
Genetic Investigation of Inverse Psoriasis


1
MTA-SZTE Dermatological Research Group, Eötvös Loránd Research Network, 6720 Szeged, Hungary
2
Department of Dermatology and Allergology, University of Szeged, 6720 Szeged, Hungary
3
Department of Medical Genetics, University of Szeged, 6720 Szeged, Hungary
4
PentaCore Laboratory, 1094 Budapest, Hungary
5
HCEMM-USZ Skin Research Group, University of Szeged, 6720 Szeged, Hungary
*
Author to whom correspondence should be addressed.
†
These two authors contributed equally to this work.
Life 2021, 11(7), 654; https://doi.org/10.3390/life11070654
Submission received: 27 May 2021
/
Revised: 27 June 2021
/
Accepted: 1 July 2021
/
Published: 5 July 2021
(This article belongs to the Special Issue Psoriasis: New Developments and Concepts in Pathogenesis and Treatment)
Abstract
:Inverse psoriasis is considered to be a rare variant of plaque-type psoriasis and is associated with significantly impaired quality of life. Clinical manifestations and treatment options are somewhat different for each subtype. Identifying genetic variants that contribute to the susceptibility of different types of psoriasis might improve understanding of the etiology of the disease. Since we have no current knowledge about the genetic background of inverse psoriasis, whole exome sequencing was used to comprehensively assess genetic variations in five patients with exclusively inverse lesions. We detected six potentially pathogenic rare (MAF < 0.01) sequence variants that occurred in all investigated patients. The corresponding mutated genes were FN1, FBLN1, MYH7B, MST1R, RHOD, and SCN10A. Several mutations identified in this study are known to cause disease, but roles in psoriasis or other papulosquamous diseases have not previously been reported. Interestingly, potentially causative variants of established psoriasis-susceptibility genes were not identified. These outcomes are in agreement with our hypothesis that the inverse subtype is a different entity from plaque-type psoriasis.
1. Introduction
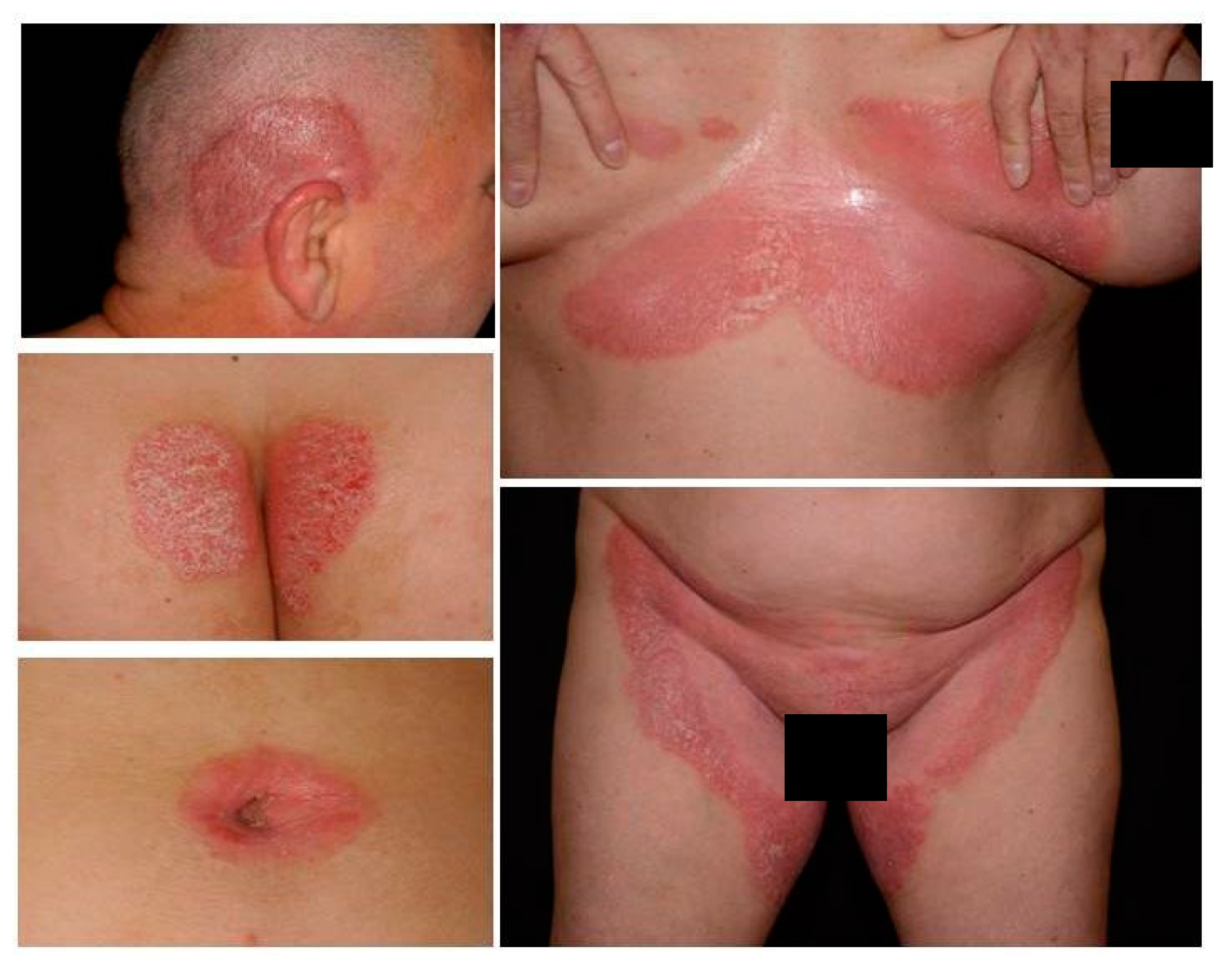
Psoriasis is a chronic inflammatory skin disorder with a significant occurrence in patients with dermatological diseases. The most common form (90% of all cases) is the plaque-type condition. Inverse psoriasis (flexural, intertriginous), an uncommon variant, is triggered by physical stimuli, infections and obesity. Despite the limited percentage of involved body areas, this variant has a significant adverse impact on quality of life especially regarding sexual behavior [1]. Due to breakthrough development in the therapeutic arsenal, complete or almost complete resolution is usually achieved for most patients with plaque-type psoriasis. Nevertheless, patients with non-plaque phenotypes often do not meet the criteria for these novel agents. As a result, this non-plaque patient population is not adequately assessed in observational and interventional studies and is, therefore, often undertreated [2]. Although up to 63% of psoriasis patients have genital involvement at some point during the course of their disease [3], psoriasis affecting exclusively the genital or flexural regions is very rare. Inverse psoriasis usually affects the skin folds, including the inguinal and axillar fold, genital area, intergluteal cleft, inframammary creases, periumbilical, retroauricular, antecubital and popliteal fossae (Figure 1). The most commonly affected area is the inguinal fold (95.8%), followed by the axillar region and genital area [4]. Morphologically, the lesions often present as well-defined, thin, erythematous plaques without scaling, which can affect both the genital skin and mucosa, and can be accompanied by fissures, ulcers, and/or erosions [5]. Fransson et al. found that patients with palmar psoriasis have increased risk for inverse psoriasis and that the presence of inverse subtypes seems to be linked with periodontitis as well [6]. In challenging cases, dermoscopy or histopathology may be necessary for diagnosis. In vivo confocal reflectance microscopy might also be helpful, although this technology is currently available only in a small number of academic centers due to high equipment costs [7].
