
Comparative Genomic Analysis of a Thermophilic Protease-Producing Strain Geobacillus stearothermophilus H6
by
 ,
,
Ruilin Lai
1,2,
Min Lin
1,2,
Yongliang Yan
2,
Shijie Jiang
1,
Zhengfu Zhou
2,* and
Jin Wang
1,2,* 1
College of Life Science and Engineering, Southwest University of Science and Technology, Mianyang 621000, China
2
Key Laboratory of Agricultural Microbiome (MARA), Biotechnology Research Institute, Chinese Academy of Agricultural Sciences, Beijing 100081, China
*
Authors to whom correspondence should be addressed.
Genes 2023, 14(2), 466; https://doi.org/10.3390/genes14020466
Submission received: 6 January 2023
/
Revised: 6 February 2023
/
Accepted: 9 February 2023
/
Published: 11 February 2023
(This article belongs to the Section Microbial Genetics and Genomics)
Abstract
:The genus Geobacillus comprises thermophilic gram-positive bacteria which are widely distributed, and their ability to withstand high temperatures makes them suitable for various applications in biotechnology and industrial production. Geobacillus stearothermophilus H6 is an extremely thermophilic Geobacillus strain isolated from hyperthermophilic compost at 80 °C. Through whole-genome sequencing and genome annotation analysis of the strain, the gene functions of G. stearothermophilus H6 were predicted and the thermophilic enzyme in the strain was mined. The G. stearothermophilus H6 draft genome consisted of 3,054,993 bp, with a genome GC content of 51.66%, and it was predicted to contain 3750 coding genes. The analysis showed that strain H6 contained a variety of enzyme-coding genes, including protease, glycoside hydrolase, xylanase, amylase and lipase genes. A skimmed milk plate experiment showed that G. stearothermophilus H6 could produce extracellular protease that functioned at 60 °C, and the genome predictions included 18 secreted proteases with signal peptides. By analyzing the sequence of the strain genome, a protease gene gs-sp1 was successfully screened. The gene sequence was analyzed and heterologously expressed, and the protease was successfully expressed in Escherichia coli. These results could provide a theoretical basis for the development and application of industrial strains.
1. Introduction
Thermophilic microorganisms are microbes that can grow at 41~122 °C, and their optimal growth temperature is 45~80 °C. Thermophilic ecological environments are distributed at sites including volcanoes, geothermal areas (terrestrial, underground and marine hot springs), compost, oil reservoirs and other extreme high-temperature areas on Earth [1]. Many thermophilic microorganisms play an important role in biotechnology and have major commercial applications in industrial production [2]. For example, they can produce a variety of thermostable enzymes [3], generate biofuels by degrading agricultural wastes [4], and they show a special leaching capacity for certain minerals [5] and a bioremediation capacity [6].
Geobacillus was separated from Bacillus in 2001 by T.N. Nazina and others as a new genus of bacteria [7]. It is mainly composed of aerobic or facultative anaerobic bacteria with the ability to form endophytic spores and is a typical thermophilic microbial group [8]. The genus is widely distributed and is found in natural environments such as oil fields, hot springs, volcanic vents, dairy plants, food processing, compost and other high-temperature environments [9]. The ability of Geobacillus to grow at high temperatures makes it suitable for various applications in biotechnology and industrial production. R.E. Cripps and others used metabolic engineering to transform two Geobacillus thermophilic bacteria to obtain a strain that can efficiently produce ethanol [10]. Geobacillus species can secrete extracellular polysaccharides and bacteriocins and show bioremediation properties [11]. They can be a source of many thermostable enzymes, such as xylanase, lipase, protease and amylase [12,13,14,15,16]. These thermostable enzymes play an important role in the commercial production of detergent, the brewing industry and the food industry [17].
Proteases are enzymes that catalyze the cleavage of protein peptide bonds and are widely found in animals, plants and microorganisms. They present important industrial applications and are widely used in the detergent, leather, medicine and other industries. Their output accounts for more than 65% of the enzyme preparation market [18]. Microbial proteases are the most important source of commercial proteases. Bacillus species can secrete a variety of proteases; this genus is the most important source of commercial production and is of great significance in commercial protease production [19]. Some Geobacillus strains have been proven to be capable of producing proteases [20]. Proteases isolated from Geobacillus can also adapt to high-temperature environments and can be used in a variety of industrial production environments [21].
Geobacillus is an important source of thermostable enzymes and thermophilic proteases. This study analyzed a thermophilic bacterial strain, G. stearothermophilus H6, isolated from hyperthermophilic compost in Beijing. After sequencing and analyzing the whole genome of G. stearothermophilus H6, we further analyzed its gene functions using bioinformatics tools.
2. Materials and Methods
2.1. Sample Collection, Strain Isolation and Culture
The samples used in this study were hyperthermophilic composting soil samples from Beijing. To isolate thermophilic bacteria, LB and R2A liquid culture media were used, and 5 g soil samples were added to 50 mL liquid culture media and were then incubated in a water bath at 80 °C for 48 h. Then, 500 μL of enrichment solution was added to 50 mL of new liquid medium for further culture in an 80 °C water bath, and the enrichment solution was collected three times. The enrichment solution was diluted to 10−1, 10−2, 10−3 and 10−4 with ddH2O, and the dilutions were spread on corresponding agar plates and cultured in an 80 °C incubator. A colony was selected for 16S rRNA sequencing, and the bacteria were preserved. The strain was isolated from an R2A agar plate.
2.2. Genomic DNA Extraction and Sequencing of G. stearothermophilus H6
G. stearothermophilus H6 was cultured in LB medium at 60 °C overnight. The bacterial solution was centrifuged at 4000 r/min at 4 °C for 10 min, the supernatant was discarded, and the bacterial cells were collected. Total bacterial DNA was extracted according to the operating instructions of a bacterial genomic DNA isolation kit (Mei5 Biotechnology Co. Ltd., Beijing, China), and the concentration and quality of the DNA were assessed with a NanoDrop 2500 system (OD260/OD280 = 1.8–2.0, ≥10 µg). The total DNA of the extracted samples was stored on dry ice and sent to Biomarker Technologies to complete the sequencing analysis.
2.3. Phylogenetic Tree and Comparative Genomic Analysis of 16S rRNA of G. stearothermophilus H6
Genomic DNA was extracted and purified with a commercial bacterial genomic DNA isolation kit. The 16S rRNA gene was amplified with the universal bacterial primers 27F and 1492R. Preliminary sequence analysis of the 16S rRNA gene was conducted using the NCBI database, and strains with high homology in the NCBI database were selected for phylogenetic tree analysis. The corresponding phylogenetic tree was constructed by using MEGA 6.0 [22] software and the maximum likelihood method (ML). The evolutionary tree was constructed based on the bootstrap values of 1000 repeats.
Five homologous strains were selected, their basic information was compared with that of G. stearothermophilus H6, and the average nucleotide identity (ANI) was calculated (www.ezbiocloud.net/tools/ani (accessed on 15 October 2022)). Using the Mauvealigner algorithm of Mauve 2.4.0 [23], the whole-genome sequence of G. stearothermophilus H6 and the whole-genome sequence of the reference located close to the source strain were analyzed for collinearity.
2.4. Detection of Protease Activity of G. stearothermophilus H6
G. stearothermophilus H6 bacterial solution (2.5 μL) was cultured to the middle logarithmic phase spotted onto a skim milk plate, and the results were compared with Bacillus velezensis, Bacillus subtilis and E. coli BL21 (DE3). The four strains were cultured at both 37 °C and 60 °C. After different culture times, the transparent circles that appeared were observed to preliminarily judge the protease production ability of the strains.
2.5. Whole-Genome Sequencing and Analysis
The original genome data were filtered and more than 2 kb of reads were retained. Hifiasm v0.16.0 2 [24] software was used to assemble the filtered reads. Circulator v1.5.5 software was used to cyclize and adjust starting sites. Pilon v1.22 software was used to further correct errors using second-generation data, and a genome with higher accuracy was obtained for subsequent analysis. Prodigal v2.6.3 [25] was used to predict coding sequences (CDSs) in the genome of the strain, and genome information obtained by assembly and prediction, such as information on tRNAs, rRNAs, repeat sequences, GC contents and gene functions, was used to draw a circular genome map with the software Circos v0.66 [26].
