

How Can Plants Help Restore Degraded Tropical Soils?
by
 , and
, and
Renaud Massoukou Pamba
1,2,
Vincent Poirier
1,*,
Pamphile Nguema Ndoutoumou
3 and
and
Terence Epule Epule
1 1
Agriculture and Agri-Food Research and Development Unit of Abitibi-Temiscamingue, University of Québec in Abitibi-Temiscamingue, Notre-Dame-du-Nord, QC JOZ 3B0, Canada
2
Ministry of Water and Forests, Sea, Environment, in Charge of the Climate Plan and the Land Use Plan, Libreville P.O. Box 199, Gabon
3
Laboratory of Plant Biotechnology, Department of General Agronomy, Institute of Agricultural and Forestry Research, National Centre for Scientific and Technological Research, Libreville P.O. Box 2246, Gabon
*
Author to whom correspondence should be addressed.
Land 2023, 12(12), 2147; https://doi.org/10.3390/land12122147
Submission received: 11 October 2023
/
Revised: 27 November 2023
/
Accepted: 30 November 2023
/
Published: 9 December 2023
(This article belongs to the Special Issue Soils for the Future)
Abstract
:In the tropics, anthropogenic activities can lead to water and wind erosion, a loss of biodiversity, and a reduction in sequestered carbon, fertility, and organic matter content in the soils concerned, potentially resulting in their degradation. This study therefore aims to identify the mechanisms used by plant species to restore degraded tropical soils and plant species characteristics that are best suited to achieve this through a critical scoping review of the peer-reviewed literature. Soil restoration leads to the re-establishment of ecosystem services and an increase in soil production potential, the regeneration of biodiversity, the stopping of organic matter losses, and the creation of favorable conditions for carbon sequestration and nitrogen fixation. The choice of appropriate plant species depends on the restoration objectives to be achieved. Five key mechanisms by which plant species contribute to restore degraded tropical soils include: (1) nitrogen fixation, (2) carbon sequestration, (3) organic matter addition, (4) structure stabilization, and (5) erosion control. The main characteristics of plant species and vegetation involved in these mechanisms are (a) the capacity to form symbiotic associations with N-fixing bacteria and mycorrhizae, (b) the production of abundant root biomass releasing litter and exudates, (c) roots having a high length density, branching intensity, and depth distribution, (d) the production of an abundant and easily decomposed above ground litter, (e) the production of a vast canopy, and (f) the presence of different vegetation strata. Targeting these characteristics will contribute to acting on several mechanisms simultaneously, which will increase the chance of success in tropical soil restoration.
1. Introduction
The world’s population is constantly growing. Estimates put the number of inhabitants at over 8.05 billion in 2023, with a projection of 9.70 billion in 2050 and 10.35 billion in 2100 [1,2]. This population growth is associated with an increased pressure on natural resources such as water and soil [3]. Indeed, soil is increasingly solicited for multiple crops to achieve food self-sufficiency. This pressure on soil opens the debate on the renewable nature of soils. Girgenti et al. [4] argue that soils are non-renewable, while several other studies show that soils can indeed be renewed in quantity and quality through agroecology and conventional options [5,6]. Soils have been described as the bedrock of life on earth [7] and are derived from the degradation of the parent rock through physical, chemical, and biological processes [8]. It is the primary resource on which life develops worldwide [9] and is a diversified and structured habitat where several resources such as water, carbon (C), nitrogen (N), and organic matter are stored [9,10]. However, soils are variable and require a great deal of effort, complex analysis, and multidisciplinary knowledge [11] to understand the processes leading to their formation, functioning, and degradation.
Soil quality can be defined as the capacity of a soil to host and provide ecosystem services [9,12,13]. Soil is said to be healthy when it can withstand stress and disturbance, recycle nutrients, transform organic matter, and support microbial life [14,15]. To assess the quality or health of a soil, we need to focus on (i) physical properties such as the stability of its structure and its level of compaction, porosity, and infiltration capacity, (ii) chemical properties such as pH and its concentrations of nutrients (particularly N), and (iii) biological properties such as root growth, biodiversity, and organic matter and C content [9]. Healthy soils perform several ecosystem services such as (1) the retention, filtration, and purification of water, (2) the production of biomass and food for humans and animals, (3) the provision of habitat for organisms and the conservation of biodiversity, (4) the regulation of climate at local and global scales, (5) the recycling of nutrients, and (6) the provision of a stable physical support for plant species [9,16,17,18]. Unfortunately, soil health can deteriorate either due to anthropogenic reasons, and is therefore linked to human activities, or due to natural causes, and is therefore linked to climate change and other related factors [9,19]. Demographic pressure leads to the intensive exploitation of soils, resulting in their degradation characterized by wind and water erosion, pollution, a loss of biodiversity and organic carbon, and a loss of macro (N, P, K) and micronutrients (Zn, Cu, Mo), as well as increased toxicity in certain elements (Al, Mn, Fe) [16]. Soil degradation causes a reduction in expected ecosystem services and leads to a decline in soil quality and productivity [9,20,21].
The tropical zone extends on either side of the equator, between the Tropic of Cancer (23°27′ north latitude) and the Tropic of Capricorn (23°27′ south latitude); it covers 45% of the world’s forest area and around 8.2 billion hectares, or around 40% of the planet’s land [17]. Due to various human activities such as deforestation and extensive agriculture, soil in the tropical zone is subject to degradation. A strong correlation between soil degradation and forest loss in the Congo Basin has been observed [19]. However, soil is of vital importance for agriculture and the environment. Hence, it is imperative to have more arable soil to produce more food. It would therefore be expedient to develop alternative ways of restoring degraded tropical soils. The remediation of these soils will make it possible to regain more arable land, limit deforestation, and protect soils and forests threatened by overexploitation. The restoration of degraded tropical soils involves stopping existing degradation processes and restoring the soil’s physiochemical characteristics [16,17,22]. Several soil restoration methods and strategies are described in the literature. One of these is phytoremediation, which involves using plants and soil microbes to obtain the twin objectives of a reduced concentration of toxic contaminants and to restore soil quantity and quality [17,23]. In this study, the term phytoremediation is used in the context of restoring soil quantity and quality. From an etymological point of view, “phyto” means “plant” in Greek and “remedium” means “restore balance” in Latin. Indeed, the cultivation of plant species promotes soil restoration [17,24,25,26,27,28] by providing organic matter and making them more fertile [29].
Currently, there are no critical reviews that examine the potential of plants in restoring tropical soils. The current studies that exist are individual based studies that focus on phytoremediation [30,31,32] without examining the actual characteristics of the plants concerned. This work is original and offers a major contribution in that it goes beyond by bringing together a critical perspective of the knowledge scattered in several studies. This literature review aims to respond to the following question: “What are the mechanisms used by plant species to restore degraded tropical soils, and which plant species characteristics are best suited to achieving this?” To this end, we will first define the key words used in this work, then provide an overview of the mechanisms used and the characteristics of species involved in the restoration of degraded tropical soils and give examples of plant species that contribute to achieving this goal.
2. Methodology
Our approach was inspired by the guidelines given to perform systematics reviews, which is a scientific method that combines the results of several independent studies on a well-defined problem to perform a rigorous secondary analysis of the primary results [33,34,35]. Our approach was inspired by the PRISMA statement [36]. However, our work is not a systemic review as we did not follow point-by-point the 27-item checklist recommended [36]. This work is a critical scoping review that synthesizes key trends on the use of plant species for the restoration of degraded tropical soils. Firstly, we based our review on a clearly formulated question. Next, we specified explicit methods and criteria for the inclusion or exclusion of articles for our critical scoping review (see Table 1).
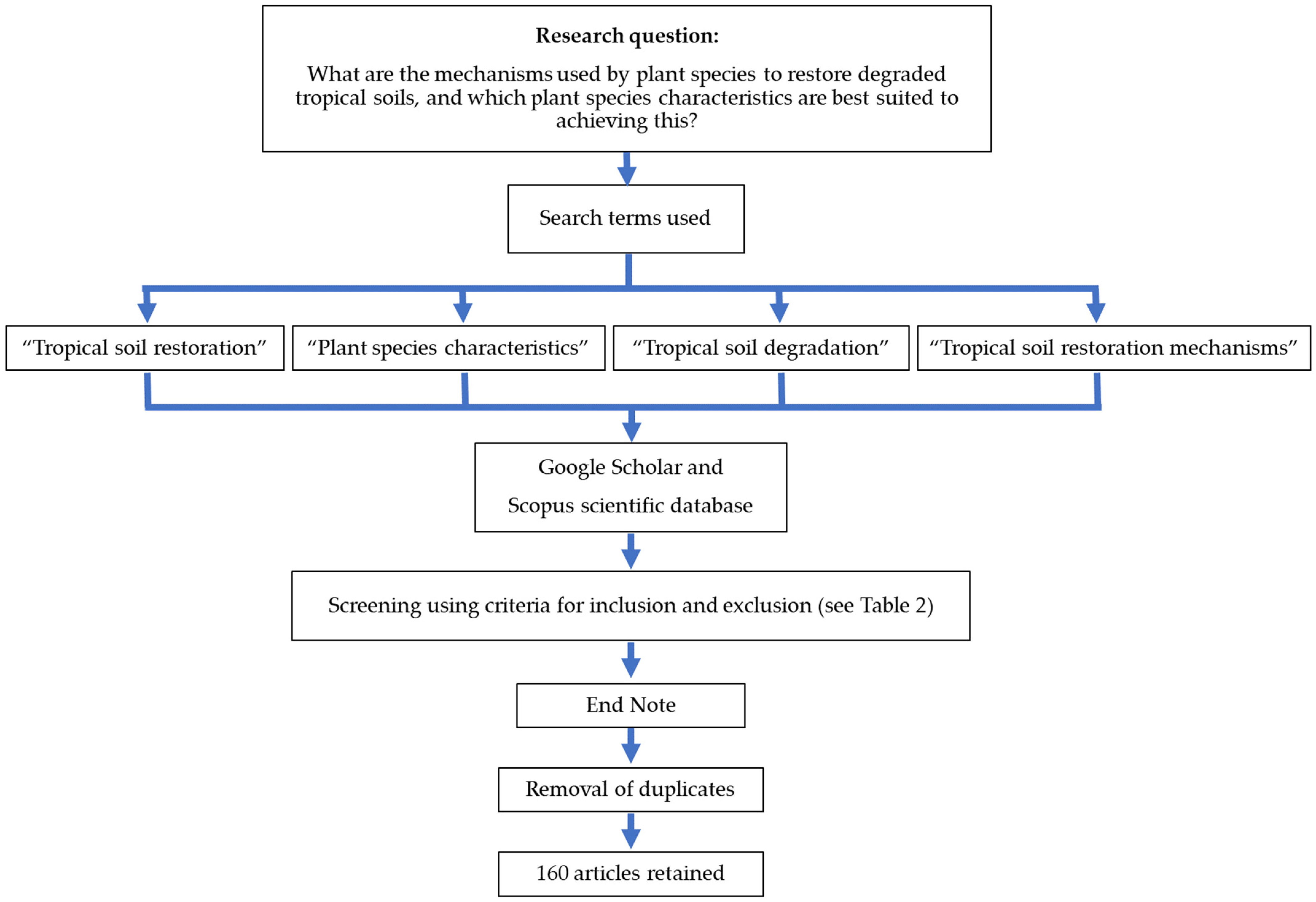
The type of literature used was solely peer-reviewed; gray and non-reviewed literature was omitted since we considered it as non-standardized knowledge. The present study is based on data collected from the French and English scientific literature documenting the restoration of degraded tropical soils. The search terms we used were “plant species characteristics”, “tropical soil degradation”, “tropical soil restoration”, and “tropical soil restoration mechanisms”. We focused on soils of the tropical zone; according to Michéli et al. [37], there are eight major soil groups found in tropical environments (see Table 2). We focused on articles published over the last 30 years, i.e., between 1993 and 2023. The articles were acquired using Scopus and Google Scholar search engines since they provide easy access to the primary literature on the topics of interest. Several potentially relevant publications were selected from the initial search and duplicates were removed after importing the articles into the EndNote software. Overall, a total of 160 references were used. A schematic representation of our approach is presented in Figure 1.
