Broadening and Enhancing Bacteriocins Activities by Association with Bioactive Substances


Abstract
:1. Bacteriocins, a Resourceful Antimicrobial Peptides
2. Antibiotics and Bacteriocins Constitute Promising Synergistic Combinations for Medical Therapy
3. Nanoparticles for Carrying and Improving the Bacteriocin Activity
4. Combined Effects of Bacteriocins and Bacteriophages
5. Bacteriocins-Essential Oils Synergy are Active Against Pathogenic Bacteria
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Ladjouzi, R.; Lucau-Danila, A.; Benachour, A.; Drider, D. A leaderless two-peptide Bacteriocin, Enterocin DD14, is involved in its own self-immunity: Evidence and insights. Front. Bioeng. Biotechnol. 2020, 8, 644. [Google Scholar] [CrossRef] [PubMed]
- Ríos Colombo, N.S.; Chalón, M.C.; Navarro, S.A.; Bellomio, A. Pediocin-like bacteriocins: New perspectives on mechanism of action and immunity. Curr. Genet. 2018, 64, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Nes, I.F.; Diep, D.B.; Håvarstein, L.S.; Brurberg, M.B.; Eijsink, V.; Holo, H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 113–128. [Google Scholar] [CrossRef]
- Drider, D.; Fimland, G.; Héchard, Y.; McMullen, L.M.; Prévost, H. The continuing story of class IIa bacteriocins. Microbiol. Mol. Biol. Rev. MMBR 2006, 70, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Line, J.E.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S.; Siragusa, G.R.; et al. Isolation and purification of enterocin E-760 with broad antimicrobial activity against gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 2008, 52, 1094–1100. [Google Scholar] [CrossRef] [Green Version]
- Messaoudi, S.; Kergourlay, G.; Dalgalarrondo, M.; Choiset, Y.; Ferchichi, M.; Prévost, H.; Pilet, M.F.; Chobert, J.M.; Manai, M.; Dousset, X. Purification and characterization of a new bacteriocin active against Campylobacter produced by Lactobacillus salivarius SMXD51. Food Microbiol. 2012, 32, 129–134. [Google Scholar] [CrossRef]
- Drider, D.; Bendali, F.; Naghmouchi, K.; Chikindas, M.L. Bacteriocins: Not only antibacterial agents. Probiotics Antimicrob. Proteins 2016, 8, 177–182. [Google Scholar] [CrossRef]
- Mokoena, M.P. Lactic acid bacteria and their bacteriocins: Classification, biosynthesis and applications against uropathogens: A mini-review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, V.Q.; Carvalho, O.V.; Paiva, J.C.; Todorov, S.D.; Júnior, A.S.; Nero, L.A. Inhibition of herpes simplex virus 1 (HSV-1) and poliovirus (PV-1) by bacteriocins from Lactococcus lactis subsp. lactis and Enterococcus durans strains isolated from goat milk. Int. J. Antimicrob. Agents 2018, 51, 33–37. [Google Scholar] [CrossRef]
- Wachsman, M.B.; Castilla, V.; de Ruiz Holgado, A.P.; de Torres, R.A.; Sesma, F.; Coto, C.E. Enterocin CRL35 inhibits late stages of HSV-1 and HSV-2 replication in vitro. Antiviral Res. 2003, 58, 17–24. [Google Scholar] [CrossRef]
