
Phylogeny of the Diploid Species of Rubus (Rosaceae)
by
 , ,
, ,
Xin-Fen Gao
1,*,†,
Xian-Hua Xiong
2,†,
David E. Boufford
3,
Yun-Dong Gao
1,
Bo Xu
1 and
Cheng Zhang
1 1
CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China
2
College of Life Science and Biotechnology, Mianyang Teachers’ College, Mianyang 621000, China
3
Harvard University Herbaria, Harvard University, 22 Divinity Avenue, Cambridge, MA 02138, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2023, 14(6), 1152; https://doi.org/10.3390/genes14061152
Submission received: 25 April 2023
/
Revised: 23 May 2023
/
Accepted: 24 May 2023
/
Published: 25 May 2023
(This article belongs to the Section Population and Evolutionary Genetics and Genomics)
Abstract
:Rubus L. (Rosaceae, Rosoideae) contains around 700 species distributed on all continents except Antarctica, with the highest species diversity in temperate to subtropical regions of the northern hemisphere. The taxonomy of Rubus is challenging due to the frequency of polyploidy, hybridization and apomixis. Previous studies mostly sampled sparsely and used limited DNA sequence data. The evolutionary relationships between infrageneric taxa, therefore, remain to be further clarified. In the present study, genotyping by sequencing (GBS) reduced-representation genome sequencing data from 186 accessions representing 65 species, 1 subspecies and 17 varieties of Rubus, with emphasis on diploid species, were used to infer a phylogeny using maximum likelihood and maximum parsimony methods. The major results were as follows: (1) we confirmed or reconfirmed the polyphyly or paraphyly of some traditionally circumscribed subgenera, sections and subsections; (2) 19 well-supported clades, which differed from one another on molecular, morphological and geographical grounds, were identified for the species sampled; (3) characteristics such as plants with dense bristles or not, leaves leathery or papyraceous, number of carpels, instead of inflorescences paniculate or not, aggregate fruits and leaves abaxially tomentose or not, may be of some use in classifying taxa whose drupelets are united into a thimble-shaped aggregate fruit that falls in its entirety from the dry receptacle; and (4) a preliminary classification scheme of diploid species of Rubus is proposed based on our results combined with those from previous phylogenetic analyses.
1. Introduction
Rubus L. (Rosaceae; Rosoideae), a species-rich and economically important genus, is characterized by shrubs, subshrubs and herbs bearing stipules, usually prickles or bristles, and a compound fruit comprising an aggregation of drupelets. Species of Rubus are on all continents except Antarctica [1,2]. Except for a fraction of the species in the southern hemisphere, most species of Rubus occur in temperate to subtropical regions of the northern hemisphere, especially in temperate Eurasia and North America [3,4,5,6,7]. The exact species number of Rubus is unclear. The estimated number has ranged from ca. 200 species [8], ca. 437 species [9,10,11], 250–700 species [2], ca. 700 species [4,6,7] and 600–1000 species [12] to 900–1000 species [13]. Various species of Rubus are economically significant as fruit crops, ornamentals, invasive weeds and as pioneers in early forest succession [1,14].
The taxonomy of Rubus has historically been difficult due to morphological diversity and overlapping characteristics, apomixis, polyploidization and hybridization [1,5,15,16,17,18,19,20,21]. Morphologically, the species of Rubus exhibit tremendous diversity, with plants ranging from woody to semi-woody, trailing, erect or climbing shrubs or subshrubs to perennial creeping dwarf herbs. The leaves range from simple to palmately or pinnately compound with 3–11(–15) leaflets [1,7,15,22]. Rubus has a basic chromosome number of seven, and ploidy levels ranging from diploid to tetradecaploid (or octadecaploid) [5,13,20,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42]. Hybridization in Rubus occurs mostly between closely related species, but also in some cases between species classified in different subgenera [1].
The most recent and the only global taxonomic treatment of Rubus was made by Focke [9,10,11] based on morphological characteristics. Focke divided Rubus into 12 subgenera: Anoplobatus (Focke) Focke, Chamaebatus (Focke) Focke, Chamaemorus (Hill) Focke, Comaropsis (Rich. ex Nestl.) Focke, Cylactis (Raf.) Focke, Dalibarda (L.) Focke, Dalibardastrum Focke, Idaeobatus (Focke) Focke, Lampobatus (Focke) Focke, Malachobatus (Focke) Focke, Orobatus (Focke) Focke and Rubus (subg. Eubatus Focke). Two subgenera, Diemenicus A. R. Bean (including only Rubus gunnianus) [43] and Micranthobatus (Firtsch) Kalkm. [44], were subsequently described. Among them, the three largest subgenera are Idaeobatus, Malachobatus and Rubus, the first two occurring mainly in Asia (especially in China); subg. Rubus is in Europe and North America [5]. The raspberries, one of the economically important crops of Rubus, mainly belong to subg. Idaeobatus, while the blackberries are primarily included in subg. Rubus. The subgenera Anoplobatus, Dalibarda and Idaeobatus are predominantly diploid, whereas subgenera Dalibardastrum, Malachobatus and Orobatus are almost entirely polyploid, and subg. Chamaebatus, subg. Cylactis and subg. Rubus contain a small number of diploid members [13,32].
Molecular data have shown Rubus to be sister to the clade comprising the genus Rosa L., tribe Sanguisorbeae DC., tribe Potentilleae Sweet and tribe Colurieae Rydb. [45]. Further studies found Rubus to be monophyletic, whereas among the long-recognized subgeneric taxa, except for subg. Orobatus, all other subgenera were paraphyletic or polyphyletic [1,42,46]. Previously, although a diversity of molecular markers, including GBSSI-1, GBSSI-2, ITS, LEAFY, ndhF, PEPC, rbcL, rpl16, rpl20-rps12, rpl32-ndhF, trnK, trnL-trnF, trnS-trnG and trnV-ndhC [1,14,21,37,46,47,48,49,50,51,52,53,54,55], were used to infer the phylogeny of Rubus, most of those studies applied to a limited number of species and molecular loci or were restricted to taxa within administrative boundaries, resulting in low phylogenetic resolution and sometimes even giving inconsistent results. The evolutionary relationships between the species of Rubus are in need of further clarification.
The rapid development of next-generation sequencing enables researchers to use genome-wide polymorphisms from different species for phylogenetic analysis. Reduced representation sequencing methods, such as genotyping by sequencing (GBS), can provide thousands of single nucleotide polymorphisms (SNPs) and have been used for phylogenetic analyses of both closely and distantly related taxa [56,57,58,59,60,61,62,63,64,65].
Given that the polyploid species of Rubus have a complex evolutionary history, this paper focuses on the phylogeny of diploid species of Rubus. We inferred a molecular phylogeny of diploid species of Rubus with thousands of SNPs identified by the GBS approach to (1) vigorously test the monophyly of subgenera, sections and subsections of Rubus based on larger sampling; (2) identify major clades within diploid species of Rubus; and (3) build the molecular foundation for future taxonomic revision of Rubus.
2. Materials and Methods
2.1. Taxon Sampling
The taxa were chosen to include as many diploids species of Rubus as possible. In total, 186 accessions, representing 65 species, 1 subspecies and 17 varieties of Rubus, constituted our ingroup. Except for subg. Chamaebatus, our sampling covered all subgenera containing diploid species and the major geographical range of the diploid species of the genus. Diploid representatives of Fragaria L. (one accession representing one species, Fragaria daltoniana) and Rosa L. (three accessions representing two species, Rosa cymosa and Rosa luciae) were selected as outgroups based on previous findings that Rosa, Rubus and three tribes (Sanguisorbeae, Potentilleae (including Fragaria) and Colurieae) constitute the super tribe Rosodae in the subfamily Rosoideae of Rosaceae. Voucher information for the sampled taxon are provided in the Appendix A.
2.2. Genotyping by sequencing (GBS) Dataset
Total genomic DNA was extracted from silica-dried material using the CTAB method [66]. The DNA was quantified, and its quality was evaluated using the following methods: (a) Agarose gel electrophoresis to test DNA purity and integrity, (b) Nanodrop spectrophotometer to test DNA purity (OD260/OD280), (c) Qubit Fluorometer to measure DNA concentration.
A reduced representation GBS library was prepared as described by Elshire et al. [56], using the restriction enzymes MseI and HaeII, and sequenced (paired-end reads) using the Illumina HiSeq platform at Novogene Bioinformatics Technology Co., Ltd., Beijing, China.
The raw data (raw reads) were first processed by fastp version 0.23.1 [67] in a series of quality control (QC) procedures using standards as follows: (a) removal of reads containing adapters; (b) removal of reads with >50% bases having phred quality < 5; and (c) removal of reads with ≥10% unidentified nucleotides (N). The remaining high quality clean reads were mapped to the reference genome (chloroplast genome, Rubus coreanus [68]; whole genome, Rubus occidentalis [69]) using the software BWA version 0.7.8 [70] with the command “mem -t 4 -k 32 -M”. In order to reduce mismatch generated by PCR amplification before sequencing, duplicated reads were removed by the help of SAMtools version 1.3.1 [71]. After alignment, we performed SNP calling on a population scale using a Bayesian approach as implemented in the SAMtools package, and then genotype likelihoods from reads for each individual at each genomic location and the allele frequencies in the sample were calculated with a Bayesian approach. The “mpileup” command was used to identify SNPs with the parameters as “-q 1 -C 50 -t SP -t DP -m 2 -F 0.002”. The resulting SNPs were filtered by script with the parameters “dp 4, miss 0.5 and maf 0.01′” to obtain high quality SNPs for subsequent phylogenetic analysis.
2.3. Phylogenetic Analysis
The chloroplast and nuclear datasets were analyzed separately. Equally weighted maximum parsimony (MP) jackknife (JK) analyses [72] were conducted using 1000 random-taxon-addition replicates and tree-bisection-reconnection (TBR) branch swapping in PAUP* version 4.0a169 [73], with MAXTREES set to 10,000 and the removal probability set to approximately 37%, and “jac” resampling emulated analyses. Insertions and deletions were coded as missing data. Jackknife 50% majority-rule consensus trees were computed.
jModelTest 2 [74,75] was used to test the models of nucleotide substitution for maximum likelihood (ML) [76]. The Akaike information criterion (AIC) [77] was used to select among models instead of the hierarchical likelihood ratio test (hLRT), following Pol [78] and Posada and Buckley [79].
Maximum Likelihood (ML) analyses were performed using RAxML-HPC2 on XSEDE version 8.2.12 [80] on the CIPRES web server [81], with 1000 rapid bootstrap analyses followed by a search for the best-scoring tree in a single run [82]. The nucleotide substitution models “GTRCAT” and “GTRGAMMA” were chosen for the bootstrapping phase for the chloroplast dataset and nuclear dataset, respectively.
3. Results
3.1. GBS Data Analysis Summary
A summary of the sequence data generated from the samples studied is included in Supplementary Material Tables S1–S3. The 190 accessions, which consisted of 186 samples of Rubus and 4 samples of outgroups, were successfully sequenced with the Illumina HiSeq sequencing platform. The mean GC content was 37.85%, which is within the normal range. The sequencing data were high-quality (Q20 ≥ 96.19%, Q30 ≥ 89.54%), and subsequent analyses could be performed. After excluding the outgroups, the average sequencing depth of the Rubus samples ranged from 10.81 to 75.48, with an average of 25.21. The average degree of sequence coverage from the samples of Rubus was 14.51% for at least single-base coverage, while the average degree of coverage of the samples with at least 4-base coverage was 8.68%. A total of 7,293,296 SNPs (nuclear genome) and 8115 SNPs (chloroplast genome) were obtained according to detection with SAMtools software; 256,954 high-quality SNPs (nuclear genome) and 1848 high-quality SNPs (chloroplast genome) were obtained after filtration.
