
The Genetic Diversity and Genetic Structure of the Germplasm Resources of the Medicinal Orchid Plant Habenaria dentata


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Methodology
2.2.1. Extraction and Quality Assay of the Total DNA
2.2.2. Synthesis and Screening of the Inter-Simple Sequence Repeat Primers
2.2.3. Inter-Simple Sequence Repeat Polymerase Chain Reaction System and Procedure
2.2.4. Data Processing
3. Results
3.1. Detection of the Total DNA Quality
3.2. Primer Screening
3.3. Inter-Simple Sequence Repeat Genetic Diversity Analysis
3.3.1. Analysis of the Genetic Diversity Parameters
3.3.2. Population Genetic Structure Analysis
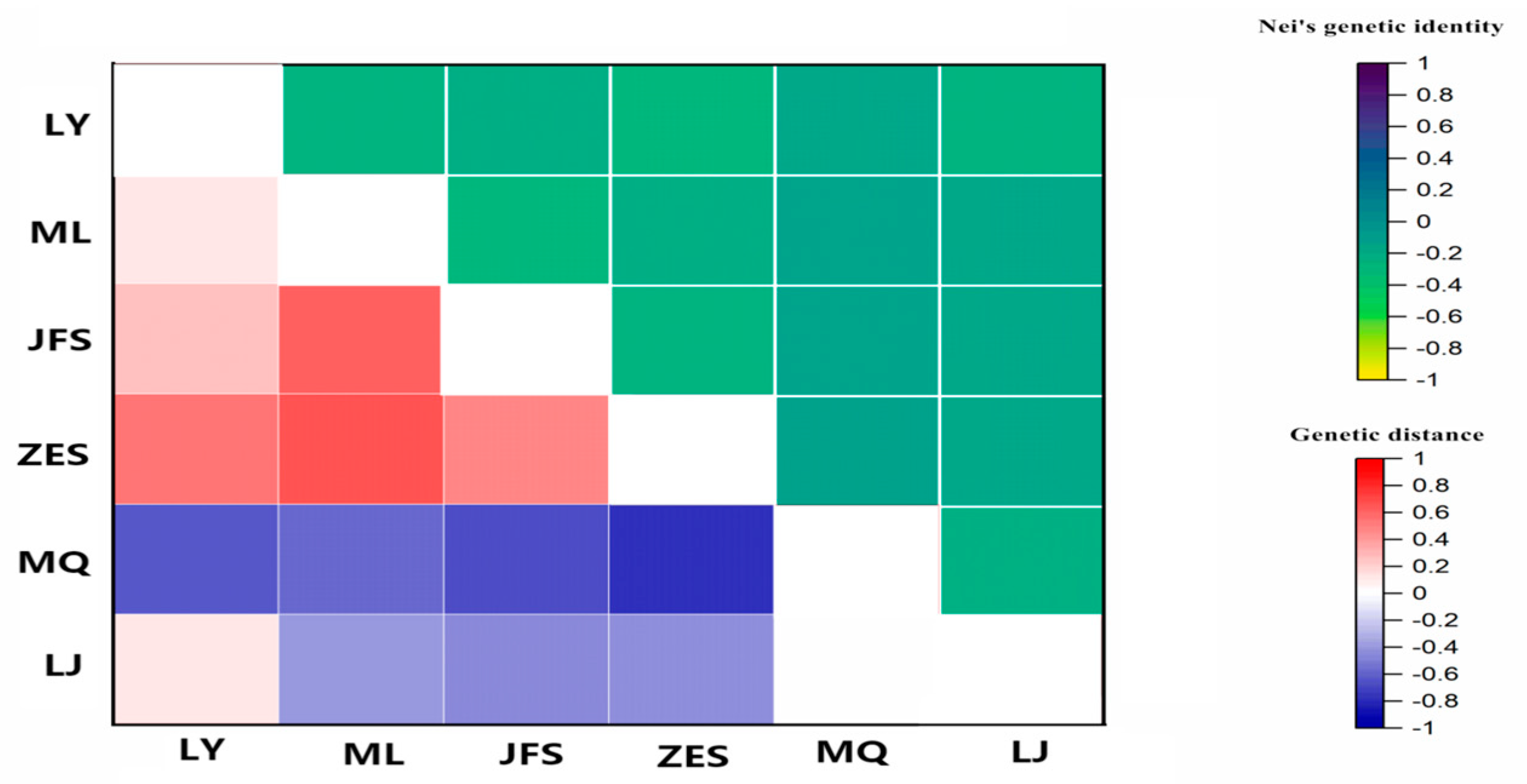
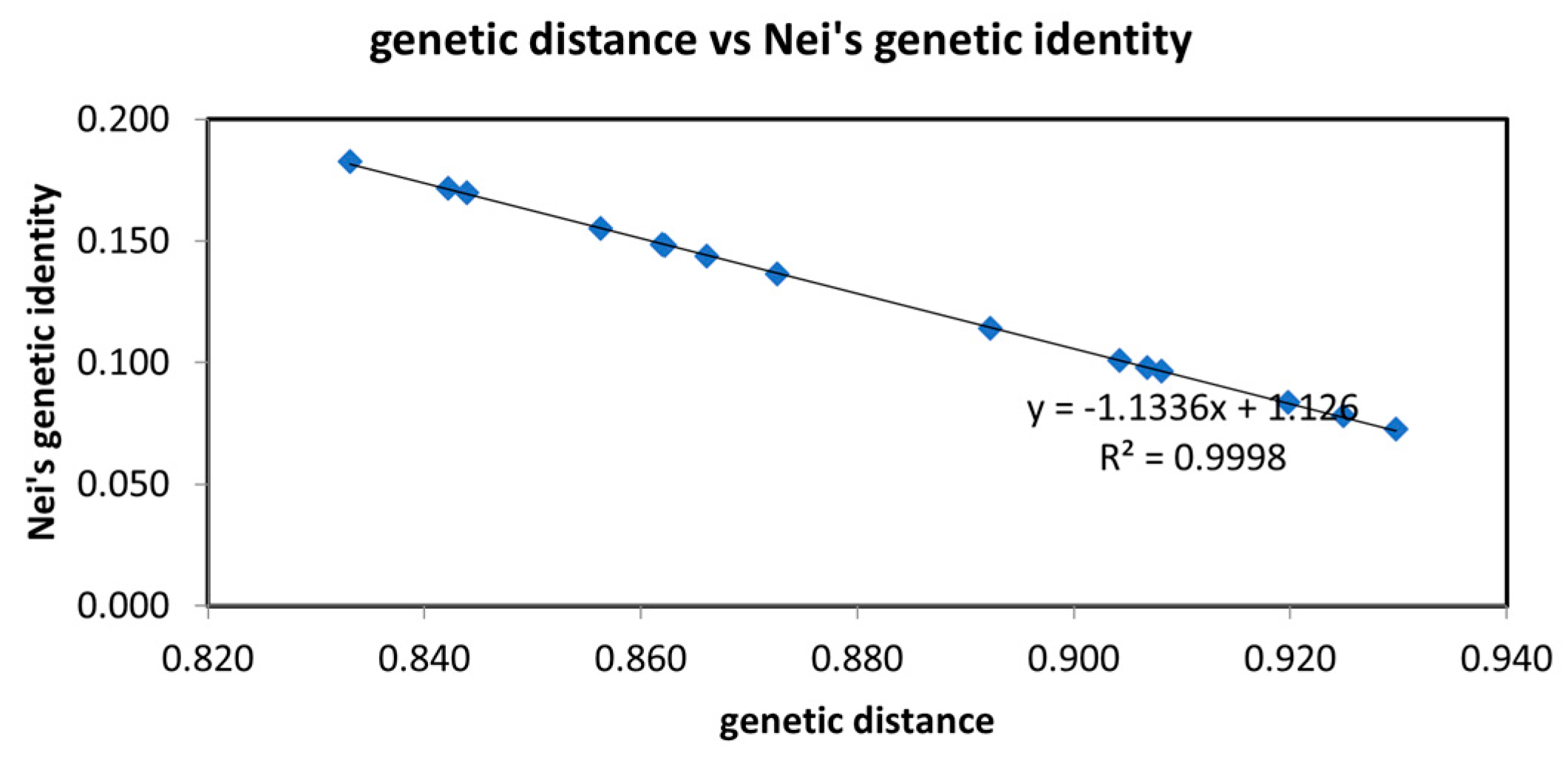
3.3.3. Genetic Distance and Clustering Analysis of H. dentata
3.3.4. Cluster Analysis of the Wild Populations
3.3.5. Principal Component Analysis of the Populations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q.; Wang, H.C.; Cheng, Z.; Wang, M.N.; Li, L.Q.; Long, C.L. Current Status of Wild Orchid Resource in China, Focusing on Their Conservation and Utilization. China Biotechnol. 2022, 11, 59–72. [Google Scholar]
- Zhao, Z.Y.; Liu, N.; Xing, X.K. Research advances in mechanisms of interaction between mycorrhizal fungi and Orchidaceae plants by using omics techniques. Mycosystema 2021, 3, 423–435. [Google Scholar]
- Tang, Y.J.; Guo, S.X.; Chen, J. Advances in the specificity of Orchid-mycorrhizal fungi. J. Cap. Norm. Univ. (Nat. Sci. Ed.) 2021, 42, 63–74. [Google Scholar]
- Zhang, Z.; Yan, Y.; Tian, Y.; Li, S.J.; He, J.S.; Tang, Z.Y. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Su, Z.H.; Li, Y.; Zhang, X.Y.; Chen, M.X.; Chen, L.J.; Li, Y.L. Ornamental Evaluation of Endemic Orchids from Guangdong Province. Chin. J. Trop. Crops 2020, 8, 1560–1565. [Google Scholar]
- Tang, H. DNA Barcoding Identification and Ecological Suitability of Important Medicinal Plants of Orchidaceae. Ph.D. Thesis, Sichuan Agricultural University, Yaan, China, 2016. [Google Scholar]
- Wei, X.; Tang, J.M.; Chai, S.F. Study on the Current Situation and Sustainable Development Strategy of Orchidaceas Resources in Guangxi. J. Guangxi Acad. Sci. 2022, 38, 99–107. [Google Scholar]
- Luo, Y.J.; Tang, J.M.; Jiang, Q.; Liu, Y.; Wei, X.; Huang, B.G. Progress in Conservation Studies of Orchids in Plant in Guangxi Yachang Reserve. J. Guangxi Acad. Sci. 2020, 1, 5–16. [Google Scholar]