We used software to predict repeat sequences, rRNAs, tRNAs, CRISPRs, and gene islands in the genome. Gene function annotation was mainly based on protein sequence comparison, performed by comparing the gene sequences in each database. The predicted gene sequences were compared with eggNOG, KEGG, Swiss-Prot, TrEMBL, Nr, GO, Pfam and other databases to obtain gene function annotation results.
2.6. Analysis of G. stearothermophilus H6 Protease
General databases such as eggNOG, KEGG, Swiss-Prot, TrEMBL, Nr, GO, and Pfam were used to predict the distribution of proteases in strain G. stearothermophilus H6, and the software SignalP v4.0 [27] was used to predict protein signal peptides and specific signal peptide excision sites for further analysis. At the same time, TMHMM v2.0 [28] was used to predict the transmembrane domains of the protease. The similarity of the protease gene between G. stearothermophilus H6 and the other two homologous strains was compared with gene sequences in the NCBI database.
2.7. Heterologous Expression of the G. stearothermophilus H6 Protease Gene in E. coli
After screening the protease genes of G. stearothermophilus H6, the GE001730 gene was selected and named gs-sp1. The thermophilic protease gene fragment was then inserted into the pET22b vector digested by Hind III and Xho I using seamless cloning and recombination technology.
PCR technology was used to amplify the gene (95 °C for 5 min, 1 cycle; 95 °C for 30 s, 58 °C for 30 s and 72 °C for 30 s, 33 cycles; 72 °C for 10 min, 1 cycle) using the following primers: F (5′-tcgagctccgtcgacaagcttATCTTCCCTCATATGAGTATAGGA-3′) and R (5′- gtggtggtggtggtgctcgagGCGCTGCAGCAGTTGCTC-3′). The PCR products were purified with a gel recovery kit (Mei5 Biotechnology Co. Ltd., Beijing, China). The PCR products were cloned into the expression plasmid pET-22b using the ClonExpress Ultra One Step Cloning Kit (Vazyme, Ninjing, China) and then transformed into E. coli BL21 (DE3) cells for further screening and verification.
The constructed E. coli BL21/pET22b/gs-sp1 strain was selected on Amp-resistant plates and cultured overnight at 37 °C. A single colony was picked and cultured overnight at 220 rpm in 20 mL liquid medium at 37 °C. Then, 2% of the volume was transferred to 20 mL LB medium, and culture was continued until reaching OD600 = 0.5~0.7. Thereafter, 0.1 mM IPTG was used to induce protein expression at 37 °C for 4 hours, and a 5 μL aliquot was spotted on a skim milk plate. The wild-type E. coli BL21 and E. coli BL21/pET22b strains were used as controls, and the transparent circles produced by the bacteria were observed.
3. Results
3.1. Phylogenetic Analysis of G. stearothermophilus H6 16S rRNA
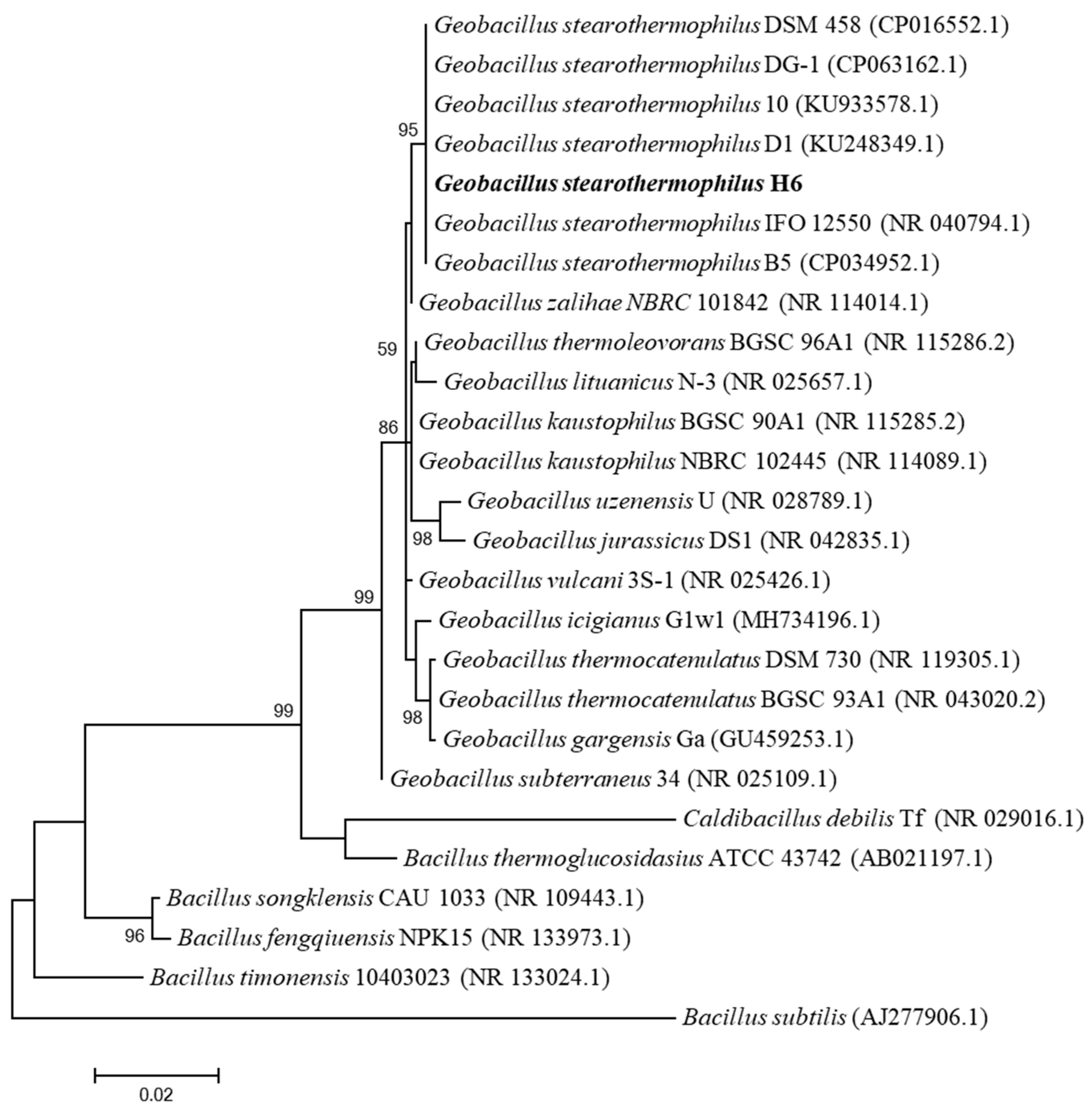
To clarify the taxonomic position of G. stearothermophilus H6, 16S rRNA analysis was used. G. stearothermophilus H6 is a thermophilic bacterium obtained from hyperthermophilic composting soil by culture in R2A medium and screening based on 80 °C culture. According to 16S rRNA gene sequence analysis, the strain is a bacterium of the genus Geobacillus showing the highest homology with G. stearothermophilus B5; therefore, the strain was named G. stearothermophilus H6. The 16S rRNA gene of G. stearothermophilus H6 presented the highest similarity with G. stearothermophilus B5 (99.97%), G. stearothermophilus D1 (99.97%), G. stearothermophilus DSM 458 (99.93%), G. stearothermophilus DG-1 (99.93%), G. stearothermophilus IFO 12550 (99.79%) and G. stearothermophilus 10 (99.78%). A phylogenetic analysis of the 16S rRNA gene with a tree constructed based on the maximum likelihood (ML) method showed that G. stearothermophilus H6 was a member of the genus Geobacillus (Figure 1).
3.2. Comparative Genome Analysis of G. stearothermophilus H6
To compare the differences between G. stearothermophilus H6 and the five most closely related strains (G. stearothermophilus DSM 458, G. stearothermophilus 10, G. stearothermophilus DG-1, G. stearothermophilus D1 and G. stearothermophilus B5), the genomic characteristics of the six strains were statistically analyzed, and the results are shown in Table 1. The average nucleotide homology (ANI) value indicates the similarity between the sequences of the conserved regions of two genomes and allows the genetic relationships between them to be analyzed. According to the whole-genome information of these six strains, the ANI of the genome of G. stearothermophilus D1 was highest (97.65%), showing good homology (Table 1). The genome sizes and GC contents of the six strains were similar, with genome sizes ranging from 2.97–3.65 Mb and GC contents from 51.66–52.6%.
