
Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications

1
Yunnan Key Laboratory of Plant Reproductive Adaptation and Evolutionary Ecology, Institute of Biodiversity, School of Ecology and Environmental Science, Yunnan University, Kunming 650500, China
2
Yunnan Key Laboratory for Integrative Conservation of Plant Species with Extremely Small Populations, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
3
The Key Laboratory of Rare and Endangered Forest Plants of National Forestry and Grassland Administration & The Key Laboratory for Silviculture and Forest Resources Development of Yunnan Province, Yunnan Academy of Forestry and Grassland, Kunming 650201, China
4
Southeastern Center for Conservation, Atlanta Botanical Garden, Atlanta, GA 30309, USA
*
Author to whom correspondence should be addressed.
Forests 2024, 15(2), 235; https://doi.org/10.3390/f15020235
Submission received: 24 December 2023
/
Revised: 18 January 2024
/
Accepted: 23 January 2024
/
Published: 26 January 2024
(This article belongs to the Section Forest Biodiversity)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Seed germination is among the most crucial and vulnerable stages in plant life cycles. Quercus marlipoensis is a critically endangered sclerophyllous oak. Only one population has ever been found worldwide in the tropical montane cloud forests of southeastern Yunnan, China, and it has shown difficulties with regeneration. However, its seed biological traits and key restrictive germination factors remain unknown. We investigate the impacts of scarification, temperature, and water potential on the seed germination of Q. marlipoensis. Results show that the seeds show typical epicotyl dormancy. The seed germination increased when removing part or all of the pericarp and part of the cotyledon (one-third and two-thirds). The seeds can germinate at 5 to 30 °C, but the highest T50 was achieved at 25 °C. When the water potential decreased from 0 to −1.0 MPa, the germination rate decreased but the germination time increased. Q. marlipoensis seeds are typically recalcitrant and highly sensitive to moisture loss, but the species can tolerate animal predation and low germination temperatures. The more frequent climatic extremes and droughts in the Indo-China region will severely degrade its natural habitats. Therefore, ex situ conservation to preserve its germplasm and introduce seedlings into a suitable habitat are essential for its conservation management.
1. Introduction
Tropical montane cloud forests (TMCFs) are unique narrow altitudinal belts in tropical highlands [1,2] between 1000 and 3000 m above sea level [3]. Their microclimate is characterized by persistent, frequent, or seasonal cloud cover at the vegetation level, with fog and mist surrounding the canopy [4]. The structure and function of TMCFs are highly vulnerable to climate change, particularly temperature increase [5], which reduces mist and causes the extinction of cloud forests on mountain peaks [6].
Quercus marlipoensis Hu (section Ilex) is an endemic evergreen sclerophyllous oak distributed in Malipo County, Yunnan Province, China [7]. This species was first reported by Hu (1951) [8] based on the voucher specimen collected in 1940 by C. W. Wang. Since then, no new specimens nor occurrence data have been reported for decades, even after three national plant inventory surveys were conducted from 1990 to 2010. However, our field expedition in 2012 rediscovered this species in the core protection area of Lao-Jun-Shan National Nature Reserve, where the specimen originated [9]. Still, no other population has been discovered in the past ten years after extensive field surveys. The population showed significant signs of regeneration difficulties, as seedlings or saplings were relatively rare. The IUCN Red List classifies this species as Critically Endangered (CR) [10]. The newly published book List of Yunnan Protected Plant Species with Extremely Small Populations (PSESP) [11] also included this species. However, the reasons for the rarity of Q. marlipoensis and the possible factors threatening its regeneration are still unknown. Understanding the important physiological and ecological characteristics of Q. marlipoensis is crucial for identifying its threatening factors and conservation strategies.
Poor regeneration is one of the factors that lead to the decline of threatened species [12]. Knowledge of the seed germination characteristics of PSESPs is important for the conservation of threatened species [12,13]. Biotic and abiotic stress can profoundly impact the seed germination and seedling establishment process. Cotyledons of oak seeds rich in starch and other nutrients are a dietary staple of rodents and insects and the foundation of the food chain in a forestry ecosystem [14]. Meanwhile, the energy reserves in cotyledons are also an important source supporting seedling development [15,16,17]. If animals in the ecosystem intensively consume oak seeds, seed mortality will dramatically increase, preventing tree populations from regenerating [18,19,20,21]. Mechanical scarification is a practice of simulating animals preying on seeds, and different treatments (pericarp or partial cotyledon removal) resemble the degree to which the seeds are eaten by animals [22,23]. Oak seeds show very different levels of sensitivity to animal predation among species. Previous mechanical scarification studies showed that losing a small part of the cotyledons in acorns of various oak species only has a minor impact on seedling establishment [24,25,26] and may even increase the germination ratio [26,27]. Under loss of cotyledons, the responses of seeds are quite different among oak species, e.g., the hypocotyl and radicle mortality dramatically increased in Q. variabilis [28], but Q. robur seeds can successfully germinate and establish seedlings even when they have suffered from severe cotyledon reductions of up to two-thirds [27]. Evaluating seed tolerance to animal predation is an essential step to untangling the factors limiting the regeneration of Q. marlipoensis.
Temperature and moisture (water potential) are the key environmental factors impacting seed germination [12,29]. Oak seeds are mostly recalcitrant, with a moisture content of 30%–55%, and seed moisture loss significantly decreases the germination ratio [30,31,32,33,34]. The majority of seeds exhibit rapid germination and continuously maintain high activity without dormancy characteristics post-maturation [35,36]; however, a small percentage of oak seeds may undergo dormancy [37], influenced by the effects of growth regulators, pericarp restrictions, or a combination of both [38,39], breaking dormancy in oak seeds typically requires stratification or pericarp abrasion [40]. As a widespread Northern Hemisphere woody clade, oaks show high ecological divergence. Their habitats range from humid temperate and tropical forests to semi-arid deserts [41,42]. Correspondingly, the best germination temperatures of oaks have typically been confirmed to match their habitats’ climatic features [43], ranging from 5 °C to 35 °C [43,44]. Although oak habitats are quite different, their seeds generally show a degree of cold tolerance, as most of them can germinate at a low base temperature (Tb) of about 5 °C [29,43,45]. Overall, the currently published seed physiological research on oaks is mainly on widespread oaks. The germination traits of endangered oaks remain largely unknown.
Considering the low regeneration capacity of Q. marlipoensis in its natural habitat, it is crucial to reveal the key factors restricting its seed germination and seedling establishment to provide conservation guidelines for this extremely endangered oak species. This study was undertaken to reveal the effects of scarification, temperature, and water potential on the seed germination of Q. marlipoensis. This study provides crucial information on this critically endangered oak in order to provide guidelines on its in situ and ex situ conservation.
2. Materials and Methods
2.1. Study Area and Acorn Collection
The habitat of Quercus marlipoensis is located at Xia-Jin-Chan village of the Lao-Jun-Shan National Nature Reserve (23.40859° N, 104.70901° E) of southeast Yunnan Province, China. It is found in TMCFs at the middle-elevation (1700–1900 m above sea level) karst areas characterized by high humidity and warm temperatures (Figure 1). The region is under the Indian summer monsoon regime with prominent warm rainy summers and mild, dry winters. The annual highest and lowest temperatures are 28.10 °C and 6.50 °C, respectively. The annual accumulated temperature greater than 10 °C is about 4500 °C to 7500 °C, and the annual average precipitation is 1318 mm [6]. In the region, we visited residents and conducted surveys of all potential forests to search for additional populations of Q. marlipoensis. However, only 64 mature and 4 young trees were scattered throughout the forested area.
Acorns of Q. marlipoensis were collected randomly in late October 2021. At least 4 kg of seeds were collected from different Q. marlipoensis adult individuals. The collected seeds were then randomly distributed into three groups for morphological measurements, dry and wet weight assessments, and seed germination experiments, respectively. Before commencing the experiments, thorough visual inspections were conducted on the acorns. Any acorns displaying abnormalities or noticeable defects were removed from the selection.
2.2. Morphological Traits of the Acorn and Seed
The length (L), cupule length (CL), width (W), and seed scar diameter (CSD) of 30 random fresh acorns were measured with a vernier caliper. The morphological variation in acorns and the longitudinal section of the seeds were captured in images using a stereo-microscope (Leica S9I, Am Leitz-Park, Wetzlar, Germany).
The acorns were divided into three batches (n = 100) to determine their fresh weight and then were dried at 103 ± 2 °C in an oven for 17 h to obtain the dry weight following a previously described protocol [46] (International Seed Testing Association 1999). The initial moisture content of acorns was calculated as (fresh weight − dry weight)/fresh weight × 100% [47].
2.3. Seed Pretreatment and Germination Test
After removing the cupule, the acorns were soaked in deionized water for two hours. The floating acorns were discarded. The sunken acorns were selected and surface-disinfected by soaking them in 1% sodium hypochlorite solution for 0.5 h, then rinsing them three times using deionized water [47] before the subsequent germination tests. All treatments in the three experiments were carried out with three replicates, and each replicate included 20 acorns. Plump acorns of similar size were selected and placed in covered plastic boxes, with two layers of filter paper on the bottom of the box as the germination substrate. Seeds germinated under conditions of 25 °C, with 16 h of light followed by 8 h of darkness; germination humidity was near 100 percent [48].
