
Somatic Embryogenesis of Representative Medicinal Trees in South America—Current Status
 , , , ,
, , , ,
Abstract
:1. Introduction
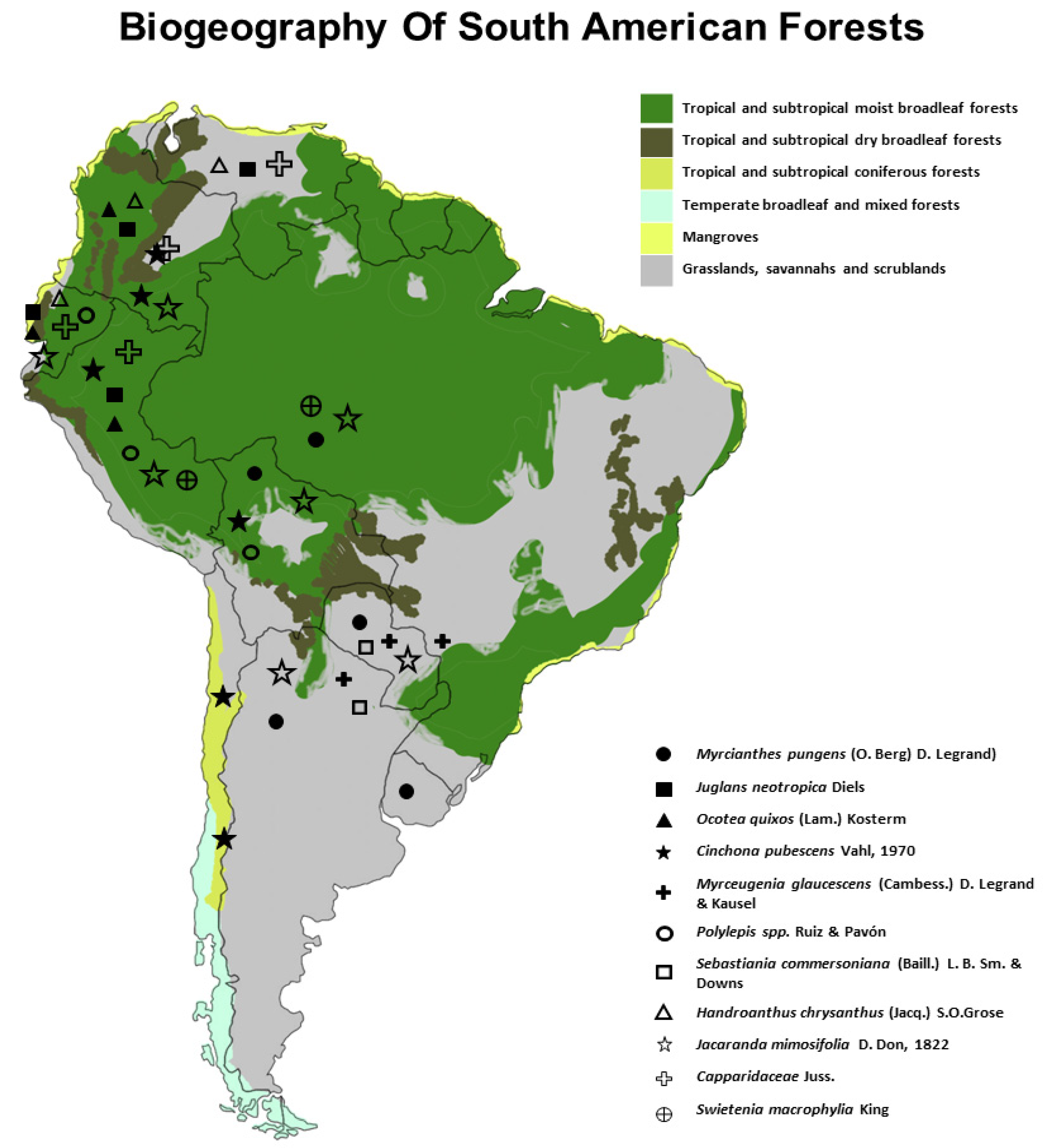
2. The Biodiversity of South America Trees and Conservation Status
3. Overview of Somatic Embryogenesis in Trees
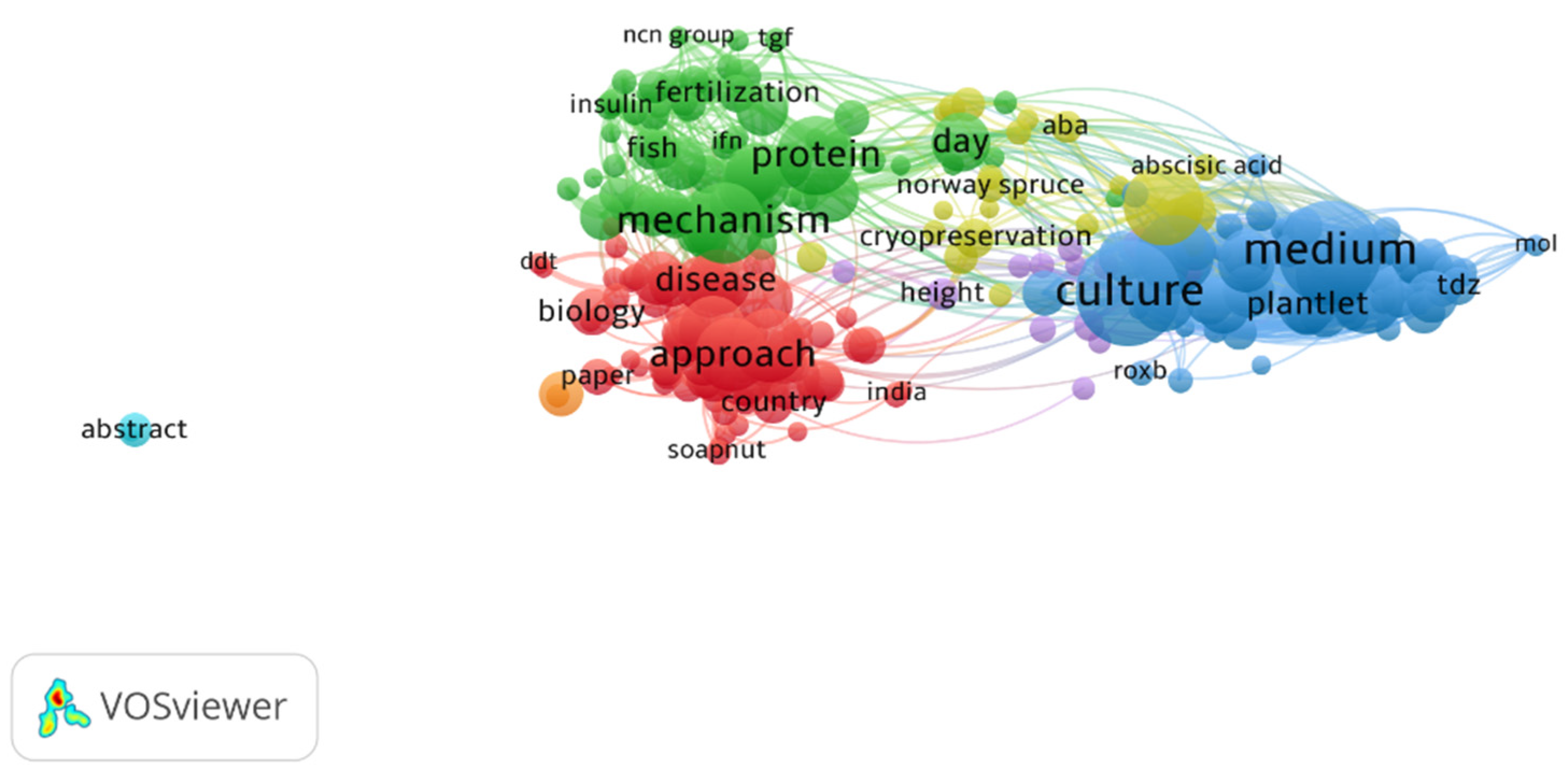
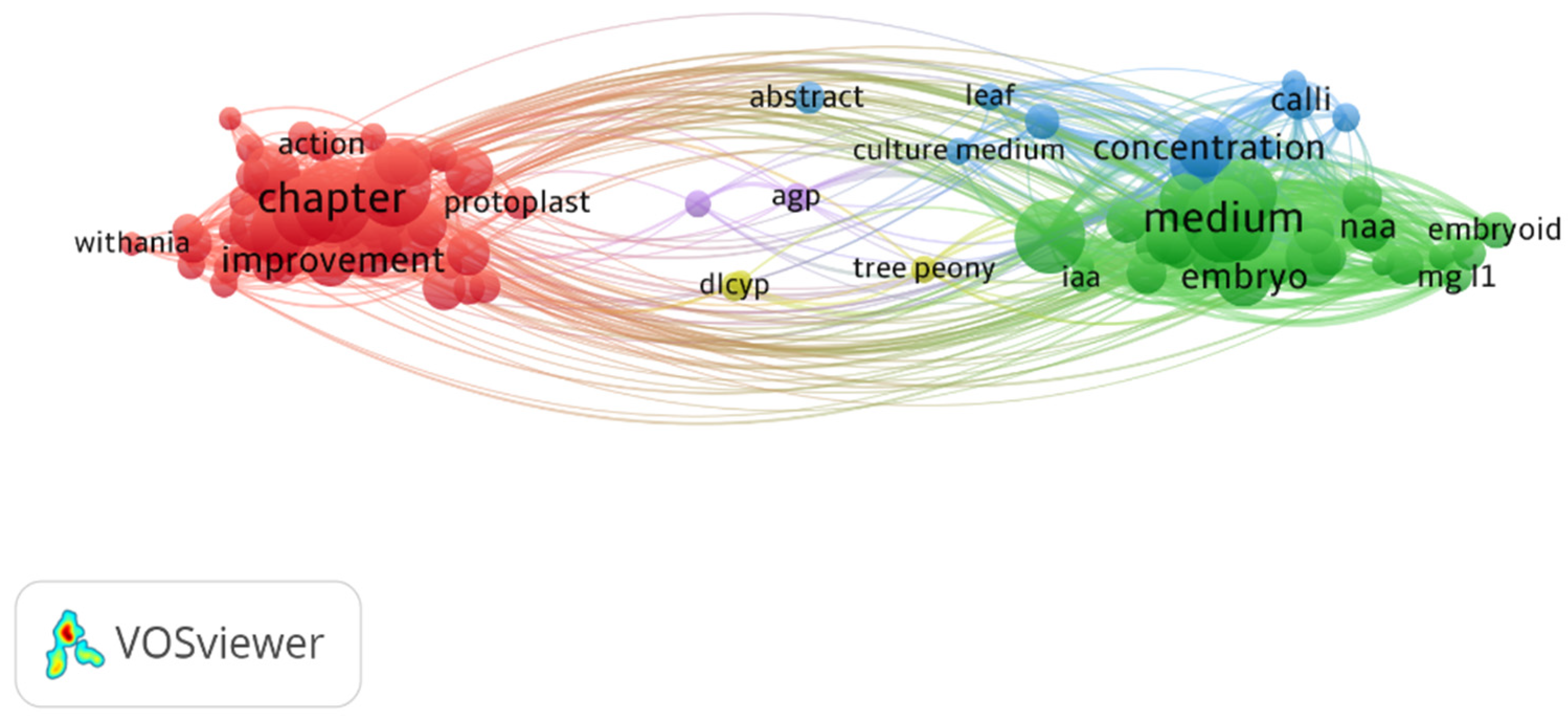
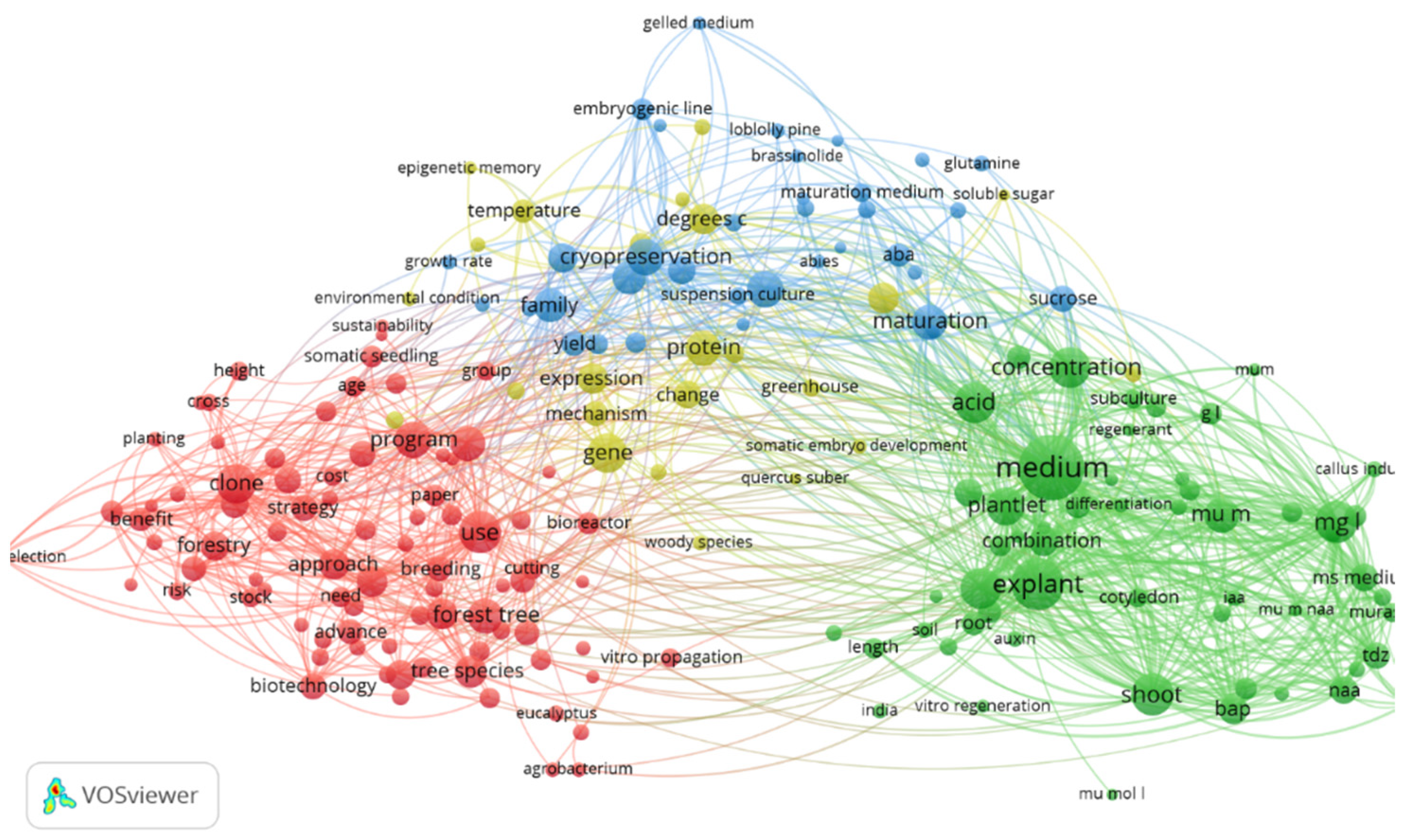
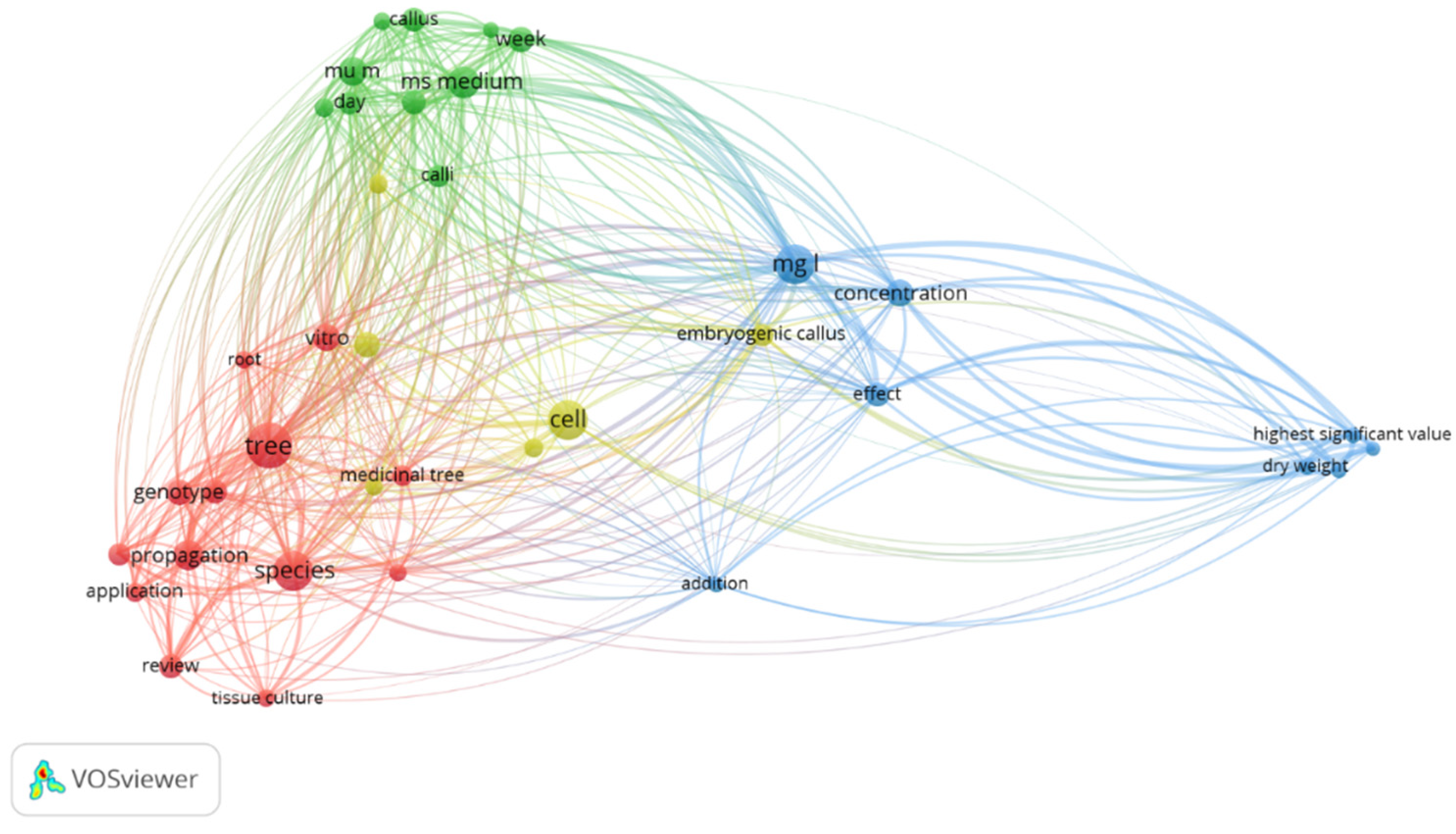
3.1. VOS Viewer for Screening and Bibliometric Analysis
3.2. Somatic Embryogenesis and Secondary Metabolites
4. Tree Species with Medicinal Properties in South America
5. Propagation of Medicinal Trees in South America
- ○
- Somatic embryogenesis induction step:
- Plant material—should be carefully collected, transported, and prepared for introduction in in vitro culture;
- Sterilization of plant material—is one of the most important steps assuring an aseptic but viable material for in vitro cultures;
- Media variant—should be added with proper plant growth hormones that are able to induce differentiation and dedifferentiation of the somatic cells;
- Control of the environmental conditions.
- ○
- Somatic embryo recovery step—for the development of the somatic embryos, these should be transferred on a media variant with a lower concentration of plant growth factors than in the first step.
