
Ecological and Biotechnological Relevance of Mediterranean Hydrothermal Vent Systems

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
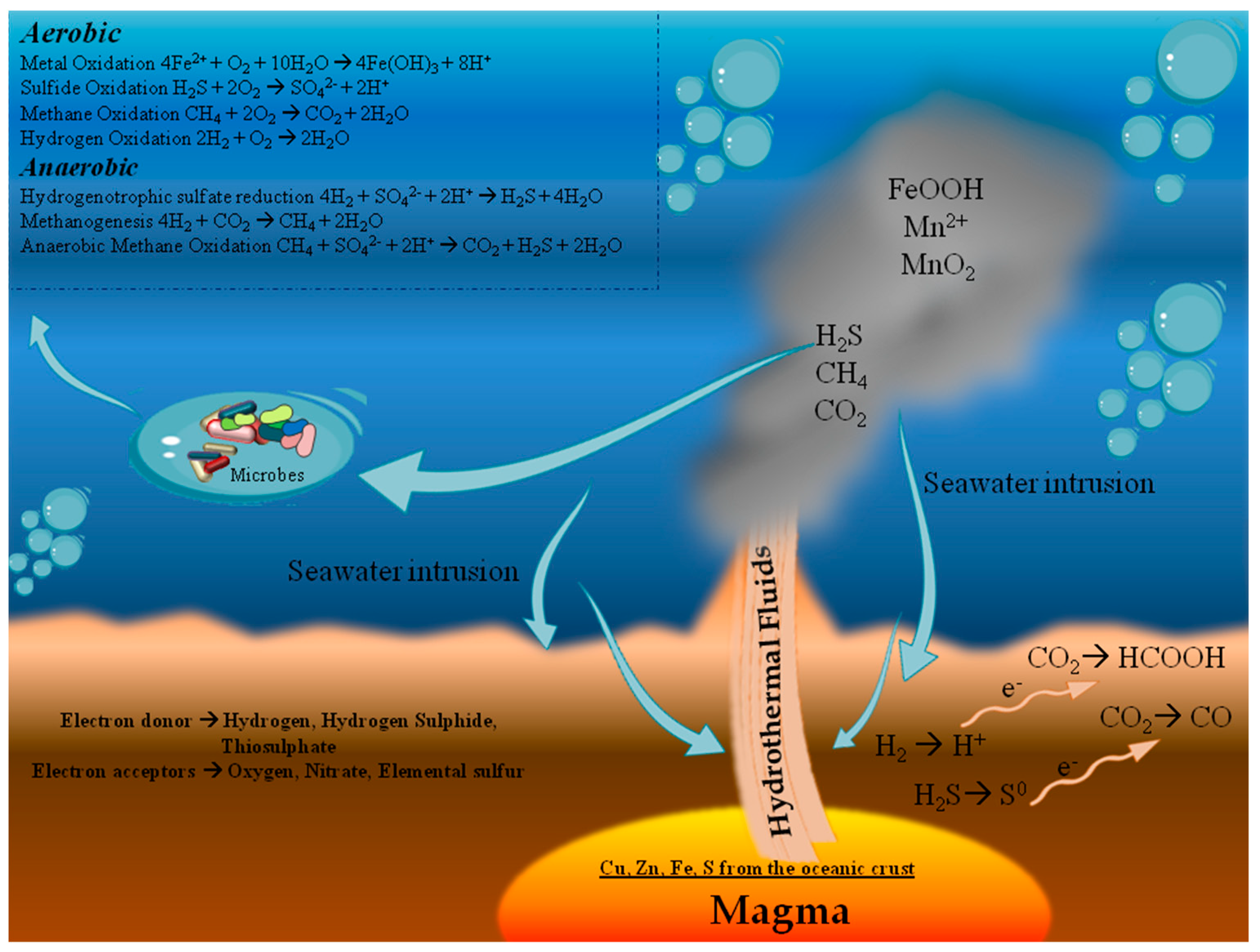
1.1. Environmental Parameters
1.1.1. Temperature
1.1.2. Light radiation
1.1.3. pH
1.1.4. Chemical Elements
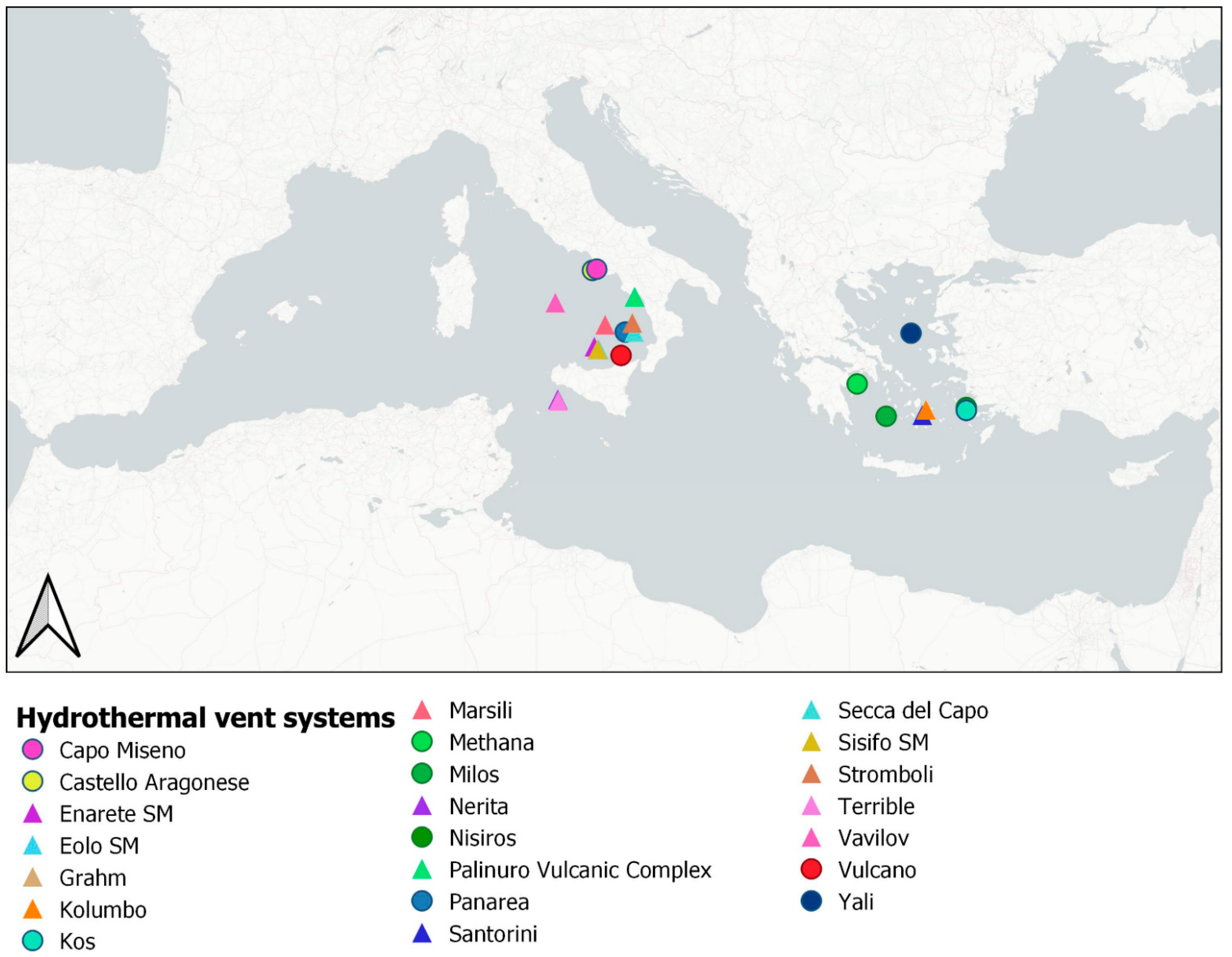
1.2. Hydrothermal Systems in the Mediterranean Sea

{kind=link}
{kind=link}
| Site | Vent | Max Temperature (°C) | Max Depth (m) | Discovery | Coordinates | References | |
|---|---|---|---|---|---|---|---|
| Shallow Mediterranean Hydrothermal vents | |||||||
| Aeolian Arc Panarea | Smoking Land Black Point Hot/Cold La Calcara Bottaro Crater | 135 | 200 | 1991, SCUBA | 38°63′ | 15°06′ | [33,38,39,51,52,53,54,55] |
| Aeolian Arc Vulcano | Levante Bay | 120 | 8 | 1969, SCUBA | 38°24′ | 14°57′ | [14,85] |
| Aeolian Arc Palinuro Vulcanic Complex | Grotta Azzurra, Grotta Sulfurea | 25 | 30 | 1990, SCUBA | 40°01′ | 15°26′ | [47,64,65] |
| Ischia Island | Castello Aragonese | ND * | 10 | 1954, from the beach | 40°44′ | 13°56′ | [10,70,71,75,76,86] |
| Naples Gulf | Capo Miseno Phlegrean Fileds | ND * | 20 | 1991, SCUBA | 40°46′ | 14°5′ | [14,61,85] |
| Hellenic Arc Milos | Milos | ND * | 200 | 1987, SCUBA 1992 Minirover ROV deeper sites | 36°41′ | 24°23′ | [45,46,76,77,78,79,80,86] |
| Hellenic Arc Kos | Brostherma Kephalos Bay Paradise Beach | ND * | 10 | 1983, SCUBA | 36°51′ | 27°14′ | [45,76,77,78,79,80,86] |
| Hellenic Arc Yali | Yali | ND * | 3 | 1990, SCUBA | 39°1′ | 25°16′ | [76,77,78,79,80,86] |
| Hellenic Arc Methana | Methana | ND * | 10 | 1997, SCUBA | 37°36′ | 23°21′ | [45,76,77,78,79,80,86] |
| Hellenic Arc Nisiros | Lefkos Bay | ND * | - | 1994, SCUBA | 36°56′ | 27°14′ | [76,77,78,79,80,86] |
| Deep Mediterranean hydrothermal vents | |||||||
| Tyrrhenian Back-Arc | Marsili | ND * | 800 | 2006, ROV Cherokee | 39°15′ | 14°23′ | [41,64,67] |
| Aeolian Arc | Enarete SM | ND * | 600 | 2011, ROV Hercules | 38°62′ | 14°0′ | [85,86] |
| Eolo SM | ND * | 800 | 2011, ROV Hercules | 38°58′ | 14°16′ | [85,86] | |
| Sisifo SM | ND * | 1200 | 2011, ROV Hercules | 38°34′ | 14°7′ | [85,86] | |
| Secca del Capo | ND * | 240 | 1995, ROV Comex | 38°63′ | 14°85′ | [54,86] | |
| Vavilov | ND * | 3500 | 1959, Akademik Valilov | 39°51′ | 12°45′ | [85,86] | |
| Stromboli | ND * | 250 | 2011, ROV Hercules | 38°78′ | 15°21′ | [46,86] | |
| Aeolian Arc Palinuro Volcanic Complex | Palinuro SM | ND * | 1000 | 2011, ROV Hercules | 39°55′ | 14°7′ | [41,47,64,67,86] |
| Hellenic Arc | Santorini | ND * | 400 | 2006, ROV Thetis | 36°43′ | 25°40′ | [76,77,78,79,80,86] |
| Santorini | |||||||
| Hellenic Arc Kolumbo | Kolumbo | 224 | 500 | 2006, ROV Hercules | 36°52′ | 25°48′ | [76,77,78,79,80,86] |
| Empedocle Banks | Grahm | ND * | 250 | ROV | 37°10′ | 12°42′ | [48] |
| Nerita | ND * | - | 37°10′ | 12°43′ | |||
| Terrible | ND * | - | 37°07′ | 12°43′ | |||
1.3. Scientific Interest
2. Necto-Benthic Communities in Hydrothermal Systems
2.1. Physiological Responses to Environmental Stressors
2.2. Abundance and Distribution
3. Prokaryotic Communities in Hydrothermal Systems
3.1. Survival Strategies and Adaptation
3.2. Abundance and Distribution
| Site | Sample | Technique | Bacteria | Archea | Reference |
|---|---|---|---|---|---|
| Taxa | Taxa | ||||
| Thyrrhenian Sea Tor Caldara | Sediment, biofilm | Illumina HiSeq | Gamma, Epsilon, Alfa | ND * | [242] |
| Thyrrhenian Sea Basiluzzo Islet | Sediment | 454 | Flavo, Gamma, Delta | ND * | [38,39,52] |
| Thyrrhenian Sea Phlegrean Fileds | Sediment | Gamma, Firmicutes, Bacillus, Actinobacteria | Euryarchaeota, Thaumarchaeota | [36] | |
| Aegean Sea Milos | Sediment | Clone libraries | Epsilon, Firmicutes, Bacteroidetes, Aquificae | Euryarchaeota, Crenarchaeota | [43,226,232] |
| Thyrrhenian Sea Panarea Hot/cold vents | Sediment | Illumina/454 | Epsilon (both), Delta (Hot), Alfa and Gamma (Cold) | Euryarchaeota (Hot Vent) Crenarchaeota (Cold Vent) | [5] |
| Thyrrhenian Sea Panarea Smoking land | Sediment | 454 | Alpha, Beta, Gamma, Delta, Zetaproteobacteria | Marine Group II (MGII), the Marine Group III (MGIII), the Marine Benthic Group E (MBGE), The Deep-Sea Hydrothermal Vent Euryarchaeotal Group 6 (Woesearchaeota), Marine Hydrothermal Vent Group (MHVG), Thaumarchaeota, Miscellaneous Crenarchaeotal Group 3 (C3) | [38,39] |
| Thyrrhenian Sea Panarea Black Point | Sediment | Illumina paired-end | Alfa, Beta, Gamma, Acidobacteria, Chlorobi | Euryarchaeota Crenarchaerota Koraarchaeota | [57,226] |
| Thyrrhenian Sea Vulcano | Sediment | Illumina MiSeq | Proteobacteria, Planctomycetes, Actinobacteria, Firmicudes, Bacteroidetes | ND * | [23,226,252] |
| Aegean Sea Kolumbo | Sediment | Illumina | Epsilon, Aquificae, Delta, Gamma, Planctomycetes, Bacteroidetes | ND * | [84] |
4. Biotechnological Relevance
4.1. Extremozymes
4.2. Exopolysaccharides
4.3. Applications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lu, G.-S.; LaRowe, D.E.; Fike, D.A.; Druschel, G.K.; Gilhooly, W.P., III; Price, R.E.; Amend, J.P. Bioenergetic characterization of a shallow-sea hydrothermal vent system: Milos Island, Greece. PLoS ONE 2020, 15, e0234175. [Google Scholar]
- Burgess, E.A.; Wagner, I.D.; Wiegel, J. Physiology and Biochemistry of Extremophiles; Gerday, C., Glansdorff, N., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 13–29. [Google Scholar]
- Gugliandolo, C.; Maugeri, T. Phylogenetic Diversity of Archaea in Shallow Hydrothermal Vents of Eolian Islands, Italy. Diversity 2019, 11, 156. [Google Scholar] [CrossRef] [Green Version]
- Bellec, L.; Cambon-Bonavita, M.-A.; Durand, L.; Aube, J.; Gayet, N.; Sandulli, R.; Brandily, C.; Zeppilli, D. Microbial Communities of the Shallow-Water Hydrothermal Vent Near Naples, Italy, and Chemosynthetic Symbionts Associated With a Free-Living Marine Nematode. Front. Microbiol. 2020, 11, 2023. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Lentini, V.; Bunk, B.; Overmann, J.; Italiano, F.; Maugeri, T.L. Changes in prokaryotic community composition accompanying a pronounced temperature shift of a shallow marine thermal brine pool (Panarea Island, Italy). Extremophiles 2015, 19, 547–559. [Google Scholar] [CrossRef]
- Tarasov, V.G. Effects of shallow-water hydrothermal venting on biological communities of coastal marine ecosystems of the Western Pacific. Adv. Mar. Biol. 2006, 50, 267–421. [Google Scholar]
- Hirayama, H.; Sunamura, M.; Takai, K.; Nunoura, T.; Noguchi, T.; Oida, H.; Furushima, Y.; Yamamoto, H.; Oomori, T.; Horikoshi, K. Culture-dependent and -independent characterization of microbial communities associated with a shallow submarine hydrothermal system occurring within a Coral Reef off Taketomi Island, Japan. Appl. Environ. Microbiol. 2007, 73, 7642–7656. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Micheli, F.; Peterson, C.H.; Mullineaux, L.S.; Fisher, C.R.; Mills, S.W.; Sancho, G.; Johnson, G.A.; Lenihan, H.S. Predation structures communities at deep-sea hydrothermal vents. Ecol. Monogr. 2002, 72, 365–382. [Google Scholar] [CrossRef]
- Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Martin, S.; Ransome, E.; Fine, M.; Turner, S.M.; Buia, M.C. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 2008, 454, 96. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microb. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Yao, H.; Dao, M.; Imholt, T.; Huang, J.; Wheeler, K.; Bonilla, A.; Ortiz, C. Protection mechanisms of the iron-plated armor of a deep-sea hydrothermal vent gastropod. Proc. Natl. Acad. Sci. USA 2010, 107, 987–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarasov, V.G.; Gebruk, A.V.; Mironov, A.N.; Moskalev, L.I. Deep-sea and shallow-water hydrothermal vent communities: Two different phenomena? Chem. Geol. 2005, 224, 5–39. [Google Scholar] [CrossRef]
- Dando, P.R.; Stüben, D.; Varnavas, S.P. Hydrothermalismin theMediterranean Sea. Prog. Oceanogr. 1999, 44, 333–367. [Google Scholar] [CrossRef]
- Dando, P.R.; Hughes, J.A.; Thiermann, F. Preliminary observations on biological communities at shallow hydrothermal vents in the Aegean Sea. In Hydrothermal Vents and Processes; Parson, L.M., Walker, C.L., Dixon, D.R., Eds.; Special Publications: London, UK, 1995; Volume 87, pp. 303–317. [Google Scholar]
- Botz, R.; Stuben, D.; Winckler, G.; Bayer, R.; Schmitt, M.; Faber, E. Hydrothermal gases from offshore Milos Island, Greece. Chem. Geol. 1996, 130, 161–173. [Google Scholar] [CrossRef]
- Fitzsimons, M.F.; Dando, P.R.; Hughes, J.A.; Thiermann, F.; Akoumianaki, I.; Pratt, S.M. Submarine hydrothermal brine seeps off Milos, Greece: Observations and geochemistry. Mar. Chem. 1997, 57, 325–340. [Google Scholar] [CrossRef]
- Tarasov, V.G.; Gebruk, A.V.; Shulkin, V.M.; Kamenev, G.M.; Fadeev, V.I.; Kosmynin, V.N.; Malakhov, V.V.; Starynin, D.A.; Obzhirov, A.I. Effect of shallow-water hydrothermal venting on the biota of Matupi Harbor (Rabaul Caldera, New Britain Island Papaua-New Guinea). Cont. Shelf Res. 1999, 19, 79–116. [Google Scholar] [CrossRef]
- Tarasov, V.G.; Propp, M.V.; Propp, L.N. Functions of the coastal marine ecosystems in relation of hydrothermal venting. Ecolog. Chim. 2002, 11, 1–15. (In Russian) [Google Scholar]
- Barriga, F.F.A.; Costa, I.M.A.; Relvas, J.M.R.; Ribeiro, A.; Fouquet, Y.; Ondreas, H.; Parson, L. The Rainbow serpentinites and serpentinite-sulphide stockwork (mid-Atlantic Ridge, AMAR-segment): A preliminary report of the FLORES results. EOS Trans. Am. Geophys. Union 1997, 78, F832. [Google Scholar]
- Baross, J.A.; Hoffman, S.E. Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life. Nav. Res. Rev. 1986, 2, 1–12. [Google Scholar] [CrossRef]
- Childress, J.J.; Fisher, C.R. The biology of hydrothermal vent animals: Physiology, biochemistry and autotrophic symbioses. Oceanogr. Mar. Biol. Annu. Rev. 1992, 30, 337–441. [Google Scholar]
- Gugliandolo, C.; Maugeri, T.L. Chemolithotrophic, sulphur-oxidizing bacteria from a marine, shallow hydrothermal vent of Vulcano (Italy). Geomicrobiol. J. 1993, 11, 109–120. [Google Scholar] [CrossRef]
- Price, R.E.; Savov, I.; Planer-Friedrich, B.; Bühring, S.; Amend, J.; Pichler, T. Processes influencing extremeas enrichment in shallow-seahydrothermal fluids of Milos Island, Greece. Chem. Geol. 2013, 348, 15–26. [Google Scholar] [CrossRef]
- Price, R.E.; London, J.; Wallschläger, D.; Ruiz-Chancho, M.J.; Pichler, T. Enhanced bioaccumulation and biotransformation of As in coral reef organisms surrounding amarine shallow-water hydrothermal vent system. Chem. Geol. 2013, 348, 48–55. [Google Scholar] [CrossRef]