For the scarification experiment, five treatments were performed on the acorns: intact seeds with pericarp (CK), removal of cup scar (RS), removal of the pericarp (RP), cutting off 1/3 of the cotyledon (TC), and removal of the pericarp and cutting off 2/3 of the cotyledon (OC).
Plump acorns of similar size were selected and placed in covered plastic boxes. The boxes were placed in growth chambers spanning a set temperature gradient (5 °C, 10 °C, 15 °C, 20 °C, 25 °C, and 30 °C).
To evaluate the effects of water stress on seed germination, different concentrations of PEG-6000 solutions (Yatai United Chemical Co., Ltd., Wuxi, China) with corresponding water potentials of 0, −0.2, −0.4, −0.6, −0.8, and −1.0 M pa [49] were added to the bottom of the plastic boxes along the edge of the filter paper until the filter paper was completely saturated. The filter papers and water/PEG solutions were replaced every other day.
2.4. Calculation of Seed Germination Indices
Seeds were considered germinated when the radicle emerged more than 2.0 mm [43]. The germinated seeds were counted and removed every other day. Counting was ended when either all of the seeds had grown or no germination was observed for 30 consecutive days [46].
The statistical analysis and visualization of germination data were performed using the ‘Germinationmetrics’ package of R version 4.1.2 [50]. The indices for evaluating seed vigor we selected were germination percentage, the initial germination time, the last germination time, time for 50% of seeds to germinate (), germination value, and germination index [51,52,53].
The x-intercept method was used to determine the base temperature (Tb) and base water potential (ψb). Germination percentage was calculated as the inverse of T50, and linear regressions were conducted to determination its relationship with temperature and water potential, respectively. The x-intercept of the linear equation was calculated as Tb or ψb [54,55].
2.5. Statistical Analyses
Analysis of variance (ANOVA) was used to assess the effects of different scarification methods, temperature, and water potential on seed germination. Germination percentages were analyzed after arcsine transformation because of the non-normal distribution of the data. The differences between means were tested using Tukey’s HSD test at a p ≤ 0.05 threshold. All statistical analyses were conducted with R version 4.1.2.
3. Results
3.1. Morphological Characteristics of the Acorn and Seed Germination
The average fresh weight of each acorn was 3.12 g (derived from 100-seed weight). The moisture content of acorns was 55.34% ± 1.56%. The acorn is ellipsoidal with a cup-shaped cupule. The purplish-red triangular bracteoles were imbricated on the outside of the cupule and more prominent at the top (Figure 2a). The mean length and width of the acorn were 1.77 ± 0.17 cm and 1.39 ± 0.08 cm, respectively. The length of the cupule was 1.01–1.45 cm (average 1.22 cm), and the cup scar diameter (also known as seed scar diameter) was 0.45–0.67 cm (average 0.56 cm). The seed cotyledon is creamy white or light yellow (Figure 2c). The radicle penetrates the seed coat at or beside the remains of the style when germinating (Figure 2d,e). The epicotyl did not elongate until 12 weeks after germination when the radicle was 7.00–9.00 cm long (Figure 2f).
3.2. Seed Germination Traits
3.2.1. Scarification and Germination
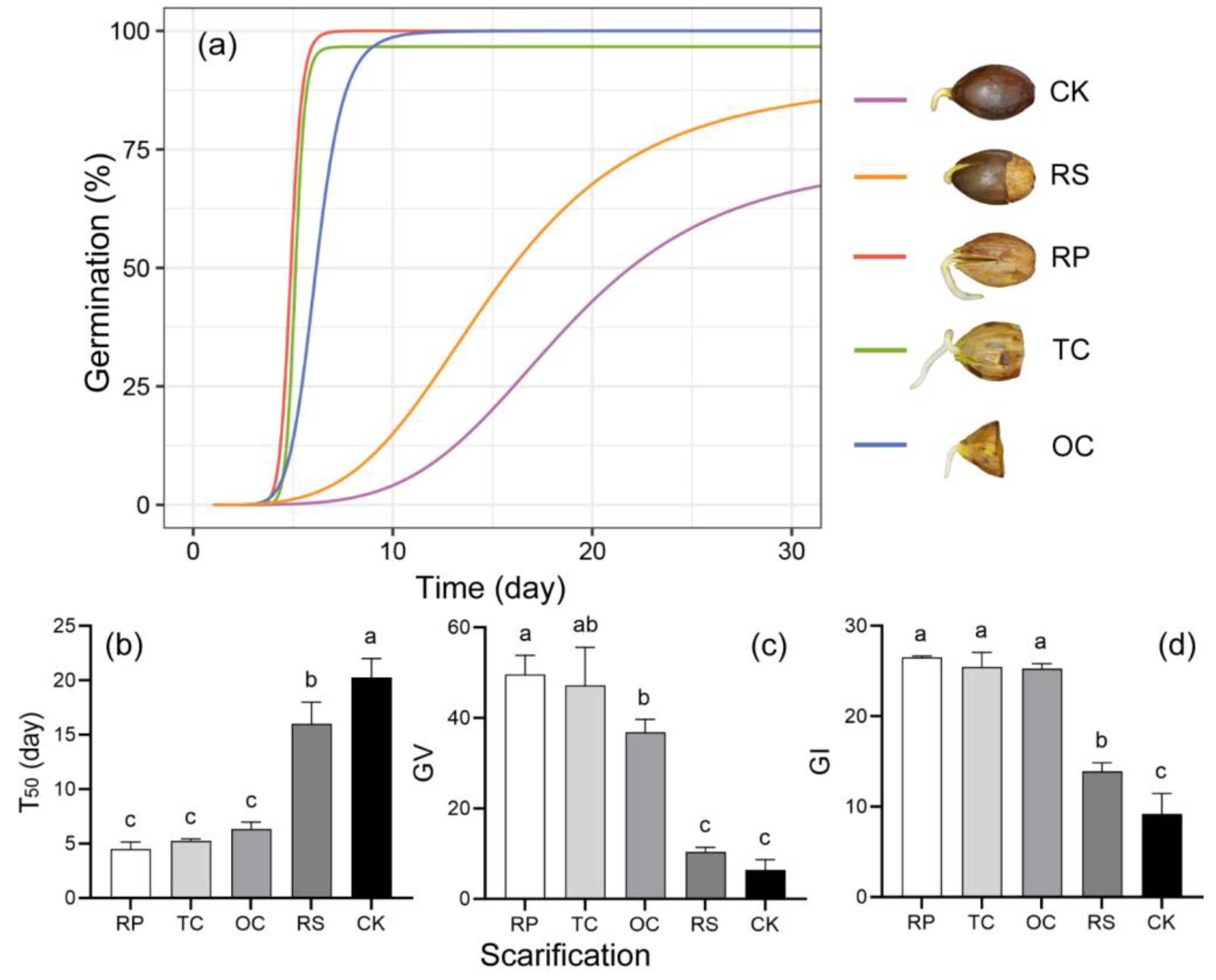
Seeds subjected to pericarp removal and/or partial cotyledon removal showed 90.00% to 100.00% germination percentages, while only 73.33% germinated in the control group. Pericarp removal also significantly accelerated the speed of germination (Table S1, RP_T50 = 4.70 ± 0.55 day, CK_T50 = 19.88 ± 1.63 day). The initial germination time showed no significant difference among the five treatments (Table S1, p > 0.05), but the last germination time of the acorns without pericarp tissue was earlier than that of the control and RS groups (Table S1, Figure 3a). The significant decrease in T50 and increase in both the germination value and the germination index (Figure 3b–d) indicated that the intact pericarp impeded germination. However, cup scar removal decreased T50 and increased the germination index less significantly than removal of the whole pericarp or cutting off part of the cotyledon (RP, TC, OC).
