
One New Species and Two New Host Records of Apiospora from Bamboo and Maize in Northern Thailand with Thirteen New Combinations

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation and Morphological Characteristic Examination
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
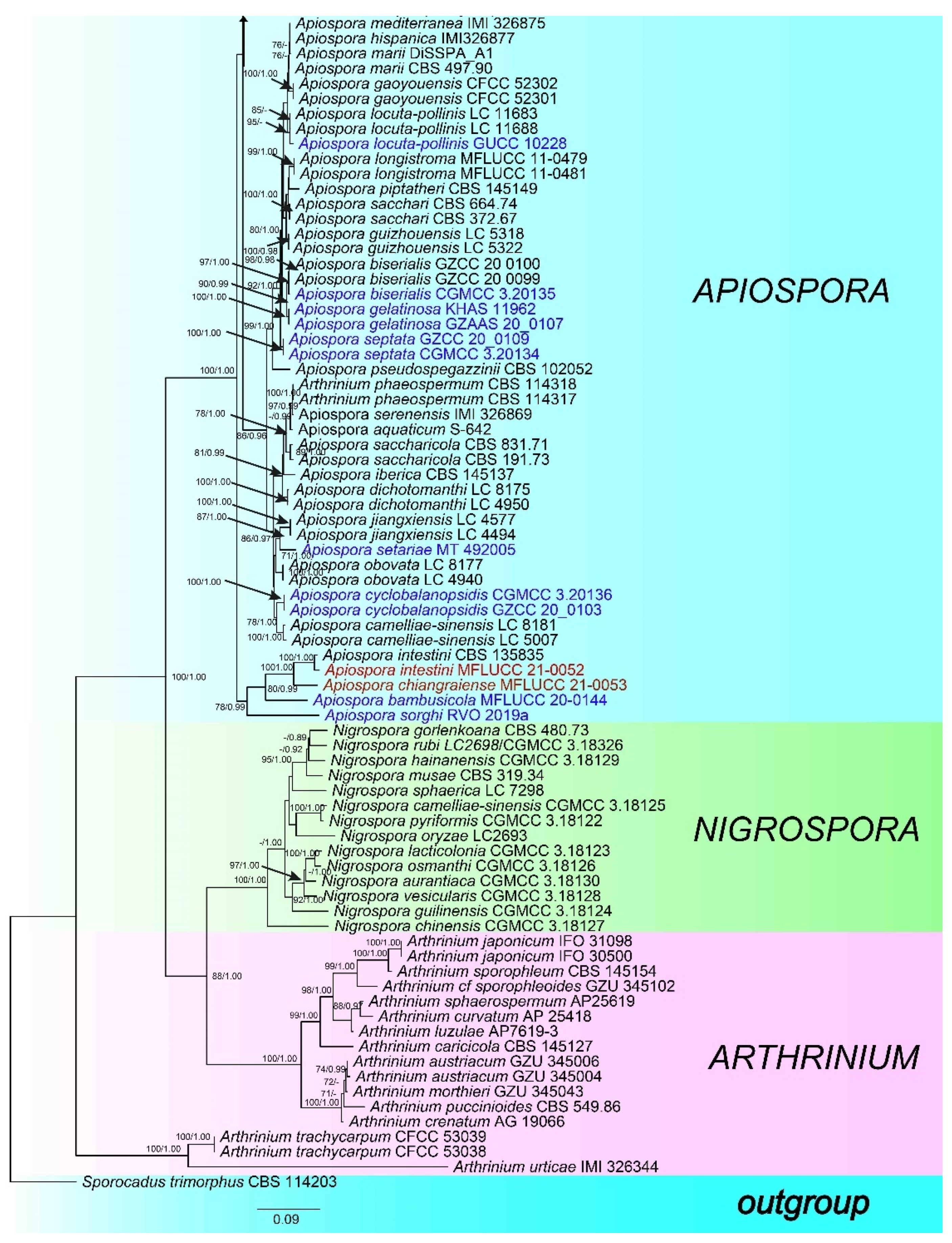
3.1. Phylogeny
3.2. Taxon Treatment
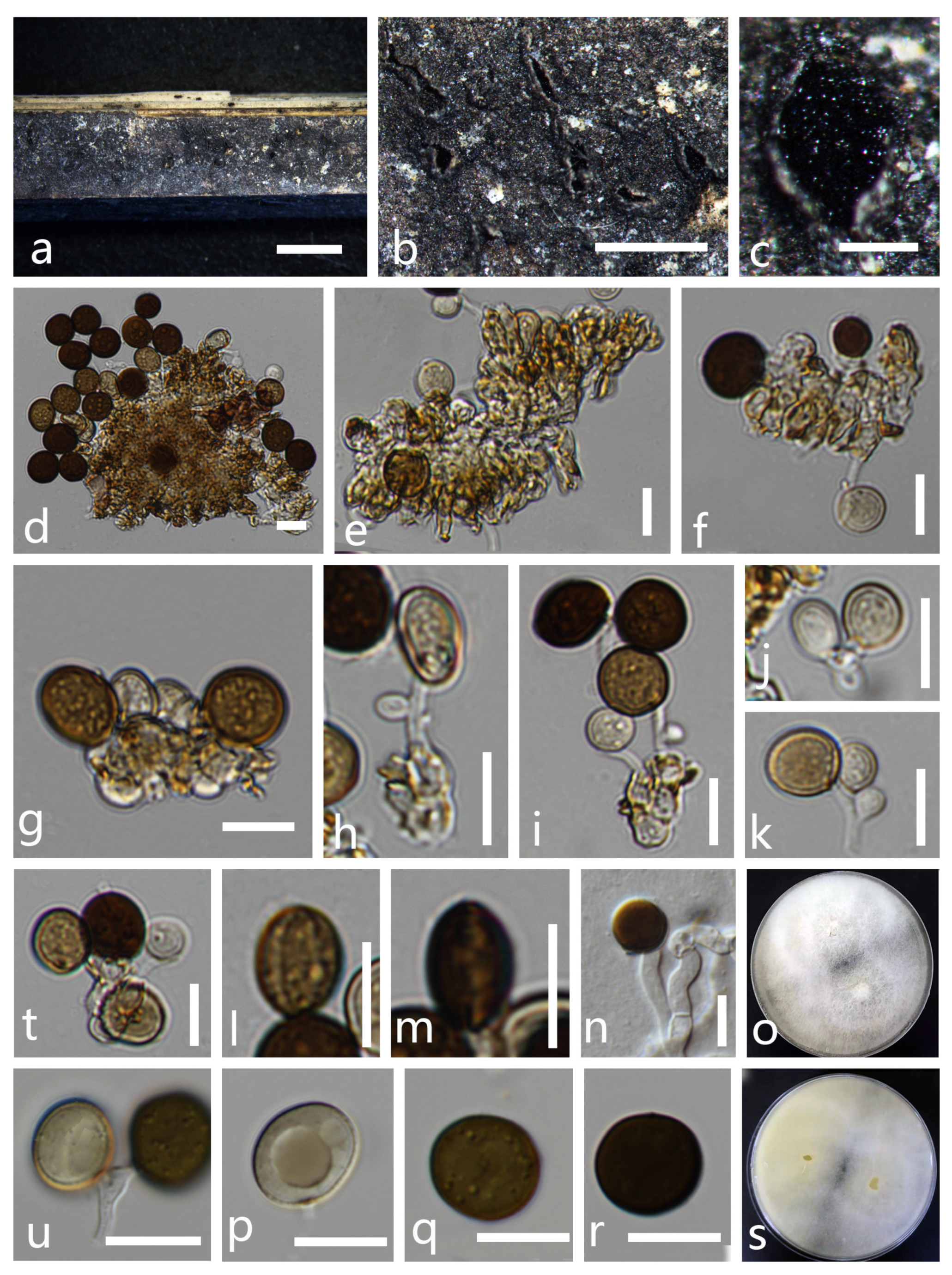
- Apiospora intestini (Kajale, Sonawane and Rohit Sharma) Pintos and P. Alvarado, Fungal Systematics and Evolution 7: 206 (2021) (Figure 3).
- Apiospora rasikravindrae (Shiv M. Singh, L.S. Yadav, P.N. Singh, Rahul Sharma and S.K. Singh) Pintos and P. Alvarado, Fungal Systematics and Evolution 7: 207 (2021) (Figure 4).
3.3. New Combinations
- Apiospora acutiapica (Senan. and Cheew) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora bambusicola (X. Tang, K.D. Hyde and J.C. Kang) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora biserialis (Y. Feng and Z.Y. Liu) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora cordylines (T.Z. Chen, Yong Wang bis and K.D. Hyde) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora cyclobalanopsidis (Y. Feng and Z.Y. Liu) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora euphorbiae (M.B. Ellis) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora gelatinosa (Y. Feng and Z.Y. Liu) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora locuta-pollinis (F. Liu and L. Cai) Pintos and P. Alvarado), X.G. Tian and Tibpromma S., comb. nov.
- Apiospora minutispora (K. Das, S.Y. Lee and H.Y. Jung) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora pseudorasikravindrae (Senan. and Cheew) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora septata (Y. Feng and Jian K. Liu) X.G. Tian and Tibpromma S., comb. nov.
- Apiosporasetariae (Jiang, N.; Tian, C.M.) X.G. Tian and Tibpromma S., comb. nov.