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviation
References
- BGCI. State of the World’s Trees; BGCI: Richmond, UK, 2021. [Google Scholar]
- Alves, R.R.; Rosa, I.M. Biodiversity, traditional medicine and public health: Where do they meet? J. Ethnobiol. Ethnomed. 2007, 3, 14. [Google Scholar] [CrossRef]
- Hazubska-Przybył, T.; Wawrzyniak, M.K.; Kijowska-Oberc, J.; Staszak, A.M.; Ratajczak, E. Somatic Embryogenesis of Norway Spruce and Scots Pine: Possibility of Application in Modern Forestry. Forests 2022, 13, 155. [Google Scholar] [CrossRef]
- Salaün, C.; Lepiniec, L.; Dubreucq, B. Genetic and Molecular Control of Somatic Embryogenesis. Plants 2021, 10, 1467. [Google Scholar] [CrossRef]
- Elhiti, M.; Stasolla, C. Somatic embryogenesis: The molecular network regulating embryo formation. In Somatic Embryogenesis in 393 Ornamentals and Its Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 217–229. [Google Scholar]
- Bogdanović, M.D.; Cuković, K.B.; Subotić, A.R.; Dragićević, M.B.; Simonović, A.D.; Filipović, B.K.; Todorović, S.I. Secondary Somatic Embryogenesis in Centaurium erythraea Rafn. Plants 2021, 10, 199. [Google Scholar] [CrossRef]
- Kriebel, H.B. Introduction. In Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Newton, R.J., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 1995; Volume 1, pp. 1–8. [Google Scholar]
- Gatti, R.C.; Reich, P.B.; Gamarra, J.G.P.; Crowther, T.; Hui, C.; Morera, A.; Bastin, J.F.; de-Miguel, S.; Nabuurs, G.J.; Svenning, J.C.; et al. The number of tree species on Earth. Proc. Natl. Acad. Sci. USA 2022, 119, e2115329119. [Google Scholar] [CrossRef]
- Sol-Sánchez, A.; Hernández-Melchor, G.; Hernández-Hernández, M. Desarrollo bioeconómico y manglares en América Latina. Rev. Iberoam. Bioeconomía Cambio Climático 2022, 8, 2007–2017. [Google Scholar] [CrossRef]
- UNEP-WCMC. The State of Biodiversity in Latin America and the Caribbean; UNEP-WCMC: Cambridge, UK, 2016. [Google Scholar]
- FAO. State of the World’s Forests; FAO: Rome, Italy, 1997. [Google Scholar]
- Villagrán, C. Biogeografía de los bosques subtropical-templados del sur de sudamérica. Hipótesis Históricas (Biogeography of the subtropical-temperate forests of southern South America. Historical Hypotheses). Magallania 2018, 46, 27–48. (In Spanish) [Google Scholar] [CrossRef]
- Silva de Miranda, P.L.; Dexter, K.G.; Swaine, M.D.; de Oliveira-Filho, A.T.; Hardy, O.J.; Fayolle, A. Dissecting the difference in tree species richness between Africa and South America. Proc. Natl. Acad. Sci. USA 2022, 119, e2112336119. [Google Scholar] [CrossRef]
- Trajano, E. Biodiversity in South America. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; Chapter 20; pp. 177–186. [Google Scholar] [CrossRef]
- Geraque, E. Biodiversity of Tropical Trees is Highest in South America. 2022. Available online: https://revistapesquisa.fapesp.br/en/biodiversity-of-tropical-trees-is-highest-in-south-america/ (accessed on 3 March 2023).
- Zalles, V.; Hansen, M.C.; Potapov, P.V.; Parker, D.; Stehman, S.V.; Pickens, A.H.; Parente, L.L.; Ferreira, L.G.; Song, X.P.; Hernandez-Serna, A.; et al. Rapid expansion of human impact on natural land in South America since 1985. Sci. Adv. 2021, 7, eabg1620. [Google Scholar] [CrossRef]
- Silva-Souza, K.J.P.; Pivato, M.G.; Silva, V.C.; Haidar, R.F.; Souza, A.F. New patterns of the tree beta diversity and its determinants in the largest savanna and wetland biomes of South America. Plant Div. 2023, 45, 369–384. [Google Scholar] [CrossRef]
- Raven, P.; Gereau, R.; Ulloa, C.; Phillipson, P.; Chatelain, C.; Jenkins, C. The distribution of biodiversity richness in the tropics. Sci. Adv. 2020, 6, eabc6228. [Google Scholar] [CrossRef] [PubMed]
- FAO: El Niño 2018-19. Available online: https://reliefweb.int/report/world/fao-el-ni-o-2018-19 (accessed on 23 April 2023).
- Beuchle, R.; Achard, F.; Bourgoin, C.; Vancutsem, C.; Eva, H.; Follador, M. Deforestation and Forest Degradation in the Amazon—Status and Trends Up to Year 2020; EUR 30727 EN; Publications Office of the European Union: Luxembourg, 2021; ISBN 978-92-76-38352-9. JRC124955. [Google Scholar] [CrossRef]
- Oldfield, S.; Lusty, C.; MacKinven, A. The World List of Threatened Trees; World Conservation Press, WCMC: Cambridge, UK, 1998. [Google Scholar]
- Available online: https://www.worldwildlife.org/industries/responsible-forestry (accessed on 3 March 2023).
- Guan, Y.; Li, S.G.; Fan, X.F.; Su, Z.H. Application of Somatic Embryogenesis in Woody Plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, N.J.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Rabechault, H.; Ahée, J.; Guenin, G. Colonies cellulaires et formes embryoides obtenues in vitro a partir de cultures d’embryons 416 de Palmier a huile (Elaeis guineensis Jacq. var. dura Becc.). Acad. Sci. Compt. Rend. Ser. D 1970, 270, 3067–3070. [Google Scholar]
- Ross, S.; Arriaga, M.E.; Pechi, E. Establecimiento in vitro de Yerba mate (Ilex paraguariensis A. St.-Hil.) nativa de Uruguay. Agrociencia 2017, 21, 15–23. [Google Scholar] [CrossRef]
- Álvarez, M. Propagación In Vitro de Guayusa (Ilex guayusa) a Partir de Segmentos Nodales Proyecto (In Vitro Propagation of Guayusa (Ilex guayusa) from Nodal Segments Project). 2018. Available online: https://repositorio.usfq.edu.ec/bitstream/23000/7968/1/141266.pdf (accessed on 3 March 2023). (In Spanish).