3.2.2. Temperature and Germination
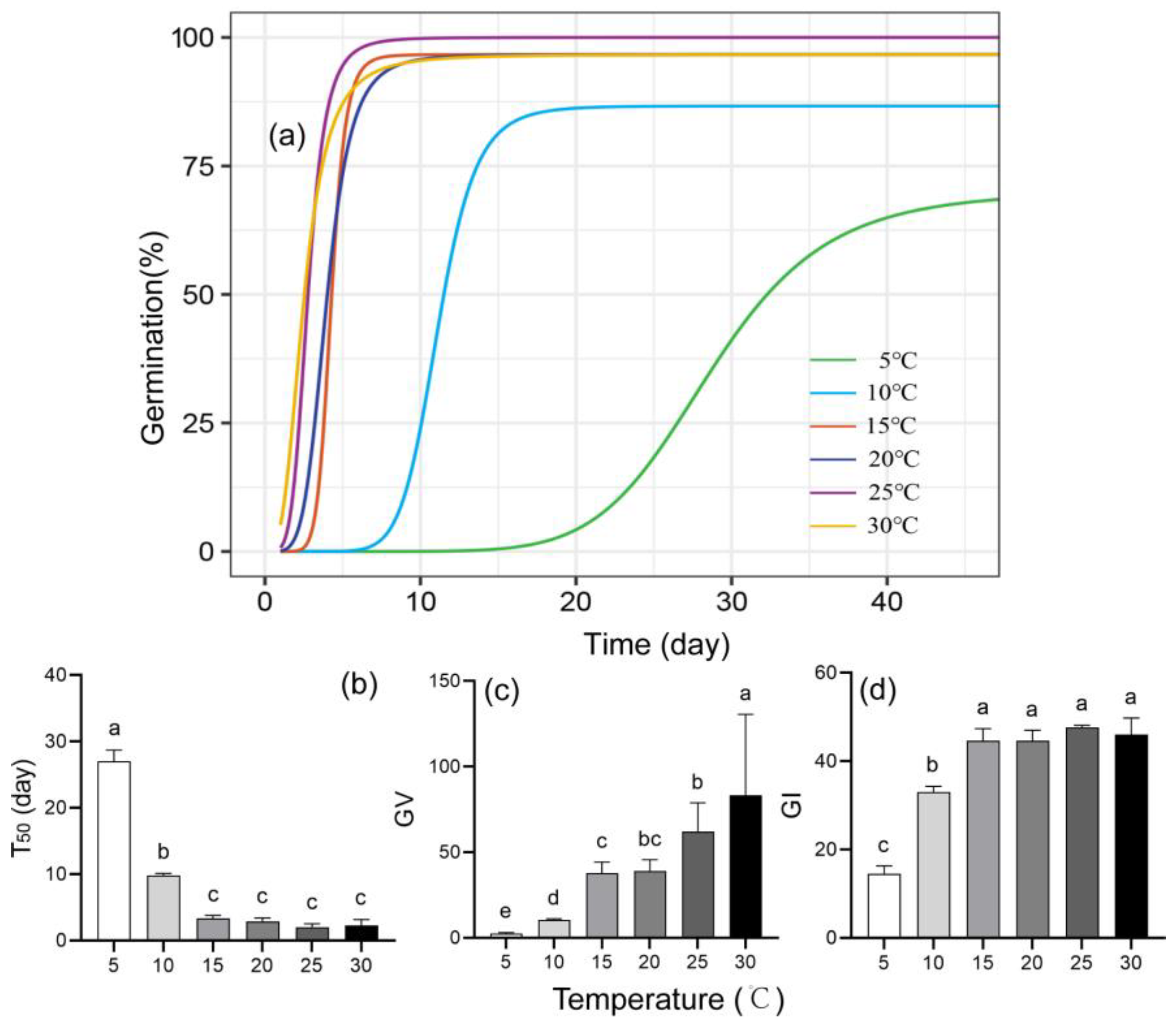
Seeds at all six of the tested temperatures (from 5 °C to 30 °C) had high cumulative germination percentages ranging from 70.00% to 100.00% (Table S2, Figure 4a). The initial and last germination times at 5 °C and 10 °C were significantly later than those at 15–30 °C (Table S2 and Figure 4a, p < 0.05). The time to complete germination was shortened significantly above 15 °C. T50 increased significantly at 5–15 °C and remained low at 20–30 °C (Table S2 and Figure 4b). The germination value and germination index increased with temperature (Figure 4c,d).
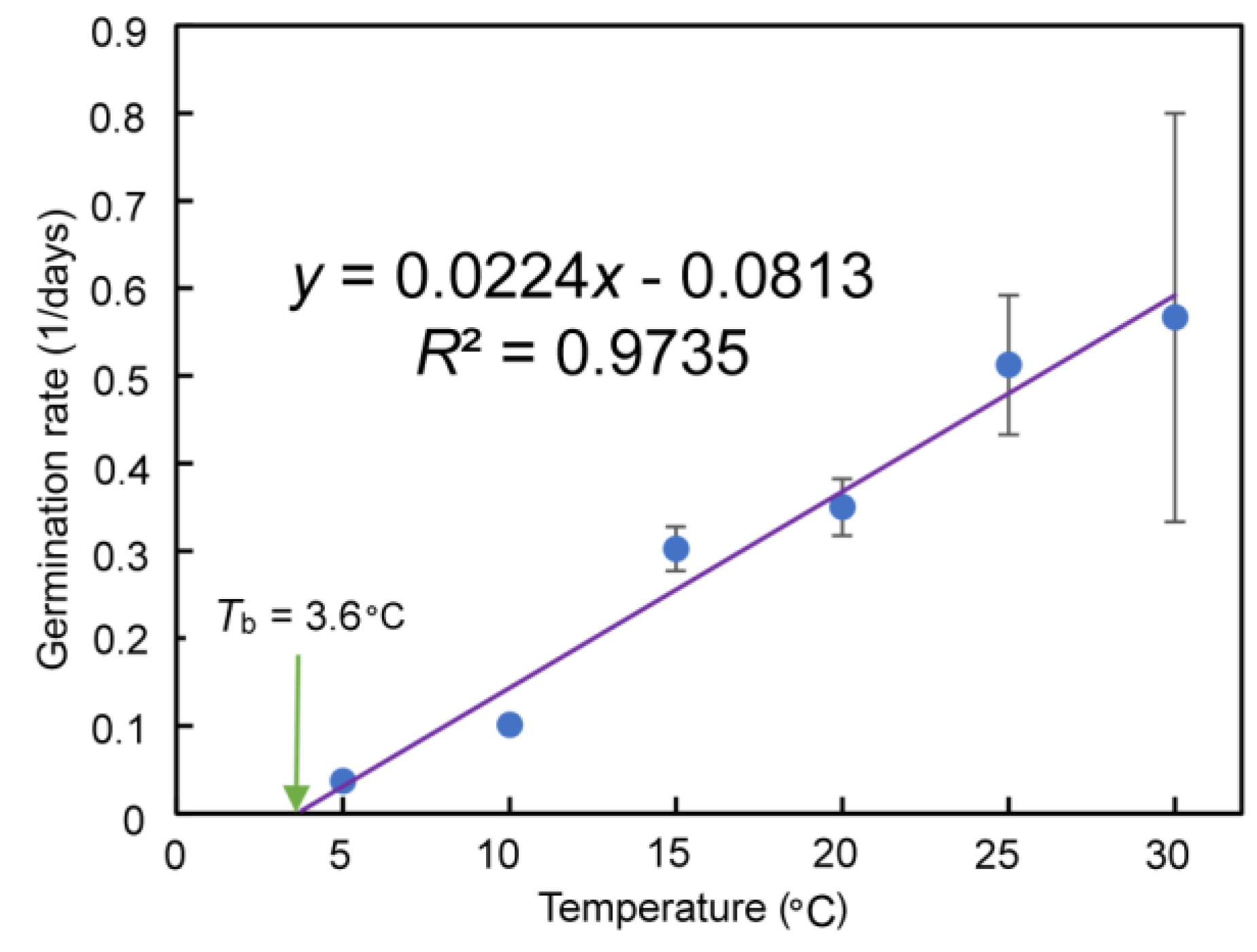
GR increased linearly with temperature, reaching 0.57 days−1 at 30 °C, though the final germination percentage did not increase from 15 °C to 30 °C. The estimated Tb was 3.60 °C (Figure 5).