- Apiospora sorghi (J.D.P. Bezerra, C.M Gonçalves and C.M. Souza-Motta) X.G. Tian and Tibpromma S., comb. nov.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saccardo, P. Conspectus generum pyrenomycetum italicorum additis speciebus fungorum Venetorum novis vel criticis, systemate carpologico dispositorum. Atti Soc. Veneziana-Trent.-Istriana Sci. Nat. 1875, 4, 77–100. [Google Scholar]
- Dai, D.Q.; Jiang, H.B.; Tang, L.Z.; Bhat, D.J. Two new species of Arthrinium (Apiosporaceae, Xylariales) associated with bamboo from Yunnan, China. Mycosphere 2016, 7, 1332–1345. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Pintos, Á.; Alvarado, P. Phylogenetic delimitation of Apiospora and Arthrinium. Fungal Syst. Evol. 2020, 4, 197–221. [Google Scholar]
- Kunze, G. Zehn neue Pilzgattungen. Mykol. Hefte 1817, 1, 1–18. [Google Scholar]
- Hyde, K.; Fröhlich, J.; Taylor, J. Fungi from palms. XXXVI. Reflections on unitunicate ascomycetes with apiospores. Sydowia 1998, 50, 21–80. [Google Scholar]
- Hyde, K.; Norphanphoun, C.; Maharachchikumbura, S.; Bhat, D.; Jones, E.; Bundhun, D.; Chen, Y.; Bao, D.; Boonmee, S.; Calabon, M. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Ramos, H.P.; Braun, G.H.; Pupo, M.T.; Said, S. Antimicrobial activity from endophytic fungi Arthrinium state of Apiospora montagnei Sacc. and Papulaspora immersa. Braz. Arch. Biol. Technol. 2010, 53, 629–632. [Google Scholar] [CrossRef]
- Wang, M.; Tan, X.M.; Liu, F.; Cai, L. Eight new Arthrinium species from China. MycoKeys 2018, 1, 1–24. [Google Scholar] [CrossRef]
- Samuels, G.; McKenzie, E.; Buchanan, D.E. Ascomycetes of New Zealand 3. Two new species of Apiospora and their Arthrinium anamorphs on bamboo. N. Z. J. Bot. 1981, 19, 137–149. [Google Scholar] [CrossRef]
- Yin, C.; Luo, F.; Zhang, H.; Fang, X.; Zhu, T.; Li, S. First Report of Arthrinium kogelbergense Causing Blight Disease of Bambusa intermedia in Sichuan Province, China. Plant Dis. 2021, 105, 214. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, C.; Chen, X. Arthrinium phaeospermum causing dermatomycosis, a new record of China. Acta Mycol. Sin. 1990, 9, 232–235. [Google Scholar]
- Piccolo, S.L.; Mondello, V.; Giambra, S.; Conigliaro, G.; Torta, L.; Burruano, S. Arthrinium phaeospermum, Phoma cladoniicola and Ulocladium consortiale, New Olive Pathogens in Italy. J. Phytopathol. 2014, 162, 258–263. [Google Scholar] [CrossRef]
- LI, S.j.; ZHU, T.h. Binding site of toxic protein from Arthrinium phaeospermum on plasmalemma of hybrid bamboo. J. Zhejiang Univ. (Agric. Life Sci.) 2012, 38, 355–361. [Google Scholar]
- Ji, Z.L.; Zhang, S.; Zhu, F.; Wan, B.; Liang, R. First Report of Arthrinium arundinis Causing Leaf Edge Spot of Peach in China. Plant Dis. 2020, 104, 3077. [Google Scholar] [CrossRef]
- Chen, K.; Wu, X.Q.; Huang, M.X.; Han, Y.Y. First report of brown culm streak of Phyllostachys praecox caused by Arthrinium arundinis in Nanjing, China. Plant Dis. 2014, 98, 1274. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Anvar, F.G. First report of leaf spot caused by Arthrinium arundinis on rosemary in Iran. J. Plant Pathol. 2014, 96, 4–126. [Google Scholar]
- Tang, X.; Goonasekara, I.D.; Jayawardena, R.S.; Jiang, H.B.; Li, J.F.; Hyde, K.D.; Kang, J.C. Arthrinium bambusicola (Fungi, Sordariomycetes), a new species from Schizostachyum brachycladum in northern Thailand. Biodivers. Data J. 2020, 8, e58755. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R. Fungal Planet description sheets: 371–399. Pers. Mol. Phylogeny Evol. Fungi 2015, 35, 264. [Google Scholar] [CrossRef]
- Sharma, R.; Kulkarni, G.; Sonawane, M.S.; Shouche, Y.S. A new endophytic species of Arthrinium (Apiosporaceae) from Jatropha podagrica. Mycoscience 2014, 55, 118–123. [Google Scholar] [CrossRef]
- Elissawy, A.M.; Ebada, S.S.; Ashour, M.L.; Özkaya, F.C.; Ebrahim, W.; Singab, A.B.; Proksch, P. Spiroarthrinols A and B, two novel meroterpenoids isolated from the sponge-derived fungus Arthrinium sp. Phytochem. Lett. 2017, 20, 246–251. [Google Scholar] [CrossRef]
- Goodenough, A.E.; Stallwood, B.; Dandy, S.; Nicholson, T.E.; Stubbs, H.; Coker, D.G. Like mother like nest: Similarity in microbial communities of adult female Pied Flycatchers and their nests. J. Ornithol. 2017, 158, 233–244. [Google Scholar] [CrossRef]
- Wang, H.; Umeokoli, B.O.; Eze, P.; Heering, C.; Janiak, C.; Müller, W.E.; Orfali, R.S.; Hartmann, R.; Dai, H.; Lin, W. Secondary metabolites of the lichen-associated fungus Apiospora montagnei. Tetrahedron Lett. 2017, 58, 1702–1705. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous Hyphomycetes: IV. Mycol. Pap. 1963, 29, 1–33. [Google Scholar]
- Pintos, Á.; Alvarado, P.; Planas, J.; Jarling, R. Six new species of Arthrinium from Europe and notes about A. caricicola and other species found in Carex spp. hosts. MycoKeys 2019, 49, 15. [Google Scholar] [CrossRef] [Green Version]
- Index Fungorum (2021). Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 12 August 2021).