- Rangel, A.M.A.; Salamanca, E.J.P.; Restrepo, D.C. Evaluación de medios de cultivo para la producción in vitro de Annona muricata mediante la técnica de microinjertación seriada (Evaluation of culture media for the in vitro production of Annona muricata using the serial micrografting technique). Acta Agronómica 2011, 60, 140–146. (In Spanish) [Google Scholar]
- Lima, N.R.; Moreno, J.S.; Eras, V.H.; Minchala, J.P.; González Zaruma, D.; Yaguana Arévalo, M.; Valarezo Ortega, C. Propagación in vitro de Cinchona officinalis L. a partir de semillas in vitro propagation of Cinchona officinalis L. from seeds Nelson. Rev. Investig. Altoandinas 2018, 20, 169–178. [Google Scholar] [CrossRef]
- Kouassi, M.K.; Kahia, J.; Kouame, C.N.; Tahi, M.G.; Koffi, E.K. Comparing the Effect of Plant Growth Regulators on Callus and Somatic Embryogenesis Induction in Four Elite Theobroma cacao L. Genotypes. HortScience Horts 2017, 52, 142–145. [Google Scholar] [CrossRef]
- Peña, D.; Rocano, M.; Salazar, J.; Torres, C. Inducción de la brotación in vitro de microplántulas de Nogal (Juglans neotropica) tratadas con Thidiazuron (TDZ) y 6-Bencilaminnopurina (BAP). MASKANA 2014, 5, 81–85. [Google Scholar] [CrossRef]
- Quintero-García, O.D.; Jaramillo-Villegas, S. Rescate y germinación in vitro de embriones inmaduros de cedro negro (Juglans neotropica Diels) (Rescue and in vitro germination of immature embryos of black cedar (Juglans neotropica Diels)). Acta Agronómica 2012, 61, 52–60. (In Spanish) [Google Scholar]
- Joseph, N.; Siril, E.A.; Nair, G.M. An efficient in vitro propagation methodology for Annatto (Bixa orellana L.). Physiol. Mol. Biol. Plants 2011, 17, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Driver, J.A.; Kuniyuki, A.H. In Vitro Propagation of Paradox Walnut Rootstock. HortScience 1984, 19, 507–509. [Google Scholar] [CrossRef]
- Fajinmi, O.O.; Olarewaju, O.O.; Van Staden, J. Propagation of Medicinal Plants for Sustainable Livelihoods, Economic Development, and Biodiversity Conservation in South Africa. Plants 2023, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The Influence of Environmental Conditions on Secondary Metabolites in Medicinal Plants: A Literature Review. Chem. Biodiv. 2021, 18, e2100345. [Google Scholar] [CrossRef]
- Isah, T. Induction of somatic embryogenesis in woody plants. Acta Physiol. Plant. 2016, 38, 118. [Google Scholar] [CrossRef]
- Rahmat, E.; Kang, Y. Adventitious root culture for secondary metabolite production in medicinal plants: A Review. J. Plant Biotechnol. 2019, 46, 143–157. [Google Scholar] [CrossRef]
- Vázquez-Flota, F.A.; Monforte-González, M.; de Lourdes Miranda-Ham, M. Application of Somatic Embryogenesis to Secondary Metabolite-Producing Plants; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 455–469. [Google Scholar] [CrossRef]
- Zhai, X.; Jia, M.; Chen, L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.-P. The regulatory mechanism of fungal elicitor-induced secondary metabolite biosynthesis in medical plants. Crit. Rev. Microbiol. 2017, 43, 238–261. [Google Scholar] [CrossRef]
- Erazo-Garcia, M.P.; Sotelo-Proaño, A.R.; Ramirez-Villacis, D.X.; Garcés-Carrera, S.; Leon-Reyes, A. Methyl jasmonate-induced resistance to Delia platura (Diptera: Anthomyiidae) in Lupinus mutabilis. Pest Manag. Sci. 2021, 77, 5382–5395. [Google Scholar] [CrossRef]
- Nora, C.D.; Danelli, D.; Souza, L.F.; Rios, A.d.O.; de Jong, E.V.; Flôres, S.H. Protective effect of guabiju (Myrcianthes pungens (O. Berg) D. Legrand) and red guava (Psidium cattleyanum Sabine) against cisplatin-induced hypercholesterolemia in rats. Braz. J. Pharm. Sci. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Cardoso, B.K.; Oliveira, H.L.M.; Zonta Melo, U.; Mariano Fernandez, C.M.; Araújo Almeida Campo, C.F.; de Gonçalves, J.E.; Laverde, A.; Barion Romagnolo, M.; Linde, G.A.; Cristiani Gazim, Z. Antioxidant activity of α and β-amyrin isolated from Myrcianthes pungens leaves. Nat. Prod. Res. 2020, 34, 1777–1781. [Google Scholar] [CrossRef] [PubMed]
- Renan Almeida, J.; Marko de Oliveira, H.L.; Campos Bortolucci, W.; De Araújo Almeida Campo, C.F.; Iecher Faria, M.G.; Gonçalves, J.E.; Barros Colauto, N.; Cristiani Gazim, Z.; Linde, G.A. Antioxidant and antibacterial activity of Myrcianthes pungens leaf essential oil. Bol. Latinoam. Caribe Plantas Med. Aromat. 2021, 20, 147–161. [Google Scholar] [CrossRef]
- Tischer, B.; Pangloli, P.; Nieto-Veloza, A.; Reeder, M.; Dia, V.P. Bioactive compounds, antioxidant capacity and anti-inflammatory activity of native fruits from Brazil. PLoS ONE 2023, 18, e0285625. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, E.T.; Rojas, I.C.R. State of the art, propagation and conservation of Juglans neotropica diels., in andean zones. Madera Bosques 2018, 24, e2411560. [Google Scholar] [CrossRef]
- Hurtado, P. Evaluación de la Actividad Gastroprotectora del Extracto Hidroalcohólico de las Hojas de Juglans neotropica Diels «Nogal Peruano» [Título de Químico Farmacéutico, Universidad Nacional Mayor de San Marcos] (Evaluation of the Gastroprotective Activity of the Hydroalcoholic Extract of the Leaves of Juglans neotropica Diels «Peruvian Walnut» [Title of Pharmaceutical Chemist, Universidad Nacional Mayor de San Marcos]. 2014. Available online: http://cybertesis.unmsm.edu.pe/bitstream/handle/20.500.12672/3748 (accessed on 3 March 2023). (In Spanish).
- Montealegre, C. Etnobotanica preliminar del Espíngo (Ocotea quixos (Lam.) Kosterm.) en la medicina tradicional indigena inga, pruebas fitoquimicas y evaluacion de la actividad antimicrobiana (Preliminary ethnobotany of Espíngo (Ocotea quixos (Lam.) Kosterm.) in traditional indigenous Inga medicine, phytochemical tests and evaluation of antimicrobial activity). Los Flavonoides Propiedades Antioxidantes 2011, 17, 271–278. (In Spanish) [Google Scholar]
- Villamizar, V.E.M. Metabolitos secundarios con actividad biológica (farmacológica), aplicación etnobotánica; y fitoquímica de algunas especies de los géneros: Ocotea, Cryptocarya, Litsea, Caryodaphnosis, Machilus Y Actinodaphne (Lauraceae) (Secondary metabolites with biological (pharmacological) activity, ethnobotanical application; and phytochemistry of some species of the genera: Ocotea, Cryptocarya, Litsea, Caryodaphnosis, Machilus and Actinodaphne (Lauraceae)). Duazary 2010, 7, 152–170. (In Spanish) [Google Scholar] [CrossRef]
- Roque, R.; Barrutia, I.; Marin, T. Germinación de semillas de Cinchona officinalis L. en tres tipos de suelos de Cajamarca, Perú. (Germination of Cinchona officinalis L. seeds in three types of soils from Cajamarca, Peru). Rev. Cuba. De Cienc. For. 2020, 8, 75–87. (In Spanish) [Google Scholar]