3.2.3. Water Potential and Germination
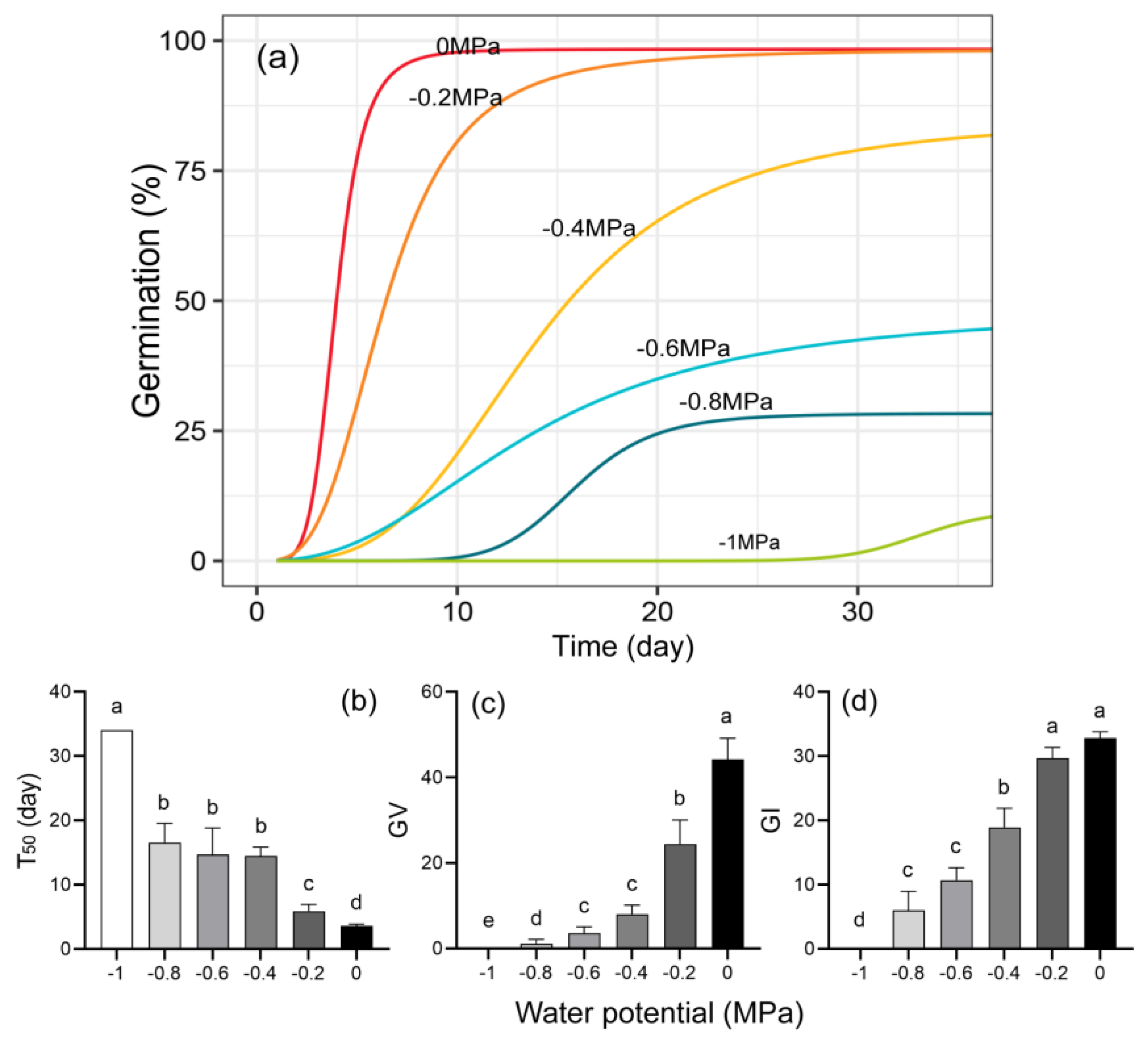
Water potential remarkably affected germination percentage, T50, germination value, and germination index (Table S3, Figure 6). Lower water potential (i.e., more negative values) reduced the final germination percentage. The seeds subjected to the control treatment (0 MPa) started to germinate within 3 days (Table S3), while those under water stress took 2 to 4 weeks to start germination (Figure 6a). T50 extended from 3.6 days to 34 days across water potential values from −0.2 MPa to −1 MPa (Figure 6b). In other words, germination significantly slowed as water potential decreased. Both germination value and germination index decreased as water potential decreased (Figure 6c,d).
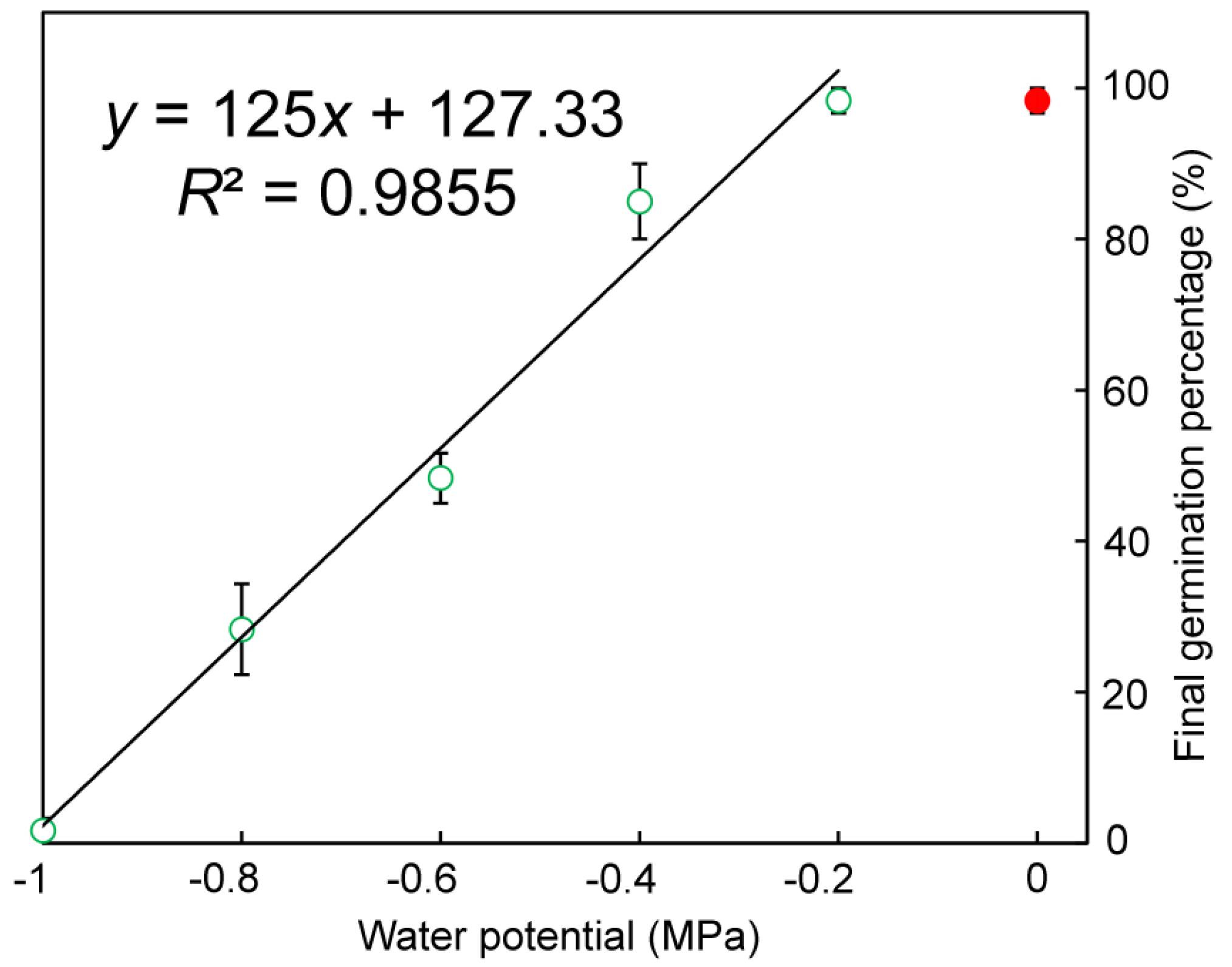
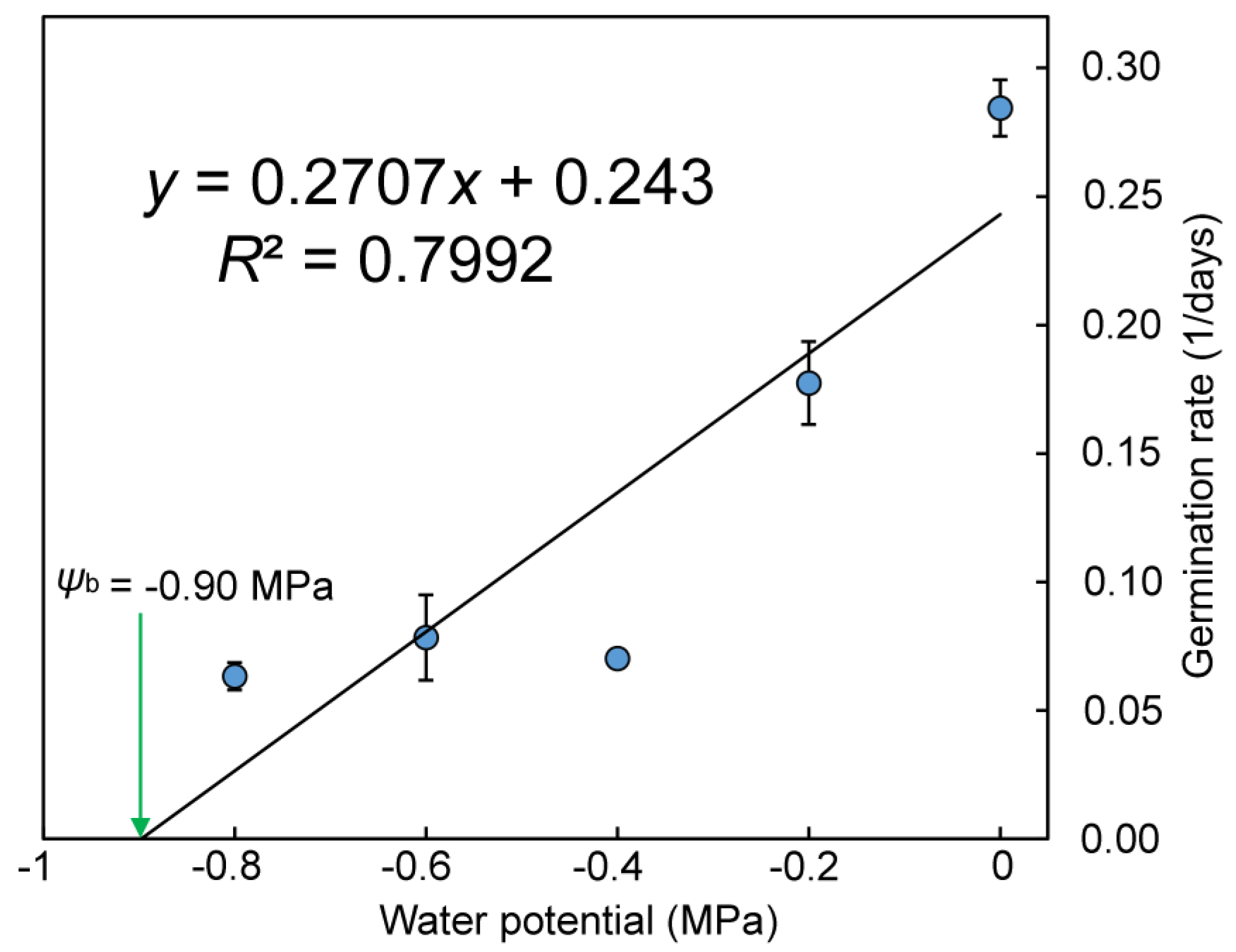
The final germination percentages decreased linearly with water potential between −0.2 and −1.0 MPa (Figure 7). Almost all the seeds germinated under 0 MPa and −0.2 MPa. The germination percentage also decreased linearly with water potential (Figure 8). The base water potential (ψb) was −0.9 MPa according to the x-intercept of the linear regression.