- Senanayake, I.; Rathnayaka, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H.; Hurdeal, V.; Pem, D.; Dissanayake, L.; Wijesinghe, S. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 18, p. 7. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Bhat, J.D.; Cheewangkoon, R.; Xie, N. Bambusicolous Arthrinium Species in Guangdong Province, China. Front. Microbiol. 2020, 11, 602773. [Google Scholar] [CrossRef]
- Gerin, D.; Nigro, F.; Faretra, F.; Pollastro, S. Identification of Arthrinium marii as Causal Agent of Olive Tree Dieback in Apulia (Southern Italy). Plant Dis. 2020, 104, 694–701. [Google Scholar] [CrossRef]
- Yuan, H.S.; Lu, X.; Dai, Y.C.; Hyde, K.D.; Kan, Y.H.; Kušan, I.; He, S.H.; Liu, N.G.; Sarma, V.V.; Zhao, C.L. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Das, K.; Lee, S.Y.; Choi, H.W.; Eom, A.H.; Cho, Y.J.; Jung, H.Y. Taxonomy of Arthrinium minutisporum sp. nov., Pezicula neosporulosa, and Acrocalymma pterocarpi: New Records from Soil in Korea. Mycobiology 2020, 48, 450–463. [Google Scholar] [CrossRef]
- JIANG, N.; TIAN, C.-M. The holomorph of Arthrinium setariae sp. nov.(Apiosporaceae, Xylariales) from China. Phytotaxa 2021, 483, 149–159. [Google Scholar] [CrossRef]
- Chen, T.Z.; Zhang, Y.; Ming, X.B.; Zhang, Q.; Long, H.; Hyde, K.D.; Li, Y.; Wang, Y. Morphological and phylogenetic resolution of Arthrinium from medicinal plants in Yunnan, including A. cordylines and A. pseudomarii spp. nov. Mycotaxon 2021, 136, 183–199. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series, London, UK, 2–6 September 1999; pp. 95–98. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Nylander, J.A.A. MrModeltest v2 Program Distributed by the Autho. Evol. Biol. Cent. Upps. Univ. Upps. Sweden. 2004. [Google Scholar]
- Rambaut, A. FigTree v1.4: Tree Figure Drawing Tool. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 12 August 2021).
- Singh, S.M.; Yadav, L.S.; Singh, P.N.; Hepat, R.; Sharma, R.; Singh, S.K. Arthrinium rasikravindrii sp. nov. from Svalbard, Norway. Mycotaxon 2013, 122, 449–460. [Google Scholar] [CrossRef]
- Rana, S.; Singh, P.N.; Gaikwad, S.B.; Singh, S.K. Morphology, phylogeny and ex situ conservation of Arthrinium rasikravindrae (Apiosporaceae: Xylariales): A new record from India. Kavaka 2017, 49, 1–5. [Google Scholar]
- Feng, Y.; Liu, J.K.J.; Lin, C.G.; Chen, Y.Y.; Xiang, M.M.; Liu, Z.Y. Additions to the genus Arthrinium (Apiosporaceae) from bamboos in China. Front. Microbiol. 2021, 12, 661281. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Jiang, N.; Liang, L.Y.; Yang, Q.; Tian, C.M. Arthrinium trachycarpum sp. nov. from Trachycarpus fortunei in China. Phytotaxa 2019, 400, 203–210. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa Names | Strain Numbers | Substrates | Countries | GenBank Accession Numbers | |||
|---|---|---|---|---|---|---|---|
| ITS | LSU | TUB2 | TEF 1-α | ||||
| Apiospora acutiapica | KUMCC 20-0210 | Bambusa bambos | China | MT946343 | MT946339 | MT947366 | MT947360 |
| Apiospora aquaticum | S-642 | Submerged wood | China | MK828608 | MK835806 | - | - |
| Apiospora arundinis | CBS 133509 | Aspergillus flavus sclerotium Buried in sandy field | USA | KF144886 | KF144930 | KF144976 | KF145018 |
| Apiospora arundinis | CBS 449.92 | Bamboo | Canada | KF144887 | KF144931 | KF144977 | KF145019 |
| Apiospora aurea | CBS 244.83 | - | Japan | AB220251 | KF144935 | KF144981 | KF145023 |
| Apiospora balearica | CBS 145129 | Undetermined poaceae | Spain | MK014869 | MK014836 | MK017975 | MK017946 |
| Apiospora bambusae | ICPM 6889 | Bamboo | New Zealand | MK014874 | MK014841 | MK017980 | MK017951 |
| Apiospora bambusae | CBS 145133 | Phyllostachys aurea | Spain | MK014875 | MK014842 | MK017981 | MK017952 |