3. Results and Discussion
3.1. Tropical Soil Degradation and Importance of Their Restoration
In the tropics, physical soil degradation takes the form of desertification, compaction, runoff, and erosion [61], chemical soil degradation leads to acidification, salinization, leaching, and pollution [9,61], and biological soil degradation results in the loss of biodiversity, organic matter, and the capacity to sequester C and fix N [9,14,21]. Anthropogenic activities such as excessive tillage, deforestation, biomass burning, compaction, and overexploitation are the main causes of tropical soil degradation [62,63]. Lal [9] and Lahmar and Ruellan [22] state that soil resilience is the ability of a soil to restructure, regenerate, and provide ecosystem services after anthropogenic or natural disturbances. Restoring degraded tropical soils involves rebuilding soil structure and the ecological processes associated with it. This involves restoring the soil’s production potential, renewing biodiversity, stopping organic matter losses, creating conditions that are favorable for C sequestration and N fixation [9,22], and improving soil aggregation and structural stability [64]. Most of the food produced to feed human beings in the tropics and elsewhere comes from the soil. The absence or deficit of cultivable soil will immediately translate into famine. To avoid this, it is important, if not essential, to examine and put into practice all possible methods of restoring degraded tropical soils. Restoring degraded tropical soils is a global priority to increase food security and regulate the climate globally [65].
3.2. Involving Plants in the Restoration of Degraded Tropical Soils
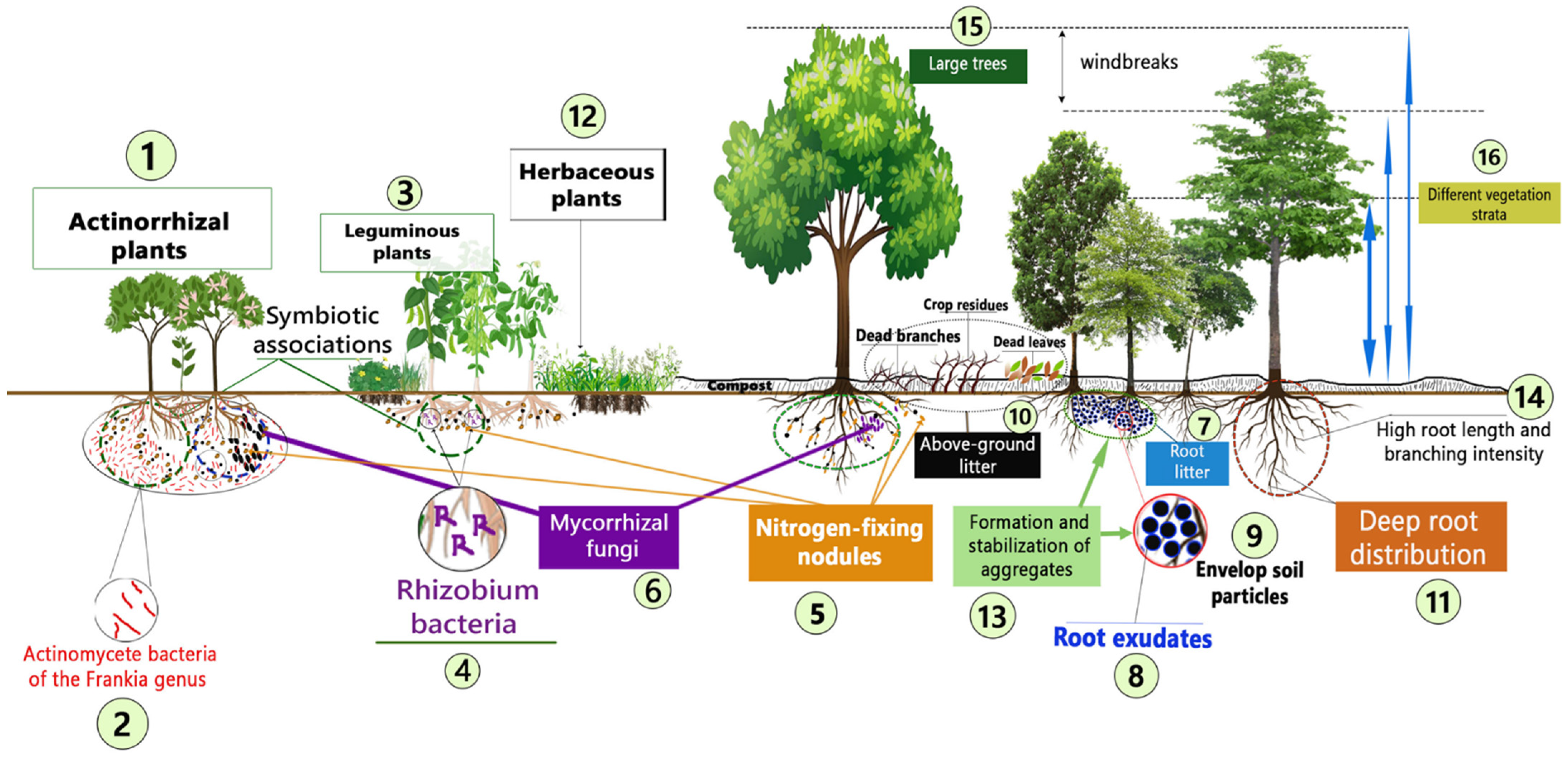
In this study, five major mechanisms by which plant species can contribute to restoring degraded tropical soils were identified (Figure 2 and Table 3). These are nitrogen fixation, carbon sequestration, organic matter addition, soil structure stabilization, and wind–water erosion control. These mechanisms will be described, followed by examples of plant species that promote them, and then the characteristics of the targeted plant species will be highlighted. Thereafter, the limits associated with each mechanism will be discussed.
3.3. Nitrogen Fixation to Restore Degraded Tropical Soils
3.3.1. Mechanisms of Nitrogen Fixation by Plant Species
Some plant species develop symbiotic mechanisms for fixing atmospheric N in the soil, thereby helping to regenerate soil fertility and improve crop yields [66]. According to Duponnois et al. [67], plants in the legume family (Fabaceae) are renowned for their ability to restore degraded soils by symbiotically associating with bacteria of the genus Rhizobium which can fix atmospheric N into soil N [27]. Actinorhizal plants form symbiotic associations with actinomycete bacteria of the Frankia genus [68] and are also capable of fixing atmospheric N into soil N. It is thanks to nodules located on the roots that leguminous and actinorhizal plants develop symbiotic nitrogen-fixing mechanisms [67]. In line with this mechanism, the characteristics of target plants are therefore the capacity to form nodules and the possession of symbiotic genes carried by plasmids, in accordance with the work of Benabdoun et al. [66]. The development of a tripartite symbiotic association between bacteria of the genus Rhizobium or Frankia and mycorrhizal fungi is also a feature developed by certain plant species. This association enriches the soil and improves its fertility thanks to the ability of Rhizobium and Frankia to fix atmospheric N and that of mycorrhizae to explore the soil’s superficial and deep horizons to acquire resources [69,70].
3.3.2. Examples of Nitrogen-Fixing Plant Species for Restoring Degraded Tropical Soils
Benbrahim et al. [70] affirmed in their study that Acacia species are atmospheric N-fixing plants which enrich and improve the fertility of tropical soils. A stand of Faidherbia albida with a density of 50 trees ha−1 produces litter that reintroduces into the soil each year the equivalent of 75 kg N ha−1 along with other nutrients, i.e., 12, 13, 20, 25, and 120 kg ha−1 of P, K, S, Mg, and Ca, respectively [70]. Cowpea (Vigna unguiculata) is a tropical legume capable of fixing between 32 and 89% of its N requirements [71]. The cultivation of groundnuts (Arachis hypogaea), a legume grown for human consumption, fixes 8 to 23 kg N ha−1 in the soil, thus increasing soil mineral N by 13 to 40% [72]. The results of a study carried out by Azontonde et al. [73] on the cultivation of Mucuna pruriens (var. utilis) as a cover legume crop show that this species contributes 290 kg N ha−1 to the soil, as well as 100, 240, 100, 5320, and 40 kg ha−1 of P, K, Mg, C, and Ca, respectively. Thus, M. pruriens (var. utilis) strongly favors the restoration of degraded tropical soils due to its high capacity for atmospheric N fixation and its strong limitation of erosion and runoff [73]. Dommergues [74] states that plants of the Alnus and Casuarina genera are among the actinorhizal plants with the greatest capacity to fix atmospheric N. Benabdoun et al. [66] have demonstrated that the tropical woody species Casuarina glauca is drought-resistant and makes a significant contribution to restoring degraded soils. In Egypt, the actinorhizal plants Casuarina equisetifolia, Casuarina cunninghamiana, and C. glauca have been shown to restore degraded soils; they can fix up to 116 kg N ha−1 yr−1 two years after sowing [66]. Plants of the Casuarina genus can therefore rapidly restore soil fertility. The findings of a study conducted by Coulibaly et al. [64] affirm that growing Jatropha curcas increases total nitrogen in the soil microaggregate fraction.
Altogether, these results indicate that the following plant characteristics are suitable to enrich tropical soil N concentration and contribute to their restoration: (i) being actinorhizal and leguminous plants, (ii) possessing symbiotic genes that maximize the formation of symbiotic associations with actinomycete bacteria of the Frankia genus and bacteria of the Rhizobium genus, and (iii) being capable of forming tripartite symbiotic association with mycorrhizal fungi.