3.2. Phylogenetic Tree Based on SNPs of Nuclear Genome
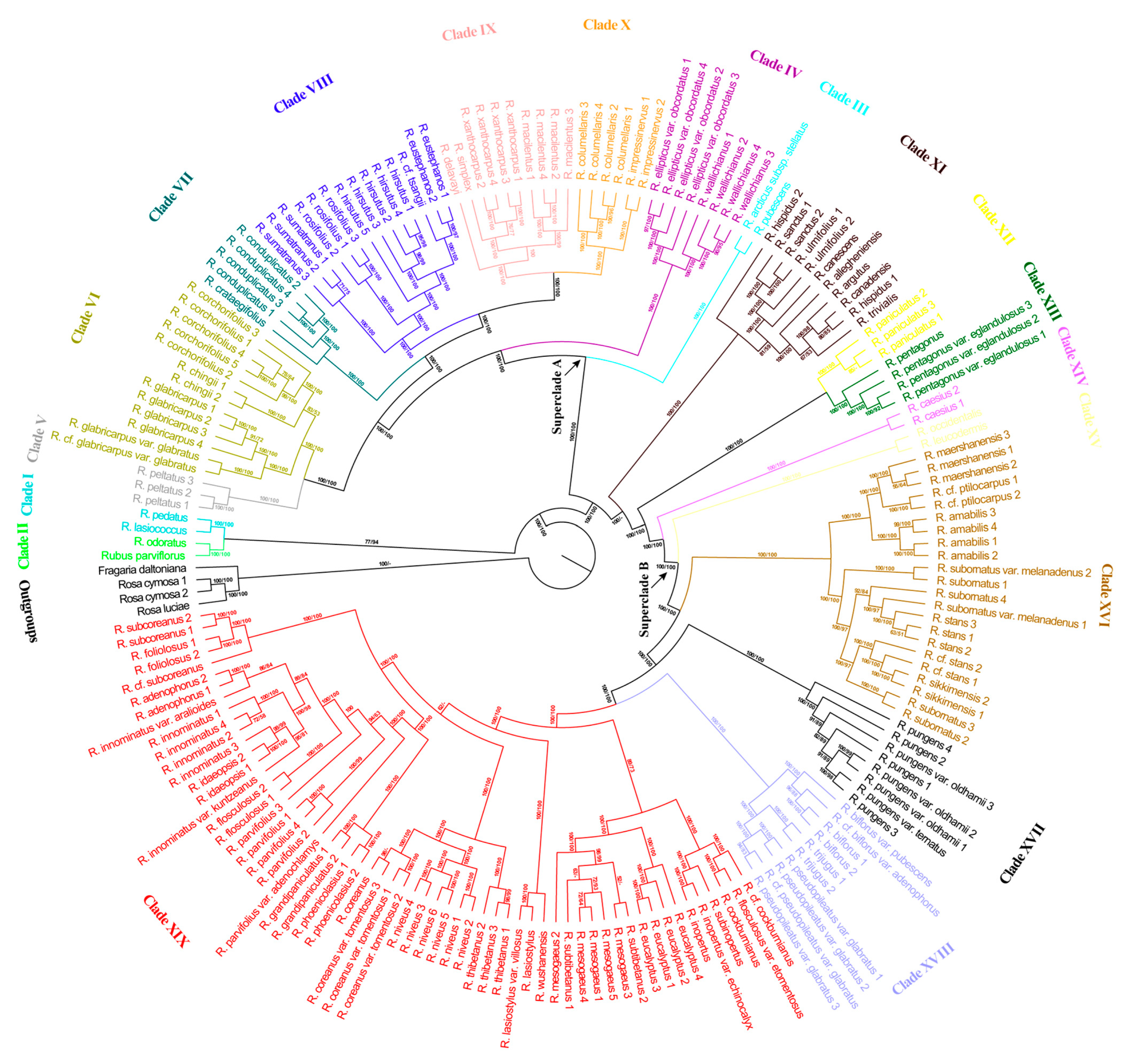
Both maximum likelihood (ML) phylogeny and maximum parsimony (MP) jackknife (JK) analyses based on high-quality SNPs from the nuclear genome showed Rubus to be monophyletic (Figure 1).
ML bootstrap (BS) support and MP JK support are shown on the phylogenetic tree from the ML analysis. Based on our reconstructed nuclear phylogeny and in consideration of morphological characteristics and distribution information, 19 maximumly supported (ML BS = 100%, MP JK = 100%) clades of Rubus were identified in this study (Figure 1). The relationships among most of the 19 clades were also well supported.
The topologies and support values from ML and MP analyses were mostly similar except for the position of Clade XI. The ML analyses resolved Clade XI as sister to the lineage containing Clade XII–Clade XIX with maximum support values. In contrast, the MP analysis resolved Clade XI as sister to the remaining clades except for Clade I and Clade II; support values were also maximum.
Clade I plus Clade II were resolved as monophyletic with 77% ML BS and 94% MP JK support and as sister to the rest of the ingroup.
Except for Clade XI, the basal Clade I plus Clade II, and the predominately polyploid Clade XII, Clade XIII and Clade XIV, the remaining clades formed two strongly supported superclades. Superclade A contained Clade III–Clade X, and Superclade B consisted of Clade XV–Clade XIX.
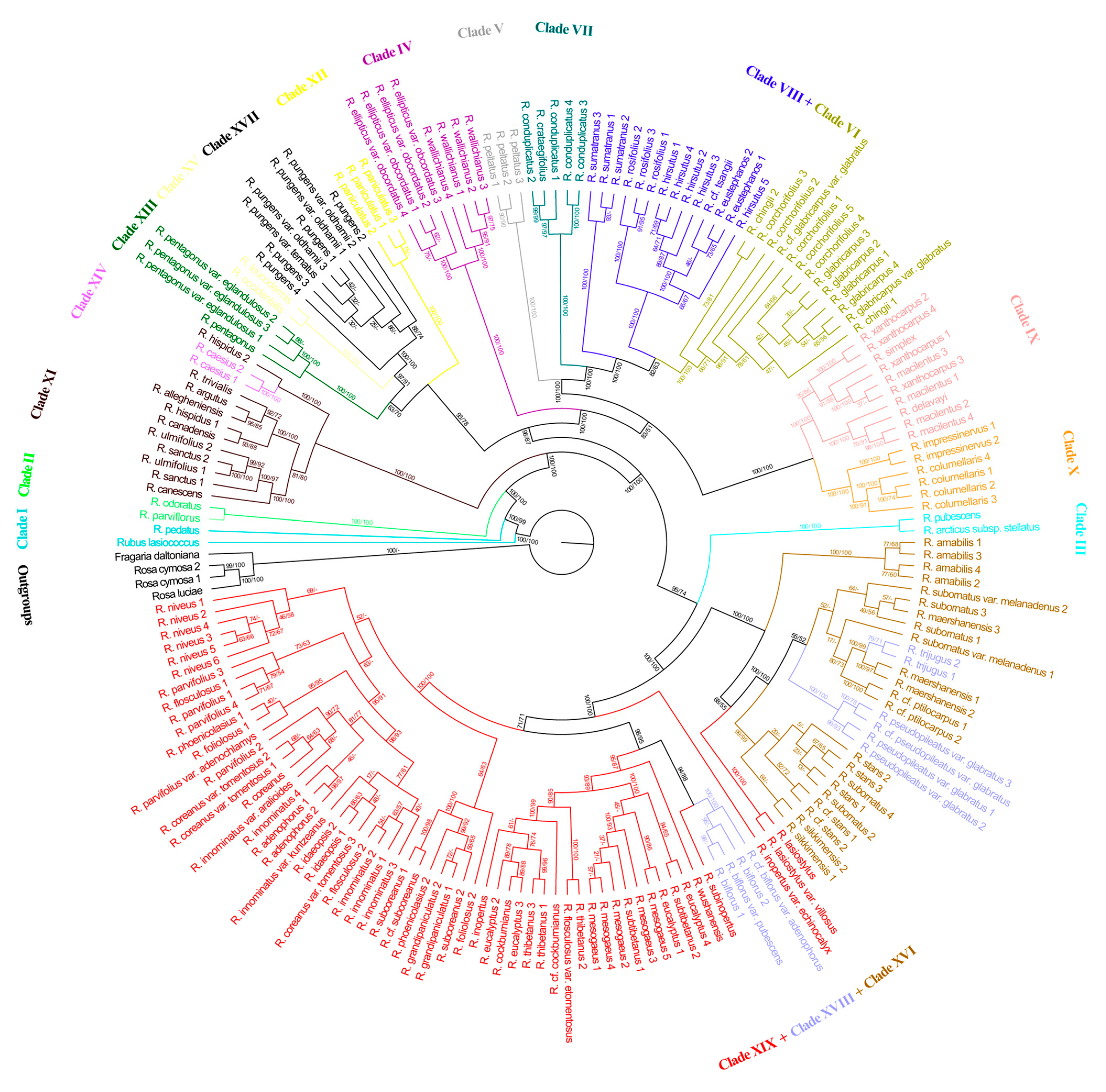
3.3. Phylogenetic Tree Based on SNPs of Chloroplast Genome
The topologies from the ML and MP analyses were similar for the chloroplast phylogeny, which also showed Rubus to be a monophyletic group. However, the topologies and support values based on the chloroplast data were not identical to those based on the nuclear data (Figure 2). First, for the topology of the chloroplast tree, 13 (Clades II, III, IV, V, VI, VII, IX, X, XII, XIII, XIV, XV and XVII) of the 19 major clades identified from the nuclear data were reconfirmed, but the monophyly of 6 clades (Clades I, VIII, XI, XVI, XVIII and XIX) was not supported. Rubus lasiococcus did not cluster with Rubus pedatus. The accessions of Clade VIII identified by nuclear data were split into two clusters on the chloroplast tree, one of which (including Rubus eustephanos, Rubus hirsutus, Rubus rosifolius, etc.) was sister to Clade VI. Clade XVI clustered with a portion of the accessions in Clade XVIII (Rubus pseudopileatus var. glabratus, Rubus trijugus, etc.) and Rubus lasiostylus and R. lasiostylus var. villosus of Clade XIX. Species in the remainder of Clade XIX clustered with some of the accessions in Clade XVIII (Rubus biflorus, etc.). Second, the two superclades were not supported by the chloroplast data. Third, Clade III was sister to the lineage containing Clades XVI, XVIII and XIX, instead of the lineage containing Clades IV to X. Clade XIV nested within Clade XI and was not sister to Superclade B as identified by nuclear data. Clade XVII was more closely related to Clade XV than to Clades XVIII and XIX.
4. Discussion
4.1. Phylogenetic Incongruence between the Chloroplast and Nuclear Phylogenies
The monophyly of Rubus in its current circumscription was maximally supported in this study (Figure 1 and Figure 2) and consistent with the results of earlier studies [1,42,46], further confirming Rubus to be a natural group. Phylogenetic incongruence, however, was detected between the chloroplast and nuclear phylogenies as the topologies based on the chloroplast data were not identical to those based on the nuclear data. The discordance between the nuclear and chloroplast phylogenetic relationships may be due to incomplete lineage sorting, genetic introgression and hybridization.
4.2. Major Evolutionary Lineages within Rubus Revealed in This Study
Within the ingroups, the 186 accessions representing 65 species, 1 subspecies and 17 varieties of Rubus included in the current study were assignable to the following 19 well-supported clades (Figure 1). Those clades are also supported by morphological characteristics.
The clades (Figure 1) can be divided into five main groups and thus may represent several major evolutionary lineages within Rubus, which is somewhat consistent with the results of Carter et al. [42]. The first group, found primarily in North America and containing Clades I and II, represents the basal lineage of Rubus. The second group was composed of Clade XI and mainly includes members of Rubus bearing aggregate fruit that falls together with the receptacle. The third group consists of Superclade A, including Clades III to X. The fourth group contains Clades XIII and XIV and represents the polyploid species of Rubus. The fifth group consists of Superclade B, including Clades XV to XIX. Unfortunately, morphological homoplasy is common in Rubus, thereby making it challenging to distinguish Superclades A and B from the other groups. That is, morphological synapomorphies of the two superclades are unclear at present.
Clade I––Clade I, composed of R. lasiococcus and R. pedatus, was shown to be sister to Clade II with high support (ML BS = 77%, MP JK = 94%; Figure 1). R. lasiococcus is native to North America and R. pedatus occurs naturally in North America and NE Asia. Both species were included in R. subg. Dalibarda by Focke [9,11]. They are morphologically similar in their creeping herbaceous habit, unarmed stems, broad stipules and filiform filaments.
