
Immunomodulation, Bioavailability and Safety of Bacteriocins

1
Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences, 117997 Moscow, Russia
2
Medical Institute, Peoples’ Friendship University of Russia (RUDN University) of the Ministry of Science and Higher Education of the Russian Federation, 117198 Moscow, Russia
Life 2023, 13(7), 1521; https://doi.org/10.3390/life13071521
Submission received: 29 May 2023
/
Revised: 1 July 2023
/
Accepted: 5 July 2023
/
Published: 7 July 2023
(This article belongs to the Special Issue Bacteriocin, Antimicrobial Peptide, and Antibiotics: Current Insights and Application)
Abstract
:The rise of antibiotic-resistant bacteria and the emergence of new pathogens have created a need for new strategies to fight against infectious diseases. One promising approach is the use of antimicrobial peptides produced by a certain species of bacteria, known as bacteriocins, which are active against other strains of the same or related species. Bacteriocins can help in the treatment and prevention of infectious diseases. Moreover, bacteriocins can be obtained in prokaryotic organisms, and contribute s to their widespread use. While the use of bacteriocins is currently limited to the food industry (for example, nisin is used as a preservative, E234), a large number of studies on their microbicidal properties suggest that their use in medicine may increase in the foreseeable future. However, for the successful use of bacteriocins in medicine, it is necessary to understand their effect on the immune system, especially in cases where immunity is weakened due to infectious processes, oncological, allergic, or autoimmune diseases. Studies on the immuno-modulatory activity of bacteriocins in animal models and human cells have revealed their ability to induce both pro-inflammatory and anti-inflammatory factors involved in the implementation of innate immunity. The influence of bacteriocins on acquired immunity is revealed by an increase in the number of T-lymphocytes with a simultaneous decrease in B-lymphocyte levels, which makes them attractive substances for reducing inflammation. The widespread use of bacteriocins in the food industry, their low toxicity, and their broad and narrow specificity are reasons for researchers to pay attention to their immunomodulatory properties and explore their medical applications. Inflammation regulation by bacteriocins can be used in the treatment of various pathologies. The aim of the review was to analyze scientific publications on the immunomodulatory activity, bioavailability, and safety of bacteriocins in order to use the data obtained to organize preclinical and clinical studies.
1. Introduction
Antibiotic resistance in bacteria is a global threat to healthcare systems. According to predictive statistical models, there were 4.95 million deaths associated with bacterial AMR in 2019, including 1.27 million deaths attributable to bacterial AMR [1]. Sub-Saharan Africa bore the highest burden of deaths, with 23.5 deaths per 100,000 population [2]. Deaths from hospital infections are increasing, with 42% of Escherichia coli, 35% of Staphylococcus aureus, and 8% of Klebsiella isolates in hospitals being resistant to antibiotics. Moreover, more than 20% of E. coli isolates, the most common causative agent of urinary tract infections, are resistant to both first-line drugs (ampicillin and co-trimoxazole) and second-line drugs (fluoroquinolones) [3,4]. Currently, over three million Americans suffer from infections caused by antibiotic-resistant strains [2]. If left unaddressed, deaths from antibiotic-resistant strains may become a leading cause by 2050, surpassing cancer and accounting for up to 10 million people per year [5].
The uncontrolled use of antibiotics in agriculture and medicine leads to the emergence of antibiotic-resistant bacterial pathogens in wastewater [6,7,8]. Residues of antibiotics used in agriculture exert selective pressure on bacterial communities in the soil–plant system, contributing to the emergence and spread of antibiotic resistance genes (ARGs) through horizontal gene transfer [9,10]. The spread of antibiotic resistance is also mediated by vesicles in the outer membrane of Gram-negative bacteria [11], conjugation between plasmids, transduction by bacteriophages, and natural transformation of extracellular DNA, which allows genetic material to move between strains and species [12].
To reduce the spread and emergence of new antibiotic-resistant strains, various approaches are being used to limit the use of antibiotics in agriculture [13,14] and medicine [15,16]. New control strategies are being developed, such as vaccines, antibodies [17,18], pattern recognition receptor ligands [19,20,21,22,23], probiotics, plant extracts [24,25], bacteriophages [26], cytokines [27], phytochemicals, silver and chitosan nanoparticles [28], antimicrobial enzymes, and peptides [29].