The genetic background of psoriasis is complex: more than sixty susceptibility loci have been identified so far, but only a sPSORS (psoriasis susceptibility) loci have been shown to be associated with the disease. Genome-wide association studies (GWAS) and/or genome-wide pathway analysis (GWPA) have shown that more than 200 genes are associated with the disease or the susceptibility to the disease [8,9,10]. Several of these genes implicated in the mechanisms of psoriasis pathology are associated with innate- and adaptive immunity, barrier function, and inflammation.
Although mild cases of inverse psoriasis can be controlled well with topical anti-inflammatory agents, the treatment of more severe symptoms is challenging. The clinical manifestation, location and responses to anti-psoriatic treatments are somewhat different for inverse psoriasis and psoriasis vulgaris, indicating possible genetic diversity for the different subtypes. Nonetheless, a unique genetic profile has not yet been established for inverse psoriasis.
We hypothesized that inverse and non-plaque-type psoriasis are different entities than plaque-type psoriasis rather than rare subtypes of the chronic plaque-type condition. To confirm our hypothesis, we identified gene polymorphisms using whole exome sequencing (WES) analysis.
2. Materials and Methods
2.1. Patients
Five patients (4 female and 1 male) presenting psoriasis exclusively in inverse areas were recruited from our database. The average age of the patients was 51.4 years. One patient had early-onset psoriasis (onset at 17 years), whereas four patients had late-onset disease (onset at 29–53 years). The affected areas included flexural and genital regions in all of the cases. Four of the patients presented retroauricular involvement, and two patients presented periumbilical involvement, as well.
2.2. Blood Samples
Peripheral blood samples were collected from patients diagnosed for inverse psoriasis (n = 5) for WES. Variants originating from WES data of other studies were used as a control in-house database. This in-house database included variants of 107 individuals (mean age: 51; without any skin diseases) of Hungarian origin. The investigation was approved by the Ethics Committee of Ministry of Human Capacities, Hungary. Written informed consent was obtained from patients and healthy individuals, and the study was conducted according to the Principles of the Declaration of Helsinki.
2.3. DNA Isolation
Genomic DNA was isolated from whole venous blood containing EDTA using the QIAamp DNA Blood Mini Kit (QIAGEN, Hilden, Germany).
2.4. Library Preparation and Exome Sequencing
An exome amplicon library was prepared using the Ion AmpliSeq Exome RDY kit (Thermo Fisher, Waltham, MA, USA). Briefly, 100 ng of genomic DNA was added to dehydrated, ultra-high multiplexed primer pairs (12 pools) in a 96-well plate and amplified with the following PCR conditions: 99 °C for 2 min; 99 °C for 15 s and 60 °C for 16 min (10 cycles); and holding at 10 °C. Primers were partially digested using a FuPa reagent, and then sequencing motifs and barcodes were ligated to the amplicons. The library was purified using the Agencourt AMPure XP Reagent (Beckmann Coulter, Brea, CA, USA). The concentration of the final library was determined using the Ion Library TaqMan Quantitation Kit (Thermo Fisher, Waltham, MA, USA) on an ABI 7500 qPCR instrument with the absolute quantification method.
