol. Rep. 2008, 20, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Hua, Y.Q.; Zhang, Z.Y.; Li, J.X.; Li, Q.; Hu, S.; Li, J.; Sun, M.X.; Cai, Z.D. Oleanolic acid derivative Dex-OA has potent anti-tumor and anti-metastatic activity on osteosarcoma cells in vitro and in vivo. Investig. New Drugs 2011, 29, 258–265. [Google Scholar] [CrossRef]
- Kapil, A.; Sharma, S. Effect of Oleanolic Acid on Complement in Adjuvant-Induced and Carrageenan-induced Inflammation in Rats. J. Pharm. Pharmacol. 1995, 47, 585–587. [Google Scholar] [CrossRef]
- Thompson, E.B. The Many Roles of c-myc in Apoptosis. Annu. Rev. Physiol. 1998, 60, 575–600. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.R.; Garner, A.L.; Yu, J.; Janda, K.D. Inhibitor of MYC identified in a krohnke pyridine library. Proc. Natl. Acad. Sci. USA 2014, 111, 12556–12561. [Google Scholar] [CrossRef] [Green Version]
- Comoglio, P.M.; Trusolino, L.; Boccaccio, C. Known and novel roles of the MET oncogene in cancer: A coherent approach to targeted therapy. Nat. Rev. Cancer 2018, 18, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, M.; Jin, K.; Wang, S.; Wei, H.; Fan, C.; Wu, Y.; Li, X.; Li, X.; Li, G.; et al. Function of the c-met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities. Mol. Cancer 2018, 17, 45. [Google Scholar] [CrossRef]
- Paik, P.K.; Felip, E.; Cortot, A.B.; Le, X. Tepotinib in non-small-cell lung cancer with MET exon 14 skipping mutations. N. Engl. J. Med. 2020, 383, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.T.; Bauer, T.M.; de Marinis, F.; Felip, E.; Goto, Y.; Liu, G.; Mazieres, J.; Kim, D.-W.; Mok, T.; Polli, A.; et al. First-Line Lorlatinib or Crizotinib in Advanced ALK-Positive Lung Cancer. N. Engl. J. Med. 2020, 383, 2018–2029. [Google Scholar] [CrossRef]
- Muller, P.A.J.; Vousden, K.H. p53 mutations in cancer. Nat. Cell Biol. 2012, 15, 2–8. [Google Scholar] [CrossRef]
- Nunez-Pons, L.; Nieto, R.M.; Avila, C.; Rodriguez, J. Mass spectrometry detection of minor new meridianins from the antarctic colonial ascidians Aplidium falklandicum and Aplidium meridianum. J. Mass Spectrom 2015, 50, 103–111. [Google Scholar] [CrossRef]
- Richardson, A.D.; Aalbersberg, W.; Ireland, C.M. The patellazoles inhibit protein synthesis at nanomolar concentrations in human colon tumor cells. Anti-Cancer Drugs 2005, 16, 533–541. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Kato, H.; Hirota, H.; Fusetani, N. Antibacterial and antifungal sulfated alkane and alkenes from the hepatopancreas of the ascidian Halocynthia roretzi. J. Nat. Prod. 1994, 57, 4. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, Y.; Liu, H.; Wei, J.; Yu, H.; Dong, B. Cultivation of gut microorganisms of the marine ascidian Halocynthia roretzi reveals their potential roles in the environmental adaptation of their host. Mar. Life Sci. Technol. 2022, 4, 7. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | δH (m, J in Hz) | δC (m) | HMBC |
|---|---|---|---|
| 1 | 36.1 | ||
| 2 | 1.42 (m) 1.81 ddd (12.4, 3.7, 2.0) | 46.0 | C-1,3,15,16 |
| 3 | 3.90 dddd (11.9, 9.3, 5.6, 3.6) | 63.7 | |
| 4 | 2.03 (m) 2.41 (m) | 40.8 | C-2,3,5,6,17 |
| 5 | 139.8 | ||
| 6 | 123.7 | ||
| 7 | 92.2 | ||
| 8 | 97.0 | ||
| 9 | 130.0 | ||
| 10 | 6.46 d (11.6) | 131.6 | C-,8,11,12,18 |
| 11 | 7.56 dd (15.3, 11.6) | 138.6 | C-12,13 |
| 12 | 6.21 d (15.3) | 129.6 | C-10,13,14 |
| 13 | 200.0 | ||
| 14 | 2.30 (s) | 26.1 | C-12,13 |
| 15 | 1.13 (s) | 27.7 | C-1,2,6,16 |
| 16 | 1.17 (s) | 29.7 | C-1,2,6,15 |
| 17 | 1.91 (s) | 21.3 | C-4,5,6,7 |
| 18 | 2.11 (s) | 17.3 | C-8,9,10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Han, S.; Zhu, Y.; Dong, B. Halorotetin A: A Novel Terpenoid Compound Isolated from Ascidian Halocynthia rotetzi Exhibits the Inhibition Activity on Tumor Cell Proliferation. Mar. Drugs 2023, 21, 51. https://doi.org/10.3390/md21010051
Li J, Han S, Zhu Y, Dong B. Halorotetin A: A Novel Terpenoid Compound Isolated from Ascidian Halocynthia rotetzi Exhibits the Inhibition Activity on Tumor Cell Proliferation. Marine Drugs. 2023; 21(1):51. https://doi.org/10.3390/md21010051
Chicago/Turabian StyleLi, Jianhui, Shanhao Han, Yuting Zhu, and Bo Dong. 2023. "Halorotetin A: A Novel Terpenoid Compound Isolated from Ascidian Halocynthia rotetzi Exhibits the Inhibition Activity on Tumor Cell Proliferation" Marine Drugs 21, no. 1: 51. https://doi.org/10.3390/md21010051