4. Discussion
4.1. Impacts of Pericarp and Cotyledon Removal on the Germination of Quercus marlipoensis
Animal predation on the pericarp and cotyledons of acorns may accelerate seed germination and increase the seed germination percentage [26,56]. Our results showed that the complete removal of the pericarp and partially cutting off the cotyledons significantly improved the final germination percentage and shortened the germination time of Q. marlipoensis. The removal of the cup scar enhanced the germination percentage and shortened the germination time, but the influence was not as great as the complete pericarp removal treatment, and this result is consistent with findings from other studies on oak seeds [23,27]. Pericarp removal improves water absorption and gas exchange, boosting the germination ratio [57]. The observed improvement in germination could be explained by the removal of both the mechanical barrier and inhibitory substances in the pericarp and/or cotyledon [48,58,59].
Although Q. marlipoensis is critically endangered, its seeds can individually tolerate a medium-high level of animal predation. The influence of cotyledon loss by animal consumption on oak seed germination and seedling survival varies by species. For example, a loss of more than 50% of the cotyledon tissue has a significant negative effect on seed germination and seedling growth in Q. variabilis [28,60] as well as Q. prinus and Q. velutina [61]. Our results showed that loss of two-thirds of the cotyledon tissue had no significant Impact on the germination of Q. marlipoensis, suggesting that Q. marlipoensis only needs a little nutrition from its cotyledons to ensure its germination and that it has good tolerance to animal predation. Likewise, similar results were obtained in Q. robur [27] and Q. mongolica [62]. During the coevolution between oaks and seed predators, oaks tend to develop large acorns or extra nutrition to ensure germination, even under high predation pressure [63,64].
4.2. Effect of Temperature on Seed Germination and the Base Temperature
Warm environmental conditions are generally suitable for seed germination, but some seeds can also tolerate low temperatures. The estimated base temperature for Q. marlipoensis seed germination is 3.60 °C. Its seeds can germinate at high percentages at temperatures above 5 °C, and germination sped up once the temperature increased. Temperatures from 15 °C to 30 °C promoted similarly high germination percentages and speeds. Similar results were found among different source populations of Q. robur from England and the Netherlands [65]. The threshold values are linked to the species’ geo-climatic origins [29]. Oak species from subalpine, subtropical, and temperate regions have been shown to have a similar low Tb, but different germination percentages [43]. In contrast, oaks in the section Cyclobalanopsis from tropical and subtropical Asia cannot germinate well at 15 °C [66]. The difference in the seed germination temperature of oak species from different biomes indicates that local adaptation indeed plays a role in the response to diverse environmental factors. Likewise, our study showed that seeds of Q. marlipoensis have a germination percentage of 70.00% at 5 °C, with Tb at 3.60 °C, which is much higher than that of evergreen oaks from subtropical lowlands. The ability to germinate at low temperatures suggests that Q. marlipoensis can cope with cool temperatures during the germination period under sufficient water content conditions, which is compatible with the microhabitat of intermediate-elevation TMCFs.
4.3. Effect of Water Potential on Seed Germination
Quercus marlipoensis is a recalcitrant-seeded species that is highly sensitive to desiccation, but its seeds can germinate quickly in moist conditions. Our results showed that Q. marlipoensis acorns are unable to tolerate osmotic potentials lower than −0.2 MPa. Therefore, its seeds can only successfully germinate and establish seedlings in humid soils. This result is similar to those found in other endangered Quercus spp. (e.g., Q. insignis, Q. sartorii, and Q. xalapensis) in TMCFs in Mexico, which can only grow vigorously in humid understory forests [67,68]. Rapid germination of various species of Quercus could be a strategy to reduce desiccation-induced mortality [43] and avoid predation [56].
Nevertheless, oaks in different habitats show different levels of resistance to water loss during germination. For example, seed germination percentage and germination percentage of Q. leucotrichophora from the dry, hot valleys of the central Himalayas were inhibited under a −0.9 MPa water potential, but the species still had a 20% final germination percentage [69]. Tropical trees growing in humid regions require a high base water potential for germination [29,57], and their tolerance to drought in the subtropical humid forests is much lower than that of seeds from tree species in arid environments [70,71].
4.4. Conservation Implications
The seed germination process of Q. marlipoensis is highly sensitive to humidity loss in the soil (i.e., low water potentials of −0.8 MPa and −1 MPa), and it can germinate rapidly under high water potentials. This characteristic is compatible with the unique cool and high-humidity microclimate of TMCFs. It is worth noting that Q. marlipoensis grows in limestone areas characterized by higher soil water evaporation rates and lower soil moisture than those of other soil types [72,73,74]. Such soil conditions could make Q. marlipoensis more vulnerable to drought stress.
Southwest China is under the Indian summer monsoon regime, with a prominent dry season beginning in September and lasting until the subsequent April [75]. The unique topography and microhabitats of TMCFs ensure the high humidity of these forests. However, the increasing length of the seasonal dry period coupled with more frequent climate extremes, e.g., drought and extreme heat events, throughout Southwest China and Indo-China as a result of global climate change profoundly impacts the local biota [76,77,78], which also degrades the TMCF habitat and profoundly impacts its natural seed germination and seedling establishment processes. However, oak acorns are not efficient propagules for long-distance dispersal that can track suitable habitats over time. Therefore, in situ conservation which first introduce the seeds to nurseries/botanical gardens and then reintroduce large seedlings back into suitable habitats with anticipated long-term stability will be an effective solution to help maintain the population size of Q. marlipoensis. Moreover, a growing body of species distribution dynamics studies have predicted a general trend of northward range expansion pattern of Asian evergreen broadleaved forest tree species [79,80,81]. Nevertheless, the other physiological characteristics of Q. marlipoensis are still unknown. Future studies coupling species distribution modeling and physiological analysis in Q. marlipoensis will provide sufficient guidance to the conservation of this endangered oak in the context of global climate change.
5. Conclusions
Quercus marlipoensis is a critically endangered tree restrictively distributed within the TMCFs of southeastern Yunnan, China. Its seeds are recalcitrant, with typical epicotyl dormancy; they germinate quickly once mature, but epicotyl development is significantly delayed. The acorns of Q. marlipoensis can each tolerate medium-high animal predation, and its seeds are viable at low germination temperatures, with a relatively high germination percentage. Its seed germination process is highly sensitive to humidity loss. The germination process of Q. marlipoensis is compatible with the microenvironments of TMCFs. But it only grows in limestone areas, and this soil type is characterized by higher soil water evaporation and low soil moisture when under environmental drought conditions. The increasing length of the seasonal dry period and more frequent climate extremes in Southwest China and Indo-China in recent years has led to habitat degradation, resulting in difficulties with seed germination and seedling establishment of Q. marlipoensis in its natural habitat. Therefore, manual ex situ germplasm conservation aiming to introduce seeds to nurseries and botanical gardens for propagation and subsequently reintroduce large seedings to suitable habitats should be carried out to conserve this extremely endangered oak species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f15020235/s1, Table S1: Seed germination indices of Quercus marlipoensis under different pericarp removal and cotyledon cut-off treatments of, Table S2: Seed germination indices of Quercus marlipoensis at different temperatures, Table S3: Seed germination indices of Quercus marlipoensis under different water potentials.