Template preparation was performed with the Ion 540 OT2 Kit (Thermo Fisher, Waltham, MA, USA) on semi-automated Ion OneTouch 2 instrument using the emPCR method. After breaking the emulsion, non-templated beads were removed from the solution during the enrichment process on the Ion OneTouch ES (Thermo Fisher, Waltham, MA, USA) machine. Subsequently, the sequencing primer and polymerase were added, the fully prepared Ion sphere particles were loaded into an Ion 540 chip, and sequencing runs were performed using the Ion S5 Sequencing kit (Thermo Fisher, Waltham, MA, USA) with 500 flows.
2.5. Data Analysis
Sequence data from the Ion torrent run were analyzed using the platform-specific pipeline software Torrent Suite, v 5.10 (Thermo Fisher, Waltham, MA, USA), for calling bases, trimming adapter and primer sequences, filtering out poor quality reads, and de-multiplexing the reads according to the barcode sequences. Briefly, the TMAP algorithm was used to align the reads to the hg19 human reference genome, and then the variant caller plug-in was executed to search for germline variants in the targeted regions. The Integrative Genomics Viewer (https://software.broadinstitute.org/software/igv/ accessed on 14 May 2020) was used for visualization of the mapped reads. Variants were annotated using the Ion Reporter software (Thermo Fisher, Waltham, MA, USA).
3. Results
Genomic DNA was isolated from peripheral blood samples using a standard technique. WES was performed with samples from five patients who had been definitively diagnosed with inverse psoriasis and were free of common plaque-type psoriasis symptoms. The WES results allowed us to identify 34,800–38,000 sequence variants per person. Many of these variants were common, neutral or synonymous; therefore, we adjusted the filtering criteria as follows: minor allele frequency of less than 0.01; coverage of more than 50; variant functions of missense, nonsense, frame shift or stop loss; and variant present in all five examined inverse psoriasis patients. A total of 186 variants met the filtering criteria and were evaluated by multiple single-nucleotide polymorphism (SNP) databases (Varsome, https://varsome.com accessed on 15 March 2021; Ensembl, https://www.ensembl.org/index.html accessed on 15 March 2021; dbSNP, https://www.ncbi.nlm.nih.gov/snp/ accessed on 15 March 2021) and pathogenicity prediction tools (SIFT, LRT, DANN, PolyPhen, Mutation Taster, FATHMM-MKL, Meta-SVM) to determine whether any could be considered pathogenic. Of 32 mutations (Supplementary Table S1) that were considered to be damaging or probably damaging, six missense variants are located in a functional domain of the corresponding protein (Table 1) and, thus, likely to have consequences at the protein level.
The six possibly causative variants occurred in the following genes: fibronectin (FN1), fibulin (FBLN1), myosin heavy chain 7B (MYH7B), macrophage stimulating 1 receptor (MST1R), ras homolog family member D (RHOD), and sodium voltage-gated channel alpha subunit 10 (SCN10A). None of these genes has been associated with psoriasis in previous large-scale genetic studies (GWAS, GWPA, etc.). All six variants were validated by Sanger sequencing. None of the identified variants were detected in control individuals (n = 107, from in-house database).
The rs1250209 missense mutation is located in the fibronectin Type I domain of the fibronectin protein. Fibronectin is a plasma protein that binds cell surfaces and various compounds including collagen, fibrin, heparin, DNA, and actin, and this domain contributes to fibrin binding. Fibronectin is involved in cell adhesion and migration processes, including embryogenesis, wound healing, blood coagulation, host defense, and metastasis. The gene has three regions subject to alternative splicing, resulting in the potential to produce 20 transcript variants. Fibronectin is believed to play a crucial role in the pathogenesis of psoriasis by influencing inflammation and keratinocyte hyperproliferation [11,12,13]. Mutations in fibronectin have been associated with spondylometaphyseal dysplasia [14] and glomerulopathy [15].
Variant rs136730 is located in the anaphylatoxin homologous domain of Fibulin 1. This anaphylatoxin-like domain was identified in the complement-derived anaphylatoxin C3a protein, which mediates smooth muscle contraction, histamine release from mast cells, enhanced vascular permeability, chemotaxis, inflammation, and generation of cytotoxic oxygen radicals [16]. Fibulin 1 localizes to basement membranes, elastic fibers, and other connective tissue structures [17] and is able to bind fibronectin, proteoglycans, tropoelastin, and various elastic fiber and basement membrane proteins [18]. The protein participates in cell adhesion and migration [19] and also contributes to the supramolecular organization of ECM architecture, affecting in particular basement membranes. Fibulin 1 is also a plasma protein capable of binding to fibrinogen [20] and may play a role in hemostasis and thrombosis; moreover, it is implicated in tumor formation and invasion processes as a tumor suppressor [21].
The rs2425015 missense mutation is located in the coiled-coil domain of the MYH7B protein. The MYH7B gene encodes the heavy chain of Myosin II, which catalyzes ATP hydrolysis, interacts with actin and is involved in muscle contraction. Several mutations in MYH7 have been associated with inherited cardiomyopathies and muscle atrophy [22,23].
The rs4930409 missense mutation is located on the RHO domain of the RHOD protein. RHOD is involved in endosome dynamics and membrane transport and participates in the reorganization of actin cytoskeleton [24,25]. No pathological mutation of RHOD has been reported in any human disease so far.