- Price, R.E.; Lesniewski, R.; Nitzsche, K.; Meyerdierks, A.; Saltikov, C.; Pichler, T.; Amend, J. Archaeal and bacterial diversity in an arsenic-rich shallow-seahydrothermal system undergoing phase separation. Front. Microbiol. 2013, 4, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manini, E.; Luna, G.M.; Corinaldesi, C.; Zeppilli, D.; Bortoluzzi, G.; Caramanna, G.; Raffa, F.; Danovaro, R. Prokaryote diversity and virus abundance in shallow hydrothermal vents of the Mediterranean Sea (Panarea Island) and the Pacific Ocean (North Sulawesi-Indonesia). Microb. Ecol. 2008, 55, 626–639. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Massoth, G.J.; McDuff, R.E.; Lupton, J.E.; Lilley, M.D. Geochemistry of hydrothermal fluids fromAxial Seamount hydrothermal vent field, Juan deFucaRidge: Subseafloor boiling and subsequent fluid-rock interaction. J. Geophys. Res. 1990, 95, 12895–12921. [Google Scholar] [CrossRef]
- Fouquet, Y.; Charlou, J.L.; Costa, l.; Donval, J.P.; Radford-Knoery, J.; Pellè, H.; Ondreas, H.; Lourenco, N.; Segonzac, M.; Tivey, M.K. A detailed study of the Lucky-Strike hydrothermal site and discovery of a new hydrothermal site:«Menez-Gwen». Preliminary results of DIVA 1 cruise (5–29 May, 1994). Preliminary Results of the DIVAlcruise. Inter-Ridge News 1994, 3, 14–17. [Google Scholar]
- Bogdanov, Y.A. Submarine hydrothermal vents as a habitatfor marine organisms. In Biology Hydrother-mal Systems; Gebruk, A.V., Ed.; KMK Press: Moscow, Russian, 2002; pp. 72–112. (In Russian) [Google Scholar]
- Andaloro, F.; Romeo, T.; Renzi, M.; Guerranti, C.; Perra, G.; Consoli, P.; Perzia, P.; Focardi, S.E. Alteration of potential harmful elements levels in sediments and biota from the central Mediterranean Sea (Aeolian Archipelago) following an episode of intense volcanic activity. Environ. Monit. Assess. 2012, 184, 4035–4047. [Google Scholar] [CrossRef]
- Stumm, W.; Morgan, J.J. Chemical Equilibria and Rates in Natural Waters, 3rd ed.; Aquatic Chemistry; Wiley: Hoboken, NJ, USA, 1995; ISBN 978-0-471-51185-4. [Google Scholar]
- Caracausi, A.; Ditta, M.; Italiano, F.; Longo, M.; Nuccio, P.M.; Paonita, A.; Rizzo, A. Changes in fluid geochemistry and physico-chemical conditions of geothermal systems caused by magmatic input: The recent abrupt outgassing off the island of Panarea (Aeolian Islands, Italy). Geochim. Cosmochim. Acta 2005, 69, 3045–3059. [Google Scholar] [CrossRef]
- Price, R.E.; Giovannelli, D. A Review of the Geochemistry and Microbiology of Marine Shallow-Water Hydrothermal Vents. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Renzi, M.; Romeo, T.; Guerranti, C.; Perra, G.; Italiano, F.; Focardi, S.E.; Esposito, V.; Andaloro, F. Temporal Trends and Matrix-Dependent Behaviors of Trace Elements Closed to a Geothermal Hot-Spot Source (Aeolian Archipelago, Italy). Procedia Earth Planet. Sci. 2011, 4, 10–28. [Google Scholar] [CrossRef] [Green Version]
- Crognale, S.; Casentini, B.; Amalfitano, S.; Fazi, S.; Petruccioli, M.; Rossetti, S. Biological As(III) oxidation in biofilters by using native groundwater microorganisms. Sci. Total Environ. 2018, 651, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Handley, K.M.; Hery, M.; Lloyd, J.R. Redox cycling of arsenic by the hydrothermal marine bacterium Marinobacter santoriniensis. Environ. Microbiol. 2009, 11, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, G.; Romeo, T.; La Cono, V.; La Spada, G.; Smedile, F.; Esposito, V.; Sabatino, G.; Di Bella, M.; Canese, S.; Scotti, G. Ferrous iron-and ammonium-rich diffuse vents support habitat-specific communities in a shallow hydrothermal field off the Basiluzzo Islet (Aeolian Volcanic Archipelago). Geobiology 2017, 15, 664–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, V.; Andaloro, F.; Canese, S.; Bortoluzzi, G.; Bo, M.; Di Bella, M.; Italiano, F.; Sabatino, G.; Battaflia, P.; Consoli, P.; et al. Exceptional discovery of a shallow-water hydrothermal site in the SW area of Basiluzzo islet (Aeolian archipelago, South Tyrrhenian Sea): An environment to preserve. PLoS ONE 2018, 13, e0190710. [Google Scholar] [CrossRef] [PubMed]
- Sciutteri, V.; Smedile, F.; Vizzini, S.; Mazzola, A.; Vetriani, C. Microbial biofilms along a geochemical gradient at the shallow-water hydrothermal system of Vulcano island, Mediterranean Sea. Front. Microbiol. 2022; in press. [Google Scholar]
- Boatta, F.; D’Alessandro, W.; Gagliano, A.L.; Liotta, M.; Milazzo, M.; Rodolfo-Metalpa, R.; Hall-Spencer, J.M.; Parello, F. Geochemical survey of Levante Bay, Vulcano Island (Italy), a natural laboratory for the study of ocean acidification. Mar. Pollut. Bull. 2013, 73, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Donnarumma, L.; Appolloni, L.; Chianese, E.; Bruno, R.; Baldrighi, E.; Guglielmo, R.; Russo, G.; Zeppilli, D.; Sandulli, R. Environmental and benthic community patterns of the shallow hydrothermal area of secca delle fumose (Baia, Naples, Italy). Front. Mar. Sci. 2019, 6, 685. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, D.; d’Errico, G.; Manini, E.; Yakimov, M.; Vetriani, C. Diversity and phylogenetic analyses of bacteria from a shallow-water hydrothermal vent in Milos island (Greece). Front. Microbiol. 2013, 4, 184. [Google Scholar] [CrossRef] [Green Version]
- Honnorez, J. La formation actuelle d’un gisement sous-marin de sulphides fumerolliens a Vulcano (Mer Tyrrhenienne). Partie I. Les Mineraux sulphures des tufs immerges a faible profondeur. Min. Depos. 1969, 4, 114–131. [Google Scholar]
- Bianchi, C.N.; Morri, C. Note sul benthos marino costiero dell’isola di Kos (Egeo Sud-Orientale). Nat. Milano 1983, 74, 96–114. [Google Scholar]
- Bonatti, E. Submarine iron deposits from the Mediterranean Sea. In The Mediterranean Sea: A Natural Sedimentation Laboratory; Stanley, D.J., Ed.; Dowden, Hutchinson & Ross, Inc.: Stroudsberg, PA, USA, 1982; pp. 701–710. [Google Scholar]
- Kidd, R.B.; Armarsson, H. Manganese and iron micronodules from a volcanic seamount in the Tyrrhenian Sea. J. Geol. Soc. 1979, 136, 71–76. [Google Scholar] [CrossRef]
- Coltelli, M.; Cavallaro, D.; D’Anna, G.; D’Alessandro, A.; Grassa, F.; Mangano, G.; Patanè, D.; Gresta, S. Exploring the submarine graham bank in the sicily channel. Ann. Geophys. 2016, 59, S0208. [Google Scholar] [CrossRef]
- Crisci, J.V.; Cigliano, M.M.; Morrone, J.J.; Roigjunen, S. Historical biogeography of southern South America. Syst. Zool. 1991, 40, 152–171. [Google Scholar] [CrossRef]
- Calanchi, N.; Peccerillo, A.; Tranne, C.; Lucchini, F.; Rossi, P.L.; Kempton, P.; Barbieri, W.; Woo, T.W. Petrology and geochemistry of volcanic rocks from the island of Panarea: Implications for mantle evolution beneath the Aeolian island arc (Southern Tyrrhenian sea). J. Volcanol. Geotherm. Res. 2002, 115, 367–395. [Google Scholar] [CrossRef]
- Voltattorni, N.; Caramanna, G.; Cinti, D.; Galli, G.; Pizzino, L.; Quattrocchi, F. Study of CO2 natural emissions in different italian geological scenarios: A refinement of natural hazard and risk assessment. In Advances in the Geological Storage of Carbon Dioxide; Lombardi, S., Altunina, L.K., Beaubien, S.E., Eds.; NATO Science Series; Springer: Berlin, Germany, 2006; Volume XV, 362p, ISBN 1-4020-4470-4. [Google Scholar]
- Esposito, A.; Giordano, G.; Anzidei, M. The 2002–2003 submarine gas eruption at Panarea volcano Aeolian Islands, Italy: Volcanology of the seafloor and implications for the hazard scenario. Mar. Geol. 2006, 227, 119–134. [Google Scholar] [CrossRef]
- Peters, M.; Strauss, H.; Petersen, S.; Kummer, N.; Thomazo, C. Hydrothermalism in the Tyrrhenian Sea: Inorganic and microbial sulphur cycling as revealed by geochemical and multiple sulphur isotope data. Chem. Geol. 2011, 280, 217–231. [Google Scholar] [CrossRef]
- Gamberi, F.; Marani, M.; Savelli, C. Tectonic volcanic and hydrothermal features of a submarine portion of the Aeolian arc (Tyrrhenian Sea). Mar. Geol. 1997, 140, 167–181. [Google Scholar] [CrossRef]
- Italiano, F. Hydrothermal fluids vented at shallow depths at the Aeolian islands: Relationships with volcanic and geothermal systems. In Proceedings of the International Workshop Research in Shallow Marine and Fresh Water Systems, Milazzo, Sicilia, Italy, 14–16 May 2009; pp. 55–60. [Google Scholar]
- Gugliandolo, C.; Spanò, A.; Lentini, V.; Arena, A.; Maugeri, T.L. Antiviral and immunomodulatory effects of a novel bacterial exopolysaccharide of shallow marine vent origin. J. Appl. Microbiol. 2014, 116, 1028–1034. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Bianconi, G.; Canganella, F.; Danovaro, R.; Gugliandolo, C.; Italiano, F.; Lentini, V.; Manini, E.; Nicolaus, B. Shallow hydrothermal vents in the southern Tyrrhenian Sea. Chem. Ecol. 2010, 26, 285–298. [Google Scholar] [CrossRef]
- Spagnoli, F.; Andaloro, F.; Canese, S.; Capaccioni, B.; Iassunto, R.A. Nuove recenti conoscenze sul sistema idrotermale del complesso vulcanico dell’Isola di Panarea (Arcipelago delle Eolie, Mar Tirreno Meridionale). Mem. Descr. Carta Geol. d’It 2020, 105, 85–90. [Google Scholar]
- Molari, M.; Giovannelli, D.; D’errico, G.; Manini, E. Factors influencing prokaryotic community structure composition in sub-surface coastal sediments. Estuar. Coast. Shelf Sci. 2012, 97, 141–148. [Google Scholar] [CrossRef]
- Noè, S.; Bellavia, C.; Calvo, S.; Mazzola, A.; Pirrotta, M.; Sciandra, M.; Vizzini, S.; Tomasello, A. Resilience of the seagrass Posidonia oceanica following pulse-type disturbance. Mar. Environ. Res. 2020, 159, 105011. [Google Scholar] [CrossRef] [PubMed]
- Peccerillo, A. Geochemical similarities between Vesuvius, Phlegraean Fields and Stromboli volcanoes: Petrogenetic, geodynamic and volcanological implications. Min. Pet. 2001, 73, 93–105. [Google Scholar] [CrossRef]
- Amend, J.P.; Rogers, K.L.; Shock, E.L.; Gurrieri, S.; Inguaggiato, S. Energetics of chemolithoautotrophy in the hydrothermal system of Vulcano Island, southern Italy. Geobiology 2003, 1, 37–58. [Google Scholar] [CrossRef] [Green Version]
- Chiodini, G.; Caliro, S.; Caramanna, G.; Granieri, D.; Minopoli, C.; Moretti, R.; Perotta, L.; Ventura, G. Geochemistry of the submarine gaseous emissions of Panarea (Aeolian Islands, southern Italy): Magmatic vs. hydrothermal origin and implications for volcanic surveillance. Pure Appl. Geophys. 2006, 163, 759–780. [Google Scholar] [CrossRef]