The use of bacteriocins, antimicrobial substances produced by bacteria, is one of the strategies to reduce extension of the antibiotic-resistant strains.
2. Resources, Gene Organization and Biosynthesis of Bacteriocins
Bacteriocins are peptide or protein antibiotics produced by bacteria that act only on strains of the same or closely related species [30]. Their history dates back to 1925, when André Gratia observed the inhibition of Escherichia coli ϕ by E. coli V [31], and Pierre Frédéricq [32] carefully described and introduced the concept of “colicin” for bacteriocins produced by E. coli [33]. Bacteriocins are now defined as peptides produced by bacteria, synthesized on ribosomes, which are active against other bacteria and against which the producer has a specific immunity mechanism [34]. Bacteriocins have several advantages, including low toxicity, a specific mechanism of action, and stability at low pH, high temperature, and over a wide range of salt concentrations [35,36]. They usually consist of 20–60 amino acids, with a positive charge and hydrophobic properties [37]. Another important trait of bacteriocins is their either linear or circular form. This has impacts on stability and activity. Bacteriocins can be degraded by proteases and digestive enzymes, limiting their oral use [38]. To preserve their activity, various delivery methods are being developed, such as liposomes [39], encapsulation [40], incorporation into coated tablets [41], starch-based matrices [42], and chemical modification of the C-terminus to protect against proteolytic enzymes [43,44].
Bacteriocins are produced by Gram-positive and Gram-negative bacteria, as well as archaea [45,46,47]. Some bacteria can produce several different bacteriocins [48,49,50,51,52] that are active against many pathogenic bacteria [53,54].
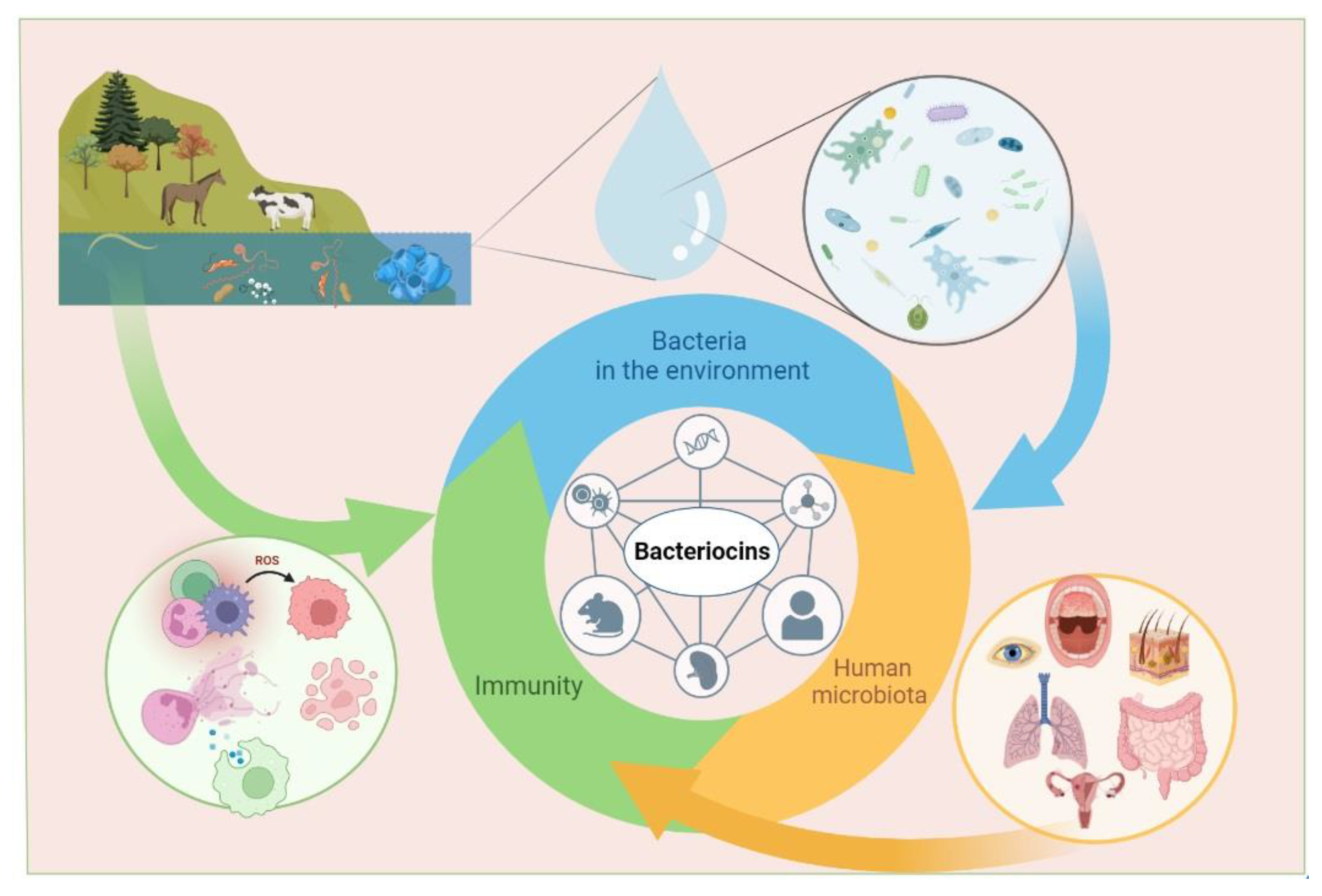
Bacteria that produce bacteriocins can be found everywhere: in water, soil, on the skin and mucous membranes of humans and animals, as well as in food and plants (Figure 1). The marine environment is considered to be one of the richest resources for the production of marine microbial bacteriocins [55]. Initially, researchers focused on studying sources such as food and animals [56,57]. In recent years, the bacteria Lactobacillus, Enterococcus, Streptococcus, and Staphylococcus, which inhabit the mucous membranes of the human oral cavity and gastrointestinal tract, have been extensively studied as potential sources of bioactive compounds, including bacteriocins [56,58].