3.4. Carbon Sequestration to Restore Degraded Tropical Soils
3.4.1. Carbon Sequestration Mechanisms
In this study, the focus is on biotic carbon (C) sequestration mechanisms, i.e., those linked to the biological activities of plants, fauna, and microorganisms [75,76,77,78]. Carbon sequestration can be defined as the fixation and stabilization of C in the soil in the form of organic matter over a long period, such as 50 years [79]. According to Lützow et al. [80], recalcitrance to decomposition, occlusion into micro- and macroaggregates, and organo-mineral interactions are the main mechanisms leading to organic C sequestration in soil. Lal [9] estimates that 10 to 15 g C kg−1 of soil corresponds to a minimal critical threshold of soil carbon concentration. Any practices that contribute to increasing organic C concentration above this critical threshold would help restore degraded soils [9,81]. Mycorrhizae located on the roots of plant species and roots themselves have the capacity to sequester organic carbon, stabilize soil structure, and thus restore it [82,83]. According to Cotrufo et al. [84], organic residues easily decomposed by microorganisms could lead to increased C sequestration; the microbial products of decomposition could promote aggregation and organo–mineral interactions.
3.4.2. Examples of Plant Species to Sequester C and Restore Degraded Tropical Soils
Belowground plant biomass plays an important role in the soil C sequestration process [85]. Jones et al. [77] argue that 27% of the C sequestered in the soil is derived from root mechanisms, of which 11% is contributed by rhizo deposits and 16% by root biomass. Herbaceous plants are monocotyledons that produce high root biomass [86], which would lead to significant organic C sequestration and potentially a good level of soil restoration. They also produce biomass that can be easily decomposed and consequently increase C concentration in superficial soil layers [87]. The characteristics of plant species that contribute to sequestering organic C in the soil are also closely linked to root traits of an architectural, morphological, chemical, physiological, and symbiotic nature [88,89]. According to [81], plant species producing a high root biomass with a high lignin and suberin content contribute to sequestering organic C in the soil through the mechanism of recalcitrance against decomposition and could consequently contribute to restoring degraded soils. Increased root depth distribution is also favorable for C sequestration in deep soil layers [88].
Plant species grown as cover crops such as sorghum (Sorghum bicolor L.), maize (Zea mays L.), soy (Glycine max. L.), and beans (Phaseolus vulgaris) protect the soil against degradation and sequester C [81]. Kang et al. [90], in a study carried out in Nigeria on an intercropping association of living hedges of two legume species, Gliricidia sepium and Leucaena leucocephala, obtained a 15% (i.e., 2.38 Mg C ha−1) improvement in soil organic C stocks between 0 and 25 cm depth in 12 years, compared with single crops. In Hawaii, the conversion of pasture soils to Eucalyptus plantations increased C stocks by 17.5% between 0 and 1 m soil depth after 7 years [91]. In Madagascar, coppicing Eucalyptus robusta in the highlands resulted in an increase in stored C of 38 Mg ha−1 compared with the original savannah, mainly via the stumps and root system [92]. The cultivation of Inga edulis in hedgerows in Peru by [93] also resulted in a 12% (i.e., 0.23 Mg C ha−1) increase in soil organic C over 6 years. In Brazil, a soil receiving fresh plant biomass mulch from sugarcane (Saccharum officinarum) residues for 6 years was richer in C than a soil cultivated with residue burning for 50 years, with total C content of 4.2 g kg−1 at a depth of 0–5 cm (+20%) and 3.0 g kg−1 at a depth of 0–10 cm (+15%), representing an average annual gain of 0.45 t C ha−1 yr−1 at a depth of 0–5 cm and 0.65 t C ha−1 yr−1 at a depth of 0–10 cm [94]. The study by Graham [95] in South Africa on a ferralitic clay soil cultivated with unburnt sugarcane and mulched with crop residues for 59 years concluded that soil C content increased by 12 g C kg−1 at a depth of 0–5 cm, of which only 0.5 g C kg−1 was in the light organic fraction (density < 1.7), made up mainly of coarse organic matter; most of the C was thus found in denser fractions where it was likely stabilized by organo–mineral interactions. The study conducted by [64] led to the conclusion that Jatropha curcas cultivation increases total soil organic carbon in the microaggregate fraction and can sequester an average of 1.33 g C kg−1 year−1. In India, a research station located in Patancheru concluded that growing Jatropha curcas in a 4-year plantation fixed 3.07 kg C per plant, an increase of 19% compared to adjacent grasslands [96]. The results of a study conducted in Senegal in 2017 on allometric equations and carbon stock in the biomass of Jatropha curcas L. trees revealed that plantations stored about 1200 kg C ha−1 in belowground biomass after 5 years [97].
Plant species characteristics that could promote C sequestration mechanisms and contribute to restore tropical soils are (i) abundant root production which provides litter and exudates stimulating microbial activity, (ii) the capacity to form symbiotic association with N-fixing bacteria and mycorrhizal fungi, (iii) increased depth distribution of the root system, and (iv) the production of abundant and easily decomposable litter aboveground.
3.5. Organic Matter Addition to Restore the Fertility of Degraded Tropical Soils
3.5.1. Soil Fertility and Organic Matter Addition Mechanisms
In the absence of exogenous organic amendments such as manure or compost, organic matter in the soil is contributed by the restitution of plant biomass, i.e., leaves, roots, stems, crop residues, and senescent organs [98]. It is difficult to dissociate soil fertility and organic matter; these two characteristics go hand in hand and are complementary, as they are regulated by the chemical, physical, and biological properties of the soil [99]. The mechanisms for rebuilding soil fertility rely on the mineralization of (i) fresh and decomposed organic matter [100,101], (ii) microbial products [99,102], and (iii) soil organic matter [102]. The two elements that come into play in the impact that the addition of organic matter can have on the soil are (i) the quantity of organic matter added and (ii) its chemical composition, which affects its capacity to be degraded. Several authors agree that plant biomass consisting mainly of leaves and roots helps to maintain or even increase soil organic matter and fertility [98,103].
3.5.2. Examples of Species That Provide Organic Matter to Restore Tropical Soils’ Fertility
The cultivation of perennial plants is useful for the restoration of degraded tropical soils, as they provide a permanent plant cover that can maintain soil fertility and combat erosion [104,105]. The diversity of species and mechanisms that take place in agroforestry systems, due to biological diversity and greater plant biomass productivity, increase soil fertility and organic matter, thus helping to restore degraded soils [104,106]. The cultivation of cover crops such as sorghum (Sorghum bicolor L.), maize (Zea mays L.), soya (Glycine max. L.), and beans (Phaseolus vulgaris) improves soil fertility and maintains its organic matter content [107]. In a cocoa (Theobroma cacao L.) plantation in Cameroon, aboveground plant biomass consisted of 60–70% leaves, 20–30% stems, and 7–10% flowers and fruits; the soil amended with this biomass benefited from about 1–2, 75–100, 30–75, 5–6, and 35–40 kg ha−1 an−1 of P, K, Mg, N, and Ca, respectively. The natural shade offered by the vegetation increases the percentage of moisture at the soil surface, thus promoting the decomposition of plant biomass [108]. The species Acacia mearnsii is a cultivated plant for the restoration of tropical degraded soils as it produces nearly 25,000 Mg ha−1 yr−1 of fresh leaves, providing 245 to 255 kg N ha−1 annually [70,109]. In a 14- and 34-year-old Pinus Kesiya forest plantation, Njoukam et al. [103] obtained plant biomass production of 8293 and 9867 kg ha−1 yr−1 made up of 74 and 86% leaves, respectively. Tree roots also increase organic matter, stimulate microbial activity, enhance mineral bioavailability, and improve soil fertility and quality [9,22]. Mulching crop residues and installing cover crops increases soil organic matter and stimulates earthworm activity [110]. Cultivation of Jatropha curcas to restore degraded soils in India yields around 550 g of dry leaves per plant annually added to the soil by litterfall, resulting in an increase in dry biomass of 305 kg C ha−1 year−1 [96]. This translates into an increase in soil organic matter but also an overall increase in soil fertility linked to a growth in microbial activity under Jatropha curcas [96].
Among the characteristics of plant species that promote the addition of organic matter to rebuild soil fertility are (i) the production of abundant litter on the soil surface via the fall of aerial organs, (ii) the production of abundant litter in the soil by root biomass, and (iii) the capacity of above- and belowground biomass to be decomposed to feed the soil.
3.6. Structure Stabilization to Restore Degraded Tropical Soils
3.6.1. Mechanisms of Soil Structure Stabilization by Plant Species
According to many authors, the roots of plant species influence the biotic mechanisms of aggregate formation and stabilization [88,111,112]. The mechanisms involved in soil structural stabilization are (i) the envelopment of macroaggregates by decomposition products, (ii) the production of root exudates and microbial polysaccharides that act as binding agents between soil particles and stimulate microbial activity, (iii) the entanglement by roots and hyphae that enhance particle and aggregate agglomeration, and (iv) the alignment and compression of soil particles that promote their adhesion [89,113,114]. Thanks to their aerial parts, plants attenuate the intensity of raindrops and their impact on the breakdown of soil aggregates, thereby improving soil structural stability [115,116]. Abiven et al. [117] believe that the decomposition of plant biomass promotes aggregate stability; the faster it decomposes, the better the stability of the aggregates.
Roots also have a major influence on soil aggregate and structural stability. They compress and align soil particles, produce exudates whose chemical composition influences their use by microorganisms, and affect the composition of bacterial fungal communities [118]. Root exudates rich in polymerized sugars act as a binder between solid particles, improving soil structural stability [115,119]. Root characteristics that help stabilize soil macroaggregates include high root length density and high root branching intensity [88,120,121]. Roots with a high concentration of polyphenols can react with soil proteins to form macromolecules that improve aggregate stability [88,114,122]. The combined effect of microbial activities in the mycorrhizosphere with root growth and hyphal length improves aggregate stability in the soil [123,124]. Aggregate stability is also enhanced by the decomposition of deciduous leaves falling from trees via the soil’s microbial biomass, which exudes microbial products that act as binding agents between soil particles [64,125].
3.6.2. Examples of Species That Stabilize Soil Structure to Restore Degraded Tropical Soils
Plant species capable of forming mycorrhizal associations such as Pinus halepensis, Shorea pinanga, Acacia mangium, and species from the genera Eucalyptus, particularly E. urophylla, contribute to the formation of stable aggregates and the restoration of degraded tropical soils [89]. Aggregate stability is also enhanced by high root length and mycelial hyphae density [126]. Soil aggregation and structural stability are enhanced by the hyphae of mycorrhizal fungi located on plant root systems [104,127]. The study carried out by Poirier et al. [88] in a Mediterranean climate attest that, on average, the 13 plants studied increased stable soil macroaggregates by 13% compared with bare soil. However, under the species Sanguisorba minor, the increase in soil macroaggregates reached 40%. In a study carried out in Tunisia, Annabi et al. [115] asserted that the association of wheat (Tritium sp.) and faba bean (Vicia faba) contributes strongly to the stability of soil aggregates and helps prevent water erosion. The effects were observed from the third month after sowing and continued for up to four months after harvest. The concentration of water-stable aggregates in soil amended with Lolium rigidum and Scutellospora calospora mulch increased to 84 and 143 g kg−1 soil; mulch addition resulted in 63% and 147% increases in the quantity of water-stable aggregates compared to the unamended control soils, respectively [128]. In Brazil, mulch made from fresh plant biomass of sugarcane (Saccharum officinarum) residues improved soil structural stability at a depth of 0–5 cm, with 17% more stable macroaggregates in unburnt sugarcane fields compared to burnt fields [94]. Short-term (10 weeks) pot cultivation of sorghum (Sorghum bicolor L.) significantly improved soil structural stability via inoculation with arbuscular mycorrhizal fungi [123,124]. Degraded soils under Jatropha curcas cultivation have observed an increase in macroaggregation, which promoted soil structural stability and contributed to soil restoration [64,129].