Based on 16S rRNA phylogenetic tree analysis, the genomes of G. stearothermophilus B5 and G. stearothermophilus 10, which show close homologous relationships with G. stearothermophilus H6, were selected. The software Mauve 2.4.0 was used to perform genome synteny analysis and quickly analyze whether large-segment sequence rearrangements existed between genomes. The squares with similar colors represent highly homologous assembly regions of the two genomes. Figure 2 shows that G. stearothermophilus H6 and G. stearothermophilus 10 had poor synteny, with many gene rearrangements, such as insertions, deletions, inversions and translocations, between them. For example, compared with G. stearothermophilus 10, there was a gene deletion at 1,177,537–1,516,081 bp in G. stearothermophilus H6, and an inversion occurred at 1,604,138–1,649,959 bp in G. stearothermophilus H6. G. stearothermophilus H6 presented good synteny with G. stearothermophilus B5, but there were also some gene rearrangements between them, such as deletions and inversions. For example, compared with G. stearothermophilus B5, a gene inversion occurred at 1,786,619–1,868,252 bp in G. stearothermophilus H6, and a deletion occurred at 2,464,988–2,718,550 bp in G. stearothermophilus H6 (Figure 2).
3.3. Detection of Protease Activity in G. stearothermophilus H6
The protease hydrolysis activity of G. stearothermophilus H6 was observed by the skimmed milk plate method and compared with that of B. velezensis, B. subtilis and E. coli BL21(DE3). The four strains were cultured at both 37 °C and 60 °C, and the results showed that G. stearothermophilus H6 could produce transparent circles that became larger with increasing culture time (Figure 3). When cultured at 37 °C, B. velezensis, B. subtilis and G. stearothermophilus H6 produced transparent circles, with B. velezensis producing the strongest degradation results. When cultured at 60 °C, G. stearothermophilus H6 produced the largest transparent circle, and the other three strains did not produce a transparent circle. Thus, G. stearothermophilus H6 produces extracellular proteases that function under high temperature and show a good effect.
3.4. Overview of the Genome Assembly and Whole Genome of G. stearothermophilus H6
Based on the specificity of the high-temperature tolerance of G. stearothermophilus H6, the whole genome of the strain was sequenced to further explore the specific coding genes associated with its high-temperature tolerance. Gene prediction was carried out with Prodigal v2.6.3 software, and a genome completion map was obtained through assembly and construction. The size of the genome sequence of G. stearothermophilus H6 was 3,054,993 bp, and the average GC content was 51.66%. It was predicted that there were 3750 coding genes with an average length of 814 bp in the genome. The results of noncoding RNA prediction showed that the genome contained 30 rRNAs and 91 tRNAs and had 17 CRISPR regions and 24 gene islands (Table 2).
Based on the genome information obtained by assembly and prediction, such as information on tRNAs, rRNAs, repeat sequences, GC contents and gene functions, Circos v0.66 software was used to obtain the circular genome map (Figure 4).
The amino acid sequence of G. stearothermophilus H6 was compared with the Nr database, and the corresponding species information was obtained from the annotation database. Through BLAST searches comparing the protein sequences of genes with the Nr database, the most similar sequences in the Nr database could be found. The corresponding annotation information of the sequences was the annotation information of the corresponding gene in the genome sequence. A total of 3699 genes were annotated.
BLAST comparisons of the protein-encoding gene sequences of the whole genome were performed against the eggNOG database, and a database of the results was generated. The database was frequently used to classify and annotate genes of newly sequenced genomes. The annotation information and classification information in the genome corresponded to the gene sequences of the sequenced genome. A total of 3186 genes were annotated in the database.
The amino acid sequences of G. stearothermophilus H6 were subjected to BLAST searches in the KEGG database to assemble databases of the biological pathways related to diseases, drugs and chemical substances in the genome. The strain has 1798 genes in the KEGG database.
The prediction results were annotated in the GO database. The number of genes dominated by GO functional classifications mainly included the highest-level functional nodes: cellular component, molecular function and biological process. A total of 2686 genes were predicted in the database (Figure 5).
3.5. Analysis of G. stearothermophilus H6 Protease
The predicted gene sequences were compared with eggNOG, GO, KEGG, Nr, Pfam, Swiss-Prot, TrEMBL and other general databases to obtain gene functional annotation results. Approximately 141 proteases were predicted, which accounted for approximately 3% of the total encoded proteins. The predicted proteases mainly consisted of serine proteases and metalloproteinases and a few cysteine proteases, aspartic proteases and threonine proteases, accounting for 19%, 27%, 1%, 2% and 1% of the predicted proteases, respectively. Other proteases could not be characterized (Table 3).
Secretory proteins are proteins secreted from the cells of living microorganisms. Through the prediction and analysis of signal peptides and secretory proteins in the genome, approximately 161 proteins with signal peptides were identified, which could form secretory proteins. The predicted proteases included 18 secretory proteins with signal peptides, and the serine and metalloproteinases included 5 secretory proteins with signal peptides.
The 18 secreted proteases were analyzed and compared with the proteases of the G. stearothermophilus B5 and G. stearothermophilus 10 genomes. Among them, GE000377 was not predicted to show homologous proteins in G. stearothermophilus B5 but showed higher homology with the proteins of G. stearothermophilus 10 (99.24%); GE003445 was not predicted to show homologous proteins in G. stearothermophilus 10 but presented homologous proteins with G. stearothermophilus B5 (76.00%). The homologous proteins encoded by the GE002130, GE003446, GE003450 and GE003532 genes in the two homologous strains exhibited low similarity. These proteins may be encoded by genes unique to G. stearothermophilus H6. Among the 18 proteases with signal peptides, most of the signal peptides were removed by the type I signal peptidase SP (Sec/SPI), and only the signal peptides of the GE000438 and GE002405 genes were removed by the type II signal peptidase LIPO (Sec/SPII) (Table 4).
The prediction and analysis of transmembrane helix structures in the genome indicated that approximately 841 proteins had transmembrane helix structures. Among these proteins, 46 proteases had transmembrane helix structures, while serine proteases, metalloproteinases and aspartic proteases had 12, 16 and 3 transmembrane helix structures, respectively.
3.6. Construction of GS-SP1 Protein Expression Vector and Verification of Secreted Proteases
The GS-SP1 protease gene was cloned into the pET22b expression vector, which contains the signal peptide pelB upstream of multiple cloning sites. Then, the constructed pET22b/gs-sp1 plasmid was transformed into E. coli BL21 (DE3) (Figure 6a), and wild-type E. coli BL21 and E. coli BL21/pET22b were used as controls. Five microliters of bacterial liquid culture was spotted onto a skimmed milk plate, and culture was performed at 37 °C for 24 h to observe transparent circle development. The results showed that E. coli BL21/pET22b/gs-sp1 could produce a transparent circle when induced by 0.1 mM IPTG, while the other two strains could not (Figure 6b). These results indicated that the GS-SP1 protein showed protease activity, and further investigation of this protein will be important for the exploitation of thermophilic proteases.
4. Discussion
In this study, we isolated a strain of Geobacillus from hyperthermophilic compost and named it G. stearothermophilus H6. Thereafter, 16S rRNA sequence analysis and comparisons showed that the strain presented the highest consistency with G. stearothermophilus B5 (99.97%). The skimmed milk plate experiment showed that G. stearothermophilus H6 could produce extracellular proteases with enzyme activity at high temperature. Whole-genome sequencing and genome annotation analysis revealed that G. stearothermophilus H6 produces a variety of enzymes with biotechnological significance, such as proteases, amylases and lipases. Thus, it may be an important source of thermophilic enzymes and has important research value.