Clade II––Clade II contained two species, Rubus odoratus and Rubus parviflorus, and was sister to Clade I (Figure 1). The sister relationship between R. odoratus and R. parviflorus was consistent with the findings by Carter et al. [42]. The two species, both native to North America and included in R. subg. Anoplobatus by Focke [9,11], are distinguished from other species of Rubus by habit (erect, prickleless shrubs) and their simple palmately lobed or divided leaves. Our study, together with previous studies [1,42], provides strong molecular evidence that R. subg. Dalibarda, R. subg. Anoplobatus and R. subg. Chamaemorus occupy basal positions in the phylogenetic trees, which conflicts with Lu’s [5] hypothesis of R. subg. Idaeobatus being the most primitive group.
Clade III––Clade III contained Rubus arcticus subsp. stellatus and Rubus pubescens and was strongly supported (ML BS = 100%, MP JK = 100%) as sister to the lineage containing Clades IV to X (Figure 1). Clade III to Clade X together form the lineage Superclade A, the morphological synapomorphy of which is unclear at present. R. arcticus subsp. stellatus occurs in North America and the Russian Far East; R. pubescens is in North America. Morphologically, they are somewhat similar to the members of Clade I, but differ from the latter in having dilated, laminar filaments.
Clade IV––Clade IV, composed of eigh samples of Rubus ellipticus var. obcordatus and Rubus wallichianus, was strongly supported (ML BS = 100%, MP JK = 100%) as sister to the clade containing Clades V to X (Figure 1 and Figure 2). The species of Clade IV are endemic to Asia [6,7] and were included in R. subg. Idaeobatus sect. Idaeanthi ser. Elliptici sensu Focke [10,11]. The morphological synapomorphies of Clade IV are shrubs with sparse, curved prickles and dense, spreading reddish brown bristles and usually three-foliolate leaves.
Clade V––This well-supported clade contained only one species, Rubus peltatus of China and Japan. Clade V was sister to Clade VI with maximum support (Figure 1). R. peltatus differs from other species of Rubus in having peltate leaves. Our molecular analysis supported the treatment of Focke [10,11], Yu and Lu [83], Lu and Yu [6] and Naruhashi [84], who placed R. peltatus in the monospecific section R. subg. Idaeobatus sect. Peltati (Lu and Yu’s section and subsection, respectively, almost corresponding to Focke’s subgenus and section).
Clade VI––In our samples, Clade VI contained Rubus corchorifolius, Rubus chingii, Rubus glabricarpus var. glabricarpus and R. glabricarpus var. glabratus. The sister relationship between Clade VI and Clade V was well supported (ML BS = 100%, MP JK = 100%) in our analyses (Figure 1). The four taxa of Clade VI are distributed in Asia and were included in R. sect. Idaeobatus subsect. Corchorifolii by Lu and Yu [6]. Morphologically, species of Clade VI share the following features: flowers usually solitary, leaves simple, stipules adnate to base of petiole, stems usually pilose or glandular (except in R. chingii), aggregate fruit nearly globose and hairy or glabrous. The members of this clade are similar in appearance to taxa included in Clade VII, with the main difference being that the latter have three to several flowers in nearly corymbiform inflorescences and glabrous aggregate fruit. Clade VI can be divided into two subclades. The first contains R. corchorifolius and R. chingii. Species of this subclade lack glandular hairs and the fruits are hairy. Members of the second subclade, represented by R. glabricarpus var. glabricarpus and R. glabricarpus var. glabratus, have glandular hairy pedicels and glabrous fruits.
Clade VII––Clade VII comprised two species—Rubus crataegifolius and Rubus conduplicatus—in our sampling and was strongly supported (ML BS = 100%, MP JK = 100%) as sister to a monophyletic lineage composed of Clades VIII, IX and X (Figure 1). Species of Clade VII are morphologically similar to each other in their nearly corymbiform inflorescences with three-to-several flowers, simple leaves, stipules adnate to the base of the petiole, aggregate fruit nearly globose and glabrous.
R. chingii, R. conduplicatus (Rubus trianthus), R. corchorifolius, R. crataegifolius, R. glabricarpus, Rubus grayanus, Rubus microphyllus, Rubus palmatus and Rubus pseudoacer were all included in R. subg. Idaeobatus sect. Corchorifolii by Focke [10,11], Yu and Lu [83] and Lu and Yu [6], a classification different from the taxonomic treatment of Naruhashi [84] and Nakai [85,86], who further segregated those species into a group with solitary flowers (R. subg. Idaeobatus sect. Villosi sensu Nakai and sect. Corchorifolii sensu Naruhashi) and a group with nearly corymbiform inflorescences (R. subg. Idaeobatus sect. Crataegifolii sensu Nakai and sect. Microphylli sensu Naruhashi). Our data (Figure 1 and Figure 2) suggested that R. corchorifolius, R. chingii, R. glabricarpus and R. glabricarpus var. glabratus, all bearing solitary flowers, form a monophyletic lineage (Clade VI). R. crataegifolius and R. conduplicatus, both bearing nearly corymbiform inflorescences, formed Clade VII. However, Clade VI + Clade VII were not shown to be monophyletic; that is, our results supported the above mentioned classification of Naruhashi [84] and Nakai [85,86], but not those of Focke [10,11], Yu and Lu [83] and Lu and Yu [6].
Clade VIII––In our sample, Clade VIII contained R. eustephanos, R. hirsutus, R. rosifolius, Rubus sumatranus and Rubus cf. tsangii, and was strongly supported (ML BS = 100%, MP JK = 100%) as sister to the monophyletic lineage formed by Clades IX and X (Figure 1). Species of Clade VIII are mainly in Asia. Morphologically, they share a shrubby habit, imparipinnate leaves and ca. 100 or more carpels usually inserted on a stipitate torus. Our molecular evidence supported the classification of Focke [10,11], Yu and Lu [83], Lu and Yu [6] and Naruhashi [84], who placed these species in R. subg. Idaeobatus sect. Rosifolii as a natural group.
Clade IX––Clade IX is composed of four endemic Asian species—Rubus delavayi, Rubus macilentus, Rubus simplex and Rubus xanthocarpus. The sister relationship between Clade IX and Clade X was well supported (ML BS = 100%, MP JK = 100%; Figure 1 and Figure 2). Morphologically, species of Clade IX are characterized by subshrubs or herbs’ leaves with three (or five) leaflets, and the abaxial surface of the calyx is pubescent and with straight needle-like or curved minute prickles (except in R. macilentus where the calyx is unarmed). R. simplex and R. xanthocarpus were both previously included in R. subg. Cylactis by Focke [9,11] for their suffruticose or nearly herbaceous habit. Yu and Lu [83], Lu and Yu [6] and Lu and Boufford [7], noting that the two species are morphologically similar to some members of R. subg. Idaeobatus in non-paniculate inflorescences and that the abaxial surface of the leaflets and aggregate fruit were not tomentose, transferred them to R. sect. Idaeobatus sensu Lu and Yu [6]. Our study (Figure 1 and Figure 2) showed that R. simplex and R. xanthocarpus are more closely related to R. macilentus of R. subg. Idaeobatus than to R. pubescens of R. subg. Cylactis, which supported the classification of Lu and Yu [6] and Lu and Boufford [7].
Clade X––Clade X consisted of four samples of Rubus columellaris and two samples of Rubus impressinervus. It was resolved as sister to the monophyletic Clade IX with maximum support (Figure 1 and Figure 2). The two species are morphologically quite different from the other species of Rubus that we sampled in having approximately leathery leaves. Our study suggests that R. sect. Idaeobatus subsect. Leucanthi sensu Lu and Yu (including R. columellaris, R. delavayi, R. impressinervus, etc.) [6] is paraphyletic and implies that the species with leathery leaves in R. subg. Idaeobatus may be a natural group and that R. delavayi might be excluded from R. subg. Idaeobatus sect. Leucanthi.
Clade XI––Clade XI was composed of Rubus allegheniensis, Rubus argutus, Rubus canadensis, etc. ML analyses of nuclear data indicated that this clade is strongly supported (BS = 100%) as sister to the monophyletic lineage formed by Clade XII–Clade XIX (Figure 1). The morphological characteristics of this clade include shrubs being often prickly, leaves usually ternate, pedately or palmately and quinately compound, persistent narrow stipules which are mostly adnate to the base of the petiole, drupelets remaining on the fleshy receptacle at maturity or falling with the receptacle, or falling separately. Geographically, the members of this clade are mainly American and Eurasian. The eight species of this clade and Rubus caesius (4x) of Clade XIV were all included in R. subg. Eubatus (= subg. Rubus) by Focke [11]. Although the nine species clustered together based on chloroplast data (Figure 2), the phylogenetic analyses based on our nuclear data (Figure 1), ITS [1] and nearly one thousand low copy nuclear genes [42] suggested that R. subg. Rubus is not monophyletic.
Clade XII––Clade XII contained three samples of Rubus paniculatus, one of the few doubtfully diploid species reported to occur in the predominately polyploid R. subg. Malachobatus [27]. This clade was strongly supported (ML BS = 100%, MP JK = 100%) as sister to Clade XIII based on the nuclear data (Figure 1). The remarkable morphological characteristics of Clade XII include shrubs that are often prickly, stems erect, arching or climbing and broad, and free stipules that are caducous or persistent on the twig near the base of the petiole.
Clade XIII––Clade XIII, comprising the Asian endemics Rubus pentagonus var. pentagonus (tetraploid [42]) and R. pentagonus var. eglandulosus, was sister to Clade XII with maximum support (Figure 1). Species of Clade XIII are morphologically distinguishable from other sampled taxa by their shrubby habit and palmately compound leaves with three or five leaflets. The close phylogenetic relationship between R. pentagonus of subg. Idaeobatus and R. paniculatus of subg. Malachobatus is consistent with the findings of Wang et al. [46] and Carter et al. [42] and may provide some support to the hypothesis that R. pentagonus may be one of the possible progenitors of the subg. Malachobatus polyploids [42,46].
Clade XIV––Clade XIV consisted of two samples of R. caesius (Figure 1), a tetraploid of subgenus Rubus occurring naturally from Europe and western Asia to western China. Our chloroplast phylogeny showed that R. caesius was nested within Clade XI (ML BS = 100%, MP JK = 100%; Figure 2), indicating that the maternal parent of R. caesius was likely from R. subg. Rubus, which is consistent with the findings of Carter et al. [42].
Clade XV––Clade XV contained two North American species, Rubus leucodermis and R. occidentalis. The nuclear phylogeny indicated that Clade XV was sister to the lineage formed by Clades XVI through XIX (ML BS = 100%, MP JK = 100%; Figure 1). Clade XV to Clade XIX together form superclade B (ML BS = 100%, MP JK = 100%; Figure 1), the morphological synapomorphy of which is unclear at present. Morphologically, the two species of Clade XV share palmately compound or ternate leaves, nearly black aggregate fruit and drupelets separating from the torus. R. leucodermis, R. occidentalis and two species, Rubus eriocarpus and Rubus glaucus, which were not sampled, were all placed in R. subg. Idaeobatus sect. Idaeanthi ser. Occidentales by Focke [9,11]. In the molecular phylogeny the three diploid species, R. leucodermis, R. occidentalis and R. eriocarpus, were closely related, while the tetraploid R. glaucus aligned with some putative blackberry × raspberry hybrids [42], suggesting this series defined by Focke is not monophyletic.
Clade XVI––Clade XVI contained Rubus amabilis, Rubus maershanensis, Rubus cf. ptilocarpus, etc., in our sampling. It was sister to the lineage containing Clade XVII–Clade XIX with strong support (ML BS = 100%, MP JK = 100%; Figure 1). Species included in Clade XVI are mainly distributed in the Himalayan region. Morphologically they share the following features: shrubs or subshrubs (R. maershanensis), flowers solitary or inflorescences with few flowers (2–6-flowered) and aggregate fruit usually red, glabrous or pubescent, not tomentose. Two well-supported subclades were identified. The first subclade was represented by Rubus subornatus var. subornatus, R. subornatus var. melanadenus, Rubus sikkimensis, Rubus stans and R. cf. stans, all usually with three-foliolate leaves. The second subclade contained R. amabilis, R. maershanensis and R. cf. ptilocarpus and was characterized by having 7–11-foliolate leaves.