- Monecke, T.; Petersen, S.; Augustin, N.; De Benedetti, A.A.; Esposito, A.; Gärtner, A.; Gemmell, B.; Gibson, H.; Hannington, M.D.; He, G.; et al. Shallow marine sulfide mineralization in the southeastern Tyrrhenian Sea, Italy. In Proceedings of the Joint Annual GAC-MAC-SEG-SGA Meeting, Quebec City, QC, Canada, 26–28 May 2008. [Google Scholar]
- Del Monte, M. Il vulcanesimo del Mar Tirreno- nota preliminare sui vulcani Marsili e Palinuro. G. Geol. 1972, 38, 231–252. [Google Scholar]
- Colantoni, P.; Lucchini, F.; Rossi, P.L.; Sartori, R.; Savelli, C. The Palinuro volcano and magmatism of the southeastern Tyrrhenian Sea (Mediterranean). Mar. Geol. 1981, 39, M1–M12. [Google Scholar] [CrossRef]
- Turley, C.M. The changing Mediterranean Sea a sensitive ecosystem? Prog. Oceanogr. 1999, 44, 387–400. [Google Scholar] [CrossRef]
- Cocchi, L.; Passaro, S.; Caratori Tontini, F.; Ventura, G. Volcanism in slab tear faults is larger than in island-arcs and back-arcs. Nat. Commun. 2017, 8, 1451. [Google Scholar] [CrossRef] [Green Version]
- Mattison, R.G.; Abbiati, M.; Dando, P.R.; Fitzsimons, M.F.; Pratt, S.M.; Southward, A.J.; Southward, E. Chemoautotrophic microbial mats in submarine caves with hydrothermal sulphidic springs at Cape Palinuro, Italy. Microb. Ecol. 1998, 35, 58–71. [Google Scholar] [CrossRef]
- De Alteriis, G.; Toscano, F. Introduzione alla geologia dei mari circostanti le isole flegree di Ischia, Procida e Vivara. In AA.VV., Ambiente Marino Costiero e Territorio delle Isole Flegree (Ischia, Procida e Vivara) e Golfo di Napoli, Risultati di uno Studio Multidisciplinare; Monografia Accademia di Scienze Matematiche, Fisiche e Naturali, Liguori: Naples, Italy, 2003; p. 77. [Google Scholar]
- Gambi, M.C. L’isola d’Ischia: Un osservatorio speciale per lo studio del cambiamento climatico globale a mare. In Ischia Patrimonio dell’Umanità. Natura e Cultura; Leone, U., Greco, P., Eds.; Doppiavoce Editore: Naples, Italy, 2014; pp. 71–97. [Google Scholar]
- De Alteriis, G.; Insinga, D.D.; Morabito, S.; Morra, V.; Chiocchi, F.L.; Terrasi, F.; Lubritto, C.; Di Benedetto, C.; Pazzanese, M. Age and submarine debris avalanches and tephrostratigraphy off shore Ischia Island, Tyrrhenian Sea, Italy. Mar. Geol. 2010, 277, 48–60. [Google Scholar]
- Tedesco, D.; Miele, G.; Sano, Y.; Toutain, J.P. Helium isotopic ratio in Vulcano island fumaroles: Temporal variations in shallow level mixing and deep magmatic supply. J. Volcanol. Geoth. Res. 1995, 64, 117–128. [Google Scholar] [CrossRef]
- Hoffmann, M.H.; Wroblewski, A.; Boyd, R. Mechanism controlling the clustering of fluvial channels and the compensational staching of cluster belts. J. Sediemntary Res. 2008, 81, 670–685. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C. Community dynamics and ecosystem simplification in a high-CO2 ocean. Proc. Natl. Acad. Sci. USA 2013, 110, 12721–12726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittman, A.; Gottini, V. L’isola d’Ischia. Geol. Boll. Serv. Geol. Ital. 1980, 101, 131–274. [Google Scholar]
- Bath, M. The seismology of Greece. Techtonophysics 1983, 98, 168–208. [Google Scholar] [CrossRef]
- Makropoulos, C.; Burton, P.W. Greek tectonics seismicity. Techtonophysics 1984, 106, 275–304. [Google Scholar] [CrossRef]
- Papanikolaou, D. Geotectonic evolution of the Aegean. Bull. Geol. Greece XXVII 1993, 28, 33–48. [Google Scholar]
- Royden, L.H.; Papanikolaou, D.J. Slab segmentation and late Cenozoic disruption of the Hellenic arc. Geochem. Geophys. Geosyst. 2011, 12, Q03010. [Google Scholar] [CrossRef]
- Morri, C.; Blanchi, C.N.; Cocito, S.; Peirano, A.; De Biase, A.M.; Aliani, S.; Pansini, M.; Boyer, M.; Ferdeghini, F.; Pestarino, M.; et al. Biodiversity of marine sessile epifauna at an Aegean island subject to hydrothermal activity: Milos, eastern Mediterranean Sea. Mar. Biol. 1998, 135, 729–739. [Google Scholar] [CrossRef]
- De Biasi, A.M.; Bianchi, C.N.; Aliani, S.; Cocito, S.; Peirano, A.; Dando, P.R.; Morri, C. Epibenthic communities in a marine shallow area with hydrothermal vents (Milos Island, Aegean Sea). Chem. Ecol. 2004, 20 (Suppl. S1), S89–S105. [Google Scholar] [CrossRef]
- De Biasi, A.M.; Aliani, S. Shallow-water hydrothermal vents in the Mediterranean sea: Stepping stones for Lessepsian migration? Hydrobiologia 2003, 503, 37–44. [Google Scholar] [CrossRef]
- Kilias, S.P.; Nomikou, P.; Papanikolaou, D.; Polymenakou, P.N.; Godelitsas, A.; Argyraki, A.; Scoullos, M. New insights into hydrothermal vent processes in the unique shallow-submarine arc-volcano, kolumbo (santorini), greece. Sci. Rep. 2013, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dando, P.R. Biological communities at marine shallow-water vent and seep sites. In The Vent and Seep Biota; Kiel, S., Ed.; Springer Science-Business Media: Dordrecht, The Netherlands, 2010; Volume 33. [Google Scholar]
- Dando, P.R.; Aliani, S.; Arab, H.; Bianchi, C.N.; Brehmer, M.; Cocito, S. Hydrothermal studies in the Aegean Sea. Phys. Chem. Earth 2000, 25, 1–8. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Jogler, M.; Wiegand, S.; Peeters, S.M.; Heuer, A.; Boedeker, C.; Jetten, M.S.M.; Rohde, M.; Jogler, C. Rubinisphaera italica sp. nov. isolated from a hydrothermal area in the Tyrrhenian Sea close to the volcanic island Panarea. Antoine Van Leeuwenhoek 2020, 113, 1727–1736. [Google Scholar] [CrossRef] [Green Version]
- Kallscheuer, N.; Jogler, M.; Wiegand, S.; Peeters, S.; Heuer, A.; Boedeker, C.; Jetten, M.; Rohde, M.; Jogler, C. Three novel Rubripirellula species isolated from artificial plastic surfaces submerged in the German part of the Baltic Sea and the estuary of the river Warnow. Antonie Van Leeuwenhoek 2019, 113, 1767–1778. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Wiegand, S.; Jogler, M.; Boedeker, C.; Peeters, S.; Rast, P.; Heuer, A.; Jetten, M.; Rohde, M.C.J. Rhodopirellula heiligendammensis sp. nov., Rhodopirellula pilleata sp. nov., and Rhodopirellula solitaria sp. nov. isolated from natural or artificial marine surfaces in Northern Germany and California, USA. Antonie Van Leeuwenhoek 2019, 113, 1737–1750. [Google Scholar] [CrossRef]
- Jogler, C.; Wiegand, S.; Boedeker, C.; Heuer, A.; Peeters, S.H.; Jogler, M.; Jetten, M.S.M.; Rohde, M.; Kallscheuer, N. Tautonia plasticadhaerens sp. nov., a novel species in the family Isosphaeraceae isolated from an alga in a hydrothermal area of the Eolian Archipelago. Antonie Van Leeuwenhoek 2020, 113, 1889–1900. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. Three novel halotolerant and thermophilic Geobacillus strains from shallow marine vents. Syst. Appl. Microbiol. 2002, 25, 450–455. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Panico, A.; Lama, L.; Gambacorta, A.; Nicolaus, B. A halophilic thermotolerant Bacillus isolated from a marine hot spring able to produce a new exopolysaccharide. Biotechnol. Lett. 2002, 24, 515–519. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Lentini, V.; Spanò, A.; Maugeri, T.L. New bacilli from shallow hydrothermal vents of Panarea Island (Italy) and their biotechnological potentialities. J. Appl. Microbiol. 2012, 112, 1102–1112. [Google Scholar] [CrossRef]
- Spanò, A.; Gugliandolo, C.; Lentini, V.; Maugeri, T.L.; Anzelmo, G.; Poli, A.; Nicolaus, B. A novel EPS-producing strain of Bacillus licheniformis isolated from a shallow vent off Panarea Island (Italy). Curr. Microbiol. 2003, 67, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Reinold, M.; Wong, H.L.; MacLeod, F.I.; Meltzer, J.; Thompson, A.; Burns, B.P. The Vulnerability of Microbial Ecosystems in a Changing Climate: Potential Impact in Shark Bay. Life 2019, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Delgado, S.; Hernandez, J.C. The importance of natural acidified systems in the study of Ocean Acidification: What have we learned? Adv. Mar. Biol. 2018, 80, 57–99. [Google Scholar] [CrossRef] [PubMed]
- Mirasole, A.; Gillanders, B.G.; Reis-Santos, P.; Grassa, F.; Capasso, G.; Scopelliti, G.; Mazzola, A.; Vizzini, S. The influence of high pCO2 on otolith shape, chemical and carbon isotope composition of six coastal fish species in a Mediterranean shallow CO2 vent. Mar. Biol. 2017, 164, 191. [Google Scholar] [CrossRef]
- Mirasole, A.; Scopelliti, G.; Tramati, C.; Signa, G.; Mazzola, A.; Vizzini, S. Evidences on alterations in skeleton composition and mineralization in a site-attached fish under naturally acidified conditions in a shallow CO2 vent. Sci. Total Environ. 2021, 761, 143309. [Google Scholar] [CrossRef] [PubMed]
- Milazzo, M.; Rodolfo-Metalpa, R.; Chan, V.B.S.; Fine, M.; Alessi, C.; Thiyagarajan, V.; Hall-Spencer, J.M.; Chemello, R. Ocean acidification impairs vermetid reef recruitment. Sci. Rep. 2014, 4, 4189. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, S.; Apostolaki, E.T.; Ricevuto, E.; Polymenakou, P.; Mazzola, A. Plant and sediment properties in seagrass meadows from two Mediterranean CO2 vents: Implications for carbon storage capacity of acidified oceans. Mar. Environ. Res. 2019, 146, 101–108. [Google Scholar] [CrossRef]
- Del Pasqua, M.; Gambi, M.C.; Caricato, R. Effects of short-term and long-term exposure to ocean acidification on T carbonic anhydrase activity and morphometric characteristics in the invasive polychaete Branchiomma boholense (Annelida: Sabellidae): A case-study from a CO2 vent system. Mar. Environ. Res. 2019, 144, 203–212. [Google Scholar] [CrossRef]
- Lucey, N.M.; Lombardi, C.; De Marchi, L.; Schulze, A.; Gambi, M.C.; Calosi, P. To brood or not to brood. Are marine organisms that protect their offspring more resilient to ocean acidification? Sci. Rep. 2015, 5, 12009. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Buia, M.C.; Palumbo, A.; Mohany, M.; Wadaan, M.A.M.; Hozzein, W.N.; Beemster, G.T.S.; AbdElgawad, H. Ocean acidification affects biological activities of seaweeds: A case study of Sargassum vulgare from Ischia volcanic CO2 vents. Environ. Pollut. 2020, 259, 113765. [Google Scholar] [CrossRef] [PubMed]
- Apostolaki, E.T.; Vizzini, S.; Hendriks, I.E.; Olsen, Y.S. Seagrass ecosystem response to long-term high CO2 in a Mediterranean volcanic vent. Mar. Environ. Res. 2014, 99, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, G.A.; Sciarretta, M.; Cassiano, C.; Buonocore, C.; Festa, C.; Mazzella, V.; Núñez Pons, L.; D’Auria, M.V.; de Pascale, D. Molecular Network and Culture Media Variation Reveal a Complex Metabolic Profile in Pantoea cf. eucrina D2 Associated with an Acidified Marine Sponge. Int. J. Mol. Sci. 2020, 21, 6307. [Google Scholar] [CrossRef]