Lactic acid bacteria (LAB) are known to produce various non-toxic bacteriocins that are pH stable and have a broad spectrum of antimicrobial activity. Bacteriocins of lactic acid bacteria do not adversely affect the intestinal microbiota as they are sensitive to the host’s digestive proteases, chymotrypsin, and trypsin [38,59]. The most common producers of bacteriocins in raw milk are Brevibacillus brevis and Bifidobacterium lactis; in yogurt and fermented soy products, L. acidophilus and L. plantarum are common, and in cheese and goat milk, L. plantarum is prevalent [60,61].
Several hundred bacteriocins have been described to date [62]. It is believed that 99% of all bacteria can produce bacteriocins, and many of them can produce more than one type, most of which are still unknown [59].
Bacteriocin genes can be located on both chromosomes and in a plasmid [63,64], with most colicin genes being found on plasmids [65]. These genes are typically arranged in clusters and contain one or more immunity proteins to prevent self-killing, and regulatory proteins provided post-translational modifications [66]. For instance, the CEA colicin gene is located on a plasmid; normally, its activity is suppressed by the LexA protein [67]. Subsequently, a high level of colicin causes the death of the producer cell. Interestingly, mutant bacteria without the LexA protein remain viable with an increase in colicin synthesis [67]. Cells that produce bacteriocins have multiple strategies to protect themselves from the lytic activity of their own bacteriocins. These strategies include the presence of specialized proteins or a system of pumps that remove bacteriocins from the cell, and sometimes a combination of these methods [68,69].
Bacteriocins are synthesized most often in the form of a precursor with further modification of the N-terminus [70]. However, circular, leaderless bacteriocins and colicins have no precursors [71,72]. Leaderless bacteriocins are active right after translation as they do not undergo any post-translational processing common to other groups of bacteriocins. Such leaderless bacteriocins may be attractive agents for medical use as their production becomes more accessible and cheaper [71].
Several databases have been developed to organize information regarding the structure, physicochemical properties, and mechanism of action of bacteriocins. One such database is BADASS, a user-friendly software package with a graphical interface that facilitates searching and analysis of bacteriocin diversity in full metagenomic shotgun sequencing data [73]. Another open-access database, BACTIBASE, provides detailed structural and functional information on 230 bacteriocins, including antimicrobial, physicochemical, and structural properties [74]. Other databases focus on specific pathologies and the regulation of immunocompetent cells by various substances, including bacteriocins, and visualize cause-and-effect relationships [75,76].
