
Mitochondriomics of Clarias Fishes (Siluriformes: Clariidae) with a New Assembly of Clarias camerunensis: Insights into the Genetic Characterization and Diversification
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Species Identification
2.2. DNA Extraction, Mitogenome Sequencing, and Assembly
2.3. Genomic Characterization and Comparative Analyses
2.4. Phylogenetic Analyses and Time Tree
3. Results and Discussion
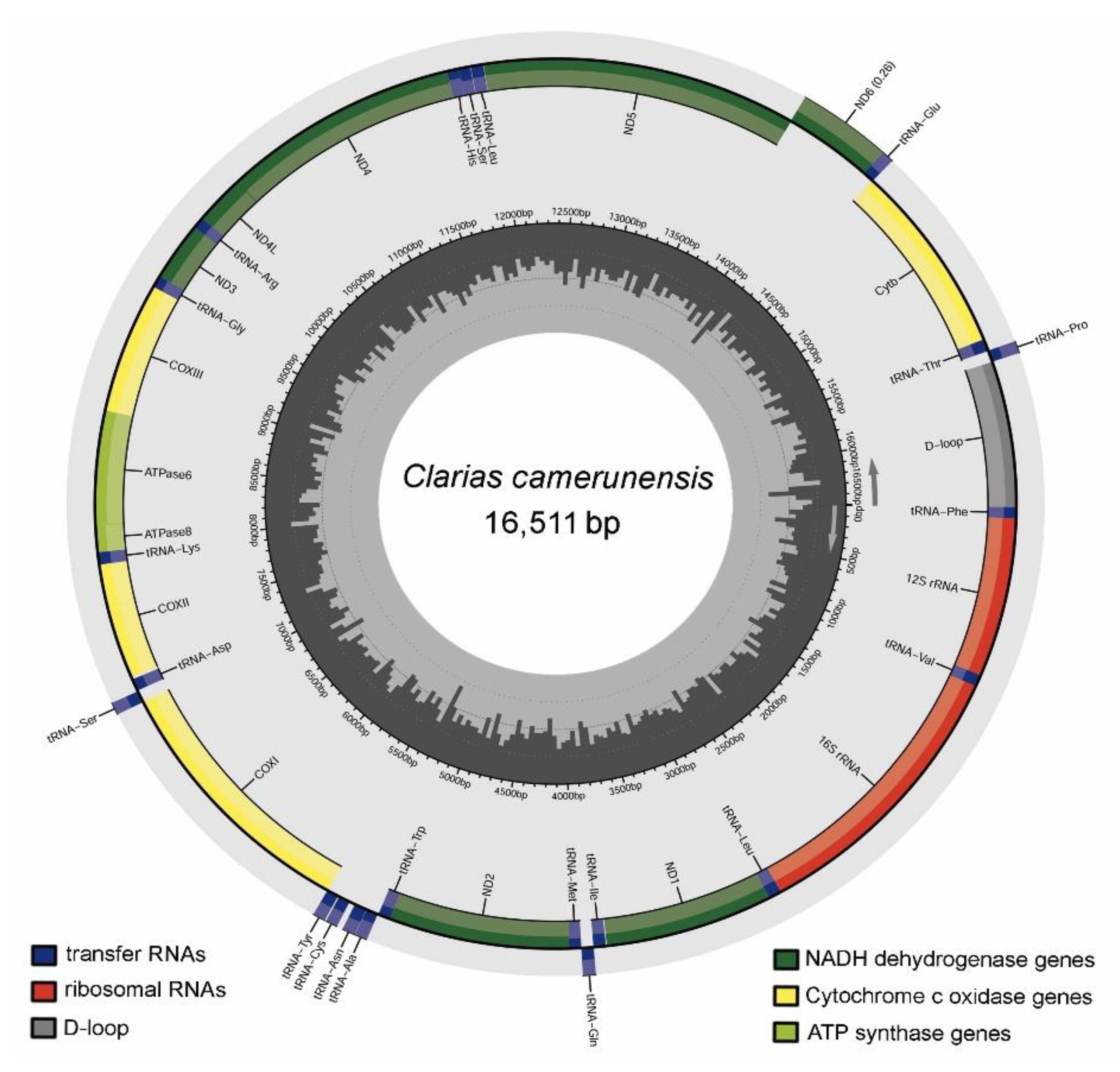
3.1. Mitogenomic Structure and Organization
3.2. Protein-Coding Genes (PCGs)
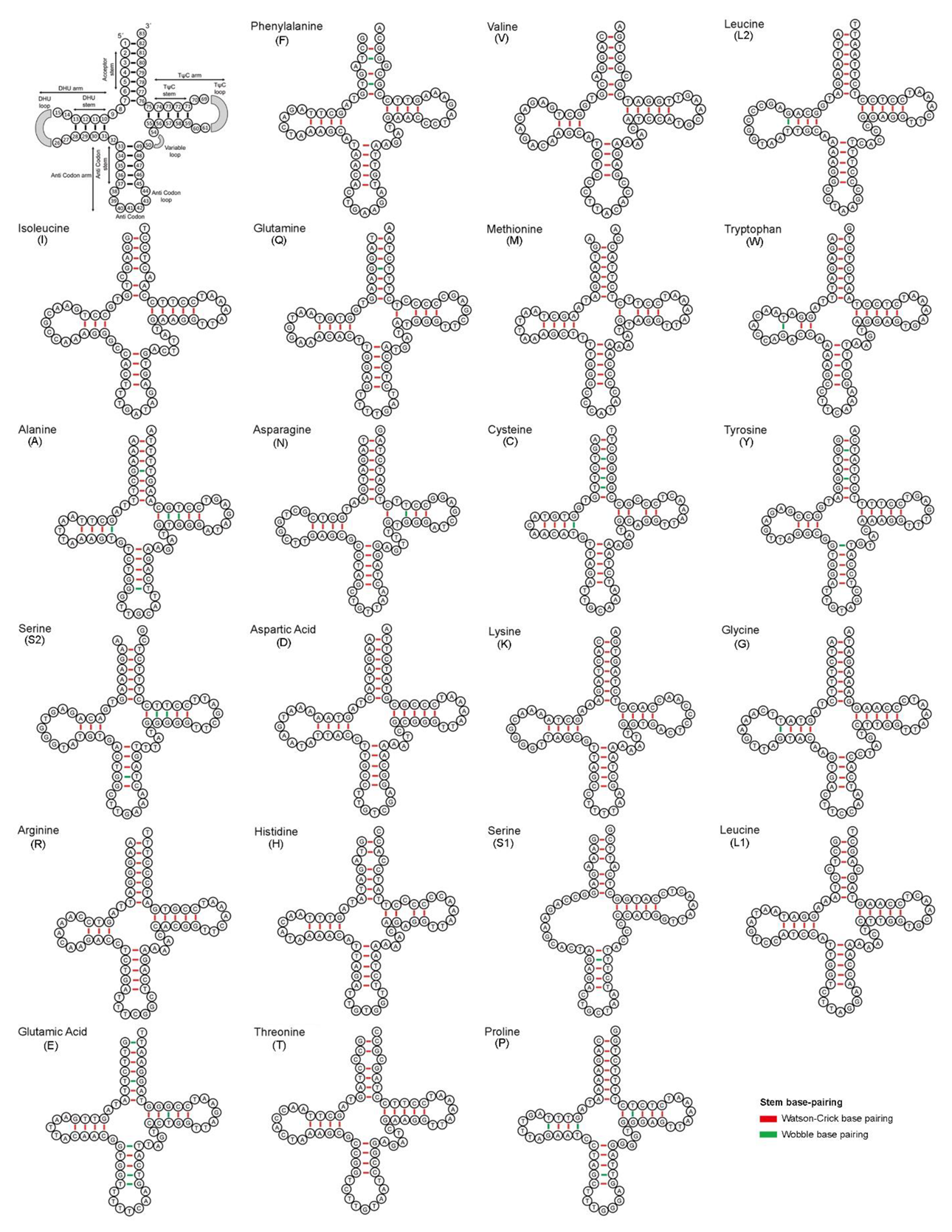
3.3. Ribosomal RNA (rRNA) and Transfer RNA (tRNA)
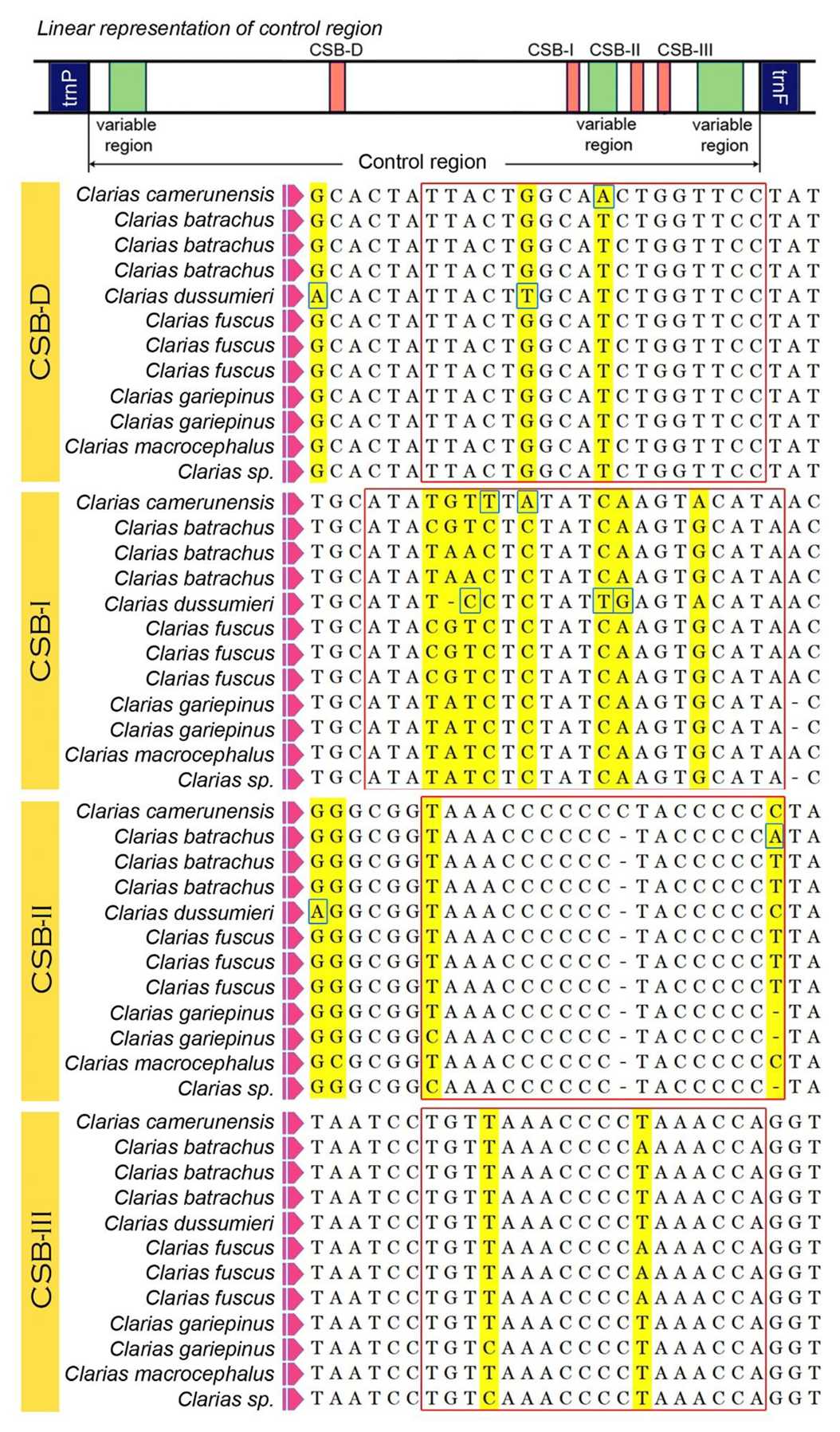
3.4. Control Regions
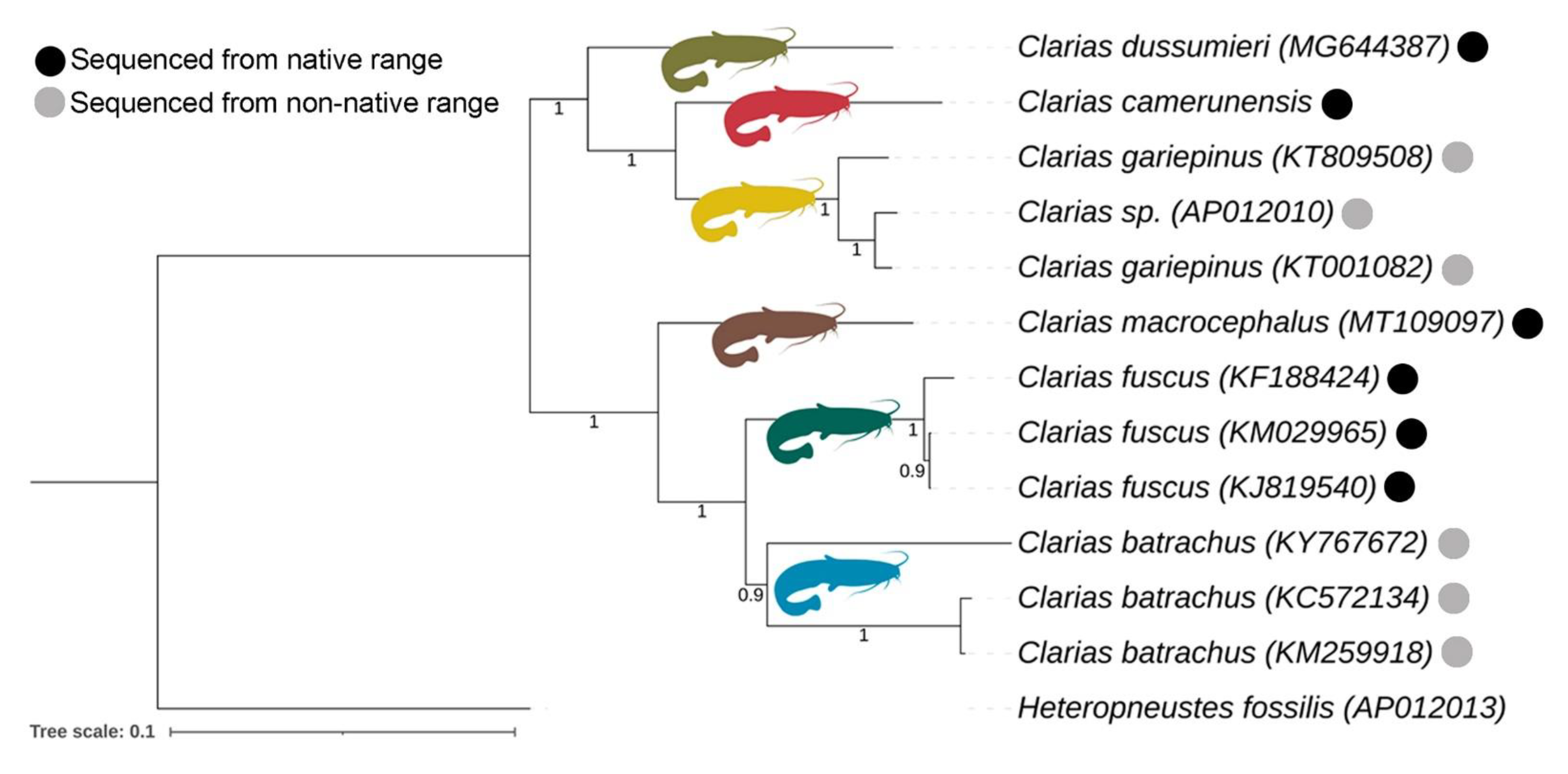
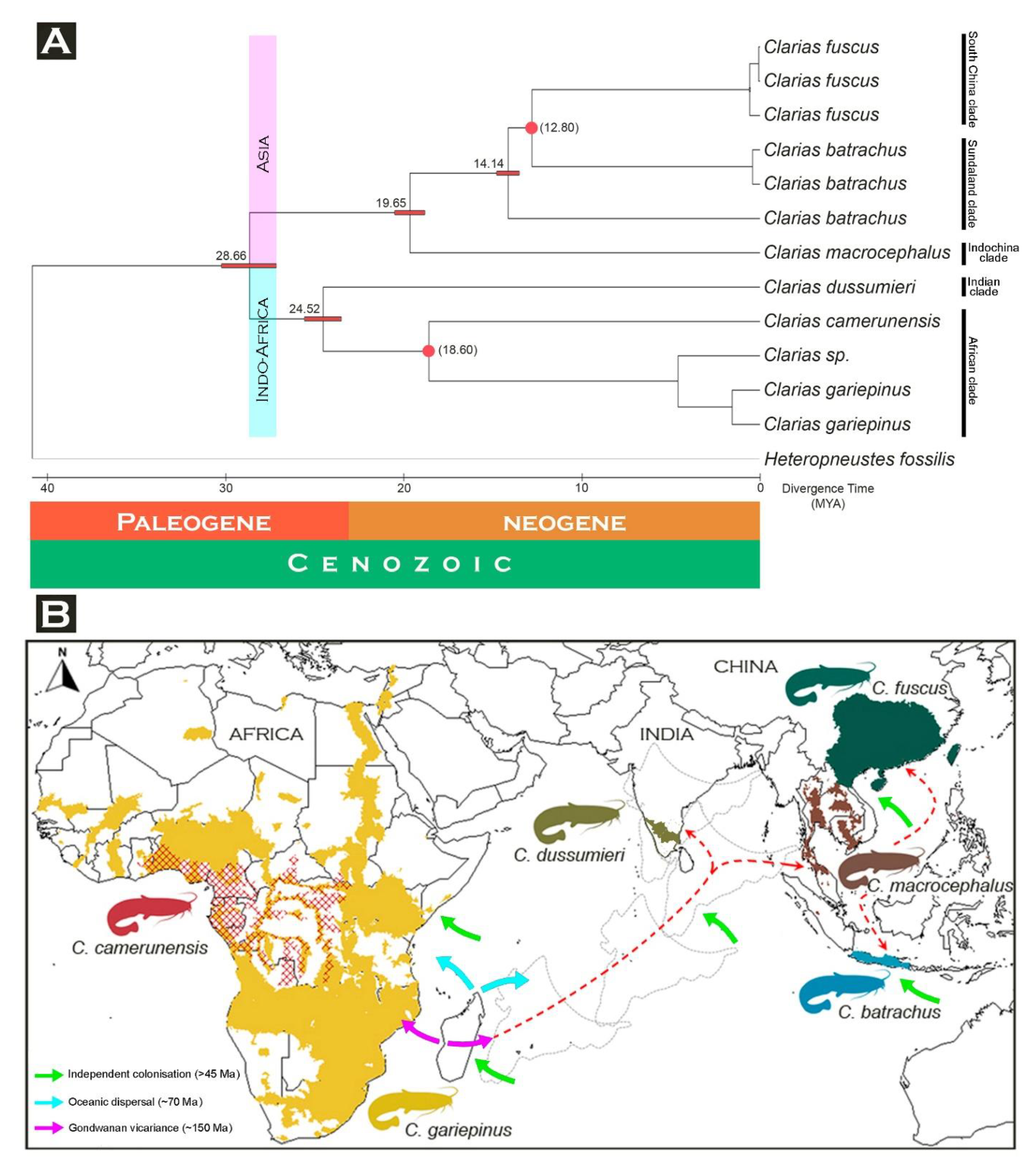
3.5. Matrilineal Phylogeny and Divergence Time
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbarossa, V.; Bosmans, J.; Wanders, N.; King, H.; Bierkens, M.F.P.; Huijbregts, M.A.J.; Schipper, A.M. Threats of global warming to the world’s freshwater fishes. Nat. Commun. 2021, 12, 1701. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication. Version (08/2022). 2022. Available online: www.fishbase.org (accessed on 26 January 2023).