Author Contributions
M.D. conceived the idea. L.L. and Y.T. performed the experiments, and L.L. collected the data. L.L. and Q.L. analyzed the data. L.L., M.D. and Q.L. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the National Natural Science Foundation of China (grant. no. 31972858), the International Oak Society (IOS) Oak Conservation and Research Fund, the Fund of Yunnan Key Laboratory for Integrative Conservation of Plant Species with Extremely Small Populations (grant. no. PSESP2021), and the Project of the Yunnan Academy of Forestry and Grassland (grant. no. KFJJ21-05), the project of the Southeast Asia Biodiversity Research Institute, Chinese Academy of Sciences (grant. no. Y4ZK111B01), and National Key Research and Development Program of China 2023YFF1305000.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy.
Acknowledgments
We are grateful to the Malipo Lao-Shan Provincial Nature Reserve of Yunnan Province, the Asian Elephant Yunnan Field Scientific Observation and Research Station, the Yunnan Asian Elephant Field Scientific Observation and Research Station of the Ministry of Education, and the Baima Snow Mountain Complex Ecosystem Vertical Transect Field Observation for their help with the fieldwork and seed collecting. We thank Lin Lin, Yuxin Ma, and Chaoying Yang for their help with data analyses.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kappelle, M.; Burley, S.; Evans, J.; Youngquist, J. Tropical montane forests. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Amsterdam, The Netherland, 2004; Volume 4, pp. 1782–1793. [Google Scholar] [CrossRef]
- Ray, D.K. Tropical montane cloud forests. In Climate Vulnerability; Pielke, R.A., Ed.; Academic Press: New York, NY, USA, 2013; Volume 5, pp. 79–85. [Google Scholar] [CrossRef]
- van de Weg, M.J.; Meir, P.; Williams, M.; Girardin, C.; Malhi, Y.; Silva-Espejo, J.; Grace, J. Gross primary productivity of a high elevation tropical montane cloud forest. Ecosystems 2014, 17, 751–764. [Google Scholar] [CrossRef]
- Fanourakis, D.; Aliniaeifard, S.; Sellin, A.; Giday, H.; Körner, O.; Rezaei Nejad, A.; Delis, C.; Bouranis, D.; Koubouris, G.; Kambourakis, E.; et al. Stomatal behavior following mid or long–term exposure to high relative air humidity: A review. Plant Physiol. Biochem. 2020, 153, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Eller, C.B.; Meireles, L.D.; Sitch, S.; Burgess, S.S.O.; Oliveira, R.S. How climate shapes the functioning of tropical montane cloud forests. Curr. For. Rep. 2020, 6, 97–114. [Google Scholar] [CrossRef]
- Shi, J.P.; Zhu, H. Tree species composition and diversity of tropical mountain cloud forest in the Yunnan, southwestern China. Ecol. Res. 2009, 24, 83–92. [Google Scholar] [CrossRef]
- Tang, C.Q. Evergreen sclerophyllous Quercus communities. In The Subtropical Vegetation of Southwestern China; Tang, C.Q., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2015; Volume 11, pp. 113–125. [Google Scholar] [CrossRef]
- Hu, H.H. Additional notes on the Fagaceae of Yunnan, II. Acta Phytotax. Sin. 1951, 2, 139. [Google Scholar]
- Deng, M.; Jiang, X.L.; Song, Y.G.; Coombes, A.; Yang, X.R.; Xiong, Y.S.; Li, Q.S. Leaf epidermal features of Quercus group Ilex (Fagaceae) and their application to species identification. Rev. Palaeobot. Palynol. 2017, 237, 10–36. [Google Scholar] [CrossRef]
- Carrero, C.; Strijk, J.S. Quercus marlipoensis. The IUCN Red List of Threatened Species 2020: E.T191481A1985007. Available online: https://www.iucnredlist.org/species/191481/1985007 (accessed on 20 January 2024).
- Sun, W.B. List of Yunnan Protected Plant Species with Extremely Small Populations (2021); Yunnan Science and Technology Press: Kunming, China, 2021. [Google Scholar]
- Iralu, V.; Barbhuyan, H.S.A.; Upadhaya, K. Ecology of seed germination in threatened trees: A review. Energy Ecol. Environ. 2019, 4, 189–210. [Google Scholar] [CrossRef]
- Wade, E.M.; Nadarajan, J.; Yang, X.Y.; Ballesteros, D.; Sun, W.B.; Pritchard, H.W. Plant species with extremely small populations (PSESP) in China: A seed and spore biology perspective. Plant Divers. 2016, 38, 209–220. [Google Scholar] [CrossRef]
- Yi, X.F.; Yang, Y.Q. Large acorns benefit seedling recruitment by satiating weevil larvae in Quercus aliena. Plant Ecol. 2010, 209, 291–300. [Google Scholar] [CrossRef]
- Tripathi, R.S.; Khan, M.L. Effects of seed weight and microsite characteristics on germination and seedling fitness in two species of Quercus in a subtropical wet hill forest. Oikos 1990, 57, 289–296. [Google Scholar] [CrossRef]
- Leishman, M.R.; Westoby, M. The role of seed size in seedling establishment in dry soil conditions-experimental evidence from semi-arid species. J. Ecol. 1994, 82, 249–258. [Google Scholar] [CrossRef]
- Xiao, Z.; Harris, M.K.; Zhang, Z. Acorn defenses to herbivory from insects: Implications for the joint evolution of resistance, tolerance and escape. For. Ecol. Manag. 2007, 238, 302–308. [Google Scholar] [CrossRef]
- Yi, X.F.; Zhang, Z.B. Influence of insect-infested cotyledons on early seedling growth of Mongolian oak, Quercus mongolica. Photosynthetica 2008, 46, 139–142. [Google Scholar] [CrossRef]
- Montes-Hernández, B.; López-Barrera, F. Seedling establishment of Quercus insignis: A critically endangered oak tree species in southern Mexico. For. Ecol. Manag. 2013, 310, 927–934. [Google Scholar] [CrossRef]
- García-Hernández, M.d.l.Á.; López-Barrera, F.; Vásquez-Reyes, V.M. Microhabitat affects acorn removal in three sympatric and endangered neotropical oak species. Ecol. Res. 2016, 31, 343–351. [Google Scholar] [CrossRef]
- Cao, L.; Guo, C.; Chen, J. Fluctuation in seed abundance has contrasting effects on the fate of seeds from two rapidly germinating tree species in an Asian tropical forest. Integr. Zool. 2017, 12, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Hopper, G.; Smith, D.; Parrish, D. Germination and seedling growth of northern red oak–effects of stratification and pericarp removal. For. Sci. 1985, 31, 31–39. [Google Scholar]
- Liu, Y.; Hou, L.Y.; Li, Q.M. Effects of different mechanical treatments on Quercus variabilis, Q. wutaishanica and Q. robur acorn germination. iForest 2015, 8, 728–734. [Google Scholar] [CrossRef]
- Armstrong, D.P.; Westoby, M. Seedlings from large seeds tolerated defoliation better: A test using phylogenetically independent contrasts. Ecology 1993, 74, 1092–1100. [Google Scholar] [CrossRef]
- Dalling, J.W.; Harms, K.E.; Aizprúa, R. Seed damage tolerance and seedling resprouting ability of Prioria copaifera in Panamá. J. Trop. Ecol. 1997, 13, 481–490. [Google Scholar] [CrossRef]
- Branco, M.; Branco, C.; Merouani, H.; Almeida, M.H. Germination success, survival and seedling vigour of Quercus suber acorns in relation to insect damage. For. Ecol. Manag. 2002, 166, 159–164. [Google Scholar] [CrossRef]
- Giertych, M.J.; Suszka, J. Consequences of cutting off distal ends of cotyledons of Quercus robur acorns before sowing. Ann. For. Sci. 2011, 68, 433–442. [Google Scholar] [CrossRef]
- Fukumoto, H.; Kajimura, H. Effects of insect predation on hypocotyl survival and germination success of mature Quercus variabilis acorns. J. For. Res. 2000, 5, 31–34. [Google Scholar] [CrossRef]
- Dürr, C.; Dickie, J.B.; Yang, X.Y.; Pritchard, H.W. Ranges of critical temperature and water potential values for the germination of species worldwide: Contribution to a seed trait database. Agric. For. Meteorol. 2015, 200, 222–232. [Google Scholar] [CrossRef]
- Schroeder, W.; Walker, D. Effects of moisture content and storage temperatures on germination of Quercus macrocarpa acorns. J. Environ. Hort. 1987, 5, 22–24. [Google Scholar] [CrossRef]
- Gosling, P.G. The effect of drying Quercus robur acorns to different moisture contents, followed by storage, either with or without imbibition. Forestry 1989, 62, 41–50. [Google Scholar] [CrossRef]
- Connor, K.F.; Sowa, S. Effects of desiccation on the physiology and biochemistry of Quercus alba acorns. Tree Physiol. 2003, 23, 1147–1152. [Google Scholar] [CrossRef]
- Dey, D.C.; Jacobs, D.; McNabb, K.; Miller, G.; Baldwin, V.; Foster, G. Artificial regeneration of major oak (Quercus) species in the eastern United States—A review of the literature. For. Sci. 2008, 54, 77–106. [Google Scholar] [CrossRef]