For instance, using a method of a random multiscale convolutional neural network, researchers found that potential bacteriocins could be HNH-endonuclease sequences from various marine bacteria. The Random Multiscale Convolutional Neural Network method, proposed by Cui Z. et al., establishes a random model for updating the scale value, and it offers a new classification system that predicts potential bacteriocin relationships [55]. Finally, state-of-the-art methods for predicting the efficacy of antimicrobial peptides, including bacteriocins, are being developed using deep learning (DL) approaches to optimize the search for new effective compounds [77].
3. Mechanism of Action on Microorganisms
Bacteriocins act in various ways, depending on their structural and physicochemical properties, as well as post-translational modifications. Most bacteriocins are cationic and interact with negatively charged bacterial membranes due to electrostatic forces, similarly to cationic antimicrobial peptides in multicellular organisms [78]. Electrostatic interactions occur between the negatively charged teichoic acids and phospholipids of Gram-positive bacteria, as well as with lipopolysaccharides of Gram-negative bacteria, and with Lipid II, which is a precursor molecule in the synthesis of the cell wall of both Gram-positive and Gram-negative bacteria [79]. This interaction represents the first stage in the manifestation of nonspecific and specific activity [34]. In the next stage, pores, ion-permeable channels, and cell death may occur [33], also due to the release of autolytic enzymes associated with lipoteichoic acids [80].
Another way for bacteriocins to enter the bacterial cell is by penetrating through a variety of transporters, which provide the necessary nutrients that ensure the vital activity of bacteria [81]. Some bacteriocins of Gram-positive bacteria, such as nisin, garvicin ML, lactococcin G, and sublancin 168, use zinc-dependent metallopeptidases, maltose ABC transporters, and undecaprenyl pyrophosphate phosphatase to enter the cell [81,82]. Gram-negative bacteria colicins use lipopolysaccharide (LPS) and transmembrane proteins in the outer membrane as receptors, allowing them to penetrate the cell through the two lipid membranes of the cell membrane. These transmembrane proteins are involved in the import of nutrients and metal ions, such as vitamins, sugars, and Fe3+. After penetrating the perioplasm, colicins interact with a translocator protein that helps establish an interaction with proteins called Tol or Ton in the host periplasm, followed by entry into the cell cytoplasm [83].
Interestingly, colicin-associated single-stranded DNA can also be transported into the bacterial periplasm. The discovered transport pathways of colicins can be utilized by bacteria to transport large molecules [84].
Studies have shown that bacteriocins from both Gram-positive and Gram-negative bacteria can use phosphotransferase sugar transport systems to enter the cell [34,85]. Once inside the bacterial cell, bacteriocins can cause the degradation of DNA and RNA, inhibit replication, transcription, and protein synthesis by specifically cleaving 16s rRNA, and stop the synthesis of the bacterial cell wall [86]. For instance, microcin C, microcin J25, and microcin B17 bind and inhibit Asp-tRNA synthetase, RNA polymerase, and DNA gyrase, respectively, which have been identified as their direct targets [45]. Ruminococcin C, isolated from the human microbiota bacterium Ruminococcus gnavus, interferes with the synthesis of nucleic acids without disrupting the membranes of target bacteria [87].
Despite the various possible mechanisms of antimicrobial strategies, the most common is pore formation in the target cell membrane [86]. Bacteriocins exhibit a bacteriostatic or bactericidal spectrum of action, mainly directed against bacteria closely related to the producing strain [88] and, in rare cases, against a wide range of bacteria [33]. It is worth noting that some bacteriocins use multiple types of mechanisms to influence the bacterial cell, significantly complicating bacterial resistance and the emergence of resistance.
Resistance to bacteriocins can be natural or acquired, appearing in previously susceptible strains [89,90]. Up to 8% of wild-type Listeria monocytogenes strains have been found to be naturally resistant to pediocin-like bacteriocins, particularly pediocin PA-1, while remaining sensitive to nisin [91,92]. Natural and acquired resistance to bacteriocins can emerge and disappear as a result of mutations in genes responsible for susceptibility [68]. For strains that do not produce bacteriocins, a common defense strategy is to synthesize extracellular proteases and peptidases [93]. For instance, Enterococcus faecalis secretes gelatinase to inactivate pediocin-like bacteriocins [94].