From the abovementioned results, we identified the following characteristics of plant species which can promote tropical soil structural stability and contribute to its restoration: (i) the production of abundant root litter and exudates, (ii) the capacity to form symbiotic association with mycorrhizal fungi, (iii) the production of roots having a high length density and branching intensity and an increased depth distribution, and (iv) the capacity for trees to grow in height with vast canopy and the combination of different vegetation strata to reduce the intensity of rain drops and the associated breakdown of soil aggregates.
3.7. Controlling Water and Wind Erosion to Restore Degraded Tropical Soils
3.7.1. Mechanisms to Control Water and Wind Erosion
The mechanisms that enhance the slowing down of water erosion are (i) the slowing of rainfall intensity by foliage strata, which reduces the force of the impact of raindrops [130], (ii) the plant biomass on the soil surface (leaves, roots, and grasses), which acts as an obstacle to the movement of water and the transport of soil particles [130], (iii) the deep roots of trees, which stabilize soil structure and limit erosion [131,132], and (iv) the presence of organic horizons in the soil, which regulates water infiltration and limits the runoff of soil particles [133]. To control wind erosion, we need to (i) grow plant species that can create windbreaks made up of trees, shrubs, or bushes which regulate and reduce the speed of wind without stopping it completely and (ii) add mulch to crops by installing a plant cover to protect the soil and thus preserve its water reserves [134].
3.7.2. Examples of Plant Species to Help Control Erosion and Restore Degraded Tropical Soils
The cultivation of plants helps to regulate the infiltration of runoff water into degraded soils and counteract the negative impact that excessive sunshine can have on the soil [107,135]. In this respect, Rey et al. [130] assert that plants with a high aboveground plant biomass intercept raindrops and reduce the intensity of their impact on the soil. Mazour et al. [136], in a study carried out in Algeria, established that natural plant cover reduces water erosion to around 0.4 Mg ha−1 year−1 of soil lost, a reduction of about 93%, compared with bare soil where soil losses are about 5.3 Mg ha−1 yr−1. This study showed that vegetation cover does not only limit soil erosion by facilitating water infiltration but also increases soil fertility and greatly improves its structural stability [136]. Similar results were obtained by Morsli et al. [137] in the same country, with a reduction in water erosion from 3 to 6 Mg ha−1 year−1 of soil lost on bare soil to around 0.8 Mg ha−1 year−1 of soil lost on soil under plant cover. Berkane et al. [138] showed that the intensity of water erosion decreased from 73 to 63 m3 ha−1 of water in western Algeria under forest cover, compared with areas without plant cover where erosion was much more intense. In Madagascar, Douzet et al. [139] achieved a reduction in runoff and erosion coefficients of less than 15% on soils cultivated with cover crops of maize (Zea mays L.), beans (Phaseolus vulgaris), and rice (Oryza sativa), despite an average rainfall of over 1300 mm, compared with soils with ploughed plots. Degraded soils under Jatropha curcas cultivation benefited from reduced water erosion [96,129,140]. This could be due to increased water infiltration in the soil since the cultivation of Jatropha curcas can increase in the weighted mean diameter of soil aggregates by 11% compared to that of the native vegetation [64]. The results of Ouattara et al. [141] showed that a quantity of 2 Mg ha−1 of mulch is necessary and sufficient to control runoff and erosion, improve infiltration, and conserve sufficient moisture. In their study, the runoff coefficient varied from 55% to 16% under plant cover compared with 94% under ploughing without mulching [141].
Mulching has proved effective also in controlling wind erosion. Millet [Pennisetum glaucum (L.) R. B] straw application rates of 2 Mg ha−1, providing a soil cover rate of 7 to 10%, could reduce wind sediment fluxes by 40 to 60% at 10 cm above ground level [142,143]. The study carried out by Tidjani et al. [144] on a sand dune bounded by Leptadenia pyrotechnica fences oriented parallel to the prevailing winds showed that the fences reduced wind flow by more than 98% over a distance 10 times its height. Plants that have deep rooting systems remain upright when winds are violent, and rainfall is heavy [145]. Moreover, the stratification of plant cover slows down rainfall velocity [146] and wind speed [134].
Overall, the characteristics of plant species that should be targeted to control water and wind erosion are (i) a vast canopy that can cover large areas above ground, (ii) an abundant production of litter that can cover the soil surface, (iii) a deep rooting system and an abundant root biomass with high length and branching intensity, and (iv) the presence of different vegetation strata.
3.8. Limitations of the Use of Plants to Help Restore Degraded Tropical Soils
Although our review demonstrates that plants can be helpful in many ways, their use to restore degraded soils in tropical environments through the five mechanisms identified also has limitations. Nitrogen fixation by plants is influenced by environmental conditions such as pH, humidity, temperature, or metal toxicity; the latter affects the plant physiologically, slows down the functioning of microbial communities, and limits the rate of N fixation [147,148]. The quantities of N fixed vary considerably from one species to another, as bacterial strains influence symbiotic activity [149,150]. Environmental conditions such as soil type, level of degradation, texture, mineralogy, water availability, acidity, and microbial community also affect C sequestration and root biomass production [9,151]. In this context, species selection is easier when we know most of the conditions listed above. Soil organic matter is an assemblage of living or dead organic constituents of plant, animal, or microbial origin, whether transformed or not, found in the soil [151,152]. It is therefore difficult to know exactly how much organic matter added from plant biomass contributes to increase soil fertility. The proportion of the organic matter added that will remain in the soil is also conditioned by similar factors to those influencing C sequestration. In many parts of Africa, several studies show that maize consumes a lot of water [153,154]. The advantages and disadvantages of growing this plant should be known to enable it to meet users’ objectives. The characteristics of the roots, leaves or foliage, and trunks of plant species differ from one species to another and are most likely unknown for many species of the tropics. Identifying plant species combining several of the desired characteristics provides better chances of restoring degraded tropical soils [155,156]. The effectiveness of plant species in stabilizing soil structure and aggregates also depends on environmental conditions such as soil moisture, temperature, rainfall, and nutrient availability [157]. These same environmental conditions influence the various living organisms in the soil that help to establish a stable soil structure. Finally, although Eucalyptus contributes to the formation of stable aggregates and the restoration of degraded tropical soils [90], this plant is no longer recommended in reforestation programs as it has extremely deep roots and consumes a lot of water, which deprives the other plants around it [158,159,160].
4. Conclusions
This work identified and described five major mechanisms by which plant species can help restore degraded tropical soils and highlighted the desired characteristics of these species. These mechanisms are (1) nitrogen fixation, (2) carbon sequestration, (3) organic matter addition, (4) structure stabilization, and (5) water and wind erosion control. The characteristics to be targeted when choosing appropriate plant species can be separated into two categories, those associated with aboveground parts on the one hand, and root parts on the other. Aboveground characteristics include the ability to grow tall, have a large leaf area to form a vast canopy, have an abundant leaf, fruit, flower, and branch production to accumulate litter on the soil, and a have a high rate of decomposition of plant residues. Belowground characteristics include high root length, roots with a large diameter and high mass density, high rooting depth, and the ability to form symbiotic associations with mycorrhizal fungi and nitrogen-fixing bacteria. Plant species to be targeted for the restoration of degraded tropical soils must possess at least one or two aerial or root characteristics favorable to at least two or three of the five mechanisms identified in this work. The cultivation of plant species has a triple benefit for degraded soils, as not only do they protect them against erosion and solar radiation, but they also contribute significantly to increase their level of fertility and structural stability. Legumes such as Acacia, as well as other plant species such as sugarcane (Saccharum officinarum), sorghum (Sorghum bicolor L.), Jatropha (Jatropha curcas L.), and cacao (Theobroma cacao), are plants to target for the restoration of degraded tropical soils because they possess favorable characteristics for promoting soil carbon sequestration and structure stability through increased biomass production and a reduction in water and wind erosion. The results of our research review can be helpful to guide researchers, farmers, and peasants on the mechanisms to target species characteristics to be selected for efficiently restoring degraded tropical soils. Future research should aim at characterizing aboveground and root characteristics of species of interest, as well as their impact on soil restoration mechanisms, from a quantitative point of view. This would greatly increase our mechanistic understanding of how plants can help restore degraded soils.
Author Contributions
Conceptualization, R.M.P.; methodology, R.M.P.; software, R.M.P.; validation, V.P. and T.E.E.; formal analysis, R.M.P.; investigation, R.M.P.; resources, V.P.; data curation, R.M.P.; writing—original draft preparation, R.M.P.; writing—review and editing, V.P., P.N.N. and T.E.E.; visualization, R.M.P.; supervision, V.P. and P.N.N. All authors have read and agreed to the published version of the manuscript.
Funding
Our most sincere thanks go to the Programme Canadien des Bourses de la Francophonie (PCBF) for providing us with a grant to R.M.P. (file number 20185529) to finance this work. Please find in these few words all my gratitude.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gu, D.; Andreev, K.; Dupre, M.E. Major trends in population growth around the world. China CDC Wkly. 2021, 3, 604. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, H. Less meat is nearly always better than sustainable meat, to reduce your carbon footprint. Our World in Data, 4 February 2020. Available online: https://ourworldindata.org/less-meat-or-sustainable-meat#article-citation(accessed on 27 November 2023).
