
Hippobosca equina L. (Hippoboscidae: Hippobosca)—An Old Enemy as an Emerging Threat in the Palearctic Zone


Abstract
:1. Introduction
2. Characteristics of the Species
3. Species Distribution in the World
4. Hosts
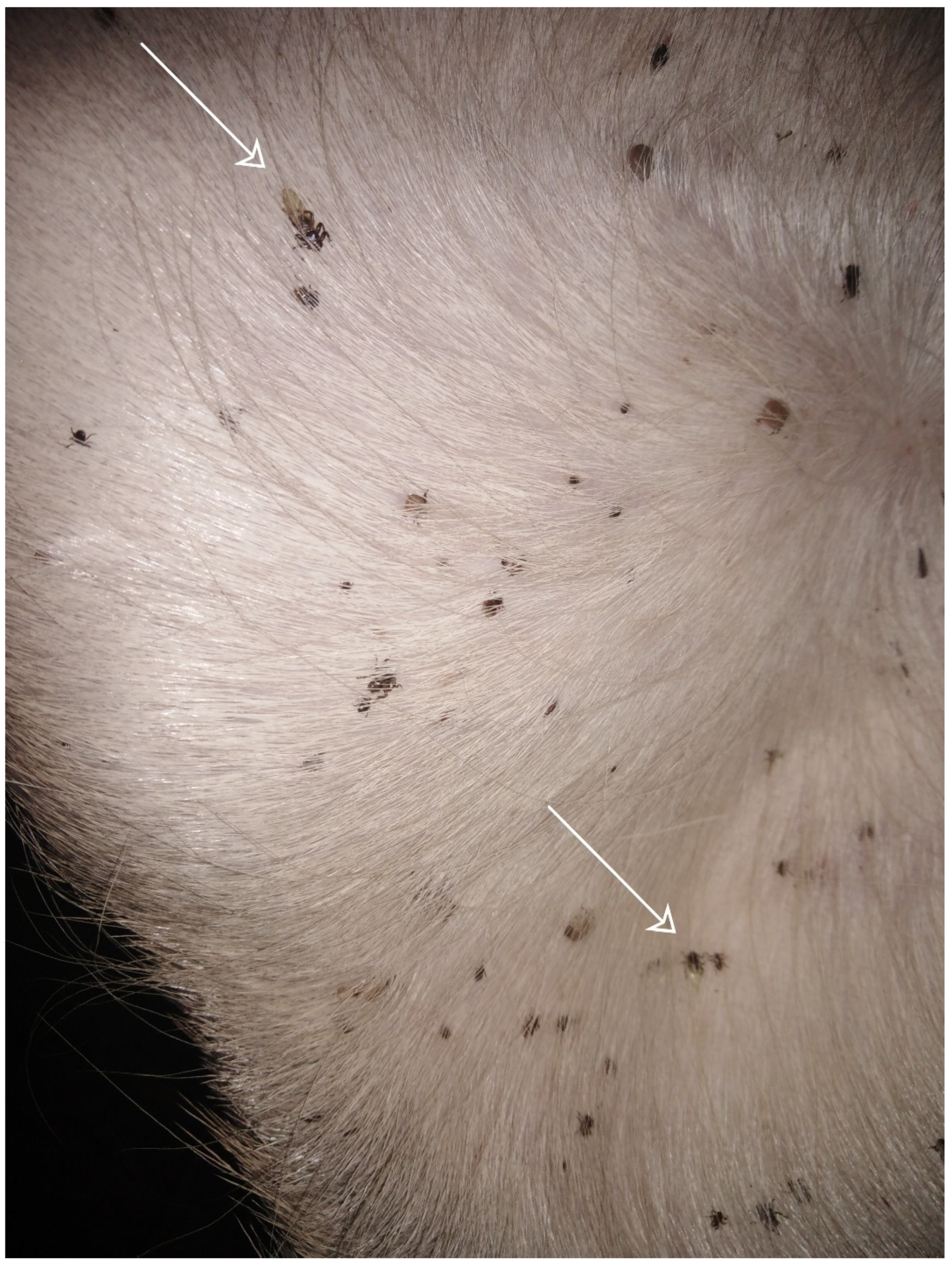
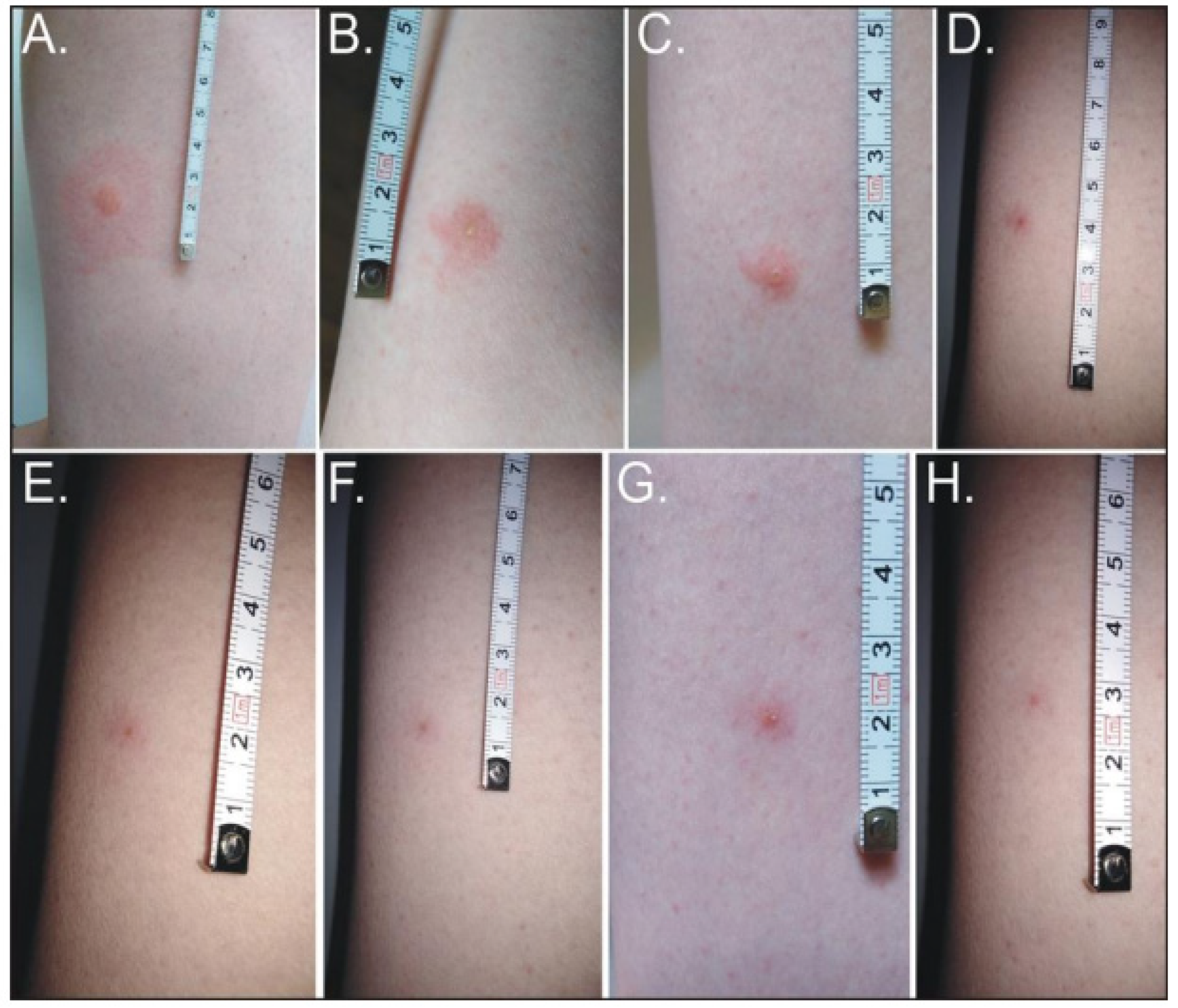
5. Medical and Veterinary Importance of Hippobosca equina
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kowal, J.; Kornaś, S.; Nosal, P.; Wajdzik, M.; Basiaga, M.; Lesiak, M. Parasite infections in red deer Cervus elaphus from Krakow area, southern Poland. Ann. Parasitol. 2015, 61, 49–52. [Google Scholar] [PubMed]
- Gałęcki, R.; Xuan, X.; Bakuła, T. Migration of deer keds to urban agglomerations—A case study. Ann Parasitol. 2022, 68, S2–S232. [Google Scholar]
- Sokół, R.; Michalski, M. Occurrence of Hippobosca equina in Polish primitive horses during the grazing season. Ann. Parasitol. 2015, 61, 118–122. [Google Scholar]
- Kadulski, S. Materiały do znajomości Hippoboscidae (Diptera-Pupipara) ssaków użytkowych Polski. Wiadomości Parazytol. 1970, 16, 473–477. [Google Scholar]
- Oboňa, J.; Krišovský, P.; Hromada, M. Short-term faunistic sampling of Louse flies (Diptera: Hippoboscidae) from Drienovec Bird Ringing Station, Slovakia. Biodivers. Environ. 2019, 11, 4–9. [Google Scholar]
- Borowiec, L. Klucze do Oznaczania Owadów Polski. Część XXVIII. Muchówki—Diptera. Zeszyt 77, Wpleszczowate—Hippoboscidae; Państwowe Wydawnictwo Naukowe PWN: Warszawa, Poland, 1984; pp. 9–30. [Google Scholar]
- Bequaert, J. Notes on Hippoboscidae. 2 The Subfamily Hippoboscinae. Psyche 1930, 37, 303–326. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liu, X.H.; Li, X.Y.; Cao, J.; Chu, H.J.; Li, K. Ultrastructural investigation of antennae in three cutaneous myiasis flies: Melophagus ovinus, Hippobosca equina, and Hippobosca longipennis (Diptera: Hippoboscidae). Parasitol. Res. 2015, 114, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Boucheikhchoukh, M.; Mechouk, N.; Benakhla, A.; Raoult, D.; Parola, P. Molecular evidence of bacteria in Melophagus ovinus sheep keds and Hippobosca equina forest flies collected from sheep and horses innortheastern Algeria. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 103–109. [Google Scholar] [CrossRef]
- Andreani, A.; Sacchetti, P.; Belcari, A. Evolutionary adaptations in four hippoboscid fly species belonging to three different subfamilies. Med. Vet. Entomol. 2020, 34, 344–363. [Google Scholar] [CrossRef]