Clade XVII––Clade XVII, composed of Rubus pungens var. pungens, R. pungens var. ternatus and R. pungens var. oldhamii, was resolved as sister to the monophyletic lineage formed by Clade XVIII and Clade XIX with maximum support (Figure 1). Generally, species of Clade XVII are widely distributed from Kashmir to Japan and exhibit complex morphological variability: prickles dense to sparse, glandular hairs present or absent and size of leaflets unstable. However, they differ in appearance from the other species of Rubus we sampled: shrubs, stems longer, climbing or trailing, with dense or sparse needle-like prickles (R. pungens var. oldhamii sometimes nearly unarmed), vegetative reproduction mainly by rooting at apex of stem, three–nine-foliolate leaves, abaxial surface of calyx with needle-like prickles, inflorescences terminal or axillary, 1-flowered or corymbose 2- to 4-flowered.
Clade XVIII––Clade XVIII included R. biflorus var. biflorus, R. cf. biflorus var. adenophorus, R. biflorus var. pubescens, R. pseudopileatus var. glabratus, R. cf. pseudopileatus var. glabratus and R. trijugus and was sister to Clade XIX with maximum support (Figure 1). Species contained in Clade XVIII occur in Asia and have the following features: shrubs, branchlets usually pruinose, pedicels distally inflated, leaves thick papyraceous or semi-leathery, aggregate fruit yellow or reddish yellow and densely gray tomentose (tomentum deciduous in R. biflorus and its varieties), flowers 1.5–3 cm in diameter and one to several in corymbose inflorescences.
Clade XIX––Clade XIX was sister to Clade XVIII and contained 20 species and seven varieties (Figure 1). All taxa of Clade XIX are shrubs occurring mainly in Asia. They have pink or purplish red petals (Rubus mesogaeus and Rubus eucalyptus have white or pink petals), which might be interpreted as morphological synapomorphies for this clade. Clade XIX can be divided into four subclades (Figure 1). The first contains Rubus cf. cockburnianus, R. cockburnianus, R. eucalyptus and Rubus flosculosus var. etomentosus, etc. with medium support (ML BS = 89%, MP JK = 73%). Most species of this subclade, except R. eucalyptus and Rubus subinopertus, have nearly purplish black aggregate fruit. R. cf. cockburnianus, R. cockburnianus, R. flosculosus var. etomentosus, together with Rubus inopertus var. inopertus, R. inopertus var. echinocalyx, R. subinopertus, all with 5–11-foliolate leaves, formed a lineage which was sister to the lineage composed of R. eucalyptus, R. mesogaeus and Rubus subtibetanus, which have 3- or 5-foliolate leaves. The second subclade contained R. lasiostylus var. lasiostylus, R. lasiostylus var. villosus and Rubus wushanensis with maximum support. Species of this subclade have broader stipules and bracts that are ovate-lanceolate, ovate or suborbicular. The third subclade contained R. coreanus var. coreanus, R. coreanus var. tomentosus, Rubus niveus and Rubus thibetanus with maximum support. The species of this subclade have 5–11 (or 13)-foliolate leaves and corymbose inflorescences. The fourth subclade contained Rubus adenophorus, R. flosculosus and Rubus foliolosus, etc. Members of this subclade exhibit significant morphological variability: three-foliolate leaves or five- or seven-foliolate leaves, inflorescences nearly corymbose, racemose or paniculate and plants with or without glandular hairs.
According to Focke’s [10,11] classification, most of the species in our study belong to R. subg. Idaeobatus. Our study showed that members of R. subg. Idaeobatus sensu Focke [10,11] are distributed among 13 clades (Clades IV–X, XIII and XV–XIX, Figure 1). The results showing that subgenus Idaeobatus is not monophyletic are consistent with the findings of previous studies [1,21,42,46,47,50,52,53]. Species of R. subg. Idaeobatus sect. Pungentes sensu Focke [10,11] occurring in Clade IX (R. macilentus), Clade XVI (R. amabilis, R. maershanensis, R. sikkimensis, R. stans), Clade XVII (R. pungens), Clade XVIII (R. biflorus) and Clade XIX (R. eucalyptus, R. lasiostylus) indicated that this section is not monophyletic. Similarly, R. subg. Idaeobatus sect. Idaeanthi sensu Focke [10,11] is not monophyletic as species of this section occurred in Clade IV (R. ellipticus var. obcordatus, R. wallichianus), Clade XVI (R. subornatus) and Clade XIX (R. adenophorus, R. cockburnianus, R. flosculosus, Rubus idaeopsis, Rubus innominatus, R. inopertus, R. niveus, Rubus parvifolius, Rubus phoenicolasius, R. thibetanus). R. sect. Idaeobatus subect. Idaeanthi sensu Lu and Yu [6] (R. eucalyptus and R. niveus) is paraphyletic, and R. sect. Idaeobatus subect. Pungentes sensu Lu and Yu [6] is not monophyletic as members of the latter subsection occurred in Clades IV (R. wallichianus), IX (R. simplex, R. xanthocarpus), XVI (R. amabilis, R. maershanensis, R. sikkimensis, R. stans), XVII (R. pungens) and XIX (R. inopertus). R. sect. Idaeobatus subsect. Pileati sensu Lu and Yu [6] is not monophyletic as members of this subsection occurred in Clades XVIII (R. pseudopileatus var. glabratus) and XIX (R. subinopertus). R. sect. Idaeobatus subect. Stimulantes sensu Lu and Yu [6] is not monophyletic as species of this subsection occurred in Clades IV (R. ellipticus var. obcordatus), XVI (R. subornatus), XVIII (R. biflorus) and XIX (R. mesogaeus, R. parvifolius, R. phoenicolasius, R. thibetanus). R. sect. Idaeobatus subect. Thyrsidaei sensu Lu and Yu [6] is paraphyletic as species of this subsection were embedded in Clade XIX (R. adenophorus, R. cockburnianus, R. flosculosus, R. idaeopsis, R. innominatus). These non-monophyletic sections and subsections indicate that morphological characteristics, such as whether inflorescences are paniculate or not, the indument of the abaxial surface of leaves and aggregate fruits, are not suitable for the classification of R. subg. Idaeobatus.
4.3. A Preliminary Classification Scheme of the Diploid Species of Rubus
According to our results, combined with previous phylogenetic analyses [1,21,37,42,46,47,49,50,52,53], a preliminary classification scheme of the diploid species of Rubus (Figure 3) is proposed herein. R. paniculatus, representing R. subg. Malachobatus, was excluded since its ploidy level is uncertain and it was not verified in this study. Before making formal taxonomic and nomenclatural decisions on an infrageneric classification, more samples and molecular data are needed to unravel and confirm the relationships and evolutionary history of these clades.
A preliminary classification scheme of the diploid species of Rubus.
- Group 1
Rubus repens (L.) Kuntze (North America)
- Group 2
R. lasiococcus A. Gray (North America)
R. pedatus Sm. (NE Asia, North America)
- Group 3
Rubus deliciosus Torr. (North America)
R. odoratus L. (North America)
R. parviflorus Nutt. (North America)
Rubus trilobus Ser. (Mesoamerica)
- Group 4
Rubus nivalis Douglas (North America)
- Group 5
R. allegheniensis Porter (North America)
R. argutus Link (North America)
R. canadensis L. (North America)
Rubus canescens DC. (SW Asia, S and C Europe)
Rubus coriifolius Liebm. (Mesoamerican)
Rubus cuneifolius Pursh (North America)
Rubus hispidus L. (North America)
Rubus robustus C. Presl (South America)
Rubus sanctus Schreb. (N Africa, W Asia, SE Europe)
Rubus setosus Bigelow (North America)
Rubus trivialis Michx. (North America)
Rubus ulmifolius Schott (N Africa, Europe)
Rubus urticifolius Poir. (Mexico to S Tropical America)
- Group 6
R. arcticus L. (Eurasia, North America)
R. arcticus L. subsp. stellatus (Sm.) B. Boivin (North America, Russian Far East)
R. pubescens Raf. (North America)
- Group 7
Rubus hawaiensis A. Gray (Hawaii)
Rubus spectabilis Pursh (North America)
- Group 8
R. ellipticus Sm. (Asia)
R. ellipticus Sm. var. obcordatus (Franch.) Focke (Asia)
R. wallichianus Wight et Arn. (Asia)
- Group 9
R. peltatus Maxim. (China, Japan)
- Group 10
R. chingii Hu (China, Japan)
R. corchorifolius L. f. (Asia)
R. glabricarpus Cheng (China)
R. glabricarpus Cheng var. glabratus C. Z. Zheng et Y. Y. Fang (China)
R. palmatus Thunb. (Japan, Korea)
- Group 11
R. conduplicatus Duthie ex J. H. Veitch (China, Japan)
R. crataegifolius Bunge (E Asia)
Rubus trifidus Thunb. (Japan, Korea)
- Group 12
R. columellaris Tutcher (China, Vietnam)
R. impressinervus F. P. Metcalf (China)
- Group 13
R. delavayi Franch. (China)
R. macilentus Jacquem. ex Cambess. (Himalaya)
R. simplex Focke (China)
R. xanthocarpus Bureau et Franch. (China)
- Group 14
Rubus croceacanthus H. Lév. (E and SE Asia)
R. eustephanos Focke (China)
R. hirsutus Thunb. (E Asia)
Rubus illecebrosus Focke (Japan)
R. rosifolius Sm. (Asia)
R. sumatranus Miq. (Asia)
R. tsangii Merr. (China)
- Group 15
Rubus idaeus L. (Africa, Eurasia, North America)
Rubus strigosus Michx. (North America)
- Group 16
R. eriocarpus Liebm. (Mesoamerica)
R. leucodermis Douglas ex Torr. et A. Gray (North America)
R. occidentalis L. (North America)
- Group 17
R. amabilis Focke (China)
R. maershanensis Huan C. Wang et H. Sun (China)
R. sikkimensis Hook. f. (E Himalaya)
R. stans Focke (China)
R. subornatus Focke (China, Myanmar)
R. subornatus Focke var. melanadenus Focke (China)
- Group 18
R. pungens Cambess. (Asia)
R. pungens Cambess. var. oldhamii (Miq.) Maxim. (E Asia)
R. pungens Cambess. var. ternatus Cardot (China)
- Group 19
R. biflorus Buch.-Ham. ex Sm. (Asia)
R. biflorus Buch.-Ham. ex Sm. var. pubescens T. T. Yu et L. T. Lu (China)
R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu (China)
R. trijugus Focke (China)
- Group 20
R. adenophorus Rolfe (China)
R. cockburnianus Hemsl. (China)
R. coreanus Miq. (E Asia)
R. coreanus Miq. var. tomentosus Cardot (China)
R. eucalyptus Focke (China)
R. flosculosus Focke (China)
R. flosculosus Focke var. etomentosus T. T. Yu et L. T. Lu (China)
R. foliolosus D. Don (Asia)
Rubus grandipaniculatus T. T. Yu et L. T. Lu (China)
R. idaeopsis Focke (China)
R. innominatus S. Moore (China)
R. innominatus S. Moore var. aralioides (Hance) T. T. Yu et L. T. Lu (China)
R. innominatus S. Moore var. kuntzeanus (Hemsl.) L. H. Bailey (China)
R. inopertus (Focke) Focke (China, Vietnam)
R. inopertus (Focke) Focke var. echinocalyx Cardot (China)
R. lasiostylus Focke (China)
R. lasiostylus Focke var. villosus Cardot (China)
R. mesogaeus Focke. (Asia)
R. niveus Thunb. (Asia)
R. parvifolius L. (E Asia, Australia)
R. parvifolius L. var. adenochlamys (Focke) Migo (China, Japan)
R. phoenicolasius Maxim. (E Asia)
Rubus subcoreanus T. T. Yu et L. T. Lu (China)
R. subinopertus T. T. Yu et L. T. Lu (China)
R. subtibetanus Hand.-Mazz. (China)
R. thibetanus Franch. (China)
R. wushanensis T. T. Yu et L. T. Lu (China)
Key to the Groups
1a. Flowers of two kinds, essentially petaliferous staminate flowers, usually with pistils abortive, fertile apetalous flowers and with few stamens; drupelets 5–10, nearly dry; low stoloniferous herbs with simple, unlobed, reniform leaves …………….…… Group 1
1b. Flowers of only one kind (in dioecious or polygamodioecious species only slightly dissimilar), mostly petaliferous, sometimes apetalous but bisexual; drupelets many or sometimes several, often fleshy; shrubs, subshrubs or herbs.