- Jiangxi Provincial Health Bureau Revolutionary Committee. Jiangxi Herbal Medicine; Xinhua Bookstore Jiangxi Province: Nanchang, China, 1970; pp. 420–421. [Google Scholar]
- Kunming Municipal Health Bureau. Commonly Used Folk Herbal Medicine in Kunming; Kunming Municipal Health Bureau: Kunming, China, 1970; p. 392. [Google Scholar]
- Liu, Y.T.; Long, C.L.; Dao, Z.L. Ethnobotanical survey on medicinal roots eaten by the local people in Simao, Yunnan Province, during the Dragon-boat Festival. J. Plant Resour. Environ. 2003, 2, 33–38. [Google Scholar]
- Wu, F.Q.; Ma, L.C.; Zheng, J.H.; Xiao, X.Y.; Zhang, X.J. Diversity of Medicinal Vascular Plant Resources from Wetland in Yunnan Province. Chin. Wild Plant Resour. 2021, 9, 75–80. [Google Scholar]
- Chen, Y.Y.; Mao, T.F.; Li, Q.K.; Liu, Z.Y. Tissue Culture and Plantlet Regeneration of Habenaria dentata. Plant Physiol. Lett. 2007, 6, 1136. [Google Scholar]
- Shao, L. Effects of different cytokinins on one-step seedling formation of aseptic seedlings of Habenaria dentata. Contemp. Hortic. 2022, 13, 15–17. [Google Scholar]
- Chen, Y.Y.; Zhu, G.S.; Mao, T.F.; Liu, Z.Y. A Preliminary Study on Habenaria dentate Endophytic Fungi. Guizhou Agric. Sci. 2008, 3, 12–13. [Google Scholar]
- Chen, Y.Y.; Yang, L.; Zhu, G.S.; Liu, Z.Y. Isolation and Identification of Mycorhiza Fungi on Habenaria dentate. Guizhou Agric. Sci. 2010, 8, 84–86. [Google Scholar]
- Chen, Y.Y.; Zhu, G.S.; Mao, T.F.; Liu, Z.Y. The Research on Habenaria dentate Mycorrhizal Fungi Acclerating the Growth of Habenaria dentate in vitro Seedling. Guizhou Agric. Sci. 2009, 12, 33–34. [Google Scholar]
- Wang, C.Y.; Hou, J.; Zhou, M.C.; Li, H.Q.; Cheng, Z.J.; Wang, Y.; Zhang, X.Y. Study in genetic diversity of Gastrodia elata. Hubei Agric. Sci. 2022, 18, 131–138. [Google Scholar]
- Ge, S.; Hong, D.Y. Genetic diversity and its detection methods. In Principles and Methods of Biological Genetic Diversity Research; Qian, Y.Q., Ma, K.P., Eds.; Beijing Science Press: Beijing, China, 1994; pp. 123–140. [Google Scholar]
- Wang, M.Y.; Wang, X.; Zhang, D.; Wang, Y. ISSR Molecular Identification of Amomum villosum and Its Adulterant Al-pinia oxyphylllae Fructus. Mol. Plant Breed. 2021, 2, 562–567. [Google Scholar]