- Mechergui, T.; Pardos, M.; Boussaidi, N.; Jacobs, D.F.; Catry, F.X. Problems and solutions to cork oak (Quercus suber L.) regeneration: A review. iForest 2023, 16, 10–22. [Google Scholar] [CrossRef]
- Romero-Rodríguez, M.C.; Archidona-Yuste, A.; Abril, N.; Gil-Serrano, A.M.; Meijón, M.; Jorrín-Novo, J.V. Germination and early seedling development in Quercus ilex recalcitrant and non-dormant seeds: Targeted transcriptional, hormonal, and sugar analysis. Front. Plant Sci. 2018, 9, 1508. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot. 2007, 101, 213–228. [Google Scholar] [CrossRef]
- Bonner, F.; Vozzo, J. Seed Biology and Technology of Quercus; US Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1987. [Google Scholar]
- Bonner, F.T. Water uptake and germination of red oak acorns. Bot. Gaz. 1968, 129, 83–85. [Google Scholar] [CrossRef]
- Amen, R.D. A model of seed dormancy. Bot. Rev. 1968, 34, 1–31. [Google Scholar] [CrossRef]
- Jones, L.; Brown, C.L. Cause of slow germination in Cherrybark and Northern Red Oak. Proc. Assoc. Off. Seed Anal. 1966, 56, 82–88. [Google Scholar]
- Abrams, M.D. Fire and the development of oak forests. Bioscience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Kremer, A.; Hipp, A.L. Oaks: An evolutionary success story. New Phytol. 2020, 226, 987–1011. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Daws, M.I.; Peng, L.L. Climate drives patterns of seed traits in Quercus species across China. New Phytol. 2022, 234, 1629–1638. [Google Scholar] [CrossRef]
- Löf, M.; Castro, J.; Engman, M.; Leverkus, A.B.; Madsen, P.; Reque, J.A.; Villalobos, A.; Gardiner, E.S. Tamm review: Direct seeding to restore oak (Quercus spp.) forests and woodlands. For. Ecol. Manag. 2019, 448, 474–489. [Google Scholar] [CrossRef]
- Amimi, N.; Dussert, S.; Vaissayre, V.; Ghouil, H.; Doulbeau, S.; Costantini, C.; Ammari, Y.; Joët, T. Variation in seed traits among mediterranean oaks in Tunisia and their ecological significance. Ann. Bot. 2020, 125, 891–904. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). International Rules for Seed Testing (2010); ISTA: Zürich, Switzerland, 2009. [Google Scholar]
- Tilki, F.; Alptekin, C.U. Variation in acorn characteristics in three provenances of Quercus aucheri Jaub. et Spach and provenance, temperature and storage effects on acorn germination. Seed Sci. Technol. 2005, 33, 441–447. [Google Scholar] [CrossRef]
- Peterson, J.K. Mechanisms involved in delayed germination of Quercus nigra L. seeds. Ann. Bot. 1983, 52, 81–92. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Aravind, J.; Vimala Devi, S.; Radhamani, J.; Jacob, S.R.; Srinivasan, K. Germinationmetrics: Seed Germination Indices and Curve Fitting, R Package Version 0.1.8. Available online: https://github.com/aravind-j/germinationmetricshttps://cran.r-project.org/package=germinationmetrics (accessed on 19 October 2023).
- Daws, M.I.; Crabtree, L.M.; Dalling, J.W.; Mullins, C.E.; Burslem, D.F.R.P. Germination responses to water potential in neotropical pioneers suggest large-seeded species take more risks. Ann. Bot. 2008, 102, 945–951. [Google Scholar] [CrossRef]
- Yan, X.F.; Zhou, L.B.; Si, B.B.; Sun, Y.; Gao, Y.F.; Wang, R.X. Stress effects of simulated drought by polyethylene glycol on the germination of Caragana korshinskii Kom. seeds under different temperature conditions. Acta Ecol. Sin. 2016, 36, 1989–1996. [Google Scholar] [CrossRef]
- Li, Z.X.; Pei, X.N.; Yin, S.P.; Lang, X.B.; Zhao, X.Y.; Qu, G.Z. Plant hormone treatments to alleviate the effects of salt stress on germination of Betula Platyphylla seeds. J. For. Res. 2019, 30, 779–787. [Google Scholar] [CrossRef]
- Arnold, C.Y. The determination and significance of the base temperature in a linear heat unit system. Proc. Am. Soc. Hortic. Sci. 1959, 74, 430–445. [Google Scholar]
- Guillemin, J.P.; Gardarin, A.; Granger, S.; Reibel, C.; Munier-Jolain, N.; Colbach, N. Assessing potential germination period of weeds with base temperatures and base water potentials. Weed Res. 2013, 53, 76–87. [Google Scholar] [CrossRef]
- Yi, X.F.; Zhang, M.M.; Bartlow, A.W.; Dong, Z. Incorporating cache management behavior into seed dispersal: The effect of pericarp removal on acorn germination. PLoS ONE 2014, 9, e92544. [Google Scholar] [CrossRef]
- Aniszewska, M.; Błuszkowska, U.; Zychowicz, W.; Brzózko, J. Impact of mechanical treatment of pedunculate oak (Quercus robur L.) seeds on germination time and seedling quality. J. For. Res. 2020, 25, 420–425. [Google Scholar] [CrossRef]
- Rawat, D.C.S.; Thapliyal, P.; Nautiyal, A.R. Pericarp delays germination in Quercus glauca seeds. J. Trop. For. Sci. 1998, 10, 472–477. [Google Scholar]
- Liu, Y.; Liu, G.Q.; Li, Q.M.; Liu, Y.; Hou, L.Y.; Li, G.L. Influence of pericarp, cotyledon and inhibitory substances on sharp tooth oak (Quercus aliena var. acuteserrata) germination. PLoS ONE 2012, 7, e47682. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.G.; Yi, X.F.; Yang, Y.Q.; Liu, W.J. Acorn germination and seedling survival of Q. variabilis: Effects of cotyledon excision. Ann. For. Sci. 2010, 67, 711. [Google Scholar] [CrossRef]
- Yi, X.F.; Bartlow, A.W.; Curtis, R.; Agosta, S.J.; Steele, M.A. Responses of seedling growth and survival to post-germination cotyledon removal: An investigation among seven oak species. J. Ecol. 2019, 107, 1817–1827. [Google Scholar] [CrossRef]
- Yi, X.F.; Wang, Z.Y.; Liu, C.Q.; Liu, G.Q.; Zhang, M.M. Acorn cotyledons are larger than their seedlings’ need: Evidence from artificial cutting experiments. Sci. Rep. 2015, 5, 8112. [Google Scholar] [CrossRef]
- Tilki, F. Influence of acorn size and storage duration on moisture content, germination and survival of Quercus petraea (Mattuschka). J. Environ. Biol. 2010, 31, 325–328. [Google Scholar] [PubMed]
- Elwell, A.L.; Gronwall, D.S.; Miller, N.D.; Spalding, E.P.; Durham Brooks, T.L. Separating parental environment from seed size effects on next generation growth and development in Arabidopsis. Plant Cell Environ. 2011, 34, 291–301. [Google Scholar] [CrossRef]
- Pritchard, H.W.; Manger, K.R. Quantal response of fruit and seed germination rate in Quercus robur L. and Castanea sativa Mill, to constant temperatures and photon dose. J. Exp. Bot. 1990, 41, 1549–1557. [Google Scholar] [CrossRef]
- Xia, K.; Daws, M.I.; Zhou, Z.K.; Pritchard, H.W. Habitat-linked temperature requirements for fruit germination in Quercus species: A comparative study of Quercus subgenus Cyclobalanopsis (Asian evergreen oaks) and Quercus subgenus Quercus. S. Afr. J. Bot. 2015, 100, 108–113. [Google Scholar] [CrossRef]
- Muñiz-Castro, M.A.; Williams-Linera, G.; Benítez-Malvido, J. Restoring montane cloud forest: Establishment of three Fagaceae species in the old fields of central Veracruz, Mexico. Restor. Ecol. 2015, 23, 26–33. [Google Scholar] [CrossRef]
- Valdés-Rodríguez, O.A.; Cruz, Y.G.-D.L.; Frey, B.R. Survival and growth of three endangered oak species in a Mexican montane cloud forest. Ann. For. Sci. 2017, 60, 89–100. [Google Scholar] [CrossRef]
- Pandey, R.; Bargali, K.; Bargali, S.S. Does seed size affect water stress tolerance in Quercus Leucotrichophora a. Camus at germination and early seedling growth stage? Biodivers. Int. J. 2017, 1, 24–30. [Google Scholar] [CrossRef]
- Tozer, M.G.; Ooi, M.K.J. Humidity-regulated dormancy onset in the Fabaceae: A conceptual model and its ecological implications for the australian wattle Acacia saligna. Ann. Bot. 2014, 114, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, C.; Lazzaro, L.; Calamassi, R.; Fico, G.; Foggi, B.; Mariotti Lippi, M. Induced water stress affects seed germination response and root anatomy in Robinia pseudoacacia (Fabaceae). Trees-Struct. Funct. 2019, 33, 1627–1638. [Google Scholar] [CrossRef]
- Tuyet, D. Characteristics of karst ecosystems of Vietnam and their vulnerability to human impact. Acta Geol. Sin.-Engl. Ed. 2010, 75, 325–329. [Google Scholar] [CrossRef]
- Tang, J.W.; Lü, X.T.; Yin, J.X.; Qi, J.F. Diversity, composition and physical structure of tropical forest over limestone in Xishuangbanna, South-West China. J. Trop. For. Sci. 2011, 23, 425–433. [Google Scholar]