2a. Styles club-shaped; stigmas slightly 2-lobed; receptacle flat; unarmed shrubs with exfoliating bark and simple digitately ribbed and lobed leaves ……….………… Group 3
2b. Styles filiform; stigmas capitate; receptacle convex, hemispheric or nipple-shaped; stems unarmed, bristly or prickly; leaves simple or compound.
3a. Stem herbaceous, never prickly, rarely bristly; stipules broad, free or nearly so; floral branches arising directly from rootstock or from stolons.
4a. Filaments filiform, not dilated ……………………………………………… Group 2
4b. Filaments dilated, laminar ……………………………………………..…… Group 6
3b. Stem more or less woody, usually prickly; if unarmed or merely bristly, the stipules narrow and more or less adnate to the petiole.
5a. Stipules broad, nearly free, at junction of stem and petiole; plants with needle-like prickles or bristles; subshrubs, stems creeping ………………………………..…… Group 4
5b. Stipules narrow, linear-lanceolate or subulate, more or less adnate to the petiole; plants with or without needle-like prickles or bristles; shrubs, subshrubs or herblike.
6a. Drupelets remaining on fleshy receptacle at maturity or falling together with receptacle, or falling separately ………………………………………………..………..Group 5
6b. Drupelets united into a thimble-shaped aggregate fruit, falling off entire from dry receptacle.
7a. Flowers solitary or few, large, 2–3.5 cm in diameter; petals pink or magenta, much exceeding sepals; filaments laminar; leaves ternate, papyraceous ………..……… Group 7
7b. Flowers solitary or in inflorescences, small to large, if large (> 3 cm in diameter), the petals white; petals white, pink or purplish; leaves simple or compound, papyraceous or leathery.
8a. Plants with dense, spreading, reddish brown or purplish brown mostly eglandular bristles, 3–7 mm long; aggregate fruit golden yellow or reddish yellow; leaves three-foliolate ……………………………………………………………………………….… Group 8
8b. Plants usually without dense, spreading eglandular bristles; aggregate fruit color varied; leaves simple, 3–11 (or 13)-foliolate.
9a. Leaves leathery ……………………………………………………………… Group 12
9b. Leaves papyraceous or membranous.
10a. Leaves simple.
11a. Leaves peltate; carpels ca. 100 or slightly more; aggregate fruit terete or cylindrical …………………………………………………………………………….………..… Group 9
11b. Leaves not peltate; carpels ca. 10–60, rarely slightly more; aggregate fruit subglobose or ovoid-globose.
12a. Flowers solitary ………………………………………………………….… Group 10
12b. Flowers in nearly corymbiform inflorescences ……………………….… Group 11
10b. Leaves compound.
13a. Carpels ca. 100 or more, usually inserted on stipitate torus…………… Group 14
13b. Carpels 10–70 or slightly more, inserted on sessile torus.
14a. Subshrubs or nearly so; three-foliolate leaves; semi-leathery, terminal leaflet much or slightly longer than lateral leaflets; petals white; fruit orange (R. macilentus with orange or red fruit) …………………………………………………………………… Group 13
14b. Shrubs, if nearly subshrubs, leaflets more than three or petals pink or purplish; leaves 3–11-foliolate; petals white, pink, purplish; fruit red or black or sometimes orange.
15a. Stems scrambling or trailing, 1–3 m long, with dense or sparse needle-like prickles; abaxial surface of calyx with dense, straight needle-like prickles; leaves abaxially pubescent or nearly glabrous ……………………………………….…………….…… Group 18
15b. Stems mostly erect or arching, sometimes scandent or trailing, mostly without needle-like prickles; calyx unarmed or its prickles mostly not needle-like; leaves abaxially tomentose, pubescent or glabrous.
16a. Petals pink or purplish red (R. mesogaeus and R. eucalyptus with white and pink petals); mainly Asian, especially in China ………………………………………… Group 20
16b. Petals white (R. subornatus, R. subornatus var. melanadenus and R. sikkimensis with purplish red petals); occurring in Africa, Eurasia and North America.
17a. Aggregate fruit densely gray tomentose, yellow or reddish yellow; branchlets pruinose; pedicels distally inflated ………………………………………………… Group 19
17b. Aggregate fruit mostly glabrous or pilose, not gray tomentose, dark purplish, black, red, orangish red or sometimes yellow; branchlets pruinose or not.
18a. Aggregate fruit usually dark purplish or black; leaves palmately compound or ternate ………………………………………………………………………………… Group 16
18b. Aggregate fruit red, orangish red, or yellow; leaves pinnately compound or ternate.
19a. Native to Himalayan regions …………………………………………… Group 17
19b. Native to North America (R. strigosus), or Africa, Eurasia and North America (R. idaeus) …………………………………………………………………………….…… Group 15
5. Conclusions
The taxonomy of Rubus is challenging and the phylogenetic relationships within Rubus remain to be clarified. This study inferred the phylogeny of Rubus with emphasis on diploid species based on GBS data with comprehensive taxon sampling. Our results provided useful information for deducing the phylogeny of Rubus, especially providing important insights into the evolution of Rubus in China. We reconfirmed that R. subg. Idaeobatus, recognized by Focke, is not monophyletic. We found that characteristics such as leathery or papyraceous leaves may be of some use in classifying the raspberries. Based on our results, and combined with previous phylogenetic analyses, a preliminary classification scheme of the diploid species of Rubus is proposed here. Further studies, however, including more samples and additional molecular data, are still needed to better unravel and confirm the complicated evolutionary history of Rubus.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14061152/s1, Table S1: Quality statistics of sequencing data of 190 samples; Table S2: Statistics of sequencing depth and coverage of nuclear genome of 190 samples; Table S3: Statistics of sequencing depth and coverage of chloroplast genome of 190 samples.
Author Contributions
Conceptualization: X.-F.G., X.-H.X. and Y.-D.G.; methodology: X.-F.G., Y.-D.G., X.-H.X., B.X. and C.Z.; investigation: X.-H.X. and X.-F.G.; writing—original draft: X.-H.X. and X.-F.G.; writing—review and editing: D.E.B., X.-H.X. and X.-F.G.; supervision: X.-F.G.; funding acquisition: X.-F.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Second Tibetan Plateau Scientific Expedition and Research (STEP) program (Grant No. 2019QZKK0502), the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment, China (Grant No. 2019HJ2096001006), China-Croatia “Belt and Road” Joint Laboratory on Biodiversity and Ecosystem Services (Grant No. 2020YFE0203200) and the Scientific Research Starting Foundation of Mianyang Teachers’ College (Grant No. QD2022A04).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data from this study are available in the manuscript and Supplementary Materials.
Acknowledgments
We thank Li-Bing Zhang, Kim Hummer, Jill Bushakra, Timothee Le Pechon, Bing-Yang Ding, Chen-Chen Zheng, Qi Yu, Xin-Mao Zhou, Meng Li, Wen-Bin Ju, Heng-Ning Deng, Wen-Tai Dai, National Botanic Garden of Belgium (Meise), Missouri Botanical Garden (MO) and the U.S. National Plant Germplasm System (NPGS) for help with samples, and the anonymous reviewers for their helpful comments.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
Table A1.
List of taxa sampled with information related to taxonomy, ploidy and vouchers.
| Taxon | Subgeneric Classification of Focke [9, 10,11] | Sectional Classification of Yu and Lu [83], Lu and Yu [6], and Lu and Boufford [7] | Ploidy Level | Voucher |
|---|---|---|---|---|
| Rubus odoratus L. | R. subg. Anoplobatus | — | 2x | USA: Massachusetts, Northampton, Ano. s. n. (BR, Living Plant Collections Accession 19941489-35) |
| R. parviflorus Nutt. | R. subg. Anoplobatus | — | 2x | USA: W. R. Smith 29200 (MO) |
| R. arcticus L. subsp. stellatus (Sm.) B. Boivin | R. subg. Cylactis | — | 2x | USA: Alaska, Wasilla, K. Hummer et C. I. Wright s. n. (NPGS, Accession PI 606535) |
| R. pubescens Raf. | — | — | 2x | USA: D. Atha 7040 (MO) |
| R. simplex Focke | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Leibo, X. H. Xiong 2036 (CDBI) |
| R. xanthocarpus Bureau et Franch. 1 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1159 (CDBI) |
| R. xanthocarpus Bureau et Franch. 2 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 1316 (CDBI) |
| R. xanthocarpus Bureau et Franch. 3 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 2002 (CDBI) |
| R. xanthocarpus Bureau et Franch. 4 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 2020 (CDBI) |
| R. lasiococcus A. Gray | R. subg. Dalibarda | — | 2x | USA: Oregon, Deschutes County, O. L. Jahn s. n. (NPGS, Accession PI 553659) |
| R. pedatus Sm. | R. subg. Dalibarda | — | 2x | USA: Alaska, Sitka, H. Chambers s. n. (NPGS, Accession PI618503) |
| R. adenophorus Rolfe 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1967 (CDBI) |
| R. adenophorus Rolfe 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1968 (CDBI) |
| R. amabilis Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1146-3 (CDBI) |
| R. amabilis Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1146-7 (CDBI) |
| R. amabilis Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Shimian, X. H. Xiong 1863 (CDBI) |
| R. amabilis Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Wenchuan, X. H. Xiong 1870 (CDBI) |
| R. biflorus Buch.-Ham. ex Sm. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Xiaojin, X. H. Xiong 1860 (CDBI) |
| R. biflorus Buch.-Ham. ex Sm. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Yajiang, X. H. Xiong 2156 (CDBI) |
| R. biflorus Buch.-Ham. ex Sm. var. pubescens T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Sichuan, Muli, X. H. Xiong 1365 (CDBI) |
| R. cf. biflorus Buch.-Ham. ex Sm. var. adenophorus Franch. | — | R. sect. Idaeobatus | NA | China: Xizang, Linzhi, X. H. Xiong 1054 (CDBI) |
| R. chingii Hu 1 | — | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1966 (CDBI) |
| R. chingii Hu 2 | — | R. sect. Idaeobatus | 2x | China: Anhui, Huangshan, X. H. Xiong 831 (CDBI) |
| R. cockburnianus Hemsl. | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Maoxian, X. F. Gao et al. 4983 (CDBI) |
| R. cf. cockburnianus Hemsl. | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Bomi, Y. D. Gao et M. Li YLZB0219 (CDBI) |
| R. columellaris Tutcher 1 | — | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1389 (CDBI) |
| R. columellaris Tutcher 2 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1735 (CDBI) |
| R. columellaris Tutcher 3 | — | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1789 (CDBI) |
| R. columellaris Tutcher 4 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1970 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 1 (R. trianthus Focke) | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Lushan, X. H. Xiong 1648 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Wenzhou, X. H. Xiong 1836-3 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1912 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1964 (CDBI) |
| R. corchorifolius L. f. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Lushan, X. H. Xiong 1634 (CDBI) |
| R. corchorifolius L. f. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1733 (CDBI) |
| R. corchorifolius L. f. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1742 (CDBI) |
| R. corchorifolius L. f. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Wenzhou, X. H. Xiong 1828-2 (CDBI) |
| R. corchorifolius L. f. 5 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1918 (CDBI) |
| R. coreanus Miq. | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Xiong s. n. (CDBI) |