Sequence variant rs6599241 is located in the transmembrane domain of the SCN10A protein. The SCN10A gene encodes the alpha subunit of a voltage-gated sodium channel which is an integral membrane glycoprotein responsible for the initial rising phase of action in most excitable cells. Sodium channels have been found to accumulate in regions of peripheral nerve injury and may be important in chronic pain [26]. Gain-of-function mutations in SCN10A can cause an episodic pain disorder [27]. In the imiquimod-induced, IL (interleukin)-23-dependent, psoriasis-like skin of mice, SCN10A+ nociceptors interact with DDCs, regulate the IL-23/IL-17 pathway, and control cutaneous immune responses [28].
The rs7433231 missense mutation is located on the tyrosine kinase catalytic domain of the MST1R protein. MST1R is a receptor tyrosine kinase that transduces signals from the extracellular matrix into the cytoplasm by binding to MST1 ligand. This binding regulates many physiological processes, including cell survival, migration and differentiation [29,30]. Dai and coworkers found a significant association between variation in the MST1R gene and development of nasopharyngeal carcinoma [31].
We also filtered the WES results for rare variants of psoriasis susceptibility genes (OMIM database, Singh et al., 2019; Tsoi et al., 2017; Supplementary Table S2). The filtering criteria included a coverage of more than 50, minor allele frequency of less than 0.01, and the variant function. All of the 21 rare variants we identified in individual patients are missense mutations and all were predicted to be benign with SNP pathogenicity prediction tools (Supplementary Table S3). Additionally, we filtered our results for Th17 pathway genes (12 genes, Supplementary Table S4) and identified 58 rare and common SNPs. Although several of these variants have a wide range of variant effect, only two of the sequence variants, each identified in only one patient, were predicted to be possibly damaging (IL-12B, rs3213119, p.Val298Phe and IL-17F, rs2397084, p.Glu126Gly).
4. Discussion
We performed WES analysis with samples taken from five adult patients of inverse psoriasis. Careful clinical and histopathological data confirmed the definitive diagnosis inverse psoriasis, and the patients were without common plaque-type psoriasis symptoms. Six of the identified missense mutations are localized in a functional domain of the corresponding protein and were predicted to be pathogenic according to data from SNP databases and computational pathogenicity prediction tools. Notably, none of the identified sequence variants is related to psoriasis-susceptibility genes or genes in pathways associated to psoriasis pathogenesis. Only non-pathogenic variants were found for psoriasis susceptibility genes from the investigated individuals, which agrees well with the absence of classical psoriatic symptoms.
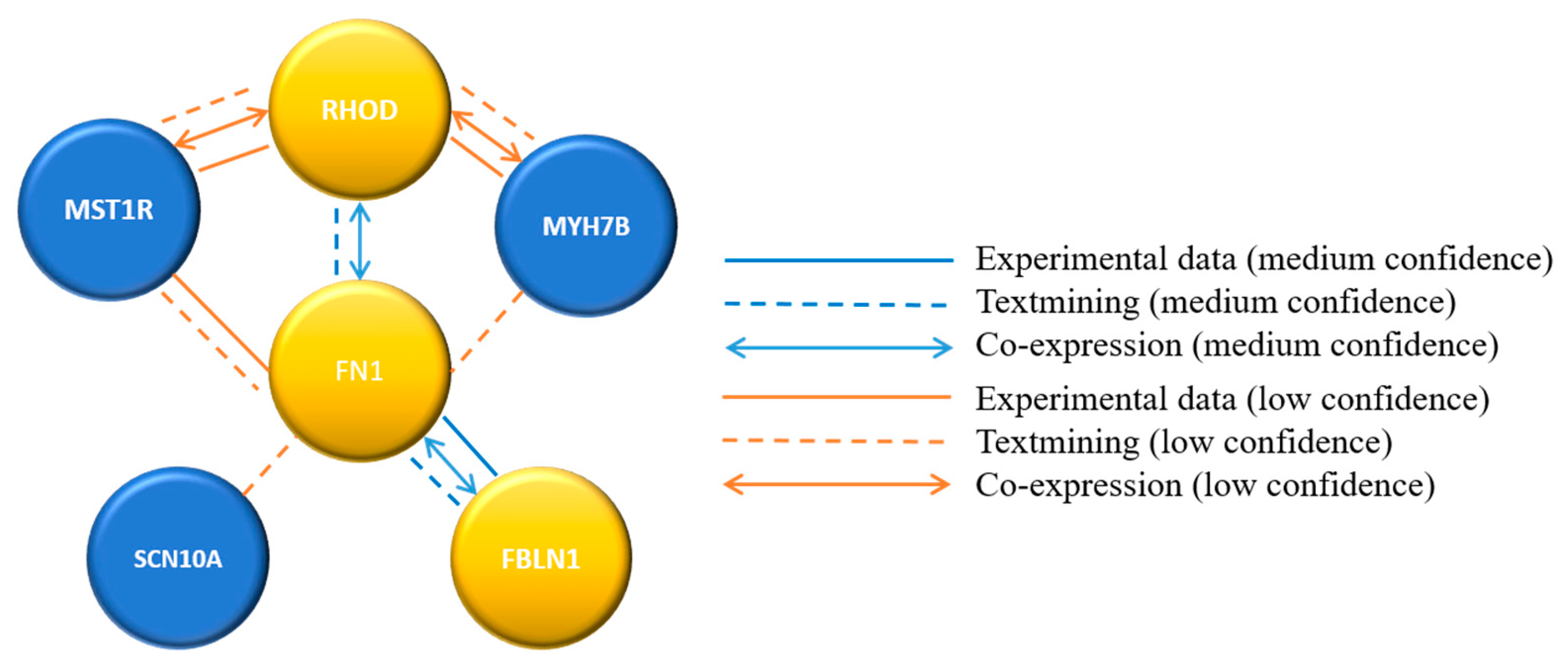
Sequence variants in six annotated genes presumed to be pathogenic were identified in all five inverse psoriatic patients. These variants were further analyzed for functional protein association networks by the STRING database (STRING, https://string-db.org/ accessed on 23 March 2021). Based on the STRING results, a confidence-based protein-interaction network was calculated for the six identified genes. The analysis predicted medium-confidence association between FBLN1, FN1, and RHOD genes and low-confidence interactions between the other genes (Figure 2).