- Zupo, V. Relevance of wound-activated compounds produced by diatoms as toxins and infochemicals for benthic invertebrates. Mar. Biol. 2014, 161, 1639–1652. [Google Scholar]
- Zupo, V.; Mutalipassi, M.; Ruocco, N.; Glaviano, F.; Pollio, A.; Langellotti, A.L.; Romano, G.; Costantini, M. Distribution of Toxigenic Halomicronema spp. in Adjacent Environments on the Island of Ischia: Comparison of Strains from Thermal Waters and Free Living in Posidonia Oceanica Meadows. Toxins 2019, 11, 99. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Boni, R.; Buia, M.C.; Monfrecola, V.; Esposito, M.C.; Tosti, E. Ocean acidification impact on ascidian Ciona robusta spermatozoa: New evidence for stress resilience. Sci. Total Environ. 2019, 697, 134100. [Google Scholar] [CrossRef]
- Biagi, E.; Caroselli, E.; Barone, M.; Pezzimenti, M.; Teixido, N.; Soverini, M.; Rampelli, S.; Turroni, S.; Gambi, M.C.; Brigidi, P.; et al. Patterns in microbiome composition differ with ocean acidification in anatomic compartments of the Mediterranean coral Astroides calycularis living at CO2 vents. Sci. Total Environ. 2020, 724, 138048. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Genet. 2019, 17, 569–586. [Google Scholar] [CrossRef] [Green Version]
- Huber, H.; Burggraf, S.; Mayer, T.; Wyschkony, I.; Rachel, R.; Stetter, K.O. Ignicoccus gen. nov., anovel genus of hyperthermophilic, chemolithoautotrophic Archaea, represented by two new species, Ignicoccus islandicus sp nov and Ignicoccus pacificus sp nov. and Ignicoccus pacificus sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 50, 2093–2100. [Google Scholar] [CrossRef]
- Menzel, P.; Gudbergsdottir, S.R.; Rike, A.G. Comparative metagenomics of eight geographically remote terrestrial hot springs. Microb. Ecol. 2015, 70, 411–424. [Google Scholar] [CrossRef]
- Colman, D.R.; Poudel, S.; Hamilton, T.L. Geobiological feedbacks and the evolution of thermoacidophiles. ISME J. 2018, 12, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrell, A.K.; van Oosterom, E.J.; Mullet, J.E.; George-Jaeggli, B.; Jordan, D.R.; Klein, P.E.; Hammer, G.L. Stay-green alleles individually enhance grain yield in sorghum under drought by modifying canopy development and water uptake patterns. New Phytol. 2014, 203, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, R.; Borell, E.M.; Yahm, R.; Shemesh, A.; Maoz, F. Natural high pCO2 increases autotrophy in Anemonia viridis (Anthozoa) as revealed from stable isotope (C, N) analysis. Sci. Rep. 2015, 5, 8779. [Google Scholar] [CrossRef] [Green Version]
- Duquette, A.; McClintock, J.B.; Amsler, C.D.; Pérez-Huerta, A.; Milazzo, M.; Hall-Spencer, J.M. Seawater carbonate chemistry and shell mineralogy, microstructure, and mechanical strength of four Mediterranean gastropod species near a CO2 seep. Mar. Pollut. Bull. 2017, 124, 917–928. [Google Scholar] [CrossRef]
- Norris, P.R.; Falagàn, C.; Moya-Beltran, A.; Castro, M.; Quatrini, R.; Johnson, D.B. Acidithiobacillus ferrianus sp. nov.: An ancestral extremely acidophilic and facultatively anaerobic chemolithoautotroph. Extremophiles 2020, 24, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Gugliandolo, C.; Maugeri, T.L.; Caccamo, D.; Stackebrandt, E. Bacillus aeolius sp. nov. a Novel Thermophilic, Halophilic Marine Bacillus Species from Eolian Islands (Italy). Syst. Appl. Microbiol. 2003, 26, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, M.; Schröder, C.; Klippel, B.; Schäfers, C.; Krüger, A.; Antranikian, G. Extremely thermoactive archaeal endoglucanase from a shallow marine hydrothermal vent from Vulcano Island. Appl. Microbiol. Biotechnol. 2019, 103, 1267–1274. [Google Scholar] [CrossRef]
- Antranikian, G.; Suleiman, M.; Schäfers, C.; Adams, M.W.W.; Bartolucci, S.; Blamey, J.M.; Birkeland, N.-K.; Bonch-Osmolovskaya, E.; da Costa, M.S.; Cowan, D.; et al. Diversity of bacteria and archaea from two shallow marine hydrothermal vents from Vulcano Island. Extremophiles 2017, 21, 733–742. [Google Scholar] [CrossRef]
- Placido, A.; Hai, T.; Ferrer, M.; Chernikova, N.T.; Distaso, M.; Armstrong, D.; Yakunin, F.A.; Toshchakov, S.V.; Yakimov, M.M.; Kublanov, I.V.; et al. Diversity of hydrolases from hydrothermal vent sediments of the Levante Bay, Vulcano Island (Aeolian archipelago) identified by activity-based metagenomics and biochemical characterization of new esterases and an arabinopyranosidase. Appl. Microbiol. Biotechnol. 2015, 99, 10031–10046. [Google Scholar] [CrossRef] [Green Version]
- Foo, S.A.; Byrne, M.; Ricevuto, E.; Gambi, M.C. The carbon dioxide vents of Ischia, Italy, a natural system to assess impacts of ocean acidification on marine ecosystems: An overview of research and comparisons with other vent systems. Oceanogr. Mar. Biol. 2018, 56, 237–310. [Google Scholar]
- Hilário, A.; Comas, M.C.; Azevedo, L.; Pinheiro, L.; Ivanov, M.K.; Cunha, M.R. First record of a Vestimentifera (Polychaeta: Siboglinidae) from chemosynthetic habitats in the western Mediterranean Sea-Biogeographical implications and future exploration. Deep-Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 200–207. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Ceregato, A.; Foglini, F.; Froglia, C.; Trincardi, F. The Gela Basin pockmark field in the strait of Sicily (Mediterranean Sea): Chemosymbiotic faunal and carbonate signatures of postglacial to modern cold seepage. Biogeosciences 2013, 10, 4653–4671. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, J.; Lott, C.; Weber, M.; Ramette, A.; Bright, M.; Dubilier, N.; Petersen, J.M. Dual symbiosis with co-occurring sulphur-oxidizing symbionts in vestimentiferan tubeworms from a Mediterranean hydrothermal vent. Environ. Microbiol. 2014, 16, 3638–3656. [Google Scholar] [CrossRef]
- Gaglioti, M.; Gambi, M.C. The natural laboratory of the CO2 vents off Panarea Aeolian islands, Italy: A special ecological setting and a further stepping stone for some alien macrophytes. Not. SIBM 2018, 75, 58–65. [Google Scholar]
- Morri, C.; Vafidis, D.; Peirano, A.; Chintiroglou, C.C.; Bianchi, C.N. Anthozoa from a subtidal hydrothermal area of milos island (aegean sea), with notes on the construction potential of the scleractinian coral madracis pharensis. Ital. J. Zool. 2000, 67, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Calosi, P.; Rastrick, S.P.S.; Graziano, M.; Thomas, S.C.; Baggini, C.; Carter, H.A.; Hall-Spencer, J.M.; Milazzo, M.; Spicer, J.I. Distribution of sea urchins living near shallow water CO2 vents is dependent upon species acid-base and ion-regulatory abilities. Mar. Pollut. Bull. 2013, 73, 470–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cigliano, M.; Gambi, M.C.; Rodolfo-Metalpa, R.; Patti, F.P.; Hall-Spencer, J.M. Effects of ocean acidification on invertebrate settlement at volcanic CO2 vents. Mar. Biol. 2010, 157, 2489–2502. [Google Scholar] [CrossRef] [Green Version]
- Garilli, V.; Rodolfo-Metalpa, R.; Scuderi, D.; Brusca, L.; Parrinello, D.; Rastrick, S.P.S.; Foggo, A.; Twitchett, R.J.; Hall-Spencer, J.M.; Milazzo, M. Physiological advantages of dwarfing in surviving extinctions in high-CO2 oceans. Nat. Clim. Chang. 2015, 5, 678–682. [Google Scholar] [CrossRef]
- Harvey, B.P.; McKeown, N.J.; Rastrick, S.P.S.; Bertolini, C.; Foggo, A.; Graham, H.; Hall-Spencer, J.M.; Milazzo, M.; Shaw, P.W.; Small, D.P.; et al. Individual and population-level responses to ocean acidification. Sci. Rep. 2016, 6, 20194. [Google Scholar] [CrossRef] [Green Version]
- Calosi, P.; Rastrick, S.P.S.; Lombardi, C.; de Guzman, H.J.; Davidson, L.; Jahnke, M.; Giangrande, A.; Hardege, J.D.; Schulze, A.; Spicer, J.I.; et al. Adaptation and acclimatization to ocean acidification in marine ectotherms: An in situ transplant experiment with polychaetes at a shallow CO2 vent system. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120444. [Google Scholar] [CrossRef] [Green Version]
- Rodolfo-Metalpa, R.; Houlbrèque, F.; Tambutté, É.; Boisson, F.; Baggini, C.; Patti, F.P.; Jeffree, R.; Fine, M.; Foggo, A.; Gattuso, J.P.; et al. Coral and mollusc resistance to ocean acidification adversely affected by warming. Nat. Clim. Chang. 2011, 1, 308–312. [Google Scholar] [CrossRef]
- Mucci, A. The solubility of calcite and aragonite in seawater at various salinities, temperatures, and one atmosphere total pressure. Am. J. Sci. 1983, 283, 780–799. [Google Scholar] [CrossRef]
- Weiner, S.; Addadi, L. Design strategies in mineralized biological materials. J. Mater. Chem. 1997, 7, 689–702. [Google Scholar] [CrossRef]
- Ries, J.B. Skeletal mineralogy in a high-CO2 world. J. Exp. Mar. Biol. Ecol. 2011, 403, 54–64. [Google Scholar] [CrossRef]
- Teixidó, N.; Caroselli, E.; Alliouane, S.; Ceccarelli, C.; Comeau, S.; Gattuso, J.P.; Fici, P.; Micheli, F.; Mirasole, A.; Monismith, S.G.; et al. Ocean acidification causes variable trait-shifts in a coral species. Glob. Chang. Biol. 2020, 26, 6813–6830. [Google Scholar] [CrossRef]
- Ziveri, P.; Passaro, M.; In Carbona, A.; Milazzo, M.; Rodolfo-Metalpa, R.; Hall-Spencer, J.M. Decline in coccolithophore diversity and impact on coccolith morphogenesis along a natural CO2 gradient. Biol. Bull. 2014, 226, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Ingrassia, M.; Di Bella, L.; Chiocci, F.L.; Martorelli, E. Influence of fluid emissions on shallow-water benthic habitats of the Pontine Archipelago (Tyrrhenian Sea, Italy). Alp. Mediterr. Quat. 2015, 28, 99–110. [Google Scholar]
- Di Bella, L.; Ingrassia, M.; Frezza, V.; Chiocci, F.L.; Pecci, R.; Bedini, R.; Martorelli, E. Spiculosiphon oceana (foraminifera) a new bio-indicator of acidic environments related to fluid emissions of the Zannone hydrothermal field (Central Tyrrhenian Sea). Mar. Environ. Res. 2018, 136, 89–98. [Google Scholar] [CrossRef]
- Martin, S.; Rodolfo-Metalpa, R.; Ransome, E.; Rowley, S.; Buia, M.-C.; Gattuso, J.P.; Hall-Spencer, J. Effects of naturally acidified seawater on seagrass calcareous epibionts. Biol. Lett. 2008, 4, 689–692. [Google Scholar] [CrossRef] [Green Version]
