

Toxic Algae in Inland Waters of the Conterminous United States—A Review and Synthesis
by
 , , ,
, , ,
Reynaldo Patiño
1,*,
Victoria G. Christensen
2,
Jennifer L. Graham
3,
Jane S. Rogosch
1 and
Barry H. Rosen
4 1
U.S. Geological Survey, Texas Cooperative Fish and Wildlife Research Unit, Texas Tech University, 1312 Boston Ave., Lubbock, TX 79409, USA
2
U.S. Geological Survey, 2280 Woodale Drive, Mounds View, MN 55112, USA
3
U.S. Geological Survey, 425 Jordan Road, Troy, NY 12180, USA
4
Department of Ecology and Environmental Studies, Florida Gulf Coast University, 10501 FGCU Blvd. S., Fort Meyers, FL 33965, USA
*
Author to whom correspondence should be addressed.
Water 2023, 15(15), 2808; https://doi.org/10.3390/w15152808
Submission received: 1 June 2023
/
Revised: 24 July 2023
/
Accepted: 29 July 2023
/
Published: 3 August 2023
Abstract
:Cyanobacteria are the most common toxigenic algae in inland waters. Their toxins can affect the health of aquatic and terrestrial organisms, including humans. Other algal groups, such as haptophytes (e.g., Prymnesium parvum) and euglenoids (e.g., Euglena sanguinea), can also form harmful algal blooms (HABs) whose toxins cause injury to aquatic biota but currently have no known effects on human health. Prymnesium parvum, however, is responsible for some of the worst HAB-related ecological disasters recorded in inland waters. Here, we provide an overview of the primary toxigenic algae found in U.S. inland waters: cyanobacteria (planktonic forms), P. parvum, and E. sanguinea with the objective of describing their similarities and differences in the areas of HAB ecology, algal toxins, and the potential for future range expansion of HABs. A detailed account of bloom habitats and their known associations with land cover and use is provided from the perspective of water quality. This review revealed that salinity may have an influence on inland cyanobacterial blooms and cyanotoxins that had not been fully recognized previously.
1. Introduction
Toxic algae occur in inland waters worldwide, with the first reports in the scientific literature dating back to the 17th century [1]. Cyanobacteria are the most commonly occurring algae with known toxin-producing strains in inland waters and the only group known to produce toxins that affect the health of aquatic and terrestrial organisms, including humans [2]. The term cyanobacterial harmful algal bloom (cyanoHAB) is deeply engrained in the literature and is used in this review to describe harmful blooms associated with cyanobacteria. However, because cyanobacteria are technically not algae but prokaryotic organisms, the term harmful cyanobacterial bloom is also sometimes used. Eukaryotic taxa, such as haptophytes and euglenoids can produce toxins during bloom events that have caused fish kills in the United States (USA), but their toxins have no known effects on human health ([3]; Section 3). In addition to their impacts on environmental and human health, HABs can also have considerable economic consequences [4,5]. This review focuses on planktonic HABs whose harmful actions are mediated by toxin production.
Cyanobacteria comprise a diverse group of organisms present in oceanic, coastal, and inland waters. They are abundant in today’s environment as well as in the fossil record. There are over 2700 species of cyanobacteria [6], of which about 100 have known toxin-producing strains [7]. Toxic cyanobacteria were first reported in the USA in 1882 after livestock deaths in Minnesota were associated with a bloom [8]. While not common, livestock and wildlife mortalities after cyanobacterial bloom exposure were reported occasionally, particularly in the mid-continent region, throughout the late 19th century and much of the 20th century [8,9]. High-profile pet mortalities in the late 20th century and early 21st century led to an increase in synoptic surveys and state monitoring programs [9], and cyanobacteria-related incidents have now been reported in most U.S. states [10,11]. Human illnesses associated with toxic cyanobacteria have occurred after exposure through drinking water and recreational activities. Worldwide, the first report of human illness associated with toxic cyanobacteria was from West Virginia, USA, in 1930; widespread gastroenteritis occurred during a bloom in communities using the Ohio River as a drinking-water supply [12]. Cyanobacteria-related human illnesses have now been reported from at least 18 states in the USA [13,14,15,16,17]. Human deaths associated with toxic cyanobacteria are rare but have been documented [18,19]. The U.S. Centers for Disease Control has a voluntary One Health Harmful Algal Bloom System [20] in which states can report animal and human illnesses associated with cyanobacterial and other algal blooms. However, reporting is not consistent, and creating a national summary relating bloom events to illnesses remains a challenge.
Four planktonic cyanobacterial genera commonly associated with toxic incidents in the USA and worldwide are: Aphanizomenon, Dolichospermum (formerly Anabaena), Microcystis, and Planktothrix (formerly Oscillatoria) [8,21,22,23]. The taxonomy of cyanobacteria is under revision, and the former Anabaena and Oscillatoria genera were split into planktonic (Dolichospermum and Planktothrix) and benthic (Anabaena and Oscillatoria) growth forms [24,25,26]. These changes in nomenclature were not consistently adopted by algal taxonomists or in the literature until recently. Herein, we reference each genus individually to the extent possible; when it was not possible to distinguish between genera, both the former and new naming conventions are referenced. A recent survey of U.S. states identified Microcystis, Dolichospermum/Anabaena, and Aphanizomenon as the genera of most concern in recreational waters; Planktothrix and Oscillatoria were also of concern but were not indicated as commonly as other taxa [27]. In a national survey of 1161 lakes conducted in the 2007 National Lake Assessment, NLA [28], Microcystis and Dolichospermum/Anabaena were present in more than 55% of lakes, Planktothrix/Oscillatoria were present in about 38%, and Aphanizomenon in about 28% [29]. The 2017 NLA [30] separated planktonic from benthic forms and showed that Microcystis was present in 25% of the surveyed lakes with Dolichospermum present in 29%, Planktothrix in 8%, and Aphanizomenon in 24%.
In contrast to cyanobacteria, the haptophyte lineage is predominantly marine with relatively few representatives in inland waters [31,32]. About eleven species have been recognized in U.S. inland waters [32,33], of which at least two can form harmful blooms. Prymnesium breviturrita (formerly Chrysochromulina breviturrita [32]) is a nuisance alga found in soft-water lakes in the northeastern region of the country [34]. Although P. breviturrita blooms can generate foul odors, there is no concrete evidence that this species can produce toxins [32,35]. The only toxigenic haptophyte documented to date in U.S. inland waters is Prymnesium parvum. It has been reported in 25 U.S. states (including Hawaii), and toxic blooms have occurred in 21 states (Section 2.1.2).
Blooms of P. parvum can be singularly catastrophic. In Texas (USA), over two million fish were estimated to have died in a single lake during a P. parvum bloom in 2001 [36]. More recently (2022), a major P. parvum bloom event in the Oder River (Poland and Germany), resulted in “one of the largest ecological disasters in recent European river history” [37]. Prymnesium parvum was first described in U.S. inland waters in the mid-1980s [38] following massive fish kills in large segments of the Pecos River (New Mexico and Texas, USA). Rhodes and Hubbs [39] hypothesized that toxic P. parvum blooms may have occurred in the Pecos River as early as the 1960s—a hypothesis based on anecdotal reports of a fish kill in conjunction with some typical signs of P. parvum blooms, such as golden-colored and foamy water conditions. Rhodes and Hubbs [39] additionally proposed that P. parvum is a native inhabitant of the Pecos River and that the appearance of blooms in the 1980s was the result of anthropogenic modifications to the riverine environment. However, a subsequent study on genetic relationships by Lutz-Carrillo et al. [40] found that U.S. strains of P. parvum are closely related to European strains and concluded that the presence of P. parvum in the USA is the result of relatively recent introductions.
Euglenoids are a diverse group of organisms that include photosynthetic and non-photosynthetic species, and most photosynthetic species reside in inland waters [31,41]. Photosynthetic genera, including Euglena, were first described in the early 1800s [41]. Although toxigenicity in the laboratory has been demonstrated for eight or nine photosynthetic euglenoids [42,43], an association with toxic blooms in the field has been confirmed for only one, Euglena sanguinea [42,44]. Euglena sanguinea is a globally distributed species [45] that was first described in U.S. inland waters in the early 1900s [46,47,48,49]. However, the first incidence of a toxic bloom associated with E. sanguinea was not documented until 2002 in a fish culture pond in North Carolina, USA [42]. This event was soon followed by multiple reports of toxic blooms of E. sanguinea in aquaculture facilities throughout the South Central and South Eastern USA [44]. Zimba et al. [43] subsequently observed blooms composed of mixed assemblages of photosynthetic euglenoids—including E. sanguinea and another potentially toxic species, Lepocinclis acus—in a wider variety of habitats that included ponds, lakes, rivers, and estuaries in a total of 17 U.S. states. Euglenoid blooms accompanied by ichthyotoxicity have not been reported outside the USA.
Recent reviews on toxic algae have mostly focused on cyanobacteria, with topics including specific genera (e.g., Aphanizomenon [50]; Dolichospermum [51]; Microcystis [52]), benthic cyanobacteria [53], and cyanobacteria in oligotrophic lakes [54]. The last comprehensive review of toxic cyanobacteria in the USA [9] was published over 15 years ago before cyanobacteria and cyanotoxins were broadly included in state and national monitoring programs. The haptophyte P. parvum and several euglenoid species are currently the only known eukaryotic taxa in the USA to produce toxins and have not been as widely studied or reviewed as cyanobacteria. Here, we provide an overview of these three toxigenic taxa with an emphasis on the most commonly occurring planktonic forms. Detailed descriptions of the toxigenic members of the cyanobacteria, haptophyte, and euglenoid algal groups could help facilitate comparative assessments of their HAB ecology (with emphasis on physicochemical factors; Section 2), algal toxins (Section 3), and the potential for future range expansion of HABs (Section 4). For all taxa, we present a comprehensive account of bloom habitats and their known associations with land cover and use from the perspective of water quality, including novel analyses of the potential influence of salinity on inland cyanobacteria.
2. Ecology
2.1. Biology and Biogeography
2.1.1. Cyanobacteria
Although cyanobacteria are prokaryotic organisms (i.e., no nucleus or membrane-bound organelles), most species contain chlorophyll-a and are capable of fixing carbon and producing oxygen by CO2-dependent photosynthesis [55]. When dense cyanobacterial blooms occur, they can change the color of the water, and buoyant species may also form floating scums or mats (Figure 1). There is no quantitative index that is generally applicable to a definition of what constitutes a cyanobacterial bloom, but coupled with visual changes in the appearance of water, chlorophyll-a concentrations ≥20 µg L−1 [56], densities ≥10,000 cells mL−1 [57], or biovolumes ≥0.3 mm3 L−1 [19] have been proposed as base values. Toxigenic and nontoxigenic strains have been described within the same species—for example, in M. aeruginosa [58]—but it is not possible to distinguish these strains in the field or under a microscope.
There is little understanding of what regulates cyanobacterial diversity and biogeography [59]. While some taxa are relatively cosmopolitan in distribution, others are considered temperate, tropical, or endemic [60]. Many cyanobacteria have traits that help them survive even in the most extreme environments. Some can fix dissolved nitrogen (N2; diazotrophic species) through a special cell called a heterocyte (e.g., Anabaena, Aphanizomenon, Dolichospermum); some can regulate buoyancy with the aid of gas vesicles to move to a preferred location in the water column (e.g., Aphanizomenon, Dolichospermum, Microcystis, Planktothrix); and some have cells called akinetes (e.g., Anabaena, Aphanizomenon, Dolichospermum) that help them survive dormant in sediments until conditions are adequate for growth [50,61,62]. Given this wide range of variation, it follows that cyanobacteria have survived since ancient times in environments ranging from clear, low-nutrient temperate lakes to deserts and hot springs [63]. The ecological success of cyanobacteria may be due to the ability of individual species to selectively exploit various environmental conditions [64,65], suggesting niche specificity for individual species [59].
Cyanobacteria reproduce by binary or multiple cell fission. Most species are very small—the largest are barely visible to the naked eye [66]. They are shaped as spheres, rods, and spirals; some are unicellular, but others have secondary morphology that can include the formation of filaments or colonies that can be up to several millimeters in size [66,67]. Cellular shape and dimension can be influenced by the development of heterocytes [68], akinetes [61], or motile filaments [69,70]. These differences can in turn be caused by certain environmental cues including nutrients [69], oxygen, or light availability [71]. Stressors, such as changes in salinity and temperature, could also be the cause of morphological differences among cells [72]. Microcystis spp. are examples of small, spherical cyanobacteria that form colonies, but many other toxigenic taxa in U.S. waters are filamentous (Figure 2).
Buoyancy regulation is especially important for species residing in deep waters in order to minimize sinking and be able to access sunlight in the near-surface layers of a water body. The tendency to settle depends not only on buoyancy regulation but also on shape, size, and density of the cyanobacterial cell. Thus, variations in buoyancy regulation, shape, size, and density “coincide with typical habitats”; cyanobacteria with low intrinsic velocities occur in well-mixed environments, whereas filamentous forms tend to occur in optically deep shallow lakes or in deep lakes with seasonal mixing [64]. Reynolds et al. [64] reported the speed of travel for some common cyanobacteria as Microcystis > Dolichospermum > Planktothrix > Synechococcus. Reynolds et al. [64] also reported that the preferred habitat of Dolichospermum flos-aquae and Aphanizomenon flos-aquae is the epilimna of temperate lakes in summer, and the preferred habitat of M. aeruginosa is systems that stratify on a diel basis. Reynolds et al. [64] stated that it may take several hours to inflate gas vesicles in cyanobacteria, and buoyancy also is influenced by nutrients and CO2. Taxa such as Anabaena and Oscillatoria lack gas vesicles and primarily consist of benthic forms because they cannot regulate buoyancy, in contrast to planktonic taxa. Due to wind, large lakes generally have large fluctuations in mixing depth, with cyanobacteria dispersed throughout this layer, whereas smaller lakes generally have persistent periods of near-surface stability [64], where surface blooms may flourish. This could also be related to wind speed and the orientation of the lake (long and narrow, along the wind direction).
Cyanobacterial blooms have been recorded in surface waters of nearly every U.S. state [10]. Toxigenic cyanobacteria in inland waters are generally described as members of freshwater biotic communities (≤0.5 psu)—the salinity classification of Mitsch and Gosselink [73] is used in this review. However, based on taxonomic and salinity data collected during the 2017 NLA campaign [30], Anabaena, Aphanizomenon, Dolichospermum, Microcystis, and Planktothrix are present in U.S. inland lakes with salinities as high as 4.9, 3.8, 3.8, 7.2, and 3.8 psu, respectively (salinity was estimated using specific conductance [74,75]; see also Section 2.2.1). Species of potentially toxic cyanobacteria that are more typical of marine or brackish environments have been found in hypersaline inland lakes in the USA, such as Salton Sea (Synechococcus and Oscillatoria spp. [76]) and Great Salt Lake (e.g., Nodularia spumigena [77]). A brackish lake in Nevada (USA), Pyramid Lake, has experienced recurring blooms of potentially toxic N. spumigena [78]. Nodularia spumigena, Sphaerospermopsis aphanizomenoides, Anabaenopsis spp., and Microcystis spp. were also found at bloom conditions in a brackish lagoon (4 psu) in California, USA (B. Rosen, personal observations, 2020–2022).
2.1.2. Haptophytes
Haptophytes are typically unicellular, flagellated, and bear a haptonema—a flagellum-like structure that may serve in attachment, feeding, and avoidance responses [31]. Haploid and diploid life stages have been recognized in some haptophytes, suggesting the existence of sexual life stages, but haploid and diploid forms can also reproduce by mitosis [31]. The life cycle of Prymnesium parvum, the most common of the Prymnesium spp., has four distinct stages: two biflagellated haploid types, one biflagellated diploid type, and a non-motile (unflagellated) cyst believed to be a resting stage [79]. Three major clades of P. parvum have been identified worldwide, each producing only one of three known types of prymnesin toxin (Section 3), indicating the possible existence of cryptic species within this taxon [80,81].
Flagellated forms of P. parvum are generally considered to be planktonic, but they can also attach to macroalgae and benthic substrates [82]. There is evidence suggesting that P. parvum (most likely the cyst stage) can survive in dry sediment exposed to high levels of heat and irradiance [3]. Prymnesium parvum contains two golden-brown chloroplasts where photosynthesis occurs (Figure 3) and can also derive nourishment by heterotrophic means via osmotrophy and phagotrophy [3,79]. High-density blooms of P. parvum can be near-monospecific and, under these conditions, the water can take a golden-brown coloration (Figure 3)—thus, the common name for this species in the USA, “golden alga” (singular), should not be confused with “golden algae” (plural), a term commonly used to describe a higher-ranked taxon that includes haptophytes and other algae [83]. A density of ≥10,000 cells mL−1 has been used as a quantitative criterion to define P. parvum bloom conditions [84]. However, the association between cell density and toxicity can vary among water bodies within and across river basins [85]. High- and low-toxicity strains of P. parvum can coexist in time and space [86], further complicating our understanding of the association between cell density and toxicity. Indeed, although they seem to be relatively uncommon, high-density blooms with no overt signs of ichthyotoxicity (e.g., dead or dying fish) have been reported in the USA [85,87,88] and in England [89]. The true frequency of non-toxic blooms is unknown because they can go unnoticed and unreported. In this regard, the NLA campaigns are not a good source of information for P. parvum bloom incidence because the surveys are conducted in the summer when cell densities are typically low (Section 2.4).
Prymnesium parvum is highly euryhaline and found in marine, estuarine, and inland waters worldwide [32,79]. There is some confusion in the literature regarding the distribution of P. parvum and its blooms in the USA. Until recently, its presence had been reported in 22 states of the conterminous USA and in Hawaii [3]. However, in two of these states (Washington [90] and Maine [91]), the records were from open coastal waters, and there is no evidence they were associated with bloom conditions. In Texas, P. parvum was also detected in the tidal reach of some rivers that drain into the Gulf of Mexico [92]. Except for one case (0.14 psu, Waco Lake, Texas; [93]), all inland reports of P. parvum occurrence were associated with brackish waters. The 2017 NLA [30] recorded the presence of P. parvum in additional freshwater bodies in three states: one in Washington (0.03 psu, Panther Lake) and the other two in states where the species had not been previously documented, including Utah (0.13 psu, Gooseberry Reservoir) and Maryland (0.01 psu, Morsell Creek). The increasing number of detections in freshwater sites suggests that P. parvum is more widely distributed in freshwater than previously thought. At the time of this writing, however, toxic blooms in the USA have occurred only in inland brackish waters (>0.5 psu; Section 2.2.2). Texas is the state that has been most severely impacted by P. parvum blooms followed by Arizona, New Mexico, and Oklahoma [36,87,94,95]. One of the latest states to report toxic blooms is California, where, since about 2014, blooms have occurred with some regularity in small brackish lakes in the southern region [96]. Figure 4 shows an updated historical distribution of P. parvum toxic blooms or presence by state.
2.1.3. Euglenoids
Euglenoids are unicellular, flagellated organisms. They can form cysts—including non-motile reproductive palmella—in response to changes in environmental conditions [97]. Their main mode of reproduction is asexual by cellular division [98]. Euglena spp. have a single emergent flagellum, are fusiform in shape with a round anterior end and a tapered posterior end (Figure 5), and exhibit typical shapeshifting euglenoid movement (metaboly) [31]. Their blooms can form scums on the water surface, and some species, such as E. sanguinea, can turn the water “blood red” (Figure 5)—thus, its species name (sanguis, Latin word for blood)—due to the large accumulation of carotenoid pigments, primarily astaxanthin [31]. Quantitative indices of euglenoid bloom conditions do not seem to be available. Toxigenic and nontoxigenic strains of E. sanguinea have been described [43], and not all of its blooms are toxic (Section 2.2.3). Photosynthetic euglenoids are mostly found in freshwater, shallow, eutrophic, and lentic ecosystems in tropical and temperate climates [31,41]. Most species that have been examined typically reside in slightly acidic to slightly basic waters ([99,100,101,102]; see also Section 2.2.3).
2.2. Toxic Bloom Habitats
2.2.1. Cyanobacteria
Toxic cyanoHABs generally occur in nutrient-rich, alkaline (basic pH) waters at temperatures of ≥15 °C [103,104]. However, blooms have also been documented in oligotrophic systems due to unique physiological adaptations found in some species that favor their growth over most eukaryotic taxa, such as the ability to fix nitrogen, use organic forms of nutrients, store nutrients intracellularly, and others [54]. Toxigenic planktonic cyanobacteria are present in lentic and lotic ecosystems, but they are relatively less abundant in the latter [105,106]. Field studies on cyanoHABs in the USA have not always reported the salinity of their sampling sites, but considering their geographic locations, most were probably conducted in freshwater (≤0.5 psu). Due to the scarcity of published reports on toxic cyanoHABs in inland brackish waters, it has been generally assumed that they preferentially occur in freshwater.
As part of this review, the assumption that most inland blooms occur in freshwater was examined using taxonomic and salinity data collected during the 2017 NLA campaign [30]. The NLA campaigns collect data from a single site in or near the lake center, and the entire set of lakes is sampled in the summer and only once [107]. This sampling design could lead to underestimating the true incidence of bloom conditions; however, it is assumed that these sampling biases generally apply across lakes and, consequently, do not affect analyses of the association between salinity and bloom conditions. Five taxa were evaluated: Anabaena, Aphanizomenon, Dolichospermum, Microcystis, and Planktothrix. Although their incidence was low, Anabaena were reported in the water column, and we included this taxon in the analysis. Records of cyanobacterial cell density representing bloom conditions (≥10,000 cells mL−1; Section 2.1.1) were selected and grouped according to lake salinity into freshwater (≤0.5 psu), oligohaline (>0.5 and ≤5 psu), mesohaline (>5 and ≤18 psu), and polyhaline (>18 and ≤30 psu) [73]. Oligohaline, mesohaline, and polyhaline waters together represent the brackish salinity range [73]. Practical salinity units were estimated using specific conductance values. A small fraction of lakes in the NLA datasets includes data collected during a second visit; these data were not used in the present analysis to avoid biasing our results. Results of these analyses showed that 19 to 48 percent of bloom records for individual taxa were associated with oligohaline lakes (Table 1). These rates of bloom occurrence are all disproportionally higher than the percentage of brackish lakes in the NLA database—11.7 percent of a total of 1090 individual records. Especially noteworthy was Planktothrix, for which one-half of all bloom conditions were observed in brackish waters (Table 1). Also notable was that of the ten highest-density records for each taxon, three (Anabaena, Dolichospermum, Microcystis), four (Aphanizomenon), or eight (Planktothrix) cases were associated with brackish lakes. Using the distribution of freshwater (≤0.5 psu; 88.3 percent) and brackish lakes (>0.5 psu; 11.7 percent) in the NLA database as the expected distribution, differences between observed and expected bloom distributions by salinity were statistically evaluated using binomial tests (GraphPad Prism version 9.5.0, GraphPad Software, San Diego, California USA)—the null hypothesis being that inland cyanobacterial blooms have no preference between freshwater and brackish waters. Relative rates of bloom occurrence in oligohaline waters were significantly higher than expected (p < 0.05) for Anabaena, Aphanizomenon, Microcystis, and Planktothrix but not Dolichospermum (Table 1). It should also be noted that a Microcystis bloom was observed during the second visit to a mesohaline lake (5.9 psu; NLA17-SD-10010) that was not in a bloom condition during the first visit; this is the highest-salinity lake where a Microcystis bloom was recorded during the 2017 NLA campaign. These observations indicate that blooms of potentially toxic cyanobacteria in U.S. inland waters are not limited to freshwaters but are also observed in brackish waters, where blooms of several taxa may preferentially occur. Similar observations were made regarding relative rates of cyanotoxin detections in brackish versus freshwater lakes (Section 3).