The miRNAs regulating the six genes were predicted using the GeneCodis tool [32,33,34]. Using a gene set from the inverse psoriasis patients, the co-occurrence annotations found by GeneCodis3 identified the following three miRNAs. miR892a modulates the FBLN1, MYH7B and MST1R genes and is known to promote proliferation and invasion of hepatocellular carcinoma cells via targeting CD226 [35]. miR892b modulates the FBLN1, MYH7B and MST1R genes and is known to influence proliferation, migration, and invasion of bladder cancer cells. In addition, silencing miR892b activates NF-κB in breast cancer [36,37]. miR647 modulates FBLN1, FN1, and MST1R genes and has a tumor-promoting role in gastric cancer via repression of TP73 [38]. This miRNA also promotes cancer progression by downregulating nuclear factor IX in colorectal cancer [39] and has a role in the suppression of human gastric cancer [40].
In conclusion, we detected six potentially pathogenic and rare sequence variants that occurred in all five investigated patients. Notably, none of the potentially causative variants known for psoriasis susceptibility genes were identified, suggesting that the psoriasis of patients with exclusively inverse psoriasis that have lesions only on the skin folds is a different entity from plaque-type psoriasis. It would be important to compare the frequency of the identified variants in inverse and plaque type psoriasis, but unfortunately we did not find any data about these variants in plaque type psoriasis. The major limitation of this study in the low number of examined patients. Although the sequence variants in the six annotated genes were found in all five patients, these findings can not be considered statistically significant due to the small number of enrolled patients. Interestingly our five patients were extremely resistant to antipsoriatic therapies. Topical anti-inflammatory treatments, such as low potent to potent corticosteroids, calcineurin-inhibitors, Vitamin D analogues, did not result in significant improvement of the lesions. Of the conventional therapies, the best results were seen with Cyclosporin A, which was used in three cases; this treatment had to be stopped after two years of use. Three of the patients also received acitretin with no improvement. Methotrexate has been used for all patients with moderate results, and one patient remains on this therapy. After the failures with topical and conventional treatments, biological therapies have been initiated. Biological therapies are antibodies directed to block cytokines responsible for the inflammation associated with psoriasis, such as IL-17 and 23 [41,42]. Treatment using TNF-alfa, IL-12/23, and IL-17 inhibitors had limited efficacy. Two of the five patients are currently receiving IL-23 inhibitors with convincing clearing of the lesions. The results of our genetic investigation and the pattern of treatment resistance of patients suggest that inverse psoriasis has a different pathogenesis than plaque-type psoriasis.
In human genetics, translating genotype data for clinical use is an important step [43]. The biological and clinical interpretation of genetic studies resulted in some therapies already being tested in clinical trials. Understanding how associated variants modulate disease risk and severity, and how they impact cellular phenotypes resulting in more effective drug discovery [44]. Therefore, further large-scale genetic and functional studies are needed to better understand the pathogenesis of this rare form of psoriasis.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/life11070654/s1, Supplementary Table S1: damaging or probably damaging sequence variants found in five inverse psoriatic patients; Supplementary Table S2: psoriasis-associated genes based on GWAS and GWPA studies and the OMIM database; Supplementary Table S3: rare variants (MAF < 0.01) located in psoriasis-susceptibility genes from individual inverse psoriasis patients; Supplementary Table S4: rare variants (MAF < 0.01) located in genes involved in the Th17 pathway from individual inverse psoriasis patients.
Author Contributions
Conceptualization, A.G. and E.V.; methodology, A.G. and K.Á.; software, A.G.; validation, A.G. and K.Á.; formal analysis, A.G.; investigation, A.G.; resources, L.K.; data curation, A.G. and K.F.; writing—original draft preparation, A.G. and E.V.; writing—review and editing, A.G. and K.F.; visualization, A.G.; supervision, L.K.; project administration, A.G.; funding acquisition, L.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the GINOP-2.3.2-15-2016-00015 grant and has received funding from the EU’s Horizon 2020 Research and Innovation Program under Grant Agreement No. 739593.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and was approved by the Ethics Committee of Ministry of Human Capacities, Hungary (Protocol Code PSO-INVRS-001, date of approval: 6 November 2017).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patients to publish this paper.
Data Availability Statement
Data is contained within the article or Supplementary Materials.
Acknowledgments
We thank Kornélia Tripolszki and Judit Danis for their helpful advice. We thank the patients and control individuals for participating and providing samples for our research.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, the collection, analyses, or interpretation of data, the writing of the manuscript, or the decision to publish the results.