- Teugels, G.G.; Adriaens, D. Taxonomy and phylogeny of Clariidae—An overview. In Catfishes; Arratia, G., Kapoor, B.G., Chardon, M., Diogo, R., Eds.; Science Publishers: Enfield, NH, USA, 2003; pp. 465–487. [Google Scholar]
- Seegers, L. The Catfishes of Africa: A Handbook for Identification and Maintenance; Aqualog Verlag A.C.S. GmbH: Rodgau, Germany, 2008; 604p. [Google Scholar]
- Hanssens, M. A review of the Clarias species (Pisces; Siluriformes) from the Lower Congo and the Pool Malebo. J. Afr. Zool. 2009, 5, 27–40. [Google Scholar]
- Pasch, J.; Palm, H.W. Economic Analysis and Improvement Opportunities of African Catfish (Clarias gariepinus) Aquaculture in Northern Germany. Sustainability 2021, 13, 13569. [Google Scholar] [CrossRef]
- Ekawati, A.W.; Ulfa, S.M.; Dewi, C.S.U.; Amin, A.A.; Salamah, L.N.; Yanuar, A.T.; Kurniawan, A. Analysis of Aquaponic Recirculation Aquaculture System (A-Ras) Application in the Catfish (Clarias gariepinus) Aquaculture in Indonesia. Aquac. Stud. 2021, 21, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Chala, B.; Erko, B.; Animut, A.; Degarege, A.; Petros, B. Assessment of Clarias gariepinus as a biological control agent against mosquito larvae. BMC Ecol. 2016, 16, 27. [Google Scholar] [CrossRef]
- Parvez, I.; Rumi, R.A.; Ray, P.R.; Hassan, M.M.; Sultana, S.; Pervin, R.; Suwanno, S.; Pradit, S. Invasion of African Clarias gariepinus Drives Genetic Erosion of the Indigenous C. batrachus in Bangladesh. Biology 2022, 11, 252. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Zimmermann, S.; Smit, N.J.; Malherbe, W.; Nachev, M.; Sures, B.; Wepener, V. Human health risks associated with consumption of fish contaminated with trace elements from intensive mining activities in a peri-urban region. Sci. Total Environ. 2022, 825, 154011. [Google Scholar] [CrossRef]
- Lakra, K.C.; Banerjee, T.K.; Lal, B. Coal mine effluent-induced metal bioaccumulation, biochemical, oxidative stress, metallothionein, and histopathological alterations in vital tissues of the catfish, Clarias batrachus. Environ. Sci. Pollut. Res. 2021, 28, 25300–25315. [Google Scholar] [CrossRef]
- Okonkwo, J.C.; Obiakor, M.O. Karyological and chromosomal study of catfish (Clariidae, Clarias gariepinus, Burchell, 1822) from Anambra River, Anambra state, Nigeria. Pak. J. Nutr. 2010, 9, 112–115. [Google Scholar] [CrossRef]
- Rahman, M.A.; Arshad, A.; Marimuthu, K.; Ara, R.; Amin, S.M.N. Inter-specific hybridization and its potential for aquaculture of fin fishes. Asian J. Anim. Vet. Adv. 2013, 8, 139–153. [Google Scholar]
- Wachirachaikarn, A.; Na-Nakorn, U. Genetic diversity of the North African catfish, Clarias gariepinus (Burchell, 1822) hatchery stocks in Thailand. Sci. Asia 2019, 45, 301–308. [Google Scholar] [CrossRef]
- Nguyen, D.H.M.; Panthum, T.; Ponjarat, J.; Laopichienpong, N.; Kraichak, E.; Singchat, W.; Ahmad, S.F.; Muangmai, N.; Peyachoknagul, S.; Na-Nakorn, U.; et al. An Investigation of ZZ/ZW and XX/XY Sex Determination Systems in North African Catfish (Clarias gariepinus, Burchell, 1822). Front. Genet. 2021, 11, 562856. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.L.; Zheng, B.S.; Dai, D.Y. Fauna Sinica, Class Teleostei, Siluriformes; Scientific Press: Beijing, China, 1999. [Google Scholar]
- Das, S.K. Seed production of magur (Clarias batrachus) using a rural model portable hatchery in Assam, India-A farmer proven technology. Aquac. Asia 2002, 7, 19–21. [Google Scholar]
- Ng, H.H.; Kottelat, M. The identity of Clarias batrachus (Linnaeus 1758) with the designation of a neotype (Teleostei: Clariidae). Zool. J. Linn. Soc. 2008, 153, 725–732. [Google Scholar] [CrossRef]
- Emiroğlu, Ö.; Atalay, M.A.; Ekmekçi, F.G.; Aksu, S.; Başkurt, S.; Keskin, E.; Tarkan, A.S.; Ünal, E.M.; Yoğurtçuoğlu, B. One of the world’s worst invasive species, Clarias batrachus (Actinopterygii: Siluriformes: Clariidae), has arrived and established a population in turkey. Acta Ichthyol. Piscat. 2020, 50, 391–400. [Google Scholar] [CrossRef]
- Akoll, P.; Mwanja, W.W. Fish health status, research and management in East Africa: Past and present. Afr. J. Aquat. Sci. 2012, 37, 117–129. [Google Scholar] [CrossRef]
- Taukhid, P.U.; Sugiani, D.; Sumiati, T.; Lusiastuti, A.M. Efficacious of in-active bacterial vaccines: Aeromonas hydrophila-AHL0905-2 (Hydrovac) and Streptococcus agalactiae-N14G (Streptovac) for prevention of bacterial diseases on freshwater aquaculture. J. Ris. Akuakultur 2015, 10, 541–551. [Google Scholar] [CrossRef]
- Suprapto, R.; Alimuddin, A.; Nuryati, S.; Imron, I.; Marnis, H.; Iswanto, B. MHC-II marker potential linked to Motile Aeromonad Septicaemia disease resistance in African catfish (Clarias gariepinus). Indones. Aquac. J. 2017, 12, 21–28. [Google Scholar] [CrossRef]