| R. coreanus Miq. var. tomentosus Cardot 1 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wushan, X. H. Xiong 1173 (CDBI) |
| R. coreanus Miq. var. tomentosus Cardot 2 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wushan, X. H. Xiong 1176 (CDBI) |
| R. coreanus Miq. var. tomentosus Cardot 3 | — | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1771 (CDBI) |
| R. crataegifolius Bunge | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Beijing, X. H. Xiong 1991 (CDBI) |
| R. delavayi Franch. | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Yunnan, Shizong, X. H. Xiong 1824 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1427 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, Maguan, X. H. Xiong 1709 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1737 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1758 (CDBI) |
| R. eucalyptus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1304 (CDBI) |
| R. eucalyptus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1839 (CDBI) |
| R. eucalyptus Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Dayi, X. H. Xiong 1856 (CDBI) |
| R. eucalyptus Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1993 (CDBI) |
| R. eustephanos Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1534 (CDBI) |
| R. eustephanos Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1784 (CDBI) |
| R. flosculosus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Xianju, S. Q. Xu 1299 (CDBI) |
| R. flosculosus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Suichang, S. Q. Xu 1344 (CDBI) |
| R. flosculosus Focke var. etomentosus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2014 (CDBI) |
| R. foliolosus D. Don 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 4x | China: Yunnan, Malipo, X. H. Xiong 1712 (CDBI) |
| R. foliolosus D. Don 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 4x | China: Yunnan, X. H. Xiong 1744 (CDBI) |
| R. glabricarpus Cheng 1 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Wenzhou, X. H. Xiong 1838-1 (CDBI) |
| R. glabricarpus Cheng 2 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Wenzhou, X. H. Xiong 1838-3 (CDBI) |
| R. glabricarpus Cheng 3 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1947 (CDBI) |
| R. glabricarpus Cheng 4 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1949 (CDBI) |
| R. glabricarpus Cheng var. glabratus C. Z. Zheng et Y. Y. Fang | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1933 (CDBI) |
| R. cf. glabricarpus Cheng var. glabratus C. Z. Zheng et Y. Y. Fang | — | R. sect. Idaeobatus | NA | China: Fujian, Wuyishan, M. Li LMWYS-45 (CDBI) |
| R. grandipaniculatus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wuxi, X. H. Xiong 2080 (CDBI) |
| R. grandipaniculatus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wuxi, X. H. Xiong 2087 (CDBI) |
| R. hirsutus Thunb. 1 | — | R. sect. Idaeobatus | 2x | China: Zhejiang, Hangzhou, X. H. Xiong 1594 (CDBI) |
| R. hirsutus Thunb. 2 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Lushan, X. H. Xiong 1635 (CDBI) |
| R. hirsutus Thunb. 3 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1734 (CDBI) |
| R. hirsutus Thunb. 4 | — | R. sect. Idaeobatus | 2x | China: Zhejiang, Wenzhou, X. H. Xiong 1837-1 (CDBI) |
| R. hirsutus Thunb. 5 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1983 (CDBI) |
| R. idaeopsis Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1627 (CDBI) |
| R. idaeopsis Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1628 (CDBI) |
| R. impressinervus F. P. Metcalf 1 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1930 (CDBI) |
| R. impressinervus F. P. Metcalf 2 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1935 (CDBI) |
| R. innominatus S. Moore 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Wuxi, Ano. THP-WX-1601 (CDBI) |
| R. innominatus S. Moore 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Chengkou, X. H. Xiong 1220 (CDBI) |
| R. innominatus S. Moore 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 1300 (CDBI) |
| R. innominatus S. Moore 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1788 (CDBI) |
| R. innominatus S. Moore var. aralioides (Hance) T. T. Yu et L. T. Lu | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Jiangxi, Lushan, X. H. Xiong 1645 (CDBI) |
| R. innominatus S. Moore var. kuntzeanus (Hemsl.) L. H. Bailey | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Xuyong, X. H. Xiong 1551 (CDBI) |
| R. inopertus (Focke) Focke | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1985 (CDBI) |
| R. inopertus (Focke) Focke var. echinocalyx Cardot | — | R. sect. Idaeobatus | NA | China: Sichuan, Xuyong, X. H. Xiong 1562 (CDBI) |
| R. lasiostylus Focke | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Xian, C. Zhang et S. Z. Yang zc12087 (CDBI) |
| R. lasiostylus Focke var. villosus Cardot | — | R. sect. Idaeobatus | NA | China: Chongqing, Chengkou, X. H. Xiong 1209 (CDBI) |
| R. leucodermis Douglas ex Hook. | R. subg. Idaeobatus | — | 2x | Canada: British Columbia, H. A. Daubeny s. n. (NPGS, Accession PI 618562) |
| R. macilentus Jacquem. ex Cambess. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1418 (CDBI) |
| R. macilentus Jacquem. ex Cambess. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Shimian, X. H. Xiong 1430 (CDBI) |
| R. macilentus Jacquem. ex Cambess. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1535 (CDBI) |
| R. macilentus Jacquem. ex Cambess. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1763 (CDBI) |
| R. maershanensis Huan C. Wang et H. Sun 1 (R. lutescens Franch.) | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1259 (CDBI) |
| R. maershanensis Huan C. Wang et H. Sun 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Muli, X. H. Xiong 1362 (CDBI) |
| R. maershanensis Huan C. Wang et H. Sun 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1786 (CDBI) |
| R. mesogaeus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1080 (CDBI) |
| R. mesogaeus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Cuona, X. H. Xiong 1575 (CDBI) |
| R. mesogaeus Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Shimian, X. H. Xiong 1861 (CDBI) |
| R. mesogaeus Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 2007 (CDBI) |
| R. mesogaeus Focke 5 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Jinyang, X. H. Xiong 2030 (CDBI) |
| R. niveus Thunb. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 1268 (CDBI) |
| R. niveus Thunb. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Yajiang, X. H. Xiong 1345 (CDBI) |
| R. niveus Thunb. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, Malipo, X. H. Xiong 1718 (CDBI) |
| R. niveus Thunb. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1741 (CDBI) |
| R. niveus Thunb. 5 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1787 (CDBI) |
| R. niveus Thunb. 6 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luding, X. H. Xiong 1862 (CDBI) |
| R. occidentalis L. | R. subg. Idaeobatus | — | 2x | USA: Oregon, Ano. s. n. (NPGS, Accession PI 672663) |
| R. parvifolius L. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Sichuan, Chengdu, X. H. Xiong 1528 (CDBI) |
| R. parvifolius L. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Zhejiang, Hangzhou, X. H. Xiong 1591 (CDBI) |
| R. parvifolius L. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Anhui, Susong, X. H. Xiong 1703 (CDBI) |
| R. parvifolius L. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Sichuan, Luzhou, X. H. Xiong 1804 (CDBI) |
| R. parvifolius L. var. adenochlamys (Focke) Migo | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Shaanxi, Taibai, X. H. Xiong 1333 (CDBI) |
| R. peltatus Maxim. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1938 (CDBI) |
| R. peltatus Maxim. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1942 (CDBI) |
| R. peltatus Maxim. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1987 (CDBI) |
| R. pentagonus Wall. | R. subg. Idaeobatus | R. sect. Idaeobatus | 4x | China: Xizang, Motuo, X. H. Xiong 1409 (CDBI) |
| R. pentagonus Wall. var. eglandulosus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Xizang, Motuo, B. Xu et X. H. Xiong YLZB1151 (CDBI) |
| R. pentagonus Wall. var. eglandulosus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Xizang, Motuo, B. Xu et X. H. Xiong YLZB1174 (CDBI) |
| R. pentagonus Wall. var. eglandulosus T. T. Yu et L. T. Lu 3 | — | R. sect. Idaeobatus | NA | China: Xizang, Motuo, X. H. Xiong 1406 (CDBI) |
| R. phoenicolasius Maxim. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Hanzhong, X. H. Xiong 1322 (CDBI) |
| R. phoenicolasius Maxim. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Taibai, X. H. Xiong 1334 (CDBI) |
| R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 1147-1 (CDBI) |
| R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 1147-27 (CDBI) |
| R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu 3 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 1147-8 (CDBI) |
| R. cf. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Sichuan, Luding, X. H. Xiong 1868 (CDBI) |
| R. cf. ptilocarpus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2003 (CDBI) |
| R. cf. ptilocarpus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2005 (CDBI) |
| R. pungens Cambess. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1162 (CDBI) |
| R. pungens Cambess. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Wushan, X. H. Xiong 1186 (CDBI) |
| R. pungens Cambess. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Chengkou, X. H. Xiong 1200 (CDBI) |
| R. pungens Cambess. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Milin, B. Xu et X. M. Zhou YLZB1613 (CDBI) |
| R. pungens Cambess. var. oldhamii (Miq.) Maxim. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Taibai, X. H. Xiong 1332 (CDBI) |
| R. pungens Cambess. var. oldhamii (Miq.) Maxim. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Gansu, Longnan, X. H. Xiong 1337 (CDBI) |
| R. pungens Cambess. var. oldhamii (Miq.) Maxim. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 2008 (CDBI) |
| R. pungens Cambess. var. ternatus Cardot | — | R. sect. Idaeobatus | NA | China: Chongqing, Chengkou, X. H. Xiong 1197 (CDBI) |
| R. rosifolius Sm. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1732 (CDBI) |
| R. rosifolius Sm. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1913 (CDBI) |
| R. rosifolius Sm. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1975 (CDBI) |
| R. sikkimensis Hook. f. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Motuo, X. H. Xiong 984 (CDBI) |
| R. sikkimensis Hook. f. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Motuo, W. B. Ju et H. N. Deng YLZB0833 (CDBI) |
| R. stans Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, Shangri-la, X. H. Xiong 1996 (CDBI) |
| R. stans Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Jinyang, X. H. Xiong 2021 (CDBI) |
| R. stans Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Muli, X. H. Xiong 2031 (CDBI) |
| R. cf. stans Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Cuona, X. H. Xiong 1577 (CDBI) |
| R. cf. stans Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Milin, X. H. Xiong 1584 (CDBI) |
| R. subcoreanus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Shaanxi, Hanzhong, X. H. Xiong 1327 (CDBI) |
| R. subcoreanus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Shaanxi, Hanzhong, X. H. Xiong 1329 (CDBI) |
| R. cf. subcoreanus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Shaanxi, Hanzhong, X. H. Xiong 1328 (CDBI) |
| R. subinopertus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1407 (CDBI) |