- Fahey, T.J.; Sherman, R.E.; Tanner, E.V.J. Tropical montane cloud forest: Environmental drivers of vegetation structure and ecosystem function. J. Trop. Ecol. 2016, 32, 355–367. [Google Scholar] [CrossRef]
- Nie, Y.P.; Chen, H.S.; Wang, K.L.; Yang, J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of southwest China. J. Hydrol. 2012, 420, 264–274. [Google Scholar] [CrossRef]
- Lin, W.; Wen, C.; Zhou, W.; Huang, G. Drought in southwest China: A review. Atmos. Ocean. Sci. Lett. 2015, 8, 339–344. [Google Scholar] [CrossRef]
- Feng, W.J.; Leung, M.Y.T.; Wang, D.X.; Zhou, W.; Zhang, O.Y.W. An extreme drought over South China in 2020/21 concurrent with an unprecedented warm northwest Pacific and La Niña. Adv. Atmos. Sci. 2022, 39, 1637–1649. [Google Scholar] [CrossRef]
- You, Q.L.; Jiang, Z.H.; Yue, X.; Guo, W.D.; Liu, Y.G.; Cao, J.; Li, W.; Wu, F.Y.; Cai, Z.Y.; Zhu, H.H.; et al. Recent frontiers of climate changes in East Asia at global warming of 1.5 °C and 2 °C. NPJ Clim. Atmos. Sci. 2022, 5, 1–17. [Google Scholar] [CrossRef]
- Rathore, P.; Roy, A.; Karnatak, H. Assessing the vulnerability of oak (Quercus) forest ecosystems under projected climate and land use land cover changes in western Himalaya. Biodivers. Conserv. 2019, 28, 2275–2294. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Meng, Q.X.; Wang, Y.Z.; Zhang, X.L.; Wang, W. Climate change-induced migration patterns and extinction risks of Theaceae species in China. Ecol. Evol. 2020, 10, 4352–4361. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Jiang, X.-L.; Guo, K.Q.; Byrne, A.; Deng, M. Climate change impacts the distribution of Quercus section Cyclobalanopsis (Fagaceae), a keystone lineage in East Asian evergreen broadleaved forests. Plant Divers. 2023, 45, 552–568. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The natural habitats of Q. marlipoensis in Xia-Jin-Chang village, Lao-Jun-Shan National Nature Reserve, Malipo County, southeastern Yunnan, China. (a) The landscape of tropical montane cloud forests (TMCF); (b) A close view of the community showing the limestone rocky soil outcrops.
Figure 1.
The natural habitats of Q. marlipoensis in Xia-Jin-Chang village, Lao-Jun-Shan National Nature Reserve, Malipo County, southeastern Yunnan, China. (a) The landscape of tropical montane cloud forests (TMCF); (b) A close view of the community showing the limestone rocky soil outcrops.

Figure 2.
Acorn and seed germination morphology of Q. marlipoensis. Scale bar = 1.0 cm. (a,b) Mature acorn with cupule (a) and without cupule (b); CL indicates the cupule length measurement; L and W represent the acorn length and width, respectively; CSD indicates the cupule scar. (c) Embryo position in the seed. (d,e) Radicle emergence at day 2 of germination, showing that embryo position varies from near the remains of the style (d) to beside the remains of the style (e). (f) Seed after two weeks of germination, an elongated radicle and emergence of the plumule. St: remains of the style; Cu: cupule; Co: cotyledon; EoA: embryo axis; CoP: cotyledon petiole; Plu: plumule; Ra: radicle.
Figure 2.
Acorn and seed germination morphology of Q. marlipoensis. Scale bar = 1.0 cm. (a,b) Mature acorn with cupule (a) and without cupule (b); CL indicates the cupule length measurement; L and W represent the acorn length and width, respectively; CSD indicates the cupule scar. (c) Embryo position in the seed. (d,e) Radicle emergence at day 2 of germination, showing that embryo position varies from near the remains of the style (d) to beside the remains of the style (e). (f) Seed after two weeks of germination, an elongated radicle and emergence of the plumule. St: remains of the style; Cu: cupule; Co: cotyledon; EoA: embryo axis; CoP: cotyledon petiole; Plu: plumule; Ra: radicle.

Figure 3.
Seed germination patterns of Q. marlipoensis under different pericarp removal and cotyledon cut-off treatments. (a) Fitted germination curves, where CK represents seeds with pericarp, RS represents removal of cup scar, RP removal of the pericarp, TC and OC respectively represent seeds with pericarp removal and cutting off 1/3 of cotyledons and 2/3 of the cotyledon; (b) ; (c) germination value (GV); (d) germination index (GI). Means followed by the same letter are not significantly different at p = 0.05 (Tukey HSD test).
Figure 3.
Seed germination patterns of Q. marlipoensis under different pericarp removal and cotyledon cut-off treatments. (a) Fitted germination curves, where CK represents seeds with pericarp, RS represents removal of cup scar, RP removal of the pericarp, TC and OC respectively represent seeds with pericarp removal and cutting off 1/3 of cotyledons and 2/3 of the cotyledon; (b) ; (c) germination value (GV); (d) germination index (GI). Means followed by the same letter are not significantly different at p = 0.05 (Tukey HSD test).

Figure 4.
Seed germination patterns of Q. marlipoensis at different temperatures. (a) Germination curves; (b) ; (c) germination value (GV); (d) germination index (GI). Means followed by the same letter are not significantly different at p = 0.05 (Tukey HSD test).
Figure 4.
Seed germination patterns of Q. marlipoensis at different temperatures. (a) Germination curves; (b) ; (c) germination value (GV); (d) germination index (GI). Means followed by the same letter are not significantly different at p = 0.05 (Tukey HSD test).

Figure 5.
Effect of temperature (x) on the germination rate (GR, y) of Q. marlipoensis. The germination base temperature (Tb) was calculated as the x-intercept of the linear regression (y = 0.0224x − 0.0813, R2 = 0.9735). GR was calculated as the inverse of .
Figure 5.
Effect of temperature (x) on the germination rate (GR, y) of Q. marlipoensis. The germination base temperature (Tb) was calculated as the x-intercept of the linear regression (y = 0.0224x − 0.0813, R2 = 0.9735). GR was calculated as the inverse of .

Figure 6.
Seed germination pattern of Q. marlipoensis under different water potentials. (a) Fitted germination curves; (b) ; (c) germination value (GV); (d) germination index (GI). Means followed by the same letter are not significantly different at p = 0.05 (Tukey HSD test).
Figure 6.
Seed germination pattern of Q. marlipoensis under different water potentials. (a) Fitted germination curves; (b) ; (c) germination value (GV); (d) germination index (GI). Means followed by the same letter are not significantly different at p = 0.05 (Tukey HSD test).

Figure 7.
Effect of water potential (ψ) on the final germination percentage (%) of Q. marliponesis. The linear equation (y = 125x + 127.33, R2 = 0.9855) fit the data well when data points for 0 MPa treatment (which had the same germination percentage as −0.2 MPa treatment) were omitted.
Figure 7.
Effect of water potential (ψ) on the final germination percentage (%) of Q. marliponesis. The linear equation (y = 125x + 127.33, R2 = 0.9855) fit the data well when data points for 0 MPa treatment (which had the same germination percentage as −0.2 MPa treatment) were omitted.

Figure 8.
Effect of water potential (ψ) on the germination rate (1/days) of Q. marliponesis. The linear equation (y = 0.2707x + 0.33, R2 = 0.7992) was fitted to the data, and the x intercept was calculated to estimate the base water potential (ψb).
Figure 8.
Effect of water potential (ψ) on the germination rate (1/days) of Q. marliponesis. The linear equation (y = 0.2707x + 0.33, R2 = 0.7992) was fitted to the data, and the x intercept was calculated to estimate the base water potential (ψb).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, L.; Tu, Y.; Li, Q.; Deng, M. Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications. Forests 2024, 15, 235. https://doi.org/10.3390/f15020235
AMA Style
Liu L, Tu Y, Li Q, Deng M. Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications. Forests. 2024; 15(2):235. https://doi.org/10.3390/f15020235
Chicago/Turabian StyleLiu, Luting, Yu Tu, Qiansheng Li, and Min Deng. 2024. "Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications" Forests 15, no. 2: 235. https://doi.org/10.3390/f15020235
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.