Blooms dominated by inland taxa such as Microcystis (including M. aeruginosa) have been observed in the upstream reaches of some U.S. estuaries [108,109,110,111,112,113]. Although the presence of toxigenic cyanobacteria and their toxins in open coastal waters could be at least partly due to seaward transport in river flow, a study on the San Francisco Estuary in California reported the formation of a large bloom dominated by Microcystis spp. during a period of extreme drought and minimal flow when salinity in the affected areas was ~1–1.8 psu [114]. The authors of the study also suggested that Microcystis populations may have become established in the upper reaches of the estuary and that these resident populations may now be serving as endemic sources of propagules [114] (see also Section 4). The salinity of the San Francisco Estuary area where the bloom occurred is well within the salinity range where Microcystis blooms are observed in inland waters (Table 1).
Toxigenic cyanobacteria typical of inland waters have also been reported at high densities in brackish waters outside the USA. For example, potentially toxic diazotrophic taxa (A. flos-aquae, Dolichospermum/Anabaena) have been found at salinities as high as ~6 psu (estimated from conductivity values) in saline inland lakes in Alberta, Canada [115,116]. In coastal systems, blooms of M. aeruginosa have been observed in Lake Shinji (0.5–8 psu), a coastal lagoon in Japan [117]; Kucukcekmece Lagoon (~8.8 psu), a coastal lagoon in Turkey [118]; and in brackish areas (7–27 psu) in the Saint Eloi River Estuary, France [119]. Microcystis spp. were reported in brackish waters (up to ~7.6 psu) in the Mekong Delta, Vietnam [120], and blooms of A. flos-aquae and Dolichospermum/Anabaena have occurred in brackish areas (~3–12 psu) in the Baltic Sea [121,122].
Salinity represents the total sum of dissolved ions in water, and research has shown that the ionic composition and major ion concentrations of surface waters may affect the spatial distribution of inland cyanoHABs, and in some cases, independently of salinity. Cations such as calcium [123,124,125,126,127], magnesium [126,127], and potassium [128] can influence the growth of cyanobacteria in the laboratory. At low concentrations, calcium stimulates unicellular growth of M. aeruginosa with little effect on colony formation, but at high concentrations (especially ≥60 mg L−1), calcium inhibits unicellular growth while stimulating colony formation [124,125]—colony formation is considered advantageous for bloom formation [129]. If vigorous growth and colony formation were both necessary for toxic bloom development, calcium alone would seem insufficient to achieve bloom conditions given its opposing effects on said processes. Magnesium also stimulates unicellular growth and colony formation in M. aeruginosa, and it has been suggested that this ion works interactively with calcium to facilitate bloom formation [126,127]. Because potassium concentrations above 2.5 mM (~98 mg L−1) inhibit M. aeruginosa growth in the laboratory, this ion also has been proposed as a water quality factor affecting bloom distribution [128]. However, based on data collected during the 2017 NLA campaign [30], this potassium concentration cutoff for growth inhibition is high and rarely observed in U.S. inland waters.
Among major anions, there is information suggesting that sulfate may play an important role in shaping the biogeography of cyanoHABs. The amount of phosphorus that is released into the water column from sediment is positively associated with sulfate concentration, and this is one of the reported reasons why nitrogen limitation is more prevalent in saline, sulfate-rich environments (e.g., estuaries) than in freshwaters [130]. Although conditions of nitrogen limitation would theoretically favor growth of diazotrophic cyanobacteria, sulfate also reduces the bioavailability of molybdenum, a cofactor of the enzyme nitrogenase that is necessary for nitrogen fixation [131,132]. Therefore, as its concentration increases, sulfate would be expected to slow down growth of these species by hindering their ability to fix nitrogen [130,132]. Marino et al. [116] examined the association between sulfate, molybdenum, and diazotrophic cyanobacteria in inland saline lakes in Alberta (Canada) and found that the sulfate:molybdenum ratio is in fact inversely associated with the abundance of A. flos-aquae and Dolichospermum/Anabaena spp. A more recent study on different saline lakes in the same general area (Alberta) found that lakes with relatively high cyanobacterial abundance were generally nitrogen-limited (TN:TP < 16) and had relatively low sulfate:molybdenum (the study did not distinguish diazotrophic from non-diazotrophic cyanobacteria) [115]. These observations seem to support the notion that sulfate concentrations typical of inland brackish waters help create ecological conditions (nitrogen deficiency) that favor growth of diazotrophic cyanobacteria while also physiologically limiting their growth (molybdenum deficiency). Another inference from these observations is that growth of diazotrophic cyanobacteria in nitrogen-limited environments may not be possible past a certain high concentration of sulfate. Indeed, an earlier review on this topic noted that diazotrophic cyanobacteria are not found in coastal environments above salinities of 10–12 psu, or about one-third of seawater salinity [130]. The review did not mention the sulfate concentration in these environments, but salinity and sulfate are positively associated in coastal waters. Our analysis of the 2017 NLA datasets showed that the highest salinity at which blooms of diazotrophic taxa (Anabaena, Aphanizomenon, and Dolichospermum) occur is 2–3 psu, which is associated with sulfate concentrations as high as ~1500–2340 mg L−1 (Table 1). These sulfate concentrations are high relative to their corresponding salinity (and well above sulfate concentration in one-third of seawater), and yet diazotrophic taxa seem to be able to grow and develop blooms in these brackish lakes. Data for water-borne molybdenum are not available in the NLA datasets, and associations among sulfate, molybdenum, and cyanobacterial abundance could not be examined.
There is evidence suggesting that sulfate may also influence the distribution of non-diazotrophic cyanobacteria. Although sulfur seems necessary for growth and microcystin production in M. aeruginosa [133], reduced growth of a freshwater strain was observed at sulfate concentrations ranging from 40 to 300 mg L−1 (above unspecified culture medium levels) via mechanisms involving oxidative stress and impaired photosynthesis [134]. Whether growth inhibition by sulfate also occurs in strains isolated from brackish waters is unknown but, based on data from the 2017 NLA, this seems unlikely. The median sulfate concentration at which Microcystis blooms were recorded during the 2017 NLA campaign was low at 18 mg L−1 (in freshwaters), but blooms were also observed at sulfate concentrations of 1525 mg L−1 in brackish waters (Table 1). For Planktothrix, the median and maximum sulfate concentrations at which blooms occurred were 128 and 2888 mg L−1, respectively (Table 1). Like the diazotrophic species, Microcystis and Planktothrix strains residing in brackish lakes seem to grow well under the sulfate-rich conditions in these lakes.
2.2.2. Haptophytes
Toxic blooms of P. parvum in U.S. inland waters largely occur during the cooler months of the year, with peak bloom conditions often coinciding with temperatures in the range of 8 to 15 °C [36,85,87,88,94,135] Although rare, there are a few instances of toxic blooms occurring in the summer. For example, a highly toxic bloom was recorded in Moss Creek Lake, Texas, at a temperature of ~29 °C in the summer [135]. Blooms typically occur in eutrophic-to-hypereutrophic, brackish, and alkaline water bodies [3,87,135,136,137,138]. They have been observed in flowing reaches of large rivers [38,39] and streams [139], suggesting that this species can grow and produce toxins in lotic ecosystems. However, blooms happen more frequently in standing waters (including aquaculture ponds), and for this reason, ecological studies have largely focused on lentic water bodies.
Most ecological studies on P. parvum that included nutrient measurements in their sampling designs reported total or inorganic nutrient fractions but ignored organic fractions. The results of the few studies that provided this information indicate that organic nutrients may play a role in determining not only where P. parvum is found but also when. For example, in the Upper Colorado River (Texas), reservoirs impacted by P. parvum have a higher concentration of total (dissolved + particulate) organic nitrogen than reservoirs without a history of toxic blooms [135]. In the Pecos River, the spatiotemporal presence of P. parvum was positively associated with total organic phosphorus, and the spatiotemporal abundance was positively associated with total organic nitrogen [95]. In an impounded urban stream system in the upper reaches of the Brazos River (Texas), the spatiotemporal presence of P. parvum was positively associated with the total organic nitrogen [88]. These observations are generally consistent with a scenario where elevated concentrations of organic nutrients may favor the growth of P. parvum, a mixotrophic species. A similar scenario was described previously for an organic matter-rich coastal site in Norway with a history of toxic P. parvum blooms [140].
In U.S. inland waters, P. parvum has been recorded at salinities ranging from freshwater levels as low as 0.01 psu (Section 2.1.2) to a high-mesohaline value of ~18–19 psu [95]. Toxic blooms, however, usually occur in oligohaline (>0.5 psu) to mid-mesohaline (~10 psu) waters [3,95]. This salinity range is comparable to the range of 1 to 12 psu reported during toxic P. parvum blooms in inland or coastal environments elsewhere around the world [37,82,89,141,142]. In the USA, toxic blooms of P. parvum have not been reported in inland freshwaters (≤0.5 psu) or in open estuarine/coastal waters.
Like cyanoHABs (Section 2.2.1), the inland distribution of toxic P. parvum blooms may be influenced by the ionic composition of water. The hardness cations calcium and magnesium are typically higher in water bodies that have experienced toxic blooms than in those that have not, and their potential role in modulating the potency of P. parvum toxins (Section 3) was proposed long ago and reviewed by Roelke et al. [3]. In addition, there is evidence suggesting that magnesium supports P. parvum growth under conditions of high salinity stress [143].
Sulfate has also emerged as a potentially important variable associated with the distribution of P. parvum blooms. Reservoirs in the Brazos River and Colorado River (Texas) that have experienced toxic blooms are not only of higher salinity but also have much higher concentrations of sulfate (~164–1307 mg L−1) than reservoirs without a history of blooms [137,138,144]. In the Pecos River, high sulfate concentrations (e.g., Red Bluff Reservoir, 3225 mg L−1) have been recorded throughout its P. parvum-affected reaches [95]. In fact, sulfate concentrations in P. parvum-impacted sites in the Brazos River, Colorado River (Texas), and Pecos River are comparable to or even higher than chloride concentrations [95,137]. In Dunkard Creek (West Virginia and Pennsylvania, USA), a freshwater stream where P. parvum blooms had never been recorded, discharge of treated mine wastewater resulted in large increases in sulfate concentration (from <200 mg L−1 pre-spill to 812 mg L−1 post-spill) and salinity but not in chloride concentration, and this event was followed within a few weeks by the formation of a toxic P. parvum bloom [139]. In an endorheic, man-made lake in southern California where P. parvum blooms have repeatedly occurred since 2014 [96], sulfate concentrations are three times higher than chloride concentrations (T. Buckowski, lake biologist, written communication, 2022). In China, sulfate concentrations in brackish inland lakes where P. parvum is present are similar to chloride concentrations [141]. In the Oder River (Poland and Germany), the site of a recent toxic bloom of P. parvum that covered hundreds of river kilometers, sulfate concentrations of ~500 mg L−1 were recorded in its upstream reaches (Gliwice Canal, Poland) [37]. Rashel and Patiño [143] showed that P. parvum growth is enhanced by sulfate over an environmentally relevant range of concentrations (up to 1000 mg L−1) and under conditions of constant salinity (5 psu), suggesting a causal association between sulfate and P. parvum growth. It appears, therefore, that the relatively high sulfate concentrations typical of P. parvum bloom habitats are ecophysiologically relevant. The specific mechanisms underlying these ion-growth associations are at present unclear.
2.2.3. Euglenoids
Blooms of photosynthetic euglenoids generally occur in eutrophic waters during the warmer months of the year, and many of the reported cases were associated with aquaculture ponds [42,44,45,100,101]. With some exceptions, euglenoid blooms have occurred at slightly basic to slightly acidic pH (Section 2.1.3). Toxic euglenoid blooms have been sufficiently documented only for E. sanguinea and only in the South Central and South Eastern regions of the USA [42,44]. In other countries, blooms of E. sanguinea have caused fish injury in aquaculture ponds but via nontoxic mechanisms such as clogging of the gills or shifts in phytoplankton communities to less nutritious assemblages for cultured herbivorous fishes [100,101].
On a global scale, most of the information available suggests that blooms of E. sanguinea—toxic [42,44] or non-toxic [45,100,101,145]—occur in freshwater habitats (Section 2.1.3). Zimba et al. [43] reported blooms composed of mixed euglenoid assemblages in estuaries but did not provide water quality information for their study sites. Our evaluation of Euglena records collected during the 2017 NLA campaign showed that 19 percent of the detections came from oligohaline lakes, where the highest salinity was 3.8 psu (Table 1). However, because differences between observed and expected distributions in brackish and freshwaters were not statistically significant (Table 1), Euglena spp. seem to have equal preference for freshwater and oligohaline lakes. This finding is consistent with the earlier report that photosynthetic euglenoids typical of inland waters can be found in estuarine waters [43].
2.3. Influence of Land Cover and Use
Planktonic taxa in inland waters typically bloom in eutrophic environments (Section 2.2). A historical association between the cultural eutrophication of aquatic habitats and the increased geographic range and intensity of phytoplankton blooms is well established on a global scale [146,147,148]. Surveys of sedimentary records of cyanobacterial pigments and DNA across north-temperate and subarctic lakes have revealed an increase in cyanobacterial abundance beginning about ~200 years ago, a trend which was further accelerated in the mid-20th century [148,149]. Major sources of anthropogenic nutrients in surface waters include runoff from agricultural lands, cities, and towns and municipal and industrial wastewater effluent discharges. Atmospheric deposition of nitrogen can also be regionally important [150,151].
Land cover studies on the conterminous USA have shown a positive association between agricultural land cover and lake cyanobacterial abundance [152,153] and cyanotoxin concentration [154]. These studies also reported that the cyanobacterial abundance [152,153,154] and cyanotoxin concentration [154] are positively associated with nutrient loading [152,153]. Compared with high-elevation lakes, low-elevation lakes have higher percentages of agricultural and developed lands over their watersheds as well as higher levels of cyanobacteria [155]. Watershed size—presumably due to increased nutrient availability—and lake nutrient concentrations were shown to be positively correlated with cyanobacterial bloom incidence in Oregon and Washington, USA [156]. Although these observations together point to nutrients from agricultural and developed lands as primary drivers of cyanobacterial bloom trends, nutrient loading alone may not be a sufficient explanatory variable in all cases. A time-series global survey of large lakes reported that trends in summer bloom intensity since the 1980s (period of record: 1984 to 2012) have not always tracked changes in fertilizer use over their watersheds or other proposed bloom drivers such as temperature or geomorphological traits of individual lakes [157]. The authors of the study suggested that local ambient conditions—such as lake water quality—may interact with other bloom drivers to influence temporal trend patterns in individual lakes [157]. A recent study of Texas reservoirs also reported the lack of an association between agricultural land coverage and the occurrence of P. parvum blooms [138]. A better understanding of the association between land cover and HAB incidence may be gained by examining land uses other than fertilizer application and land cover types other than agricultural and developed lands.
Beyond fertilizers, the use of chemicals in agriculture has increased greatly over the last several decades [158], and mounting evidence suggests that the presence of these chemicals in surface waters may increase the risk of HAB occurrence. In the simplest scenario, agrochemical-tolerant algal species would have a reproductive advantage over intolerant species and consequently grow to higher densities. Evidence in favor of this scenario may be strongest for herbicides [158,159], and among herbicides, glyphosate-based herbicides have received much attention in part because they are the most widely used herbicides in the USA and globally [160,161]. Glyphosate acid contains 18.3% phosphorus by mass and, under certain circumstances, may also contribute to eutrophication [161]. Thus, the selective growth of glyphosate-tolerant species may be further enhanced with the added nutrient, especially in phosphorus-deficient environments.
Cyanobacteria are generally resistant to a variety of agrochemicals and other pollutants [158,159]. In the specific case of glyphosate, some cyanobacteria (e.g., Anabaena sp., M. aeruginosa, Planktothrix sp.) are not only resistant to the herbicide but their growth is also stimulated at low concentrations, particularly in phosphorus-deficient media [162,163,164,165]. A recent study on M. aeruginosa found that growth and microcystin production are both increased in the presence of glyphosate at a very low concentration of 0.1 µg L−1 [164]. Tolerance to—and growth stimulation with—glyphosate is not unique to cyanobacteria. Glyphosate-induced growth has been documented in P. parvum at a concentration of 10 µg L−1 in phosphorus-sufficient media [166]. In Chlorella vulgaris, an extremophile green alga [167], glyphosate and its metabolite aminomethylphosphonic acid (AMPA) stimulate growth at concentrations of 50 and 100 µg L−1, respectively [168]. Glyphosate has a relatively short half-life, and its concentration in U.S. inland waters generally is ≤1 µg L−1 [169,170,171,172]; however, far higher concentrations have been measured in streams receiving agricultural runoff—as high as 430 and 49 µg L−1 for glyphosate and AMPA, respectively [170]. In addition, glyphosate can be taken into biofilm where its concentration (and that of AMPA) is ~3 orders of magnitude higher than in water [173]. Considering these observations, planktonic and benthic algae in water bodies associated with agricultural lands where glyphosate is used may be experiencing exposure to biologically active concentrations of the herbicide and its metabolites, especially following rainfall events. This situation may contribute to an increased risk of HAB occurrence by providing a growth advantage to glyphosate-resistant species, such as potentially toxic cyanobacteria and P. parvum. Consistent with this scenario, a series of large range expansions of P. parvum blooms that occurred in brackish Texas reservoirs during the first decade of the 21st century generally coincided with a large rise in the use of glyphosate-based herbicides over the state’s agricultural lands [138].
Wetlands store, release, and transform a variety of natural and anthropogenic chemicals and play a key role in the maintenance of aquatic ecosystem integrity [73]. They are known to retain pesticides, including glyphosate [174]. Degradation and losses of wetlands due to anthropogenic activities have occurred globally over many centuries, but the pace of destruction increased greatly in the early 1900s, and more so in inland environments compared to coastal areas [175]. Because of their ability to retain nutrients and other pollutants, treatment (constructed) wetlands have been used or proposed as a means to reduce the incidence of HABs [176,177,178]. Although only a handful of studies have addressed the historical or contemporaneous association between wetland cover and HABs in the conterminous USA, the information available is consistent with the notion that wetland or forest cover influences the risk of HABs. In lakes of Oregon and Washington, for example, evergreen forest cover at the watershed level negatively correlates with cyanoHAB occurrence [156]. In Texas, the percentage of wetland cover at the riparian or whole-watershed levels is much lower in reservoirs impacted by P. parvum than in those without a history of blooms [138]. In New York, water quality attributes such as stream salinity and total phosphorus concentration negatively correlate with riparian forest cover [179]. A national study examining the association between cyanobacteria and land cover at the county level found a negative association between cyanobacterial abundance and the extent of forested area [153].
Additional landcover studies with emphasis on wetlands and riparian areas and on water quality (e.g., salinity, major ions, agrochemicals) may help understand the variability in HAB trends among lakes noted by some investigators, e.g., [157]. Such studies would also help identify watersheds and water bodies that would benefit most from enhanced land management practices and wetland construction or restoration projects.
2.4. Seasonal Bloom Dynamics
Seasonal changes in the composition and abundance of plankton communities are driven by external factors and internal interactions. A review by Sommer et al. [180] concluded that the beginning and end of the phytoplankton growth period are defined primarily by physical factors such as light availability (day length) and that overall phytoplankton abundance is limited by nutrients. Changes in phytoplankton abundance during the growth period can be due to multiple factors including nutrient depletion, protist and metazoan (zooplankton) grazing, and parasitism (fungal and viral infections). The classic annual profile of phytoplankton abundance for a eutrophic temperate water body with limited fish predation and little overwintering of metazoan grazers includes a spring bloom, a clear water phase, and a summer bloom with a relatively high proportion of species inedible for zooplankton [180]. Here, we discuss bloom seasonality in the context of physicochemical variables.
Patterns in the seasonal growth of cyanobacteria generally fit within the model of Sommer et al. [180]. Toxic cyanoHABs mostly occur at temperatures of ≥15 °C (Section 2.2.1), however, and temperature also has been used to explain the seasonal succession of phytoplankton communities that generally ends with cyanobacterial blooms in the summer. Using the 2007 NLA survey data, Paerl and Otten [104] found a negative association between water temperature and abundance of diazotrophic cyanobacteria (e.g., Dolichospermum/Anabaena, Aphanizomenon) and a positive association with the abundance of non-diazotrophic species (e.g., Microcystis, Planktothrix/Oscillatoria). These observations are consistent with the seasonal shift from diazotrophic to non-diazotrophic cyanobacteria reported by several studies [104,181,182]. More broadly, eukaryotic algae often dominate in the winter/spring but subsequently give way to cyanobacterial dominance as the summer approaches or progresses, reflecting, at least in part, differences in thermal optima among various taxa [104,183,184]. This eukaryote-to-prokaryote succession may also be partly due to the relatively large increase in pH (basification) that occurs during intense cyanobacterial blooms in the summer coupled with the high tolerance of cyanobacteria to basic water conditions [185,186,187]. Cyanobacterial blooms in U.S. inland waters can last up to several months in the summer [188]. Likewise, cyanotoxin levels are generally highest in the summer (during the stratified period), but increased levels have been observed in the fall (Figure 6 [189]). Outside the USA, cyanobacterial bloom durations of up to five months starting in the summer (artificial ponds in southern Poland) [190] or eight months starting in the spring (Taihu, China) [191] have been reported. It should be noted, however, that cyanobacterial blooms can also occur during the cooler months of the year (<15 °C) including the winter [10,192]. The relative frequency of cold-water cyanobacterial blooms is at present uncertain but there is evidence suggesting that the strains involved have acquired adaptative mechanisms to survive and grow at the lower temperatures [192].
The timing of P. parvum’s growth in U.S. inland waters is not what would be expected if light (increased daylength and light intensity) were its primary driver. With rare exceptions, toxic blooms of P. parvum typically occur during the cooler months of the year (<15 °C), either in the fall or spring [88] or from the fall to the spring (Figure 7). The growth period of this species thus has been described as beginning in the fall and ending in the spring, with variability in population density and toxicity (including the absence of blooms) along this timeline being a function of other environmental variables [88]. In view of this information, temperature seems to be more important than light as driver of P. parvum’s growth period in U.S. inland waters. (For comparison purposes, although P. parvum blooms elsewhere around the world have occurred at various times of the year including the summer [37,82,89,142], most toxic blooms appear to have been recorded outside of the summer [82,89,142]). Prymnesium parvum can utilize organic forms of nitrogen and phosphorus but still requires light to meet its carbon demands for growth [193]. Light intensities during the bloom season in the USA, however, are sufficient to support growth in the field as well as the laboratory [194]. Blooms can last from a few weeks [88,139,195] to several months [144,196], including an extreme case of a toxic bloom that lasted eight months from initiation in the fall to termination in the spring [144] (Figure 7). Geographic differences in bloom duration and ecological impact have been noted [144,197], with lakes in watersheds receiving lower amounts of precipitation [198] generally faring worse.