- Albán-Castillo, J.; Chilquillo, E.; Melchor-Castro, B.; Arakaki, M.; León, B.; Suni, M.; Cinchona, L. Árbol de la Quina (Quina Tree). Rev. Peru. Biol. 2020, 27, 423–426. [Google Scholar] [CrossRef]
- Remuzgo Foronda, J.R.; Alvarez Melo, J.B.; Sales Dávila, F.; Valdivieso Arenas, G. Caracterización Taxonómica y Fitoquímica de Cinchona pubescens y Ladenbergia oblongifolia en el Ámbito del Valle Alto Huallaga—Región Huánuco (Taxonomic and Phytochemical Characterization of Cinchona pubescens and Ladenbergia oblongifolia in the Alto Huallaga Valley Area—Huánuco Region). REBIOL 2020, 42, 242–255. (In Spanish) [Google Scholar] [CrossRef]
- Waizel-Bucay, J.; Waizel-Haiat, S. Las plantas con principios amargos y su uso medicinal. ¿Un futuro dulce? (Plants with bitter principles and their medicinal use. A sweet future?) An. Orl. Mex. 2019, 64, 202–228. (In Spanish) [Google Scholar]
- de Araújo Rodrigues, P.; de Morais, S.M.; Aguiar, L.A.; Vila-Nova, N.S.; Benjamin, S.R. Effect of Byrsonima sericea DC. leaf extracts on mice gastrointestinal tract. Toxicol. Rep. 2019, 6, 1182–1187. [Google Scholar] [CrossRef] [PubMed]
- Godinho, W.M.; Farnezi, M.M.; Pereira, I.M.; Gregório, L.E.; Grael, C.F. Volatile constituents from leaves of Blepharocalyx salicifolius (Kunth) O. Berg (Myrtaceae). Boletín Latinoam. Caribe Plantas Med. Aromáticas 2014, 13, 249–253. [Google Scholar]
- Canales Gutiérrez, Á.; Huarasa Vilca, Y.R. Poder germinativo de Polylepis incana con aplicación de diferentes tratamientos de agua (Germinative power of Polylepis incana with application of different water treatments). Rev. Cuba. De Cienc. For. 2020, 8, 495–506. (In Spanish) [Google Scholar]
- González, J.A.; Gallardo, M.G.; Boero, C.; Liberman Cruz, M.; Prado, F.E. Altitudinal and seasonal variation of protective and photosynthetic pigments in leaves of the world’s highest elevation trees Polylepis tarapacana (Rosaceae). Acta Oecologica 2007, 32, 36–41. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, D. Secondary metabolites as a survival strategy in plants of high mountain habitats. Bol. Latinoam. Caribe Plantas Med. Aromat. 2019, 18, 444–458. [Google Scholar] [CrossRef]
- Hnatyszyn, O.; Juárez, S.; Ouviña, A.; Martino, V.; Zacchino, S.; Ferraro, G. Phytochemical analysis and antifungal evaluation of Sebastiania commersoniana extracts. Pharm. Biol. 2007, 45, 404–406. [Google Scholar] [CrossRef]
- Pérez-González, M.Z.; Macías-Rubalcava, M.L.; Hernández-Ortega, S.; Siordia-Reyes, A.G.; Jiménez-Arellanes, M.A. Additional compounds and the therapeutic potential of Cnidoscolus chayamansa (McVaugh) against hepatotoxicity induced by antitubercular drugs. Biomed. Pharmacother. 2019, 117, 109140. [Google Scholar] [CrossRef]
- Tovar, V. Actividad Antibacteriana y Antioxidante del Aceite Esencial Extraído del Tronco y Corteza de la Especie Handroanthus chrysanthus (Guayacán). [Universidad Central del Ecuador] (Antibacterial and Antioxidant Activity of the Essential Oil Extracted from the Trunk and Bark of the Handroanthus chrysanthus (Guayacán) Species. [Central University of Ecuador]). 2018. Available online: https://www.dspace.uce.edu.ec/bitstream/25000/15263/1/T-UCE-0008-QF057-2018.pdf (accessed on 3 March 2023). (In Spanish).
- Arbeláez, J.S.M. Metabolitos secundarios en el guayacán amarillo y en el guayacán rosado (Secondary metabolites in yellow guayacán and pink guayacán). Dialnet 2011, 1, 297–301. (In Spanish) [Google Scholar]
- Ramírez-Pisco, R.; Velázquez-Pomar, D.; Acosta-Baena, E. Efecto de la aplicación de biosolidos en el crecimiento de Jacaranda mimosifolia (Gualanday) y en las condiciones físicas y químicas de un suelo degradado. Facultad Nacional de Agronomía Medellín. 2011. Available online: https://www.revistas.unal.edu.co/index.php/refame/article/view/24393 (accessed on 3 March 2023).
- Játiva, C. Separación y Posible Identificación de Metabolitos Secundarios de la Jacaranda (Jacaranda mimosifolia) con Fines de Aporte a una Técnica de Análisis Químico. 2013. Available online: https://dspace.espoch.edu.ec/handle/123456789/2560 (accessed on 3 March 2023).
- Coronel, E.; Alvarenga, N.; Caballero, S.; Mereles, L. Potencial nutritivo y medicinal de recursos fitogenéticos del género Capparis L. y especies relacionadas en el Gran Chaco (Nutritional and medicinal potential of plant genetic resources of the genus Capparis L. and related species in the Gran Chaco). Rojasiana 2020, 19, 21–42. (In Spanish) [Google Scholar]
- Torres, A.; Salinas, Y.; Valle, S.; Soto, R.M.; Alia, I. Proantocianidinas y actividad enzimática en fruto de mamey zapote (Pouteria sapota) durante su maduración (Proanthocyanidins and enzymatic activity in mamey sapote fruit (Pouteria sapota) during ripening). Rev. Bio Cienc. 2019, 6, e565. (In Spanish) [Google Scholar] [CrossRef]
- Lating, M. Forest Tree Species with Medical Uses. 2002. Available online: https://erdb.denr.gov.ph/wp-content/uploads/2015/06/denr_v11.pdf (accessed on 3 March 2023).
- Telrandhe, U.; Kosalge, S.; Parihar, D.; Sharma, D.; Lade, S.N. Phytochemistry and Pharmacological Activities of Swietenia macrophylla King (Meliaceae). Sch. Acad. J. Pharm. 2022, 11, 6–12. [Google Scholar] [CrossRef]
- Dos Santos De Souza, L.; Avrella, E.D.; De Campos, S.S.; Fior, C.S.; Schwarz, S.F. Clonagem de espécime adulto de Myrcianthes pungens (Berg) Legrand através da estaquia (Cloning of adult Myrcianthes pungens (Berg) Legrand specimen by cutting). Iheringia-Ser. Bot. 2018, 73, 336–341. [Google Scholar] [CrossRef]
- Rocano, C.; Mélida, N.; Peña, T.D.F. Evaluación de los sistemas de cultivo semisólido y BIT en la multiplicación in vitro de Juglans neotrópica (Evaluation of semi-solid culture systems and BIT in the in vitro multiplication of Juglans neotropica). Maskana 2017, 8, 103–109. (In Spanish) [Google Scholar] [CrossRef]
- Gonzales, P. (2021, 11 febrero). Un Proyecto con Visión Sustentable (A Project with a Sustainable Vision). Revistalideres.com. (A Project with a Sustainable Vision. Revistalideres.com). Available online: https://www.revistalideres.ec/lideres/proyecto-vision-sustentable-amazonia-lideres.html (accessed on 15 June 2023). (In Spanish).
- Narciso, P.L.M. Tesis Para Optar el Título de Ingeniera Forestal. Universidad Nacional Agraria la Molina. 2021. Available online: https://repositorio.lamolina.edu.pe/bitstream/handle/20.500.12996/4723/monzon-narciso-paolalicet.pdf (accessed on 22 June 2023).