- Ermolenko, E.I.; Desheva, Y.A.; Kolobov, A.A.; Kotyleva, M.P.; Sychev, I.A.; Suvorov, A.N. Anti-influenza activity of Enterocin B in vitro and protective effect of bacteriocinogenic enterococcal probiotic strain on influenza infection in mouse model. Probiotics Antimicrob. Proteins 2019, 11, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Małaczewska, J.; Kaczorek-Łukowska, E.; Wójcik, R.; Siwicki, A.K. Antiviral effects of nisin, lysozyme, lactoferrin and their mixtures against bovine viral diarrhoea virus. BMC Vet. Res. 2019, 15, 318. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Kaur, S. Bacteriocins as potential anticancer agents. Front. Pharmacol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Baindara, P.; Korpole, S.; Grover, V. Bacteriocins: Perspective for the development of novel anticancer drugs. Appl. Microbiol. Biotechnol. 2018, 102, 10393–10408. [Google Scholar] [CrossRef]
- Baindara, P.; Gautam, A.; Raghava, G.P.S.; Korpole, S. Anticancer properties of a defensin like class IId bacteriocin Laterosporulin10. Sci. Rep. 2017, 7, 46541. [Google Scholar] [CrossRef] [Green Version]
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocin-antimicrobial synergy: A medical and food perspective. Front. Microbiol. 2017, 8, 1205. [Google Scholar] [CrossRef] [Green Version]
- Namasivayam, K.R.S.; Samrat, K.; Debnath, S.; Jayaprakash, C. Biocompatible chitosan nanoparticles incorporated bacteriocin (CSNps-B) preparation for the controlled release and improved anti-bacterial activity against food borne pathogenic bacteria Listeria monocytogenes. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 625–631. [Google Scholar]
- Rodríguez-Rubio, L.; García, P.; Rodríguez, A.; Billington, C.; Hudson, J.A.; Martínez, B. Listeriaphages and coagulin C23 act synergistically to kill Listeria monocytogenes in milk under refrigeration conditions. Int. J. Food Microbiol. 2015, 205, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Issouffou, C.; Suwansri, S.; Salaipeth, L.; Domig, K.J.; Hwanhlem, N. Synergistic effect of essential oils and enterocin KT2W2G on the growth of spoilage microorganisms isolated from spoiled banana peel. Food Control 2018, 89, 260–269. [Google Scholar] [CrossRef]
- Todorov, S.D.; de Paula, O.A.L.; Camargo, A.C.; Lopes, D.A.; Nero, L.A. Combined effect of bacteriocin produced by Lactobacillus plantarum ST8SH and vancomycin, propolis or EDTA for controlling biofilm development by Listeria monocytogenes. Rev. Argent. Microbiol. 2018, 50, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, A.M.; Fialho, A.M. (Eds.) Microbial Infections and Cancer Therapy; Jenny Stanford Publishing: Singapore, 2019; ISBN 978-1-351-04190-4. [Google Scholar]
- Iseppi, R.; Camellini, S.; Sabia, C.; Messi, P. Combined antimicrobial use of essential oils and bacteriocin bacLP17 as seafood biopreservative to control Listeria monocytogenes both in planktonic and in sessile forms. Res. Microbiol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.; Kim, M.G.; Kwon, M.; Lee, H.S.; Kim, G.B. Inhibition of Clostridium perfringens using bacteriophages and Bacteriocin producing strains. Korean J. Food Sci. Anim. Resour. 2018, 38, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Al Atya, A.K.; Belguesmia, Y.; Chataigne, G.; Ravallec, R.; Vachée, A.; Szunerits, S.; Boukherroub, R.; Drider, D. Anti-MRSA activities of enterocins DD28 and DD93 and evidences on their role in the inhibition of biofilm formation. Front. Microbiol. 2016, 7, 817. [Google Scholar] [CrossRef] [Green Version]