There is no established answer to the question of why most toxic P. parvum blooms occur when temperatures are seasonally low, which are suboptimal for their growth [194,199]. However, using a common P. parvum strain found in U.S. inland waters, Baker et al. [194] reported that toxin production (Section 3) increases as temperature decreases at salinities typical of P. parvum bloom habitats. Also, the pH of water can increase to relatively high levels during intense P. parvum blooms in the field (>9 [88]), and, like cyanobacteria, this haptophyte is relatively resistant to high pH—P. parvum growth in batch cultures can continue until pH reaches values ≥10 [200]. In addition, the potency of P. parvum toxin increases with increasing pH [3]. Thus, the ability of P. parvum to produce toxins at cool temperatures coupled with the toxin’s increased potency as pH increases during bloom development may deter grazing, reduce competition, and consequently confer a reproductive advantage to P. parvum over other phytoplankton [196]. Indeed, intense P. parvum blooms in the USA [3] and elsewhere [79] can be near-monospecific. It should be noted that toxic blooms of P. parvum do not occur every year in many, if not most, of the affected water bodies. The reasons for this interannual variability are uncertain, but external factors other than temperature may in some instances include the timing (relative to bloom season) and amount of stormwater runoff [88].
Blooms of photosynthetic euglenoids typically occur during the warmer months of the year (Section 2.2.3), and in the USA, E. sanguinea blooms have been recorded between the mid-spring and early fall [42,44,145]. These observations are consistent with the classical model [180] of the primary role of light in defining the annual growth period of photosynthetic euglenoids. Euglenoid blooms are generally restricted to slightly basic/slightly acidic environments and, unlike most cyanobacteria and P. parvum, with some exemptions, photosynthetic euglenoids do not tolerate high pH (Section 2.1.3 and Section 2.2.3; [201]). Although the growth periods of cyanobacteria and euglenoids are similar and these taxa can coexist in time and space, differences in pH preferences or tolerances may partly explain why euglenoid abundance (E. sanguinea) seems to be negligible during intense cyanobacterial (e.g., M. aeruginosa) blooms (associated with high pH), while cyanobacteria can coexist with euglenoids during euglenoid-dominated (e.g., E. sanguinea) blooms [202]. Information on euglenoid bloom duration in U.S. inland waters is unavailable or difficult to find. In Bangladesh, blooms lasting up to 5 months from initiation to termination have been recorded in aquaculture ponds [203].
3. Toxins
The toxin classes covered herein are those that are known or suspected to cause harm, occur in the USA, and are produced by primarily planktonic cyanobacteria, haptophytes, and euglenoids. In U.S. water bodies used for recreation and drinking water, microcystin was cited as the cyanotoxin of most concern; anatoxins and cylindrospermopsin were also frequently cited as a concern [27]. Of note is the large number of cyanobacterial taxa and strains known to produce toxins compared to haptophyte and euglenoid taxa. Toxins discussed include anatoxin, cylindrospermopsin, euglenophycin, microcystin, prymnesin, saxitoxin, and emerging toxin classes (β-methylamino-L-alanine (BMAA), anabaenopeptins, and cyanopeptolins). Currently (2023), the classification of BMAA as a human neurotoxin is controversial, and further studies would be beneficial to clarify its role in human disease [19,204]. Although there are numerous variants in each toxin class with a range of toxicities, those discussed here fall into general categories: neurotoxins, hepatotoxins, and ichthyotoxins. These toxins generally have different modalities and thus behave differently in the environment and organisms. Neurotoxins that are produced in inland waters include anatoxins, saxitoxins, and BMAA [205] and act upon the nervous system; hepatotoxins include microcystin and cylindrospermopsin and act upon the liver; and prymnesins and euglenophycin are ichthyotoxins, which are toxic to fishes and other aquatic—primarily gilled—organisms.
Because relative rates of cyanobacterial bloom occurrence were generally higher in brackish than in freshwater lakes (Section 2.2.1), an analysis was conducted to determine if this situation also applies to cyanotoxin detections. Microcystins (sum of congeners), cylindrospermopsin, and salinity data from the 2017 NLA [30] were used for this analysis. All cases of toxin concentrations greater than the method detection limit (MDL) or the national recreational water quality criteria (microcystins > 8 µg L−1; cylindrospermopsin > 15 µg L−1) were classified according to lake salinity into freshwater, oligohaline, mesohaline, and polyhaline for a general assessment of salinity-dependent distributions. In addition, differences between observed and expected cyanotoxin distributions in freshwater and brackish sites were evaluated with binomial tests, as described in Section 2.2.1. Microcystins were detected over a wide range of salinities, from freshwater to polyhaline (0.00–21.7 psu), and one-half of the cases above the national criterion were recorded in oligohaline lakes (Figure 8, Table 2). Cylindrospermopsin concentrations in the 2017 NLA database all fell below the national criterion, but this toxin was recorded at salinities ranging from freshwater to mesohaline (0.00 to 7.2 psu; Figure 8, Table 2). Differences in relative rates of toxin detections in freshwater and brackish lakes were highly statistically significant; namely, cyanotoxin detections occurred at relatively higher rates in brackish than freshwater lakes (Table 2).
Across the USA, some toxins (i.e., microcystin, see Figure 9) appear more cosmopolitan than others [29,206]. However, data on many toxins are limited and the variation in data availability may be due to monitoring or study design rather than actual variability in occurrence. Some cyanobacterial species have shown regional or local dominance, and it follows that their toxins may have a regional or local pattern as well. Associations have been documented for microcystin and watershed land use [154] and microcystin and ecoregion [207]. Many cyanobacterial taxa show preferences for certain extreme environments [63]. It follows that if taxa are adapted to certain niches, toxins that are produced by those taxa may have a higher occurrence in those niches.
Bioaccumulation occurs when organism tissue concentrations exceed environmental concentrations [208]. Organisms may be exposed to toxins through skin contact, direct consumption of contaminated water, or secondary consumption through contaminated fish, crops, or other food sources. Algal toxins have varying degrees of bioaccumulation potential.
3.1. Anatoxins
Anatoxins consist of a group of neurotoxins, including anatoxin-a, homoanatoxin-a, and dihydroanatoxin-a. Anatoxin-a(s) is structurally unrelated to anatoxin-a [205,209], and a new name, guanitoxin, has been adopted [210]. Most field studies [211] on this group of toxins focus on the anatoxin-a analog. Organisms that are orally exposed are affected because anatoxin-a mimics acetylcholine, a neurotransmitter, and binds to acetylcholine receptors, effectively overstimulating muscles, including those responsible for respiration, leading to fatigue and possibly death by respiratory failure [212,213,214]. Chronic and sublethal effects of anatoxin-a are understudied.
A recent study indicated that the global distribution of anatoxin-a-producing cyanobacteria is expanding [215], although many instances are due to benthic cyanobacteria, e.g., [216]. Reporting on blooms in Midwestern USA, Graham et al. [206] noted that anatoxin-a occurred more frequently than reported in other studies both in the USA and worldwide. Some multiple-toxin studies reported that seasonal anatoxin-a occurrence patterns were not the same as microcystin patterns [217,218].
Bioaccumulation of anatoxin-a is a possible exposure route for larger organisms. Anatoxin-a was shown to have negative allelopathic effects on macrophytes [219], and the effects of anatoxin-a include altered morphology and inhibited photosynthesis [220]. When compared with microcystin and cylindrospermopsin, anatoxin-a showed greater bioaccumulation capacity in aquatic plants [220]. In one study [221], anatoxin-a was shown to bioaccumulate in fish, with the highest concentrations occurring in the liver. This would indicate that humans, who typically eat fish fillets, may not be as at-risk as wildlife that consume fish whole [211]. Global occurrence, toxicity, and environmental fate of anatoxins are extensively reviewed in Christensen and Khan [211].
3.2. Cylindrospermopsin
Until recently, cylindrospermopsin was not monitored routinely in the USA [222], yet it has become a freshwater toxin of increasing interest due to the severity of its human health impacts and multiple toxicity endpoints—damaging the liver, digestive system, and DNA, as well as being a possible carcinogen [223]. Thus, cylindrospermopsin could also be considered a cytotoxin, affecting cells throughout an organism. Sublethal effects also can include changes in growth, reproduction, and behavior [222]. There are five known analogs: cylindrospermopsin, 7-epi-cylindrospermopsin, 7-deoxy-cylindrospermopsin, 7-deoxy-desulpho-cylindrospermopsin, and 7-deoxy-desulpho-12-acetylclylindrospermopsin [19,224]. Cylindrospermopsin is water soluble, typically extracellular [19], and relatively stable but decomposes rapidly when exposed to sunlight (half-life of 1.5 h; [224,225]).
Some of the challenges for tracking and detecting cylindrospermopsin are that some cyanobacteria that produce it have expanded from tropical and subtropical areas into temperate zones [208] and some (e.g., Raphidiopsis raciborskii; formerly Cylindrospermopsis raciborskii [226]) can bloom in the winter when many monitoring programs are on hold. Data on cylindrospermopsin occurrence due to planktonic cyanobacteria is limited, although there are reports of cylindrospermopsin occurrences attributed to benthic cyanobacteria [227]. Raphidiopsis raciborskii, a planktonic cyanobacteria known to produce cylindrospermopsin, can be found in many different environments including flowing water, and it is able to tolerate a wide range of temperatures, light, and other cyanobacteria species [228], potentially contributing to its expansion into temperate climates. Based on the 2017 NLA data, the distribution of cylindrospermopsin across the USA is concentrated primarily in the Midwestern states including the eastern edge of the Missouri watershed, the Arkansas–White–Red watershed and the Texas–Gulf watershed (Figure 10). Other watersheds with notable numbers of detections include the South Atlantic–Gulf watershed and the Mid-Atlantic watershed (Figure 10). Midwestern Plains states and North Central Texas appear to be affected by the highest concentrations.
Evidence of cylindrospermopsin bioaccumulation, even at very low field concentrations, was reported by Kinnear [208], who reviewed more than ten studies on cylindrospermopsin bioaccumulation. These studies included evidence of bioaccumulation in snails, mussels, crayfish, fish, tadpoles, duckweed, hydrilla, mice, and more. Sundaravadivelu et al. [229] noted that cylindrospermopsin is likely to be taken up by a variety of aquatic organisms. Some evidence indicates that bioaccumulation in fish is species-specific [227], and bioaccumulation also varied between different snails [230]. Cylindrospermopsin has been shown to damage gut linings, which may accelerate absorption and accumulate preferentially into certain tissues, such as the pancreas and liver [208]. However, organisms like snails, which are lower in the trophic chain, can accumulate high concentrations of cylindrospermopsin without negative effects [208]. Global occurrence, detection, toxicity, and degradation of cylindrospermopsin is are extensively reviewed in De La Cruz et al. [223].
3.3. Euglenophycin
Euglenophycin, produced by euglenoids, can cause fish kills, affect the growth of microalgae, and inhibit mammalian cancer tissue growth [43], having demonstrated cytotoxic activity against colon and neuroblastoma cancer cells [231]. Euglenophycin has a structure similar to solenopsin, an alkaloid found in fire ant (Solenopsis invicta) venom [43]. In laboratory experiments, euglenophycin was found to be more stable at room temperature than at 8 °C or −80 °C [232]. Fish exposed to euglenophycin in the laboratory exhibited behavioral changes that included loss of equilibria and disorientation, see [42]. The discovery of euglenophycin was relatively recent, and methods for its detection are emerging [232,233].
Information on the geographic range of euglenophycin is limited, but the toxin has been detected in farmed fish ponds in the South Central and South Eastern USA [44]. The geographic range of euglenophycin is likely smaller than the range of euglenoids (see Section 4.3), as many euglenoids do not produce euglenophycin [43]. No evidence of bioaccumulation of the toxin was found in the literature, possibly due to the relatively recent discovery of euglenophycin.
3.4. Microcystins
Among the cyanobacterial toxins, microcystins are the most widely investigated and best described in the literature [234,235] with at least 246 known variants [7,236]. Microcystins are stable and water-soluble [237] and quickly concentrate in the liver of exposed organisms [236]. Once in the liver, microcystins bind to protein phosphatase, which may lead to cell necrosis, hemorrhage, and death, e.g., [15,238]. Microcystins are stable and can persist for weeks after being released from cyanobacterial cells [239].
In a study on 1156 U.S. water bodies [154], high microcystin concentrations (>1 mg L−1) were limited to three ecoregions where agriculture has a strong influence (Corn Belt and Northern Great Plains, Mostly Glaciated Dairy Region, and South Central Cultivated Great Plains). The NLA 2017 data support this finding, with most microcystin detections in Central USA, within the Mississippi River drainage (Figure 9), and the highest concentrations in North Dakota and South Dakota, with one high detection in Oklahoma. Numerous field studies have reported microcystin concentrations across the USA, e.g., [29], and one study noted differences in the distribution of microcystin within bays of the Great Lakes, with intracellular concentrations lower in the center and higher near the edges. However, extracellular concentrations did not retain this pattern but rather had an east–west pattern with higher concentrations on the western shores [239]. Svirčev et al. [23] reviewed blooms and associated poisonings worldwide, noting that the highest percentage of poisonings occurred in North and Central America.
Microcystin has been shown to bioaccumulate in fishes, although one study indicated detectable concentrations occurred in the liver and gut and not in muscle tissues [240]. Preece et al. [241] not only documented freshwater to marine transfer of microcystins but also showed subsequent bioaccumulation in mussels. A number of studies have shown additional evidence of bioaccumulation of microcystin, including in terrestrial insects [242] and plants [243]. More information on the global occurrence, detection, toxicity, and congeners of microcystin can be found in a variety of reviews [29,213,234,236].
3.5. Prymnesins
Blooms of P. parvum have caused mortality in a wide variety of aquatic organisms including vertebrate (fishes, amphibians) and invertebrate (crustaceans, shellfish) animals and microalgae [3,79]. A large number of substances have been proposed as toxins produced by P. parvum. It is not clear if all of these substances are biologically relevant, but a consensus seems to be emerging that the prymnesins are important. Prymnesins are a group of compounds that can be cytotoxic, neurotoxic, or ichthyotoxic [244]. Neurotoxic effects include rapid changes in swimming behavior and lethargy of fish exposed to prymnesin in laboratory experiments [245]. In the field, lethargic fish can be seen near the surface of the water prior to the onset of mortality (Figure 3). In terms of ichthyotoxic effects, these toxins are believed to permit ion leakage in plasma membranes and are particularly harmful to gill-breathing organisms [244].
Prymnesium parvum seems to be the most widely studied prymnesin-producing species, and 26 toxin-producing strains of P. parvum have been identified [80]. Scientists have characterized 51 prymnesins [80], including prymnesin-1, prymnesin-B1, prymnesin-2 [229]. The toxins have been grouped into 3 types: prymnesin-A, prymnesin-B, and prymnesin-C based on the length of the carbon chain, with both A and B types identified in the USA [80,81,246]). The toxicity of prymnesin-A is higher than that of prymnesin-B [246]. An unusually toxic strain of P. parvum was recently isolated from a small lake in southern California [96], but the toxins produced by this strain have not been characterized.
Salinity, dissolved major ions, pH, and other water quality characteristics have been linked to the activation and potency of prymnesins [244], and the organisms that produce prymnesins are thought to be of marine origin. Despite this observation, prymnesins have been detected in sites from a wide geographic range, primarily in temperate and subtropical zones (see map from [244]). No geographical pattern in prymnesin-producing P. parvum has been identified [80].
Limited research is available on the bioaccumulation of prymnesins. Valenti et al. [247] noted the importance of considering pH when determining the bioavailability of prymnesin to organisms. Sundaravadivelu et al. [229] noted the need for a method to detect prymnesins in matrices that include flesh. While additional information is available on the global distribution of P. parvum, e.g., [244]), less information is available on the distribution of prymnesin, but the toxin likely occurs primarily in the same temperate and subtropical zones.
3.6. Saxitoxins
Saxitoxins are a group of toxins, including saxitoxin and neosaxitoxin, that block sodium channels along nerve cells and suppress signals from nerves to the brain [213,248], impairing muscle stimulation [18] and leading to possible respiratory failure. Much research exists on the effects of saxitoxin in marine waters, and saxitoxins are associated with paralytic shellfish poisoning. However, in freshwater, saxitoxin analogs may be hydrophobic [249,250], which can result in a decrease in binding to sodium channel receptors, lowering the toxicity [251].
In Brazil, higher saxitoxin production was associated with lower water temperatures [252]. A U.S. study [253] also indicated an association between saxitoxin and colder water, although the association was with a benthic species of cyanobacteria. A summary of global saxitoxin occurrence in freshwater [211] indicated detections in lakes, rivers, reservoirs, and cobbled streams, although the authors indicated that many factors may affect occurrence and detection (altitude, latitude, sample depth, seasonality, and trophic state). In an Australian study [254], saxitoxin was detected frequently (24 of 31 samples) in reservoirs and farm ponds. Riverine saxitoxin occurrence seems to be associated with benthic, rather than planktonic species, e.g., [243,245,253,255]). In the United States, some researchers noted that saxitoxins are rare compared to other cyanotoxins [29,206], but relatively high levels of occurrence have been observed in places such as Ohio, e.g., [14].
It is well known that saxitoxins bioaccumulate in marine waters, concentrating in fish and shellfish [256,257]. Fewer freshwater studies are available on saxitoxin, but research indicates that saxitoxin is lipophilic, increasing the concern for bioaccumulation in fat tissue [250,258]. Some research studies [259,260] show bioaccumulation in freshwater fish and shellfish, whereas other studies, e.g., [240], show bioaccumulation of microcystin, but not saxitoxin. It is possible that bioaccumulation is species dependent [211], and the presence of saxitoxin-producing cyanobacteria throughout freshwater environments may indicate that ecological effects occur throughout the food web [258]. The global occurrence, toxicity, and environmental fate of saxitoxin are extensively reviewed in Christensen and Khan [211].
3.7. Lesser Studied Toxins and Toxin Co-Occurrence
Emerging toxins or new variants of existing toxin classes are often detected in U.S. lakes. Toxins such as BMAA, anabaenopeptins, and cyanopeptolins have been an emerging concern in lakes across the USA [110,209,261]. Although BMAA is associated with soil cyanobacteria [243], Cox et al. [262] reported BMAA production by numerous bloom-forming cyanobacteria, including Microcystis, Dolichospermum/Anabaena, and Planktothrix species. However, other researchers have questioned the production of BMAA by cyanobacteria and its correct identification [263,264,265]. Anabaenopeptins can be produced by several cyanobacteria, including Anabaena, Nostoc, Microcystis, Planktothrix, Lyngbya, and Brasilonema [266,267], whereas cyanopeptolins can be produced by Microcystis, Planktothrix, Tychonema bourrellyi, and Nostoc edaphicum [268,269]. These two toxins may occur as frequently as microcystins and at similar concentrations [270].
BMAA was shown to bioaccumulate in mammals and seafood [271]. Limited bioaccumulation data are available for anabaenopeptins, cyanopeptolins, and other cyanopeptides, although cyanopeptides have been detected in fish, frog, snail, and mussel tissue [270]. Sublethal effects of BMAA have been debated, e.g., [272]. Sublethal effects of anabaenopeptolin include inhibition of protease and effects on reproduction [261,270]. Cyanopeptolins have induced DNA damage and produced neurodevelopmental effects in zebrafish [273,274].
Exposure to multiple toxins may be more common than single-toxin exposure [224]. Toxin co-occurrence in U.S. water bodies has been investigated by several researchers, e.g., [206,275]. The literature includes evidence of low co-occurrence from a large nationwide study (0.32 to 5.0% of samples having more than one of four toxins; [29]), with more frequent co-occurrence in some individual lakes (e.g., 84% of samples in two California lakes) [276].
Co-occurrence has been reported for relatively common toxins, such as anatoxin-a, microcystin, and saxitoxins [218,277], as well as lesser studied toxins, such as anabaenopeptins [275,278] and BMAA [214]. Anabaenopeptins and cyanopeptolins, although infrequently analyzed in water-quality studies, can occur at similar concentrations and just as frequently as microcystins [270], and synergistic exposure to multiple toxins may increase exposure risk [209,279].
4. Potential for Future Expansion of Toxic Blooms in the USA
The potential for HAB expansion and intensification in inland waters throughout the USA and globally has been addressed by multiple reviews and analyses. The primary contexts of previous efforts have been climate change and eutrophication trends. For example, globally rising temperatures are projected to enhance growth and extend the growth period of thermo-tolerant species (e.g., harmful cyanobacteria) while reducing growth of thermo-intolerant species [280,281,282]. In nutrient-rich environments, high rates of carbon fixation by rapidly growing blooms could result in carbon deficiency, but the increasing concentration of air CO2 may provide some relief to this limitation due to steeper air-to-water CO2 gradients and fluxes [185,280]—under this scenario, blooms could further grow in intensity in some places. The combined effects of rising seawater levels and changes in freshwater riverine flows could alter the salinity of coastal ecosystems and the composition of phytoplankton and either expand the inland range of marine HABs or the seaward reach of inland HABs [108,283]. The following sections present perspectives on the potential for future HAB expansions not often addressed in the prior literature, namely, barriers to expansion and adaptation.
4.1. Cyanobacteria
Although the primary taxa associated with inland cyanoHABs are commonly referred to as freshwater cyanobacteria, analyses of the 2017 NLA data revealed that for most taxa, relative bloom frequencies are higher in brackish than in freshwater lakes (Section 2.2.1). The label “freshwater” to describe all inland cyanoHABs is, therefore, a misnomer. This is not a simple issue of semantics. Recent studies have shown that salt-tolerant Microcystis strains isolated from brackish coastal lagoons in the Netherlands [284,285] and Japan [117,286] bear genes that produce the compatible osmolyte sucrose, which is believed to help confer tolerance to saline environments. These brackish strains include one (PCC 7806) that was incorrectly regarded by earlier studies as being of freshwater origin [287]. As salinity is experimentally increased using NaCl, sucrose gene expression and sucrose production also increase in these salt-tolerant strains, and their growth remains relatively stable up to about 7–10 g NaCl L−1 [117,287,288]. Importantly, salt-intolerant Microcystis strains isolated from freshwater habitats do not have sucrose genes [285,286]. Also, a freshwater bloom composed of a mixed-species cyanobacterial assemblage seemed to have a limited capacity for long-term maintenance in experimental saline conditions, but Microcystis showed higher tolerance to acutely increased salinity than Dolichospermum [289]. These observations indicate that freshwater (salt-intolerant) and brackish (salt-tolerant) strains are genetically and physiologically distinct. There is also information suggesting that the acquisition of osmolyte genes may occur relatively quickly via intra- or inter-specific horizontal gene transfers [117,284,286] and that further strain diversification could occur by osmolyte gene losses and reacquisitions [286]. Genetic and comparative physiological studies on cyanoHAB strains found across salinity gradients in the USA are, to our knowledge, unavailable. This limits our understanding of their current biogeography as well as efforts to assess potential future changes in their distribution within inland waters and towards the sea. The increasing frequency of typically inland cyanoHABs and their toxins in coastal waters is in fact an emerging concern in the USA and globally [241,290]. The occurrence of cyanoHABs in coastal waters may be associated with contemporaneous transport in riverine flow in many cases, but there is also evidence suggesting that they can develop from endemic sources of propagules in some U.S. estuaries [114]. Further study seems warranted to determine if adaptation can be a mechanism for freshwater strains to overcome salinity barriers to seaward expansion and if transfer of pre-adapted, salt-tolerant inland strains can also lead to the same endpoint.