- Oberle, B.; Bringezu, S.; Hatfield-Dodds, S.; Hellweg, S.; Schandl, H.; Clement, J. Global Resources Outlook; International Resource Panel, United Nations Envio: Paris, France, 2019. [Google Scholar]
- Girgenti, V.; Peano, C.; Baudino, C.; Tecco, N. From “farm to fork” strawberry system: Current realities and potential innovative scenarios from life cycle assessment of non-renewable energy use and greenhouse gas emissions. Sci. Total Environ. 2014, 473, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Sarandón, S.; Flores, C. L’agroécologie: Un nouveau paradigme pour l’élaboration du développement rural durable. In L’Agroécologie en Argentine et en France. Regards Croisés; L’Harmattan: Paris, France, 2012; pp. 91–120. [Google Scholar]
- Baret, P.; Stassart, P.M.; Vanloqueren, G.; Van Damme, J. Dépasser les verrouillages de régimes socio-techniques des systèmes alimentaires pour construire une transition agroécologique. In Actes du Premier Congrès Interdisciplinaire sur le Développement Durable: Quelle Transition Pour Nos Sociétés? Thème 2: Alimentation, Agriculture, Élevage; Université Catholique de Louvain: Namur, Belgium, 2013; pp. 5–14. [Google Scholar]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Moebius-Clune, B.; Van Es, H.; Idowu, O.; Schindelbeck, R.; Kimetu, J.; Ngoze, S.; Lehmann, J.; Kinyangi, J. Long-term soil quality degradation along a cultivation chronosequence in western Kenya. Agric. Ecosys Environ. 2011, 141, 86–99. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef]
- Bachir, A.S.; Hafner, H.; Tourenq, J.N.; Doumandji, S. Structure de l’habitat et biologie de reproduction du Héron garde-bœuf, Bubulcus ibis, dans une colonie de la vallée de la Soummam (Petite Kabylie, Algérie). Rev. Ecol. Terre Et Vie 2000, 55, 33–43. [Google Scholar] [CrossRef]
- Sheng, D.; Fredlund, D.G.; Gens, A. A new modelling approach for unsaturated soils using independent stress variables. Can. Geotech. J. 2008, 45, 511–534. [Google Scholar] [CrossRef]
- Cannavacciulo, M.; Cassagne, N.; Riou, V.; Mulliez, P.; Prévost-Bouré, N.C.; Dequiedt, S.; Villenave, C.; Cérémonie, H.; Cluzeau, D.; Cylly, D. Validation d’un tableau de bord d’indicateurs sur un réseau national de fermes en grande culture et en viticulture pour diagnostiquer la qualité biologique des sols agricoles. Innov. Agron. 2017, 55, 41–54. [Google Scholar] [CrossRef]
- Tóth, B.; Weynants, M.; Nemes, A.; Makó, A.; Bilas, G.; Tóth, G. New generation of hydraulic pedotransfer functions for Europe. Eur. J. Soil Sci. 2015, 66, 226–238. [Google Scholar] [CrossRef]
- Tecon, R.; Or, D. Biophysical processes supporting the diversity of microbial life in soil. FEMS Microbiol. Rev. 2017, 41, 599–623. [Google Scholar] [CrossRef] [PubMed]
- Lehman, R.M.; Acosta-Martinez, V.; Buyer, J.S.; Cambardella, C.A.; Collins, H.P.; Ducey, T.F.; Halvorson, J.J.; Jin, V.L.; Johnson, J.M.F.; Kremer, R.J.; et al. Soil biology for resilient, healthy soil. J. Soil Water Conserv. 2015, 70, 12A–18A. [Google Scholar] [CrossRef]
- Calvaruso, C.; Blanchart, A.; Bertin, S.; Grand, C.; Pierart, A.; Eglin, T. Quels paramètres du sol mesurer pour évaluer les fonctions et les services écosystémiques associés? Étud. Gest. Sols 2021, 28, 3–29. [Google Scholar]
- Lal, R. Restorative effects of Mucuna utilis on soil organic C pool of a severely degraded Alfisol in western Nigeria. In Global Climate Change and Tropical Ecosystems; CRC Press: Boca Raton, FL, USA, 2019; pp. 147–156. [Google Scholar]
- Abrahams, P.W. Soils: Their implications to human health. Sci. Total Environ. 2002, 291, 1–32. [Google Scholar] [CrossRef]
- Drake, T.W.; Podgorski, D.C.; Dinga, B.; Chanton, J.P.; Six, J.; Spencer, R.G. Land-use controls on carbon biogeochemistry in lowland streams of the Congo Basin. Glob. Chang. Biol. 2020, 26, 1374–1389. [Google Scholar] [CrossRef] [PubMed]
- Mama, A.; Sinsin, B.; De Cannière, C.; Bogaert, J. Anthropisation et dynamique des paysages en zone soudanienne au nord du Bénin. Tropicultura 2013, 31, 78–88. [Google Scholar]
- Lal, R. Soil degradation as a reason for inadequate human nutrition. Food Secur. 2009, 1, 45–57. [Google Scholar] [CrossRef]
- Lahmar, R.; Ruellan, A. Dégradation des sols et stratégies coopératives en Méditerranée: La pression sur les ressources naturelles et les stratégies de développement durable. Cah. Agric. 2007, 16, 318–323. [Google Scholar] [CrossRef]
- Schulte, R.P.; Bampa, F.; Bardy, M.; Coyle, C.; Creamer, R.E.; Fealy, R.; Gardi, C.; Ghaley, B.B.; Jordan, P.; O’Donoghue, C.; et al. Making the most of our land: Managing soil functions from local to continental scale. Front. Environ. Sci. 2015, 3, 81. [Google Scholar] [CrossRef]
- Krupenikov, I.A.; Boincean, B.P.; Dent, D. Humus–Guardian of Fertility and Global Carbon Sink. In The Black Earth; Springer: Dordrecht, The Netherlands, 2011; pp. 39–50. [Google Scholar]
- Frossard, P.-A.; Évette, A. Le génie végétal pour la lutte contre l’érosion en rivière: Une tradition millénaire en constante évolution. Sci. Eaux. Territ. (Spécial Ingénieries-EAT) 2009, 29, 99–109. [Google Scholar]
- Aïdoud, A.; Le Floc’h, É.; Le Houérou, H.N. Les steppes arides du nord de l’Afrique. Sci. Chang. Planét/Sécher. 2006, 17, 19–30. [Google Scholar]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Karlen, D.L.; Mausbach, M.; Doran, J.W.; Cline, R.; Harris, R.; Schuman, G. Soil quality: A concept, definition, and framework for evaluation (a guest editorial). Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef]
- Dabre, A.; Hien, E.; Some, D.; Drevon, J.J. Effets d’amendements organiques et phosphatés sous zaï sur les propriétés chimiques et biologiques du sol et la qualité de la matière organique en zone soudano-sahélienne du Burkina Faso. Int. J. Biol. Chem. Sci. 2017, 11, 473–487. [Google Scholar] [CrossRef]
- Bennett, T. Culture: A Reformer’s Science; Sage Publications Ltd.: Thousand Oaks, CA, USA, 1998. [Google Scholar]
- Futughe, A.E.; Purchase, D.; Jones, H. Phytoremediation using native plants. In Phytoremediation: In-Situ Applications; Springer: Cham, Switzerland, 2020; pp. 285–327. [Google Scholar]
- Lee, J.; Kaunda, R.B.; Sinkala, T.; Workman, C.F.; Bazilian, M.D.; Clough, G. Phytoremediation and phytoextraction in Sub-Saharan Africa: Addressing economic and social challenges. Ecotoxicol. Environ. Saf. 2021, 226, 112864. [Google Scholar] [CrossRef]
- Colombet, I. Revue systématique et méta-analyse en médecine palliative. Médecine Palliat. Soins De Support-Accompagnement-Ethique 2015, 14, 240–253. [Google Scholar] [CrossRef]
- Maison, P. La méta-analyse sur données résumées. Rech. En Soins Infirm. 2010, 101, 18–24. [Google Scholar] [CrossRef]
- Beaucher, V.; Jutras, F. Étude comparative de la métasynthèse et de la méta-analyse qualitative. Rech. Qual. 2007, 27, 58–77. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Intern. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Michéli, E.; Fuchs, M.; Hegymegi, P.; Stefanovits, P. Classification of the major soils of Hungary and their correlation with the World Reference Base for Soil Resources (WRB). Agrokémia És Talajt. 2006, 55, 19–28. [Google Scholar] [CrossRef]
- Cavalli, J.P.; Reichert, J.M.; Rodrigues, M.F.; de Araújo, E.F. Composition and functional soil properties of Arenosols and Acrisols: Effects on eucalyptus growth and productivity. Soil Tillage Res. 2020, 196, 104439. [Google Scholar] [CrossRef]
- Ilstedt, U.; Nordgren, A.; Malmer, A. Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biol. Biochem. 2000, 32, 1591–1599. [Google Scholar] [CrossRef]
- Mwamba, S.; Kaluba, P.; Moualeu-Ngangue, D.; Winter, E.; Chiona, M.; Chishala, B.H.; Munyinda, K.; Stützel, H. Physiological and morphological responses of cassava genotypes to fertilization regimes in chromi-haplic acrisols soils. Agron. J. 2021, 11, 1757. [Google Scholar] [CrossRef]
- IUSS Working Group, W.R.B. World Reference Base for Soil Resources 2006, 2nd ed.; World Soil Resources Reports, No. 103; FAO: Rome, Italy, 2006; 145p. [Google Scholar]
- Cueto, O.G.; Coronel, C.E.I.; Bravo, E.L.; Morfa CA, R.; Suárez, M.H. Modelling in FEM the soil pressures distribution caused by a tyre on a Rhodic Ferralsol soil. J. Terramech. 2016, 63, 61–67. [Google Scholar] [CrossRef]
- Dick, D.P.; Gonçalves, C.N.; Dalmolin, R.S.; Knicker, H.; Klamt, E.; Kögel-Knabner, I.; Simões, M.L.; Martin-Neto, L. Characteristics of soil organic matter of different Brazilian Ferralsols under native vegetation as a function of soil depth. Geoderma 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Madari, B.E.; Reeves, J.B., III; Machado, P.L.; Guimarães, C.M.; Torres, E.; McCarty, G.W. Mid-and near-infrared spectroscopic assessment of soil compositional parameters and structural indices in two Ferralsols. Geoderma 2006, 136, 245–259. [Google Scholar] [CrossRef]
- Özcan, H. Fluvisols. In The Soils of Turkey; Springer: Cham, Switzerland, 2018; pp. 129–137. [Google Scholar]
- Rodrigo-Comino, J.; Keshavarzi, A.; Senciales-González, J.M. Evaluating soil quality status of fluvisols at the regional scale: A multidisciplinary approach crossing multiple variables. River Res. Appl. 2023, 39, 1367–1381. [Google Scholar] [CrossRef]
- Dawoe, E.K.; Quashie-Sam, J.S.; Oppong, S.K. Effect of land-use conversion from forest to cocoa agroforest on soil characteristics and quality of a Ferric Lixisol in lowland humid Ghana. Agrofor. Syst. 2014, 88, 87–99. [Google Scholar] [CrossRef]
- Oorts, K.; Vanlauwe, B.; Merckx, R. Cation exchange capacities of soil organic matter fractions in a Ferric Lixisol with different organic matter inputs. Agric. Ecosyst. Environ. 2003, 100, 161–171. [Google Scholar] [CrossRef]