- Al-Seraih, A.; Belguesmia, Y.; Baah, J.; Szunerits, S.; Boukherroub, R.; Drider, D. Enterocin B3A-B3B produced by LAB collected from infant faeces: Potential utilization in the food industry for Listeria monocytogenes biofilm management. Antonie Van Leeuwenhoek 2017, 110, 205–219. [Google Scholar] [CrossRef]
- Field, D.; Seisling, N.; Cotter, P.D.; Ross, R.P.; Hill, C. Synergistic Nisin-Polymyxin combinations for the control of Pseudomonas biofilm formation. Front. Microbiol. 2016, 7, 1713. [Google Scholar] [CrossRef] [Green Version]
- Hanchi, H.; Hammami, R.; Gingras, H.; Kourda, R.; Bergeron, M.G.; Ben Hamida, J.; Ouellette, M.; Fliss, I. Inhibition of MRSA and of Clostridium difficile by durancin 61A: Synergy with bacteriocins and antibiotics. Future Microbiol. 2017, 12, 205–212. [Google Scholar] [CrossRef]
- Montalbán-López, M.; Cebrián, R.; Galera, R.; Mingorance, L.; Martín-Platero, A.M.; Valdivia, E.; Martínez-Bueno, M.; Maqueda, M. Synergy of the Bacteriocin AS-48 and antibiotics against uropathogenic Enterococci. Antibiotics 2020, 9, 567. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Baah, J.; Hober, D.; Jouy, E.; Rubrecht, C.; Sané, F.; Drider, D. Synergistic effect between colistin and bacteriocins in controlling Gram-negative pathogens and their potential to reduce antibiotic toxicity in mammalian epithelial cells. Antimicrob. Agents Chemother. 2013, 57, 2719–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naghmouchi, K.; Le Lay, C.; Baah, J.; Drider, D. Antibiotic and antimicrobial peptide combinations: Synergistic inhibition of Pseudomonas fluorescens and antibiotic-resistant variants. Res. Microbiol. 2012, 163, 101–108. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Belguesmia, Y.; Baah, J.; Teather, R.; Drider, D. Antibacterial activity of class I and IIa bacteriocins combined with polymyxin E against resistant variants of Listeria monocytogenes and Escherichia coli. Res. Microbiol. 2011, 162, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Naghmouchi, K.; Drider, D.; Baah, J.; Teather, R. Nisin A and Polymyxin B as synergistic inhibitors of gram-positive and gram-negative bacteria. Probiotics Antimicrob. Proteins 2010, 2, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.M.; Brown, R.M.; Ashcraft, D.S.; Pankey, G.A. Synergistic effect between nisin and polymyxin B against pandrug-resistant and extensively drug-resistant Acinetobacter baumannii. Int. J. Antimicrob. Agents 2019, 53, 663–668. [Google Scholar] [CrossRef]
- Leffler, D.A.; Lamont, L. Clostridium difficile Infection. Available online: https://pubmed.ncbi.nlm.nih.gov/25875259/ (accessed on 16 September 2020).
- Lay, C.L.; Dridi, L.; Bergeron, M.G.; Ouellette, M.; Fliss, I.L. Nisin is an effective inhibitor of Clostridium difficile vegetative cells and spore germination. J. Med. Microbiol. 2016, 65, 169–175. [Google Scholar] [CrossRef]
- Mathur, H.; O’Connor, P.M.; Hill, C.; Cotter, P.D.; Ross, R.P. Analysis of anti-Clostridium difficile activity of Thuricin CD, Vancomycin, Metronidazole, Ramoplanin, and Actagardine, both singly and in paired combinations. Antimicrob. Agents Chemother. 2013, 57, 2882–2886. [Google Scholar] [CrossRef] [Green Version]