- Cui, X.Q.; Tang, X.; Huang, C.Y.; Deng, J.L.; Li, X.L.; Zhang, Z.B.; Luo, J.S. Analysis of Genetic Diversity of Cattleya Germplasms by Using ISSR Markers. Southwest China J. Agric. Sci. 2020, 7, 1383–1388. [Google Scholar]
- Wang, H.L.; Bu, Z.Y.; Zeng, Y.H.; Long, Q.Y. Study on Genetic Diversity of Cymbidium ensifolium Germplasm Based on ISSR Marker. Chin. J. Trop. Crops 2021, 9, 2526–2534. [Google Scholar]
- Wang, J.; Peng, X.; Sun, X. Primary Exploration of Reform in Education About Training of Communal Nutritionist Courses. J. Ludong Univ. (Nat. Sci. Ed.) 2015, 31, 32–38. [Google Scholar]
- Huang, J.C.; Wan, S.Q.; Chen, Y.; Li, L.H.; Zhang, Z.L.; Zhu, J.J.; Wu, M.; Xing, B.C.; Shao, Q.S.; Lu, C.F. Genet-ic diversity of Anoectochilus roxburghii based on ISSR and SRAP molecular markers. J. Zhejiang A F Univ. 2023, 1, 22–29. [Google Scholar]
- Sun, F.F. Research on Genetic Diversity and Development and Utilization of Germplasm Resource of Medicinal Plant Tetrapanax Papyrifer. Ph.D. Thesis, Guilin Medical University, Guilin, China, 2022. [Google Scholar]
- Lu, J.S.; Bu, C.Y.; Lv, W.L.; He, J.Z.; Su, J.M.; He, C.Y. Analysis on Genetic Diversity of 20 Plants of Orchidaceae by ISSR Molecular Marker. Southwest China J. Agric. Sci. 2012, 6, 2252–2257. [Google Scholar]
- El-Haggar, M.A.; Mahgoub, Y.A.; Aly, H.M.; Ghazy, N.M.; El-Fiky, F.K.; El-Hawiet, A.M. DNA barcodes, ISSR, RAPD and SCAR markers as potential quality control tools for molecular authentication of black and white mulberry. Food Control 2023, 152, 109821. [Google Scholar] [CrossRef]
- Li, Z.H.; He, L.F.; Wang, X.J.; Guo, L.F.; Luo, C.Y.; Li, Q.W. Genetic diversity analysis and fingerprints of Chrysanthemum×morifolium based on SSR molecular markers. Chin. J. Biotechnol. 2023, 7, 2839–2860. [Google Scholar]
- Zafar, P.M.; Shahbazi, E.; Golkar, P.; Shiran, B. Genetic variation of Eruca sativa L. genotypes revealed by agro-morphological traits and ISSR molecular markers. Ind. Crops Prod. 2020, 145, 111992. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Xie, X.Y. ISSR Analysis of Genetic Diversity of Wild Zizania Latifolia Germplasm Resources in China. Master’s Thesis, Jiangxi University of Finance and Economics, Nanchang, China, 2019. [Google Scholar]
- Wang, T.J.; Li, W.Q.; Zhang, S.Y.; Meng, X.F. Genetic Diversity and Differentiation of Five Natural Populations of Ar-temisia halodendron. Sci. Silvae Sin. 2010, 12, 171–175. [Google Scholar]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef]