| Apiospora bambusicola | MFLUCC20-0144 | Schizostachyum brachycladum | Thailand | MW173030 | MW173087 | - | MW183262 |
| Apiospora biserialis | CGMCC 3.20135 | Bamboo | China | MW481708 | MW478885 | MW522955 | MW522938 |
| Apiospora biserialis | GZCC 20_0099 | Bamboo | China | MW481709 | MW478886 | MW522956 | MW522939 |
| Apiospora biserialis | GZCC 20_0100 | Bamboo | China | MW481710 | MW478887 | MW522957 | MW522940 |
| Apiospora camelliae-sinensis | LC 5007 | Camellia sinensis | China | KY494704 | KY494780 | KY705173 | KY705103 |
| Apiospora camelliae-sinensis | LC 8181 | Brassica rapa | China | KY494761 | KY494837 | KY705229 | KY705157 |
| Apiospora chromolaenae | MFLUCC17-1505 | Chromolaena odorata | Thailand | MT214342 | MT214436 | - | MT235802 |
| Apiospora chiangraiense▲ | MFLUCC21-0053 | Dead culms of bamboo | Thailand | MZ542520 | MZ542524 | MZ546409 | - |
| Apiospora cordylinae | GUCC 10026 | Cordyline fruticosa | China | MT040105 | - | MT040147 | MT040126 |
| Apiospora cyclobalanopsidis | CGMCC 3.20136 | Cyclobalanopsidis glauca | China | MW481713 | MW478892 | MW522962 | MW522945 |
| Apiospora cyclobalanopsidis | GZCC 20_0103 | Cyclobalanopsidis glauca | China | MW481714 | MW478893 | MW522963 | MW522946 |
| Apiospora descalsii | CBS 145130 | Ampelodesmos mauritanicus | Spain | MK014870 | MK014837 | MK017976 | MK017947 |
| Apiospora dichotomanthi | LC 4950 | Dichotomanthes tristaniicarpa | China | KY494697 | KY494773 | KY705167 | KY705096 |
| Apiospora dichotomanthi | LC 8175 | Dichotomanthes tristaniicarpa | China | KY494755 | KY494831 | KY705223 | KY705151 |
| Apiospora esporlensis | CBS 145136 | Phyllostachys aurea | Spain | MK014878 | MK014845 | MK017983 | MK017954 |
| Apiospora euphorbiae | IMI 285638b | Bambusa sp. | Bangladesh | AB220241 | AB220335 | AB220288 | - |
| Apiospora gaoyouensis | CFCC 52301 | Phragmites australis | China | MH197124 | - | MH236789 | MH236793 |
| Apiospora gaoyouensis | CFCC 52302 | Phragmites australis | China | MH197125 | - | MH236790 | MH236794 |
| Apiospora garethjonesii | KUMCC 16-0202 | Dead culms of bamboo | China | KY356086 | KY356091 | - | - |
| Apiospora gelatinosa | KHAS 11962 | Bamboo | China | MW481706 | MW478888 | MW522958 | MW522941 |
| Apiospora gelatinosa | GZAAS 20-0107 | Bamboo | China | MW481707 | MW478889 | MW522959 | MW522942 |
| Apiospora guizhouensis | LC 5318 | Air in karst cave | China | KY494708 | KY494784 | KY705177 | KY705107 |
| Apiospora guizhouensis | LC 5322 | Air in karst cave | China | KY494709 | KY494785 | KY705178 | KY705108 |
| Apiospora hispanica | IMI 326877 | Beach sand | Spain | AB220242 | AB220336 | AB220289 | - |
| Apiospora hydei | CBS 114990 | Bambusa tuldoides | China | KF144890 | KF144936 | KF144982 | KF145024 |
| Apiospora hydei | KUMCC 16-0204 | Dead culms of bamboo | China | KY356087 | KY356092 | - | - |
| Apiospora hyphopodii | MFLUCC15-0003 | Bambusa tuldoides | China | KR069110 | - | - | - |
| Apiospora hyphopodii | KUMCC 16-0201 | Culms of bamboo | China | KY356088 | KY356093 | - | - |
| Apiospora iberica | CBS 145137 | Arundo donax | Portugal | MK014879 | MK014846 | MK017984 | MK017955 |
| Apiospora intestini | CBS 135835 | Gut of a grasshopper | India | KR011352 | MH877577 | KR011350 | KR011351 |
| Apiospora intestini▲ | MFLUCC 21-0052 | Dead culms of bamboo | Thailand | MZ542521 | MZ542525 | MZ546410 | MZ546406 |
| Apiospora italica | CBS 145138 | Arundo donax | Italy | MK014880 | MK014847 | MK017985 | MK017956 |
| Apiospora italica | CBS 145139 | Phragmites australis | Spain | MK014881 | MK014848 | MK017986 | - |
| Apiospora jatrophae | AMH-9557 | Jatropha podagrica | India | JQ246355 | - | - | - |