- Fernandez-Zarate, F.H.; Huaccha-Castillo, A.E.; Quiñones-Huatangari, L.; Vaca-Marquina, S.P.; Sanchez-Santillan, T.; Morales-Rojas, E.; Seminario-Cunya, A.; Guelac-Santillan, M.; Barturén-Vega, L.M.; Coronel-Bustamante, D. Effect of arbuscular mycorrhiza on germination and initial growth of Cinchona officinalis L. (Rubiaceae). For. Sci. Technol. 2022, 18, 182–189. [Google Scholar] [CrossRef]
- Espinosa, C.I.; Ríos, G. Patrones de crecimiento de Cinchona officinalis in vitro y ex vitro; respuestas de plántulas micropropagadas y de semillas (Growth patterns of Cinchona officinalis in vitro and ex vitro; responses of micropropagated seedlings and seeds). Rev. Ecuat. Med. Cienc. Biol. 2014, 35, 73–82. (In Spanish) [Google Scholar]
- Moreno Serrano, J.A.; Pérez Ruíz, C.; Moreno Fierro, I.M.; Moreno Fierro, J. Effect of culture medium on morphogenic processes in vitro in Cinchona officinalis L. Rev. FCA UNCuyo 2019, 51, 55–68. [Google Scholar]
- Eras, V.G.; Yaguana, M.P.; Paredes, D.M. Balance hormonal para la fase de brotación y enraizamiento in vitro de explantes de Cinchona Officinalis L., provenientes de relictos boscosos de la provincia de Loja (Hormonal balance for the in vitro budding and rooting phase of Cinchona Officinalis L. explants, from forest relics in the province of Loja). Bosques Latid. Cero 2019, 9, 58–68. (In Spanish) [Google Scholar]
- Vega, C.; Quezada, J.; Rocabado, P.; Villegas, G.; Bermejo, J.C. Assessment of Seed Quality and Germination Response in the Species of the Genus Polylepis. In Seed Dormancy and Germination; Jimenez-Lopez, J.C., Ed.; IntechOpen: Rijeka, Croatia, 2020; Available online: https://books.google.com.ec/books?id=KXL8DwAAQBAJ&hl=es&source=gbs_navlinks_s (accessed on 22 June 2023).
- Durand, B.F.; Sevillano, S. Experiences of Community Rehabilitation with Queñual (Polylepis sp.) in the Department of Ancash, Peru. 2017. Available online: https://www.researchgate.net/publication/320274684_Experiences_of_community_rehabilitation_with_quenual_Polylepis_sp_in_the_Department_of_Ancash_Peru (accessed on 22 June 2023).
- Huarhua Chipani, T.; Rojas, B.J.; Virgilio, E.; Justo, B. Propagación Vegetativa de Esquejes de Queñua (Polylepis incana) con la Aplicación de dos Enraizadores naturales y Tres Tipos de Sustratos en Condiciones de vivero Cuajone, Torata-Moquegua (Vegetative Propagation of Queñua (Polylepis incana) Cuttings with the Application of Two Natural Rooters and Three Types of Substrates in Nursery Conditions Cuajone, Torata-Moquegua). 2020. Available online: https://revistas.ujcm.edu.pe/index.php/rctd/article/viewFile/168/150 (accessed on 22 June 2023). (In Spanish).
- Mollohuanca Ccuno, C.; Mayta Anco, L.; Bardales Alvarez, R. Growth Regulators (BAP and ANA) for in vitro Propagation of Queñoa (Polylepis rugulosa Bitter). Manglar 2021, 18, 207–213. [Google Scholar] [CrossRef]
- Boene, H.C.A.M.; Nogueira, C.A.; Sousa, J.N.; Kratz, D.; Dutra de Souza, P.V. Efeitos de diferentes substratos na produção de mudas de Sebastiania commersoniana (Effects of different substrates on the production of Sebastiania commersoniana seedlings. Floresta 2013, 43, 407–420. (In Spanish) [Google Scholar] [CrossRef]
- Dos Santos, J.; Davide, A.; Fonseca, L.; Santos, A.; De Melo, L. Enraizamento de Estacas Lenhosas de Espécies Florestais (Rooting of Flemish Cuttings of Florestai Species). CERNE 2011, 17, 293–301. [Google Scholar] [CrossRef]
- Carranza Patiño, M.; Ibarra, O.C.; Rodríguez, E.N.; Aguiar, S.S.; Falquez, O.C.; Troya, A.E.; Chancay, X.R.; Morante Carriel, J. Propagation of Tabebuia donnell-smithii rose (white guayacán) using rootings hormones. Cien. Tecnol. 2012, 5, 17–26. [Google Scholar] [CrossRef]
- Indacochea, B.V.Á.; Parrales, J.; Hernández, A.V.; Castro, C.; Vera, M.; Zhindón, A.C.R.; Gabriel, J. Evaluación de medios de cultivo in vitro para especies forestales nativas en peligro de extinción en Ecuador (Evaluation of in vitro culture media for native forest species in danger of extinction in Ecuador). Agron. Costarric. 2018, 42, 63–89. (In Spanish) [Google Scholar] [CrossRef]
- Sabah, S.; Hosni, A.; Abdul Raziq, A.; Hewidy, M. A Comprehensive Protocol for In Vitro Propagation and Acclimatization of Jacaranda mimosifolia Trees under the Egyptian Conditions. Egypt. J. Agric. Sci. 2023, 4, 79–92. [Google Scholar] [CrossRef]
- Maruyama, E. Tissue Culture of Swietenia macrophylla King (Big-Leaf Mahogany). In Plantation Technology in Tropical Forest Science; Suzuki, K., Ishii, K., Sakurai, S., Sasaki, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 131–136. [Google Scholar] [CrossRef]
- Gatica-Arias, A.; Vargas-Corrales, K.; Benavides-Acevedo, M.; Bolívar-González, A.; Sánchez-Chacón, E.; García-Díaz, E.; Delgado-Rodríguez, F.; Huang, N.T.W.; Hegele, M.; Wünsche, J.-N.; et al. Morphological and biochemical changes during somatic embryogenesis in mahogany, Swietenia macrophylla (Meliaceae). Rev. De Biol. Trop. 2019, 67, 406–418. [Google Scholar] [CrossRef]
- Pérez, J.; Quenta, E. Germinación in vitro de embriones de Nogal (Juglans boliviana). J. Selva Andin. Biosph. 2015, 3, 15–23. [Google Scholar] [CrossRef]
- Minocha, S.C.; Minocha, R. Historical Aspects of Somatic Embryogenesis in Woody plants. In Somatic Embryogenesis in Woody Plants; Jain, S., Gupta, P., Newtorz, R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; Volume 1, pp. 9–22. [Google Scholar]
- Campos Ruiz, J.; Arteaga Cuba, M.; Campos Ruiz, S.; Chico Ruíz, J.; Cerna Rebaza, L.; Campos Ruiz, J.; Arteaga Cuba, M.; Campos Ruiz, S.; Chico Ruíz, J.; Cerna Rebaza, L. Establecimiento de un protocolo de desinfección y micropropagación in vitro de «caoba» Swietenia macrophylla King (Meliaceae). Arnaldoa 2020, 27, 141–156. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Common Name | Origin | Medicinal Properties | Secondary Metabolites |
|---|---|---|---|
| Endangered Species | |||