- Collard, M.; Rastrick, S.P.S.; Calosi, P.; Demolder, Y.; Dille, J.; Findlay, H.S.; Hall-Spencer, J.M.; Milazzo, M.; Moulin, L.; Widdicombe, S.; et al. The impact of ocean acidification and warming on the skeletal mechanical properties of the sea urchin Paracentrotus lividus from laboratory and field observations Marie. ICES J. Mar. Sci. 2016, 73, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Cattano, C.; Giomi, F.; Milazzo, M. Effects of ocean acidification on embryonic respiration and development of a temperate wrasse living along a natural CO2 gradient. Conserv. Physiol. 2016, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A.; Giordano, M.; Beardall, J.; Maberly, S.C. Algal evolution in relation to atmospheric CO2: Carboxylases, carbon-concentrating mechanisms and carbon oxidation cycles. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 493–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornwall, C.E.; Revill, A.T.; Hall-Spencer, J.M.; Milazzo, M.; Raven, J.A.; Hurd, C.L. Inorganic carbon physiology underpins macroalgal responses to elevated CO2. Sci. Rep. 2017, 7, 46297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, V.R.; Russell, B.D.; Fabricius, K.E.; Brownlee, C.; Hall-Spencer, J.M. Temperate and tropical brown macroalgae thrive, despite decalcification, along natural CO2 gradients. Glob. Chang. Biol. 2012, 18, 2792–2803. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, S.; Tomasello, A.; Maida, G.D.; Pirrotta, M.; Mazzola, A.; Calvo, S. Effect of explosive shallow hydrothermal vents on δ13C and growth performance in the seagrass Posidonia oceanica. J. Ecol. 2010, 98, 1284–1291. [Google Scholar] [CrossRef]
- Lauritano, C.; Ruocco, M.; Dattolo, E.; Buia, M.C.; Silva, J.; Santos, R.; Olive, I.; Costa, M.M.; Procaccini, G. Response of key stress-related genes of the seagrass Posidonia oceanica in the vicinity of submarine volcanic vents. Biogeosciences 2015, 12, 4185–4194. [Google Scholar] [CrossRef] [Green Version]
- Arnold, T.; Mealey, C.; Leahey, H.; Miller, A.W.; Hall-Spencer, J.M.; Milazzo, M.; Maers, K. Ocean acidification and the loss of phenolic substances in marine plants. PLoS ONE 2012, 7, e35107. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, S.; Di Leonardo, R.; Costa, V.; Tramati, C.D.; Luzzu, F.; Mazzola, A. Trace element bias in the use of CO2 vents as analogues for low pH environments: Implications for contamination levels in acidified oceans. Estuar Coast Shelf Sci. 2013, 134, 19–30. [Google Scholar] [CrossRef]
- McClintock, J.B.; Amsler, C.D.; Amsler, M.O.; Duquette, A.; Angus, R.A. Trace elements in shells of common gastropods in the near vicinity of a natural CO2 vent: No evidence of pH-dependent contamination. Biogeosciences 2012, 11, 5215–5237. [Google Scholar] [CrossRef]
- Ricevuto, E.; Vizzini, S.; Gambi, M.C. Ocean acidification effects on stable isotope signatures and trophic interactions of polychaete consumers and organic matter sources at a CO2 shallow vent system. J. Exp. Mar. Biol. Ecol. 2015, 468, 105–117. [Google Scholar] [CrossRef]
- Ricevuto, E.; Lorenti, M.; Patti, F. Temporal trends of benthic invertebrate settlement along a gradient of ocean acidification at natural CO2 vents (Tyrrhenian sea). Biol. Mar. Mediterr. 2012, 19, 49–52. [Google Scholar]
- Petersen, S.; Monecke, T.; Augustin, N.; De Benedetti, A.A.; Esposito, A.; Gardeler, A.; Gemmell, J.B.; Gibson, H.; He, G.; Kayser, A.; et al. Drilling submarine hydrothermal systems in the Tyrrhenian Sea, Italy. InterRidge News 2008, 17, 21–23. [Google Scholar]
- Southward, A.J.; Southward, E.C.; Dando, P.R.; Hughes, J.A.; Kennicutt, M.C.; Herrera-Alcala, J.; Leahy, Y. Behaviour and feeding of the nassariid gastropod Cyclope neritea, abundant at hydrothermal brine seeps off Milos (Aegean sea). J. Mar. Biol. Assoc. UK 1997, 77, 753–771. [Google Scholar] [CrossRef]
- Thiel, V.; Hügler, M.; Blümel, M.; Baumann, H.I.; Gärtner, A.; Schmaljohann, R.; Strauss, H.; Garbe-Schönberg, D.; Petersen, S.; Cowart, D.A.; et al. Widespread occurrence of two carbon fixation pathways in tubeworm endosymbionts: Lessons from hydrothermal vent associated tubeworms from the Mediterranean Sea. Front. Microbiol. 2012, 3, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrazin, J.V.; Robigou, S.K.; Juniper, J.R. Delaney, Biological and geological dynamics over four years on a high-temperature sulphide structure at the Juan de Fuca Ridge hydrothermal observatory. Mar. Ecol. Prog. Ser. 1997, 153, 5–24. [Google Scholar] [CrossRef] [Green Version]
- Dando, P.R.; Hughes, J.A.; Leathy, Y.; Niven, S.J.; Smith, C. Gas venting rates from submarine hydrothermal areas around the Island of Milos, Hellenic Volcanic Arc. Cont. Shelf Res. 1995, 15, 913–929. [Google Scholar] [CrossRef]
- Thiermann, F.; Akoumianaki, I.; Hugjes, A.J.; Giere, O. Benthic fauna of a shallow-water gaseohydrothermal vent area in the Aegean Sea (Milos, Greece). Mar. Biol. 1997, 128, 149–159. [Google Scholar] [CrossRef]
- Carey, S.; Bell, K.L.C.; Rosi, M.; Marani, M.; Nornikou, P.; Walker, S.L.; Faure, K.; Kelly, J. Submarine volcanoes of the Aeolian Arc, Tyrrhenian Sea. Oceanography 2012, 25, 32–33. [Google Scholar]
- Taviani, M. Marine chemosynthesis in the Mediterranean Sea. In The Mediterranean Sea; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 69–83. [Google Scholar]
- Acunto, S.; Rini, F.; Cinelli, F. Osservazioni sui popolamenti fitobentici della Baia di Levante dell’Isola di Vulcano (isole Eolie): Variabilità spaziale in relazione ad attività idrotermali. Caratterizzazione Ambientale Marine del Sistema Eolie e dei Bacini Limitrofi di Cefalù e Gioia (EOCUMM95). Data Rep. Consorzio Naz. Interuniv. Sci. Mare 1996, 323–326. [Google Scholar]
- Vizzini, S.; Martínez-Crego, B.; Andolina, C.; Massa-Gallucci, A.; Connell, S.D.; Gambi, M.C. Ocean acidification as a driver of community simplification via the collapse of higher-order and rise of lower-order consumers. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Teixidó, N.; Gambi, M.C.; Parravacini, V.; Kroeker, K.; Micheli, F.; Villéger, S.; Ballesteros, E. Functional biodiversity loss along natural CO2 gradients. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, K.E.; Langdon, C.; Uthicke, S.; Humphrey, C.; Noonan, S.; De’ath, G.; Okazaki, R.; Muehllehner, N.; Glas, M.S.; Lough, J.M. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nat. Clim. Chang. 2011, 1, 165–169. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Kluibenschedl, A.; Harrington, L.; Noonan, S.; De’ath, G. In situ changes of tropical crustose coralline algae along carbon dioxide gradients. Sci. Rep. 2015, 5, 9537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goffredo, S.; Prada, F.; Caroselli, E.; Capaccioni, B.; Zaccanti, F.; Fantazzini, P.; Fermani, S.; Reggi, M.; Levy, O.; Fabricius, K.E.; et al. Biomineralization control related to population density under ocean acidification. Nat. Clim. Chang. 2014, 4, 293–297. [Google Scholar] [CrossRef] [Green Version]
- Auriemma, R.; De Vittor, C.; Esposito, V.; Gaglioti, M.; Gambi, M.C. Motile fauna associated to Cystoseira brachycarpa var. balearica along a gradient of ocean acidification at a vent system off Panarea (Aeolian Islands, Italy). In Proceedings of the 50° Congresso della Società Italiana di Biologia Marina, Livorno, Italy, 10–14 June 2019; pp. 180–183. [Google Scholar]
- Auriemma, R.; De Vittor, C.; Gaglioti, M.; Esposito, V.; Teixido, N.; Gambi, M.C. An overview of the benthic habitat of the Bottaro crater hydrothermal vent system at Panarea (Aeolian Islands, Italy). In Proceedings of the International Conference of Gas Geochemistry—ICGG15, Palermo & Milazzo, Italy, 29 September–5 October 2019. [Google Scholar]
- Molari, M.; Guilini, K.; Lott, C.; Weber, M.; de Beer, D.; Meyer, S.; Ramette, A.; Wegener, G.; Wenzhöfer, F.; Martin, D.; et al. CO2 leakage alters biogeochemical and ecological functions of submarine sands. Sci. Adv. 2018, 4, eaao2040. [Google Scholar] [CrossRef] [Green Version]
- Molari, M.; Guilini, K.; Lins, L.; Ramette, A.; Vanreusel, A. CO2 leakage can cause loss of benthic biodiversity in submarine sands. Mar. Environ. Res. 2019, 144, 213–229. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Dando, P.R.; Morri, C. Increased diversity of sessile epibenthos at subtidal hydrothermal vents: Seven hypotheses based on observations at Milos Island, Aegean Sea. Adv. Oceanogr. Limnol. 2011, 2, 1–31. [Google Scholar] [CrossRef]
- Nielsen, C. A review of the solitary entoprocts reported from sponges from Napoli (Italy), with designation of a neotype of Loxosoma pes Schimdt, 1878. J. Nat. Hist. 2008, 42, 1573–1579. [Google Scholar] [CrossRef]
- Valvassori, G.; Massa-Gallucci, A.; Gambi, M.C. Reappraisal of Platynereis massiliensis (Moquin- Tandon) (Annelida, Nereididae), a neglected sibling species of Platynereis dumerilii (Audouin & Milne Edwards). Biol. Mar. Mediterr. 2015, 22, 113–116. [Google Scholar]
- Wäge, J.; Valvassori, G.; Hardege, J.D.; Shulze, A.; Gambi, M.C. The sibling polychaetes Platynereis dumerilii and Platynereis massiliensis in the Mediterranean Sea: Are phylogeographic patterns related to exposure to ocean acidification? Mar. Biol. 2017, 164, 199. [Google Scholar] [CrossRef]
- Scipione, M.B. On the presence of the Mediterranean endemic Microdeutopus sporadhi Myers, 1969 (Crustacea: Amphipoda: Aoridae) in the Gulf of Naples (Italy) with a review on its distribution and ecology. Mediterr. Mar. Sci. 2013, 4, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Esposito, V.; Giacobbe, S.; Cosentino, A.; Minerva, C.S.; Romeo, T.; Canese, S.; Andaloro, F. Distribution and ecology of the tube-dweller Ampelisca ledoyeri (Amphipoda: Ampeliscidae) associated to the hydrothermal field off Panarea Island (Tyrrhenian Sea, Mediterranean). Mar. Biodivers. 2015, 45, 763–768. [Google Scholar] [CrossRef]
- Maldonado, M.; López-Acosta, M.; Sitjà, C.; Aguilar, R.; García, S.; Vacelet, J. A giant foraminifer that converges to the feeding strategy of carnivorous sponges: Spiculosiphon oceana sp. nov. (foraminifera, Astrorhizida). Zootaxa 2013, 3669, 571–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, V.; Canese, S.; Scotti, G.; Bo, M.; De Vittor, C.; Andaloro, F.; Romeo, T. Spiculosiphon oceana (foraminifera) and its affinity to intermediate stress conditions in the Panarea hydrothermal complex (Mediterranean Sea). Mar. Biodivers. Rec. 2019, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, K.S.; Wallberg, A.; Jondelius, U. New species of Acoela from the Mediterranean, the Red Sea, and the South Pacific. Zootaxa 2011, 2867, 1–31. [Google Scholar] [CrossRef]