- Andreani, A.; Sacchetti, P.; Belcari, A. Keds and Bat Flies (Hippoboscidae, Nycteribiidae and Streblidae). In Reference Module in Biomedical Sciences, Encyklopedia of Infection and Immunity; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 1–18. [Google Scholar]
- Andreani, A.; Belcari, A.; Sacchetti, P.; Romani, R. Antennal Morphology and Fine Structure of Flagellar Sensilla in Hippoboscid Flies with Special Reference to Lipoptena fortisetosa (Diptera: Hippoboscidae). Insects 2022, 13, 236. [Google Scholar] [CrossRef]
- Jedlička, L.; Stloukalová, V. Červený (ekosozoologický) zoznam dvojkrídlovcov (Diptera) Slovenska. In Červený Zoznam Rastlín Aživočíchov Slovenska; Supplementum 20; Baláž, D., Marhold, K., Urban, P., Eds.; Ochrana Prírody: Banská Bystrica, Slovakia, 2001; pp. 139–142. [Google Scholar]
- Oboňa, J.; Oldřich, S.; Greš, S.; Heřman, P.; Manko, P.; Roháček, J.; Šestáková, A.; Šlapák, J.J.; Hromada, M. A revised annotated checklist of louse flies (Diptera, Hippoboscidae) from Slovakia. ZooKeys 2019, 862, 129–152. [Google Scholar] [CrossRef] [PubMed]
- Romaniuk, K.; Gad, K.; Kiszka, W. Występowanie muchówki Hippobosca equina u koników polskich. Med. Weter. 2007, 63, 1100–1101. [Google Scholar]
- Johnsen, P. Notes on the Danish louse-flies (Diptera: Hippoboscidae). Entomol. Medd. 1948, 25, 278–298. [Google Scholar]
- Mehlhorn, H. Hippoboscidae. In Encyclopedia of Parasitology; Springer: Berlin/Heidelberg, Germany, 2016; p. 1252. [Google Scholar] [CrossRef]
- Haarløv, N. Life Cycle and Distribution Pattern of Lipoptena cervi (L.) (Dipt., Hippobosc.) on Danish Deer. Oikos 1964, 15, 93–129. [Google Scholar] [CrossRef]
- Kowal, J.; Nosal, P.; Kornaś, S.; Wajdzik, M.; Matysek, M.; Basiaga, M. Różnorodność i znaczenie muchówek z rodziny narzępikowatych—Pasożytów jeleniowatych. Med. Weter. 2016, 72, 745–749. [Google Scholar] [CrossRef]
- Soliman, S.M.; Attia, M.M.; Al-Harbi, M.S.; Saad, A.M.; El-Saadony, M.T.; Salem, H.M. Low host specificity of Hippobosca equina infestation in different domestic animals and pigeon. Saudi J. Biol. Sci. 2022, 29, 2112–2120. [Google Scholar] [CrossRef]
- Kulczycki, W. Owady Pasorzytujące u Ludzi i Zwierząt Domowych; Główny Skład w Księgarni Gubrynowicza i Schmidta: Lwów, Poland, 1892; pp. 1–68. [Google Scholar]