- Rea, M.C.; Clayton, E.; O’Connor, P.M.; Shanahan, F.; Kiely, B.; Ross, R.P.; Hill, C. Antimicrobial activity of lacticin 3,147 against clinical Clostridium difficile strains. J. Med. Microbiol. 2007, 56, 940–946. [Google Scholar] [CrossRef]
- Boakes, S.; Ayala, T.; Herman, M.; Appleyard, A.N.; Dawson, M.J.; Cortés, J. Generation of an actagardine A variant library through saturation mutagenesis. Appl. Microbiol. Biotechnol. 2012, 95, 1509–1517. [Google Scholar] [CrossRef]
- Danesh, A.; Ljungh, Å.; Mattiasson, B.; Mamo, G. Synergistic effect of haloduracin and chloramphenicol against clinically important gram-positive bacteria. Biotechnol. Rep. 2016, 13, 37–41. [Google Scholar] [CrossRef]
- Sarhan, S.R.; Ibrahim, O.M.S. In-vitro study the antibacterial activity of Bacteriocin against Stenotrophomonas maltophilia and evaluation its synergism with some antibiotics. Adv. Anim. Vet. Sci. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.C.; Ross, R.P.; Hill, C. Nisin Z. and Lacticin 3147 improve efficacy of antibiotics against clinically significant bacteria. Future Microbiol. 2019, 14, 1573–1587. [Google Scholar] [CrossRef]
- Kitazaki, K.; Koga, S.; Nagatoshi, K.; Kuwano, K.; Zendo, T.; Nakayama, J.; Sonomoto, K.; Ano, H.; Katamoto, H. In vitro synergistic activities of cefazolin and nisin A against mastitis pathogens. J. Vet. Med. Sci. 2017, 79, 1472–1479. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, Z.A.; Jamal, A.M.; Malik, A.H.; Zaidi, S.M.J.; Abdul Rahim, N.U.; Arshad, D. Exploring antimicrobial resistance in agents causing urinary tract infections at a tertiary care hospital in a developing country. Cureus 2020, 12, e9735. [Google Scholar] [CrossRef] [PubMed]
- Bono, M.J.; Reygaert, W.C. Urinary tract infection. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Wagenlehner, F.M.E.; Bjerklund Johansen, T.E.; Cai, T.; Koves, B.; Kranz, J.; Pilatz, A.; Tandogdu, Z. Epidemiology, definition and treatment of complicated urinary tract infections. Nat. Rev. Urol. 2020, 17, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Balto, H.A.; Shakoor, Z.A.; Kanfar, M.A. Combined effect of a mixture of tetracycline, acid, and detergent, and Nisin against Enterococcus faecalis and Actinomyces viscosus biofilms. Saudi Med. J. 2015, 36, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Holo, H. Synergistic antimicrobial activity between the broad spectrum Bacteriocin Garvicin KS and Nisin, Farnesol and Polymyxin B against gram-positive and gram-negative bacteria. Curr. Microbiol. 2018, 75, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Biswas, K.; Upadhayay, S.; Rapsang, G.F.; Joshi, S.R. Antibacterial and synergistic activity against β-Lactamase-producing nosocomial bacteria by bacteriocin of LAB isolated from lesser known traditionally fermented products of India. HAYATI J. Biosci. 2017, 24, 87–95. [Google Scholar] [CrossRef]
- Tremblay, Y.D.N.; Hathroubi, S.; Jacques, M. Les biofilms bactériens: Leur importance en santé animale et en santé publique. Can. J. Vet. Res. 2014, 78, 110–116. [Google Scholar]
- Cao, Y.; Naseri, M.; He, Y.; Xu, C.; Walsh, L.J.; Ziora, Z.M. Non-antibiotic antimicrobial agents to combat biofilm-forming bacteria. J. Glob. Antimicrob. Resist. 2020, 21, 445–451. [Google Scholar] [CrossRef]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El Jaziri, M. The Formation of Biofilms by Pseudomonas aeruginosa: A Review of the Natural and Synthetic Compounds Interfering with Control Mechanisms. Available online: https://www.hindawi.com/journals/bmri/2015/759348/ (accessed on 16 September 2020).