It should be noted that sucrose gene-positive cyanobacterial strains isolated from brackish coastal environments have been identified that seem to be intolerant to NaCl-based increases in salinity in the laboratory, e.g., M. aeruginosa [286] and Dolichospermum sp. [122]. The salt intolerance of these strains is difficult to reconcile with their saline origin and the presence of sucrose genes in their genome. Insights into these conflicting observations may be gained by exploring the effect of salinity on the growth of these strains using media of differing ionic compositions. In the euryhaline P. parvum, for example, the effect of salinity on growth is greatly influenced by the ionic composition of the culture medium (e.g., sulfate, magnesium; Section 4.2). Other studies have proposed that the acquisition of salt tolerance in strictly freshwater cyanobacterial strains may occur via spontaneous mutations under salinity stress [291]. Genetic characterization of the putative mutants could help evaluate this proposal.
4.2. Haptophytes
Although P. parvum was initially believed to be a native taxon in the USA [39], subsequent genetic analyses led to the conclusion that it is an invasive species [40]. Retrospective studies of water quality in Texas reservoirs showed that among the ambient traits commonly associated with P. parvum blooms, brackish conditions may have existed since shortly after the reservoirs were constructed (due to watershed and aquifer geochemistry), and their eutrophication preceded first bloom occurrences by at least 10 years or longer [137,198]. These water quality studies concluded that after the original invasion and establishment of P. parvum in Texas waters (per [40]), subsequent bloom expansions within Texas and beyond were due to P. parvum dispersal into pre-existing, bloom-favorable habitats. Yet, another study suggested that environmental conditions (favorable habitat), not dispersal, best explained the occurrence and distribution of P. parvum blooms in real time within a single lake; i.e., “everything is everywhere, but, the environment selects” [136]. For an invasive species, however, range expansions would require dispersal as an explanatory variable, especially for expansions across river basins.
Prymnesium parvum blooms now cover a large area of the conterminous USA (Figure 4), and whether the first blooms in Texas were due to invasion or environmental change may be a moot point from the perspective of applied HAB research. However, a better understanding of environmental conditions that facilitate expansions or trigger blooms could help prevent or predict and manage future blooms. Like cyanoHABs, the risk of P. parvum blooms expanding into estuaries is of particular concern. Prymnesium parvum has been detected within the tidal reach of some Texas rivers [92], and its ability to grow and become toxic in Texas estuarine waters has been demonstrated [292]. So why have blooms not occurred in Texas estuaries? In and of itself, increased salinity (based on NaCl addition) into the polyhaline range greatly suppresses P. parvum growth, but in the presence of elevated concentrations of specific ions (e.g., magnesium), growth is fully restored [143]. At lower salinities more typical of P. parvum’s inland habitats (5 psu), sulfate can also stimulate its growth [143]. Lundgren et al. [292] looked into biotic factors that may influence bloom formation in Texas estuarine waters and reported that the seasonal composition of planktonic communities may complicate P. parvum’s establishment in these waters. Based on these observations, Richardson and Patiño [293] suggested that biotic and abiotic conditions in the upper reach of intertidal zones may be serving as barriers to downstream expansion. Research into these and other possible scenarios could help understand—and maintain—current barriers to the seaward expansion of P. parvum blooms in Texas. Prymnesium parvum has been found in coastal waters of Washington and Maine, but there are no records of toxic blooms or detailed information about the species biogeography in these states (Section 2.1.2).
4.3. Euglenoids
The relatively recent discovery of toxigenic strains of E. sanguinea [42] took the field by surprise [43,44,45]. Although Euglena spp. were formally described in the early 1800s and were first recorded in U.S. inland waters since at least the early 1900s, toxic blooms were not documented until 2002, when E. sanguinea was identified as the cause of a fish-kill event in a North Carolina aquaculture pond [42]. Zimba et al. [43,44] provided several potential reasons for why toxic blooms were not detected earlier. For example, rapid shifts among life stages (e.g., from the flagellated stage to cysts or reproductive palmella) caused by acute weather changes could complicate identification of the causative toxic agent; the possibility of toxin production by cysts after they sink to the sediment; and the lack of chemical biomarkers and limited visual methods to identify cysts. These proposed explanations, although plausible, imply that toxic euglenoid blooms occurred prior to their first documentation but were erroneously ascribed to blooms of other toxigenic algae or to undetermined causes. Misidentification of toxic euglenoid blooms presumably would have occurred not only in the USA but also globally. There are alternative explanations that should also be considered.
Multiple reports of toxic blooms of E. sanguinea emerged from several U.S. states soon after the first toxic bloom was reported in 2002 [42,43,44]. Although increased attention to toxic euglenoids could partly explain the increased number of reports, their geographic scope to date has remained remarkably limited to the South Central and South Eastern regions of the country (Section 2.2.3). One scenario that could explain these observations is that a toxigenic strain or strains of E. sanguinea appeared for the first time around 2000 in the South Central or South Eastern USA, perhaps in a nutrient-rich aquaculture pond, and in the years that followed, they spread regionally to other aquaculture facilities raising warmwater fishes (e.g., catfish, tilapia, and striped bass [42,44]). This scenario is consistent with the existence of toxigenic and non-toxigenic strains of E. sanguinea and with the global scope of non-toxigenic strains combined with the far narrower, regional range of toxigenic strains. If this scenario is correct, management practices to prevent the spread of toxigenic strains would be advisable. Additional research on the biogeography and genetics of euglenoid strains responsible for toxic and non-toxic blooms seems warranted.
5. Summary and Conclusions
Blooms of the toxigenic cyanobacteria Anabaena, Aphanizomenon, Microcystis, and Planktothrix occur at higher relative frequency in oligohaline than freshwater lakes. Dolichospermum shows no significant salinity preferences, but its relative bloom rates are numerically higher in oligohaline waters. The relative detection rates of two cyanotoxins, microcystins and cylindrospermopsin, are also significantly higher in brackish than in freshwater lakes. These observations suggest that efforts to assess the risk of cyanoHAB occurrence and cyanotoxin exposure should account for lake salinity. Although blooms of the toxigenic haptophyte P. parvum have been documented exclusively in brackish waters, there is increasing evidence that this species may commonly occur in low-salinity freshwaters—perhaps explaining a case where a singular discharge of saline wastewater into a freshwater stream without a history of P. parvum blooms led to the acute formation of a toxic bloom. Potentially toxic Euglena spp. have the same relative rates of occurrence in freshwater and oligohaline lakes, showing that, like cyanobacteria, the inland distribution of this taxon is not restricted to freshwater lakes.
Studies on HAB biogeography should consider the ionic composition of inland waters separately from salinity. While little information is available for Euglena, major ions such as calcium, magnesium, and sulfate have all been associated with the growth and toxicity of cyanoHAB species and P. parvum. Sulfate presents a particularly interesting suite of possible effect scenarios in which this anion may directly or indirectly inhibit or stimulate (or both) harmful algal growth.
Nutrient inputs from agricultural and developed lands are well-established drivers of cultural lake eutrophication and HABs, but other land cover types may also influence HAB trends. Most harmful algae, especially those of widespread distribution, are “hardy” species that tolerate not only weather and climate extremes but also exposure to a variety of anthropogenic pollutants, including agricultural herbicides. Moreover, some commonly used herbicides such as glyphosate can stimulate the growth of HAB species (e.g., M. aeruginosa, P. parvum) at low, environmentally relevant concentrations. Thus, inputs of agrochemicals other than nutrients into surface waters could increase the risk of HABs. In addition, the loss of wetlands and riparian buffers may contribute to HAB occurrence because of the loss of multiple services that these lands provide to aquatic habitats, including the removal of nutrients and herbicides. A better understanding of HAB trends and their drivers from the individual lake to the global scale may be gained by paying increased attention to land uses other than fertilizer application and land cover types other than agriculture and development.
Like most other phytoplankton, cyanobacteria and Euglena normally grow between the spring and fall when day length and light intensity are relatively increased. Prymnesium parvum is unique in that its typical growth period in the USA is the reverse, from the fall to spring, even though temperature and light conditions are suboptimal. The growth season of P. parvum may not be the result of environmental preferences but of growth opportunities (assisted by its toxin production and mixotrophy) at a time when most other taxa are dormant. Cold water blooms of some cyanobacteria may also be of increasing importance, and research has shown that saxitoxin may be associated with lower water temperatures and that some cylindrospermopsin-producing species bloom in winter.
The toxin classes covered herein include neurotoxins, hepatotoxins, and ichthyotoxins. However, some toxins may have multiple modes of action, and blooms may contain organisms that produce multiple toxins. The neurotoxins, anatoxin and saxitoxin, have a wide global distribution but many of the literature examples indicate production from benthic taxa. The distribution of the hepatotoxin cylindrospermopsin is concentrated primarily in Mid-Atlantic States, Midwestern States including Texas, and Florida; however, detections seem to have expanded from tropical and subtropical areas into temperate zones. Microcystin is more widely distributed than cylindrospermopsin but seems to be concentrated in Central USA. The ichthyotoxins, euglenophycin and prymnesins, can cause harm to fish and other aquatic organisms, along with sublethal and ecological effects. Information on the geographic range of euglenophycin is limited, but the toxin has been detected in fish culture ponds in the South Central and Southeastern USA. Prymnesins have been detected in sites from a wide geographic range, primarily in the temperate and subtropical zones. Neurotoxins and hepatotoxins can bioaccumulate in organisms exposed to these toxins, which suggests that bioaccumulation is a possible exposure route to larger organisms. Little evidence of bioaccumulation of euglenophycin or prymnesins was found in the literature.
There is evidence suggesting that inland cyanoHAB species may be gaining a foothold in U.S. estuaries. The basis for this apparent seaward expansion is uncertain. However, inland strains isolated from saline coastal waters elsewhere have acquired genes whose function may be to allow their growth in brackish waters, thus suggesting that adaptation may be a means for freshwater strains to overcome instream salinity barriers. Dispersal of inland brackish strains into estuaries via natural or anthropogenic vectors could also result in the seaward expansion of inland cyanoHABs. Genetic and comparative physiological studies of strains from freshwater and brackish environments (inland and estuarine) would help evaluate the feasibility of these scenarios. Additional research could also improve our understanding of why blooms of the highly euryhaline P. parvum have not occurred in U.S. estuaries and why the distribution of toxic E. sanguinea blooms is restricted to a specific region of the conterminous USA despite the global distribution of the species and its blooms.
Funding
This work was not supported by grant funding.
Data Availability Statement
No original data were generated during this research.
Acknowledgments
The authors wish to thank David Creamer for assistance with the map in Figure 4. The Texas Cooperative Fish and Wildlife Research Unit is jointly supported by the U.S. Geological Survey, Texas Tech University, Texas Parks and Wildlife Department, The Wildlife Management Institute, and U.S. Fish and Wildlife Service. This journal article has been peer reviewed and approved for publication consistent with USGS Fundamental Science Practices (https://pubs.usgs.gov/circ/1367/; accessed on 1 July 2023). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. All authors have read and agreed to the published version of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Codd, G.A.; Pliński, M.; Surosz, W.; Hutson, J.; Fallowfield, H.J. Publication in 1672 of Animal Deaths at the Tuchomskie Lake, Northern Poland and a Likely Role of Cyanobacterial Blooms. Toxicon 2015, 108, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.B.; Jewett, E.B.; Dortch, Q.; Walton, B.T.; Hudnell, H.K. Scientific Assessment of Freshwater Harmful Algal Blooms; Interagency Working Group on Harmful Algal Blooms, Hypoxia, and Human Health of the Joint Subcommittee on Ocean Science and Technology: Washington, DC, USA, 2008. [Google Scholar]
- Roelke, D.L.; Barkoh, A.; Brooks, B.W.; Grover, J.P.; Hambright, K.D.; Laclaire, J.W.; Moeller, P.D.R.; Patino, R. A Chronicle of a Killer Alga in the West: Ecology, Assessment, and Management of Prymnesium parvum Blooms. Hydrobiologia 2016, 764, 29–50. [Google Scholar] [CrossRef]
- Trainer, V.L. GlobalHAB: Evaluating, Reducing, and Mitigating the Cost of Harmful Algal Blooms: A Compendium of Case Studies. PICES Sci. Rep. 2020, 59, 107. [Google Scholar]
- Pindilli, E.J.; Loftin, K. What ’s It Worth? Estimating the Potential Value of Early Warnings of Cyanobacterial Harmful Algal Blooms for Managing Freshwater Reservoirs in Kansas, United States. Front. Environ. Sci. 2022, 10, 805165. [Google Scholar] [CrossRef]
- Nabout, J.C.; da Silva Rocha, B.; Carneiro, F.M.; Sant’Anna, C.L. How Many Species of Cyanobacteria Are There? Using a Discovery Curve to Predict the Species Number. Biodivers. Conserv. 2013, 22, 2907–2918. [Google Scholar] [CrossRef]
- Bernard, C.; Ballot, A.; Thomazeau, S.; Maloufi, S.; Furey, A.; Mankiewicz-Boczek, J.; Pawlik-Skowrońska, B.; Capelli, C.; Salmaso, N. Appendix 2: Cyanobacteria Associated with the Production of Cyanotoxins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 501–525. [Google Scholar] [CrossRef] [Green Version]
- Yoo, R.S.; Carmichael, W.W.; Hoehn, R.C.; Hrudey, S.E. Cyanobacterial (Blue-Green Algal) Toxins: A Resource Guide; AWWA Foundation: Denver, CO, USA, 1995. [Google Scholar]
- Hudnell, H.K. Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 9780387758640. [Google Scholar]
- Graham, J.L.; Foster, G.M.; Williams, T.J.; Kramer, A.R.; Harris, T.D. Occurrence of Cyanobacteria, Microcystin, and Taste-and-Odor Compounds in Cheney Reservoir, Kansas, 2001–2016; US Geological Survey: Lawrence, KS, USA, 2017. [Google Scholar]
- Anderson, D.M.; Fensin, E.; Gobler, C.J.; Hoeglund, A.E.; Hubbard, K.A.; Kulis, D.M.; Landsberg, J.H.; Lefebvre, K.A.; Provoost, P.; Richlen, M.L.; et al. Marine Harmful Algal Blooms (HABs) in the United States: History, Current Status and Future Trends. Harmful Algae 2021, 102, 101975. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, E.S. Epidemic of Intestinal Disorders in Charlston, W. VA., Occuring Simulataneously with Unprecedented Water Supply Conditions. Public Health Eng. 1931, 21, 198–200. [Google Scholar] [CrossRef]
- Lippy, E.C.; Erb, J. Gastrointestinal Illness at Sewickley, Pa. AWWA J. 1976, 68, 606–610. [Google Scholar] [CrossRef]
- Hilborn, E.D.; Roberts, V.A.; Backer, L.; DeConno, E.; Egan, J.S.; Hyde, J.B.; Nicholas, D.C.; Wiegert, E.J.; Billing, L.M.; DiOrio, M.; et al. Algal Bloom-Associated Disease Outbreaks among Users of Freshwater Lakes—United States, 2009–2010. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 11–15. [Google Scholar]
- Trevino-Garrison, I.; Dement, J.; Ahmed, F.S.; Haines-Lieber, P.; Langer, T.; Ménager, H.; Neff, J.; Van Der Merwe, D.; Carney, E. Human Illnesses and Animal Deaths Associated with Freshwater Harmful Algal Blooms—Kansas. Toxins 2015, 7, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Figgat, M.; Muscatiello, N.; Wilson, L.; Dziewulski, D. Harmful Algal Bloom–Associated Illness Surveillance: Lessons from Reported Hospital Visits in New York, 2008–2014. Am. J. Public Health 2016, 106, 440–442. [Google Scholar] [CrossRef] [PubMed]
- U.S. Centers for Disease Control and Prevention OHHAB Data. Available online: https://www.cdc.gov/habs/data/index.html (accessed on 19 April 2023).
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; World Health Organization Report; World Health Organization: London, UK, 1999. [Google Scholar]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press on Behalf of the World Health Organization: Boca Raton, FL, USA, 2021; ISBN 9781626239777. [Google Scholar]
- U.S. Center for Disease Control and Prevention One Health Harmful Algal Bloom System (OHHABs). Available online: https://www.cdc.gov/habs/ohhabs.html (accessed on 19 April 2023).
- Rose, E.T. Toxic Algae in Iowa Lakes. Proc. Iowa Acad. Sci. 1953, 60, 738–745. [Google Scholar]
- Carmichael, W.W. The Toxins of Cyanobacteria. Sci. Am. 1994, 270, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global Geographical and Historical Overview of Cyanotoxin Distribution and Cyanobacterial Poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Wacklin, P.; Hoffmann, L.; Komárek, J. Nomenclatural Validation of the Genetically Revised Cyanobacterial Genus Dolichospermum (Ralfs Ex Bornet et Flahault) Comb, Nova. Fottea 2009, 9, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J. Modern Taxonomic Revision of Planktic Nostocacean Cyanobacteria: A Short Review of Genera. Hydrobiologia 2010, 639, 231–243. [Google Scholar] [CrossRef]
- Suda, S.; Watanabe, M.M.; Otsuka, S.; Mahakahant, A.; Yongmanitchai, W.; Nopartnaraporn, N.; Liu, Y.; Day, J.G. Taxonomic Revision of Water-Bloom-Forming Species of Oscillatorioid Cyanobacteria. Int. J. Syst. Evol. Microbiol. 2002, 52, 1577–1595. [Google Scholar] [CrossRef]
- Hardy, F.J.; Preece, E.; Backer, L. Status of State CyanoHAB Outreach and Monitoring Efforts, United States. Lake Reserv. Manag. 2021, 37, 246–260. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. National Lakes Assessment: A Collaborative Survey of the Nation’s Lakes; U.S. Environmental Protection Agency: Washington, DC, USA, 2009. Available online: https://www.epa.gov/national-aquatic-resource-surveys/nla (accessed on 15 August 2022).
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in Inland Lakes of the United States: Occurrence and Potential Recreational Health Risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [Green Version]
- U.S. Environmental Protection Agency. National Lakes Assessment: A Collaborative Survey of the Nation’s Lakes; U.S. Environmental Protection Agency: Washington, DC, USA, 2017. Available online: https://www.epa.gov/national-aquatic-resource-surveys/nla (accessed on 15 August 2022).