- Giangrande, A.; Gambi, M.C.; Micheli, F.; Kroeker, K.J. Fabriciidae (Annelida, Sabellida) from a naturally acidified coastal system (Italy) with description of two new species. J. Mar. Biol. Assoc. UK 2014, 94, 1417–1427. [Google Scholar] [CrossRef]
- Cookson, L.J.; Lorenti, M. A new species of limnoriid seagrass borer (Isopoda) from the Mediterranean. Crustaceana 2001, 74, 339–346. [Google Scholar]
- Giangrande, A.; Putignano, M.; Licciano, M.; Gambi, M.C. The Pandora’s box: Morphological diversity within the genus Amphiglena Claparède, 1864 (Sabellidae, Annelida) in the Mediterranean Sea, with description of nine new species. Zootaxa 2021, 4949, zootaxa-4949. [Google Scholar] [CrossRef]
- Colangelo, M.A.; Bertasi, F.; Dall’Olio, P.; Ceccherelli, V.H. Meiofaunal Biodiversity on Hydrothermal Seepage off Panarea (Aeolian Islands, Tyrrhenian Sea). In Mediterranean Ecosystems; Faranda, F.M., Guglielmo, L., Spezie, G., Eds.; Springer: Milano, Italy, 2001. [Google Scholar] [CrossRef]
- Baldrighi, E.; Zeppilli, D.; Appolloni, L.; Donnarumma, L.; Chianese, E.; Russo, G.F.; Sandulli, R. Meiofaunal communities and nematode diversity characterizing the Secca delle Fumose shallow vent area (Gulf of Naples, Italy). PeerJ 2020, 8, e9058. [Google Scholar] [CrossRef]
- Appolloni, L.; Zeppilli, D.; Donnarumma, L.; Baldrighi, E.; Chianese, E.; Russo, G.F.; Sandulli, R. Seawater Acidification Affects Beta-Diversity of Benthic Communities at a Shallow Hydrothermal Vent in a Mediterranean Marine Protected Area (Underwater Archaeological Park of Baia, Naples, Italy). Diversity 2020, 12, 464. [Google Scholar] [CrossRef]
- Milazzo, M.; Cattano, C.; Alonzo, S.H.; Foggo, A.; Gristina, M.; Rodolfo-Metalpa, R.; Sinopoli, M.; Spatafora, D.; Stiver, K.A.; Hall-Spencer, J.M. Ocean acidification affects fish spawning but not paternity at CO2 seeps. Proc. Biol. Sci. 2016, 283, 20161021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattano, C.; Claudet, J.; Domenici, P.; Milazzo, M. Living in a high CO2 world: A global meta-analysis shows multiple trait-mediated fish responses to ocean acidification. Ecol. Monogr. 2018, 88, 320–335. [Google Scholar] [CrossRef]
- Mirasole, A.; Signa, G.; Gianguzza, P.; Bonaviri, C.; Mazzola, A.; Vizzini, S. Fish assemblages cope with ocean acidification in a shallow volcanic CO2 vent benefiting from an adjacent recovery area. Mar. Environ. Res. 2020, 157, 104851. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Crego, B.; Vizzini, S.; Califano, G.; Massa-Gallucci, A.; Andolina, C.; Gambi, M.; Santos, R. Resistance of seagrass habitats to ocean acidification via altered interactions in a tri-trophic chain. Sci. Rep. 2020, 10, 5103. [Google Scholar] [CrossRef] [Green Version]
- Giaccone, G. Associazioni algali e fenomeni secondari di vulcanismo nelle acque marine di Vulcano (Mar Tirreno). G. Bot. Ital. 1969, 103, 353–366. [Google Scholar] [CrossRef]
- Mazzella, L.; Buia, M.C. Strategie evolutive nelle fanerogame marine del Mediterraneo. Nova Thalass. 1986, 8, 651. [Google Scholar]
- Aliani, S.; Bianchi, C.N.; Cocito, S.; Dando, P.R.; Meloni, R.; Morri, C.; Niemeyer, A.; Peirano, A.; Ziebis, W. A map of seagrass meadows in Palaeochori Bay (Milos Island, Greece), a marine area with hydrothermal activity. Rapp. Proces-Verbaux Réunions Comm. Int. L’Explor. Sci. Mer Méditerranée 1998, 35, 512–513. [Google Scholar]
- Porzio, L.; Buia, M.C.; Hall-Spencer, J.M. Effects of ocean acidification on macroalgal communities. J. Exp. Mar. Biol. Ecol. 2011, 400, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Linares, C.; Vidal, M.; Canals, M.; Kersting, D.K.; Amblas, D.; Aspillaga, E.; Cebrián, E.; Delgado-Huertas, A.; Díaz, D.; Garrabou, J.; et al. Persistent natural acidification drives major distribution shifts in marine benthic ecosystems. Proc. R. Soc. B 2015, 282, 2015058720150587. [Google Scholar] [CrossRef]
- Bang, C.; Dagan, T.; Deines, P.; Dubilier, N.; Duschl, W.J.; Fraune, S.; Hentschel, U.; Hirt, H.; Hülter, N.; Lachnit, T.; et al. Metaorganisms in extreme environments: Do microbes play a role in organismal adaptation? Zoology 2018, 127, 1–19. [Google Scholar] [CrossRef]
- Reysenbach, A.L.; Banta, A.B.; Boone, D.R.; Cary, S.C.; Luther, G.W. Microbial essentials at hydrothermal vents. Nature 2000, 404, 835. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, B.N.; Sylvan, J.B.; Knab, N.J.; Edwards, K.J. Microbial ecology of the dark ocean above, at, and below the seafloor 2011. Microbiol. Mol. Biol. Rev. 2011, 75, 361–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milojevic, T.; Weckwerth, W. Molecular Mechanisms of Microbial Survivability in Outer Space: A Systems Biology Approach. Front. Microbiol. 2020, 15, 923. [Google Scholar] [CrossRef]
- Yucel, M.; Sievert, S.M.; Vetriani, C.; Foustoukos, D.I.; Giovannelli, D.; LeBris, N. Eco-geochemical dynamics of ashallow-water hydrothermal vent system at Milos Island, Aegean Sea(Eastern Mediterranean). Chem. Geol. 2013, 356, 11–20. [Google Scholar] [CrossRef]
- Price, R.E.; Amend, J.P.; Pichler, T. Enhanced geochemical gradientsinamarineshallow-water hydrothermal system: Unusual arsenic speciation in horizontal and vertical pore water profiles. Appl. Geochem. 2007, 22, 2595–2605. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Lentini, V.; Gugliandolo, C.; Italiano, F.; Cousin, S.; Stackebrandt, E. Bacterial and archaeal populations at two shallow hydrothermal vents off Panarea Island (Eolian Islands, Italy). Extremophiles 2008, 13, 199–212. [Google Scholar] [CrossRef]
- Sievert, S.M.; Kuever, J.; Muyzer, G. Identification of 16Sribosomal DNA-defined bacterial populations at a shallow submarine hydrothermal vent near Milos Island (Greece). Appl. Environ. Microbiol. 2000, 66, 3102–3109. [Google Scholar] [CrossRef] [Green Version]
- Alfredsson, G.A.; Kristjansson, J.K.; Hjorleifsdottir, S.; Stetter, K.O. Rhodothermus marinus, gen. nov., sp. nov., a thermophilic, halophilic bacterium from submarine hot springs in Iceland. J. Gen. Microbiol. 1988, 134, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Perez-Rodriguez, I.; Rawls, M.; Coykendall, D.K.; Foustoukos, D.I. Deferrisoma paleochoriense sp. nov., athermophilic, iron (III)-reducing bacteriumfroma shallow-water hydrothermal vent in the Mediterranean Sea. Int. J. Syst. Evol. Microb. 2015, 66, 830–836. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Giuliano, L.; Cappello, S.; Denaro, R.; Golyshin, P.N. Microbial Community of a Hydrothermal Mud Vent Underneath the Deep-Sea Anoxic Brine Lake Urania (Eastern Mediterranean). Orig. Life Evol. Biosph. 2007, 37, 177–188. [Google Scholar] [CrossRef]
- L’Haridon, S.; Chalopin, M.; Colombo, D.; Toffin, L. Methanococcoides vulcani sp. nov., a marine methylotrophic methanogen that uses betaine, choline and N,N-dimethylethanolamine for methanogenesis, isolated from a mud volcano, and emended description of the genus Methanococcoides. Int. J. Syst. Evol. Microb. 2014, 64, 1978–1983. [Google Scholar] [CrossRef]
- Golyshina, O.V.; Yakimov, M.M.; Lünsdorf, H.; Ferrer, M.; Nimtz, M.; Timmis, K.; Brian, W.; Tindall, J.V.; Golyshin, P.N. Acidiplasma aeolicum gen. nov., sp. nov., a euryarchaeon of the family Ferroplasmaceae isolated from a hydrothermal pool, and transfer of Ferroplasma cupricumulans to Acidiplasma cupricumulans comb. Nov. Int. J. Syst. Evol. Microb. 2009, 59, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.J.; Lewis, H.; Trejo, E.; Winston, V.; Evilia, C. Protein adaptations in archaeal extremophiles. Archaea 2013, 2013, 373275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Yogo, K.; Furuike, S.; Sutoh, K.; Kikuchi, A.; Kinosita, K., Jr. Direct observation of DNA overwinding by reverse gyrase. Proc. Natl. Acad. Sci. USA 2015, 16, 7495–7500. [Google Scholar] [CrossRef] [Green Version]
- Siliakus, M.F.; van der Oost, J.; Kengen, S.W.M. Adaptations of archaeal and bacterial membranes to variations in temperature, pH and pressure. Extremophiles 2017, 21, 651–670. [Google Scholar] [CrossRef]
- Kashefi, K.; Lovley, D.R. Extending the upper temperature limit for life. Science 2003, 301, 934. [Google Scholar] [CrossRef] [PubMed]
- Mora, M.; Bellack, A.; Ugele, M.; Hopf, J.; Wirth, R. The temperature gradient- forming device, an accessory unit for normal light microscopes to study the biology of hyperthermophilic microorganisms. Appl. Environ. Microbiol. 2014, 80, 4764–4770. [Google Scholar] [CrossRef] [Green Version]
- Reysenbach, A.L.; Shock, E. Merging genomes with geochemistry in hydrothermal ecosystems. Science 2002, 296, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T.A. Alkaline pH homeostasis in bacteria: New insights. Nov. Biochim. Biophys. Acta 2005, 30, 67–88. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Thermodynamic limits to microbial life at high salt concentrations. Environ. Microbiol. 2011, 13, 1908–1923. [Google Scholar] [CrossRef]
- Reveillaud, J.; Reddington, E.; McDermott, J.; Algar, C.; Meyer, J.; Sylva, S.; Huber, J.A. Subseafloor microbial communities in hydrogen- rich vent fluids from hydrothermal systems along the Mid- Cayman Rise. Environ. Microbiol. 2016, 18, 1970–1987. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. AT3-60 Shipboard Party. An off-axis hydrothermal vent field near themid-atlantic ridgeat 30 N. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Hugler, M.; Sievert, S.M. Beyond the Calvin Cycle: Autotrophic Carbon Fixation in theOcean. Annu. Rev. Mar. Sci. 2011, 3, 261–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konn, C.; Charlou, J.-L.; Donval, J.-P.; Holm, N.; Dehairs, F.; Bouillon, S. Hydrocarbons and oxidized organic compounds in hydrothermal fluids from Rainbow and Lost City ultramafic-hosted vents. Chem. Geol. 2009, 258, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, Z.; Zeng, L.; Dong, C.; Shao, Z. The oxidation of hydrocarbons by diverse heterotrophic and mixotrophic bacteria that inhabit deep-sea hydrothermal ecosystems. ISME J. 2020, 14, 1994–2006. [Google Scholar] [CrossRef]
- Galambos, D.; Anderson, R.E.; Reveillaud, J.; Huber, J.A. Genome-resolved metagenomics and metatranscriptomics reveal niche differentiation in functionally redundant microbial communities at deep-sea hydrothermal vents. Environ. Microbiol. 2019, 21, 4395–4410. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.; Liu, K.; Jiao, N.; Zhang, Y.; Chen, C. Functional metagenomic investigations of microbial communities in a shallow-sea hydrothermal system. PLoS ONE 2013, 8, e72958. [Google Scholar] [CrossRef] [Green Version]