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Fighting biofilms with lantibiotics and other groups of bacteriocins. NPJ Biofilms Microbiomes 2018, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, A.; Field, D.; Pérez-Ibarreche, M.; Warda, A.K.; Hill, C.; Ross, R.P. Vancomycin and nisin A are effective against biofilms of multi-drug resistant Staphylococcus aureus isolates from human milk. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Naveen, S.V.; Kalaivani, K. Cancer stem cells and evolving novel therapies: A paradigm shift. Stem Cell Investig. 2018, 5, 4. [Google Scholar] [CrossRef]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef]
- Kamarajan, P.; Hayami, T.; Matte, B.; Liu, Y.; Danciu, T.; Ramamoorthy, A.; Worden, F.; Kapila, S.; Kapila, Y. Nisin ZP, a Bacteriocin and food preservative, inhibits head and neck cancer tumorigenesis and prolongs survival. PLoS ONE 2015, 10, e0131008. [Google Scholar] [CrossRef] [Green Version]
- Preet, S.; Bharati, S.; Panjeta, A.; Tewari, R.; Rishi, P. Effect of nisin and doxorubicin on DMBA-induced skin carcinogenesis--a possible adjunct therapy. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 8301–8308. [Google Scholar] [CrossRef] [PubMed]
- Boverhof, D.R.; Bramante, C.M.; Butala, J.H.; Clancy, S.F.; Lafranconi, M.; West, J.; Gordon, S.C. Comparative assessment of nanomaterial definitions and safety evaluation considerations. Regul. Toxicol. Pharmacol. 2015, 73, 137–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarschler, K.; Rocks, L.; Licciardello, N.; Boselli, L.; Polo, E.; Garcia, K.P.; De Cola, L.; Stephan, H.; Dawson, K.A. Ultrasmall inorganic nanoparticles: State-of-the-art and perspectives for biomedical applications. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1663–1701. [Google Scholar] [CrossRef]
- Kwizera, E.A.; Chaffin, E.; Shen, X.; Chen, J.; Zou, Q.; Wu, Z.; Gai, Z.; Bhana, S.; O’Connor, R.; Wang, L.; et al. Size and shape-controlled synthesis and properties of magnetic-plasmonic core-shell nanoparticles. J. Phys. Chem. C Nanomater. Interfaces 2016, 120, 10530. [Google Scholar] [CrossRef] [Green Version]
- Betzer, O.; Shilo, M.; Opochinsky, R.; Barnoy, E.; Motiei, M.; Okun, E.; Yadid, G.; Popovtzer, R. The effect of nanoparticle size on the ability to cross the blood-brain barrier: An in vivo study. Nanomedicine 2017, 12, 1533–1546. [Google Scholar] [CrossRef]
- Lalwani, G.; Sitharaman, B. Multifunctional fullerene- and metallofullerene-based nanobiomaterials. Nano LIFE 2013, 3, 1342003. [Google Scholar] [CrossRef]
- Lu, J.; Tang, M.; Zhang, T. Review of toxicological effect of quantum dots on the liver. J. Appl. Toxicol. 2019, 39, 72–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AbdelAllah, N.H.; Gaber, Y.; Rashed, M.E.; Azmy, A.F.; Abou-Taleb, H.A.; AbdelGhani, S. Alginate-coated chitosan nanoparticles act as effective adjuvant for hepatitis A vaccine in mice. Int. J. Biol. Macromol. 2020, 152, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; El-Banna, H.A.; Elzorba, H.Y.; Galal, A.M. Application of some nanoparticles in the field of veterinary medicine. Int. J. Vet. Sci. Med. 2019, 7, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Oh, J. Marine polysaccharide-based nanomaterials as a novel source of nanobiotechnological applications. Int. J. Biol. Macromol. 2016, 82, 315–327. [Google Scholar] [CrossRef]