| R. subornatus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1314 (CDBI) |
| R. subornatus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Linzhi, X. H. Xiong 1374 (CDBI) |
| R. subornatus Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Milin, X. H. Xiong 1582 (CDBI) |
| R. subornatus Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Jinyang, X. H. Xiong 2028 (CDBI) |
| R. subornatus Focke var. melanadenus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1264 (CDBI) |
| R. subornatus Focke var. melanadenus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2011 (CDBI) |
| R. subtibetanus Hand.-Mazz. 1 | — | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1762 (CDBI) |
| R. subtibetanus Hand.-Mazz. 2 | — | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1999 (CDBI) |
| R. sumatranus Miq. 1 | R. subg. Malachobatus (1910, erroneous), R. subg. Idaeobatus (1911, correct) | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1392 (CDBI) |
| R. sumatranus Miq. 2 | R. subg. Malachobatus (1910, erroneous), R. subg. Idaeobatus (1912, correct) | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1805 (CDBI) |
| R. sumatranus Miq. 3 | R. subg. Malachobatus (1910, erroneous), R. subg. Idaeobatus (1913, correct) | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1989 (CDBI) |
| R. thibetanus Franch. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Gansu, Longnan, X. H. Xiong 1336 (CDBI) |
| R. thibetanus Franch. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1529 (CDBI) |
| R. thibetanus Franch. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1764 (CDBI) |
| R. trijugus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Danba, X. H. Xiong 2167 (CDBI) |
| R. trijugus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Jinchuan, X. H. Xiong 2175 (CDBI) |
| R. cf. tsangii Merr. | — | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1783 (CDBI) |
| R. wallichianus Wight et Arn. 1 (R. pinfaensis H. Lév. et Vaniot) | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1743 (CDBI) |
| R. wallichianus Wight et Arn. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1785 (CDBI) |
| R. wallichianus Wight et Arn. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1799 (CDBI) |
| R. wallichianus Wight et Arn. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Guizhou, X. H. Xiong 1820 (CDBI) |
| R. wushanensis T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Chongqing, Wushan, X. H. Xiong 1183 (CDBI) |
| R. paniculatus Sm. 1 | R. subg. Malachobatus | R. sect. Malachobatus | 2x | China: Xizang, Motuo, X. H. Xiong 966 (CDBI) |
| R. paniculatus Sm. 2 | R. subg. Malachobatus | R. sect. Malachobatus | 2x | China: Xizang, Motuo, X. H. Xiong 969 (CDBI) |
| R. paniculatus Sm. 3 | R. subg. Malachobatus | R. sect. Malachobatus | 2x | China: Xizang, Motuo, X. H. Xiong 971 (CDBI) |
| R. allegheniensis Porter | R. subg. Rubus | — | 2x, 3x, 4x | USA: Pennsylvania, Lehighton, T. A. Merkel s. n. (NPGS, Accession PI 553970) |
| R. argutus Link | R. subg. Rubus | — | 2x, 3x | USA: South Carolina, J. R. Ballington et J. A. Payne s. n. (NPGS, Accession PI 606490) |
| R. caesius L. 1 | R. subg. Rubus | R. sect. Rubus | 4x | Russia: Stavropol, H. J. Brooks s. n. (NPGS, Accession PI 324058) |
| R. caesius L. 2 | R. subg. Rubus | R. sect. Rubus | 4x | Spain: Zaragoza, Erickson s. n. (NPGS, Accession PI 420407) |
| R. canadensis L. | R. subg. Rubus | — | 2x, 3x | USA: Georgia, J. R. Ballington s. n. (NPGS, Accession PI 553141) |
| R. canescens DC. | R. subg. Rubus | — | 2x | Former Serbia and Montenegro: L. Aladzajkov s. n. (NPGS, Accession PI 379537) |
| R. hispidus L. 1 | R. subg. Rubus | — | 2x | USA: North Carolina, Beaufort County, J. R. Ballington s. n. (NPGS, Accession PI 606507) |
| R. hispidus L. 2 | R. subg. Rubus | — | 2x | USA: Tennessee, Cumberland County, Ano. s. n. (NPGS, Accession PI 618281) |
| R. sanctus Schreb. 1 | R. subg. Rubus | — | 2x | Pakistan: Chitral District, D. Brenner et M. Thompson s. n. (NPGS, Accession PI 553879) |
| R. sanctus Schreb. 2 | R. subg. Rubus | — | 2x | Syria: Damascus, M. Thompson s. n. (NPGS, Accession PI 553930) |
| R. trivialis Michx. | R. subg. Rubus | — | 2x | USA: Texas, Austin, L. Whiting s. n. (NPGS, Accession PI 553704) |
| R. ulmifolius Schott 1 | R. subg. Rubus | — | 2x | Syria: Damascus, M. Thompson s. n. (NPGS, Accession PI 553931) |
| R. ulmifolius Schott 2 | R. subg. Rubus | — | 2x | USA: California, Napa County, L. Clark s. n. (NPGS, Accession PI 672650) |
| Outgroups | ||||
| Fragaria daltoniana J. Gay | 2x | China: Xizang, Milin, X. H. Xiong 1590 (CDBI) | ||
| Rosa cymosa Tratt. 1 | 2x | China: Jiangxi, Dayu, X. H. Xiong 803-2 (CDBI) | ||
| R. cymosa Tratt. 2 | 2x | China: Jiangxi, Xingan, X. H. Xiong 813-1 (CDBI) | ||
| R. luciae Franch. et Rochebr. | 2x | China: Zhejiang, Wenzhou, C. Zhang s. n. (CDBI) |
References
- Alice, L.A.; Campbell, C.S. Phylogeny of Rubus (Rosaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences. Am. J. Bot. 1999, 86, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Alice, L.A.; Goldman, D.H.; Macklin, J.A.; Moore, G. Rubus L. In Flora of North America North of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2014; Volume 9, pp. 28–56. [Google Scholar]
- Bailey, L.H. Gentes Herbarum: Rubus in North America; Ithaca: New York, NY, USA, 1945; Volume 5, pp. 1–932. [Google Scholar]
- Thuan, N. Flore du Cambodge du Laos et du Vietnam; Fascicule 7; Museum National D’Histoire Naturelle: Paris, France, 1968; pp. 1–83. [Google Scholar]
- Lu, L.T. A study on the genus Rubus of China. Acta Phytotax. Sin. 1983, 21, 13–25. [Google Scholar]
- Lu, L.T.; Yu, T.T. Rubus L. In Flora Reipublicae Popularis Sinicae; Yu, T.T., Ed.; Science Press: Beijing, China, 1985; Volume 37, pp. 10–218. [Google Scholar]
- Lu, L.T.; Boufford, D.E. Rubus L. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2003; Volume 9, pp. 195–285. [Google Scholar]
- Abrams, L. Illustrated Flora of the Pacific States, Washington, Oregon, and California; Stanford University Press: Stanford, CA, USA, 1944; Volume Ⅱ, pp. 454–458. [Google Scholar]
- Focke, W.O. Species Ruborum, monographiae generis Rubi prodromus, Pars I. Biblioth. Bot. 1910, 17, 1–120. [Google Scholar]
- Focke, W.O. Species Ruborum, monographiae generis Rubi Prodromus, Pars II. Biblioth. Bot. 1911, 17, 121–223. [Google Scholar]
- Focke, W.O. Species Ruborum, monographiae generis Rubi Prodromus, Pars III. Biblioth. Bot. 1914, 19, 1–274. [Google Scholar]
- Royen, P. The Alpine Flora of New Guinea; Strauss et Cramer: Hirschberg, Germany, 1983; Volume 4, pp. 2458–2490. [Google Scholar]
- Thompson, M.M. Survey of chromosome numbers in Rubus (Rosaceae: Rosoideae). Ann. Mo. Bot. Gard. 1997, 84, 128–164. [Google Scholar] [CrossRef]
- Howarth, D.G.; Gardner, D.E.; Morden, C.W. Phylogeny of Rubus subgenus Idaeobatus (Rosaceae) and its implications toward colonization of the Hawaiian Islands. Syst. Bot. 1997, 22, 433–441. [Google Scholar] [CrossRef]
- Gustafsson, A. The genesis of the European blackberry flora. Lunds Univ. Arsskrift. 1943, 39, 1–200. [Google Scholar]
- Aalders, L.E.; Hall, I.V. A cytotaxonomic survey of the native blackberries of Nova Scotia. Canad. J. Genet. Cytol. 1966, 8, 528–532. [Google Scholar] [CrossRef]
- Richards, A.J.; Kirschner, J.; Stepanek, J.; Marhold, K. Apomixis and taxonomy: An introduction. Folia Geobot. Phytotax. 1996, 31, 281–282. [Google Scholar] [CrossRef]
- Weber, H.E. Former and modern taxonomic treatment of the apomictic Rubus complex. Folia Geobot. Phytotax. 1996, 31, 373–380. [Google Scholar] [CrossRef]
- Robertson, K.R. The genera of Rosaceae in the southeastern United States. J. Arnold Arbor. 1974, 55, 352–360. [Google Scholar] [CrossRef]
- Thompson, M.M. Chromosome numbers of Rubus species at the National Clonal Germplasm Repository. Hortscience 1995, 30, 1447–1452. [Google Scholar] [CrossRef]
- Alice, L.A.; Dodson, T.M.; Sutherland, B.L. Diversity and relationships of Bhutanese Rubus. Acta Hort. 2008, 777, 63–69. [Google Scholar] [CrossRef]
- Waugh, R.; van de Ven, W.T.G.; Phillips, M.S.; Powell, W. Chloroplast DNA diversity in the genus Rubus (Rosaceae) revealed by Southern hybridization. Plant Syst. Evol. 1990, 172, 65–75. [Google Scholar] [CrossRef]
- Gustafsson, A. Chromosomenzahlen in der gattung Rubus. Hereditas 1933, 18, 77–80. [Google Scholar] [CrossRef]
- Gustafsson, A. Differential polyploidy within the blackberries. Hereditas 1939, 25, 33–47. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y. Cytological studies in the Genus Rubus L. Ⅰ. Chromosome numbers in the British Rubus Flora. New Phytol. 1953, 52, 22–39. [Google Scholar] [CrossRef]
- Bammi, R.K. Complement fractionation in a natural hybrid between Rubus procerus Muell. and R. laciniatus Willd. Nature 1965, 208, 608. [Google Scholar] [CrossRef]
- Malik, C.P. Cytology of some Indian species of Rosaceae. Caryologia 1965, 18, 139–149. [Google Scholar] [CrossRef]
- Nybom, H. Chromosome numbers in Rubus species from Sri Lanka. Bot. Not. 1980, 133, 47–48. [Google Scholar]
- Nybom, H. Chromosome numbers and reproduction in Rubus subgen. Malachobatus. Plant Syst. Evol. 1986, 152, 211–218. [Google Scholar] [CrossRef]
- Dale, A.; Ingram, R. Chromosome numbers of some South-American blackberries. Hort. Res. 1981, 21, 107. [Google Scholar]
- Subramanian, D. Cytotaxonomic studies of South Indian Rosaceae. Cytologia 1987, 52, 395–403. [Google Scholar] [CrossRef]
- Thompson, M.M.; Zhao, C.M. Chromosome numbers of Rubus species in Southwest China. Acta Hort. 1993, 352, 493–502. [Google Scholar] [CrossRef]
- Lin, S.H.; Zhang, B.B.; Fang, C.Q.; Lin, F.Q.; Pu, F.S. Chromosome numbers and karyotypes of 8 Rubus species in China. Acta Hort. Sin. 1994, 21, 313–319. [Google Scholar]
- Iwatsubo, Y.; Naruhashi, N.; Weber, H.E. Chromosome numbers of European blackberries (Rubus subg Rubus, Rosaceae). Plant Syst. Evol. 1995, 198, 143–149. [Google Scholar] [CrossRef]
- Meng, R.G.; Finn, C. Determining ploidy level and nuclear DNA content in Rubus by flow cytometry. J. Am. Soc. Hortic. Sci. 2002, 127, 767–775. [Google Scholar] [CrossRef]
- Naruhashi, N.; Iwatsubo, Y.; Peng, C.I. Chromosome numbers in Rubus (Rosaceae) of Taiwan. Bot. Bull. Acad. Sin. 2002, 43, 193–201. [Google Scholar] [CrossRef]
- Michael, K. Clarification of Basal Relationships in Rubus (Rosaceae) and the Origin of Rubus chamaemorus. Master’s Thesis, Western Kentucky University, Bowling Green, KY, USA, 2006. [Google Scholar]
- Wang, X.R.; Tang, H.R.; Duan, J.; Li, L. A comparative study on karyotypes of 28 taxa in Rubus sect. Idaeobatus and sect. Malachobatus (Rosaceae) from China. J. Syst. Evol. 2008, 46, 505–515. [Google Scholar] [CrossRef]
- Wang, X.R.; Tang, H.R.; Fu, H.Q.; Luo, Y.; Deng, Q.X.; Dong, X.L.; Li, L.; Duan, J. Chromosome numbers and karyotypes of 10 wild bramble (Rubus L.) species from Southwest of China. Acta Hort. Sin. 2008, 35, 343–350. [Google Scholar]
- Krahulcova, A.; Travnicek, B.; Sarhanova, P. Karyological variation in the genus Rubus subgenus Rubus: New data from the Czech Republic and synthesis of the current knowledge of European species. Preslia 2013, 85, 19–39. [Google Scholar]
- Wang, Y.; Wang, X.R.; Chen, Q.; Zhang, L.; Tang, H.R.; Luo, Y.; Liu, Z.J. Phylogenetic insight into subgenera Idaeobatus and Malachobatus (Rubus, Rosaceae) inferring from ISH analysis. Mol. Cytogenet. 2015, 8, 11–23. [Google Scholar] [CrossRef]