- Amend, J.; Mc Collom, T.; Hentscher, M.; Bach, W. Catabolic and anabolic energy for chemolithoautotrophs in deep-sea hydrothermal systems hosted in different rock types. Geochim. Cosmochim. Acta 2011, 75, 5736–5748. [Google Scholar] [CrossRef]
- Huber, J.A.; Mark Welch, D.B.; Morrison, H.G.; Huse, S.M.; Neal, P.R.; Butterfield, D.A.; Sogin, M.L. Microbial population structures in the deep marine biosphere. Science 2007, 318, 97–100. [Google Scholar] [CrossRef] [Green Version]
- Lentini, V.; Gugliandolo, C.; Bunk, B.; Overmann, J.; Maugeri, T.L. Diversity of Prokaryotic Community at a Shallow Marine Hydrothermal Site Elucidated by Illumina Sequencing Technology. Curr. Microbiol. 2014, 69, 457–466. [Google Scholar] [CrossRef]
- Dávila-Ramos, S.; Estradas-Romero, A.; Prol-Ledesma, R.M.; Juárez-López, K. Bacterial Populations (First Record) at Two Shallow Hydrothermal Vents of the Mexican Pacific West Coast. Geomicrobiol. J. 2015, 32, 657–665. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Italiano, F.; Maugeri, T.; Inguaggiato, S.; Caccamo, D.; Amend, J.P. Submarine Hydrothermal Vents of the Aeolian Islands: Relationship between Microbial Communities and Thermal Fluids. Geomicrobiol. J. 1999, 16, 105–117. [Google Scholar] [CrossRef]
- Miranda, P.J.; Mc Lain, N.K.; Hatzenpichler, R.; Orphan, V.J.; Dillon, J.G. Characterization of Chemosynthetic Microbial Mats Associated with Intertidal Hydrothermal Sulphur Vents in White Point, San Pedro, CA, USA. Front. Microbiol. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kuever, J.; Sievert, S.M.; Stevens, H.; Brinkhoff, T.; Muyzer, G. Microorganisms of theoxidativeand reductivepart of thesulphur cycleat ashallow-water hydrothermal vent in the Aegean Sea (Milos, Greece). Cah. Biol. Mar. 2002, 43, 413–416. [Google Scholar]
- Sievert, S.M.; Brinkhoff, T.; Muyzer, G.; Ziebis, W.; Kuever, J. Spatial heterogeneity of bacterial populations along an environmental gradient at a shallow submarine hydrothermal vent near Milos Island (Greece). Appl. Environ. Microbiol. 1999, 65, 3834–3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dando, P.R.; Thomm, M.; Arab, H.; Brehmer, M.; Hooper, L.E.; Jochimsen, B.; Fowler, S.W. Microbiology of shallow hydrothermal sites off palaeochori bay, milos (hellenic volcanic arc). Cah. Biol. Mar. 1998, 39, 369–372. [Google Scholar]
- Zhang, Y.; Zhao, Z.; Chen, C.T.A.; Tang, K.; Su, J.; Jiao, N. Sulphur Metabolizing Microbes Dominate Microbial Communities in Andesite-Hosted Shallow-Sea Hydrothermal Systems. PLoS ONE 2012, 7, e44593. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.J.; Engel, A.S.; Porter, M.L.; Takai, K. The versatile e-proteobacteria: Key players in sulphidic habitats. Nat. Rev. Microbiol. 2006, 4, 458–468. [Google Scholar] [CrossRef]
- Sievert, S.; Vetriani, C. Chemoautotrophy at Deep-Sea vents: Past, present, and future. Oceanography 2012, 25, 218–233. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Maugeri, T.L. Temporal Variations in Heterotrophic Mesophilic Bacteria from a Marine Shallow Hydrothermal Vent off the Island of Vulcano (Eolian Islands, Italy). Microb. Ecol. 1998, 36, 13–22. [Google Scholar] [CrossRef]
- Kamenev, G.M.; Fadeev, V.I.; Selin, N.I.; Tarasov, V.G.; Malakhov, V.V. Composition and distribution of macro- and meio benthos around sublittoral hydrothermal vents in the Bay of Plenty, New Zealand. New Zealand. J. Mar. Freshw. Res. 1993, 27, 407–418. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Lentini, V.; Spano, A.; Gugliandolo, C. Abundance and Diversity of Picocyanobacteria in Shallow Hydrothermal Vents of Panarea Island (Italy). Geomicrobiol. J. 2013, 30, 93–99. [Google Scholar] [CrossRef]
- Thornburg, C.C.; Zabriskie, T.M.; McPhail, K.L. Deep-sea hydrothermal vents: Potential hot spots for natural products discovery? J. Nat. Prod. 2010, 73, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Italiano, F.; Maugeri, T.L. The submarine hydrothermal system of Panarea (Southern Italy): Biogeochemical processes at the thermal fluids-sea bottom interface. Ann. Geophys. 2006, 49, 783–792. [Google Scholar] [CrossRef]
- Vetriani, C.; Voordeckers, J.W.; Crespo-Medina, M.; O’Brien, C.E.; Giovannelli, D.; Lutz, R.A. Deep-sea hydrothermal vent Epsilonproteobacteria encode a conserved and widespread nitrate reduction pathway (Nap). ISME J. 2014, 8, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, S.; Smedile, F.; Giovannelli, D.; Vetriani, C. Metaproteogenomic Profiling of Chemosynthetic Microbial Biofilms Reveals Metabolic Flexibility During Colonization of a Shallow-Water Gas Vent. Front. Microbiol. 2021, 6, 638300. [Google Scholar] [CrossRef]
- Emerson, D.; Rentz, J.A.; Lilburn, T.G.; Davis, R.E.; Aldrich, H.; Chan, C.; Moyer, C.L. A novel lineage of Proteobacteria involved in for- mation of marine Fe-oxidizing microbial mat communities. PLoS ONE 2007, 2, e667. [Google Scholar] [CrossRef] [Green Version]
- McAllister, S.M.; Moore, R.M.; Gartman, A.; Luther, G.W., III; Emerson, D.; Chan, C.S. The Fe(II)-oxidizing Zetaproteobacteria: Historical, ecological and genomic perspectives. FEMS Microbiol. Ecol. 2019, 95, fiz015. [Google Scholar] [CrossRef] [Green Version]
- DeLong, E.F.; Taylor, L.T.; Marsh, T.L.; Preston, C.M. Visualization and enumeration of marine planktonic archaea and bacteria by using polyribonucleotide probes and fluorescent in situ hybridization. Appl. Environ. Microbiol. 1999, 65, 5554–5563. [Google Scholar] [CrossRef] [Green Version]
- Biddanda, B.; Ogdahl, M.; Cotner, J. Dominance of bacterial metabolism in oligotrophic relative to eutrophic waters. Limnol. Oceanogr. 2001, 46, 730–739. [Google Scholar] [CrossRef]
- Teira, E.; Reinthaler, T.; Pernthaler, A.; Pernthaler, J.; Herndl, G.J. Combining catalyzed reporter deposi- tion fluorescence in situ hybridization and microautoradi- ography to detect substrate utilization by bacteria and Archaea in deep ocean. Appl. Environ. Microbiol. 2004, 70, 4411–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipp, J.S.; Morono, Y.; Inagaki, F.; Hinrichs, K.U. Significant contribution of Archaea to extant biomass in marine subsurface sediments. Nature 2008, 454, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Karner, M.B.; DeLong, E.F.; Karl, D.M. Archaeal dominance in the mesopelagic zone of the Pacific Ocean. Nature 2001, 409, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Flores, G.E.; Shakya, M.; Meneghin, J.; Yang, Z.K.; Seewald, J.S.; Wheat, C.G.; Podar, M.; Reysenbach, A.L. Inter-field variability in the microbial communities of hydrothermal vent deposits from a back-arc basin. Geobiology 2012, 10, 333–346. [Google Scholar] [CrossRef]
- Leoni, C.; Volpicella, M.; Fosso, B.; Manzari, C.; Piancone, E.; Dileo, M.C.; Ceci, L.R. A differential metabarcoding approach to describe taxonomy profiles of bacteria and archaea in the saltern of margherita di savoia (Italy). Microorganisms 2020, 8, 936. [Google Scholar] [CrossRef]
- Fagorzi, C.; Del Duca, S.; Venturi, S.; Chiellini, C.; Bacci, G.; Fani, R.; Tassi, F. Bacterial Communities from Extreme Environments: Vulcano Island. Diversity 2019, 11, 140. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, D.; Maugeri, T.L.; Gugliandolo, C. Identification of thermophilic and marine bacilli from shallow thermal vents by restriction analysis of their amplified 16S rDNA. J. Appl. Microb. 2001, 91, 520–524. [Google Scholar] [CrossRef]
- Caccamo, D.; Gugliandolo, C.; Stackebrandt, E.; Maugeri, T.L. Bacillus vulcani sp. nov., a novel thermophilic species isolated from a shallow marine hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2020, 50, 2009–2012. [Google Scholar] [CrossRef] [Green Version]
- Lentini, V.; Gugliandolo, C.; Maugeri, T.L. Identification of enzyme-producing thermophilic bacilli isolated from marine vents of Aeolian Islands (Italy). Ann. Microbiol. 2007, 57, 355–361. [Google Scholar] [CrossRef]
- Eythorsdottir, A.; Omarsdottir, S.; Einarsson, H. Antimicrobial Activity of Marine Bacterial Symbionts Retrieved from Shallow Water Hydrothermal Vents. Mar. Biotech. 2016, 18, 293–300. [Google Scholar] [CrossRef]
- Giordano, D. Bioactive Molecules from Extreme Environments. Mar. Drugs 2020, 18, 640. [Google Scholar] [CrossRef] [PubMed]
- Illanes, A.; Cauerhff, A.; Wilson, L.; Castro, G.R. Recent trends in biocatalysis engineering. Bioresour. Technol. 2012, 115, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, A.; Harani Devi, P.; Chattopadhyay, S.; Kavitha, M. Commercial applications of microbial enzymes. In Microbial Enzymes: Roles and Applications in Industries; Arora, N.K., Mishra, J., Mishra, V., Eds.; Springer: Singapore, 2020; pp. 137–184. [Google Scholar]
- Anastas, P.; Eghbali, N. Green chemistry: Principles and practice. Chem. Soc. Rev. 2010, 39, 301–312. [Google Scholar] [CrossRef]
- Sheldon, R.A. Green chemistry and resource efficiency: Towards a green economy. Green Chem. 2016, 18, 3180–3183. [Google Scholar] [CrossRef]
- Sysoev, M.; Grötzinger, S.W.; Renn, D.; Eppinger, J.; Rueping, M.; Karan, R. Bioprospecting of Novel Extremozymes From Prokaryotes—The Advent of Culture-Independent Methods. Front. Microbiol. 2021, 12, 630013. [Google Scholar] [CrossRef]
- Gatti-Lafranconi, P.; Natalello, A.; Rehm, S.; Doglia, S.M.; Pleiss, J.; Lotti, M. Evolution of stability in a cold-active enzyme elicits specificity relaxation and highlights substrate-related effects on temperature adaptation. J. Mol. Biol. 2010, 395, 155–166. [Google Scholar] [CrossRef]
- Siddiqui, K.S.; Parkin, D.M.; Curmi, P.M.; De Francisci, D.; Poljak, A.; Barrow, K.; Noble, M.H.; Trewhella, J.; Cavicchioli, R. A novel approach for enhancing the catalytic efficiency of a protease at low temperature: Reduction in substrate inhibition by chemical modification. Biotechnol. Bioeng. 2009, 103, 676–686. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Dasgupta, A.K.; Chakrabarti, K. Enhanced functionality and stabilization of a cold active laccase using nanotechnology based activation-immobilization. Bioresour. Technol. 2015, 179, 573–584. [Google Scholar] [CrossRef]
- Sarmiento, F.; Peralta, R.; Blamey, J.M. Cold and hot extremozymes: Industrial relevance and current trends. Front. Bioeng. Biotechnol. 2015, 3, 148. [Google Scholar] [CrossRef] [Green Version]