- Ngwuluka, N.C. 19 responsive polysaccharides and polysaccharides-based nanoparticles for drug delivery. In Stimuli Responsive Polymeric Nanocarriers for Drug Delivery Applications; Makhlouf, A.S.H., Abu-Thabit, N.Y., Eds.; Woodhead Publishing Series in Biomaterials; Woodhead Publishing: Cambridge, UK, 2018; Volume 1, pp. 531–554. ISBN 978-0-08-101997-9. [Google Scholar]
- No, D.S.; Algburi, A.; Huynh, P.; Moret, A.; Ringard, M.; Comito, N.; Drider, D.; Takhistov, P.; Chikindas, M.L. Antimicrobial efficacy of curcumin nanoparticles against Listeria monocytogenes is mediated by surface charge. J. Food Saf. 2017, 37, e12353. [Google Scholar] [CrossRef]
- Perinelli, D.R.; Fagioli, L.; Campana, R.; Lam, J.K.W.; Baffone, W.; Palmieri, G.F.; Casettari, L.; Bonacucina, G. Chitosan-based nanosystems and their exploited antimicrobial activity. Eur. J. Pharm. Sci. 2018, 117, 8–20. [Google Scholar] [CrossRef]
- Akbar, A.; Shakeel, A. A review on chitosan and its nanocomposites in drug delivery. Int. J. Biol. Macromol. 2018, 109, 273–286. [Google Scholar] [CrossRef]
- Borel, T.; Sabliov, C.M. Nanodelivery of bioactive components for food applications: Types of delivery systems, properties, and their effect on ADME profiles and toxicity of nanoparticles. Annu. Rev. Food Sci. Technol. 2014, 5, 197–213. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoscale nutrient delivery systems for food applications: Improving bioactive dispersibility, stability, and bioavailability. J. Food Sci. 2015, 80, N1602–N1611. [Google Scholar] [CrossRef] [PubMed]
- Filli, M.S.; Ibrahim, A.A.; Aquib, M.; Abbas, A.B.; Morshed, A.; Boakye-Yiadom, K.O.; Kesse, S.; Farooq, M.A.; Bazezy, M.; Wang, B. The impact of physicochemical characteristics on therapeutic efficacy of anticancer nanomaterials: A review. Int. J. Pharm. Sci. Drug Res. 2019, 11, 61–70. [Google Scholar] [CrossRef]
- Sidhu, P.K.; Nehra, K. Bacteriocin-nanoconjugates as emerging compounds for enhancing antimicrobial activity of bacteriocins. J. King Saud Univ. Sci. 2019, 31, 758–767. [Google Scholar] [CrossRef]
- Zimet, P.; Mombrú, Á.W.; Faccio, R.; Brugnini, G.; Miraballes, I.; Rufo, C.; Pardo, H. Optimization and characterization of Nisin-loaded Alginate-Chitosan nanoparticles with antimicrobial activity in lean beef. LWT 2018, 91, 107–116. [Google Scholar] [CrossRef]
- Niaz, T.; Shabbir, S.; Noor, T.; Imran, M. Antimicrobial and antibiofilm potential of bacteriocin loaded nano-vesicles functionalized with rhamnolipids against foodborne pathogens. LWT 2019, 116, 108583. [Google Scholar] [CrossRef]
- Goudarzi, F.; Asadi, A.; Afsharpour, M.; Jamadi, R.H. In vitro characterization and evaluation of the cytotoxicity effects of Nisin and Nisin-loaded PLA-PEG-PLA nanoparticles on gastrointestinal (AGS and KYSE-30), hepatic (HepG2) and blood (K562) cancer cell lines. AAPS PharmSciTech 2018, 19, 1554–1566. [Google Scholar] [CrossRef]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primer 2018, 4, 18033. [Google Scholar] [CrossRef]