- Graham, L.E.; Graham, J.M.; Wilcox, L.W.; Cook, M.E. Algae, 3rd ed.; LJLM Press: Upper Saddle River, NJ, USA, 2016; ISBN 9780986393532. [Google Scholar]
- Nicholls, H.H. Haptophyte Algae. In Freshwater Algae of North America; Wehr, J.D., Sheath, R.G., Kociolek, J.P., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 587–605. [Google Scholar]
- Reifel, K.M.; McCoy, M.P.; Tiffany, M.A.; Rocke, T.E.; Trees, C.C.; Barlow, S.B.; Faulkner, D.J.; Hurlbert, S.H. Pleurochrysis pseudoroscoffensis (Prymnesiophyceae) Blooms on the Surface of the Salton Sea, California. Hydrobiologia 2001, 466, 177–185. [Google Scholar] [CrossRef]
- Nicholls, K.H.; Beaver, J.L.; Estabrook, R.H. Lakewide Odours in Ontario and New Hampshire Caused by Chrysochromulina breviturrita Nich. (Prymnesiophyceae). Hydrobiologia 1982, 96, 91–95. [Google Scholar] [CrossRef]
- Moestrup, Ø. Economic Aspects: ‘Blooms,’ Nuisance Species and Toxins. In The Haptophyte Algae; Green, J.C., Leadbeater, B.S.C., Eds.; Clarendon Press: Oxford, UK, 1994; pp. 265–285. [Google Scholar]
- Southard, G.M.; Fries, L.T.; Barkoh, A. Prymnesium parvum: The Texas Experience. J. Am. Water Resour. Assoc. 2010, 46, 14–23. [Google Scholar] [CrossRef]
- Free, G.; Van de Bund, W.; Gawlik, B.M.; Van Wijk, L.; Wood, M.; Guagnini, E.; Koutelos, K.; Annunziato, A.; Grizzetti, B.; Vigiak, O.; et al. An EU Analysis of the Ecological Disaster in the Oder River of 2022; Publications Office of the European Union: Luxembourg, 2023; ISBN 9789276993148. [Google Scholar]
- James, T.L.; De La Cruz, A. Prymnesium parvum Carter (Chrysophyceae) as a Suspect of Mass Mortalities of Fish and Shellfish Communities in Western Texas. Texas J. Sci. 1989, 41, 429–430. [Google Scholar]
- Rhodes, K.; Hubbs, C. Recovery of Pecos River Fishes from a Red Tide Fish Kill. Southwest. Nat. 1992, 37, 178–187. [Google Scholar] [CrossRef]
- Lutz-Carrillo, D.J.; Southard, G.M.; Fries, L.T. Global Genetic Relationships among Isolates of Golden Alga (Prymnesium parvum). J. Am. Water Resour. Assoc. 2010, 46, 24–32. [Google Scholar] [CrossRef]
- Triemer, R.E.; Zakryś, B. Photosynthetic Euglenoids. In Freshwater Algae of North America; Wehr, J.D., Sheath, R.G., Kociolek, J.P., Eds.; Academic Press: San Diego, California, USA, 2015; pp. 459–483. [Google Scholar]
- Zimba, P.V.; Rowan, M.; Triemer, R. Identification of Euglenoid Algae That Produce Ichthyotoxin(S). J. Fish Dis. 2004, 27, 115–117. [Google Scholar] [CrossRef]
- Zimba, P.V.; Huang, I.S.; Gutierrez, D.; Shin, W.; Bennett, M.S.; Triemer, R.E. Euglenophycin Is Produced in at Least Six Species of Euglenoid Algae and Six of Seven Strains of Euglena sanguinea. Harmful Algae 2017, 63, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Zimba, P.V.; Moeller, P.D.; Beauchesne, K.; Lane, H.E.; Triemer, R.E. Identification of Euglenophycin—A Toxin Found in Certain Euglenoids. Toxicon 2010, 55, 100–104. [Google Scholar] [CrossRef]
- Janse van Vuuren, S.; Levanets, A. Mass Developments of Euglena sanguinea Ehrenberg in South Africa. Afr. J. Aquat. Sci. 2021, 46, 110–122. [Google Scholar] [CrossRef]
- Bevel, N. Some Notes on the Protozoa of Reelfoot Lake. J. Tennessee Acad. Sci. 1937, 13, 137–159. [Google Scholar]
- Johnson, L.P. Euglenae of Iowa. Trans. Am. Microsc. Soc. 1944, 2, 97–135. [Google Scholar] [CrossRef]
- Zakryś, B.; Walne, P.L. Floristic, Taxonomic and Phytogeographic Studies of Green Euglenophyta from the Southeastern United States, with Emphasis on New and Rare Species. Arch. Fur Hydrobiol. 1994, 72, 71–114. [Google Scholar]
- Gojdics, M. Some Observations of Euglena Sanguinea EHRBG. Trans. Am. Microsc. Soc. 1939, 58, 241–248. [Google Scholar] [CrossRef]
- Cirés, S.; Ballot, A. A Review of the Phylogeny, Ecology and Toxin Production of Bloom-Forming Aphanizomenon Spp. and Related Species within the Nostocales (Cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef]
- Li, X.; Dreher, T.W.; Li, R. An Overview of Diversity, Occurrence, Genetics and Toxin Production of Bloom-Forming Dolichospermum (Anabaena) Species. Harmful Algae 2016, 54, 54–68. [Google Scholar] [CrossRef] [Green Version]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A Review of the Global Ecology, Genomics, and Biogeography of the Toxic Cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.A.; Kelly, L.T.; Bouma-Gregson, K.; Humbert, J.F.; Laughinghouse, H.D.; Lazorchak, J.; McAllister, T.G.; McQueen, A.; Pokrzywinski, K.; Puddick, J.; et al. Toxic Benthic Freshwater Cyanobacterial Proliferations: Challenges and Solutions for Enhancing Knowledge and Improving Monitoring and Mitigation. Freshw. Biol. 2020, 65, 1824–1842. [Google Scholar] [CrossRef]
- Reinl, K.L.; Brookes, J.D.; Carey, C.C.; Harris, T.D.; Ibelings, B.W.; Morales-Williams, A.M.; De Senerpont Domis, L.N.; Atkins, K.S.; Isles, P.D.F.; Mesman, J.P.; et al. Cyanobacterial Blooms in Oligotrophic Lakes: Shifting the High-Nutrient Paradigm. Freshw. Biol. 2021, 66, 1846–1859. [Google Scholar] [CrossRef]
- Komárek, J.; Johansen, J.R. Coccoid Cyanobacteria. In Freshwater Algae of North America; Wehr, J.D., Sheath, R.G., Kociolek, J.P., Eds.; Academic Press: London, UK; Elsevier: Amsterdam, The Netherlands, 2015; pp. 75–133. [Google Scholar]
- Krztoń, W.; Kosiba, J.; Pociecha, A.; Wilk-Woźniak, E. The Effect of Cyanobacterial Blooms on Bio- and Functional Diversity of Zooplankton Communities. Biodivers. Conserv. 2019, 28, 1815–1835. [Google Scholar] [CrossRef] [Green Version]
- Falconer, I.R. Algal Toxins and Human Health. Qual. Treat. Drink. Water II 1998, 5, 53–82. [Google Scholar] [CrossRef]
- Meissner, K.; Dittmann, E.; Börner, T. Toxic and Non-Toxic Strains of the Cyanobacterium Microcystis aeruginosa Contain Sequences Homologous to Peptide Synthetase Genes. FEMS Microbiol. Lett. 1996, 135, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Bertos-Fortis, M.; Farnelid, H.M.; Lindh, M.V.; Casini, M.; Andersson, A.; Pinhassi, J.; Legrand, C. Unscrambling Cyanobacteria Community Dynamics Related to Environmental Factors. Front. Microbiol. 2016, 7, 625. [Google Scholar] [CrossRef]
- Ribeiro, K.F.; Duarte, L.; Crossetti, L.O. Everything Is Not Everywhere: A Tale on the Biogeography of Cyanobacteria. Hydrobiologia 2018, 820, 23–48. [Google Scholar] [CrossRef]
- Kaplan-Levy, R.N.; Hadas, O.; Summers, M.L.; Rucker, J.; Sukenik, A. Akinetes: Dormant Cells of Cyanobacteria. In Dormancy and Resistance in Harsh Environments, Topics in Current Genetics; Lubenz, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 21, ISBN 978-3-642-12421-1. [Google Scholar]
- Moustaka-Gouni, M.; Hiskia, A.; Genitsaris, S.; Katsiapi, M.; Manolidi, K.; Zervou, S.K.; Christophoridis, C.; Triantis, T.M.; Kaloudis, T.; Orfanidis, S. First Report of Aphanizomenon favaloroi Occurrence in Europe Associated with Saxitoxins and a Massive Fish Kill in Lake Vistonis, Greece. Mar. Freshw. Res. 2017, 68, 793–800. [Google Scholar] [CrossRef]
- Cirés, S.; Casero, M.C.; Quesada, A. Toxicity at the Edge of Life: A Review on Cyanobacterial Toxins from Extreme Environments. Mar. Drugs 2017, 15, 233. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S.; Oliver, R.L.; Walsby, A.E. Cyanobacterial Dominance: The Role of Buoyancy Regulation in Dynamic Lake Environments. N. Z. J. Mar. Freshw. Res. 1987, 21, 379–390. [Google Scholar] [CrossRef]
- Reynolds, C.S. What Factors Influence the Species Composition of Phytoplankton in Lakes of Different Trophic Status? Hydrobiologia 1998, 369–370, 11–26. [Google Scholar] [CrossRef]
- Graham, J.L.; Loftin, K.A.; Ziegler, A.C.; Meyer, M.T. Guidelines for Design and Sampling for Cyanobacterial Toxin and Taste-and-Odor Studies in Lakes and Reservoirs; U.S. Geological Survey Scientific Investigations Report 2008-5038; U.S. Geological Survey: Reston, VA, USA, 2008. [Google Scholar]
- Carmichael, W.W.; Boyer, G.L. Health Impacts from Cyanobacteria Harmful Algae Blooms: Implications for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial Heterocysts. Cold Spring Harb. Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef] [Green Version]
- Meeks, J.C.; Elhai, J. Regulation of Cellular Differentiation in Filamentous Cyanobacteria in Free-Living and Plant-Associated Symbiotic Growth States. Microbiol. Mol. Biol. Rev. 2002, 66, 94–121. [Google Scholar] [CrossRef] [Green Version]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the Coverage of the Cyanobacterial Phylum Using Diversity-Driven Genome Sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, B.L. Light-Dependent Governance of Cell Shape Dimensions in Cyanobacteria. Front. Microbiol. 2015, 6, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.P.; Montgomery, B.L. Determining Cell Shape: Adaptive Regulation of Cyanobacterial Cellular Differentiation and Morphology. Trends Microbiol. 2011, 19, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Fofonoff, N.P.; Millard, R.C., Jr. Algorithms for Computation of Fundamental Properties of Seawater; UNESCO: Paris, France, 1983. [Google Scholar]
- Hill, K.D.; Dauphinee, T.M.; Woods, D.J. The Extension of the Practical Salinity Scale 1978 to Low Salinities. IEEE J. Ocean. Eng. 1986, 11, 109–112. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Li, R. Cyanobacteria Toxins in the Salton Sea. Saline Syst. 2006, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Marcarelli, A.M.; Wurtsbaugh, W.A.; Griset, O. Salinity Controls Phytoplankton Response to Nutrient Enrichment in the Great Salt Lake, Utah, USA. Can. J. Fish. Aquat. Sci. 2006, 63, 2236–2248. [Google Scholar] [CrossRef] [Green Version]
- Galat, D.L.; Verdin, J.P.; Sims, L.L. Large-Scale Patterns of Nodularia spumigena Blooms in Pyramid Lake, Nevada, Determined from Landsat Imagery: 1972–1986. Hydrobiologia 1990, 197, 147–164. [Google Scholar] [CrossRef]
- Granéli, E.; Edvardsen, B.; Roelke, D.L.; Hagström, J.A. The Ecophysiology and Bloom Dynamics of Prymnesium spp. Harmful Algae 2012, 14, 260–270. [Google Scholar] [CrossRef]
- Binzer, S.B.; Svenssen, D.K.; Daugbjerg, N.; Alves-de-Souza, C.; Pinto, E.; Hansen, P.J.; Larsen, T.O.; Varga, E. A-, B- and C-Type Prymnesins Are Clade Specific Compounds and Chemotaxonomic Markers in Prymnesium parvum. Harmful Algae 2019, 81, 10–17. [Google Scholar] [CrossRef]
- Wisecaver, J.H.; Auber, R.P.; Pendleton, A.L.; Manning, S.R.; Moore, B.S.; Driscoll, W.W.; Wisecaver, J.H.; Auber, R.P.; Pendleton, A.L.; Watervoort, N.F.; et al. Article Extreme Genome Diversity and Cryptic Speciation in a Harmful Algal-Bloom-Forming Eukaryote. Curr. Biol. 2023, 33, 2246–2259.e8. [Google Scholar] [CrossRef]
- Edvardsen, B.; Paasche, E. Bloom Dynamics and Physiology of Prymnesium and Chrysochromulina. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998; pp. 193–208. [Google Scholar]
- Medlin, L.K.; Kooistra, W.H.C.F.; Potter, D.; Saunders, G.W.; Andersen, R.A. Phylogenetic Relationships of the ‘Golden Algae’ (Haptophytes, Heterokont Chromophytes) and Their Plastids. In Origins of Algae and their Plastids. Plant Systematics and Evolution; Bhattacharya, D., Ed.; Springer: Vienna, Austria, 1997; pp. 187–219. [Google Scholar]
- Roelke, D.L.; Errera, R.M.; Riesling, R.; Brooks, B.W.; Grover, J.P.; Schwierzke, L.; Ureña-Boeck, F.; Baker, J.; Pinckney, J.L. Effects of Nutrient Enrichment on Prymnesium parvum Population Dynamics and Toxicity: Results from Field Experiments, Lake Possum Kingdom, USA. Aquat. Microb. Ecol. 2007, 46, 125–140. [Google Scholar] [CrossRef]
- VanLandeghem, M.M.; Denny, S.; Patiño, R. Predicting the Risk of Toxic Blooms of Golden Alga from Cell Abundance and Environmental Covariates. Limnol. Oceanogr. Methods 2015, 13, 568–586. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, W.W.; Espinosa, N.J.; Eldakar, O.T.; Hackett, J.D. Allelopathy as an Emergent, Exploitable Public Good in the Bloom-Forming Microalga Prymnesium parvum. Evolution 2013, 67, 1582–1590. [Google Scholar] [CrossRef] [Green Version]
- Hambright, K.D.; Zamor, R.M.; Easton, J.D.; Glenn, K.L.; Remmel, E.J.; Easton, A.C. Temporal and Spatial Variability of an Invasive Toxigenic Protist in a North American Subtropical Reservoir. Harmful Algae 2010, 9, 568–577. [Google Scholar] [CrossRef]
- Clayton, J.B.; Patiño, R.; Rashel, R.H.; Tábora-Sarmiento, S. Water Quality Associations and Spatiotemporal Distribution of the Harmful Alga Prymnesium parvum in an Impounded Urban Stream System. J. Urban Ecol. 2021, 7, juab011. [Google Scholar] [CrossRef]
- Wagstaff, B.A.; Pratscher, J.; Rivera, P.P.L.; Hems, E.S.; Brooks, E.; Rejzek, M.; Todd, J.D.; Murrell, J.C.; Field, R.A. Assessing the Toxicity and Mitigating the Impact of Harmful Prymnesium Blooms in Eutrophic Waters of the Norfolk Broads. Environ. Sci. Technol. 2021, 55, 16538–16551. [Google Scholar] [CrossRef]
- Green, J.C.; Hibberd, D.J.; Pienaar, R.N. The Taxonomy of Prymnesium (Prymnesiophyceae) Including a Description of a New Cosmopolitan Species, P. Patellifera Sp. Nov., and Further Observations on P. Parvum N. Carter. Br. Phycol. J. 1982, 17, 363–382. [Google Scholar] [CrossRef] [Green Version]
- Li, W.K.W.; Andersen, R.A.; Gifford, D.J.; Incze, L.S.; Martin, J.L.; Pilskaln, C.H.; Rooney-Varga, J.N.; Sieracki, M.E.; Wilson, W.H.; Wolff, N.H. Planktonic Microbes in the Gulf of Maine Area. PLoS ONE 2011, 6, e20981. [Google Scholar] [CrossRef]
- Nelson, J.; Byrd, M. Occurrence of Prymnesium parvum (Golden Alga) in Texas Intertidal Waters; Texas Parks and Wildlife Department Management Data Series 264; 2011. Available online: https://texashistory.unt.edu/ark:/67531/metapth577898/m2/1/high_res_d/txca-0908.pdf (accessed on 1 March 2023).
- Roelke, D.L.; Schwierzke, L.; Brooks, B.W.; Grover, J.P.; Errera, R.M.; Valenti, T.W.; Pinckney, J.L. Factors Influencing Prymnesium parvum Population Dynamics during Bloom Initiation: Results from in-Lake Mesocosm Experiments. J. Am. Water Resour. Assoc. 2010, 46, 76–91. [Google Scholar] [CrossRef]
- Sager, R.; Barkoh, A.A.; Buzan, D.L.; Fries, L.T.; Guss, J.A.; Kurten, G.L.; Ralph, J.J.; Srnghurst, E.J.; Southard, G.M.; Swanson, E. Prymnesium parvum: A Potential Threat to U.S. Reservoirs. Am. Fish. Soc. Symp. 2008, 62, 261–273. [Google Scholar]
- Israël, N.M.D.; VanLandeghem, M.M.; Denny, S.; Ingle, J.; Patiño, R. Golden Alga Presence and Abundance Are Inversely Related to Salinity in a High-Salinity River Ecosystem, Pecos River, USA. Harmful Algae 2014, 39, 81–91. [Google Scholar] [CrossRef]
- Caron, D.A.; Lie, A.A.Y.; Buckowski, T.; Turneer, J.; Frabotta, K. The Effect of pH and Salinity on the Toxicity and Growth of the Golden Alga, Prymnesium parvum. Protist 2023, 174, 125927. [Google Scholar] [CrossRef]
- Hindák, F.; Wołowski, K.; Hindáková, A. Cysts and Their Formation in Some Neustonic Euglena Species. Ann. Limnol. 2000, 36, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.L.; Dubrovsky, N.M.; Eberts, S.M. Cyanobacterial Harmful Algal Blooms and U.S. Geological Survey Science Capabilities: U.S. Geological Survey Open-File Report 2016-1174; US Department of the Interior, US Geological Survey: Reston, VA, USA, 2016. [Google Scholar]
- Binoy, T.T.; Ponnachan, V.T.; Mavila, S.S. Outbreak of Harmful Algal Blooms (HABs) in the Sacred River of Achenkoil, Kerala, India during the Post Flood Season. Indian Hydrobiol. 2020, 19, 229–240. [Google Scholar]
- Rahman, M.M.; Jewel, M.A.S.; Khan, S.; Haque, M.M. Study of Euglenophytes Bloom and It’s Impact on Fish Growth in Bangladesh. Algae 2007, 22, 185–192. [Google Scholar] [CrossRef]
- Xavier, M.B.; Mainardes Pinto, C.S.R.; Takino, M. Euglena sanguinea Ehrenberg Bloom in a Fish-Breeding Tank (Pindamonhangaba, São Paulo, Brazil). Arch. Hydrobiol. Suppl. Algol. Stud. 1991, 26, 133–142. [Google Scholar]
- Zakryś, B.; Karnkowska-Ishikawa, A.; ŁUkomska-Kowalczyk, M.; Milanowski, R. A New Photosynthetic Euglenoid Isolated in Poland: Euglenaria clepsydroides Sp. Nov. (Euglenea). Eur. J. Phycol. 2013, 48, 260–267. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful Freshwater Algal Blooms, with an Emphasis on Cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Otten, T.G. Duelling “CyanoHABs”: Unravelling the Environmental Drivers Controlling Dominance and Succession among Diazotrophic and Non-N2-Fixing Harmful Cyanobacteria. Environ. Microbiol. 2016, 18, 316–324. [Google Scholar] [CrossRef]
- Graham, J.L.; Dubrovsky, N.M.; Foster, G.M.; King, L.R.; Loftin, K.A.; Rosen, B.H.; Stelzer, E.A. Cyanotoxin Occurrence in Large Rivers of the United States. INL Waters 2020, 10, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Nietch, C.T.; Gains-germain, L.; Lazorchak, J.; Keely, S.P.; Youngstrom, G.; Urichich, E.M.; Astifan, B.; Dasilva, A.; Mayfield, H. Development of a Risk Characterization Tool for Harmful Cyanobacteria Blooms on the Ohio River. Water 2022, 14, 644. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. National Lakes Assessment 2017 Field Operations Manual EPA 841-B-16-002; U.S. Environmental Protection Agency: Washington, DC, USA, 2017. [Google Scholar]
- Bargu, S.; Justic, D.; White, J.R.; Lane, R.; Day, J.; Paerl, H.; Raynie, R. Mississippi River Diversions and Phytoplankton Dynamics in Deltaic Gulf of Mexico Estuaries: A Review. Estuar. Coast. Shelf Sci. 2019, 221, 39–52. [Google Scholar] [CrossRef]
- Lehman, P.W.; Boyer, G.; Hall, C.; Waller, S.; Gehrts, K. Distribution and Toxicity of a New Colonial Microcystis aeruginosa Bloom in the San Francisco Bay Estuary, California. Hydrobiologia 2005, 541, 87–99. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Banack, S.A.; Powell, J.T.; Tymm, F.J.M.; Murch, S.J.; Brand, L.E.; Cox, P.A. Public Health Responses to Toxic Cyanobacterial Blooms: Perspectives from the 2016 Florida Event. Water Policy 2018, 20, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, J.S.; Banack, S.A.; Wessel, R.A.; Lester, M.; Pim, J.G.; Cassani, J.R.; Cox, P.A. Toxin Analysis of Freshwater Cyanobacterial and Marine Harmful Algal Blooms on the West Coast of Florida and Implications for Estuarine Environments. Neurotox. Res. 2021, 39, 27–35. [Google Scholar] [CrossRef]
- Sellner, K.G.; Lacouture, R.V.; Parrish, C.R. Effects of Increasing Salinity on a Cyanobacteria Bloom in the Potomac River Estuary. J. Plankton Res. 1988, 10, 49–61. [Google Scholar] [CrossRef]
- Ross, C.; Santiago-Vázquez, L.; Paul, V. Toxin Release in Response to Oxidative Stress and Programmed Cell Death in the Cyanobacterium Microcystis aeruginosa. Aquat. Toxicol. 2006, 78, 66–73. [Google Scholar] [CrossRef]
- Lehman, P.W.; Kurobe, T.; Teh, S.J. Impact of Extreme Wet and Dry Years on the Persistence of Microcystis Harmful Algal Blooms in San Francisco Estuary. Quat. Int. 2022, 621, 16–25. [Google Scholar] [CrossRef]
- Joshi, S.M.; Jackson, L.J. How Might Changing Climate Limit Cyanobacteria Growth in Shallow Prairie Lakes? An Empirical Space-for-Time Evaluation of the Potential Role of Increasing Sulfate. Adv. Environ. Eng. Res. 2022, 3. [Google Scholar] [CrossRef]
- Marino, R.; Howarth, R.W.; Shames, J.; Prepas, E. Molybdenum and Sulfate as Controls on the Abundance of Nitrogen-Fixing Cyanobacteria in Saline Lakes in Alberta. Limnol. Oceanogr. 1990, 35, 245–259. [Google Scholar] [CrossRef]