Geobacillus is a genus of thermophilic Gram-positive bacteria belonging to Bacillaceae, including denitrifying bacteria, facultative anaerobes and obligate aerobic bacteria, which can grow at 45–80 °C [29]. The members of the genus can form endophytic spores, which can diffuse through global atmospheric circulation [30] and are widely distributed in environments such as in soil, hot springs, dairy plants or other food processing plants [8]. The chromosomes and plasmids of Geobacillus species exhibit significant genetic diversity. Bezuidt et al. [31] analyzed the pangenome of 29 genome sequences of Geobacillus sp. and found that the core genome was relatively small, mainly consisting of Bacillus-related genes, indicating that these bacteria originated from an ancestor of Bacillus; it contained a large number of dispensable genomes, which showed that Geobacillus spp. can achieve extensive genomic diversity through horizontal gene transfer, which is the key mechanism whereby Geobacillus spp. adapt to different environmental niches. For example, G. stearothermophilus obtained the lac operon through horizontal gene transfer, enabling it to survive in dairy products [32]. This feature provides a new way to produce thermostable enzymes for industrial use through the evolution of thermoadaptive directed enzymes, thus expanding the biotechnological application of Geobacillus spp. For example, G. kaustophilus HTA42 producing thermostable variants of rRNA methyltransferase was generated through thermal-adaptation-directed evolution [33]. G. stearothermophilus H6 shows potential as a host for whole-cell applications and a biological tool in evolutionary engineering.
The characteristics and distribution of proteases in the G. stearothermophilus H6 genome indicate that its proteases consist of serine proteases, metalloproteinases, cysteine proteases and aspartic proteases, and the proportion of proteases from PDB entries distributed in all Bacillus genomes is similar [34,35]. The exploration of the proteases of this strain may provide knowledge for the discovery of new potential proteases with various potential industrial applications. By analyzing G. stearothermophilus H6 genome proteases, we screened 18 proteases with signal peptides, selected the gs-sp1 gene for heterologous expression, and successfully expressed the protease in E. coli BL21. Compared with the homologous strains G. stearothermophilus B5 and G. stearothermophilus 10, the gs-sp1 protease had higher homology. In addition, G. stearothermophilus H6 had unique protease genes, like the GE003532 gene, which had low similarity among homologous strains. The G. stearothermophilus H6 genome also contains a variety of other enzymes which may have high thermal stability and broad application prospects in biotechnology applications. The ability of the thermophilic Geobacillus microorganisms to grow under high temperatures makes them a valuable resource for the development of new biotechnological applications [36]. They can be a source of many thermophilic enzymes, such as proteases, xylanases, amylases and lipases, and can be used for the synthesis of biofuels, such as bioethanol, isobutanol, biogas and biodiesel [37,38,39].
Currently, many species of Geobacillus are used to produce thermophilic enzymes either naturally or through the introduction of genetic engineering. Thermophilic enzymes are mainly used in biotechnology [40], including the food industry, detergent industry, leather industry, and medical industry [41]. The proteases isolated from Geobacillus sp. are extremely heat-resistant and can be used to improve the biodegradation of sewage sludge [42]. The optimum conditions for Geobacillus sp. YMTC 1049 to produce serine protease are 85 °C and pH 7.5 [43]. Due to the decreasing reserves of natural fossil fuels, the world needs to produce biofuels to develop alternative energy sources or fuels [8]. Geobacillus is used to biodegrade agricultural and industrial residues such as beet, soybean, barley, sugarcane, corn, sorghum and other biomass and produce biofuels through modern processes [44]. G. stearothermophilus has been employed to produce bioethanol using sucrose as a carbon source at approximately 70 °C, and the product yield is the same as that of yeast [45]. When Geobacillus strain AT1 is added to methanogenic sludge, it could effectively improve biogas production due to protease activity [46].
G. stearothermophilus is an important species of Geobacillus that can be employed as a source of various thermophilic enzymes and is widely used in a variety of biotechnology industries. Thermophilic enzymes produced by G. stearothermophilus SR74 α-amylase can be used in the papermaking, food and other industries [15]. G. stearothermophilus strain RM is used for the mass production of α-glucosidase at high temperature [47]. G. stearothermophilus PS11 can produce thermophilic and stable lipase under high temperature and alkali conditions, which is used for the production of biodiesel [17]. A protease cloned from G. stearothermophilus strain B-1172 has been used in the detergent and many other industries due to its catalytic domain and good activity [20]. G. stearothermophilus H6 isolated from hyperthermophilic compost can produce a protease with good activity at high temperature, which has broad application prospects in biotechnology applications.
Author Contributions
Conceptualization, R.L., Y.Y. and J.W.; data curation, R.L. and Y.Y.; formal analysis, R.L., Y.Y. and Z.Z.; funding acquisition, M.L. and J.W.; methodology, Z.Z. and J.W.; project administration, M.L.; writing—original draft, R.L.; writing—review and editing, S.J., Z.Z. and J.W. All authors will be informed about each step of manuscript processing, including submission, revision, revision reminder, etc., via emails from our system or assigned Assistant Editor. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China (Nos: 2018YFA0901000, 2018YFA0901003), the National Natural Science Foundation of China (Grant Nos: 31930004 and 32150021), and the Third Xinjiang Scientific Expedition (2022xjkk020602). We also appreciate the support of the Agricultural Science and Technology Innovation Program of CAAS.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The whole genome sequence data reported in this paper have been deposited in the Genome Warehouse in the National Genomics Data Center, Beijing Institute of Genomics, Chinese Academy of Sciences/China National Center for Bioinformation, under accession number GWHBQTK01000000, which is publicly accessible at https://ngdc.cncb.ac.cn/gwh (accessed on 5 January 2023).
Acknowledgments
Special thanks go to Zhengfu Zhou and Jin Wang for their constructive suggestions for the revision of this article and other authors for their excellent technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhuang, Y.T.; Liu, R.C.; Chen, Y.L.; Yang, C.Y.; Yan, Y.U.; Wang, T.; Wang, Y.L.; Song, Y.J.; Teng, Y. Extremophiles and their applications. Sci. Sin. Vitae 2022, 52, 204–222. [Google Scholar] [CrossRef]
- Mehta, R.; Singhal, P.; Singh, H.; Damle, D.; Sharma, A.K. Insight into thermophiles and their wide-spectrum applications. 3 Biotech 2016, 6, 81. [Google Scholar] [CrossRef]
- Singh, D.N.; Sood, U.; Singh, A.K.; Gupta, V.; Shakarad, M.; Rawat, C.D.; Lal, R. Genome Sequencing Revealed the Biotechnological Potential of an Obligate Thermophile Geobacillus thermoleovorans Strain RL Isolated from Hot Water Spring. Indian J. Microbiol. 2019, 59, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Vilcaez, J.; Suto, K.; Inoue, C. Bioleaching of chalcopyrite with thermophiles: Temperature-pH-ORP dependence. Int. J. Miner. Process. 2008, 88, 37–44. [Google Scholar] [CrossRef]
- Najar, I.N.; Sherpa, M.T.; Das, S.; Verma, K.; Dubey, V.K.; Thakur, N. Geobacillus yumthangensis sp nov., a thermophilic bacterium isolated from a north-east Indian hot spring. Int. J. Syst. Evol. Microbiol. 2018, 68, 3430–3434. [Google Scholar] [CrossRef]