Acquired resistance can be developed through various strategies, such as increasing hydrophobicity to reduce membrane permeability, synthesizing the bacterial cell wall or cytoplasmic membrane [33,68]. For instance, L. monocytogenes can develop resistance to nisin by altering the composition of fatty acids that enter the membrane [95], while Clostridium difficile undergoes cell wall reorganization and changes in central cellular processes, such as carbohydrate metabolism, as well as an increase in the number of flagella per cell, without any change in cell mobility [96].
Moreover, certain bacterial strains can acquire multiple modes of resistance simultaneously, resulting in a shared resistance phenotype [97]. Understanding the mechanisms underlying the development of protection against bacteriocins is crucial for devising new strategies for the clinical use of bacteriocins, while also taking into account the potential adverse effects of resulting resistant strains.
4. Immunomodulatory Activity of Bacteriocins
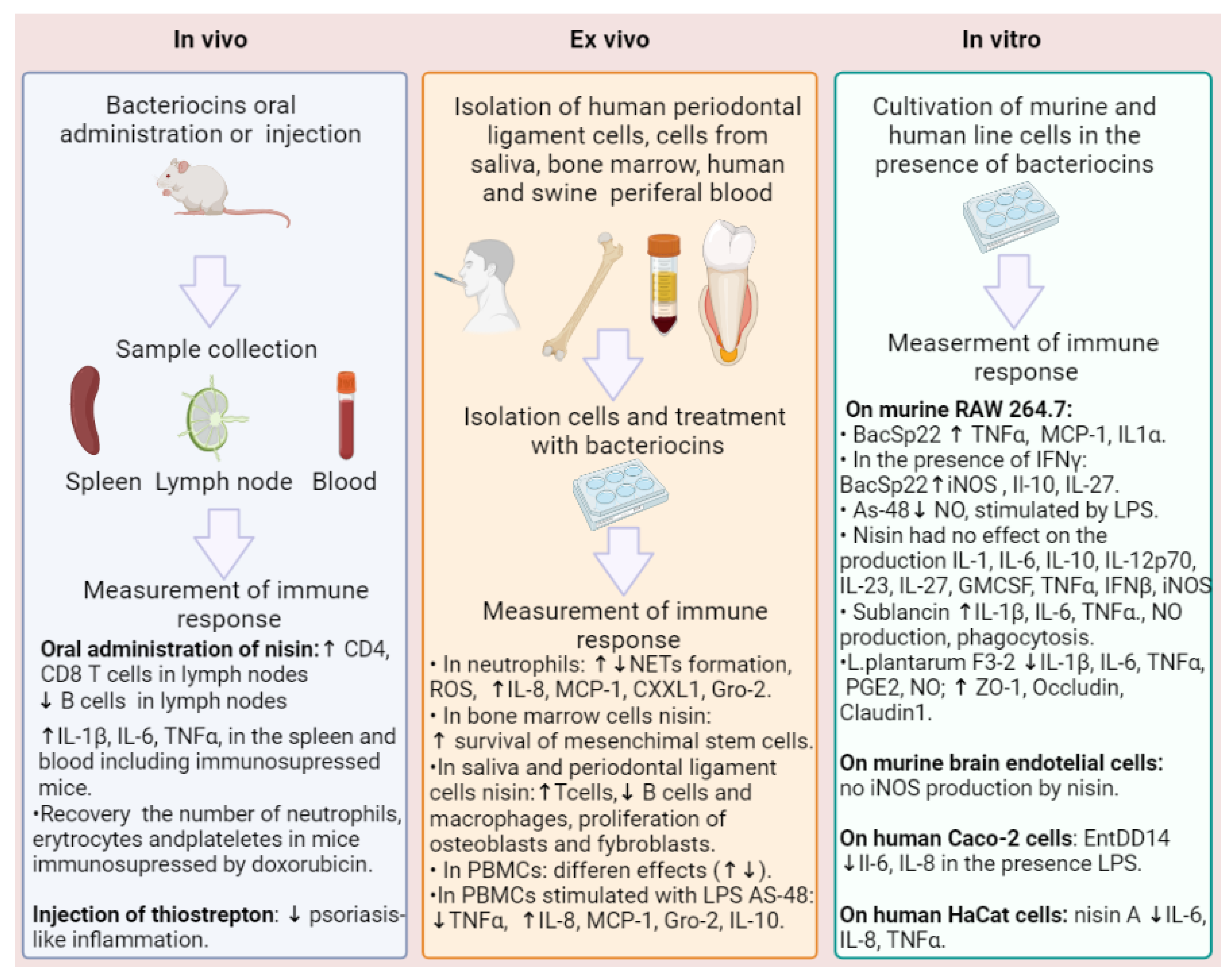
Numerous studies have investigated the bactericidal, bacteriostatic, antiviral, antiparasitic, and antitumor properties of bacteriocins [33,38]. However, research on the immunomodulatory activity of bacteriocins is represented by few studies, making it difficult to characterize their overall immunomodulatory and allergenic effects. Nevertheless, these studies suggest that the biological activity of bacteriocins may be similar to that of human antimicrobial peptides, based on comparable physicochemical properties [98]. For instance, the lanthiobiotic nisin Z has been found to induce the secretion of chemokines MCP-1, Gro-α, and IL-6 in human peripheral blood mononuclear cells (PBMCs) in a dose-dependent manner (concentrations of nisin Z 50, 100, 150 µg/mL), wherein nisin Z significantly reduces TNF-α induction in response to bacterial LPS (concentration of 2 ng/mL) PBMCs. The inhibition of nisin Z-mediated IL-6 secretion was effected by the inhibition of mitogen-activated protein kinase (MAPK) signaling and specific receptor tyrosine kinases. The inhibition of p38 (SB-202190), MAPK kinase (MEK) (U-0126), components of the MAPK: extracellular signal-regulated kinases 1 and 2 (ERK1/2) pathway (c-Raf) ZM 336372, and the Src family kinases (PP1 and PP2) all significantly inhibited IL-6 induction by nisin Z. This suggested a strong correlation between ERK/MAPK signaling and the induction of immunomodulatory responses by nisin Z [99]. Furthermore, nisin Z has been shown to provide protection against both Gram-positive microorganisms S. aureus and Gram-negative Salmonella enterica sv. Typhimurium and Escherichia coli in mouse infection models. The authors of the study suggest that nisin Z modulates host immunity through mechanisms similar to those of human natural host defense peptides, involving multiple signal transduction pathways and growth factor receptors [99].
An increase in the levels of CD4 and CD8 T-lymphocytes was observed in mice with short-term dietary intake of nisin, along with a simultaneous decrease in B-lymphocyte levels. After long-term dieting, the number of T-cells returned to the control level. The period of administration of nisin was either 30, 75, or 100 days [100]. Nisin’s ability to lower the number of B-cells may prove useful in the treatment of periodontal and peri-implantitis [101,102,103], as increased amounts of T-cells are found in inflammatory infiltrate in peri-implant soft tissues [104]. Additionally, nisin was shown to disrupt oral pathogenic biofilms and restore microbiome composition towards healthy control levels in a peri-implantitis setting [101,102,103]. The authors conclude that nisin is a perspective agent in the treatment of periodontal and peri-implantitis inflammation [101,102,103].
Nisin is effective when used topically on mucous membranes due to its ability to restore the structure of the endometrium in a rat model and normalize the number of neutrophils to control levels, improve levels of IFN-γ, IL-2, IL-8, and inhibit the formation of S. aureus biofilms [103,104]. Additionally, nisin has been found to significantly reduce not only the levels of several periodontal pathogens but also bone loss and the oral and systemic inflammatory response of the host. At the same time, nisin increased the population of fibroblasts and osteoblasts and mediated the proliferation of human periodontal ligament cells in a dose-dependent manner by increasing the proliferation marker Ki-67 [105].