- Li, Z.Y.; Guan, M.Y.; Li, J.; Li, M.Y. Genetic Diversity of Paphiopedilum micranthum Detected by ISSR Data. Acta Bot. Boreali-Occident. Sin. 2016, 7, 1351–1356. [Google Scholar]
- Jiang, Y.W.; Sun, X.Q.; Luo, H.L.; Yang, B.Y.; Xiong, D.J. Studies on Genetic Diversity of Cymbidium kanran Populations from the Main Mountains in Jiangxi Province Based on ISSR Marker. Acta Hortic. Sin. 2017, 10, 1993–2000. [Google Scholar]
- Ren, X.F.; Deng, Y.B.; Zang, G.C.; Zheng, T.Q. A SSR marker analysis of genetic diversity and population genetic structure of bermudagrass in Henan Province. Acta Prataculturae Sin. 2022, 3, 60–70. [Google Scholar]
- Huang, M.; Huang, X.P.; Chen, X.C.; Wu, M.J.; Wen, J.Y.; Chen, F.X. Genetic diversity analysis of Liquidambar formosana based on ISSR molecular marker. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2022, 4, 524–532. [Google Scholar]
- Nybom, H.; Bartish, I.V. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Perspect. Plant Ecol. Evol. Syst. 2000, 2, 93–114. [Google Scholar] [CrossRef]
- Trapnell, D.W.; Hamrick, J.L.; Ishibashi, C.D.; Kartzinel, T.R. Genetic inference of epiphytic orchid colonization; it may only take one. Mol. Ecol. 2013, 22, 3680–3692. [Google Scholar] [CrossRef]
- Arditti, J.; Ghani, A.K.A. Numerical and physical proper-ties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef] [PubMed]
- Brzosko, E.; Ostrowiecka, B.; Kotowicz, J.; Bolesta, M.; Gromotowicz, A.; Gromotowicz, M.; Orzechowska, A.; Orzołek, J.; Wojdalska, M. Seed dispersal in six species of terrestrial orchids in Biebrza National Park (NE Poland). Acta Soc. Bot. Pol. 2017, 86, 3. [Google Scholar] [CrossRef]
- Lee, S.R.; Choi, T.Y.; Jung, S.Y. Genetic Diversity on a Rare Terrestrial Orchid, Habenaria linearifolia in South Korea: Im-plications for Conservation Offered by Genome-Wide Single Nucleotide Polymorphisms. Front. Plant Sci. 2022, 13, 772621. [Google Scholar] [CrossRef] [PubMed]
- Maki, M.; Matsumua, S.; Yamashiro, T. Allozyme diversity and history of distribution expansion in the maritime peren-nial plant Hedyotis strigulosa (Rubiaceae), distributed over the wide latitudes in the Japanese Archipelago. Biol. J. Linn. Soc. 2008, 93, 679–688. [Google Scholar] [CrossRef]