- Doss, J.; Culbertson, K.; Hahn, D.; Camacho, J.; Barekzi, N. A review of Phage therapy against bacterial pathogens of aquatic and terrestrial organisms. Viruses 2017, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baños, A.; García-López, J.D.; Núñez, C.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Biocontrol of Listeria monocytogenes in fish by Enterocin AS-48 and Listeria lytic bacteriophage P100. LWT Food Sci. Technol. 2016, 66, 672–677. [Google Scholar] [CrossRef]
- Kim, S.G.; Lee, Y.D.; Park, J.H.; Moon, G.S. Synergistic inhibition by Bacteriocin and Bacteriophage against Staphylococcus aureus. Food Sci. Anim. Resour. 2019, 39, 1015–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firenzuoli, F.; Jaitak, V.; Horvath, G.; Bassolé, I.H.N.; Setzer, W.N.; Gori, L. Essential oils: New perspectives in human health and wellness. Evid.-Based Complement. Altern. Med. ECAM 2014, 2014, 467363. [Google Scholar] [CrossRef]
- Ay, Z.; Tuncer, Y. Combined antimicrobial effect of Nisin, Carvacrol and EDTA against Salmonella Typhimurium in TSBYE at 4°C and 37°C. Rom. Biotechnol. Lett. 2016, 21, 11666–11674. [Google Scholar]
- Mehdizadeh, T.; Hashemzadeh, M.S.; Nazarizadeh, A.; Neyriz-Naghadehi, M.; Tat, M.; Ghalavand, M.; Dorostkar, R. Chemical composition and antibacterial properties of Ocimum basilicum, Salvia officinalis and Trachyspermum ammi essential oils alone and in combination with nisin. Res. J. Pharmacogn. 2016, 3, 51–58. [Google Scholar]
- Shi, C.; Zhang, X.; Zhao, X.; Meng, R.; Liu, Z.; Chen, X.; Guo, N. Synergistic interactions of nisin in combination with cinnamaldehyde against Staphylococcus aureus in pasteurized milk. Food Control 2017, 71, 10–16. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.K.; Draper, L.A.; Hazelhoff, P.J.; Cotter, P.D.; Ross, R.P.; Hill, C.A. Bioengineered Nisin derivative, M21A, in combination with food grade additives eradicates biofilms of Listeria monocytogenes. Front. Microbiol. 2016, 7, 1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Bacteriocins Classification Scheme According to: | ||
|---|---|---|
| Klaenhammer [1] | Drider et al. [6] | Alvarez-Sieiro et al. [7] |
| Class I Small modified peptides < 5 kDa; containing specific amino acids (lanthionine and dehydrated residues. | Class I Small peptides with post-translational modification (lanthionines and dehydrated residues) Remark: This class was subdivided in two types: Type A (elongated molecules with molecular mass < 4 kDa) Type B (globular molecules with molecular mass between 1.8 and 2.1 kDa) | Class I Ribosomally produced and post-translationally modified peptides (RiPPs) (<10 kDa) Remark: Five subclasses were assigned to class I bacteriocins: Ia: lanthipeptides (types I, II, III, and IV) Ib: Cyclized peptides (Enterocin AS-48) Ic: Sactibiotics Id: linear azol(in)e-containing peptides Ie: Glycocins If: Lasso peptides |