- Ouattara, K.; Ouattara, B.; Assa, A.; Sédogo, P.M. Long-term effect of ploughing, and organic matter input on soil moisture characteristics of a Ferric Lixisol in Burkina Faso. Soil Tillage Res. 2006, 88, 217–224. [Google Scholar] [CrossRef]
- Araújo, M.S.; Schaefer, C.E.; Sampaio, E.V. Soil phosphorus fractions from toposequences of semi-arid Latosols and Luvisols in northeastern Brazil. Geoderma 2004, 119, 309–321. [Google Scholar] [CrossRef]
- Dyck, M.F.; Sorenson, P.T.; Lejoly, J.D.; Quideau, S.A. Gray Luvisols are polygenetic. Can. J. Soil Sci. 2022, 103, 121–133. [Google Scholar] [CrossRef]
- Walmsley, A.; Azadi, H.; Tomeckova, K.; Sklenicka, P. Contrasting effects of land tenure on degradation of Cambisols and Luvisols: The case of Central Bohemia Region in the Czech Republic. Land Use Policy 2020, 99, 104956. [Google Scholar] [CrossRef]
- Agegnehu, G.; Nelson, P.N.; Bird, M.I. Crop yield, plant nutrient uptake and soil physicochemical properties under organic soil amendments and nitrogen fertilization on Nitisols. Soil Tillage Res. 2016, 160, 1–13. [Google Scholar] [CrossRef]
- Cooper, M.; Vidal-Torrado, P.; Chaplot, V. Origin of microaggregates in soils with ferralic horizons. Sci. Agric. 2005, 62, 256–263. [Google Scholar] [CrossRef]
- Dorel, M.; Roger-Estrade, J.; Manichon, H.; Delvaux, B. Porosity and soil water properties of Caribbean volcanic ash soils. Soil Use Manag. 2000, 16, 133–140. [Google Scholar] [CrossRef]
- Gura, I.; Mnkeni, P.; Du Preez, C.; Barnard, J. Short-term effects of conservation agriculture strategies on the soil quality of a Haplic Plinthosol in Eastern Cape, South Africa. Soil Tillage Res. 2022, 220, 105378. [Google Scholar] [CrossRef]
- Ikazaki, K.; Nagumo, F.; Simporé, S.; Barro, A. Soil toposequence, productivity, and a simple technique to detect petroplinthites using ground-penetrating radar in the Sudan Savanna. Soil Sci. Plant Nutr. 2018, 64, 623–631. [Google Scholar] [CrossRef]
- Wildemeersch, J.C.; Garba, M.; Sabiou, M.; Sleutel, S.; Cornelis, W. The effect of water and soil conservation (WSC) on the soil chemical, biological, and physical quality of a Plinthosol in Niger. Land Degrad. Dev. 2015, 26, 773–783. [Google Scholar] [CrossRef]
- Bhattacharyya, T.; Pal, D.; Deshpande, S. Genesis and transformation of minerals in the formation of red (Alfisols) and black (Inceptisols and Vertisols) soils on Deccan basalt in the Western Ghats, India. J. Soil Sci. 1993, 44, 159–171. [Google Scholar] [CrossRef]
- Kurtzman, D.; Baram, S.; Dahan, O. Soil–aquifer phenomena affecting groundwater under vertisols: A review. Hydrol. Earth Syst. Sci. 2016, 20, 1–12. [Google Scholar] [CrossRef]
- Doamba, S.M.F.; Nacro, H.B.; Sanon, A.; Sedogo, M. Effet des cordons pierreux sur l’activité biologique d’un sol ferrugineux tropical lessivé (Province du Kouritenga au Burkina Faso). Int. J. Biol. Chem. Sci. 2011, 5, 304–313. [Google Scholar] [CrossRef]
- Soulama, S.; Kadeba, A.; Nacoulma, B.M.; Traoré, S.; Bachmann, Y.; Thiombiano, A. Impact des activités anthropiques sur la dynamique de la végétation de la réserve partielle de faune de Pama et de ses périphéries (sud-est du Burkina Faso) dans un contexte de variabilité climatique. J. Appl. Biosci. 2015, 87, 8047–8064. [Google Scholar] [CrossRef]
- Diallo, H.; Bamba, I.; Barima, Y.S.S.; Visser, M.; Ballo, A.; Mama, A.; Vranken, I.; Maiga, M.; Bogaert, J. Effets combinés du climat et des pressions anthropiques sur la dynamique évolutive de la végétation d’une zone protégée du Mali (Réserve de Fina, Boucle du Baoulé). Sci. Chang. Planét/Sécher. 2011, 22, 97–107. [Google Scholar] [CrossRef]
- Coulibaly, N.D.; Kone, M.; Koné, T.; Hilairekouakou, T.; Kouadio, Y.J. Influence des conditions pédo-climatiques sur les composantes du rendement de Jatropha curcas L. dans les zones Nord de savane et Sud forestière de la Côte d’Ivoire. Int. J. Biol. Chem. Sci. 2015, 9, 2346–2361. [Google Scholar] [CrossRef]
- Clark, K.H.; Nicholas, K.A. Introducing urban food forestry: A multifunctional approach to increase food security and provide ecosystem services. Landsc. Ecol. 2013, 28, 1649–1669. [Google Scholar] [CrossRef]
- Benabdoun, M.; Gherbi, H.; Djekoun, A.; Bogusz, D.; Franche, C.; Ykhlef, N. Fixation biologique de l’azote: La symbiose actinorhizienne Casuarina-Frankia. Sci. Technol. C Biotechnol. 2012, 35, 15–19. [Google Scholar]
- Duponnois, R.; Plenchette, C.; Bâ, A.M. Growth stimulation of seventeen fallow leguminous plants inoculated with Glomus aggregatum in Senegal. Eur. J. Soil Biol. 2001, 37, 181–186. [Google Scholar] [CrossRef]
- Hocher, V.; Auguy, F.; Bogusz, D.; Doumas, P.; Franche, C.; Gherbi, H.; Laplaze, L.; Obertello, M.; Svistoonoff, S. Les symbioses actinorhiziennes fixatrices d’azote: Un exemple d’adaptation aux contraintes abiotiques du sol. Cah. Agric. 2009, 18, 498–505. [Google Scholar] [CrossRef]
- Benbrahim, K.F.; Ismaili, M.; Benbrahim, S.F.; Tribak, A. Land degradation by desertification and deforestation in Morocco. Sci. Chang. Planét/Sécher. 2004, 15, 307–320. [Google Scholar]
- Benbrahim, K.F.; Berrada, H.; El Ghachtouli, N.; Ismaili, M. Les acacias: Des plantes fixatrices d’azote prometteuses pour le développement durable des zones arides et semi-arides [Acacia: Promising Nitrogen fixing trees for sustainable development in arid and semi-arid areas]. Int. J. Innov. Appl. Studies 2014, 8, 46. [Google Scholar]
- Amadji, G.; Aholoukpè, H. Impact du niébé (Vigna unguiculata) et de la fumure minérale sur les propriétés chimiques de la terre de barre du Bénin. Étud. Gest. Sols 2008, 15, 147–160. [Google Scholar]
- Bado, B.V. Rôle des Légumineuses sur la Fertilité des Sols Ferrugineux Tropicaux des Zones Guinéenne et Soudanienne du Burkina Faso. Ph.D. Thesis, Université de Laval, Québec, QC, Canada, 2002; 166p. [Google Scholar]
- Azontonde, A.; Feller, C.; Ganry, F.; Remy, J.C. La mucuna et la restauration des propriétés d’un sol ferrallitique au Sud du Bénin. Plante de couverture. Agric. Dev. 1998, 18, 55–62. [Google Scholar]
- Dommergues, Y. La fixation d’azote chez les plantes actinorhiziennes et ses applications. Acta Bot. Gall. 1996, 143, 663–679. [Google Scholar] [CrossRef]
- Balesdent, J.; Basile-Doelsch, I.; Chadoeuf, J.; Cornu, S.; Fekiacova, Z.; Fontaine, S.; Guenet, B.; Hatté, C. Renouvellement du carbone profond des sols cultivés: Une estimation par compilation de données isotopiques. Biotechnol. Agron. Soc. Environ. 2016, 21, 181–190. [Google Scholar] [CrossRef]
- Balesdent, J.; Derrien, D.; Fontaine, S.; Kirman, S.; Klumpp, K.; Loiseau, P.; Marol, C.; Nguyen, C.; Péan, M.; Personeni, E.; et al. Contribution de la rhizodéposition aux matières organiques du sol, quelques implications pour la modélisation de la dynamique du carbone. Étud Gest. Sols 2011, 18, 201–216. [Google Scholar]
- Jones, D.L.; Nguyen, C.; Finlay, R.D. Carbon flow in the rhizosphere: Carbon trading at the soil–root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Lal, R. Carbon sequestration. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 815–830. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions–A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon dynamics in cropland and rangeland. Environ. Pollut. 2002, 116, 353–362. [Google Scholar] [CrossRef]
- Johnson, N.C.; Hoeksema, J.D.; Bever, J.D.; Chaudhary, V.B.; Gehring, C.; Klironomos, J.; Koide, R.T.; Michael Miller, R.; Moore, J.; Moutoglis, P.; et al. From Lilliput to Brobdingnag: Extending models of mycorrhizal function across scales. Bioscience 2006, 56, 889–900. [Google Scholar] [CrossRef]
- Lal, R.; Bruce, J. The potential of world cropland soils to sequester C and mitigate the greenhouse effect. Environ. Sci. Policy 1999, 2, 177–185. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-M atrix S tabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.; De Jong, R. Root-to-straw ratios-influence of moisture and rate of N fertilizer. Can. J. Soil Sci. 2001, 81, 39–43. [Google Scholar] [CrossRef]
- Poorter, H.; Jagodzinski, A.M.; Ruiz-Peinado, R.; Kuyah, S.; Luo, Y.; Oleksyn, J.; Usoltsev, V.A.; Buckley, T.N.; Reich, P.B.; Sack, L. How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continents. New Phytol. 2015, 208, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.; Diaz, R.A.; Barbero, N.; Santanatoglia, O.J.; Blotta, L. Soil organic carbon, microbial biomass, and CO2-C pro- duction from three tillage systems. Soil Tillage Res. 1995, 33, 17–28. [Google Scholar] [CrossRef]
- Poirier, V.; Roumet, C.; Angers, D.A.; Munson, A.D. Species, and root traits impact macroaggregation in the rhizo spheric soil of a Mediterranean common garden experiment. Plant Soil 2018, 424, 289–302. [Google Scholar] [CrossRef]
- Derrien, D.; Dignac, M.; Basile-Doelsch, I.; Barot, S.; Cécillon, L.; Chenu, C.; Chevallier, T.; Freschet, G.T.; Garnier, P.; Guenet, B.; et al. Stocker du C dans les sols: Quels mécanismes, quelles pratiques agricoles, quels indicateurs? Étud. Gest. Sols 2016, 23, 193–224. [Google Scholar]
- Kang, B.; Caveness, F.; Tian, G.; Kolawole, G. Longterm alley cropping with four hedgerow species on an Alfisol in southwestern Nigeria–effect on crop performance, soil chemical properties and nematode population. Nutr. Cycl. Agroecosyst. 1999, 54, 145–155. [Google Scholar] [CrossRef]
- Crow, S.E.; Reeves, M.; Turn, S.; Taniguchi, S.; Schubert, O.S.; Koch, N. Carbon balance implications of land use change from pasture to managed eucalyptus forest in Hawaii. Carbon Manag. 2016, 7, 171–181. [Google Scholar] [CrossRef]