- Pfadt, R.E.; Roberts, I.H. Louse flies (family Hippoboscidae). In Surveillance and Collection of Arthropods of Veterinary Importance; Agriculture Handbook No. 518; Bram, R.A., Ed.; U.S. Department of Agriculture: Washington, DC, USA, 1978; pp. 60–71. [Google Scholar]
- Zittra, C.; Schoener, E.R.; Wagner, R.; Heddergott, M.; Duscher, G.G.; Fuehrer, H.-P. Unnoticed arrival of two dipteran species in Austria: The synanthropic moth fly Clogmia albipunctata (Williston, 1893) and the parasitic bird louse fly Ornithoica turdi (Olivier in Latreille, 1811). Parasitol. Res. 2020, 119, 737–740. [Google Scholar] [CrossRef] [Green Version]
- Wallach, A.; Shanas, U.; Mumcuoglu, K.Y.; Inbar, M. Ectoparasites on Reintroduced Roe Deer Capreolus capreolus in Israel. J. Wildl. Dis. 2008, 44, 693–696. [Google Scholar] [CrossRef] [Green Version]
- El-Hawagry, M.S.; Khalil, M.W.; Sharaf, M.R.; Fadl, H.H.; Aldawood, A.S. A preliminary study on the insect fauna of Al-Baha Province, Saudi Arabia, with descriptions of two new species. ZooKeys 2013, 274, 1–88. [Google Scholar] [CrossRef]
- Romaniuk, K. Występowanie muchówek u krów i koników polskich przebywających na pastwisku. Med. Weter. 2004, 61, 332–334. [Google Scholar]
- Łabędzki, A. Krwiopijcy z lasów i łąk. Wiedza Życie 2012, 9, 28–31. [Google Scholar]
- Abubakar, B.A.; Falmata, K.; ThankGod, O.E.; Abdulmalik, A.; Ali, M. Survey of flies (order: Diptera) of Medical and Veterinary importance infesting livestock in Maiduguri, Borno state, Nigeria. J. Sci. Agric. 2018, 2, 97–100. [Google Scholar] [CrossRef]
- Kameneva, E.P. B—Diptera—Ulidiidae. In 11. Lista dos Artrópodes (Arthropoda); Borges, P.A.V., Costa, A., Cunha, R., Gabriel, R., Gonçalves, V., Martins, A.F., Melo, I., Parente, M., Raposeiro, P., Rodrigues, P., et al., Eds.; Princípia, Cascais: Parede, Portugal, 2010; pp. 179–246. [Google Scholar]
- Vázquez, L.; Dacal, V.; Pato, F.J.; Díaz, P.; Painceira, A.; Fernández, G.; Morrondo, P.; Diez-Baños, P. Ectoparásitos presentes en corzos (Capreolus capreolus) de Galicia (NO España). Galemys 2010, 22, 243–253. [Google Scholar]
- Halos, L.; Jamal, T.; Maillard, R.; Girard, B.; Guillot, J.; Chomel, B.; Vayssier-Taussat, M.; Boulouis, H.J. Role of Hippoboscidae flies as potential vectors of Bartonella spp. infecting wild and domestic ruminants. Appl. Environ. Microbiol. J. 2004, 70, 302–305. [Google Scholar] [CrossRef] [Green Version]