- Zeigler, D.R. The Geobacillus paradox: Why is a thermophilic bacterial genus so prevalent on a mesophilic planet? Microbiology 2014, 160, 1–11. [Google Scholar] [CrossRef]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.A.; Ivanova, A.E.; Lysenko, A.M.; Petrunyaka, V.V.; Osipov, G.A.; Belyaev, S.S.; et al. Taxonomic study of aerobic thermophilic bacilli: Descriptions of Geobacillus subterraneus gen. nov., sp. nov. and Geobacillus uzenensis sp. nov. from petroleum reservoirs and transfer of Bacillus stearothermophilus, Bacillus thermocatenulatus, Bacillus thermoleovorans, Bacillus kaustophilus, Bacillus thermodenitrificans to Geobacillus as the new combinations G. stearothermophilus, G. th. Int. J. Syst. Evol. Microbiol. 2001, 51, 433–446. [Google Scholar]
- Khaswal, A.; Chaturvedi, N.; Mishra, S.K.; Kumar, P.R.; Paul, P.K. Current status and applications of genus Geobacillus in the production of industrially important products—A review. Folia Microbiol. 2022, 67, 389–404. [Google Scholar] [CrossRef]
- Novik, G.; Savich, V.; Meerovskaya, O. Geobacillus Bacteria: Potential Commercial Applications in Industry, Bioremediation, and Bioenergy Production. In Growing and Handling of Bacterial Cultures; IntechOpen: London, UK, 2018. [Google Scholar]
- Cripps, R.E.; Eley, K.; Leak, D.J.; Rudd, B.; Taylor, M.; Todd, M.; Boakes, S.; Martin, S.; Atkinson, T. Metabolic engineering of Geobacillus thermoglucosidasius for high yield ethanol production. Metab. Eng. 2009, 11, 398–408. [Google Scholar] [CrossRef]
- Mir, M.Y.; Hamid, S.; Rohela, G.K.; Parray, J.A.; Kamili, A.N. Composting and Bioremediation Potential of Thermophiles. In Soil Bioremediation: An Approach Towards Sustainable Technology; Wiley: Hoboken, NJ, USA, 2021. [Google Scholar]
- Sari, B.; Faiz, O.; Genc, B.; Sisecioglu, M.; Adiguzel, A.; Adiguzel, G. New xylanolytic enzyme from Geobacillus galactosidasius BS61 from a geothermal resource in Turkey. Int. J. Biol. Macromol. 2018, 119, 1017–1026. [Google Scholar] [CrossRef]
- Abol-Fotouh, D.; AlHagar, O.E.A.; Hassan, M.A. Optimization, purification, and biochemical characterization of thermoalkaliphilic lipase from a novel Geobacillus stearothermophilus FMR12 for detergent formulations. Int. J. Biol. Macromol. 2021, 181, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, W.; Tayyab, M.; Aftab, M.N.; Hashmi, A.; Ahmad, M.D.; Firyal, S.; Wasim, M.; Awan, A.R. Optimization of Conditions for the Higher Level Production of Protease: Characterization of Protease from Geobacillus SBS-4S. Waste Biomass Valorization 2020, 11, 6613–6623. [Google Scholar] [CrossRef]
- Gandhi, S.; Salleh, A.B.; Rahman, R.N.; Chor Leow, T.; Oslan, S.N. Expression and Characterization of Geobacillus stearothermophilus SR74 Recombinant α-Amylase in Pichia pastoris. Biomed. Res. Int. 2015, 2015, 529059. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zha, S.; Ma, Y.; Xiao, L.; Yang, H. The whole-genome sequencing and sequence analysis of Geobacillus sp. YHL. J. Jiangxi Norm. Univ. (Nat. Sci.) 2022, 46, 147–155. [Google Scholar]
- Sarkar, P.; Lepcha, K.; Ghosh, S. Purification and Characterization of Solvent Stable Lipase from a Solvent Tolerant Strain of Geobacillus Stearothermophilus Ps 11. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 602–605. [Google Scholar] [CrossRef]
- Han, S.; Zhang, J.; Jing, Y. Progress in microbial-derived proteases. Food Ind. Technol. 2020, 41, 321–327. [Google Scholar]
- Ye, S.; Zhao, Y. Production and application of microbial-derived alkaline protease. Qinghai Sci. Technol. 2018, 25, 4. [Google Scholar]
- Iqbal, I.; Aftab, M.N.; Afzal, M.; Ur-Rehman, A.; Aftab, S.; Zafar, A.; Ud-Din, Z.; Khuharo, A.R.; Iqbal, J.; Ul-Haq, I. Purification and characterization of cloned alkaline protease gene of Geobacillus stearothermophilus. J. Basic Microbiol. 2015, 55, 160–171. [Google Scholar] [CrossRef]
- Chen, X.G.; Stabnikova, O.; Tay, J.H.; Wang, J.Y.; Tay, S.T. Thermoactive extracellular proteases of Geobacillus caldoproteolyticus, sp. nov., from sewage sludge. Extremophiles 2004, 8, 489–498. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Darling, A.; Mau, B.; Blattner, F.R.; Perna, A. Mauve: Multiple Alignment of Conserved Genomic Sequence With Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Concepcion, G.T.; Feng, X.; Zhang, H.; Li, H. Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat. Methods 2021, 18, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; Heijne, G.V.; Nielsen, H.H. SIGNALP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Heijne, G.V.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes—ScienceDirect. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Studholme, D.J. Some (bacilli) like it hot: Genomics of Geobacillus species. Microb. Biotechnol. 2015, 8, 40–48. [Google Scholar] [CrossRef]
- Suzuki, H. Peculiarities and biotechnological potential of environmental adaptation by Geobacillus species. Appl. Microbiol. Biotechnol. 2018, 102, 10425–10437. [Google Scholar] [CrossRef]
- Bezuidt, O.K.; Rian, P.; Gomri, A.M.; Fiyin, A.; Makhalanyane, T.P.; Karima, K.; Cowan, D.A. The Geobacillus Pan-Genome: Implications for the Evolution of the Genus. Front. Microbiol. 2016, 7, 723. [Google Scholar] [CrossRef]
- Burgess, S.A.; Flint, S.H.; Lindsay, D.; Cox, M.P.; Biggs, P.J. Insights into the Geobacillus stearothermophilus species based on phylogenomic principles. BMC Microbiol. 2017, 17, 140. [Google Scholar] [CrossRef]
- Wada, K.; Kobayashi, J.; Furukawa, M.; Doi, K.; Ohshiro, T.; Suzuki, H. A thiostrepton resistance gene and its mutants serve as selectable markers in Geobacillus kaustophilus HTA426. Biosci. Biotechnol. Biochem. 2016, 80, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Contesini, F.J.; Melo, R.R.; Sato, H.H. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol. 2018, 38, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Pudova, D.S.; Toymentseva, A.A.; Gogoleva, N.E.; Shagimardanova, E.I.; Mardanova, A.M.; Sharipova, M.R. Comparative Genome Analysis of Two Bacillus pumilus Strains Producing High Level of Extracellular Hydrolases. Genes 2022, 13, 409. [Google Scholar] [CrossRef]
- Hussein, A.H.; Lisowska, B.K.; Leak, D.J. The genus Geobacillus and their biotechnological potential. Adv. Appl. Microbiol. 2015, 92, 1–48. [Google Scholar] [PubMed]
- Logan, N.A.; Vos, P.D. Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- Peralta-Yahya, P.P.; Zhang, F.; del Cardayre, S.B.; Keasling, J.D. Microbial engineering for the production of advanced biofuels. Nature 2012, 488, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Semenova, E.M.; Sokolova, D.S.; Grouzdev, D.S.; Poltaraus, A.B.; Vinokurova, N.G.; Tourova, T.P.; Nazina, T.N. Geobacillus proteiniphilus sp. nov., a thermophilic bacterium isolated from a high-temperature heavy oil reservoir in China. Int. J. Syst. Evol. Microbiol. 2019, 69, 3001–3008. [Google Scholar] [CrossRef]
- Wang, J.; Goh, K.M.; Salem, D.R.; Sani, R.K. Genome analysis of a thermophilic exopolysaccharide-producing bacterium—Geobacillus sp. WSUCF1. Sci. Rep. 2019, 9, 1608. [Google Scholar] [CrossRef] [PubMed]
- SanthaKalaikumari, S.; Sivakumar, R.; Gunasekaran, P.; Rajendhran, J. Whole-genome Sequencing and Mining of Protease Coding Genes in Bacillus paralicheniformis MKU3, and its Degradomics in Feather Meal Medium. Curr. Microbiol. 2021, 78, 206–217. [Google Scholar] [CrossRef]
- Venugopal, V.; Alur, M.D.; Nerkar, D.P. Solubilization of fish proteins using immobilized microbial cells. Biotechnol. Bioeng. 1989, 33, 1098–1103. [Google Scholar] [CrossRef]
- Zhu, W.; Cha, D.M.; Cheng, G.Y.; Peng, Q.; Shen, P. Purification and characterization of a thermostable protease from a newly isolated Geobacillus sp. YMTC 1049. Enzym. Microb. Technol. 2007, 40, 1592–1597. [Google Scholar] [CrossRef]
- Forster, A.H.; Gescher, J. Metabolic Engineering of Escherichia coli for Production of Mixed-Acid Fermentation End Products. Front. Bioeng. Biotechnol. 2014, 2, 16. [Google Scholar] [PubMed]
- Hartley, B.S.; Payton, M.A. Industrial prospects for thermophiles and thermophilic enzymes. Biochem. Soc. Symp. 1983, 48, 133–146. [Google Scholar] [PubMed]
- Miah, M.S.; Tada, C.; Sawayama, S. Enhancement of Biogas Production from Sewage Sludge with the Addition of Geobacillus sp. Strain AT1 Culture. Jpn. J. Water Treat. Biol. 2010, 40, 97–104. [Google Scholar] [CrossRef]
- Mohamed, R.A.; Salleh, A.B.; Noor, R.; Raja, Z.; Leow, T.C. Isolation of the encoding gene for a thermostable -glucosidase from Geobacillus stearothermophilus strain RM and its expression in Escherichia coli. Afr. J. Microbiol. Res. 2012, 6, 2909–2917. [Google Scholar]
Figure 1.