- Tanabe, Y.; Hodoki, Y.; Sano, T.; Tada, K.; Watanabe, M.M. Adaptation of the Freshwater Bloom-Forming Cyanobacterium Microcystis aeruginosa to Brackish Water Is Driven by Recent Horizontal Transfer of Sucrose Genes. Front. Microbiol. 2018, 9, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albay, M.; Matthiensen, A.; Codd, G.A. Occurrence of Toxic Blue-Green Algae in the Kucukcekmece Lagoon (Istanbul, Turkey). Environ. Toxicol. 2005, 20, 277–284. [Google Scholar] [CrossRef]
- Bormans, M.; Savar, V.; Legrand, B.; Mineaud, E.; Robert, E.; Lance, E.; Amzil, Z. Cyanobacteria and Cyanotoxins in Estuarine Water and Sediment. Aquat. Ecol. 2020, 54, 625–640. [Google Scholar] [CrossRef]
- Trung, B.; Dao, T.S.; Faassen, E.; Lürling, M. Cyanobacterial Blooms and Microcystins in Southern Vietnam. Toxins 2018, 10, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, M.; Suikkanen, S.; Kobos, J.; Wasmund, N.; Karlson, B. Basin-Specific Changes in Filamentous Cyanobacteria Community Composition across Four Decades in the Baltic Sea. Harmful Algae 2020, 91, 101685. [Google Scholar] [CrossRef] [PubMed]
- Teikari, J.E.; Popin, R.V.; Hou, S.; Wahlsten, M.; Hess, W.R.; Sivonen, K. Insight into the Genome and Brackish Water Adaptation Strategies of Toxic and Bloom-Forming Baltic Sea Dolichospermum sp. UHCC 0315. Sci. Rep. 2019, 9, 4888. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Lürling, M. Calcium Promotes Formation of Large Colonies of the Cyanobacterium Microcystis by Enhancing Cell-Adhesion. Harmful Algae 2020, 92, 101768. [Google Scholar] [CrossRef]
- Gu, P.; Li, Q.; Zhang, W.; Zheng, Z.; Luo, X. Effects of Different Metal Ions (Ca, Cu, Pb, Cd) on Formation of Cyanobacterial Blooms. Ecotoxicol. Environ. Saf. 2020, 189, 109976. [Google Scholar] [CrossRef]
- Shi, J.Q.; Wu, Z.X.; Song, L.R. Physiological and Molecular Responses to Calcium Supplementation in Microcystis aeruginosa (Cyanobacteria). N. Z. J. Mar. Freshw. Res. 2013, 47, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Wei, K.; Jung, S.; Amano, Y.; Machida, M. Control of the Buoyancy of Microcystis aeruginosa via Colony Formation Induced by Regulating Extracellular Polysaccharides and Cationic Ions. SN Appl. Sci. 2019, 1, 1573. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Lu, L.; Li, M.; Xu, Z.; Zhu, W. Effects of Ca and Mg Levels on Colony Formation and EPS Content of Cultured M. aeruginosa. Procedia Environ. Sci. 2011, 10, 1452–1458. [Google Scholar] [CrossRef] [Green Version]
- Parker, D.L.; Kumar, H.D.; Rai, L.C.; Singh, J.B. Potassium Salts Inhibit Growth of the Cyanobacteria Microcystis Spp. in Pond Water and Defined Media: Implications for Control of Microcystin-Producing Aquatic Blooms. Appl. Environ. Microbiol. 1997, 63, 2324–2329. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony Formation in the Cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howarth, R.W.; Marino, R. Nitrogen as the Limiting Nutrient for Eutrophication in Coastal Marine Ecosystems: Evolving Views over Three Decades. Limnol. Oceanogr. 2006, 51, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.J.; Lane, J.M.; Marino, R.; Howarth, R.W. Molybdenum Assimilation by Cyanobacteria and Phytoplankton in Freshwater and Salt Water. Limnol. Oceanogr. 1993, 38, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Marino, R.; Howarth, R.W.; Chan, F.; Cole, J.J.; Likens, G.E. Sulfate Inhibition of Molybdenum-Dependent Nitrogen Fixation by Planktonic Cyanobacteria under Seawater Conditions: A Non-Reversible Effect. Hydrobiologia 2003, 500, 277–293. [Google Scholar] [CrossRef]
- Long, B.M. Evidence That Sulfur Metabolism Plays a Role in Microcystin Production by Microcystis aeruginosa. Harmful Algae 2010, 9, 74–81. [Google Scholar] [CrossRef]
- Chen, L.; Gin, K.Y.H.; He, Y. Effects of Sulfate on Microcystin Production, Photosynthesis, and Oxidative Stress in Microcystis aeruginosa. Environ. Sci. Pollut. Res. 2016, 23, 3586–3595. [Google Scholar] [CrossRef]
- VanLandeghem, M.M.; Farooqi, M.; Southard, G.M.; Patiño, R. Spatiotemporal Associations of Reservoir Nutrient Characteristics and the Invasive, Harmful Alga Prymnesium parvum in West Texas. J. Am. Water Resour. Assoc. 2015, 51, 487–501. [Google Scholar] [CrossRef]
- Hambright, K.D.; Beyer, J.E.; Easton, J.D.; Zamor, R.M.; Easton, A.C.; Hallidayschult, T.C. The Niche of an Invasive Marine Microbe in a Subtropical Freshwater Impoundment. ISME J. 2015, 9, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Patiño, R.; Dawson, D.; VanLandeghem, M.M. Retrospective Analysis of Associations between Water Quality and Toxic Blooms of Golden Alga (Prymnesium parvum) in Texas Reservoirs: Implications for Understanding Dispersal Mechanisms and Impacts of Climate Change. Harmful Algae 2014, 33, 1–11. [Google Scholar] [CrossRef]
- Tábora-Sarmiento, S.; Patiño, R.; Portillo-Quintero, C.; Coldren, C. Air, Land, and Water Variables Associated with the First Appearance and Current Spatial Distribution of Toxic Prymnesium parvum Blooms in Reservoirs of the Southern Great Plains, USA. Sci. Total Environ. 2022, 836, 155567. [Google Scholar] [CrossRef] [PubMed]
- Hartman, K.J.; Wellman, D.I.; Kingsbury, J.W.; Cincotta, D.A.; Clayton, J.L.; Eliason, K.M.; Jernejcic, F.A.; Owens, N.V.; Smith, D.M. A Case Study of a Prymnesium parvum Harmful Algae Bloom in the Ohio River Drainage: Impact, Recovery and Potential for Future Invasions/Range Expansion. Water 2021, 13, 3233. [Google Scholar] [CrossRef]
- Johnsen, T.M.; Eikrem, W.; Olseng, C.D.; Tollefsen, K.E.; Bjerknes, V. Prymnesium parvum: The Norwegian Experience. J. Am. Water Resour. Assoc. 2010, 46, 6–13. [Google Scholar] [CrossRef]
- Guo, M.; Harrison, P.J.; Taylor, F.J.R. Fish Kills Related to Primnesium parvum N. Carter (Haptophyta) in the People’s Republic of China. J. Appl. Phycol. 1996, 8, 111–117. [Google Scholar] [CrossRef]
- Lemley, D.A.; Adams, J.B.; Rishworth, G.M.; Bouland, C. Phytoplankton Responses to Adaptive Management Interventions in Eutrophic Urban Estuaries. Sci. Total Environ. 2019, 693, 133601. [Google Scholar] [CrossRef]
- Rashel, R.H.; Patiño, R. Growth Response of the Ichthyotoxic Haptophyte, Prymnesium parvum Carter, to Changes in Sulfate and Fluoride Concentrations. PLoS ONE 2019, 14, e0223266. [Google Scholar] [CrossRef]
- VanLandeghem, M.M.; Farooqi, M.; Southard, G.M.; Patiño, R. Associations between Water Physicochemistry and Prymnesium parvum Presence, Abundance, and Toxicity in West Texas Reservoirs. J. Am. Water Resour. Assoc. 2015, 51, 471–486. [Google Scholar] [CrossRef]
- Ohio Environmental Protection Agency. Seeing Red: Emerging Harmful Algal Blooms; Spigot News; Ohio Environmental Protection Agency: Columbus, OH, USA, 2013; Volume 13. [Google Scholar]
- Paerl, H.W.; Otten, T.G.; Kudela, R. Mitigating the Expansion of Harmful Algal Blooms across the Freshwater-to-Marine Continuum. Environ. Sci. Technol. 2018, 52, 5519–5529. [Google Scholar] [CrossRef]
- Schindler, D.W.; Carpenter, S.R.; Chapra, S.C.; Hecky, R.E.; Orihel, D.M. Reducing Phosphorus to Curb Lake Eutrophication Is a Success. Environ. Sci. Technol. 2016, 50, 8923–8929. [Google Scholar] [CrossRef] [Green Version]
- Taranu, Z.E.; Gregory-Eaves, I.; Leavitt, P.R.; Bunting, L.; Buchaca, T.; Catalan, J.; Domaizon, I.; Guilizzoni, P.; Lami, A.; Mcgowan, S.; et al. Acceleration of Cyanobacterial Dominance in North Temperate-Subarctic Lakes during the Anthropocene. Ecol. Lett. 2015, 18, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Heathcote, A.J.; Taranu, Z.E.; Tromas, N.; MacIntyre-Newell, M.; Leavitt, P.R.; Pick, F.R. Sedimentary DNA and Pigments Show Increasing Abundance and Toxicity of CyanoHABs during the Anthropocene. Freshw. Biol. 2023, 1–16. [Google Scholar] [CrossRef]
- Glibert, P.M. Harmful Algae at the Complex Nexus of Eutrophication and Climate Change. Harmful Algae 2020, 91, 101583. [Google Scholar] [CrossRef] [PubMed]
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, Eutrophication and Harmful Algal Blooms along the Freshwater to Marine Continuum. Wiley Interdiscip. Rev. Water 2019, 6, e1373. [Google Scholar] [CrossRef]
- Doubek, J.P.; Carey, C.C.; Cardinale, B.J. Anthropogenic Land Use Is Associated with N-Fixing Cyanobacterial Dominance in Lakes across the Continental United States. Aquat. Sci. 2015, 77, 681–694. [Google Scholar] [CrossRef]
- Marion, J.W.; Zhang, F.; Cutting, D.; Lee, J. Associations between County-Level Land Cover Classes and Cyanobacteria Blooms in the United States. Ecol. Eng. 2017, 108, 556–563. [Google Scholar] [CrossRef]
- Beaver, J.R.; Manis, E.E.; Loftin, K.A.; Graham, J.L.; Pollard, A.I.; Mitchell, R.M. Land Use Patterns, Ecoregion, and Microcystin Relationships in U.S. Lakes and Reservoirs: A Preliminary Evaluation. Harmful Algae 2014, 36, 57–62. [Google Scholar] [CrossRef]
- Iiames, J.S.; Salls, W.B.; Mehaffey, M.H.; Nash, M.S.; Christensen, J.R.; Schaeffer, B.A. Modeling Anthropogenic and Environmental Influences on Freshwater Harmful Algal Bloom Development Detected by MERIS over the Central United States. Water Resour. Res. 2021, 57, e2020WR028946. [Google Scholar] [CrossRef]
- Rose, V.J.; Forney, W.M.; Norton, R.A.; Harrison, J.A. Catchment Characteristics, Water Quality, and Cyanobacterial Blooms in Washington and Oregon Lakes. Lake Reserv. Manag. 2019, 35, 51–63. [Google Scholar] [CrossRef]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread Global Increase in Intense Lake Phytoplankton Blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef]
- Brêda-Alves, F.; de Oliveira Fernandes, V.; Chia, M.A. Understanding the Environmental Roles of Herbicides on Cyanobacteria, Cyanotoxins, and CyanoHABs. Aquat. Ecol. 2021, 55, 347–361. [Google Scholar] [CrossRef]
- Harris, T.D.; Smith, V.H. Do Persistent Organic Pollutants Stimulate Cyanobacterial Blooms? INL Waters 2016, 6, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Benbrook, C.M. Trends in Glyphosate Herbicide Use in the United States and Globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hébert, M.P.; Fugère, V.; Gonzalez, A. The Overlooked Impact of Rising Glyphosate Use on Phosphorus Loading in Agricultural Watersheds. Front. Ecol. Environ. 2019, 17, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Forlani, G.; Pavan, M.; Gramek, M.; Kafarski, P.; Lipok, J. Biochemical Bases for a Widespread Tolerance of Cyanobacteria to the Phosphonate Herbicide Glyphosate. Plant Cell Physiol. 2008, 49, 443–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Zhang, Z.; Chen, Y.; Zhang, Q.; Ke, M.; Lu, T.; Qian, H. The Mechanism of Different Cyanobacterial Responses to Glyphosate. J. Environ. Sci. 2023, 125, 258–265. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Zhang, J.; Gao, B. Proteomic Mechanisms for the Combined Stimulatory Effects of Glyphosate and Antibiotic Contaminants on Microcystis aeruginosa. Chemosphere 2021, 267, 129244. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, H.; Li, Z.; Zhu, J.; Zhou, C.; Zhao, M. Effects of Glyphosate at Environmentally Relevant Concentrations on the Growth of and Microcystin Production by Microcystis aeruginosa. Environ. Monit. Assess. 2016, 188, 632. [Google Scholar] [CrossRef]
- Dabney, B.L.; Patiño, R. Low-Dose Stimulation of Growth of the Harmful Alga, Prymnesium parvum, by Glyphosate and Glyphosate-Based Herbicides. Harmful Algae 2018, 80, 130–139. [Google Scholar] [CrossRef]
- Sydney, E.B.; Schafranski, K.; Barretti, B.R.V.; Sydney, A.C.N.; Zimmerman, J.F.D.A.; Cerri, M.L.; Mottin Demiate, I. Biomolecules from Extremophile Microalgae: From Genetics to Bioprocessing of a New Candidate for Large-Scale Production. Process Biochem. 2019, 87, 37–44. [Google Scholar] [CrossRef]
- Qu, M.; Wang, L.; Xu, Q.; An, J.; Mei, Y.; Liu, G. Influence of Glyphosate and Its Metabolite Aminomethylphosphonic Acid on Aquatic Plants in Different Ecological Niches. Ecotoxicol. Environ. Saf. 2022, 246, 114155. [Google Scholar] [CrossRef] [PubMed]
- Battaglin, W.A.; Meyer, M.T.; Kuivila, K.M.; Dietze, J.E. Glyphosate and Its Degradation Product AMPA Occur Frequently and Widely in U.S. Soils, Surface Water, Groundwater, and Precipitation. J. Am. Water Resour. Assoc. 2014, 50, 275–290. [Google Scholar] [CrossRef]
- Coupe, R.H.; Kalkhoff, S.J.; Capel, P.D.; Gregoire, C. Fate and Transport of Glyphosate and Aminomethylphosphonic Acid in Surface Waters of Agricultural Basins. Pest Manag. Sci. 2011, 68, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Mahler, B.J.; Van Metre, P.C.; Burley, T.E.; Loftin, K.A.; Meyer, M.T.; Nowell, L.H. Similarities and Differences in Occurrence and Temporal Fluctuations in Glyphosate and Atrazine in Small Midwestern Streams (USA) during the 2013 Growing Season. Sci. Total Environ. 2017, 579, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Medalie, L.; Baker, N.T.; Shoda, M.E.; Stone, W.W.; Meyer, M.T.; Stets, E.G.; Wilson, M. Influence of Land Use and Region on Glyphosate and Aminomethylphosphonic Acid in Streams in the USA. Sci. Total Environ. 2020, 707, 136008. [Google Scholar] [CrossRef]
- Beecraft, L.; Rooney, R. Bioconcentration of Glyphosate in Wetland Biofilms. Sci. Total Environ. 2021, 756, 143993. [Google Scholar] [CrossRef]
- Vymazal, J.; Březinová, T. The Use of Constructed Wetlands for Removal of Pesticides from Agricultural Runoff and Drainage: A Review. Environ. Int. 2015, 75, 11–20. [Google Scholar] [CrossRef]
- Davidson, N.C. How Much Wetland Has the World Lost? Long-Term and Recent Trends in Global Wetland Area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Bullerjahn, G.S.; McKay, R.M.; Davis, T.W.; Baker, D.B.; Boyer, G.L.; D’Anglada, L.V.; Doucette, G.J.; Ho, J.C.; Irwin, E.G.; Kling, C.L.; et al. Global Solutions to Regional Problems: Collecting Global Expertise to Address the Problem of Harmful Cyanobacterial Blooms. A Lake Erie Case Study. Harmful Algae 2016, 54, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Mitsch, W.J. Solving Lake Erie’s Harmful Algal Blooms by Restoring the Great Black Swamp in Ohio. Ecol. Eng. 2017, 108, 406–413. [Google Scholar] [CrossRef]
- Mitsch, W.J. Restoring the Florida Everglades: Comments on the Current Reservoir Plan for Solving Harmful Algal Blooms and Restoring the Florida Everglades. Ecol. Eng. 2019, 3, 100009. [Google Scholar] [CrossRef]
- Tran, C.P.; Bode, R.W.; Smith, A.J.; Kleppel, G.S. Land-Use Proximity as a Basis for Assessing Stream Water Quality in New York State (USA). Ecol. Indic. 2010, 10, 727–733. [Google Scholar] [CrossRef]
- Sommer, U.; Adrian, R.; De Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lürling, M.; Molinero, J.C.; Mooij, W.M.; et al. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Shan, K.; Song, L.; Chen, W.; Li, L.; Liu, L.; Wu, Y.; Jia, Y.; Zhou, Q.; Peng, L. Analysis of Environmental Drivers Influencing Interspecific Variations and Associations among Bloom-Forming Cyanobacteria in Large, Shallow Eutrophic Lakes. Harmful Algae 2019, 84, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, L.; Zheng, L.; Dai, G.; Ma, H.; Shan, K.; Wu, H.; Zhou, Q.; Song, L. Patterns of Succession between Bloom-Forming Cyanobacteria Aphanizomenon flos-aquae and Microcystis and Related Environmental Factors in Large, Shallow Dianchi Lake, China. Hydrobiologia 2016, 765, 1–13. [Google Scholar] [CrossRef]
- Becker, V.; Caputo, L.; Ordóñez, J.; Marcé, R.; Armengol, J.; Crossetti, L.O.; Huszar, V.L.M. Driving Factors of the Phytoplankton Functional Groups in a Deep Mediterranean Reservoir. Water Res. 2010, 44, 3345–3354. [Google Scholar] [CrossRef]
- Christensen, V.G.; Maki, R.P.; Stelzer, E.A.; Norland, J.E.; Khan, E. Phytoplankton Community and Algal Toxicity at a Recurring Bloom in Sullivan Bay, Kabetogama Lake, Minnesota, USA. Sci. Rep. 2019, 9, 16129. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial Blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nakahara, H. The Formation and Degradation of Cyanobacterium Aphanizomenon flos-aquae Blooms: The Importance of pH, Water Temperature, and Day Length. Limnology 2005, 6, 1–6. [Google Scholar] [CrossRef]
- Zepernick, B.N.; Gann, E.R.; Martin, R.M.; Pound, H.L.; Krausfeldt, L.E.; Chaffin, J.D.; Wilhelm, S.W. Elevated pH Conditions Associated with Microcystis Spp. Blooms Decrease Viability of the Cultured Diatom Fragilaria crotonensis and Natural Diatoms in Lake Erie. Front. Microbiol. 2021, 12, 598736. [Google Scholar] [CrossRef]
- Graham, J.L.; Jones, J.R.; Jones, S.B.; Clevenger, T.E. Spatial and Temporal Dynamics of Microcystin in a Missouri Reservoir. Lake Reserv. Manag. 2006, 22, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.R.; Argerich, A.; Obrecht, D.V.; Thorpe, A.P.; North, R.L. Missouri Lakes and Reservoirs Long-Term Limnological Dataset Ver 1. Environmental Data Initiative. Available online: https://portal.edirepository.org/nis/mapbrowse?packageid=edi.520.1 (accessed on 10 July 2023).
- Krztoń, W.; Kosiba, J. Variations in Zooplankton Functional Groups Density in Freshwater Ecosystems Exposed to Cyanobacterial Blooms. Sci. Total Environ. 2020, 730, 139044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Duan, H.; Shi, X.; Yu, Y.; Kong, F. Contributions of Meteorology to the Phenology of Cyanobacterial Blooms: Implications for Future Climate Change. Water Res. 2012, 46, 442–452. [Google Scholar] [CrossRef]
- Reinl, K.L.; Harris, T.D.; North, R.L.; Almela, P.; Berger, S.A.; Bizic, M.; Burnet, S.H.; Grossart, H.P.; Ibelings, B.W.; Jakobsson, E.; et al. Blooms Also like It Cold. Limnol. Oceanogr. Lett. 2023, 8, 546–564. [Google Scholar] [CrossRef]
- Brutemark, A.; Granéli, E. Role of Mixotrophy and Light for Growth and Survival of the Toxic Haptophyte Prymnesium parvum. Harmful Algae 2011, 10, 388–394. [Google Scholar] [CrossRef]
- Baker, J.W.; Grover, J.P.; Brooks, B.W.; Ureña-Boeck, F.; Roelke, D.L.; Errera, R.; Kiesling, R.L. Growth and Toxicity of Prymnesium parvum (Haptophyta) as a Function of Salinity, Light, and Temperature. J. Phycol. 2007, 43, 219–227. [Google Scholar] [CrossRef]
- VanLandeghem, M.M.; Meyer, M.D.; Cox, S.B.; Sharma, B.; Patiño, R. Spatial and Temporal Patterns of Surface Water Quality and Ichthyotoxicity in Urban and Rural River Basins in Texas. Water Res. 2012, 46, 6638–6651. [Google Scholar] [CrossRef]
- Roelke, D.L.; Brooks, B.W.; Grover, J.P.; Gable, G.M.; Schwierzke-Wade, L.; Hewitt, N.C. Anticipated Human Population and Climate Change Effects on Algal Blooms of a Toxic Haptophyte in the South-Central USA. Can. J. Fish. Aquat. Sci. 2012, 69, 1389–1404. [Google Scholar] [CrossRef]
- VanLandeghem, M.M.; Farooqi, M.; Farquhar, B.; Patiño, R. Impacts of Golden Alga Prymnesium parvum on Fish Populations in Reservoirs of the Upper Colorado River and Brazos River Basins, Texas. Trans. Am. Fish. Soc. 2013, 142, 581–595. [Google Scholar] [CrossRef]
- Dawson, D.; VanLandeghem, M.M.; Asquith, W.H.; Patiño, R. Long-Term Trends in Reservoir Water Quality and Quantity in Two Major River Basins of the Southern Great Plains. Lake Reserv. Manag. 2015, 31, 254–279. [Google Scholar]
- Rashel, R.H.; Patiño, R. Influence of Genetic Background, Salinity, and Inoculum Size on Growth of the Ichthyotoxic Golden Alga (Prymnesium parvum). Harmful Algae 2017, 66, 97–104. [Google Scholar] [CrossRef]
- Rashel, R.H. Influence of Water Quality and Climate Variables on Growth of the Harmful Alga, Prymnesium parvum. Ph.D. Thesis, Texas Tech University, Lubbock, TX, USA, 2020. Available online: https://hdl.handle.net/2346/85858 (accessed on 1 March 2023).