References
- Dattola, A.; Silvestri, M.; Bennardo, L.; Del Duca, E.; Longo, C.; Bianchi, L.; Nisticò, S. Update of calcineurin inhibitors to treat inverse psoriasis: A systematic review. Dermatol. Ther. 2018, 31, e12728. [Google Scholar] [CrossRef]
- Kaskas, N.; Merola, J.F.; Qureshi, A.A.; Paek, S.Y. Polyphenotypic Psoriasis: A Report from the GRAPPA 2016 Annual Meeting. J. Rheumatol. 2017, 44, 695–696. [Google Scholar] [CrossRef]
- Meeuwis, K.A.P.; Bleakman, A.P.; Van De Kerkhof, P.C.M.; Dutronc, Y.; Henneges, C.; Kornberg, L.J.; Menter, A. Prevalence of genital psoriasis in patients with psoriasis. J. Dermatol. Treat. 2018, 29, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, C.; Gao, T.; Liu, Y. Clinical analysis of 48 cases of inverse psoriasis: A hospital-based study. Eur. J. Dermatol. 2005, 15, 176–178. [Google Scholar] [PubMed]
- Meeuwis, K.; Hullu, J.; Massuger, L.; Kerkhof, P.; Rossum, M. Genital Psoriasis: A Systematic Literature Review on this Hidden Skin Disease. Acta Derm. Venereol. 2011, 91, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Fransson, J.; Storgårds, K.; Hammar, H. Palmoplantar lesions in psoriatic patients and their relation to inverse psoriasis, tinea infection and contact allergy. Acta Derm. Venereol. 1985, 65, 218–223. [Google Scholar] [PubMed]
- Micali, G.; Verzì, A.E.; Giuffrida, G.; Panebianco, E.; Musumeci, M.L.; Lacarrubba, F. Inverse Psoriasis: From Diagnosis to Current Treatment Options. Clin. Cosmet. Investig. Dermatol. 2020, 12, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Stuart, P.E.; Tian, C.; Gudjonsson, J.E.; Das, S.; Zawistowski, M.; Ellinghaus, E.; Barker, J.N.; Chandran, V.; Dand, N.; et al. Large scale meta-analysis characterizes genetic architecture for common psoriasis associated variants. Nat. Commun. 2017, 8, 15382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Pradhan, D.; Puri, P.; Ramesh, V.; Aggarwal, S.; Nayek, A.; Jain, A. Genomic alterations driving psoriasis pathogenesis. Gene 2019, 683, 61–71. [Google Scholar] [CrossRef]
- Mahil, S.K.; Capon, F.; Barker, J.N. Genetics of Psoriasis. Dermatol. Clin. 2015, 33, 1–11. [Google Scholar] [CrossRef]
- Széll, M.; Bata-Csörgő, Z.; Koreck, A.; Pivarcsi, A.; Polyánka, H.; Szeg, C.; Gaál, M.; Dobozy, A.; Kemény, L. Proliferating Keratinocytes Are Putative Sources of the Psoriasis Susceptibility-Related EDA+(Extra Domain A of Fibronectin) Oncofetal Fibronectin. J. Investig. Dermatol. 2004, 123, 537–546. [Google Scholar] [CrossRef]
- Sandig, H.; McDonald, J.; Gilmour, J.; Arno, M.; Lee, T.H.; Cousins, D. Fibronectin is a TH1-specific molecule in human subjects. J. Allergy Clin. Immunol. 2009, 124, 528–535.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubán, B.; Vas, K.; Balog, Z.; Manczinger, M.; Bebes, A.; Groma, G.; Szell, M.; Kemény, L.; Batacsorgő, Z. Abnormal regulation of fibronectin production by fibroblasts in psoriasis. Br. J. Dermatol. 2015, 174, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.S.; Fu, H.; Baratang, N.; Rousseau, J.; Kumra, H.; Sutton, V.R.; Niceta, M.; Ciolfi, A.; Yamamoto, G.; Bertola, D.; et al. Mutations in Fibronectin Cause a Subtype of Spondylometaphyseal Dysplasia with “Corner Fractures”. Am. J. Hum. Genet. 2017, 101, 815–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelletti, F.; Donadelli, R.; Banterla, F.; Hildebrandt, F.; Zipfel, P.F.; Bresin, E.; Otto, E.; Skerka, C.; Renieri, A.; Todeschini, M.; et al. Mutations in FN1 cause glomerulopathy with fibronectin deposits. Proc. Natl. Acad. Sci. USA 2008, 105, 2538–2543. [Google Scholar] [CrossRef] [Green Version]
- Gennaro, R.; Simonic, T.; Negri, A.; Mottola, C.; Secchi, C.; Ronchi, S.; Romeo, D. C5a fragment of bovine complement. Purification, bioassays, amino-acid sequence and other structural studies. J. Biol. Inorg. Chem. 1986, 155, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-Y.; Timpl, R.; Sasaki, T.; Chu, M.-L.; Ekblom, P. Fibulin-1 and fibulin-2 expression during organogenesis in the developing mouse embryo. Dev. Dyn. 1996, 205, 348–364. [Google Scholar] [CrossRef]
- Timpl, R.; Sasaki, T.; Kostka, G.; Chu, M.-L. Fibulins: A versatile family of extracellular matrix proteins. Nat. Rev. Mol. Cell Biol. 2003, 4, 479–489. [Google Scholar] [CrossRef]