Phylogenetic tree of the 16S rRNA gene sequence based on maximum likelihood (ML). This tree shows the phylogenetic relationships between G. stearothermophilus H6 and closely related species. The GenBank registration number of the 16S rRNA gene sequence is shown in parentheses. Bootstrap test: the percentage is based on 1000 duplicates (bootstrap value > 70, indicating that the branch is reliable, and less than 50% will not be displayed).
Figure 1.
Phylogenetic tree of the 16S rRNA gene sequence based on maximum likelihood (ML). This tree shows the phylogenetic relationships between G. stearothermophilus H6 and closely related species. The GenBank registration number of the 16S rRNA gene sequence is shown in parentheses. Bootstrap test: the percentage is based on 1000 duplicates (bootstrap value > 70, indicating that the branch is reliable, and less than 50% will not be displayed).

Figure 2.
Genomic synteny analysis of G. stearothermophilus H6, G. stearothermophilus B5 and G. stearothermophilus 10. The same color block represents two genomes with high homology.
Figure 2.
Genomic synteny analysis of G. stearothermophilus H6, G. stearothermophilus B5 and G. stearothermophilus 10. The same color block represents two genomes with high homology.

Figure 3.
The experimental strains of bacteria were cultured on skimmed milk plates at 37 °C and 60 °C for 12 h, 24 h, 36 h and 48 h: (a) cultured at 37 °C; (b) cultured at 60 °C.
Figure 3.
The experimental strains of bacteria were cultured on skimmed milk plates at 37 °C and 60 °C for 12 h, 24 h, 36 h and 48 h: (a) cultured at 37 °C; (b) cultured at 60 °C.

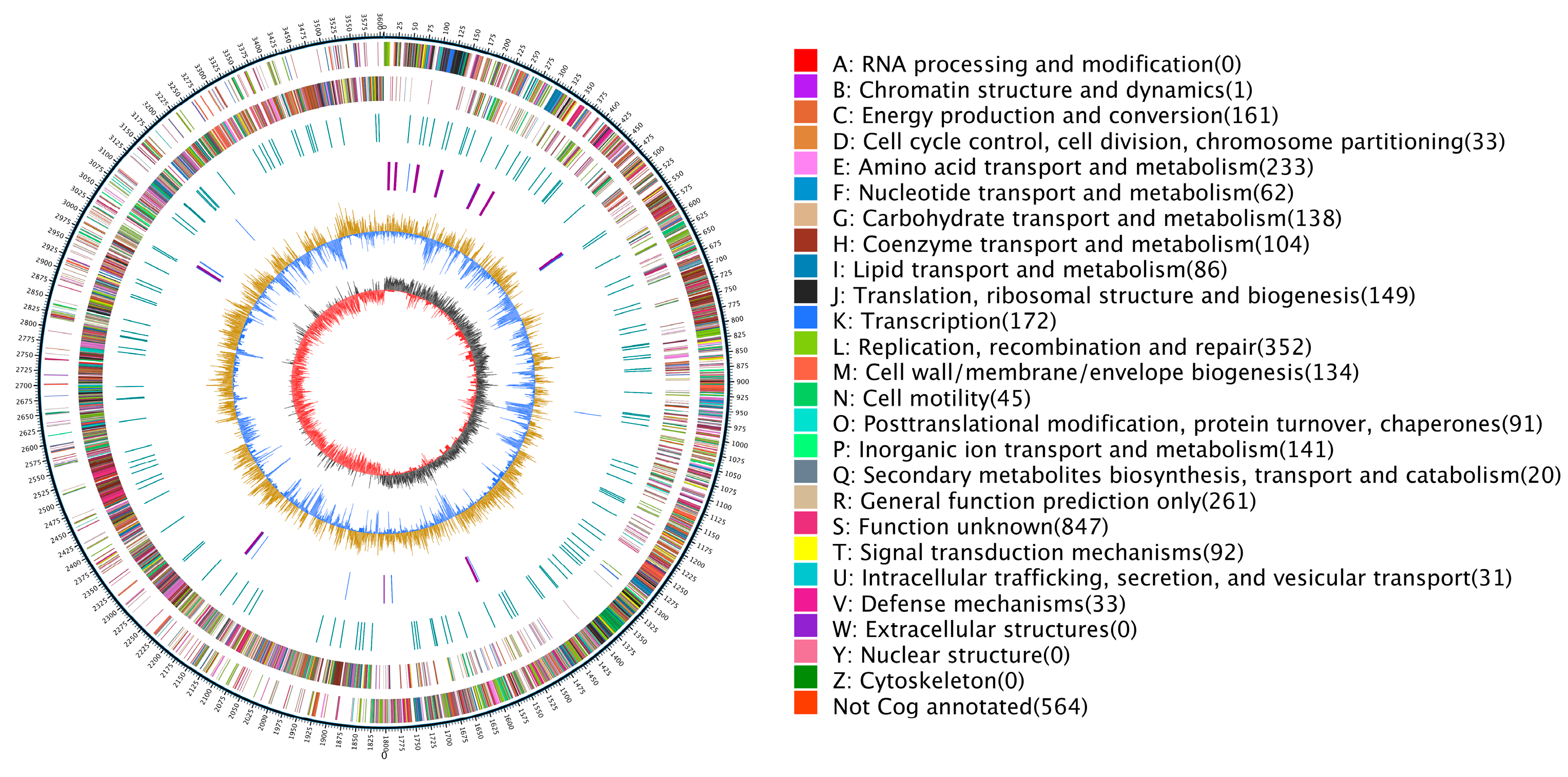
Figure 4.
Circular genome map of G. stearothermophilus H6. Note: The outermost circle is a mark of genome size, each scale is 5 kb; the second and third circles are the gene on the positive and negative chains of the genome, respectively. Different colors represent different COG functional classifications; the fourth circle is the repeat sequence; the fifth circle is tRNA and rRNA, where blue is tRNA and purple is rRNA; the sixth circle is the GC content. The straw yellow part indicates that the GC content in this region is higher than the average GC content of the genome. The higher the peak value is, the greater the difference between the GC content and the average GC content is. The blue part indicates that the GC content in this region is lower than the average GC content of the genome; the innermost circle is GC skew. Dark grey represents the area where G content is greater than C, and red represents the area where C content is greater than G.
Figure 4.
Circular genome map of G. stearothermophilus H6. Note: The outermost circle is a mark of genome size, each scale is 5 kb; the second and third circles are the gene on the positive and negative chains of the genome, respectively. Different colors represent different COG functional classifications; the fourth circle is the repeat sequence; the fifth circle is tRNA and rRNA, where blue is tRNA and purple is rRNA; the sixth circle is the GC content. The straw yellow part indicates that the GC content in this region is higher than the average GC content of the genome. The higher the peak value is, the greater the difference between the GC content and the average GC content is. The blue part indicates that the GC content in this region is lower than the average GC content of the genome; the innermost circle is GC skew. Dark grey represents the area where G content is greater than C, and red represents the area where C content is greater than G.