Nisin was found to significantly increase the survival of mesenchymal stem cells (MSCs) from human bone marrow in vitro [106]. MSCs are used for transplantation, but their low survival rate after transplantation is a significant disadvantage. The survival and anti-inflammatory effects of nisin were assessed by cultivating MSCs against the background of exposure to H2O2 or in a serum-free medium using MTT analysis, ELISA, and real-time PCR. It was discovered that 250 and 500 IU/mL of nisin had a significant anti-apoptotic effect on MSCs, increasing cell viability and proliferation. The expression of IL-10, fibroblast growth factor 2 (FGF-2) and transforming growth factor-β (TGF-β) genes, as well as the synthesis of TGF-β and FGF-2 proteins, increased, indicating that nisin can have anti-inflammatory effects. During wound healing therapy with mesenchymal stem cells (MSC), an important limitation is that the MSCs are sensitive and short-lived in stress conditions. Preconditioning is effective to increase cellular resistance and survival, and nisin is a good choice as nisin improves the stability of MSCs. Long-lived MSCs produce more anti-inflammatory and less inflammatory cytokines and growth factors, which help cell repair and differentiation into fibroblasts at the site of tissue damage [106]. Nisin showed opposite pro-inflammatory effects on unstimulated and stimulated porcine PBMCs [107]. Nisin in a concentration of 50 µg/mL exhibited the proliferative activity, increasing the production of IL-1β and IL-6 and increasing the percentage of CD4+ CD8+ T in porcine PBMCs. After cell stimulation with E. coli, nisin showed antiproliferative activity, decreased phagocytosis, and inhibited the synthesis of IL-6 [107].
The zoonotic pathogen Staphylococcus pseudintermedius 222, able to infect humans, produces the unmodified form of the bacteriocidin BacSp222, as well as two post-translationally modified forms via succinylation and the cleavage of formylmethionine [108]. The production of such modified forms occurs in response to environmental changes, protects the cells of the producing bacteria from auto-toxicity of the secreted bacteriocin and limits the pathogenicity of the strain [109]. A study investigated the effects of these three bacteriocins and nisin A, used as a reference, on murine monocyte-macrophage-like Il- and murine brain endothelial (MBE) cell lines, as well as human polymorphonuclear neutrophils (hPMN) [108]. The results demonstrated that all tested BacSp222 compounds increased NO production and the expression of inducible nitric oxide synthase (iNOS) in combination with IFN-gamma in monocyte/macrophage-like cell lines P388.D1 and RAW 264.7, and they do not potentiate NO production by endothelial cells. Furthermore, all natural forms of BacSp222, either alone or with IFN-gamma, stimulated the production of TNF-α, MCP-1, and IL-1α. When combined with IFN-γ, the levels of IL-10 and IL-27 were increased. This studies on murine monocyte-macrophage cells revealed that bacteriocin BacSp222 and its forms activated the NF-κB transcription factor, leading to the increased expression of proteins associated with inflammation, such as iNOS and TNFα. In contrast, the IFN-β production was increased after the exposure of the cells to all forms of bacteriocin in the presence of IFN-γ. However, nisin A did not cause any changes in the production of the cytokines studied (IL-1α, IL-1β, IL-6, IL-10, IL-12p70, IL-17A, IL-23, IL-27, GMCSF, IFN-β, TNF-α) and did not influence the NO production and iNOS expression regardless of the presence or absence of IFN-γ on murine monocyte-macrophage-like cell lines. In human neutrophils, all forms of BacSp222 bacteriocin upregulated IL-8, but they did not induce ROS production or the formation of neutrophilic extracellular networks. In contrast, nisin A did not stimulate IL-8 production by human PMNs. Nisin A (1 μM nisin A for 4 h) also did not induce ROS production or the formation of neutrophilic extracellular networks. BacSp222 enhanced the expression of the iNOS only in the macrophages but not in the endothelial cells. Notably, in all experiments, the deformylated bacteriocin exhibited lower activity compared to the other forms of the peptide. BacSp222 and its succinylated form could be recognized as a novel peptide inducer of NO production by immune cells [108]. Another bacteriocin, AS-48, produced by different strains of Enterococcus, decreased nitric oxide (NO) production induced by LPS (up to 13.5 µM) (>96.51 µg/mL) on RAW macrophages and demonstrated an absence of pro-inflammatory effects [110].
Interestingly, the effects of nisin A on the activation of neutrophils in the experiment of other researchers revealed the ability to form neutrophil extracellular traps (NETs) [111]. Nisin A’s ability to activate neutrophils (concentrations of 75 and 150 µM) was demonstrated using scanning electron microscopy and fluorescence microscopy to observe the formation of NETs, well known for their ability to neutralize virulence factors and destroy bacterial pathogens. In addition, the presence of nisin A increased the intracellular level of neutrophil superoxide, which is normally produced by activated NADPH oxidase and is a prerequisite for the formation of NETs (Table 1) [111].