- Liénard, E.; Salem, A.; Grisez, C.; Prévot, F.; Bergeaud, J.P.; Franc, M.; Gottstein, B.; Alzieu, J.P.; Lagalisse, Y.; Jacquiet, P. A longitudinal study of Besnoitia besnoiti infections and seasonal abundance of Stomoxys calcitrans in a dairy cattle farm of southwest France. Vet. Parasitol. 2011, 177, 20–27. [Google Scholar] [CrossRef]
- Hutson, A.M. Keds, flat-flies and bat-flies. Diptera, Hippoboscidae and Nycteribiidae. In Handbooks for the Identification of British Insects; Fitton, M.G., Ed.; Royal Entomological Society of London: London, UK, 1984; Volume 10, part 7; pp. 1–40. [Google Scholar]
- Turner, C.R.; Mann, D.J. Recent observations of Hippobosca equina L. (Diptera: Hippoboscidae) in South Devon. Br. J. Entomol. Nat. Hist. 2004, 17, 1274. [Google Scholar]
- Chandler, P.J. An Update of the 1998 Checklist of Diptera of the British Isles; Updated 28 July 2021; Royal Entomological Society of London: London, UK, 2021; pp. 1–440. [Google Scholar]
- Grootaert, P.; De Bruyn, L.; De Meyer, M. Catalogue of the Diptera of Belgium; Koninklijk Belgisch Instituut voor Natuurwetenschappen: Brussel, Belgium, 1991; Volume 70, pp. 1–338. [Google Scholar]
- Beuk, P.L.T. (Ed.) Family Hippoboscidae. In Checklist of the Diptera of the Netherlands. 2001. Available online: https://diptera-info.nl/news.php?fam=Hippoboscidae (accessed on 20 October 2022).
- Jentzsch, M.; Müller, J. Lausfliegen (Diptera: Hippoboscidae). In Pflanzen und Tiere in Sachsen-Anhalt; Frank, D., Schnitter, P., Eds.; Vom Landesamt für Umweltschutz Sachsen-Anhalt: Halle, Germany, 2016; pp. 1129–1132. [Google Scholar]
- Jentzsch, M.; Knauthe, C.H. Louse fly collections of institutes and museums in Saxony (Diptera, Hippoboscidae). Entomol. Nachr. Ber. 2019, 63, 134–141. [Google Scholar]
- Schröter, S.; Freick, M.; Vogt, I.; Jentzsch, M. Lausfliegen (Hippoboscidae: Diptera) als Vektoren für Bakterien mit Zoonosepotential bei Säugetieren. Beiträge Jagd Wildforschung 2020, 45, 193–198. [Google Scholar]
- Draber-Mońko, A.; Durska, E.; Klasa, A.; Kownacki, A.; Krzemiński, W. 28. Diptera—Muchówki. In Checklist of Animals of Poland; Razowski, J., Ed.; Wydawnictwo Polskiej Akademii Nauk: Wroclav, Poland, 1991; Volume 2, pp. 77–269. [Google Scholar]
- Siebke, H. Catalogum Dipterorum Continentem. Programmatis Nomine Edidit Universitas Regia Fredericiana. 1877. Available online: http://www.entomologi.no/journals/enumeratio/Enumeratio_04_1877.pdf (accessed on 13 October 2022).