- Razakamanarivo, R.H.; Razafindrakoto, M.A.; Albrecht, A. Fonction puits de carbone des taillis d’eucalyptus à Madagascar. Bois Forets Trop. 2010, 305, 5–19. [Google Scholar] [CrossRef]
- Alegre, J.; Rao, M. Soil and water conservation by contour hedging in the humid tropics of Peru. Agric. Ecosyst. Environ. 1996, 57, 17–25. [Google Scholar] [CrossRef]
- Razafimbelo, T.; Barthes, B.; De Luca, E.; Larré-Larrouy, M.-C.; Laurent, J.; Cerri, C.; Feller, C. Effet du paillis des résidus de canne à sucre sur la séquestration de carbone dans un sol ferrallitique argileux du Bresil. Étud. Gest. Sols 2003, 10, 191–200. [Google Scholar]
- Graham, M.; Haynes, R.; Meyer, J. Soil organic matter content and quality: Effects of fertilizer applications, burning and trash retention on a long-term sugarcane experiment in South Africa. Soil Biol. Biochem. 2002, 34, 93–102. [Google Scholar] [CrossRef]
- Wani, S.P.; Chander, G.; Sahrawat, K.L.; Rao, C.S.; Raghvendra, G.; Susanna, P.; Pavani, M. Carbon sequestration and land rehabilitation through Jatropha curcas (L.) plantation in degraded lands. Agric. Ecosyst. Environ. 2012, 161, 112–120. [Google Scholar] [CrossRef]
- Diédhiou, I.; Diallo, D.; Mbengue, A.; Hernandez, R.; Bayala, R.; Diémé, R.; Diédhiou, P.M.; Sène, A. Allometric equations and carbon stocks in tree biomass of Jatropha curcas L. in Senegal’s Peanut Basin. Glob. Ecol. Conserv. 2017, 9, 61–69. [Google Scholar] [CrossRef]
- Recous, S.; Lashermes, G.; Bertrand, I. Couplages et contrôles des cycles du carbone et de l’azote par les communautés microbiennes dans les sols cultivés. In Les Sols et la Vie Souterraine: Des Enjeux Majeurs en Agroécologie; Briat, J.-F., Job, D., Eds.; Éditions Quae: Versaille, France, 2017; pp. 43–62. [Google Scholar]
- Janssen, B.H.; Guiking, F.; van der Eijk, D.; Smaling, E.M.; Wolf, J.; van Reuler, H. A system for quantitative evaluation of the fertility of tropical soils (QUEFTS). Geoderma 1990, 46, 299–318. [Google Scholar] [CrossRef]
- Bonneau, M. Evolution of the mineral fertility of an acidic soil during a period of ten years in the Vosges mountains (France). Impact of humus mineralisation. Ann. For. Sci. 2005, 62, 253–260. [Google Scholar] [CrossRef]
- Yan, J.; Wang, L.; Hu, Y.; Tsang, Y.F.; Zhang, Y.; Wu, J.; Fu, X.; Sun, Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma 2018, 319, 194–203. [Google Scholar] [CrossRef]
- Ouattara, B.; Coulibaly, K.; Kohio, E.; Doumbia, S.; Ouédraogo, S.; Nacro, H.B. Effets du Système de Culture sous couverture Végétale (SCV) sur les flux hydriques d’un sol ferrugineux à l’Ouest du Burkina Faso. Int. J. Biol. Chem. Sci. 2018, 12, 1770–1783. [Google Scholar] [CrossRef]
- Njoukam, R.; Oliver, R.; Peltier, R. Restitution minérale au sol par la litière dans les plantations de Pinus kesiya Royle ex-Gordon dans l’ouest du Cameroun. Ann. For. Sci. 1999, 56, 431–439. [Google Scholar] [CrossRef]
- Cardoso, I.M.; Kuyper, T.W. Mycorrhizas and tropical soil fertility. Agric. Ecosyst. Environ. 2006, 116, 72–84. [Google Scholar] [CrossRef]
- Altieri, M.A. Agroecology: The science of natural resource management for poor farmers in marginal environments. Agric. Ecosyst. Environ. 2002, 93, 1–24. [Google Scholar] [CrossRef]
- Ewel, J.J. Natural systems as models for the design of sustainable systems of land use. Agrofor. Syst. 1999, 45, 1–21. [Google Scholar] [CrossRef]
- Roose, É. Évolution historique des stratégies de lutte antiérosive. Vers la gestion conservatoire de l ‘eau, de la biomasse et de la fertilité des sols (GCES). Sci. Chang. Planét/Sécher. 2004, 15, 9–18. [Google Scholar]
- Hugron, S.; Bussières, J.; Rochefort, L. Plantations d’Essences Forestières Dans le Contexte de la Restauration Écologique des Tourbières: Un Guide Pratique; Groupe de Recherche en Écologie des Tourbières, Université Laval: Québec, QC, Canada, 2011; 88p. [Google Scholar]
- Seigler, D.S. Phytochemistry of Acacia—Sensu lato. Biochem. Syst. Ecol. 2003, 31, 845–873. [Google Scholar] [CrossRef]
- Araujo, J.H.R. Rôle de la Diversité des Vers de Terre Dans le Fonctionnement des Sols: Application à l’Ingénierie Pédologique. Ph.D. Thesis, Université Paris-Est Créteil Val-de-Marne-Paris 12, Créteil, France, 2022; 175p. [Google Scholar]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Poirier, V.; Angers, D.A.; Whalen, J.K. Formation of millimetric-scale aggregates and associated retention of 13C–15N-labelled residues are greater in subsoil than topsoil. Soil Biol. Biochem. 2014, 75, 45–53. [Google Scholar] [CrossRef]
- Martens, D.A. Plant residue biochemistry regulates soil carbon cycling and carbon sequestration. Soil Biol. Biochem. 2000, 32, 361–369. [Google Scholar] [CrossRef]
- Annabi, M.; Haithem, B. Influence des racines sur la stabilité structurale d’un sol du Nord de la Tunisie. Étud. Gest. Sols 2011, 18, 63–74. [Google Scholar]
- Gyssels, G.; Poesen, J.; Bochet, E.; Li, Y. Impact of plant roots on the resistance of soils to erosion by water: A review. Prog. Phys. Geogr. 2005, 29, 189–217. [Google Scholar] [CrossRef]
- Abiven, S.; Menasseri, S.; Chenu, C. The effects of organic inputs over time on soil aggregate stability–A literature analysis. Soil Biol. Biochem. 2009, 41, 1–12. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C. Rhizodeposition of organic C by plant: Mechanisms and controls. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 97–123. [Google Scholar]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Bodner, G.; Leitner, D.; Kaul, H.-P. Coarse and fine rooted plant species have distinct effect on soil pore size distribution. Plant Soil 2014, 380, 133–151. [Google Scholar] [CrossRef]
- Kraus, T.E.; Dahlgren, R.A.; Zasoski, R.J. Tannins in nutrient dynamics of forest ecosystems-a review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Bearden, B.N. Influence of arbuscular mycorrhizal fungi on soil structure and soil water characteristics of vertisols. Plant Soil 2001, 229, 245–258. [Google Scholar] [CrossRef]
- Bearden, B.N.; Petersen, L. Influence of arbuscular mycorrhizal fungi on soil structure and aggregate stability of a vertisol. Plant Soil 2000, 218, 173–183. [Google Scholar] [CrossRef]
- Haynes, R.; Beare, M. Influence of six crop species on aggregate stability and some labile organic matter fractions. Soil Biol. Biochem. 1997, 29, 1647–1653. [Google Scholar] [CrossRef]
- Erktan, A.; Cécillon, L.; Graf, F.; Roumet, C.; Legout, C.; Rey, F. Increase in soil aggregate stability along a Mediterranean successional gradient in severely eroded gully bed ecosystems: Combined effects of soil, root traits and plant community characteristics. Plant Soil 2016, 398, 121–137. [Google Scholar] [CrossRef]
- Newsham, K.; Fitter, A.; Watkinson, A. Multi-functionality and biodiversity in arbuscular mycorrhizas. Trends Ecol. Evol. 1995, 10, 407–411. [Google Scholar] [CrossRef]
- Degens, B.; Sparling, G.; Abbott, L. Increasing the length of hyphae in a sandy soil increases the amount of water-stable aggregates. Appl. Soil Ecol. 1996, 3, 149–159. [Google Scholar] [CrossRef]
- Ogunwole, J.; Chaudhary, D.; Ghosh, A.; Daudu, C.; Chikara, J.; Patolia, J. Contribution of Jatropha curcas to soil quality improvement in a degraded Indian entisol. Acta Agric. Scand. Sect. B 2008, 58, 245–251. [Google Scholar] [CrossRef]
- Rey, F.; Ballais, J.-L.; Marre, A.; Rovéra, G. Rôle de la végétation dans la protection contre l’érosion hydrique de surface. Comptes Rendus Geosci. 2004, 336, 991–998. [Google Scholar] [CrossRef]
- Sidle, R.C.; Ziegler, A.D.; Negishi, J.N.; Nik, A.R.; Siew, R.; Turkelboom, F. Erosion processes in steep terrain—Truths, myths, and uncertainties related to forest management in Southeast Asia. For. Ecol. Manag. 2006, 224, 199–225. [Google Scholar] [CrossRef]
- Wondzell, S.M.; King, J.G. Postfire erosional processes in the Pacific Northwest and Rocky Mountain regions. For. Ecol. Manag. 2003, 178, 75–87. [Google Scholar] [CrossRef]
- Vennetier, M.; Ladier, J.; Rey, F. Le contrôle de l’érosion des sols forestiers par la végétation face aux changements globaux. Rev. For. Française 2014, 4, 517–530. [Google Scholar] [CrossRef]
- Lamers, J.; Feil, P. Farmers’ knowledge and management of spatial soil and crop growth variability in Niger, West Africa. Neth. J. Agric. Sci. 1995, 43, 375–389. [Google Scholar] [CrossRef]
- Cam, C.; Froger, D.; Moulin, J.; Rassineux, J.; Servant, J. Representation cartographique de la sensibilite des sols l’infiltration hydrique verticale. Étud. Gest. Sols 1996, 3, 97–112. [Google Scholar]
- Mazour, M.; Roose, E. Influence de la couverture végétale sur le ruissellement et l’érosion des sols sur parcelles d’érosion dans les bassins versants du Nord-ouest de l’Algérie. Bull. Réseau Erosion 2002, 21, 320–330. [Google Scholar]
- Morsli, B.; Mazour, M.; Mededjel, N.; Hamoudi, A.; Roose, E. Influence de l’utilisation des terres sur les risques de ruissellement et d’érosion sur les versants semi-arides du nord-ouest de l’Algérie. Sci. Chang. Planét/Sécher. 2004, 15, 96–104. [Google Scholar]
- Berkane, A.; Yahiaou, A. L’érosion dans les Aurès. Sci. Chang. Planét/Sécher. 2007, 18, 213–216. [Google Scholar]
- Douzet, J.-M.; Scopel, E.; Muller, B.; Rakotoarisoa, J.; Albrecht, A.; Razafindramanana, N. Effets des systèmes de cultures en semis direct avec couverture végétale sur le ruissellement et l’érosion des cultures pluviales des Hautes Terres de Madagascar. Étud. Gest. Sols 2010, 17, 131–142. [Google Scholar]