- Twal, O.W.; Czirok, A.; Hegedus, B.; Knaak, C.; Chintalapudi, M.R.; Okagawa, H.; Sugi, Y.; Argraves, W.S. Fibulin-1 suppression of fibronectin-regulated cell adhesion and motility. J. Cell Sci. 2001, 114, 4587–4598. [Google Scholar] [CrossRef]
- Argraves, W.S.; Tanaka, A.; Smith, E.P.; Twal, W.O.; Argraves, K.M.; Fan, D.; Haudenschild, C.C. Fibulin-1 and fibrinogen in human atherosclerotic lesions. Histochem. Cell Biol. 2009, 132, 559–565. [Google Scholar] [CrossRef]
- Hayashido, Y.; Lucas, A.; Rougeot, C.; Godyna, S.; Argraves, W.S.; Rochefort, H. Estradiol and fibulin-1 inhibit motility of human ovarian- and breast-cancer cells induced by fibronectin. Int. J. Cancer 1998, 75, 654–658. [Google Scholar] [CrossRef]
- Geisterfer-Lowrance, A.A.; Kass, S.; Tanigawa, G.; Vosberg, H.-P.; McKenna, W.; Seidman, C.E.; Seidman, J. A molecular basis for familial hypertrophic cardiomyopathy: A β cardiac myosin heavy chain gene missense mutation. Cell 1990, 62, 999–1006. [Google Scholar] [CrossRef]
- Park, J.-M.; Kim, Y.J.; Yoo, J.H.; Bin Hong, Y.; Park, J.H.; Koo, H.; Chung, K.W.; Choi, B.-O. A novel MYH7 mutation with prominent paraspinal and proximal muscle involvement. Neuromuscul. Disord. 2013, 23, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Gad, A.; Nehru, V.; Ruusala, A.; Aspenström, P. RhoD regulates cytoskeletal dynamics via the actin nucleation–promoting factor WASp homologue associated with actin Golgi membranes and microtubules. Mol. Biol. Cell 2012, 23, 4807–4819. [Google Scholar] [CrossRef] [PubMed]
- Nehru, V.; Voytyuk, O.; Lennartsson, J.; Aspenström, P. RhoD Binds the Rab5 Effector Rabankyrin-5 and has a Role in Trafficking of the Platelet-derived Growth Factor Receptor. Traffic 2013, 14, 1242–1254. [Google Scholar] [CrossRef] [Green Version]
- Okuse, K.; Malik-Hall, M.; Baker, M.D.; Poon, W.-Y.L.; Kong, H.; Chao, M.V.; Wood, J.N. Annexin II light chain regulates sensory neuron-specific sodium channel expression. Nat. Cell Biol. 2002, 417, 653–656. [Google Scholar] [CrossRef]
- Faber, C.G.; Lauria, G.; Merkies, I.S.J.; Cheng, X.; Han, C.; Ahn, H.-S.; Persson, A.-K.; Hoeijmakers, J.G.J.; Gerrits, M.M.; Pierro, T.; et al. Gain-of-function Nav1.8 mutations in painful neuropathy. Proc. Natl. Acad. Sci. USA 2012, 109, 19444–19449. [Google Scholar] [CrossRef] [Green Version]
- Riol-Blanco, L.; Ordovas-Montanes, J.; Perro, M.; Naval, E.; Thiriot, A.; Alvarez, D.; Paust, S.; Wood, J.N.; Von Andrian, U.H. Nociceptive sensory neurons drive interleukin-23-mediated psoriasiform skin inflammation. Nature 2014, 510, 157–161. [Google Scholar] [CrossRef]
- Nanney, L.B.; Skeel, A.; Luan, J.; Polis, S.; Richmond, A.; Wang, M.-H.; Leonard, E.J. Proteolytic Cleavage and Activation of pro-Macrophage-Stimulating Protein and Upregulation of its Receptor in Tissue Injury. J. Investig. Dermatol. 1998, 111, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.H.; Ronsin, C.; Gesnel, M.C.; Coupey, L.; Skeel, A.; Leonard, E.J.; Breathnach, R. Identification of the ron gene product as the receptor for the human macrophage stimulating protein. Science 1994, 266, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zheng, H.; Cheung, A.K.L.; Tang, C.; Ko, J.; Wong, B.W.Y.; Leong, M.M.L.; Sham, P.C.; Cheung, F.; Kwong, D.L.-W.; et al. Whole-exome sequencing identifies MST1R as a genetic susceptibility gene in nasopharyngeal carcinoma. Proc. Natl. Acad. Sci. USA 2016, 113, 3317–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas-Madrid, D.; Nogales-Cadenas, R.; Pascual-Montano, A.D. GeneCodis3: A non-redundant and modular enrichment analysis tool for functional genomics. Nucleic Acids Res. 2012, 40, W478–W483. [Google Scholar] [CrossRef] [PubMed]
- Nogales-Cadenas, R.; Carmona-Saez, P.; Vazquez, M.; Vicente, C.; Yang, X.; Tirado, F.; Carazo, J.M.; Pascual-Montano, A. GeneCodis: Interpreting gene lists through enrichment analysis and integration of diverse biological information. Nucleic Acids Res. 2009, 37, W317–W322. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Saez, P.; Chagoyen, M.; Tirado, F.; Carazo, J.M.; Pascual-Montano, A. GENECODIS: A web-based tool for finding significant concurrent annotations in gene lists. Genome Biol. 2007, 8, R3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, B.; Tan, L.; Jin, Z.; Jiao, Y.; Fu, Y.; Liu, Y. MiR-892a Promotes Hepatocellular Carcinoma Cells Proliferation and Invasion Through Targeting CD226. J. Cell. Biochem. 2017, 118, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.-S.; Park, S.-S.; Hwang, B.; Moon, B.; Kim, W.T.; Kim, W.-J.; Moon, S.-K. MicroRNA-892b influences proliferation, migration and invasion of bladder cancer cells by mediating the p19ARF/cyclin D1/CDK6 and Sp-1/MMP-9 pathways. Oncol. Rep. 2016, 36, 2313–2320. [Google Scholar] [CrossRef]