- Andersson, H. De svenska lusflugorna. The Swedish louse-flies (Diptera: Hippoboscidae). Entomol. Tidskr. 1985, 106, 15–25. [Google Scholar]
- Pohjoismäki, J.; Kahanpää, J. Checklist of the superfamilies Oestroidea and Hippoboscoidea of Finland (Insecta, Diptera). ZooKeys 2014, 441, 383–408. [Google Scholar] [CrossRef] [Green Version]
- Teder, T.; Tammaru, T. Insects of Britain and Western Europe. In Euroopa Putukad; Chinery, M., Ed.; Estonian Encyclopaedia Publishers: Tallinn, Estonia, 2005; p. 212. [Google Scholar]
- Karpa, A. Catalogue of Latvian Flies (Diptera: Brachycera). Latv. Entomol. 2008, 46, 4–43. [Google Scholar]
- Pakalniškis, S.; Bernotienė, R.; Lutovinovas, E.; Petrašiūnas, A.; Podėnas, S.; Rimšaitė, J.; Sæther, O.A.; Spungis, V. Checklist Of Lithuanian Diptera. New Rare Lith. Insect Species 2006, 18, 1–154. [Google Scholar] [CrossRef]
- Trilar, T.; Krčmar, S. Contribution to the knowledge of louse flies of Croatia (Diptera: Hippoboscidae). Nat. Croat. 2005, 14, 131–140. [Google Scholar]
- Fois, F.; Mereu Piras, P.; Cappai, S.; Cillo, D.; Culurgioni, J.; Deiana, A.M.; Mandas, L.; Rolesu, S. Contribution to the knowledge of Diptera Hippoboscidae in Sardinia. Mappe Parassitol. 2012, 18, 108. [Google Scholar]
- Гембiцкi, А.С.; Давыдава, Г.П. Весцi Акадэмii навук БССР. In Крывасысучыя Двухкрылыя Басейна Вoзера Нарач; Гембiцкi, А.С., Ed.; Yanka Kupala State University of Grodno: Grodno, Belarus, 1983; Volume 3, pp. 99–103. [Google Scholar]
- Billeter, S.A.; Levy, M.G.; Chomel, B.B.; Breitschwerdt, E.B. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med. Vet. Entomol. 2008, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Prelezov, P.; Nizamov, N. A case of multiple mixed invasion with ectoparasites in goats. Tradit. Mod. Vet. Med. 2020, 5, 73–78. [Google Scholar]
- Alexiou, S.; Gavalas, I. Calyptrate flies (Diptera) of Cyclades, Greece—I. Families Calliphoridae, Hippoboscidae, Oestridae, Polleniidae, Rhiniidae, Rhinophoridae and Scathophagidae. Studia Dipterol. 2017, 24, 99–105. [Google Scholar]
- Nartshuk, E.P.; Oboňa, J. The distribution of genus Hippobosca in Transcaucasia. Acta Musei Sil. Sci. Nat. 2019, 68, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Sazmand, A.; Bahari, A.; Papi, S.; Otranto, D. Parasitic diseases of equids in Iran (1931–2020): A literature review. Parasites Vectors 2020, 13, 586. [Google Scholar] [CrossRef]
- Selmi, R.; Dhibi, M.; Ben Said, M.; Ben Yahia, H.; Abdelaali, H.; Ameur, H.; Baccouche, S.; Gritli, A.; Mhadhbi, M. Evidence of natural infections with Trypanosoma, Anaplasma and Babesia spp. in military livestock fromTunisia. Trop. Biomed. 2019, 36, 742–757. [Google Scholar]
- Abdullah, H.H.A.M.; Aboelsoued, D.; Farag, T.K.; Abdel-Shafy, S.; Abdel Megeed, K.N.; Parola, P.; Raoult, D.; Mediannikov, O. Molecular characterization of some equine vector-borne diseases and associated arthropods in Egypt. Acta Trop. 2022, 227, 106274. [Google Scholar] [CrossRef] [PubMed]
- Seok, D.M. The Insect Fauna of the Island Quelpart; Po-Chin-Chai Co: Seoul, Republic of Korea, 1970. [Google Scholar]
- Kim, H.C.; Chong, S.T.; Chae, J.-S.; Lee, H.; Klein, T.A.; Suh, S.J.; Rueda, L.M. New Record of Lipoptena cervi and Updated Checklist of the Louse Flies (Diptera: Hippoboscidae) of the Republic of Korea. J. Med. Entomol. 2010, 47, 1227–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, P.A.M.; Coleman, G.T.; Irwin, P.J.; Traub, R.J. Hippobosca longipennis—A potential intermediate host of a species of Acanthocheilonema in dogs in northern India. Parasit Vectors 2011, 4, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maity, A.; Naskar, A.; Mitra, S.; Chakraborty, A.; Hazra, S.; Banerjee, D. Checklist of Indian Louse Flies (Insecta: Diptera: Hippoboscidae). Zsi Publ. 2014, 1–7. Available online: https://www.researchgate.net/profile/Aniruddha-Maity/publication/304580676_checklist_of_Indian_Louse_Flies_Insecta_Diptera_Hippoboscidae/links/5774104608aeb9427e241df5/checklist-of-Indian-Louse-Flies-Insecta-Diptera-Hippoboscidae.pdf (accessed on 13 October 2022).