- Garg, K.K.; Karlberg, L.; Wani, S.P.; Berndes, G. Jatropha production on wastelands in India: Opportunities and trade-offs for soil and water management at the watershed scale. Biofuels Bioprod. Biorefining 2011, 5, 410–430. [Google Scholar] [CrossRef]
- Ouattara, V.T.; Konate, Z.; Messoum, G.F.; Kassin, E.K.; Tahi, M.G.; Koko, L.A.; Kone, B.; Dick, B.A.; et Camara, M. Effets de la fertilisation organo-phosphatée sur la fertilité en matière organique et complexe adsorbant d’un ferralsol sous cacaoyers dans la région de Divo (Côte d’Ivoire). Int. J. Biol. Chem. Sci. 2018, 12, 2901–2921. [Google Scholar] [CrossRef]
- Bielders, C.L.; Michels, K.; Rajot, J.-L. On-farm evaluation of ridging and residue management practices to reduce wind erosion in Niger. Soil Sci. Soc. Am. J. 2000, 64, 1776–1785. [Google Scholar] [CrossRef]
- Michels, K.; Sivakumar, M.; Allison, B. Wind erosion control using crop residue I. Effects on soil flux and soil properties. Field Crops Res. 1995, 40, 101–110. [Google Scholar] [CrossRef]
- Tidjani, A.D.; Ambouta, K.; Bielders, C. Réhabilitation d’une dune vive par fixation mécanique: Flux éoliens, fertilité du sol et biodiversité des herbacées. Geo-Eco-Trop 2009, 33, 81–98. [Google Scholar]
- Danjon, F.; Bert, D.; Godin, C.; Trichet, P. Structural root architecture of 5-year-old Pinus pinaster measured by 3D digitising and analysed with AMAPmod. Plant Soil 1999, 217, 49–63. [Google Scholar] [CrossRef]
- Mouisat, A.; Harrouni, C.; Chaouki, A.; Douaik, A.; Derradji, A.; Tlemcani, N.-E.B. Efficacité des méthodes du génie biologique pour lutter contre l’érosion hydrique des talus marneux de l’autoroute Fès-Taza. Rev. Maroc. Sci. Agron. Vét. 2020, 8, 67–77. [Google Scholar]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- Kante, M. Rhizodéposition du Carbone et de l’Azote Chez Quatre Espèces de Fabacées, Conséquences sur la Structure et l’Activité du Microbiote du sol. Ph.D. Thesis, Université de Normandie, Caen, France, 2022; 395p. [Google Scholar]
- Maj, D.; Wielbo, J.; Marek-Kozaczuk, M.; Skorupska, A. Response to flavonoids as a factor influencing competitiveness and symbiotic activity of Rhizobium leguminosarum. Microbiol. Res. 2010, 165, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Azib, S. La Symbiose Rhizobium-Luzerne: Étude de la Diversité Rhizobienne et Essais d’Inoculation en vue d’Améliorer les Rendements en Fourrage Dans le Sahara Septentrional Est-Algérien. Ph.D. Thesis, Université Kasdi Merbah Ouargla, Ouargla, Algeria, 2020; 143p. [Google Scholar]
- Girard, M.C.; Walter, C.; Rémy, J.C.; Berthelin, J.; Morel, J.L. Sols et Environnement-2e Édition—Cours, Exercices et Études de Cas; Dunod: Malakoff, France, 2011. [Google Scholar]
- Gobat, J.M.; Aragno, M.; Matthey, W. Le sol Vivant: Bases de Pédologie, Biologie des Sols-3e Édition; EPFL Press: Lausanne, Switzerland, 2010; 848p. [Google Scholar]
- Barrière, Y. Le maïs et l’eau: Une situation aujourd’hui paradoxale, mais des progrès génétiques à attendre d’un idéotype redéfini. Fourrages 2001, 168, 477–489. [Google Scholar]
- Kabongo Tshiabukole, J.P. Evaluation de la Sensibilité aux Stress Hydriques du Maïs (Zea mays L.) Cultivé Dans la Savane du Sud-Ouest de la RD Congo, cas de Mvuazi. Ph.D. Thesis, Université Pédagogique Nationale, Kinshasa, Democratic Republic of the Congo, 2018; 162p. [Google Scholar]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Ntenga, R. Modélisation Multi-Échelle et Caractérisation de l’Anisotropie Élastique de Fibres Végétales pour le Renforcement de Matériaux Composites. Ph.D. Thesis, Université Blaise Pascal, Clermont-Ferrand, France, Université de Yaoundé, Yaoundé, Cameroon, 2007; 185p. [Google Scholar]
- Razanamalala, K. Priming Effect: Vers un Outil de Gestion de la Fertilité des Sols Cultivés à Madagascar. Ph.D. Thesis, Montpellier SupAgro, Montpellier, France, Université d’Antananarivo, Antananarivo, Madagascar, 2017; 212p. [Google Scholar]
- Germon, A. Conséquences de la coupe rase sur la production de racines fines, CO2, CH4 et N2O jusqu’à la nappe phréatique dans une plantation d’Eucalyptus grandis menée en taillis sur un dispositif d’exclusion de pluie. Bois For. Trop. 2020, 346, 79–80. [Google Scholar] [CrossRef]
- Pradier, C. Rôles Fonctionnels des Racines Fines Profondes en Plantation d’Eucalyptus au Brésil sur Sols Pauvres en Nutriments. Réponse à une Situation Hydrique Limitante. Ph.D. Thesis, Montpellier SupAgro, Montpellier, France, 2016; 235p. [Google Scholar]
- Chew, M.K. The monstering of tamarisk: How scientists made a plant into a problem. J. Hist. Biol. 2009, 42, 231–266. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the methodological approach used to select articles in this critical scoping review.
Figure 1.
Schematic representation of the methodological approach used to select articles in this critical scoping review.

Figure 2.
Characteristics and mechanisms of plants species that play a positive role in the restoration of degraded tropical soil. See Table 3 for a detailed explanation associated with each reference number.
Figure 2.
Characteristics and mechanisms of plants species that play a positive role in the restoration of degraded tropical soil. See Table 3 for a detailed explanation associated with each reference number.


{kind=link}
{kind=link}
Table 1.
Summary of inclusion and exclusion criteria used in this review.
| Category | Inclusion Criteria | Exclusion Criteria |
|---|---|---|
| 1. Literature type | Peer-reviewed publications | Gray literature and non-reviewed documents |
| 2. Languages | Documents published in French and English | Documents in all other languages |
| 3. Topics | Documents on the restoration of degraded tropical soils using plant species | All other themes |
| 4. Geography | Tropical zone | All other parts of the globe |
| 5. Period | Documents published between 1993 and 2023 | Documents published before 1993 |
| 6. Database | Google Scholar and Scopus | All other databases |
Table 2.
Classification of tropical soils into eight main groups.
| Soil Group | General Description | Geographical Distribution | References |
|---|---|---|---|
| Acrisols | Soils with a higher clay content in the subsoil than in the topsoil. At certain depths, they have a low base saturation and low clay content. | Humid tropical, humid subtropical, and warm temperate regions, widespread in Southeast Asia, the Amazon, the southeastern United States, and east and west Africa. | [38,39,40,41]. |
| Ferralsols | Deeply weathered, red, or yellow soils typical of the humid tropics, rich in primary minerals and kaolinite. | Worldwide extent estimated at around 750 million hectares, almost exclusively in the humid tropics on the continental shields of South America and Africa. | [41,42,43,44]. |
| Fluvisols | Young soils formed in alluvial deposits, having a profile with signs of stratification and little horizon differentiation. | Present on every continent and in every climate, they occupy some 350 million hectares, with more than half of them in the tropics. | [41,45,46]. |
| Lixisols | Soils with a higher clay content in subsoil than topsoil. They have a high base saturation and low clay content at certain depths. | Found in tropical, subtropical, and warm temperate regions, seasonally dry, on Pleistocene and older surfaces. | [41,47,48,49]. |
| Luvisols | Soils with a higher clay content in subsoil than topsoil. They have highly active clays in the clay horizon and a high base saturation at certain depths. | Covers 500 M to 600 M ha worldwide, mainly in temperate, subtropical, and tropical regions. | [41,50,51,52]. |
| Nitisols | Deep, well-drained, red tropical soils with >30% clay subsoil and a stable structure. | Around 200 M ha worldwide, more than half of which are in tropical Africa, notably on the high plateaus (>1000 m) of Ethiopia, Kenya, Congo, and Cameroon. | [41,53,54,55]. |
| Plinthosols | Soils containing plinthite, a mixture of kaolinitic clays rich in Fe and/or Mn. | Found in the humid tropics, notably in the eastern Amazon basin, the central Congo basin, and parts of southeast Asia. | [41,56,57,58]. |
| Vertisols | Clay soils, with a high proportion of swelling clays, forming wide, deep fissures when they dry out. | Found mainly in semi-arid tropical regions but also in humid tropical regions such as Trinidad. | [41,59,60]. |
Table 3.
Mechanisms by which plants can help restore degraded tropical soils and characteristics of associated species.
Table 3.
Mechanisms by which plants can help restore degraded tropical soils and characteristics of associated species.
| Restoration Mechanisms | Plant Characteristics (Numbers in Parenthesis Refer to Those Shown in Figure 2) |
|---|---|
| Nitrogen fixation |
|
| Carbon sequestration |
|
| Organic matter addition |
|
| Structure stabilization |
|
| Erosion control |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Massoukou Pamba, R.; Poirier, V.; Nguema Ndoutoumou, P.; Epule, T.E. How Can Plants Help Restore Degraded Tropical Soils? Land 2023, 12, 2147. https://doi.org/10.3390/land12122147
AMA Style
Massoukou Pamba R, Poirier V, Nguema Ndoutoumou P, Epule TE. How Can Plants Help Restore Degraded Tropical Soils? Land. 2023; 12(12):2147. https://doi.org/10.3390/land12122147
Chicago/Turabian StyleMassoukou Pamba, Renaud, Vincent Poirier, Pamphile Nguema Ndoutoumou, and Terence Epule Epule. 2023. "How Can Plants Help Restore Degraded Tropical Soils?" Land 12, no. 12: 2147. https://doi.org/10.3390/land12122147
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.