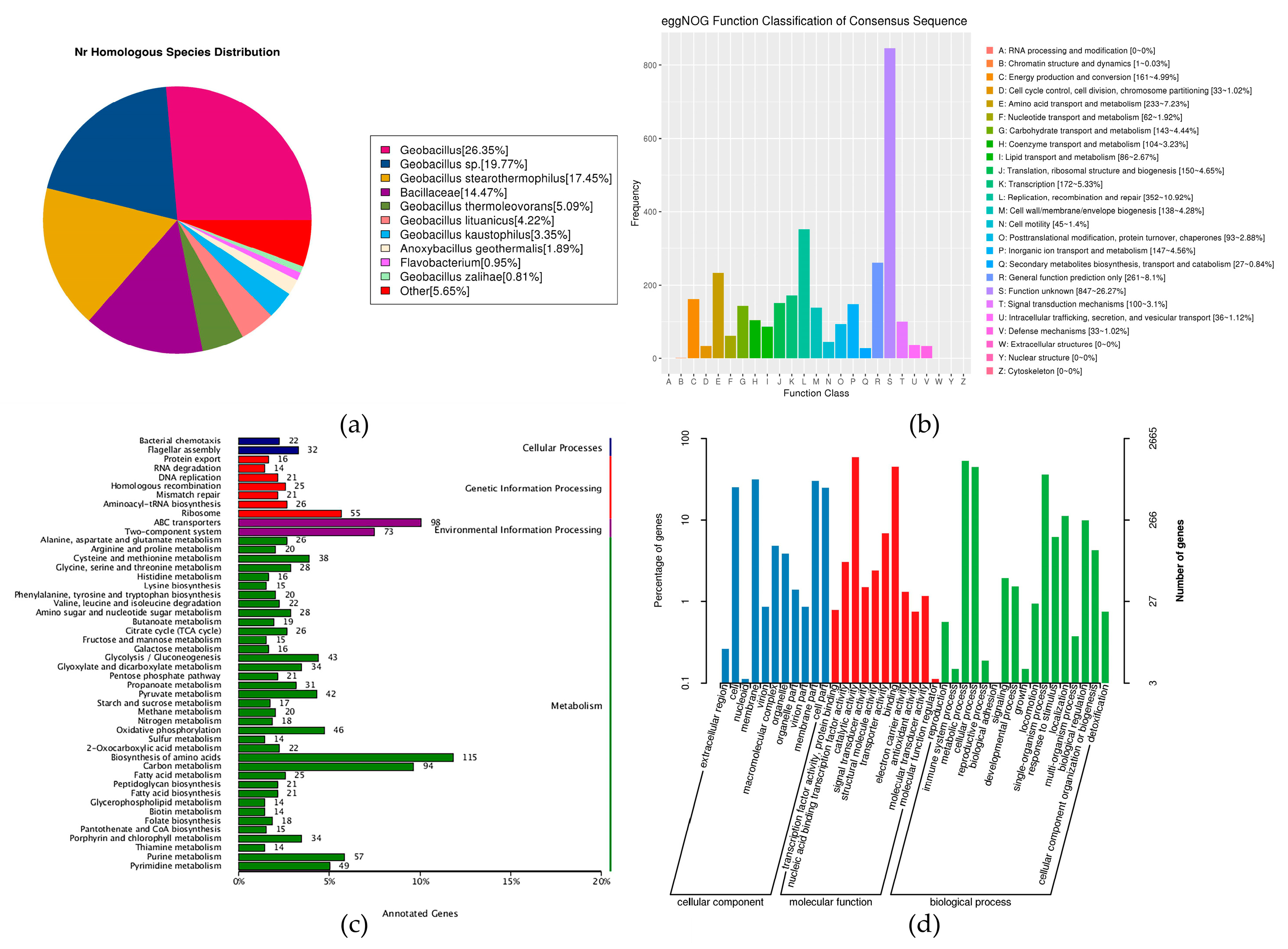
Figure 5.
G. stearothermophilus H6 general protein prediction database: (a) Species distribution map of sequences compared to the Nr database: this map reflects the species distribution of the sequences compared to the Nr database. Different colors represent different species. (b) Statistical chart of the functional classification of eggNOG functional genes: the abscissa represents the classification content of eggNOG, and the ordinate represents the relative contents of the number of corresponding functional genes. (c) Statistical chart of KEGG annotation classifications: the ordinate represents the KEGG secondary classification, and the abscissa represents percentage. (d) Statistical chart of GO function annotation classifications: the abscissa represents the GO classification content, the left side of the ordinate represents the percentage of the number of genes, and the right side represents the number of genes. This figure shows the gene enrichment of each secondary GO function against the background of all genes, reflecting the status of each secondary function against this background.
Figure 5.
G. stearothermophilus H6 general protein prediction database: (a) Species distribution map of sequences compared to the Nr database: this map reflects the species distribution of the sequences compared to the Nr database. Different colors represent different species. (b) Statistical chart of the functional classification of eggNOG functional genes: the abscissa represents the classification content of eggNOG, and the ordinate represents the relative contents of the number of corresponding functional genes. (c) Statistical chart of KEGG annotation classifications: the ordinate represents the KEGG secondary classification, and the abscissa represents percentage. (d) Statistical chart of GO function annotation classifications: the abscissa represents the GO classification content, the left side of the ordinate represents the percentage of the number of genes, and the right side represents the number of genes. This figure shows the gene enrichment of each secondary GO function against the background of all genes, reflecting the status of each secondary function against this background.

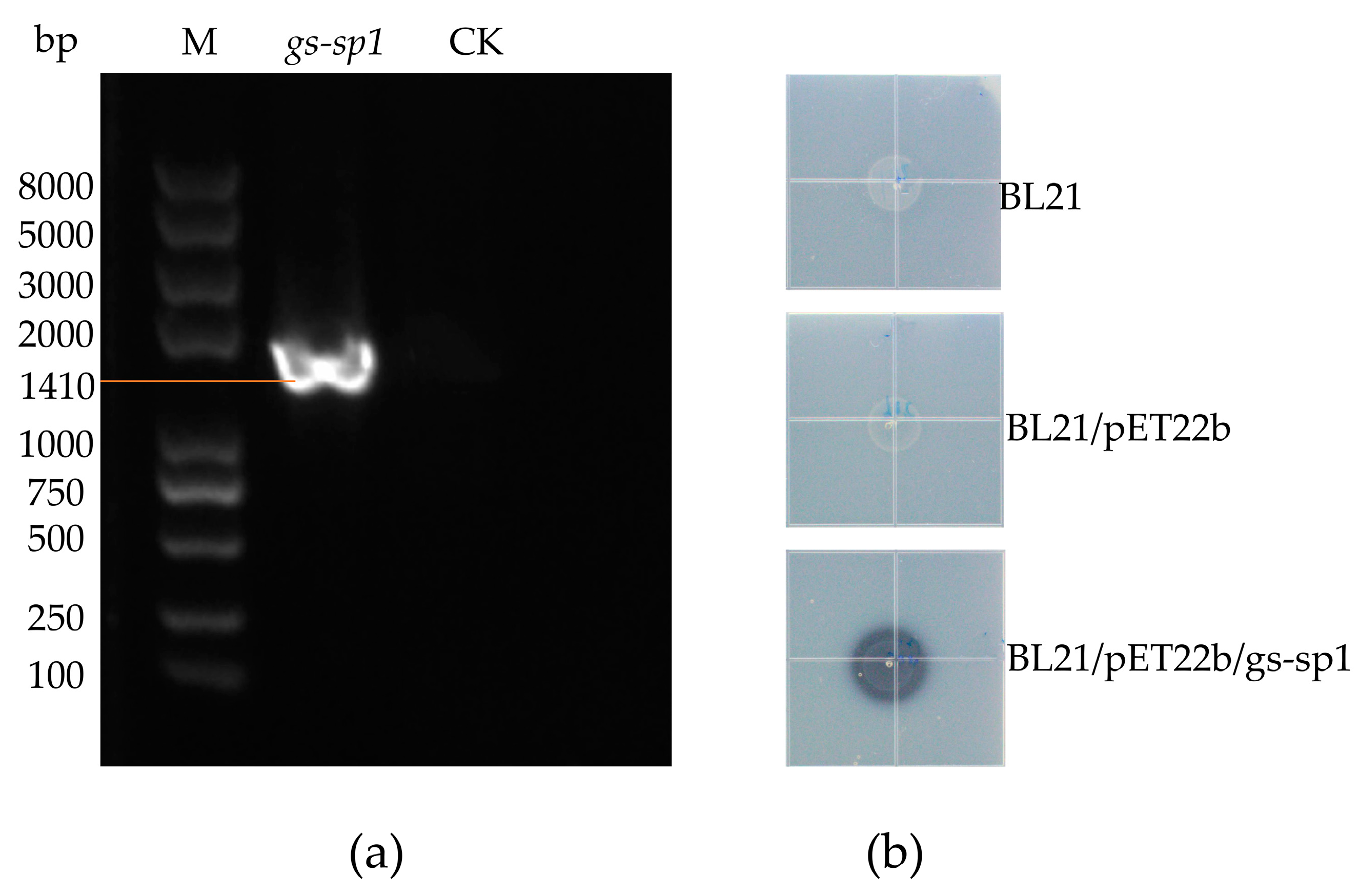
Figure 6.
pET22b/gs-sp1 recombinant vector and protease activity observation: (a) PCR validation of the pET22b/gs-sp1 plasmid, M: Trans2K Plus DNA Marker; (b) after the strain was induced by 0.1 mM IPTG for 24 h, the transparent circle.