- Danilov, R.A.; Ekelund, N.G.A. Effects of pH on the Growth Rate, Motility and Photosynthesis in Euglena gracilis. Folia Microbiol. 2001, 46, 549–554. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Y.; Yang, T.; He, Z.; Yan, Q. Size-Fractioned Aggregates within Phycosphere Define Functional Bacterial Communities Related to Microcystis aeruginosa and Euglena sanguinea Blooms. Aquat. Ecol. 2020, 54, 609–623. [Google Scholar] [CrossRef]
- Rahman, M.S.; Shahjahan, M.; Haque, M.M.; Khan, S. Control of Euglenophyte Bloom and Fish Production Enhancement Using Duckweed and Lime. Iran. J. Fish. Sci. 2012, 11, 602–617. [Google Scholar]
- Chernoff, N.; Hill, D.J.; Diggs, D.L.; Faison, B.D.; Francis, B.M.; Lang, J.R.; Larue, M.M.; Le, T.T.; Loftin, K.A.; Lugo, J.N.; et al. A Critical Review of the Postulated Role of the Non-Essential Amino Acid, β-N-Methylamino-L-Alanine, in Neurodegenerative Disease in Humans. J. Toxicol. Environ. Health Part B Crit. Rev. 2017, 20, 183–229. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, M.; Płotka-Wasylka, J.; Majchrzak, T.; Wojnowski, W.; Mazur-Marzec, H.; Namieśnik, J. Recent Trends in Determination of Neurotoxins in Aquatic Environmental Samples. TrAC Trends Anal. Chem. 2019, 112, 112–122. [Google Scholar] [CrossRef]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin Mixtures and Taste-and-Odor Compounds in Cyanobacterial Blooms from the Midwestern United States. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Beaver, J.R.; Tausz, C.E.; Scotese, K.C.; Pollard, A.I.; Mitchell, R.M. Environmental Factors Influencing the Quantitative Distribution of Microcystin and Common Potentially Toxigenic Cyanobacteria in U.S. Lakes and Reservoirs. Harmful Algae 2018, 78, 118–128. [Google Scholar] [CrossRef]
- Kinnear, S. Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research. Mar. Drugs 2010, 8, 542–564. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.R.; Beversdorf, L.J.; Weirich, C.A.; Bartlett, S.L. Cyanobacterial Toxins of the Laurentian Great Lakes, Their Toxicological Effects, and Numerical Limits in Drinking Water. Mar. Drugs 2017, 15, 160. [Google Scholar] [CrossRef] [Green Version]
- Fiore, M.F.; de Lima, S.T.; Carmichael, W.W.; McKinnie, S.M.K.; Chekan, J.R.; Moore, B.S. Guanitoxin, Re-Naming a Cyanobacterial Organophosphate Toxin. Harmful Algae 2020, 92, 101737. [Google Scholar] [CrossRef]
- Christensen, V.G.; Khan, E. Freshwater Neurotoxins and Concerns for Human, Animal, and Ecosystem Health: A Review of Anatoxin-a and Saxitoxin. Sci. Total Environ. 2020, 736, 139515. [Google Scholar] [CrossRef] [PubMed]
- D’Anglada, L.V.; Hilborn, E.D.; Backer, L.C. Harmful Algal Blooms (HABs) and Public Health: Progress and Current Challenges. In Toxins; MDPI: Basel, Switzerland, 2016; ISBN 9783038421559. [Google Scholar]
- Kotak, B.G.; Zurawell, R.W. Cyanobacterial Toxins in Canadian Freshwaters: A Review. Lake Reserv. Manag. 2007, 23, 109–122. [Google Scholar] [CrossRef]
- Al-Sammak, M.A.; Hoagland, K.D.; Cassada, D.; Snow, D.D. Co-Occurrence of the Cyanotoxins BMAA, DABA and Anatoxin-a in Nebraska Reservoirs, Fish, and Aquatic Plants. Toxins 2014, 6, 488–508. [Google Scholar] [CrossRef] [Green Version]
- John, N.; Baker, L.; Ansell, B.R.E.; Newham, S.; Crosbie, N.D.; Jex, A.R. First Report of Anatoxin-a Producing Cyanobacteria in Australia Illustrates Need to Regularly up-Date Monitoring Strategies in a Shifting Global Distribution. Sci. Rep. 2019, 9, 10894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, T.G.; Wood, S.A.; Hawes, I. The Rise of Toxic Benthic Phormidium Proliferations: A Review of Their Taxonomy, Distribution, Toxin Content and Factors Regulating Prevalence and Increased Severity. Harmful Algae 2016, 55, 282–294. [Google Scholar] [CrossRef]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean Warming since 1982 Has Expanded the Niche of Toxic Algal Blooms in the North Atlantic and North Pacific Oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef]
- Christensen, V.G.; Stelzer, E.A.; Eikenberry, B.C.; Olds, H.T.; LeDuc, J.F.; Maki, R.P.; Saley, A.M.; Norland, J.E.; Khan, E. Cyanotoxin Mixture Models: Relating Environmental Variables and Toxin Co-Occurrence to Human Exposure Risk. J. Hazard. Mater. 2021, 415, 125560. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Pflugmacher, S.; James, K.J.; Furey, A. Anatoxin-a Elicits an Increase in Peroxidase and Glutathione S-Transferase Activity in Aquatic Plants. Aquat. Toxicol. 2004, 68, 185–192. [Google Scholar] [CrossRef]
- Zhang, Y.; Vo Duy, S.; Munoz, G.; Sauvé, S. Phytotoxic Effects of Microcystins, Anatoxin-a and Cylindrospermopsin to Aquatic Plants: A Meta-Analysis. Sci. Total Environ. 2022, 810, 152104. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Toporowska, M.; Rechulicz, J. Simultaneous Accumulation of Anatoxin-a and Microcystins in Three Fish Species Indigenous to Lakes Affected by Cyanobacterial Blooms. Oceanol. Hydrobiol. Stud. 2012, 41, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Scarlett, K.R.; Kim, S.; Lovin, L.M.; Chatterjee, S.; Scott, J.T.; Brooks, B.W. Global Scanning of Cylindrospermopsin: Critical Review and Analysis of Aquatic Occurrence, Bioaccumulation, Toxicity and Health Hazards. Sci. Total Environ. 2020, 738, 139807. [Google Scholar] [CrossRef] [PubMed]
- De La Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.A.; O’Shea, K.; Zhao, C.; et al. A Review on Cylindrospermopsin: The Global Occurrence, Detection, Toxicity and Degradation of a Potent Cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef] [PubMed]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Méjean, A.; Ploux, O. Biosynthesis of Cylindrospermopsin and 7-Epicylindrospermopsin in Oscillatoria Sp. Strain PCC 6506: Identification of the Cyr Gene Cluster and Toxin Analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar] [CrossRef] [Green Version]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of Cylindrospermopsin, the Toxin from the Cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, Temperature, and Sunlight on Decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Aguilera, A.; Haakonsson, S.; Martin, M.V.; Salerno, G.L.; Echenique, R.O. Bloom-Forming Cyanobacteria and Cyanotoxins in Argentina: A Growing Health and Environmental Concern. Limnologica 2018, 69, 103–114. [Google Scholar] [CrossRef]
- Seifert, M.; McGregor, G.; Eaglesham, G.; Wickramasinghe, W.; Shaw, G. First Evidence for the Production of Cylindrospermopsin and Deoxy-Cylindrospermopsin by the Freshwater Benthic Cyanobacterium, Lyngbya wollei (Farlow Ex Gomont) Speziale and Dyck. Harmful Algae 2007, 6, 73–80. [Google Scholar] [CrossRef]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the Distribution, Phylogeography, and Ecophysiology of a Global Invasive Species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef] [Green Version]
- Sundaravadivelu, D.; Sanan, T.T.; Venkatapathy, R.; Mash, H.; Tettenhorst, D.; Danglada, L.; Frey, S.; Tatters, A.O.; Lazorchak, J. Determination of Cyanotoxins and Prymnesins in Water, Fish Tissue, and Other Matrices: A Review. Toxins 2022, 14, 213. [Google Scholar] [CrossRef]
- White, S.H.; Duivenvoorden, L.J.; Fabbro, L.D.; Eaglesham, G.K. Influence of Intracellular Toxin Concentrations on Cylindrospermopsin Bioaccumulation in a Freshwater Gastropod (Melanoides Tuberculata). Toxicon 2006, 47, 497–509. [Google Scholar] [CrossRef]
- Cabang, A.B.; De Mukhopadhyay, K.; Meyers, S.; Morris, J.; Zimba, P.V.; Wargovich, M.J. Therapeutic Effects of the Euglenoid Ichthyotoxin, Euglenophycin, in Colon Cancer. Oncotarget 2017, 8, 104347–104358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, D.B.; Rafalski, A.; Beauchesne, K.; Moeller, P.D.; Triemer, R.E.; Zimba, P.V. Quantitative Mass Spectrometric Analysis and Post-Extraction Stability Assessment of the Euglenoid Toxin Euglenophycin. Toxins 2013, 5, 1587–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulczycka, A.; Łukomska-Kowalczyk, M.; Zakryś, B.; Milanowski, R. PCR Identification of Toxic Euglenid Species Euglena sanguinea. J. Appl. Phycol. 2018, 30, 1759–1763. [Google Scholar] [CrossRef] [Green Version]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and Toxicity of Microcystin Congeners Other than MC-LR and MC-RR: A Review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of Microcystins with Reference to Cases of Human Intoxications and Epidemiological Investigations of Exposures to Cyanobacteria and Cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- Chernoff, N.; Hill, D.; Lang, J.; Schmid, J.; Le, T.; Farthing, A.; Huang, H. The Comparative Toxicity of 10 Microcystin Congeners Administered Orally to Mice: Clinical Effects and Organ Toxicity. Toxins 2020, 12, 403. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; Chorus, I., Bartram, J., Eds.; World Health Organization: London, UK, 1999; pp. 41–112. [Google Scholar]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver Tumor Promotion by the Cyanobacterial Cyclic Peptide Toxin Microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef]
- Dyble, J.; Fahnenstiel, G.L.; Litaker, R.W.; Millie, D.F.; Tester, P.A. Microcystin Concentrations and Genetic Diversity of Microcystis in the Lower Great Lakes. Environ. Toxicol. 2008, 23, 507–516. [Google Scholar] [CrossRef]
- Hardy, F.J.; Johnson, A.; Hamel, K.; Preece, E. Cyanotoxin Bioaccumulation in Freshwater Fish, Washington State, USA. Environ. Monit. Assess. 2015, 187, 667. [Google Scholar] [CrossRef]
- Preece, E.P.; Hardy, F.J.; Moore, B.C.; Bryan, M. A Review of Microcystin Detections in Estuarine and Marine Waters: Environmental Implications and Human Health Risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Woller-Skar, M.M.; Russell, A.L.; Gaskill, J.A.; Luttenton, M.R. Microcystin in Multiple Life Stages of Hexagenia limbata, with Implications for Toxin Transfer. J. Great Lakes Res. 2020, 46, 666–671. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial Toxins: Modes of Actions, Fate in Aquatic and Soil Ecosystems, Phytotoxicity and Bioaccumulation in Agricultural Crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef]
- Manning, S.R.; La Claire, J.W. Prymnesins: Toxic Metabolites of the Golden Alga, Prymnesium parvum Carter (Haptophyta). Mar. Drugs 2010, 8, 678–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindholm, T.; Öhman, P.; Kurki-Helasmo, K.; Kincaid, B.; Meriluoto, J. Toxic Algae and Fish Mortality in a Brackish-Water Lake in Aland, SW Finland. Hydrobiologia 1999, 397, 109–120. [Google Scholar] [CrossRef]
- Rasmussen, S.A.; Meier, S.; Andersen, N.G.; Blossom, H.E.; Duus, J.Ø.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Chemodiversity of Ladder-Frame Prymnesin Polyethers in Prymnesium parvum. J. Nat. Prod. 2016, 79, 2250–2256. [Google Scholar] [CrossRef] [Green Version]
- Valenti, T.W.; James, S.V.; Lahousse, M.; Schug, K.A.; Roelke, D.L.; Grover, J.P.; Brooks, B.W. Influence of pH on Amine Toxicology and Implications for Harmful Algal Bloom Ecology. Toxicon 2010, 55, 1038–1043. [Google Scholar] [CrossRef]
- O’Neill, K.; Musgrave, I.F.; Humpage, A.R. Low Dose Extended Exposure to Saxitoxin and Its Potential Neurodevelopmental Effects: A Review. Environ. Toxicol. Pharmacol. 2016, 48, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Bernard, C.; Fastner, J. Saxitoxin and Analogues. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley and Sons, Ltd.: Chichester, UK, 2017; pp. 148–154. [Google Scholar]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic Alkaloids: Saxitoxin and Its Analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [Green Version]
- Onodera, H.; Satake, M.; Oshima, Y.; Yasumoto, T.; Carmichael, W.W. New Saxitoxin Analogues from the Freshwater Filamentous Cyanobacterium Lyngbya wollei. Nat. Toxins 1997, 5, 146–151. [Google Scholar] [CrossRef]
- Casali, S.P.; Dos Santos, A.C.A.; De Falco, P.B.; Do Carmo Calijuri, M. Influence of Environmental Variables on Saxitoxin Yields by Cylindrospermopsis raciborskii in a Mesotrophic Subtropical Reservoir. J. Water Health 2017, 15, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Smith, Z.J.; Martin, R.M.; Wei, B.; Wilhelm, S.W.; Boyer, G.L. Spatial and Temporal Variation in Paralytic Shellfish Toxin Production by Benthic Microseira (Lyngbya) wollei in a Freshwater New York Lake. Toxins 2019, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, A.P.; Jones, G.; Blackburn, S.I.; Oshima, Y.; Onodera, H. Effect of Culture and Bloom Development and of Sample Storage on Paralytic Shellfish Poisons in the Cyanobacterium Anabaena circinalis. J. Phycol. 1997, 33, 26–35. [Google Scholar] [CrossRef]
- Belykh, O.I.; Tikhonova, I.V.; Kuzmin, A.V.; Sorokovikova, E.G.; Fedorova, G.A.; Khanaev, I.V.; Sherbakova, T.A.; Timoshkin, O.A. First Detection of Benthic Cyanobacteria in Lake Baikal Producing Paralytic Shellfish Toxins. Toxicon 2016, 121, 36–40. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Jones, C.L.A.; Mahmood, N.A.; Theiss, W.C. Algal Toxins and Water-Based Diseases. Crit. Rev. Environ. Control 1985, 15, 275–313. [Google Scholar] [CrossRef]
- Cusick, K.D.; Sayler, G.S. An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [Green Version]
- Negri, A.P.; Jones, G.J. Bioaccumulation of Paralytic Shellfish Poisoning (PSP) Toxins from the Cyanobacterium Anabaena circinalis by the Freshwater Mussel Alathyria condola. Toxicon 1995, 33, 667–678. [Google Scholar] [CrossRef]
- De Calado, S.L.M.; Santos, G.S.; Wojciechowski, J.; de Magalhães, V.F.; Silva de Assis, H.C. The Accumulation Dynamics, Elimination and Risk Assessment of Paralytic Shellfish Toxins in Fish from a Water Supply Reservoir. Sci. Total Environ. 2019, 651, 3222–3229. [Google Scholar] [CrossRef]
- Giovannardi, S.; Pollegioni, L.; Pomati, F.; Rossetti, C.; Sacchi, S.; Sessa, L.; Calamari, D. Toxic Cyanobacterial Blooms in Lake Varese (Italy): A Multidisciplinary Approach. Environ. Toxicol. 1999, 14, 127–134. [Google Scholar] [CrossRef]
- Lenz, K.A.; Miller, T.R.; Ma, H. Anabaenopeptins and Cyanopeptolins Induce Systemic Toxicity Effects in a Model Organism the Nematode Caenorhabditis elegans. Chemosphere 2019, 214, 60–69. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse Taxa of Cyanobacteria Produce Beta-N-Methylamino-L-Alanine, a Neurotoxic Amino Acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef]
- Faassen, E.J.; Harkema, L.; Begeman, L.; Lurling, M. First Report of (Homo)Anatoxin-a and Dog Neurotoxicosis after Ingestion of Benthic Cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.A. Analytical Techniques for the Detection of α-Amino-β-Methylaminopropionic Acid. Analyst 2012, 137, 1991–2005. [Google Scholar] [CrossRef]
- Monteiro, P.R.; Do Amaral, S.C.; Siqueira, A.S.; Xavier, L.P.; Santos, A.V. Anabaenopeptins: What We Know so Far. Toxins 2021, 13, 522. [Google Scholar] [CrossRef] [PubMed]
- Beversdorf, L.J.; Rude, K.; Weirich, C.A.; Bartlett, S.L.; Seaman, M.; Kozik, C.; Biese, P.; Gosz, T.; Suha, M.; Stempa, C.; et al. Analysis of Cyanobacterial Metabolites in Surface and Raw Drinking Waters Reveals More than Microcystin. Water Res. 2018, 140, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Fastner, J.; Dittmann, E.; Christiansen, G.; Vasconcelos, V.M. Variation between Strains of the Cyanobacterium Microcystis aeruginosa Isolated from a Portuguese River. J. Appl. Microbiol. 2005, 99, 749–757. [Google Scholar] [CrossRef]
- Cerasino, L.; Capelli, C.; Salmaso, N. A Comparative Study of the Metabolic Profiles of Common Nuisance Cyanobacteria in Southern Perialpine Lakes. Adv. Oceanogr. Limnol. 2017, 8, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Marzec, H.; Fidor, A.; Cegłowska, M.; Wieczerzak, E.; Kropidłowska, M.; Goua, M.; Macaskill, J.; Edwards, C. Cyanopeptolins with Trypsin and Chymotrypsin Inhibitory Activity from the Cyanobacterium Nostoc edaphicum CCNP1411. Mar. Drugs 2018, 16, 220. [Google Scholar] [CrossRef] [Green Version]
- Janssen, E.M.-L. Cyanobacterial Peptides beyond Microcystins—A Review on Co-Occurrence, Toxicity, and Challenges for Risk Assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Weirich, C.A.; Miller, T.R. Freshwater Harmful Algal Blooms: Toxins and Children’s Health. Curr. Probl. Pediatr. Adolesc. Health Care 2014, 44, 2–24. [Google Scholar] [CrossRef]
- Funari, E.; Testai, E. Human Health Risk Assessment Related to Cyanotoxins Exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef]
- Faltermann, S.; Zucchi, S.; Kohler, E.; Blom, J.F.; Pernthaler, J.; Fent, K. Molecular Effects of the Cyanobacterial Toxin Cyanopeptolin (CP1020) Occurring in Algal Blooms: Global Transcriptome Analysis in Zebrafish Embryos. Aquat. Toxicol. 2014, 149, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhao, Y. Reduced Zebrafish Transcriptome Atlas toward Understanding Environmental Neurotoxicants. Environ. Sci. Technol. 2018, 52, 7120–7130. [Google Scholar] [CrossRef] [PubMed]
- Christensen, V.G.; Olds, H.T.; Norland, J.; Khan, E. Phytoplankton Community Interactions and Cyanotoxin Mixtures in Three Recurring Surface Blooms within One Lake. J. Hazard. Mater. 2022, 427, 128142. [Google Scholar] [CrossRef]
- Howard, M.D.A.; Kudela, R.M.; Hayashi, K.; Tatters, A.O.; Caron, D.A.; Theroux, S.; Oehrle, S.; Roethler, M.; Donovan, A.; Loftin, K.; et al. Multiple Co-Occurring and Persistently Detected Cyanotoxins and Associated Cyanobacteria in Adjacent California Lakes. Toxicon 2021, 192, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Smith, Z.J.; Conroe, D.E.; Schulz, K.L.; Boyer, G.L. Limnological Differences in a Two-Basin Lake Help to Explain the Occurrence of Anatoxin-a, Paralytic Shellfish Poisoning Toxins, and Microcystins. Toxins 2020, 12, 559. [Google Scholar] [CrossRef] [PubMed]
- Beversdorf, L.J.; Weirich, C.; Bartlett, S.; Miller, T. Variable Cyanobacterial Toxin and Metabolite Profiles across Six Eutrophic Lakes of Differing Physiochemical Characteristics. Toxins 2017, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, D.R.; Fischer, A.; Michel, C.; Hoeger, S. Toxin Mixture in Cyanobacterial Blooms—A Critical Comparison of Reality with Current Procedures Employed in Human Health Risk Assessment. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: New York, NY, USA, 2008; pp. 885–912. [Google Scholar]
- Burford, M.A.; Carey, C.C.; Hamilton, D.P.; Huisman, J.; Paerl, H.W.; Wood, S.A.; Wulff, A. Perspective: Advancing the Research Agenda for Improving Understanding of Cyanobacteria in a Future of Global Change. Harmful Algae 2020, 91, 101601. [Google Scholar] [CrossRef]
- Paerl, H.W. Impacts of Climate Change on Cyanobacteria in Aquatic Environments. In Climate Change and Microbial Ecology: Current Research and Future Trends; Marxsen, J., Ed.; Caister Academic Press: Wymondham, UK, 2016. [Google Scholar]
- Chapra, S.C.; Boehlert, B.; Fant, C.; Bierman, V.J.; Henderson, J.; Mills, D.; Mas, D.M.L.; Rennels, L.; Jantarasami, L.; Martinich, J.; et al. Climate Change Impacts on Harmful Algal Blooms in U. S. Freshwaters: A Screening-Level Assessment. Environ. Sci. Technol. 2017, 51, 8933–8943. [Google Scholar] [CrossRef] [Green Version]
- Brandenburg, K.M.; de Senerpont Domis, L.N.; Wohlrab, S.; Krock, B.; John, U.; van Scheppingen, Y.; van Donk, E.; Van de Waal, D.B. Combined Physical, Chemical and Biological Factors Shape Alexandrium ostenfeldii Blooms in the Netherlands. Harmful Algae 2017, 63, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Kolman, M.A.; Torres, L.L.; Martin, M.L.; Salerno, G.L. Sucrose Synthase in Unicellular Cyanobacteria and Its Relationship with Salt and Hypoxic Stress. Planta 2012, 235, 955–964. [Google Scholar] [CrossRef]
- Sandrini, G.; Huisman, J.; Matthijs, H.C.P. Potassium Sensitivity Differs among Strains of the Harmful Cyanobacterium Microcystis and Correlates with the Presence of Salt Tolerance Genes. FEMS Microbiol. Lett. 2015, 362, fnv121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, Y.; Yamaguchi, H.; Sano, T.; Kawachi, M. A Novel Salt-Tolerant Genotype Illuminates the Sucrose Gene Evolution in Freshwater Bloom-Forming Cyanobacterium Microcystis aeruginosa. FEMS Microbiol. Lett. 2019, 366, fnz190. [Google Scholar] [CrossRef] [PubMed]
- Tonk, L.; Bosch, K.; Visser, P.M.; Huisman, J. Salt Tolerance of the Harmful Cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2007, 46, 117–123. [Google Scholar] [CrossRef]
- Des Aulnois, M.G.; Roux, P.; Caruana, A.; Réveillon, D.; Briand, E.; Hervé, F.; Savar, V.; Bormans, M.; Amzil, Z. Physiological and Metabolic Responses of Freshwater and Brackish-Water Strains of Microcystis aeruginosa Acclimated to a Salinity Gradient: Insight into Salt Tolerance. Appl. Environ. Microbiol. 2019, 85, e01614-19. [Google Scholar] [CrossRef]
- Rosen, B.H.; Loftin, K.A.; Graham, J.L.; Stahlhut, K.N.; Riley, J.M.; Johnson, B.D.; Senegal, S. Understanding the Effect of Salinity Tolerance on Cyanobacteria Associated with a Harmful Algal Bloom in Lake Okeechobee, Florida; U.S. Geological Survey Scientific Investigations Report 2018-5092; US Geological Survey: Reston, VA, USA, 2018. [Google Scholar]
- Tatters, A.O.; Smith, J.; Kudela, R.M.; Hayashi, K.; Da, M.; Donovan, A.R.; Loftin, K.A.; Caron, D.A. The Tide Turns: Episodic and Localized Cross-Contamination of a California Coastline with Cyanotoxins. Harmful Algae 2021, 103, 102003. [Google Scholar] [CrossRef]
- Melero-Jiménez, I.J.; Martín-Clemente, E.; García-Sánchez, M.J.; Bañares-España, E.; Flores-Moya, A. The Limit of Resistance to Salinity in the Freshwater Cyanobacterium Microcystis aeruginosa Is Modulated by the Rate of Salinity Increase. Ecol. Evol. 2020, 10, 5045–5055. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, V.M.; Roelke, D.L.; Brooks, B.W.; Granéli, E.; Davis, S.L.; Baty, T.; Scott, W.C. Prymnesium parvum Invasion Success into Coastal Bays of the Gulf of Mexico: Galveston Bay Case Study. Harmful Algae 2015, 43, 31–45. [Google Scholar] [CrossRef]
- Richardson, E.T.; Patiño, R. Growth of the Harmful Alga, Prymnesium parvum (Prymnesiophyceae), after Gradual and Abrupt Increases in Salinity. J. Phycol. 2021, 57, 1335–1344. [Google Scholar] [CrossRef]
Figure 1.
Inland cyanobacterial blooms. (A) Aphanizomenon-dominated bloom, Kabetogama Lake, MN, USA. (B) Dolichospermum-dominated bloom, Timothy Lake, OR, USA. (C) Microcystis-dominated bloom, Milford Lake, KS, USA. (D) Planktothrix agardhii/prolifica-dominated bloom, Fleeing Horse Lake, AB, Canada. USGS, U.S. Geological Survey.
Figure 1.
Inland cyanobacterial blooms. (A) Aphanizomenon-dominated bloom, Kabetogama Lake, MN, USA. (B) Dolichospermum-dominated bloom, Timothy Lake, OR, USA. (C) Microcystis-dominated bloom, Milford Lake, KS, USA. (D) Planktothrix agardhii/prolifica-dominated bloom, Fleeing Horse Lake, AB, Canada. USGS, U.S. Geological Survey.

Figure 2.