| Apiospora jatrophae | AMH-9556 | Jatropha podagrica | India | HE981191 | - | - | - |
| Apiospora jiangxiensis | LC 4494 | Phyllostachys sp. | China | KY494690 | KY494766 | KY705160 | KY705089 |
| Apiospora jiangxiensis | LC 4577 | Maesa sp. | China | KY494693 | KY494769 | KY705163 | KY705092 |
| Apiospora kogelbergensis | CBS 113332 | Cannomois virgata | South Africa | KF144891 | KF144937 | KF144983 | KF145025 |
| Apiospora kogelbergensis | CBS 113333 | Dead culms of Restionaceae | South Africa | KF144892 | KF144938 | KF144984 | KF145026 |
| Apiospora locuta-pollinis | LC 11688 | Bee bread | China | MF939596 | - | MF939623 | MF939618 |
| Apiospora locuta-pollinis | LC 11683 | Brassica campestris | China | MF939595 | - | MF939622 | MF939616 |
| Apiospora longistroma | MFLUCC 11-0479 | Dead culms of bamboo | Thailand | KU940142 | KU863130 | - | - |
| Apiospora longistroma | MFLUCC11-0481 | Dead culms of bamboo | Thailand | KU940141 | KU863129 | - | - |
| Apiospora malaysiana | CBS 102053 | Macaranga hullettii | Malaysia | KF144896 | KF144942 | KF144988 | KF145030 |
| Apiospora marii | CBS 497.90 | Beach sands | Spain | AB220252 | KF144947 | KF144993 | KF145035 |
| Apiospora marii | DiSSPA_A1 | Oleaeuropaea | Italy | MK602320 | - | MK614695 | MK645472 |
| Apiospora mediterranea | IMI 326875 | Air | Spain | AB220243 | AB220337 | AB220290 | - |
| Apiospora minutispora | 1.70E-41 | Mountain soil | Korea | LC517882 | - | LC518888 | LC518889 |
| Apiospora mytilomorpha | DAOM 214595 | Andropogon sp. | India | KY494685 | - | - | - |
| Apiospora neobambusae | LC 7106 | Leaves of bamboo | China | KY494718 | KY494794 | KY705186 | KY806204 |
| Apiospora neobambusae | LC 7124 | Leaves of bamboo | China | KY494727 | KY494803 | KY705195 | KY806206 |
| Apiospora neochinensis | CFCC 53036 | Fargesia qinlingensis | China | MK819291 | - | MK818547 | MK818545 |
| Apiospora neochinensis | CFCC 53037 | Fargesia qinlingensis | China | MK819292 | - | MK818548 | MK818546 |
| Apiospora neogarethjonesii | DQD 2019a | Bamboo | China | MK070897 | MK070898 | - | - |
| Apiospora neosubglobosa | JHB 006 | Bamboo | China | KY356089 | KY356094 | - | - |
| Apiospora neosubglobosa | KUMCC 16-0203 | Bamboo | China | KY356090 | KY356095 | - | - |
| Apiospora obovata | LC 4940 | Lithocarpus sp. | China | KY494696 | KY494772 | KY705166 | KY705095 |
| Apiospora obovata | LC 8177 | Lithocarpus sp. | China | KY494757 | KY494833 | KY705225 | KY705153 |
| Apiospora ovata | CBS 115042 | Arundinaria hindsii | China | KF144903 | KF144950 | KF144995 | KF145037 |
| Apiospora paraphaeosperma | MFLUCC13-0644 | Dead culms of bamboo | Thailand | KX822128 | KX822124 | - | - |
| Apiospora phragmitis | CPC 18900 | Phragmites australis | Italy | KF144909 | KF144956 | KF145001 | KF145043 |
| Apiospora phyllostachydis | MFLUCC18-1101 | Phyllostachys heteroclada | China | MK351842 | MH368077 | MK291949 | MK340918 |
| Apiospora piptatheri | CBS 145149 | Piptatherum miliaceum | Spain | MK014893 | MK014860 | - | MK017969 |
| Apiospora pseudomarii | GUCC 10228 | Aristolochia debilis | China | MT040124 | - | MT040166 | MT040145 |
| Apiospora pseudoparenchymatica | LC 7234 | Leaves of bamboo | China | KY494743 | KY494819 | KY705211 | KY705139 |
| Apiospora pseudoparenchymatica | LC 8173 | Leaves of bamboo | China | KY494753 | KY494829 | KY705221 | KY705149 |
| Apiospora pseudorasikravindrae | KUMCC 20-0208 | Bambusa dolichoclada | China | MT946344 | - | MT947367 | MT947361 |
| Apiospora pseudosinensis | CPC 21546 | Leaves of bamboo | Netherlands | KF144910 | KF144957 | - | KF145044 |
| Apiospora pseudospegazzinii | CBS 102052 | Macaranga hullettii | Malaysia | KF144911 | KF144958 | KF145002 | KF145045 |