- Bruins, M.E.; Janssen, A.E.; Boom, R.M. Thermozymes and their applications: A review of recent literature and patents. Appl. Biochem. Biotechnol. 2001, 90, 155–186. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. A Polyphasic Taxonomic Study of Thermophilic Bacilli from Shallow, Marine Vents. System. Appl. Microbiol. 2001, 24, 572–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirjian, D.; Moris Varas, F.; Cassidy, C.S. Enzymes from extremophiles. Curr. Opin. Chem. Biol. 2001, 5, 144–151. [Google Scholar] [CrossRef]
- Yavuz, E.; Gunes, H.; Harsa, S.; Yenidunya, A.F. Identification of extracellular enzyme producing thermophilic bacilli from Balcova (Agamemnon) geothermal site by ITS rDNA RFLP. J. Appl. Microbiol. 2004, 97, 810–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, T.D. The value of basic research: Discovery of Thermus aquaticus and other extreme thermophiles. Genetics 1997, 146, 1207–1210. [Google Scholar] [CrossRef]
- Irwin, J.A.; Baird, A.W. Extremophiles and their application to veterinary medicine. Ir. Vet. J. 2004, 57, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Damiano, V.; Bocchini, D.; Gomes, E.; Da Silva, R. Application of crude xylanase from Bacillus licheniformis 77-2 to the bleaching of eucalyptus Kraft pulp. World J. Microbiol. Biotechnol. 2003, 19, 139–144. [Google Scholar] [CrossRef]
- Atalah, J.; Cáceres-Moreno, P.; Espina, G.; Blamey, J.M. Thermophiles and the applications of their enzymes as new biocatalysts. Bioresour. Technol. 2019, 280, 478–488. [Google Scholar] [CrossRef]
- Jin, M.; Gai, Y.; Guo, X.; Hou, Y.; Zeng, R. Properties and applications of extremozymes from deep-sea extremophilic microorganisms: A mini review. Mar. Drugs 2019, 17, 656. [Google Scholar] [CrossRef] [Green Version]
- Di Donato, P.; Buono, A.; Poli, A.; Finore, I.; Abbamondi, G.R.; Nicolaus, B.; Lama, L. Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses. Sustainability 2019, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial Exopolysaccharides from Extreme Marine Habitats: Production, Characterization and Biological Activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Caccamo, D.; Gugliandolo, C.; Zammuto, V.; Magazù, S. Thermal properties of an exopolysaccharide produced by a marine thermotolerant Bacillus licheniformis by ATR-FTIR spectroscopy. J. Biol. Macromol. 2020, 145, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Nicolaus, B.; Panico, A.; Manca, M.C.; Lama, L.; Gambacorta, A.; Maugeri, T.L.; Gugliandolo, C.; Caccamo, D. A Thermophilic Bacillus Isolated From an Eolian Shallow Hydrothermal Vent, Able to Produce Exopolysaccharides. Syst. Appl. Microbiol. 2000, 23, 426–432. [Google Scholar] [CrossRef]
- Nicolaus, B.; Lama, L.; Panico, A.; Schiano Moriello, V.; Romano, I.; Gambacorta, A. Production and characterization of exopolysaccharides excreted by thermophilic bacteria from shallow, marine hydrothermal vents of flegrean areas (Italy). Syst. Appl. Microbiol. 2002, 25, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Rougeaux, H.; Kervarec, N.; Pichon, R.; Guerzennec, J. Structure of the exopolysaccharide of Vibrio diabolicus isolated from a deep–sea hydrothermal vent. Carbohydr. Res. 1999, 322, 40–45. [Google Scholar] [CrossRef]
- Delbarre-Ladrat, C.; Salas, M.L.; Sinquin, C.; Zykwinska, A.; Colliec-Jouault, S. Bioprospecting for Exopolysaccharides from Deep-Sea Hydrothermal Vent Bacteria: Relationship between Bacterial Diversity and Chemical Diversity. Microorganisms 2017, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Guezennec, J.G.; Pignet, P.; Raguenes, G.; Deslandes, E.; Lijour, Y.; Gentric, E. Preliminary chemical characterization of unusual eubacterial exopolysaccharides of deep-sea origin. Carbohydr. Polym. 1994, 24, 287–294. [Google Scholar] [CrossRef]
- König, H.; Skorko, R.; Zillig, W.; Reiter, W.-D. Glycogen in thermoacidophilic archaebacteria of the genera Sulfolobus, Thermoproteus, Desulphurococcus and Thermococcus. Arch. Microbiol. 1982, 132, 297–303. [Google Scholar] [CrossRef]
- Antón, J.; Meseguer, I.; Rodrı’guez-Valera, F. Production of an extracellular polysaccharide by Haloferax mediterranei. Appl. Environ. Microbiol. 1988, 54, 2381–2386. [Google Scholar] [CrossRef] [Green Version]
- Sowers, K.R.; Gunsalus, R.P. Adaptation for growth at various saline concentrations by the archaebacterium Methanosarcina thermophila. J. Bacteriol. 1988, 170, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- Rinker, K.D.; Kelly, R.M. Growth Physiology of the Hyperthermophilic Archaeon Thermococcus litoralis: Development of a Sulphur-Free Defined Medium, Characterization of an Exopolysaccharide, and Evidence of Biofilm Formation. Appl. Environ. Microbiol. 1996, 62, 4478–4485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raguenes, G.; Pignet, P.; Gauthier, G.; Peres, A.; Christen, R.; Gougeaux, H.; Barbier, G.; Guezennec, J. Description of a new polymer-secreting bacterium from a deep-sea hydrothermal vent, Alteromonas macleodii subsp. fijiensis, and preliminary characterization of the polymer. Appl. Environ. Microbiol. 1995, 62, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaus, B.; Manca, M.C.; Romano, I.; Lama, L. Production of an exopolysaccharide from two thermophilic archaea belonging to the genus Sulfolobus. FEMS Microbiol. Lett. 1993, 109, 203–206. [Google Scholar] [CrossRef]
- Alcaide, M.; Tchigvintsev, A.; Martínez-Martínez, M.; Popovic, A.; Reva, O.N.; Lafraya, Á.; Bargiela, R.; Nechitaylo, T.Y.; Matesanz, R.; Cambon-Bonavita, M.A.; et al. MAMBA Consortium. Identification and characterization of carboxyl esterases of gill chamber-associated microbiota in the deep-sea shrimp Rimicaris exoculata using functional metagenomics. Appl. Environ. Microbiol. 2014, 81, 2125–2136. [Google Scholar] [CrossRef] [Green Version]
- Egorova, K.; Antranikian, G. Industrial relevance of thermophilic Archaea. Curr. Opin. Microbiol. 2005, 8, 649–655. [Google Scholar] [CrossRef]
- Trincone, A. Enzymatic Processes in Marine Biotechnology. Mar. Drugs 2017, 15, 93. [Google Scholar] [CrossRef] [Green Version]
- Bornscheuer, U.T. Microbial carboxyl esterases: Classification, properties and application in biocatalysis. FEMS Microbiol. Rev. 2002, 26, 73–81. [Google Scholar] [CrossRef]
- Frazzetto, G. White biotechnology: The application of biotechnology to industrial production holds many promises for sustainable development, but many products still have to pass the test of economic viability. EMBO 2003, 4, 835–837. [Google Scholar] [CrossRef]
- Panda, T.; Gowrishankar, B.S. Production and applications of esterases. Appl. Microbiol. Biotechnol. 2005, 67, 160–169. [Google Scholar] [CrossRef]
- Kennedy, J.; O’Leary, N.D.; Kiran, G.S.; Morrissey, J.P.; O’Gara, F.; Selvin, J.; Dobson, A.D.W. Functional metagenomic strategies for thediscovery of novel enzymes and biosurfactants with biotechnological applications from marine ecosystems. J. Appl. Microbiol. 2011, 111, 787–799. [Google Scholar] [CrossRef]
- Raddadi, N.; Cherif, A.; Da_onchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biot. 2015, 99, 7907–7913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arena, A.; Gugliandolo, C.; Stassi, G.; Pavone, B.; Iannello, D.; Bisignano, G.; Maugeri, T.L. An exopolysaccharide produced by Geobacillus thermodenitrificans strain B3-72: Antiviral activity on immunocompetent cells. Immunol. Lett. 2009, 123, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Arena, A.; Maugeri, T.L.; Pavone, B.; Jannello, D.; Gugliandolo, C.; Bisignano, G. Antiviral and immunoregulatory effect of a novel exopolysaccharide from a marine thermotolerant Bacillus licheniformis. Int. Immunopharmacol. 2006, 6, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Caccamo, M.T.; Zammuto, V.; Gugliandolo, C.; Madeleine-Perdrillat, C.; Spanò, A.; Magazù, S. Thermal restraint of a bacterial exopolysaccharide of shallow vent origin. Int. J. Biol. Macromol. 2018, 114, 649–655. [Google Scholar] [CrossRef]
- Scala, A.; Piperno, A.; Hada, A.; Astilean, S.; Vulpoi, A.; Ginestra, G.; Marino, A.; Nostro, A.; Zammuto, V.; Gugliandolo, C. Marine bacterial exopolymers-mediated green synthesis of noble metal nanoparticles with antimicrobial properties. Polymers 2019, 11, 1157. [Google Scholar] [CrossRef] [Green Version]
- Raguénès, G.H.C.; Peres, A.; Ruimy, R.; Pignet, P.; Christen, R.; Loaec, M.; Rougeaux, H.; Barbier, G.; Guezennec, J.G. Alteromonas infernus sp. nov., a new polysaccharide-producing bacterium isolated from a deep-sea hydrothermal vent. J. Appl. Microbiol. 1997, 82, 422–430. [Google Scholar] [CrossRef]
- Roger, O.; Kervarec, N.; Ratiskol, J.; Colliec-Jouault, S.; Chevolot, L. Structural studies of the main exopolysaccharide produced by the deep-sea bacterium Alteromonas infernus. Carbohydr. Res. 2004, 339, 2371–2380. [Google Scholar] [CrossRef] [Green Version]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Extracellular polymeric substances with metal adsorption capacity produced by Pseudoalteromonas sp. MER144 from Antarctic seawater. Environ. Sci. Pollut. Res. 2017, 25, 4667–4677. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Finore, I.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Production and biotechnological potential of extracellular polymeric substances from sponge-associated Antarctic bacteria. Appl. Environ. Microbiol. 2018, 84, e01624-17. [Google Scholar] [CrossRef] [Green Version]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from Marine and Marine Extremophilic Bacteria: Structures, Properties, Ecological Roles and Applications. Mar. Drugs 2018, 16, 69. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzo, C.; Arcadi, E.; Calogero, R.; Sciutteri, V.; Consoli, P.; Esposito, V.; Canese, S.; Andaloro, F.; Romeo, T. Ecological and Biotechnological Relevance of Mediterranean Hydrothermal Vent Systems. Minerals 2022, 12, 251. https://doi.org/10.3390/min12020251
Rizzo C, Arcadi E, Calogero R, Sciutteri V, Consoli P, Esposito V, Canese S, Andaloro F, Romeo T. Ecological and Biotechnological Relevance of Mediterranean Hydrothermal Vent Systems. Minerals. 2022; 12(2):251. https://doi.org/10.3390/min12020251
Chicago/Turabian StyleRizzo, Carmen, Erika Arcadi, Rosario Calogero, Valentina Sciutteri, Pierpaolo Consoli, Valentina Esposito, Simonepietro Canese, Franco Andaloro, and Teresa Romeo. 2022. "Ecological and Biotechnological Relevance of Mediterranean Hydrothermal Vent Systems" Minerals 12, no. 2: 251. https://doi.org/10.3390/min12020251