| Class II Small unmodified peptides (<10 kDa); thermostables; acting by pore formation in the membranes | Class II Small heat-stable unmodified peptides (<10 kDa) Remark: Three subclasses were retained IIa: Pediocin-like bacteriocins IIb: Two-peptide bacteriocins IIc: Other peptide bacteriocins including cyclized ones (enterocin AS-48) | Class II Unmodified peptides (<10 kDa) Remark: Four subclasses were proposed IIa: Pediocin-like bacteriocins IIb: Two-peptides bacteriocins IIc: Leaderless bacteriocins IId: Non pediocin-like single-peptide bacteriocins (ex: laterosporulin) including cyclized ones (enterocin AS–48) |
| Class III Proteins with molecular weight > 30 kDa; enzymatic activity | Class III Large, heat-labile (>10 kDa) protein | Class III Thermo-labile (>10 kDa) bacteriolysins and non-lytic bacteriocins |
| Class IV Proteins attached to sugars or lipids | ||
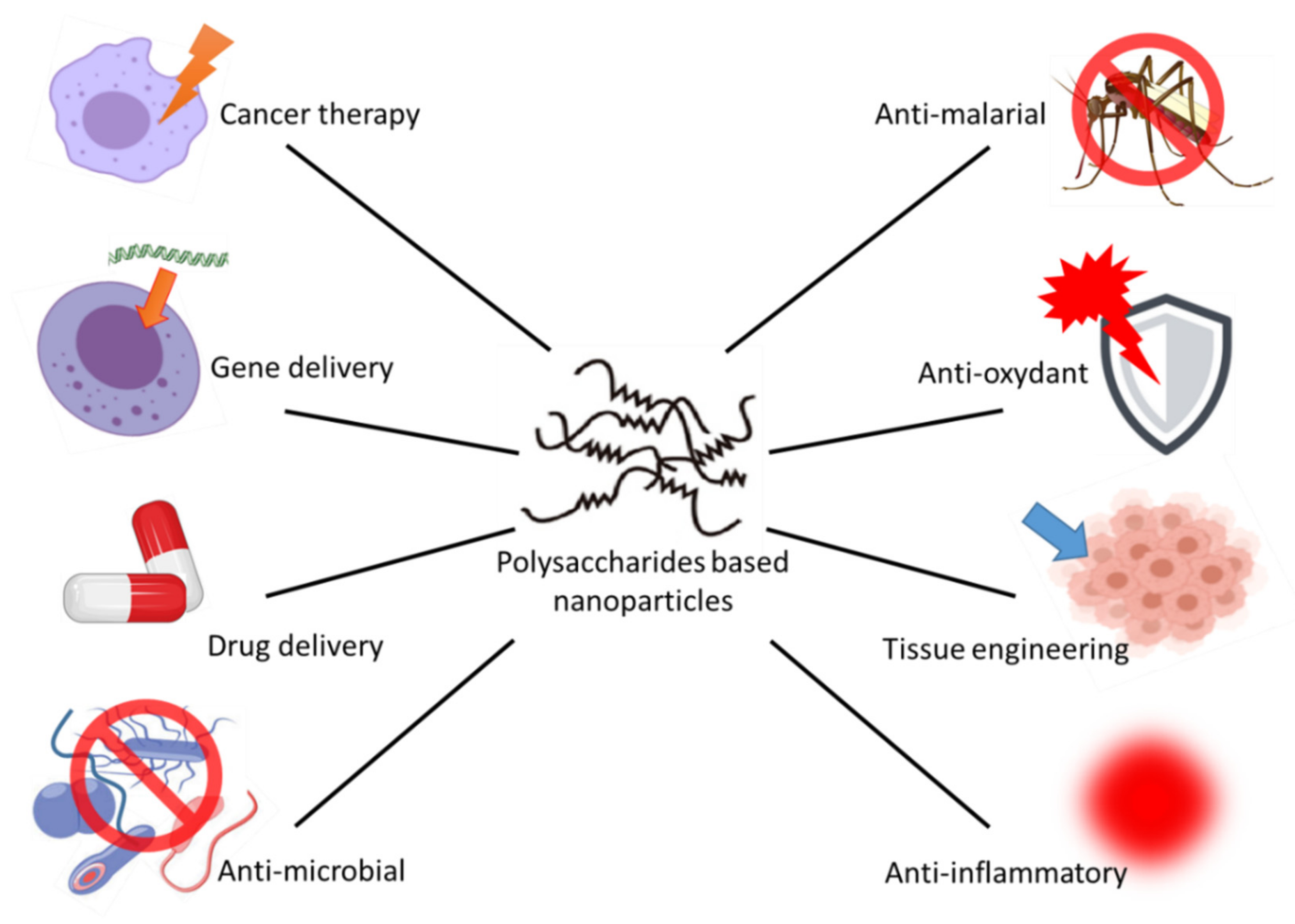
| Polysaccharides | Origin | Applications of Its NPs |
|---|---|---|
| Alginate | Extracted from brown algae (Phaeophyceae) | Gene and drug delivery system, tissue engineering, etc. |
| Cyclodextrin | Enzymatic conversion of starch | Gene and drug delivery, controlled drug release |
| Starch | Higher plants | Insulin controlled release & drug delivery |
| Pectin | Extracted from the middle lamellae of plant cells | Wound healing, delivery of amino acids and drugs |
| Chitosan | Extracted from the cell walls of fungi and the exoskeleton of crustaceans (crabs, shrimp) | Nanocarrier in drug delivery and antimicrobial |
| Cellulose | Cell wall of green plants | Nanodelivery for oral protein, for anticancer drugs, etc. |
| Bacteriocin-NPs Combinations | Applications | Targets | References |
|---|---|---|---|
| Nisin-alginate-chitosan NPs | Antibacterial | L. monocytogenes | [79] |
| Nisin-alginate-chitosan NPs | Antibacterial | S. aureus L. monocytogenes E. coli S. Typhimurium | [82] |
| Nisin-nano vesicles | Antibiofilm | S. aureus L. monocytogenes E. coli, P. aeruginosa | [80] |
| Nisin-PLA-PEG-PLA NPs | Anticancer | Cancer cell lines: Gastrointestinal (AGS and KYSE-30) Hepatic (HepG2) Blood (K562) | [81] |
| Bacteriocin-EO Combinations | Target Bacteria/Yeast |
|---|---|
| Nisin-Salvia officinalis | E. coli O 157 |
| Nisin-Carvacrol | S. Typhimurium |
| Nisin-Cinnamaldehyde | S. aureus |
| M21A- Cinnamaldehyde | L. monocytogenes |
| Bacteriocin bacLP17- Thymus vulgaris, Salvia officinalis | L. monocytogenes |
| Enterocin KT2W2G- Cinnamon | Bacteria:Serratia marcescens, Klebsiella variicola, Enterococcus faecalis, Lactococcus lactis subsp. lactis, Klebsiella pneumoniae. Yeasts: Hanseniaspora opuntiae, Pichia aff. fermentans, Candida metapsilosis, Pichia kundriavzevii and Kodamaea ohmeri. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zgheib, H.; Drider, D.; Belguesmia, Y. Broadening and Enhancing Bacteriocins Activities by Association with Bioactive Substances. Int. J. Environ. Res. Public Health 2020, 17, 7835. https://doi.org/10.3390/ijerph17217835
Zgheib H, Drider D, Belguesmia Y. Broadening and Enhancing Bacteriocins Activities by Association with Bioactive Substances. International Journal of Environmental Research and Public Health. 2020; 17(21):7835. https://doi.org/10.3390/ijerph17217835
Chicago/Turabian StyleZgheib, Hassan, Djamel Drider, and Yanath Belguesmia. 2020. "Broadening and Enhancing Bacteriocins Activities by Association with Bioactive Substances" International Journal of Environmental Research and Public Health 17, no. 21: 7835. https://doi.org/10.3390/ijerph17217835