| Apiospora pterosperma | CBS 123185 | Machaerina sinclairii | New Zealand | KF144912 | KF144959 | KF145003 | - |
| Apiospora pterosperma | CPC 20193 | Lepidosperma gladiatum | Australia | KF144913 | KF144960 | KF145004 | KF145046 |
| Apiospora qinlingensis | CFCC 52303 | Fargesiaqinlingensis | China | MH197120 | - | MH236791 | MH236795 |
| Apiospora qinlingensis | CFCC 52304 | Fargesia qinlingensis | China | MH197121 | - | MH236792 | MH236796 |
| Apiospora rasikravindrae | LC 8179 | Brassica rapa | China | KY494759 | KY494835 | KY705227 | KY705155 |
| Apiospora rasikravindrae | NFCCI 2144 | Soil | Norway | JF326454 | - | - | - |
| Apiospora rasikravindrae▲ | MFLUCC 21-0051 | Dead culms of bamboo | Thailand | MZ542523 | MZ542527 | MZ546412 | MZ546408 |
| Apiospora rasikravindrae▲ | MFLUCC 21-0054 | Dead culms of Maize | Thailand | MZ542522 | MZ542526 | MZ546411 | MZ546407 |
| Apiospora sacchari | CBS 372.67 | Air | - | KF144918 | KF144964 | KF145007 | KF145049 |
| Apiospora sacchari | CBS 664.74 | Soil under Calluna vulgaris | Netherlands | KF144919 | KF144965 | KF145008 | KF145050 |
| Apiospora saccharicola | CBS 191.73 | Air | Netherlands | KF144920 | KF144966 | KF145009 | KF145051 |
| Apiospora saccharicola | CBS 831.71 | - | Netherlands | KF144922 | KF144969 | KF145012 | KF145054 |
| Apiospora septata | CGMCC 3.20134 | bamboo | China | MW481711 | MW478890 | MW522960 | MW522943 |
| Apiospora septata | GZCC 20_0109 | bamboo | China | MW481712 | MW478891 | MW522961 | MW522944 |
| Apiospora serenensis | IMI 326869 | Food, pharmaceutical excipients, atmosphere and home dust | Spain | AB220250 | AB220344 | AB220297 | - |
| Apiospora setariae | MT492005 | Setaria viridis | China | MT492005 | - | MT497467 | MW118457 |
| Apiospora setostroma | KUMCC 19-0217 | Dead branches of bamboo | China | MN528012 | MN528011 | - | MN527357 |
| Apiospora sorghi | URM 93000 | Sorghum bicolor | Brazil | MK371706 | - | MK348526 | - |
| Apiospora subglobosa | MFLUCC11-0397 | Dead culms of bamboo | Thailand | KR069112 | KR069113 | - | - |
| Apiospora subrosea | LC 7291 | Leaves of bamboo | China | KY494751 | KY494827 | KY705219 | KY705147 |
| Apiospora subrosea | LC 7292 | Leaves of bamboo | China | KY494752 | KY494828 | KY705220 | KY705148 |
| Apiospora thailandica | MFLUCC 15-0199 | Dead culms of bamboo | Thailand | KU940146 | KU863134 | - | - |
| Apiospora thailandica | MFLUCC15-0202 | Dead culms of bamboo | Thailand | KU940145 | KU863133 | - | - |
| Apiospora vietnamensis | IMI 99670 | Citrus sinensis | Vietnam | KX986096 | KX986111 | KY019466 | - |
| Apiospora xenocordella | CBS 478.86 | Soil from roadway | Zimbabwe | KF144925 | KF144970 | KF145013 | KF145055 |
| Apiospora xenocordella | CBS 595.66 | Soil | Austria | KF144926 | KF144971 | - | - |
| Apiospora yunnana | DDQ 00281 | Phyllostachys nigra | China | KU940148 | KU863136 | - | - |
| Apiospora yunnana | MFLUCC15-1002 | Phyllostachys nigra | China | KU940147 | KU863135 | - | - |
| Arthrinium austriacum | GZU 345004 | Carex pendula | Austria | MW208928 | - | - | - |
| Arthrinium austriacum | GZU 345006 | Carex pendula | Austria | MW208929 | MW208860 | - | - |
| Arthrinium cf. sporophleoides | GZU 345102 | Carex firma | Austria | MW208944 | MW208866 | - | - |
| Arthrinium caricicola | CBS 145127 | Carex ericetorum | China | MK014871 | MK014838 | MK017977 | MK017948 |
| Arthrinium crenatum | AG 19066 | Poaceae, Carex | France | MW208931 | MW208861 | - | - |
| Arthrinium curvatum | AP 25418 | Leaves of Carex sp. | China | MK014872 | MK014839 | MK017978 | MK017949 |
| Arthrinium japonicum | IFO 30500 | - | Japan | AB220262 | AB220356 | AB220309 | - |