- Sychra, O.; Literák, I.; Najer, T.; Čapek, M.; Koubek, P.; Procházka, P. Chewing lice (Insecta: Phthiraptera) from estrildid finches (Aves: Passeriformes: Estrildidae) and louse-flies (Insecta: Diptera: Hippoboscidae) from birds in Senegal, with descriptions of three new species of the genus Brueelia. Zootaxa 2010, 2714, 59–68. [Google Scholar] [CrossRef]
- Sychra, O.; Halajian, A.; Engelbrecht, D.; Symes, C.T.; Oschadleus, H.D.; de Swardt, D.H.; Papousek, I. Louse-flies (Diptera: Hippoboscidae) of birds from South Africa: Prevalence and diversity. Afr. Entomol. 2020, 28, 249–261. [Google Scholar] [CrossRef]
- Oboňa, T.J.; Zeegers, T.; Wamiti, W.; Njoroge, N. Additions to the Checklist of the Louse Flies (Diptera: Hippoboscidae) of Kenya. Afr. Entomol. 2016, 24, 393–397. [Google Scholar] [CrossRef]
- Rachola, N.; Goodman, S.M.; Robert, V. The Hippoboscidae (Insecta: Diptera) from Madagascar, with new records from the “Parc National de Midongy Befotaka”. Parasite 2011, 18, 127–140. [Google Scholar] [CrossRef]
- Kazimierczak, K.; Górski, P. Narzępikowate w Polsce—biologia i znaczenie. Życie Weter. 2007, 82, 51–53. [Google Scholar]
- Bezerra-Santos, M.A.; Otranto, D. Keds, the enigmatic flies and their role as vectors of pathogens. Acta Trop. 2020, 209, 105521. [Google Scholar] [CrossRef]
- Dik, B.; Ceylan, O.; Ceylan, C.; Tekindal, M.A.; Semassel, A.; Sönmez, G.; Ekici, Ö.D. Ectoparasites of feral horses (Equus ferus caballus (Linnaeus., 1758)) on Karadağ Mountain, Karaman, Turkey. J. Parasit. Dis. 2020, 44, 590–596. [Google Scholar] [CrossRef]
- Maa, T.C. A revised checklist and concise host index of Hippoboscidae (Diptera). Pac. Insects Monogr. 1969, 20, 261–299. [Google Scholar]
- Olafsson, E. A heron carrying louse flies to Iceland. Bliki 1985, 3, 12–14. [Google Scholar]
- Krištofík, J.; Štefan, P. K poznaniu čelade Hippoboscidae (Diptera) na Slovensku. Biológia Bratisl. 1980, 35, 137–140. [Google Scholar]
- Salem, H.M.; Yehia, N.; Al-Otaibi, S.; El-Shehawi, A.M.; Elrys, A.A.M.E.; El-Saadony, M.T.; Attia, M.M. The prevalence and intensity of external parasites in domestic pigeons (Columba livia domestica) in Egypt with special reference to the role of deltamethrin as insecticidal agent. Saudi J. Biol. Sci. 2021, 29, 1825–1831. [Google Scholar] [CrossRef] [PubMed]
- Karbowiak, G.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Wita, I.; Moskwa, B.; Werszko, J.; Bień, J.; Goździk, K.; Lachowicz, J.; Cabaj, W. The parasitic fauna of the European bison (Bison bonasus) (Linnaeus, 1758) and their impact on the conservation. Part 1. The summarising list of parasites noted. Acta Parasitol. 2014, 59, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Karbowiak, G.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Wita, I.; Moskwa, B.; Werszko, J.; Bień, J.; Goździk, K.; Lachowicz, J.; Cabaj, W. The parasitic fauna of the European bison (Bison bonasus) (Linnaeus, 1758) and their impact on the conservation. Part 2. The structure and changes over time. Acta Parasitol. 2014, 59, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Hafez, M.; Hilali, M.; Fouda, M. Biological studies on Hippobosca equina L. (Diptera: Hippoboscidae) infesting domestic animals in Egypt. Entomology 1977, 83, 426–441. [Google Scholar] [CrossRef]
- Quercia, O.; Emiliani, F.; Foschi, F.G.; Stefanini, G.F. Anaphylactic reaction after Hippobosca equina bite. Eur. Ann. Allergy Cinical Immunol. 2005, 20, 31–33. [Google Scholar]
- Matito, A.; Bartolomé-Zavala, B.; Álvarez-Twose, I.; Sánchez-Matas, I.; Escribano, L. IgE-mediated anaphylaxis to Hippobosca equina in a patient with systemic mastocytosis. Allergy 2010, 65, 1058–1072. [Google Scholar] [CrossRef]
- Sokół, R.; Gałęcki, R. Prevalence of keds on city dogs in central Poland. Med. Vet. Entomol. 2017, 31, 114–116. [Google Scholar] [CrossRef]
- Hodžić, A.; Omeragić, J.; Alić, A.; Jažić, A.; Zuko, A. Lipoptena cervi (Diptera: Hippoboscidae) in Roe deer (Capreolus capreolus). Veterinaria 2012, 61, 17–21. [Google Scholar]
- Kaunisto, S.; Korter, R.; Härkönen, L.; Härkönen, S.; Ylönen, H.; Laaksonen, S. New bedding site examination-based method to analyse deer ked (Lipoptena cervi) infection in cervids. Parasitol. Res. 2009, 104, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Robertson, B.A.; Hutto, R.L. A framework for understanding ecological traps and an evaluation of existing evidence. Ecology 2006, 87, 1075–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Härkönen, L.; Kaitala, A. Host Dynamics and Ectoparasite Life Histories of Invasive And Non-Invasive Deer Ked Populations. In Biological Invasions in Changing Ecosystems; Canning-Clode, J., Ed.; De Gruyter Open: Warsaw, Poland, 2015; pp. 212–229. [Google Scholar]