- Agnese, J.F.; Teugels, G.G. Insight into the phylogeny of African Clariidae (Teleostei, Siluriformes): Implications for their body shape evolution, biogeography and taxonomy. Mol. Phylogenet. Evol. 2005, 36, 546–553. [Google Scholar] [CrossRef]
- Iyiola, O.A.; Nneji, L.M.; Mustapha, M.K.; Nzeh, C.G.; Oladipo, S.O.; Nneji, I.C.; Okeyoyin, A.O.; Nwani, C.D.; Ugwumba, O.A.; Ugwumba, A.A.A.; et al. DNA barcoding of economically important freshwater fish species from north-central Nigeria uncovers cryptic diversity. Ecol. Evol. 2018, 8, 6932–6951. [Google Scholar] [CrossRef]
- Pouyaud, L.; Sudarto, T.; Paradis, E. The phylogenetic structure of habitat shift and morphological convergence in Asian Clarias (Teleostei, Siluriformes: Clariidae). J. Zool. Syst. Evol. Res. 2009, 47, 344–356. [Google Scholar] [CrossRef]
- Chand, G.B.; Kumar, S.; Azad, G.K. Molecular assessment of proteins encoded by the mitochondrial genome of Clarias batrachus and Clarias gariepinus. Biochem. Biophys. Rep. 2021, 26, 100985. [Google Scholar] [CrossRef] [PubMed]
- Duong, T.Y.; Tan, M.H.; Lee, Y.P.; Croft, L.; Austin, C.M. Dataset for genome sequencing and de novo assembly of the Vietnamese bighead catfish (Clarias macrocephalus Günther, 1864). Data Brief 2020, 31, 105861. [Google Scholar] [CrossRef]
- Han, C.; Li, Q.; Xu, J.; Li, X.; Huang, J. Characterization of Clarias gariepinus mitochondrial genome sequence and a comparative analysis with other catfishes. Biologia 2015, 70, 1245–1253. [Google Scholar] [CrossRef]
- Kushwaha, B.; Kumar, R.; Agarwal, S.; Pandey, M.; Nagpure, N.S.; Singh, M.; Srivastava, S.; Joshi, C.G.; Das, P.; Sahoo, L.; et al. Assembly and variation analyses of Clarias batrachus mitogenome retrieved from WGS data and its phylogenetic relationship with other catfishes. Meta Gene 2015, 5, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Bao, L.; Zhou, T.; Yuan, Z.; Liu, S.; Dunham, R.; Li, Y.; Wang, K.; Xu, X.; Jin, Y.; et al. Genome sequence of walking catfish (Clarias batrachus) provides insights into terrestrial adaptation. BMC Genom. 2018, 19, 952. [Google Scholar] [CrossRef]
- Mohindra, V.; Singh, R.K.; Kumar, R.; Sah, R.S.; Lal, K.K. Complete mitochondrial genome sequences of two endangered Indian catfish species, Clarias batrachus and Pangasius pangasius. Mitochondrial DNA 2015, 26, 678–679. [Google Scholar] [CrossRef]
- Yang, H.; Sun, J.; Zhao, H.; Chen, Y.; Yang, Z.; Li, G.; Liu, L. The complete mitochondrial genome of the Clarias fuscus (Siluriformes, Clariidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 1255–1256. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, X.; Guan, L.; He, S. The complete mitochondrial genome of Clarias fuscus (Teleostei, Siluriformes: Clariidae). Mitochondrial DNA 2015, 26, 270–271. [Google Scholar] [CrossRef]
- Elberri, A.I.; Galal-Khallaf, A.; Gibreel, S.E.; El-Sakhawy, S.F.; El-Garawani, I.; El-Sayed Hassab ElNabi, S.; Mohammed-Geba, K. DNA and eDNA-based tracking of the North African sharptooth catfish Clarias gariepinus. Mol. Cell. Probes 2020, 51, 101535. [Google Scholar] [CrossRef]
- Miya, M.; Kawaguchi, A.; Nishida, M. Mitogenomic exploration of higher teleostean phylogenies: A case study for moderate-scale evolutionary genomics with 38 newly determined complete mitochondrial DNA sequences. Mol. Biol. Evol. 2001, 18, 1993–2009. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Miya, M.; Fukunaga, T.; Sado, T.; Iwasaki, W. MitoFish and MiFish Pipeline: A Mitochondrial Genome Database of Fish with an Analysis Pipeline for Environmental DNA Metabarcoding. Mol. Biol. Evol. 2018, 35, 1553–1555. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.P.; Miya, M.; Mabuchi, K.; Nishida, M. Structure and variation of the mitochondrial genome of fishes. BMC Genom. 2016, 17, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo Metazoan Mitochondrial Genome Annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, W.; Fukunaga, T.; Isagozawa, R.; Yamada, K.; Maeda, Y.; Satoh, T.P.; Sado, T.; Mabuchi, K.; Takeshima, H.; Miya, M.; et al. MitoFish and MitoAnnotator: A mitochondrial genome database of fish with an accurate and automatic annotation pipeline. Mol. Biol. Evol. 2013, 30, 2531–2540. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–359. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Laslett, D.; Canbäck, B. ARWEN, a program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Vences, M.; Miralles, A.; Brouillet, S.; Ducasse, J.; Fedosov, A.; Kharchev, V.; Kostadinov, I.; Kumari, S.; Patmanidis, S.; Scherz, M.D.; et al. iTaxoTools 0.1: Kickstarting a specimen-based software toolkit for taxonomists. Megataxa 2021, 6, 77–92. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Suleski, M.; Craig, J.M.; Kasprowicz, A.E.; Sanderford, M.; Sanderford, M.; Li, M.; Stecher, G.; Hedges, S.B. TimeTree 5: An Expanded Resource for Species Divergence Times. Mol. Biol. Evol. 2022, 39, msac174. [Google Scholar] [CrossRef] [PubMed]