- Jiang, L.; Yu, L.; Zhang, X.; Lei, F.; Wang, L.; Liu, X.; Wu, S.; Zhu, J.; Wu, G.; Cao, L.; et al. miR-892b Silencing Activates NF-κB and Promotes Aggressiveness in Breast Cancer. Cancer Res. 2016, 76, 1101–1111. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, M.; Wang, G.; Tian, Y.; He, X. Tumor promotor role of miR-647 in gastric cancer via repression of TP73. Mol. Med. Rep. 2018, 18, 3744–3750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Qu, D.; Li, W.; He, C.; Li, S.; Wu, G.; Zhao, Q.; Shen, L.; Zhang, J.; Zheng, J. miR-647 and miR-1914 promote cancer progression equivalently by downregulating nuclear factor IX in colorectal cancer. Mol. Med. Rep. 2017, 16, 8189–8199. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Wei, W.; Zhan, Z.; Xie, D.; Xie, Y.; Xiao, Q. Role of miR-647 in human gastric cancer suppression. Oncol. Rep. 2017, 37, 1401–1411. [Google Scholar] [CrossRef] [Green Version]
- Dattola, A.; Silvestri, M.; Tamburi, F.; Amoruso, G.F.; Bennardo, L.; Nisticò, S.P. Emerging role of anti-IL23 in the treatment of psoriasis: When humanized is very promising. Dermatol. Ther. 2020, 33, e14504. [Google Scholar] [CrossRef]
- Passante, M.; Dastoli, S.; Nisticò, S.P.; Bennardo, L.; Patruno, C. Effectiveness of brodalumab in acrodermatitis continua of Hallopeau: A case report. Dermatol. Ther. 2019, 33, e13170. [Google Scholar] [CrossRef] [PubMed]
- Ipe, J.; Swart, M.; Burgess, K.; Skaar, T. High-Throughput Assays to Assess the Functional Impact of Genetic Variants: A Road Towards Genomic-Driven Medicine. Clin. Transl. Sci. 2017, 10, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.R.; Tipney, H.; Painter, J.L.; Shen, J.; Nicoletti, P.; Shen, Y.; Floratos, A.; Sham, P.C.; Li, M.J.; Wang, J.; et al. The support of human genetic evidence for approved drug indications. Nat. Genet. 2015, 47, 856–860. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Clinical manifestation of inverse psoriasis.

Figure 2.
Confidence-based protein-interaction network for variant genes identified in inverse psoriasis.
Figure 2.
Confidence-based protein-interaction network for variant genes identified in inverse psoriasis.


{kind=link}
{kind=link}
Table 1.
Damaging or probably damaging missense mutations identified in patients with inverse psoriasis.
Table 1.
Damaging or probably damaging missense mutations identified in patients with inverse psoriasis.
| Variant ID | Location | Gene | Variant Effect | cDNA | Protein | MAF | Transcript | Domain | |
|---|---|---|---|---|---|---|---|---|---|
| rs1250209 | chr2:216235089 | FN1 | fibronectin 1 | Missense | c.6781G > A | p.Val2261Ile | <0.01 | NM_212482.2 | Fibronectin type 1 |
| rs136730 | chr22:45923827 | FBLN1 | fibulin 1 | Missense | c.422A > G | p.Gln141Arg | <0.01 | NM_006486.2 | Anaphylatoxin homologous |
| rs2425015 | chr20:33583331 | MYH7B | myosin heavy chain 7B | Missense | c.3019A > G | p.Lys1007Glu | <0.01 | NM_020884.4 | Coiled coil |
| rs4930409 | chr11:66837965 | RHOD | ras homolog family member D | Missense | c.400T > C | p.Cys134Arg | <0.01 | NM_014578.3 | RHO |
| rs6599241 | chr3:38739574 | SCN10A | sodium voltage-gated channel alpha subunit 10 | Missense | c.5137A > G | p.Met1713Val | <0.01 | NM_006514.3 | Transmembrane |
| rs7433231 | chr3:49928691 | MST1R | macrophage stimulating 1 receptor | Missense | c.3583A > G | p.Ser1195Gly | <0.01 | NM_002447.3 | Tyrosine kinase, catalytic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Göblös, A.; Varga, E.; Farkas, K.; Árvai, K.; Kemény, L. Genetic Investigation of Inverse Psoriasis. Life 2021, 11, 654. https://doi.org/10.3390/life11070654
AMA Style
Göblös A, Varga E, Farkas K, Árvai K, Kemény L. Genetic Investigation of Inverse Psoriasis. Life. 2021; 11(7):654. https://doi.org/10.3390/life11070654
Chicago/Turabian StyleGöblös, Anikó, Emese Varga, Katalin Farkas, Kristóf Árvai, and Lajos Kemény. 2021. "Genetic Investigation of Inverse Psoriasis" Life 11, no. 7: 654. https://doi.org/10.3390/life11070654
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.