- Chung, M.Y.; Moon, M.O.; López-Pujol, J.; Maki, M.; Yamashiro, T.; Yukawa, T.; Sugiura, N.; Lee, Y.I.; Chung, M.G. Was Jeju Island a glacial refugium for East Asian warm-temperate plants? Insights from the homosporous fern Selliguea hastata (Polypodiaceae). Am. J. Bot. 2013, 100, 2240–2249. [Google Scholar] [CrossRef]
- Chung, M.Y.; Le HT, Q.; Son, S.; Tian, H.Z.; Chung, M.G. Genetic diversity of the extremely rare Habenaria dentata and the rare Habenaria linearifolia (Orchidaceae) in South Korea: Implications for population history and conservation. Plant Ecol. Evol. 2018, 151, 48–60. [Google Scholar] [CrossRef]
- Guo, J.H.; Liu, S.J.; Wu, Y.H.; Guo, Y.K.; Wang, Z.; Wang, W.L. Genetic Structure of Toxicodendron vernicifluum in Southern Shanxi Province Based on SSR Marker. Mol. Plant Breed. 2019, 9, 2950–2955. [Google Scholar]
- Qin, H.Z.; Pan, B.; Zhao, J.; Zou, R.; Wei, X.; Tang, F.L. Genetic Diversity Analysis by ISSR of Paphiopedilum emersonill, a plant Species with Extremely Small Population. Guangxi Sci. 2022, 6, 1134–1140. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | North Latitude | East Longitude | Number of Samples | Biotope |
|---|---|---|---|---|
| Minqiang Village, Shanglong Township, Longzhou County, Chongzuo City (MQ) | 22°25′23″ | 106°54′43″ | 22 | Hilltop grass |
| Jianfeng Mountain, Xinzhuangxiong Village, Jiangzhou District, Chongzuo City (JFS) | 22°29′30″ | 106°55′2″ | 23 | Mountain slopes |
| Jingxi City, Lu Dong Township (ZES) | 23°8′00″ | 106°19′45″ | 20 | Roadside slopes |
| Longji Village, Luodong Township, Jingxi City (LJ) | 23°8′22 | 106°21′26″ | 20 | Roadside slopes |
| Huaping Village, Huaping Township, Leye County, Baise City (LY) | 24°50′55″ | 106°21′50″ | 24 | Roadside grasses |
| Huanjiang Maonan Autonomous County, Hechi City, Chuanshan Town, Xizaitun Mu Lun Nature Reserve (ML) | 24°55′33″ | 106°33′42″ | 24 | Grass by the river |
| Primer | Primer Sequences (5′-3′) | Annealing Temperature (°C) | Number of Total Amplified Bands | Number of Polymorphic Bands | Polymorphic Percentage (%) |
|---|---|---|---|---|---|
| U808 | AGAGAGAGAGAGAGAGGC | 48.7 | 8 | 8 | 100% |
| U823 | TCTCTCTCTCTCTCTCTCC | 48.0 | 5 | 4 | 80.0% |
| U844 | CTCTCTCTCTCTCTCTCTRC | 48.6 | 6 | 6 | 100% |
| U855 | ACACACACACACACACACACYT | 52.7 | 7 | 5 | 71.4% |
| U876 | GATAGATAGACAGACA | 38.4 | 6 | 4 | 66.7% |
| U878 | GGATGGATGGATGGAT | 47.0 | 7 | 6 | 85.7% |