- Rabosky, D.L.; Santini, F.; Eastman, J.; Smith, S.A.; Sidlauskas, B.; Chang, J.; Alfaro, M.E. Rates of speciation and morphological evolution are correlated across the largest vertebrate radiation. Nat. Commun. 2013, 4, 1958. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Chang, J.; Title, P.O.; Cowman, P.F.; Sallan, L.; Friedman, M.; Kaschner, K.; Garilao, C.; Near, T.J.; Coll, M.; et al. An inverse latitudinal gradient in speciation rate for marine fishes. Nature 2018, 559, 392–395. [Google Scholar] [CrossRef]
- Kundu, S.; Kumar, V.; Tyagi, K.; Chakraborty, R.; Chandra, K. The first complete mitochondrial genome of the Indian Tent Turtle, Pangshura tentoria (Testudines: Geoemydidae): Characterization and comparative analysis. Ecol. Evol. 2019, 9, 10854–10868. [Google Scholar] [CrossRef]
- Kundu, S.; Pakrashi, A.; Kamalakannan, M.; Singha, D.; Tyagi, K.; Banerjee, D.; Venkatraman, C.; Kumar, V. Complete mitogenome of the endangered and endemic Nicobar treeshrew (Tupaia nicobarica) and comparison with other Scandentians. Sci. Rep. 2022, 12, 877. [Google Scholar] [CrossRef]
- Da Fonseca, R.R.; Johnson, W.E.; O’Brien, S.J.; Ramos, M.J.; Antunes, A. The adaptive evolution of the mammalian mitochondrial genome. BMC Genom. 2008, 9, 119. [Google Scholar] [CrossRef]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef]
- Foote, A.D.; Morin, P.A.; Durban, J.W.; Pitman, R.L.; Wade, P.; Willerslev, E.; Gilbert, M.T.; da Fonseca, R.R. Positive selection on the killer whale mitogenome. Biol. Lett. 2011, 7, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.; Enbody, E.D.; Pettersson, M.E.; Sprehn, C.G.; Bekkevold, D.; Folkvord, A.; Laikre, L.; Kleinau, G.; Scheerer, P.; Andersson, L. Recurrent convergent evolution at amino acid residue 261 in fish rhodopsin. Proc. Natl. Acad. Sci. USA 2019, 116, 18473–18478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, N.S.; Hirabayashi, N.; Agmon, I.; Yonath, A.; Suzuki, T. Comprehensive genetic selection revealed essential bases in the peptidyl-transferase center. Proc. Natl. Acad. Sci. USA 2006, 103, 15386–15391. [Google Scholar] [CrossRef] [PubMed]
- Varani, G.; McClain, W.H. The G-U wobble base pair: A fundamental building block of RNA structure crucial to RNA function in diverse biological systems. EMBO Rep. 2000, 1, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Conroy, J.; Howell, W.H.; Kocher, T.D. Structure and evolution of teleost mitochondrial control regions. J. Mol. Evol. 1995, 41, 54–66. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, X.; Nie, L. Organization and variation of mitochondrial DNA control region in pleurodiran turtles. Zoologia 2011, 28, 495–504. [Google Scholar] [CrossRef]
- Toews, D.P.; Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907–3930. [Google Scholar] [PubMed]
- Tamura, K.; Battistuzzi, F.U.; Billing-Ross, P.; Murillo, O.; Filipski, A.; Kumar, S. Estimating Divergence Times in Large Molecular Phylogenies. Proc. Natl. Acad. Sci. USA 2012, 109, 19333–19338. [Google Scholar] [CrossRef]
- Kappas, I.; Vittas, S.; Pantzartzi, C.N.; Drosopoulou, E.; Scouras, Z.G. A Time-Calibrated Mitogenome Phylogeny of Catfish (Teleostei: Siluriformes). PLoS ONE 2016, 11, e0166988. [Google Scholar] [CrossRef]
- Che, J.; Zhou, W.W.; Hu, J.S.; Yan, F.; Papenfuss, T.J.; Wake, D.B.; Zhang, Y.P. Spiny frogs (Paini) illuminate the history of the Himalayan region and Southeast Asia. Proc. Natl. Acad. Sci. USA 2010, 107, 13765–13770. [Google Scholar] [CrossRef]
- Corn, K.A.; Friedman, S.T.; Burress, E.D.; Martinez, C.M.; Larouche, O.; Price, S.A.; Wainwright, P.C. The rise of biting during the Cenozoic fueled reef fish body shape diversification. Proc. Natl. Acad. Sci. USA 2022, 119, e2119828119. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, S. The stepwise Indian-Eurasian collision and uplift of the Himalayan-Tibetan plateau drove the diversification of high-elevation Scytodes spiders. Cladistics 2022, 38, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Anoop, V.K.; Dahanukar, N.; Philip, S.; Thomas, L.; Raghavan, R. Phylogeny of the hillstream loach genus Mesonoemacheilus reveals widespread diversification through ancient drainage connections in the Western Ghats Biodiversity Hotspot. Mol. Phylogenetics Evol. 2018, 129, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Klaus, S.; Morley, R.J.; Plath, M.; Zhang, Y.-P.; Li, J.-T. Biotic interchange between the Indian subcontinent and mainland Asia through time. Nat. Commun. 2016, 7, 12132. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Li, Y.; Klaus, S.; Rao, D.Q.; Hillis, D.M.; Zhang, Y.P. Diversification of rhacophorid frogs provides evidence for accelerated faunal exchange between India and Eurasia during the Oligocene. Proc. Natl. Acad. Sci. USA 2013, 110, 3441–3446. [Google Scholar] [CrossRef]