Figure 6.
pET22b/gs-sp1 recombinant vector and protease activity observation: (a) PCR validation of the pET22b/gs-sp1 plasmid, M: Trans2K Plus DNA Marker; (b) after the strain was induced by 0.1 mM IPTG for 24 h, the transparent circle.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of basic characteristics of whole-genome sequences of strain H6 and other strains of G. stearothermophilus.
Table 1.
Comparison of basic characteristics of whole-genome sequences of strain H6 and other strains of G. stearothermophilus.
| Type | Size (Mb) | GC% | Protein | Gene | Average Nucleotide Identity (ANI) | Isolation |
|---|---|---|---|---|---|---|
| G. stearothermophilus H6 | 3.6 | 51.66 | 3706 | 3750 | - | hyperthermophilic composting |
| G. stearothermophilus B5 (CP034952.1) | 3.39 | 52.5 | 3114 | 3446 | 97.35% | rice stack |
| G. stearothermophilus DSM 458 (CP016552.1) | 3.47 | 52.1 | 3232 | 3614 | 96.35% | sugar beet juice from extraction installations |
| G. stearothermophilus DG-1 (CP063162.1) | 3.51 | 52.5 | 3322 | 3627 | 97.07% | oilfield |
| G. stearothermophilus D1 (NZ_LDNU01000016.1) | 2.97 | 52.2 | 2954 | 3482 | 97.65% | milk powder manufacturing plant |
| G. stearothermophilus 10 (CP008934.1) | 3.65 | 52.6 | 3288 | 3473 | 89.56% | hot spring |
Table 2.
General genomic characteristics of G. stearothermophilus H6.
| Attribute | Value |
|---|---|
| Size (bp) | 3,606,258 |
| GC content (%) | 51.66 |
| Total genes | 3750 |
| RNA genes | 121 |
| rRNAs | 30 |
| tRNAs | 91 |
| Total repetitive sequence length (bp) | 5600 |
| CRISPR number | 17 |
| Number of genomic islands | 24 |
| Number of signal peptides | 161 |
| Transmembrane protein | 841 |
Table 3.
Basic characteristics of proteases in the genome of G. stearothermophilus H6.
| Types of Proteases | Number of Protease Sequences | Number of Signal Peptides | Number of Transmembrane Helixes |
|---|---|---|---|
| Serine protease | 27 | 5 | 12 |
| Metallopeptidase | 38 | 5 | 16 |
| Cysteine protease | 1 | 0 | 0 |
| Aspartic peptidase | 3 | 0 | 3 |
| Threonine peptidase | 1 | 0 | 0 |
| Other | 71 | 8 | 15 |
| Total | 141 | 18 | 46 |
Table 4.
Basic characteristics and homology analysis of proteases secreted by G. stearothermophilus H6.
Table 4.
Basic characteristics and homology analysis of proteases secreted by G. stearothermophilus H6.
| ID | Amino Acid Length | Average Molecular Weight (kDa) | Academic pl (pH) | Type of Signal Peptide | Homologous Protein and Identity | |
|---|---|---|---|---|---|---|
| G. stearothermophilus B5 | G. stearothermophilus 10 | |||||
| GE000011 | 451 | 51.00 | 9.64 | SP(Sec/SPI) | WP_160269798.1 (36.20%) | ALA70391.1 (92.22%) |
| GE000377 | 263 | 30.03 | 5.84 | SP(Sec/SPI) | - | ALA70722.1 (99.24%) |
| GE000438 | 150 | 16.72 | 10.42 | LIPO(Sec/SPII) | WP_160268695.1 (97.33%) | ALA70781.1 (86.67%) |
| GE001521 | 341 | 36.35 | 9.47 | SP(Sec/SPI) | WP_160269176.1 (90.62%) | ALA69075.1 (96.19%) |
| GE001730 | 453 | 49.85 | 6.01 | SP(Sec/SPI) | WP_160269346.1 (97.79%) | ALA68779.1 (96.24%) |
| GE001981 | 618 | 70.05 | 8.76 | SP(Sec/SPI) | WP_160269500.1 (99.03%) | ALA71875.1 (95.30%) |
| GE002130 | 437 | 49.26 | 4.05 | SP(Sec/SPI) | WP_160270355.1 (28.57%) | ALA70136.1 (28.16%) |
| GE002405 | 381 | 42.86 | 9.47 | LIPO(Sec/SPII) | WP_160269774.1 (99.48%) | ALA71443.1 (93.44%) |
| GE002430 | 391 | 43.35 | 8.92 | SP(Sec/SPI) | WP_160269798.1 (93.61%) | ALA71420.1 (90.28%) |
| GE002476 | 168 | 18.34 | 10.27 | SP(Sec/SPI) | WP_160269827.1 (98.81%) | ALA71363.1 (96.43%) |
| GE003057 | 335 | 37.00 | 5.89 | SP(Sec/SPI) | WP_160270729.1 (97.31%) | ALA69727.1 (95.52%) |
| GE003085 | 98 | 10.36 | 9.26 | SP(Sec/SPI) | WP_160270055.1 (98.98%) | ALA69758.1 (81.63%) |
| GE003250 | 329 | 37.49 | 9.17 | SP(Sec/SPI) | WP_160270168.1 (96.66%) | ALA69899.1 (93.10%) |
| GE003377 | 432 | 47.72 | 6.06 | SP(Sec/SPI) | WP_160270252.1 (92.13%) | ALA70034.1 (96.30%) |
| GE003445 | 134 | 15.01 | 9.96 | SP(Sec/SPI) | WP_160270104.1 (76.00%) | - |
| GE003446 | 1338 | 145.36 | 9.07 | SP(Sec/SPI) | WP_160270104.1 (38.24%) | ALA69775.1 (49.32%) |
| GE003450 | 452 | 49.14 | 9.67 | SP(Sec/SPI) | WP_236658918.1 (66.67%) | ALA70130.1 (67.33%) |
| GE003532 | 1447 | 156.28 | 6 | SP(Sec/SPI) | WP_160270104.1 (40.06%) | ALA69775.1 (52.21%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lai, R.; Lin, M.; Yan, Y.; Jiang, S.; Zhou, Z.; Wang, J. Comparative Genomic Analysis of a Thermophilic Protease-Producing Strain Geobacillus stearothermophilus H6. Genes 2023, 14, 466. https://doi.org/10.3390/genes14020466
AMA Style
Lai R, Lin M, Yan Y, Jiang S, Zhou Z, Wang J. Comparative Genomic Analysis of a Thermophilic Protease-Producing Strain Geobacillus stearothermophilus H6. Genes. 2023; 14(2):466. https://doi.org/10.3390/genes14020466
Chicago/Turabian StyleLai, Ruilin, Min Lin, Yongliang Yan, Shijie Jiang, Zhengfu Zhou, and Jin Wang. 2023. "Comparative Genomic Analysis of a Thermophilic Protease-Producing Strain Geobacillus stearothermophilus H6" Genes 14, no. 2: 466. https://doi.org/10.3390/genes14020466
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.