| U880 | GGAGAGGAGAGGAGA | 47.9 | 6 | 5 | 83.3% |
| U885 | BHBGAGAGAGAGAGAGAGA | 48.0 | 5 | 4 | 80.0% |
| Population | Number Of Polymorphic Loci | Number of Alleles (Na) | Effective Number of Alleles (Ne) | Nei’s Genetic Diversity (He) | Shannon’s Information Index (I) | Percentage of Polymorphic Loci (PPL) |
|---|---|---|---|---|---|---|
| ML | 33 | 1.846 ± 0.366 | 1.506 ± 0.362 | 0.294 ± 0.184 | 0.436 ± 0.256 | 84.621 |
| LY | 31 | 1.795 ± 0.409 | 1.463 ± 0.350 | 0.275 ± 0.1807 | 0.414 ± 0.251 | 79.492 |
| JFS | 27 | 1.692 ± 0.468 | 1.353 ± 0.361 | 0.211 ± 0.195 | 0.321 ± 0.275 | 69.232 |
| ZES | 30 | 1.769 ± 0.427 | 1.387 ± 0.356 | 0.235 ± 0.1806 | 0.361 ± 0.252 | 76.922 |
| LJ | 31 | 1.795 ± 0.409 | 1.430 ± 0.353 | 0.257 ± 0.1828 | 0.393 ± 0.250 | 79.493 |
| MQ | 27 | 1.692 ± 0.468 | 1.406 ± 0.365 | 0.240 ± 0.1949 | 0.360 ± 0.277 | 69.234 |
| Mean | 29.8 | 1.765 | 1.424 | 0.252 | 0.381 | 76.499 |
| p value | 0.168 | 0.101 | 0.087 | 0.089 | ||
| Species level | 2.000 | 1.539 | 0.324 | 0.493 | 100 |
| Total Genetic Diversity (Ht) | Population Genetic Diversity (Hs) | Gene Differentiation Factor (Gst) | Gene Flow (Nm) | |
|---|---|---|---|---|
| Mean | 0.326 | 0.252 | 0.227 | 1.706 |
| Standard deviation | 0.019 | 0.010 |
| Source of Variation | df | Sum of Squares | Variance Components | Variation Percentage | p Value |
|---|---|---|---|---|---|
| Among populations | 5 | 250.856 | 2.014 | 27% | <0.01 |
| Within populations | 127 | 708.618 | 5.580 | 73% | <0.01 |
| Total | 132 | 959.474 | 7.594 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Tang, J.; Zou, R.; Luo, Y.; Deng, Z.; Li, D.; Chai, S.; Wei, X. The Genetic Diversity and Genetic Structure of the Germplasm Resources of the Medicinal Orchid Plant Habenaria dentata. Genes 2023, 14, 1749. https://doi.org/10.3390/genes14091749
Yang Y, Tang J, Zou R, Luo Y, Deng Z, Li D, Chai S, Wei X. The Genetic Diversity and Genetic Structure of the Germplasm Resources of the Medicinal Orchid Plant Habenaria dentata. Genes. 2023; 14(9):1749. https://doi.org/10.3390/genes14091749
Chicago/Turabian StyleYang, Yishan, Jianmin Tang, Rong Zou, Yajin Luo, Zhenhai Deng, Dongxin Li, Shengfeng Chai, and Xiao Wei. 2023. "The Genetic Diversity and Genetic Structure of the Germplasm Resources of the Medicinal Orchid Plant Habenaria dentata" Genes 14, no. 9: 1749. https://doi.org/10.3390/genes14091749