- Meier, C.M.; Bonte, D.; Kaitala, A.; Ovaskainen, O. Invasion rate of deer ked depends on spatiotemporal variation in host density. Bull. Entomol. Res. 2014, 104, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Kynkäänniemi, S.M.; Kortet, R.; Laaksonen, S. Range expansion and reproduction of the ectoparasitic deer ked (Lipoptena cervi) in its novel host, the Arctic reindeer (Rangifer tarandus tarandus), in Finland. Parasitol. Res. 2020, 119, 3113–3117. [Google Scholar] [CrossRef] [PubMed]
- Central Statistical Office. Environment, Statistical Information and Elaborations, 2014; Central Statistical Office: Warsaw, Poland, 2014. [Google Scholar]
- Central Statistical Office. Environment, Statistical Information and Elaborations, 2019; Central Statistical Office: Warsaw, Poland, 2019. [Google Scholar]
- Mysterud, A.; Madslien, K.; Herland, A.; Viljugrein, H.; Ytrehus, B. Phenology of deer ked (Lipoptena cervi) host-seeking flight activity and its relationship with prevailing autumn weather. Parasites Vectors 2016, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaunisto, S.; Ylonen, H.; Kortet, R. Passive sinking into the snow as possible survival strategy during the off-host stage in an insect ectoparasite. Folia Parasitol. 2015, 62, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieminen, P.; Käkelä, R.; Paakkonen, T.; Halonen, T.; Mustonen, A.M. Fatty acid modifications during autumnal cold-hardening in an obligatory ectoparasite, the deer ked (Lipoptena cervi). J. Insect Physiol. 2013, 59, 631–637. [Google Scholar] [CrossRef]
- Härkönen, L.; Hurme, E.; Kaitala, A. Unexpected seasonal variation in offspring size and performance in a viviparous ectoparasite. Parasitology 2013, 140, 229–236. [Google Scholar] [CrossRef]
- El-Sayed, A.; Kamel, M. Climatic changes and their role in emergence and re-emergence of diseases. Environ. Sci. Pollut. Res. Int. 2020, 27, 22336–22352. [Google Scholar] [CrossRef]
- Vidal, C.; Armisén, M.; Bartolomé, B.; Rodriguez, V.; Luna, I. Anaphylaxis to Hippobosca equina (louse fly). Eur. Ann. Allergy Cinical Immunol. 2007, 99, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Decastello, A.; Farkas, R. Anaphylactic reaction following forest fly (Hippobosca equina) bite: A human case. Clin. Exp. Med. J. 2010, 4, 193–198. [Google Scholar] [CrossRef]
- Arafa, M.I.; Hamouda, S.M.; Rateb, H.Z.; Abdel-Hafeez, M.M.; Aamer, A.A. Oedematous Skin Disease (OSD) transmission among buffaloes. Global Journal of Medical Research: G. Vet. Sci. Vet. Med. 2019, 19, 15–19. [Google Scholar]
- Selim, S.A. Review Oedematous Skin Disease of Buffalo in Egypt. J. Vet. Med. Ser. B 2001, 48, 241–258. [Google Scholar] [CrossRef]
- Moussa, I.M.; Ali, M.S.; Hessain, A.M.; Kabli, S.A.; Hemeg, H.A.; Selim, S.A. Vaccination against Corynebacterium pseudotuberculosis infections controlling caseous lymphadenitis (CLA) and oedematous skin disease. Saudi J. Biol. Sci. 2016, 23, 718–723. [Google Scholar] [CrossRef]
- Ghoneim, M.A.; Mousa, W.M.; Ibrahim, A.K.; Amin, A.S.; Khafagy, A.; Selim, S.A. Role of Hippobosca equina as a transmitter of C. pseudotuberculosis among buffaloes by PCR and Dot blot hydridization. Egypt. Vet. Med. Assoc. 2001, 61, 165–176. [Google Scholar]
- Syame, S.M.; EL-Hewairy, H.M.; Selim, S.A. Protection of Buffaloes Against Oedematous Skin Disease by Recombinent-bacterin and Toxoid-bacterin Vaccines. Glob. Vet. 2008, 2, 151–156. [Google Scholar]
- Benelli, G.; Pavela, R. Repellence of essential oils and selected compounds against ticks—A systematic review. Acta Trop. 2018, 179, 47–54. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R. Beyond mosquitoes—Essential oil toxicity and repellency against bloodsucking insects. Ind. Crops Prod. 2018, 117, 382–392. [Google Scholar] [CrossRef]
- Lee, M.Y. Essential Oils as Repellents against Arthropods. BioMed Res. Int. 2018, 2018, 6860271. [Google Scholar] [CrossRef] [Green Version]