| Myrcianthes pungens (O. Berg) D. Legrand) synonym: Eugenia punges/ Guaviyú, Guabijú, Yguaviyú, Ibabiyú, Yva viyú, Yva poreí | BO, BR, AR, UY, PY | Diuretic, digestive, and antidiarrheal properties, for sore throats and other throat conditions [43] | α and β-amyrins, quercetin, sesquiterpenes β-caryophyllene, 1,8-cineole, bicyclogermacrene, spathulenol, 5-epi-neointermedeol (leaves), gallic acid, catechin, vanillic acid, ellagic acid (fruit) [44,45,46] |
| Juglans neotropica Diels./ Nogal, cedro negro, Tocte | VE, CO, EC, PE | Antidiabetic, antioxidant, antifungal, antimicrobial, hypoglycemic, anti-hepatic, vaginal, gastric, and respiratory infections, open skin wounds, and treat sores. [47] | Gallic acid, ellagic acid, caffeic acid, tannins (seeds), hydroxycinnamic acid derivatives, flavonoid heterosides, sterol, beta-sitosterol, isoquinolin berberine, catechin, 3-0-glucosyl-galactosidequercetin, 5-hydroxy-tryptamine (leaves), juglone quinones, 1,4-naphthoquinone, currumidicin (fruit), juglone qinone, regiolone, betulin triterpenes, betulinic acid (stem) [48] |
| Ocotea quixos (Lam.) Kosterm./ Ishpingo, Espíngo, Canelillo | CO, EC, PE | Disinfectant, local anesthetic, against arthritis, antimicrobial, and antifungal activity; antiplatelet, antithrombotic, vaso-relaxing [49] | Transcinnamaldehyde (27.9%), methylcinnamate (21.6%), 1,8-cineole (8.0%), benzaldehyde (3.6%), β-selinene (2.1%) [50] |
| Vulnerable species | |||
| Cinchona pubescens Vahl, 1970/ Cascarilla, cinchona, Quinina, Quino, Kina | S. América (Andean Region) | Malaria treatment [51,52] | Cinchonine, cinchonidine, quinine, quinidine [53], triterpene heterosides, resins, catechin, and tannins [54]. |
| Myrceugenia glaucescens (Cambess.) D. Legrand & Kausel/ Murta | UY, BR, PY, N. of AR | Astringent, antidiarrheal, and digestive [55] | It shares several characteristics with its native relative, the Anacahuita, since some members of the Myrtaceae family produce essential oils (camphor, monoterpenes, sesquiterpenes, triterpenes, phenylpropanoids) [56] |
| Polylepis spp. Ruiz & Pavón/ Quinual, Queñoa | BO, PE, EC | Antihypertensive properties, for respiratory diseases, hypoglycemic [57] | Flavonoids, anthocyanins, carotenoids, triterpenes oxygenated, and flavonoids glycosidated [58,59] |
| Sebastiania commersoniana (Baill.) L. B. Sm. & Downs/Guindillo, Palo de leche | AR, UY | Antiseptic, external use [55] | Quercetin, isoquercitrin, cumarin, kaempferol, isorhamnetin, gallicin, scopoletin, syringic, and caffeic acids [60,61] |
| Handroanthus chrysanthus (Jacq.) S.O.Grose/ Guayacán amarillo, Araguaney | VE, CO, EC | Antimicrobial, antioxidant, hepatitis, alleviate osteoporosis [62] | Flavonoids, cardiotonic glycosides, sesquiterpene lactones [63] |
| Jacaranda mimosifolia D. Don, 1822/ Jacaranda, tarco | BR, BO, PY, PE, CO, EC, N. AR | Treatment of amoebic dysentery and other acute gastrointestinal conditions, antiseptic, antitumor, spasmolytic [64] | Jacaranone, jacoumaric acid, methyl jacaranone [65] |
| Capparidaceae Juss./Sapote, Zapote | VE, CO, EC, N. PE, Antilles | Gastrointestinal, inflammation, anemia, liver dysfunction, rheumatism, analgesic, antispasmodic, vermifuge, anti-hemorrhoidal, laxative, cleansing, diuretic, expectorant, body tonic [66] | Catechin, epicatechin, gallocatechin, catechin 3-O-gallate, gallic acid, syringic acid, p-hydroxybenzoic acid, p-coumaric acid, proanthocyanidins [67] |
| Swietenia macrophylia King/ Mahogany | PE, BR | Antipyretic, astringent, tonic [68] | Scopoletin, stearic acid methyl ester, beta-siterol, swietenine, swietenolide, swietemahonin E, F, G (seeds), catechin, epicatechin, swietemacrophyllanin (bark) swietephragmin (H, I, J), germacrene (A, D), swietenine (J) (leaves), 3-hydroxycaruilignan C (stem) [69] |
| Names of Trees | Type of Explant | Media Variant for Somatic Embryogenesis Induction and PGF | Media Variant for Somatic Embryo Recovery and PGF | References |
|---|---|---|---|---|
| Myrcianthes pungens | Nodal | WPM medium and BAP | WPM medium without PGF | [70] |
| Juglans neotropica | Disinfected embryos from mechanical rupture of the seed coat [71] | WPM medium and IBA (0.832 µM) and BAP (8.32 µM) | MS medium/DKW medium + BAP + TDZ | [88] |
| Ocotea quixos | - | - | - | - |
| Cinchona pubescens | - | - | - | - |
| Polylepis spp. | - | - | - | - |
| Sebastiania commersoniana | - | - | - | - |
| Handroanthus chrysanthus | - | - | - | - |
| Jacaranda mimosifolia | - | - | - | - |
| Swietenia macrophylla | Shoot tips of aseptically germinated seedlings Vegetative material from clones | WPM (solid or liquid) and ZEA (1–10 µM) and AC WPM liquid and L-cysteines, CH, malt extract, citric acid, IBA, 2,4-S, iP, +sucrose [87] | WPM without PGF/WPM and low ZEA (0.1 µM)/WPM and glutamine, arginine, asparagine, lysine, proline WPM liquid medium and 6-BAP [88] | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihai, R.A.; Melo Heras, E.J.; Pinto Valdiviezo, E.A.; Espinoza Caiza, I.A.; Cubi Insuaste, N.S.; Mejía, J.P.; Catana, R.D.; Moldoveanu, M.M.; Florescu, L.I. Somatic Embryogenesis of Representative Medicinal Trees in South America—Current Status. Forests 2023, 14, 2066. https://doi.org/10.3390/f14102066
Mihai RA, Melo Heras EJ, Pinto Valdiviezo EA, Espinoza Caiza IA, Cubi Insuaste NS, Mejía JP, Catana RD, Moldoveanu MM, Florescu LI. Somatic Embryogenesis of Representative Medicinal Trees in South America—Current Status. Forests. 2023; 14(10):2066. https://doi.org/10.3390/f14102066
Chicago/Turabian StyleMihai, Raluca A., Erly J. Melo Heras, Eliza A. Pinto Valdiviezo, Iván A. Espinoza Caiza, Nelson S. Cubi Insuaste, Jean Pierre Mejía, Rodica D. Catana, Mirela M. Moldoveanu, and Larisa I. Florescu. 2023. "Somatic Embryogenesis of Representative Medicinal Trees in South America—Current Status" Forests 14, no. 10: 2066. https://doi.org/10.3390/f14102066