| Arthrinium japonicum | IFO 31098 | Leaves of Carex despalata | Japan | AB220264 | AB220358 | AB220311 | - |
| Arthrinium luzulae | AP7619-3 | Luzula sylvatica | Spain | MW208937 | MW208863 | - | - |
| Arthrinium morthieri | GZU 345043 | Cyperaceae carex | Austria | MW208938 | MW208864 | - | - |
| Arthrinium phaeospermum | CBS 114317 | Leaves of Hordeum vulgare | Iran | KF144906 | KF144953 | KF144998 | KF145040 |
| Arthrinium phaeospermum | CBS 114318 | Leaves of Hordeum vulgare | Iran | KF144907 | KF144954 | KF144999 | KF145041 |
| Arthrinium puccinioides | CBS 549.86 | Lepidosperma gladiatum | Germany | AB220253 | AB220347 | AB220300 | - |
| Arthrinium sphaerospermum | AP25619/CBS 146355 | Poaceae | Norway | MW208943 | MW208865 | - | - |
| Arthrinium sporophleum | CBS 145154 | Dead leaves of Juncus sp. | Spain | MK014898 | MK014865 | MK018001 | MK017973 |
| Arthrinium trachycarpum | CFCC 53038 | Trachycarpus fortune | China | MK301098 | - | MK303394 | MK303396 |
| Arthrinium urticae | IMI 326344 | - | - | AB220245 | AB220339 | AB220292 | - |
| Arthrinium trachycarpum | CFCC 53039 | Trachycarpus fortune | China | MK301099 | - | MK303395 | MK303397 |
| Nigrospora aurantiaca | CGMCC 3.18130 | Nelumbo sp. | China | KX986064 | KX986098 | KY019465 | KY019295 |
| Nigrospora camelliae-sinensis | CGMCC 3.18125 | Camellia sinensis | China | KX985986 | KX986103 | KY019460 | KY019293 |
| Nigrospora chinensis | CGMCC 3.18127 | Machilus breviflora | China | KX986023 | KX986107 | KY019462 | KY019422 |
| Nigrospora gorlenkoana | CBS 480.73 | Vitis vinifera | Kazakhstan | KX986048 | KX986109 | KY019456 | KY019420 |
| Nigrospora guilinensis | CGMCC 3.18124 | Camellia sinensis | China | KX985983 | KX986113 | KY019459 | KY019292 |
| Nigrospora hainanensis | CGMCC 3.18129 | Musa paradisiaca | China | KX986091 | KX986112 | KY019464 | KY019415 |
| Nigrospora lacticolonia | CGMCC 3.18123 | Camellia sinensis | China | KX985978 | KX986105 | KY019458 | KY019291 |
| Nigrospora musae | CBS 319.34 | Musa sp. | Australia | MH855545 | KX986110 | KY019455 | KY019419 |
| Nigrospora oryzae | LC2693 | Neolitsea sp. | China | KX985944 | KX986101 | KY019471 | KY019299 |
| Nigrospora osmanthi | CGMCC 3.18126 | Hedera nepalensis | China | KX986010 | KX986106 | KY019461 | KY019421 |
| Nigrospora pyriformis | CGMCC 3.18122 | Citrus sinensis | China | KX985940 | KX986100 | KY019457 | KY019290 |
| Nigrospora rubi | LC2698 | Rubus sp. | China | KX985948 | KX986102 | KY019475 | KY019302 |
| Nigrospora sphaerica | LC7298 | Nelumbo sp. | China | KX985937 | KX986097 | KY019606 | KY019401 |
| Nigrospora vesicularis | CGMCC 3.18128 | Musa paradisiaca | China | KX986088 | KX986099 | KY019463 | KY019294 |
| Sporocadus trimorphus | CBS 114203 | Rosa canina | Sweden | MH553977 | MH554196 | MH554636 | MH554395 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, X.; Karunarathna, S.C.; Mapook, A.; Promputtha, I.; Xu, J.; Bao, D.; Tibpromma, S. One New Species and Two New Host Records of Apiospora from Bamboo and Maize in Northern Thailand with Thirteen New Combinations. Life 2021, 11, 1071. https://doi.org/10.3390/life11101071
Tian X, Karunarathna SC, Mapook A, Promputtha I, Xu J, Bao D, Tibpromma S. One New Species and Two New Host Records of Apiospora from Bamboo and Maize in Northern Thailand with Thirteen New Combinations. Life. 2021; 11(10):1071. https://doi.org/10.3390/life11101071
Chicago/Turabian StyleTian, Xingguo, Samantha C. Karunarathna, Ausana Mapook, Itthayakorn Promputtha, Jianchu Xu, Danfeng Bao, and Saowaluck Tibpromma. 2021. "One New Species and Two New Host Records of Apiospora from Bamboo and Maize in Northern Thailand with Thirteen New Combinations" Life 11, no. 10: 1071. https://doi.org/10.3390/life11101071