- Khanikor, B.; Parida, P.; Yadav, R.N.S.; Bora, D. Comparative mode of action of some terpene compounds against octopamine receptor and acetyl cholinesterase of mosquito and human system by the help of homology modeling and Docking studies. J. Appl. Pharm. Sci. 2013, 3, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E. Repellent activity of essential oils: A review. Bioresour. Technol. 2010, 101, 372–378. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0960852409009468?via%3Dihub (accessed on 13 October 2022). [CrossRef] [PubMed]
- Khater, H.F.; Ramadan, M.Y.; El-Madawy, R.S. Lousicidal, ovicidal and repellent efficacy of some essential oils against lice and flies infesting water buffaloes in Egypt. Vet. Parasitol. 2009, 164, 257–266. [Google Scholar] [CrossRef]
- Ramsey, J.T.; Shropshire, B.C.; Nagy, T.R.; Chambers, K.D.; Li, Y.; Korach, K.S. Essential Oils and Health. Yale J. Biol. Med. 2020, 93, 291–305. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Observations of Hippobosca equina L. 1758 | ||
|---|---|---|
| Zone | Country | Data Source |
| Palearctic | Azores, Portugal | [29] |
| Spain | [30] | |
| France | [31,32] | |
| Great Britain | [33,34,35] | |
| Belgium | [36] | |
| The Netherlands | [37] | |
| Germany | [38,39,40] | |
| Poland | [6,19,41], own observations 2021, 2022 | |
| Norway | [42] | |
| Sweden | [43] | |
| Finland | [44] | |
| Estonia | [45] | |
| Latvia | [46] | |
| Lithuania | [47] | |
| Hungary | [39] | |
| Croatia | [39,48] | |
| North Macedonia | [39] | |
| Bosnia and Herzegovina | [39] | |
| Continental Italy | [10] | |
| Sardinia, Italy | [49] | |
| Slovakia | [14,39] | |
| Belarus | [50] | |
| Romania | [51] | |
| Bulgaria | [52] | |
| The Cyclades, Greece | [53] | |
| Azerbaijan | [54] | |
| Armenia | [54] | |
| Iran | [54,55] | |
| Algeria | [9] | |
| Tunisia | [56] | |
| Egypt | [57] | |
| Israel | [24] | |
| Jeju Island, Republic of Korea | [58,59] | |
| West Oriental | India | [60,61] |
| Afrotropical | Nigeria | [28] |
| Observations of Hippobosca equina L. 1758 | ||
|---|---|---|
| Name of Database | Country | Month and Year |
| GBIF 1,* | Palearctic zone | |
| Continental Portugal | 01.2015. x 164; 05.2018. x 2; 06.2018. x 3; 10.2018. x 2; 07.2020; 04.2021.; 04.2022.; 05.2022., 06.2022. | |
| Slovenia | 08.2015. x 2 | |
| Austria | 06.2017.; 05.2018.; 03.2020., 05.2022. | |
| Sweden | 10.2003.; 06.2012.; 06.2014.; 07.2015. x 4; 05.2016.; 07.2016.; 08.2017.; 06.2020.; 07.2020. x 3, 05.2022.; 06.2022. | |
| Continental Greece | 07.2012.; 08.2018.; 07.2020. | |
| The Republic of Turkey | 03.2022. | |
| Kazakhstan | 09.2020. | |
| Oriental zone | ||
| The Philippines | 05.1911. ** x 4 | |
| Australasian zone | ||
| New Caledonia | no data ** x 4 | |
| Afrotropical zone | ||
| Mozambique | 04.1928.; 01.1930.; 07.1968. | |
| iNaturalist 2,*** | Tenerife, Spain | 09.2016. |
| Portugal | 06.2020. x 2; 04.2022.; 05.2022.; 06.2022. | |
| Austria | 05.2018; 03.2020.; 05.2022. x 2 | |
| The Czech Republic | 07.2016. | |
| Corsica, France | 09.2012.; 07.2020. | |
| Continental Greece | 07.2020. | |
| Lesbos | 08.2018. | |
| North Aegean, Greece | 08.2022.; 09.2022. | |
| Romania | 09.2019. | |
| The Republic of Turkey | 03.2022. | |
| Ukraine | 07.2020. | |
| Kazakhstan | 09.2020. | |
| Fauna Europaea 3 | The Canary Islands | no data |
| Ireland | ||
| Norway | ||
| Denmark | ||
| Switzerland | ||
| Sicily, Italy | ||
| The Czech Republic | ||
| Crete, Greece | ||
| The Dodecanese Islands, Greece | ||
| PESI 4 | The Balearic Islands, SpainSwitzerland The Czech republic | no data |
| Denmark | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maślanko, W.; Szwaj, E.; Gazda, M.; Bartosik, K. Hippobosca equina L. (Hippoboscidae: Hippobosca)—An Old Enemy as an Emerging Threat in the Palearctic Zone. Int. J. Environ. Res. Public Health 2022, 19, 16978. https://doi.org/10.3390/ijerph192416978
Maślanko W, Szwaj E, Gazda M, Bartosik K. Hippobosca equina L. (Hippoboscidae: Hippobosca)—An Old Enemy as an Emerging Threat in the Palearctic Zone. International Journal of Environmental Research and Public Health. 2022; 19(24):16978. https://doi.org/10.3390/ijerph192416978
Chicago/Turabian StyleMaślanko, Weronika, Ewelina Szwaj, Michał Gazda, and Katarzyna Bartosik. 2022. "Hippobosca equina L. (Hippoboscidae: Hippobosca)—An Old Enemy as an Emerging Threat in the Palearctic Zone" International Journal of Environmental Research and Public Health 19, no. 24: 16978. https://doi.org/10.3390/ijerph192416978