Toxic bloom-forming cyanobacterial taxa in U.S. inland waters. (A) Anabaena spp. are filamentous, do not have aerotopes (gas vesicles), and are typically benthic. A nitrogen-fixing cell (heterocyte) is visible within the filament. (B) Aphanizomenon flos-aquae forms bundles of filaments (fascicles) and has aerotopes. (C) Dolichospermum lemmermannii is one of several coiled forms in this genus; the image shows akinetes on either side of single heterocytes. The arrangement, size, and location of these two cell types are used to distinguish species. (D) Oscillatoria spp. form filaments with short cells and are typically benthic. (E) Microcystis aeruginosa is a colonial form with cells embedded in a common mucilage that is not easily visible. The open spaces in the colonies (clathrate) are characteristic of this species. (F) Planktothrix spp. are filamentous, planktonic forms with abundant aerotopes. FGCU, Florida Gulf Coast University.
Figure 2.
Toxic bloom-forming cyanobacterial taxa in U.S. inland waters. (A) Anabaena spp. are filamentous, do not have aerotopes (gas vesicles), and are typically benthic. A nitrogen-fixing cell (heterocyte) is visible within the filament. (B) Aphanizomenon flos-aquae forms bundles of filaments (fascicles) and has aerotopes. (C) Dolichospermum lemmermannii is one of several coiled forms in this genus; the image shows akinetes on either side of single heterocytes. The arrangement, size, and location of these two cell types are used to distinguish species. (D) Oscillatoria spp. form filaments with short cells and are typically benthic. (E) Microcystis aeruginosa is a colonial form with cells embedded in a common mucilage that is not easily visible. The open spaces in the colonies (clathrate) are characteristic of this species. (F) Planktothrix spp. are filamentous, planktonic forms with abundant aerotopes. FGCU, Florida Gulf Coast University.

Figure 3.
Prymnesium parvum. (A) Single cell of Prymnesium parvum. Note the two golden-colored chloroplasts inside the cell and the haptonema (bottom left) flanked by two longer flagella. (B) Winter bloom of P. parvum in a Texas reservoir (USA). The lake water has taken on a golden-brown appearance and ice can be seen attached to emergent macrophytes. (C) Spring bloom of P. parvum in an impounded urban stream system in Lubbock, Texas. Fish in the affected areas appeared lethargic and unresponsive, and most died within a few days. TPWD, Texas Parks and Wildlife Department; USGS, U.S. Geological Survey.
Figure 3.
Prymnesium parvum. (A) Single cell of Prymnesium parvum. Note the two golden-colored chloroplasts inside the cell and the haptonema (bottom left) flanked by two longer flagella. (B) Winter bloom of P. parvum in a Texas reservoir (USA). The lake water has taken on a golden-brown appearance and ice can be seen attached to emergent macrophytes. (C) Spring bloom of P. parvum in an impounded urban stream system in Lubbock, Texas. Fish in the affected areas appeared lethargic and unresponsive, and most died within a few days. TPWD, Texas Parks and Wildlife Department; USGS, U.S. Geological Survey.

Figure 4.
Geographic distribution of Prymnesium parvum in the USA. With few exceptions, the majority of states where P. parvum has been found have also reported toxic blooms. There are a few states where P. parvum has been recorded in inland freshwater or in coastal environments (or both), but records of bloom occurrence are only available for inland brackish waters. The distance scale for the Hawaii (HI) inset is the same as for the main map.
Figure 4.
Geographic distribution of Prymnesium parvum in the USA. With few exceptions, the majority of states where P. parvum has been found have also reported toxic blooms. There are a few states where P. parvum has been recorded in inland freshwater or in coastal environments (or both), but records of bloom occurrence are only available for inland brackish waters. The distance scale for the Hawaii (HI) inset is the same as for the main map.

Figure 5.
Euglena sanguinea. (A) Single cell of Euglena sanguinea. Note the presence of red pigment in astaxanthin globules. The position of globules within the cell (peripheral or central) and the outward cell coloration (red or green) can be regulated by light. (B) Euglena bloom in Dillon Lake, OH, USA. Note the blood-red surface scum. FGCU, Florida Gulf Coast University; EPA, Environmental Protection Agency.
Figure 5.
Euglena sanguinea. (A) Single cell of Euglena sanguinea. Note the presence of red pigment in astaxanthin globules. The position of globules within the cell (peripheral or central) and the outward cell coloration (red or green) can be regulated by light. (B) Euglena bloom in Dillon Lake, OH, USA. Note the blood-red surface scum. FGCU, Florida Gulf Coast University; EPA, Environmental Protection Agency.

Figure 6.
Seasonal changes in microcystin concentrations. Microcystin concentration in eight reservoirs from Northwestern Missouri, USA, sampled weekly during 2004. Red circles indicate the onset and duration of thermal stratification during the summer in these midcontinent reservoirs. The figure was created using data available in Jones et al. [189].
Figure 6.
Seasonal changes in microcystin concentrations. Microcystin concentration in eight reservoirs from Northwestern Missouri, USA, sampled weekly during 2004. Red circles indicate the onset and duration of thermal stratification during the summer in these midcontinent reservoirs. The figure was created using data available in Jones et al. [189].

Figure 7.
Seasonal changes in Prymnesium parvum abundance and toxicity. Cell abundance and water ichthyotoxicity were measured from January 2010 to July 2011 in E.V. Spence Reservoir, Texas (USA). Colored lines indicate the beginning of the spring (spring equinox) and the fall (fall equinox). Ichthyotoxicity was determined using a semiquantitative fish bioassay. Cell abundance and toxicity increased in the fall and declined in the spring. Data from VanLandeghem et al. [144].
Figure 7.
Seasonal changes in Prymnesium parvum abundance and toxicity. Cell abundance and water ichthyotoxicity were measured from January 2010 to July 2011 in E.V. Spence Reservoir, Texas (USA). Colored lines indicate the beginning of the spring (spring equinox) and the fall (fall equinox). Ichthyotoxicity was determined using a semiquantitative fish bioassay. Cell abundance and toxicity increased in the fall and declined in the spring. Data from VanLandeghem et al. [144].

Figure 8.
Cyanotoxin distribution by lake salinity in the conterminous USA. Pie charts show the percent distribution of microcystins and cylindrospermopsin detections (>method detection limit, MDL) by lake salinity. The percent distribution of microcystins by salinity is also shown for values considered potentially harmful to people when swimming or participating in other activities in or on the water (>8 μg L−1; recreational criterion). No samples contained cylindrospermopsin levels above its recreational criterion (15 μg L−1). Data are from the 2017 National Lakes Assessment [30].
Figure 8.
Cyanotoxin distribution by lake salinity in the conterminous USA. Pie charts show the percent distribution of microcystins and cylindrospermopsin detections (>method detection limit, MDL) by lake salinity. The percent distribution of microcystins by salinity is also shown for values considered potentially harmful to people when swimming or participating in other activities in or on the water (>8 μg L−1; recreational criterion). No samples contained cylindrospermopsin levels above its recreational criterion (15 μg L−1). Data are from the 2017 National Lakes Assessment [30].

Figure 9.
Geographic distribution and concentrations of microcystins in the conterminous USA. The map shows hydrologic regions and distribution and relative concentrations (μg L−1) of toxin in sampled lakes from the 2017 National Lakes Assessment [30]. The darker hydrologic regions indicate watershed boundaries belonging to the Mississippi River drainage. Symbol sizes show relative toxin concentrations based on 5 quantile data distributions, with each symbol size representing 20% of the dataset. Yellow symbols (not all are visible on the map) indicate lakes with toxin levels considered potentially harmful to people when swimming or participating in other activities in or on the water (>8 μg L−1). WBDHU2, Watershed Boundary Dataset for 2-digit Hydrologic Unit.
Figure 9.
Geographic distribution and concentrations of microcystins in the conterminous USA. The map shows hydrologic regions and distribution and relative concentrations (μg L−1) of toxin in sampled lakes from the 2017 National Lakes Assessment [30]. The darker hydrologic regions indicate watershed boundaries belonging to the Mississippi River drainage. Symbol sizes show relative toxin concentrations based on 5 quantile data distributions, with each symbol size representing 20% of the dataset. Yellow symbols (not all are visible on the map) indicate lakes with toxin levels considered potentially harmful to people when swimming or participating in other activities in or on the water (>8 μg L−1). WBDHU2, Watershed Boundary Dataset for 2-digit Hydrologic Unit.

Figure 10.
Geographic distribution and concentrations of cylindrospermopsin in the conterminous USA. The map shows hydrologic regions and distribution and relative concentrations (μg L−1) of toxins in sampled lakes from the 2017 National Lakes Assessment [30]. The darker hydrologic regions indicate watershed boundaries belonging to the Mississippi River drainage. Symbol sizes show relative toxin concentrations based on 5 quantile data distributions, with each symbol size representing 20% of the dataset. No samples contained toxin levels considered potentially harmful to people swimming or participating in other activities in or on the water (>15 μg L−1). WBDHU2, Watershed Boundary Dataset for 2-digit Hydrologic Unit.
Figure 10.
Geographic distribution and concentrations of cylindrospermopsin in the conterminous USA. The map shows hydrologic regions and distribution and relative concentrations (μg L−1) of toxins in sampled lakes from the 2017 National Lakes Assessment [30]. The darker hydrologic regions indicate watershed boundaries belonging to the Mississippi River drainage. Symbol sizes show relative toxin concentrations based on 5 quantile data distributions, with each symbol size representing 20% of the dataset. No samples contained toxin levels considered potentially harmful to people swimming or participating in other activities in or on the water (>15 μg L−1). WBDHU2, Watershed Boundary Dataset for 2-digit Hydrologic Unit.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
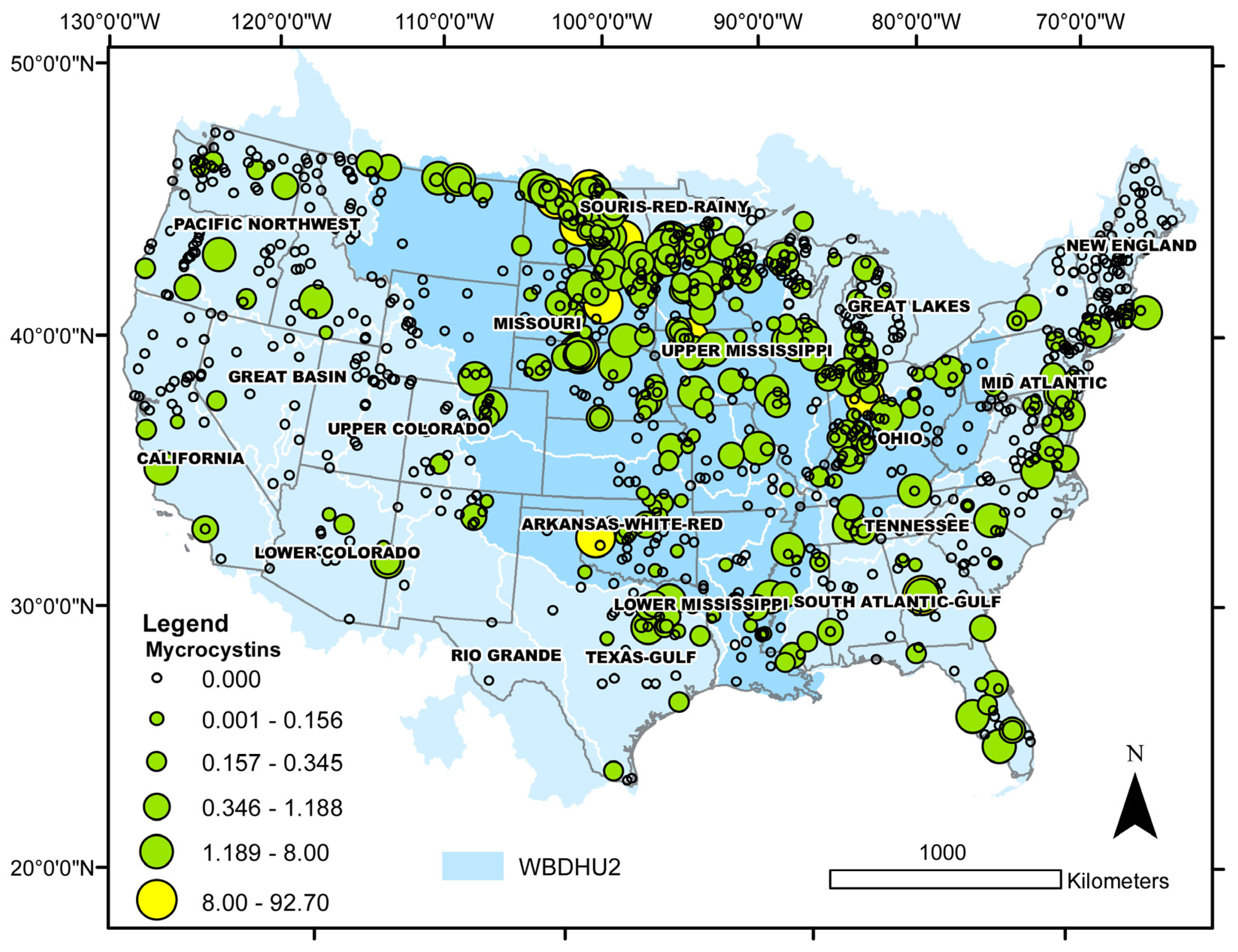{kind=link}
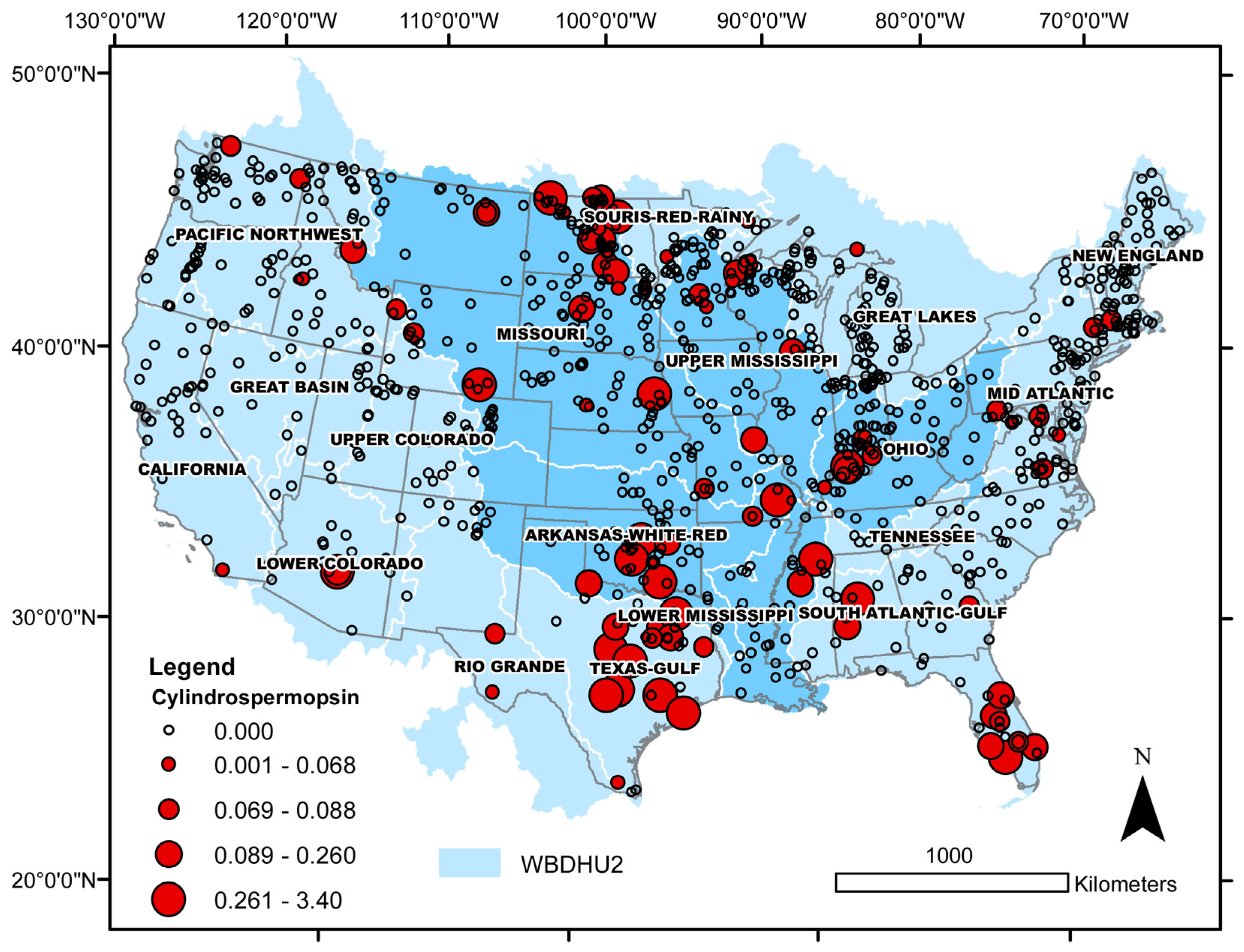{kind=link}
Table 1.
Binomial test for differences between observed and expected distributions of cyanobacteria and Euglena in freshwater (≤0.5 psu) and brackish (>0.5 psu) lakes of the conterminous USA. Cases with densities ≥10,000 cells mL−1 for cyanobacteria and >method detection limit for Euglena were considered. The expected distribution is the distribution of freshwater and brackish sites in the database: 88.3 and 11.7%, respectively (total number of cases, 1090). Also shown are the median, minimum (min), and maximum (max) cell densities (cells mL−1) for each taxon with the corresponding salinity (psu) and sulfate concentration (mg L−1) at the sampling sites. Data are from the 2017 National Lakes Assessment [30]. p-values are two-tailed.
Table 1.
Binomial test for differences between observed and expected distributions of cyanobacteria and Euglena in freshwater (≤0.5 psu) and brackish (>0.5 psu) lakes of the conterminous USA. Cases with densities ≥10,000 cells mL−1 for cyanobacteria and >method detection limit for Euglena were considered. The expected distribution is the distribution of freshwater and brackish sites in the database: 88.3 and 11.7%, respectively (total number of cases, 1090). Also shown are the median, minimum (min), and maximum (max) cell densities (cells mL−1) for each taxon with the corresponding salinity (psu) and sulfate concentration (mg L−1) at the sampling sites. Data are from the 2017 National Lakes Assessment [30]. p-values are two-tailed.
| Taxon | Number of Cases | Median Cell Density (min, max) × 104 | Median Salinity (min, max) | Median Sulfate (min, max) | Lake Salinity | Observed Distribution (%) | Binomial Test p-Value |
|---|---|---|---|---|---|---|---|
| Anabaena | 12 | 1.52 (1.09, 125) | 0.17 (0.02, 2.15) | 10.0 (0.25, 1483) | freshwater | 67 | 0.0433 |
| brackish | 33 | ||||||
| Aphanizomenon | 126 | 5.73 (1.00, 105) | 0.17 (0.00, 2.97) | 15.8 (0.02, 2343) | freshwater | 76 | <0.0001 |
| brackish | 24 | ||||||
| Dolichospermum | 81 | 2.73 (1.02, 39.2) | 0.16 (0.01, 2.25) | 12.4 (0.00, 1736) | freshwater | 81 | 0.0809 |
| brackish | 19 | ||||||
| Microcystis | 93 | 3.30 (1.03, 137) | 0.18 (0.01, 3.79) | 17.9 (0.00, 1525) | freshwater | 80 | 0.0146 |
| brackish | 20 | ||||||
| Planktothrix | 46 | 25.0 (1.07, 558) | 0.43 (0.02, 2.94) | 128 (0.27, 2888) | freshwater | 52 | <0.0001 |
| brackish | 48 | ||||||
| Euglena | 26 | 0.03 (0.006, 0.46) | 0.13 (0.01, 3.79) | 8.87 (0.22, 2888) | freshwater | 81 | 0.2218 |
| brackish | 19 |
Table 2.
Binomial test for differences between observed and expected distributions of microcystins and cylindrospermopsin detections in freshwater (≤0.5 psu) and brackish (>0.5 psu) lakes in the conterminous USA. Cases with concentrations greater than the method detection limit (MDL) for both toxins and >8 μg L−1 for microcystins were analyzed. The national recreational water quality criterion for microcystins is 8 μg L−1, and for cylindrospermopsin, it is 15 μg L−1, but no cases above the latter were recorded. The expected distribution is the distribution of freshwater and brackish sites in the database: 88.3 and 11.7%, respectively (total number of cases: 1090). Also shown are the median, minimum (min), and maximum (max) concentrations (μg L−1) for each toxin and the corresponding salinity (psu) at the sampling sites. Data are from the 2017 National Lakes Assessment [30]. p-values are two-tailed.
Table 2.
Binomial test for differences between observed and expected distributions of microcystins and cylindrospermopsin detections in freshwater (≤0.5 psu) and brackish (>0.5 psu) lakes in the conterminous USA. Cases with concentrations greater than the method detection limit (MDL) for both toxins and >8 μg L−1 for microcystins were analyzed. The national recreational water quality criterion for microcystins is 8 μg L−1, and for cylindrospermopsin, it is 15 μg L−1, but no cases above the latter were recorded. The expected distribution is the distribution of freshwater and brackish sites in the database: 88.3 and 11.7%, respectively (total number of cases: 1090). Also shown are the median, minimum (min), and maximum (max) concentrations (μg L−1) for each toxin and the corresponding salinity (psu) at the sampling sites. Data are from the 2017 National Lakes Assessment [30]. p-values are two-tailed.
| Toxin | Number of Cases | Median Toxin Concentration (min, max) | Median Salinity (min, max) | Lake Salinity | Observed Distribution (%) | Binomial Test p-Value |
|---|---|---|---|---|---|---|
| Microcystins > MDL | 295 | 0.34 (0.10, 92.7) | 0.17 (0.00, 21.5) | freshwater | 78 | <0.0001 |
| brackish | 22 | |||||
| Cylindrospermopsin > MDL | 85 | 0.09 (0.05, 3.40) | 0.11 (0.00, 7.24) | freshwater | 76 | 0.002 |
| brackish | 24 | |||||
| Microcystins > 8 μg L−1 | 12 | 16.2 (8.02, 92.7) | 0.44 (0.01, 2.10) | freshwater | 50 | 0.0013 |
| brackish | 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Patiño, R.; Christensen, V.G.; Graham, J.L.; Rogosch, J.S.; Rosen, B.H. Toxic Algae in Inland Waters of the Conterminous United States—A Review and Synthesis. Water 2023, 15, 2808. https://doi.org/10.3390/w15152808
AMA Style
Patiño R, Christensen VG, Graham JL, Rogosch JS, Rosen BH. Toxic Algae in Inland Waters of the Conterminous United States—A Review and Synthesis. Water. 2023; 15(15):2808. https://doi.org/10.3390/w15152808
Chicago/Turabian StylePatiño, Reynaldo, Victoria G. Christensen, Jennifer L. Graham, Jane S. Rogosch, and Barry H. Rosen. 2023. "Toxic Algae in Inland Waters of the Conterminous United States—A Review and Synthesis" Water 15, no. 15: 2808. https://doi.org/10.3390/w15152808
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.