- Yamahira, K.; Ansai, S.; Kakioka, R.; Yaguchi, H.; Kon, T.; Montenegro, J.; Kobayashi, H.; Fujimoto, S.; Kimura, R.; Takehana, Y. Mesozoic origin and ‘out-of-India’ radiation of ricefishes (Adrianichthyidae). Biol. Lett. 2021, 17, 20210212. [Google Scholar] [CrossRef]
- Warren, B.H.; Strasberg, D.; Bruggemann, J.H.; Prys-Jones, R.P.; Thébaud, C. Why does the biota of the Madagascar region have such a strong Asiatic flavour? Cladistics 2010, 26, 526–538. [Google Scholar] [CrossRef]
- Van Steenberge, M.W.; Vanhove, M.P.M.; Manda, A.C.; Larmuseau, M.H.D.; Swart, B.L.; Khang’Mate, F.; Arndt, A.; Hellemans, B.; van Houdt, J.; Micha, J.-C.; et al. Unravelling the evolution of Africa’s drainage basins through a widespread freshwater fish, the African sharptooth catfish Clarias gariepinus. J. Biogeogr. 2020, 47, 1739–1754. [Google Scholar] [CrossRef]
- Day, J.J.; Peart, C.R.; Brown, K.J.; Friel, J.P.; Bills, R.; Moritz, T. Continental diversification of an African Catfish Radiation (Mochokidae: Synodontis). Syst. Biol. 2013, 62, 351–365. [Google Scholar] [CrossRef]
- Elmer, K.R.; Reggio, C.; Wirth, T.; Verheyen, E.; Salzburger, W.; Meyer, A. Pleistocene desiccation in East Africa bottlenecked but did not extirpate the adaptive radiation of Lake Victoria haplochromine cichlid fishes. Proc. Natl. Acad. Sci. USA 2009, 106, 13404–13409. [Google Scholar] [CrossRef] [PubMed]
- Matschiner, M.; Böhne, A.; Ronco, F.; Salzburger, W. The genomic timeline of cichlid fish diversification across continents. Nat. Commun. 2020, 11, 5895. [Google Scholar] [CrossRef] [PubMed]
- Feijó, A.; Ge, D.; Wen, Z.; Cheng, J.; Xia, L.; Patterson, B.D.; Yang, Q. Mammalian diversification bursts and biotic turnovers are synchronous with Cenozoic geoclimatic events in Asia. Proc. Natl. Acad. Sci. USA 2022, 119, e2207845119. [Google Scholar] [CrossRef] [PubMed]
- Grobler, J.P.; Ndyogolo, S.; Barasa, J.; Abila, R.; Bindeman, H.; Schlemmer, A.F.J. Genetic Identification of Invasive Walking Catfish, Clarias batrachus, Intermingled with African Catfish, C. gariepinus, in South Africa. Afr. J. Wildl. Res. 2015, 45, 55–62. [Google Scholar] [CrossRef]
- Falaye, A.E.; Omoike, A.; Ajani, E.K.; Olanrewaju, A.N. Comparative growth performance and survival of monosex and mixed sex population of African catfish (Clarias gariepinus, Burchell, 1822) under culture system. Niger. J. Sci. 2011, 45, 75–81. [Google Scholar]
- Erondu, E.; Gabriel, U.U.; Akinrotimi, O. Genetic manipulation for enhanced aquaculture production in Nigeria. Biosci. Res. J. 2011, 23, 131–140. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Start | End | Strand | Size (bp) | Intergenic Nucleotide | Anti-Codon | Start Codon | Stop Codon |
|---|---|---|---|---|---|---|---|---|
| tRNA-Phe (F) | 1 | 69 | H | 69 | 0 | TTC | . | . |
| 12S rRNA | 70 | 1022 | H | 953 | 0 | . | . | . |
| tRNA-Val (V) | 1023 | 1094 | H | 72 | 0 | GTA | . | . |
| 16S rRNA | 1095 | 2767 | H | 1673 | 0 | . | . | . |
| tRNA-Leu (L2) | 2768 | 2842 | H | 75 | 0 | TTA | . | . |
| ND1 | 2843 | 3817 | H | 975 | 5 | . | ATG | TAA |
| tRNA-Ile (I) | 3823 | 3894 | H | 72 | −1 | ATC | . | . |
| tRNA-Gln (Q) | 3894 | 3964 | L | 71 | −1 | CAA | . | . |
| tRNA-Met (M) | 3964 | 4033 | H | 70 | 0 | ATG | . | . |
| ND2 | 4034 | 5078 | H | 1045 | 0 | . | ATG | T-- |
| tRNA-Trp (W) | 5079 | 5149 | H | 71 | 3 | TGA | . | . |
| tRNA-Ala (A) | 5153 | 5221 | L | 69 | 1 | GCA | . | . |
| tRNA-Asn (N) | 5223 | 5295 | L | 73 | 33 | AAC | . | . |
| tRNA-Cys (C) | 5329 | 5395 | L | 67 | 7 | TGC | . | . |
| tRNA-Tyr (Y) | 5403 | 5472 | L | 70 | 1 | TAC | . | . |
| COI | 5474 | 7024 | H | 1551 | 0 | . | GTG | TAA |
| tRNA-Ser (S2) | 7025 | 7095 | L | 71 | 4 | TCA | . | . |
| tRNA-Asp (D) | 7100 | 7172 | H | 73 | 14 | GAC | . | . |
| COII | 7187 | 7877 | H | 691 | 0 | . | ATG | T-- |
| tRNA-Lys (K) | 7878 | 7951 | H | 74 | 1 | AAA | . | . |
| ATP8 | 7953 | 8120 | H | 168 | −10 | . | ATG | TAA |
| ATP6 | 8111 | 8793 | H | 683 | 0 | . | ATG | TA- |
| COIII | 8794 | 9577 | H | 784 | 0 | . | ATG | T-- |