- Carter, K.A.; Liston, A.; Bassil, N.V.; Alice, L.A.; Bushakra, J.M.; Sutherland, B.L.; Mockler, T.C.; Bryant, D.W.; Hummer, K.E. Target capture sequencing unravels Rubus evolution. Front. Plant Sci. 2019, 10, 1615. [Google Scholar] [CrossRef]
- Bean, A.R. A revision of Rubus subg. Malachobatus (Focke) Focke and Rubus subg. Diemenicus A. R. Bean (Rosaceae) in Australia. Austrobaileya 1997, 5, 39–51. [Google Scholar]
- Kalkman, C. The genus Rubus (Rosaceae) in Malesia 3. The subgenus Micranthobatus. Blumea 1987, 32, 323–341. [Google Scholar]
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.H.; Smedmark, J.E.E.; Campbell, C.S. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Chen, T.; Tang, H.R.; Liu, L.; Wang, X.R. Phylogenetic insights into Chinese Rubus (Rosaceae) from multiple chloroplast and nuclear DNAs. Front. Plant Sci. 2016, 7, 968. [Google Scholar] [CrossRef]
- Morden, C.W.; Gardner, D.E.; Weniger, D.A. Phylogeny and biogeography of Pacific Rubus subgenus Idaeobatus (Rosaceae) species: Investigating the origin of the endemic Hawaiian raspberry R. macraei. Pac. Sci. 2003, 57, 181–197. [Google Scholar] [CrossRef]
- Stafne, E.T.; Clark, J.R.; Szalanski, A.L. Nuclear ribosomal ITS region sequences for differentiation of Rubus genotypes. J. Ark. Acad. Sci. 2003, 57, 176–180. [Google Scholar]
- Sutherland, B. Phylogenetics of Rubus ursinus and R. macraai (Rosaceae): Evidence of Hybrid Origin. Master’s Thesis, Western Kentucky University, Bowling Green, KY, USA, 2005. [Google Scholar]
- Yang, J.Y.; Pak, J.H. Phylogeny of Korean Rubus (Rosaceae) based on ITS (nrDNA) and trn L/F intergenic region (cpDNA). J. Plant Biol. 2006, 49, 44–54. [Google Scholar] [CrossRef]
- Imanishi, H.; Nakahara, K.; Tsuyuzaki, H. Genetic relationships among native and introduced Rubus species in Japan based on rbcL sequence. Acta Hort. 2008, 769, 195–199. [Google Scholar] [CrossRef]
- Wang, Y.N. Relationships among Rubus (Rosaceae) Species Used in Traditional Chinese Medicine. Master’s Dissertation, Western Kentucky University, Bowling Green, KY, USA, 2011. [Google Scholar]
- Yang, J.Y.; Yoon, H.S.; Pak, J.H. Phylogeny of Korean Rubus (Rosaceae) based on the second intron of the LEAFY gene. Can. J. Plant Sci. 2012, 92, 461–472. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Chen, Q.; Luo, Y.; Zhang, Y.; Tang, H.R.; Wang, X.R. Phylogenetic utility of Chinese Rubus (Rosaceae) based on ndhF sequence. Acta Hort. Sin. 2015, 42, 19–30. [Google Scholar] [CrossRef]
- Sochor, M.; Trávníček, B. Melting pot of biodiversity: First insights into the evolutionary patterns of the Colchic bramble flora (Rubus subgenus Rubus, Rosaceae). Bot. J. Linn. Soc. 2016, 181, 610–620. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, É.; Laroche, J.; Larose, S.; Jean, M.; et al. An improved genotyping by sequencing (GBS) approach offering increased versatility and efficiency of SNP discovery and genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef]
- Arbizu, C.I.; Ellison, S.L.; Senalik, D.; Simon, P.W.; Spooner, D.M. Genotyping-by-sequencing provides the discriminating power to investigate the subspecies of Daucus carota (Apiaceae). BMC Evol. Biol. 2016, 16, 234. [Google Scholar] [CrossRef]
- Ariani, A.; Berny Mier y Teran, J.C.; Gepts, P. Genome-wide identification of SNPs and copy number variation in common bean (Phaseolus vulgaris L.) using genotyping-by-sequencing (GBS). Mol. Breed. 2016, 36, 87. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Chong, C.; Bragg, J.G.; Ong, C.R.; Aitken, N.C.; Chuah, A.; Lepschi, B.; Borevitz, J.O. Population and phylogenomic decomposition via genotyping-by-sequencing in Australian Pelargonium. Mol. Ecol. 2016, 25, 2000–2014. [Google Scholar] [CrossRef]
- Stetter, M.G.; Schmid, K.J. Analysis of phylogenetic relationships and genome size evolution of the Amaranthus genus using GBS indicates the ancestors of an ancient crop. Mol. Phylogenet. Evol. 2017, 109, 80–92. [Google Scholar] [CrossRef]
- Wang, X.K.; Jiang, D.; Sun, Z.Z. Study on phylogeny of 240 mandarin accessions with Genotyping-by-Sequencing technology. Sci. Agric. Sin. 2017, 50, 1666–1673. [Google Scholar] [CrossRef]
- Zhao, P.; Zhou, H.J.; Potter, D.; Hu, Y.H.; Feng, X.J.; Dang, M.; Feng, L.; Zulfiqar, S.; Liu, W.Z.; Zhao, G.F.; et al. Population genetics, phylogenomics and hybrid speciation of Juglans in China determined from whole chloroplast genomes, transcriptomes, and genotyping-by-sequencing (GBS). Mol. Phylogenet. Evol. 2018, 126, 250–265. [Google Scholar] [CrossRef]
- Morigengaowa; Shang, H.; Liu, B.D.; Kang, M.; Yan, Y.H. One or more species? GBS sequencing and morphological traits evidence reveal species diversification of Sphaeropteris brunoniana in China. Biodivers. Sci. 2019, 27, 1196–1204. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Sun, B.; Chen, T.; Luo, Y.; Zhang, Y.; Wang, X.R.; Tang, H.R. The complete chloroplast genome sequence of Rubus coreanus, an excellent diseases-resistant resource. Mitochondrial DNA B Resour. 2019, 4, 216–217. [Google Scholar] [CrossRef]
- VanBuren, R.; Wai, C.M.; Colle, M.; Wang, J.; Sullivan, S.; Bushakra, J.M.; Liachko, I.; Vining, K.J.; Dossett, M.; Finn, C.E.; et al. A near complete, chromosome-scale assembly of the black raspberry (Rubus occidentalis) genome. GigaScience 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup, The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Farris, J.S.; Albert, V.A.; Källersjö, M.; Lipscomb, D.; Kluge, A.G. Parsimony jackknifing outperforms neighbor-joining. Cladistics 1996, 12, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Maximum likelihood and minimum-steps methods for estimating evolutionary trees from data on discrete characters. Syst. Biol. 1973, 22, 240–249. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Pol, D. Empirical problems of the hierarchical likelihood ratio test for model selection. Syst. Biol. 2004, 53, 949–962. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA, 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Yu, T.T.; Lu, L.T. Taxa nova Rosacearum sinicarum (Ⅳ). Acta Phytotaxon. Sin. 1982, 20, 295–310. [Google Scholar]
- Naruhashi, N. Rubus L. In Flora of Japan; Iwatsuki, K., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 2001; Volume Ⅱb, pp. 145–169. [Google Scholar]
- Nakai, T. Praecursores ad Floram Sylvaticam Coreanam VII. (Rosaceae). Bot. Mag. 1916, 30, 217–242. [Google Scholar] [CrossRef]
- Nakai, T. Flora Sylvatica Koreana; Pars VII; The government of Chosen: Seoul, Republic of Korea, 1918; pp. 48–77. [Google Scholar]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)—A community resource of plant chromosome numbers. New Phytol. 2015, 206, 19–26. [Google Scholar] [CrossRef]
- Holmgren, P.K.; Holmgren, N.H. Index Herbariorum, A Global Directory of Public Herbaria and Associated Staff [Continuously Updated]; New York Botanical Garden: New York, NY, USA, 1998. [Google Scholar]
Figure 1.
Maximum likelihood phylogeny of Rubus based on SNPs of nuclear genome. Maximum likelihood bootstrap support values (ML BS) and maximum parsimony jackknife support values (MP JK) are shown along the branches (symbol “-” indicates that the relationship or clade is not supported by MP analysis).
Figure 1.
Maximum likelihood phylogeny of Rubus based on SNPs of nuclear genome. Maximum likelihood bootstrap support values (ML BS) and maximum parsimony jackknife support values (MP JK) are shown along the branches (symbol “-” indicates that the relationship or clade is not supported by MP analysis).

Figure 2.
Maximum likelihood phylogeny of Rubus based on SNPs of chloroplast genome. Maximum likelihood bootstrap support values (ML BS) and maximum parsimony jackknife support values (MP JK) are shown along the branches (symbol “-” indicates that the relationship or clade is not supported by MP analysis).
Figure 2.
Maximum likelihood phylogeny of Rubus based on SNPs of chloroplast genome. Maximum likelihood bootstrap support values (ML BS) and maximum parsimony jackknife support values (MP JK) are shown along the branches (symbol “-” indicates that the relationship or clade is not supported by MP analysis).

Figure 3.
Representative species of Rubus from the 20 major lineages of the preliminary classification scheme proposed in the current study. (A) R. repens (Group 1, cited from Native Plant Trust, photo credit: Donald Cameron); (B) R. pedatus (Group 2, cited from Oregon Flora Image Project, photo credit: Gerald D. Carr); (C) R. odoratus (Group 3, photo credit: Sally Wasowski and Andy Wasowski, Lady Bird Johnson Wildflower Center); (D) R. nivalis (Group 4, cited from GBIF, photo credit: Tyson Ehlers); (E) R. ulmifolius (Group 5, cited from Flora-On, photo credit: Carlos Aguiar); (F) R. arcticus (Group 6, cited from Saxifaga Foundation–Images of European biodiversity, photo credit: Dirk Hilbers); (G) R. spectabilis (Group 7, cited from Oregon Flora Image Project, photo credit: Gerald D. Carr); (H) R. ellipticus var. obcordatus (Group 8); (I) R. peltatus (Group 9); (J) R. corchorifolius (Group 10); (K) R. conduplicatus (Group 11); (L) R. columellaris (Group 12); (M) R. xanthocarpus (Group 13); (N) R. sumatranus (Group 14); (O) R. idaeus (Group 15, cited from Saxifraga Foundation–Images of European biodiversity, photo credit: Jan Van der Straaten); (P) R. occidentalis (Group 16, cited from Discover Life, photo credit: Steven J. Baskauf); (Q) R. subornatus (Group 17); (R) R. pungens (Group 18); (S) R. biflorus (Group 19); (T) R. mesogaeus (Group 20); (U) R. niveus (Group 20); (V) R. parvifolius (Group 20).
Figure 3.
Representative species of Rubus from the 20 major lineages of the preliminary classification scheme proposed in the current study. (A) R. repens (Group 1, cited from Native Plant Trust, photo credit: Donald Cameron); (B) R. pedatus (Group 2, cited from Oregon Flora Image Project, photo credit: Gerald D. Carr); (C) R. odoratus (Group 3, photo credit: Sally Wasowski and Andy Wasowski, Lady Bird Johnson Wildflower Center); (D) R. nivalis (Group 4, cited from GBIF, photo credit: Tyson Ehlers); (E) R. ulmifolius (Group 5, cited from Flora-On, photo credit: Carlos Aguiar); (F) R. arcticus (Group 6, cited from Saxifaga Foundation–Images of European biodiversity, photo credit: Dirk Hilbers); (G) R. spectabilis (Group 7, cited from Oregon Flora Image Project, photo credit: Gerald D. Carr); (H) R. ellipticus var. obcordatus (Group 8); (I) R. peltatus (Group 9); (J) R. corchorifolius (Group 10); (K) R. conduplicatus (Group 11); (L) R. columellaris (Group 12); (M) R. xanthocarpus (Group 13); (N) R. sumatranus (Group 14); (O) R. idaeus (Group 15, cited from Saxifraga Foundation–Images of European biodiversity, photo credit: Jan Van der Straaten); (P) R. occidentalis (Group 16, cited from Discover Life, photo credit: Steven J. Baskauf); (Q) R. subornatus (Group 17); (R) R. pungens (Group 18); (S) R. biflorus (Group 19); (T) R. mesogaeus (Group 20); (U) R. niveus (Group 20); (V) R. parvifolius (Group 20).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, X.-F.; Xiong, X.-H.; Boufford, D.E.; Gao, Y.-D.; Xu, B.; Zhang, C. Phylogeny of the Diploid Species of Rubus (Rosaceae). Genes 2023, 14, 1152. https://doi.org/10.3390/genes14061152
AMA Style
Gao X-F, Xiong X-H, Boufford DE, Gao Y-D, Xu B, Zhang C. Phylogeny of the Diploid Species of Rubus (Rosaceae). Genes. 2023; 14(6):1152. https://doi.org/10.3390/genes14061152
Chicago/Turabian StyleGao, Xin-Fen, Xian-Hua Xiong, David E. Boufford, Yun-Dong Gao, Bo Xu, and Cheng Zhang. 2023. "Phylogeny of the Diploid Species of Rubus (Rosaceae)" Genes 14, no. 6: 1152. https://doi.org/10.3390/genes14061152
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.