| tRNA-Gly (G) | 9578 | 9650 | H | 73 | 0 | GGA | . | . |
| ND3 | 9651 | 9999 | H | 349 | 0 | . | ATG | T-- |
| tRNA-Arg (R) | 10,000 | 10,068 | H | 69 | 0 | CGA | . | . |
| ND4L | 10,069 | 10,365 | H | 297 | −7 | . | ATG | TAA |
| ND4 | 10,359 | 11,739 | H | 1381 | 0 | . | ATG | T-- |
| tRNA-His (H) | 11,740 | 11,809 | H | 70 | 0 | CAC | . | . |
| tRNA-Ser (S1) | 11,810 | 11,876 | H | 67 | 2 | AGC | . | . |
| tRNA-Leu (L1) | 11,879 | 11,951 | H | 73 | 2 | CTA | . | . |
| ND5 | 11,954 | 13,777 | H | 1824 | −4 | . | ATA | TAA |
| ND6 | 13,774 | 14,292 | L | 519 | 0 | . | ATG | TAA |
| tRNA-Glu (E) | 14,293 | 14,361 | L | 69 | 1 | GAA | . | . |
| Cyt b | 14,363 | 15,500 | H | 1138 | 0 | . | ATG | T-- |
| tRNA-Thr (T) | 15,501 | 15,573 | H | 73 | −2 | ACA | . | . |
| tRNA-Pro (P) | 15,572 | 15,641 | L | 70 | 0 | CCA | . | . |
| Control region | 15,642 | 16,511 | H | 870 | . | . | . | . |
| Species Name | Size (bp) | A% | T% | G% | C% | A + T% | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|---|
| Complete mitogenome | ||||||||
| C. camerunensis | 16,511 | 32.28 | 24.61 | 14.87 | 28.24 | 56.89 | 0.135 | −0.310 |
| C. batrachus | 16,511 | 32.31 | 24.97 | 15.3 | 27.42 | 57.28 | 0.128 | −0.284 |
| C. dussumieri | 16,514 | 32.52 | 26.11 | 14.39 | 26.97 | 58.63 | 0.223 | −0.304 |
| C. fuscus | 16,525 | 32.17 | 25.34 | 14.95 | 27.54 | 57.51 | 0.119 | −0.296 |
| C. gariepinus | 16,508 | 32.53 | 24.68 | 14.84 | 27.96 | 57.21 | 0.137 | −0.307 |
| C. macrocephalus | 16,511 | 32.2 | 25.35 | 14.87 | 27.58 | 57.55 | 0.119 | −0.299 |
| PCGs | ||||||||
| C. camerunensis | 11,404 | 30.66 | 26.39 | 14.6 | 28.35 | 57.05 | 0.075 | −0.320 |
| C. batrachus | 11,417 | 30.61 | 26.84 | 14.95 | 27.6 | 57.45 | 0.066 | −0.297 |
| C. dussumieri | 11,408 | 30.86 | 28.18 | 13.99 | 26.96 | 59.04 | 0.441 | −0.317 |
| C. fuscus | 11,422 | 30.54 | 27.33 | 14.56 | 27.57 | 57.87 | 0.055 | −0.309 |
| C. gariepinus | 11,408 | 30.96 | 26.51 | 14.42 | 28.11 | 57.47 | 0.077 | −0.322 |
| C. macrocephalus | 11,408 | 30.51 | 27.32 | 14.54 | 27.62 | 57.83 | 0.055 | −0.310 |
| rRNAs | ||||||||
| C. camerunensis | 2626 | 34.69 | 20.26 | 19.46 | 25.59 | 54.95 | 0.263 | −0.136 |
| C. batrachus | 2660 | 34.51 | 20.53 | 20.23 | 24.74 | 55.04 | 0.254 | −0.261 |
| C. dussumieri | 2627 | 34.64 | 20.86 | 19.68 | 24.82 | 55.5 | 0.325 | −0.115 |
| C. fuscus | 2618 | 34.45 | 20.82 | 19.9 | 24.83 | 55.27 | 0.247 | −0.110 |
| C. gariepinus | 2627 | 34.64 | 20.25 | 19.68 | 25.43 | 54.89 | 0.262 | −0.127 |
| C. macrocephalus | 2622 | 34.48 | 20.59 | 19.87 | 25.06 | 55.07 | 0.252 | −0.115 |
| tRNAs | ||||||||
| C. camerunensis | 1561 | 29.34 | 27.61 | 22.42 | 20.63 | 56.95 | 0.030 | 0.042 |
| C. batrachus | 1560 | 29.42 | 27.56 | 22.31 | 20.71 | 56.98 | 0.033 | 0.037 |
| C. dussumieri | 1560 | 29.17 | 27.95 | 22.37 | 20.51 | 57.12 | 0.190 | 0.043 |
| C. fuscus | 1560 | 29.04 | 27.56 | 22.56 | 20.83 | 56.6 | 0.026 | 0.040 |
| C. gariepinus | 1560 | 28.85 | 27.69 | 22.82 | 20.64 | 56.54 | 0.020 | 0.050 |
| C. macrocephalus | 1561 | 29.08 | 28.12 | 22.55 | 20.24 | 57.2 | 0.017 | 0.054 |
| CRs | ||||||||
| C. camerunensis | 870 | 30.69 | 31.03 | 14.14 | 24.14 | 61.72 | -0.006 | −0.261 |
| C. batrachus | 871 | 31.46 | 31.23 | 13.89 | 23.42 | 62.69 | 0.004 | −0.255 |
| C. dussumieri | 870 | 33.22 | 32.87 | 12.41 | 21.49 | 66.09 | 0.503 | −0.268 |
| C. fuscus | 871 | 30.08 | 31.23 | 14.24 | 24.45 | 61.31 | -0.019 | −0.264 |
| C. gariepinus | 863 | 32.21 | 30.48 | 14.6 | 22.71 | 62.69 | 0.028 | −0.217 |
| C. macrocephalus | 869 | 31.99 | 30.38 | 14.15 | 23.48 | 62.37 | 0.026 | −0.248 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Alwis, P.S.; Kundu, S.; Gietbong, F.Z.; Amin, M.H.F.; Lee, S.-R.; Kim, H.-W.; Kim, A.R. Mitochondriomics of Clarias Fishes (Siluriformes: Clariidae) with a New Assembly of Clarias camerunensis: Insights into the Genetic Characterization and Diversification. Life 2023, 13, 482. https://doi.org/10.3390/life13020482
De Alwis PS, Kundu S, Gietbong FZ, Amin MHF, Lee S-R, Kim H-W, Kim AR. Mitochondriomics of Clarias Fishes (Siluriformes: Clariidae) with a New Assembly of Clarias camerunensis: Insights into the Genetic Characterization and Diversification. Life. 2023; 13(2):482. https://doi.org/10.3390/life13020482
Chicago/Turabian StyleDe Alwis, Piyumi S., Shantanu Kundu, Fantong Zealous Gietbong, Muhammad Hilman Fu’adil Amin, Soo-Rin Lee, Hyun-Woo Kim, and Ah Ran Kim. 2023. "Mitochondriomics of Clarias Fishes (Siluriformes: Clariidae) with a New Assembly of Clarias camerunensis: Insights into the Genetic Characterization and Diversification" Life 13, no. 2: 482. https://doi.org/10.3390/life13020482