Thus, a comparison of the data obtained by Śmiałek J. et al. and Begde D. et al. revealed that nisin A in concentrations of 1 µM did not have an influence on the NETs formation, while in concentrations of 75 µM and 150 µM, the formation of NETs was observed [108,111].
In a study examining the immunomodulatory activity of sublancin, isolated from Bacillus subtilis 168, an increase in CXCL1 and MCP-1 chemokine levels and a decrease in TNF-alpha production were observed in murine macrophage cells and neutrophils [112]. However, sublancin increased production of IL-1β, IL-6, TNF-alpha, nitric oxide, phagocytic, and microbicidal activity against MRSA in murine peritoneal macrophages and RAW264.7 cells (Figure 2) [113]. Oral administration of sublancin (1.0 mg/kg body weight) led to an increase in the expression of IL-1β, IL-6, and TNF-α mRNA in the spleen of BALB/c mice, including immunosuppressed mice treated with cyclophosphamide, and accelerated the recovery of peripheral leukocytes, erythrocytes, hemoglobin, and platelets while increasing the phagocytic activity of macrophages that had decreased after cyclophosphamide treatment. Together, these findings suggest that sublancin plays a crucial role in protecting against immunosuppression in mice treated with cyclophosphamide and may be a potential candidate for use in immunotherapy [114]. The study revealed that macrophage activation by sublancin is to some extent carried out through TLR4 with the participation of NF-κB and MAPK. When administered orally to mice, sublancin increased CD4 and CD8 T-lymphocytes in the mesenteric lymph nodes, indicating that it is capable of exerting an immunomodulatory effect by activating macrophage and T-cell immunity [113].
When studying bacteriocin-producing strains of Lactiplantibacillus plantarum, researchers found that L. plantarum YRL45 significantly reduced the elevated levels of IL1β, IL6, TNFα, nitric oxide, and prostaglandin E2 induced by LPS in RAW264.7 cells, thereby reducing the severity of the inflammatory process [115]. Additionally, L. plantarum F3-2 significantly increased the expression levels of ZO-1, Occludin, and Claudin 1, all of which are involved in the formation of intercellular contacts in damaged cells of the intestinal epithelium, which is necessary to maintain the integrity of the epithelium [115].
On the other hand, acidocin A, produced by Lactobacillus acidophilus TK9201, was found to induce the production of a number of inflammatory mediators (IL-6, TNFα, MIG/CXCL9, MCP-1/CCL2, MCP-3/CCL7, and MIP-1β) in primary human monocytes, while also inhibiting the production of some anti-inflammatory factors such as IL-1RA and MDC/CCL22. This demonstrates its pronounced pro-inflammatory properties [116].
The enterocin DD14 (Ent DD14), which is produced by Enterococcus faecalis 14 strain isolated from newborn’s meconium, exerts the anti-inflammatory effect on the secretion of pro-inflammatory interleukins, including IL-6 and IL-8. The results show that EntDD14 is able to significantly decrease the secretion of both interleukins on Caco-2 cells following their treatments with LPS [117].
Thiostrepton is a ribosomally synthesized and post-translationally modified peptide (RiPP) produced by bacteria of Streptomyces genus [118,119]. Thiostrepton was reported to exhibit activity against Gram-positive bacteria and against various human cancer cells [120,121,122]. This bacteriocin has been characterized as a potent chemical inhibitor of the oncogenic transcription factor FoxM1, frequently overexpressed in cancers or other diseases [123,124,125]. The mechanism of action was defined through upregulation of heat shock proteins HspA1A, Hsp70, Hsp90α, or Hsp105 and triggering apoptosis in human cancer cells [126]. Thiostrepton inhibited TLR7-9 activation in mouse dendritic cells and did not inhibit NF-κB activation induced by TNF-α, IL-1, and other TLRs, as it inhibits TLR9 localization in endosomes via proteasome inhibition and via endosomal acidification. Moreover, in different murine models, thiostrepton attenuated LL37- and imiquimod-induced psoriasis-like inflammation. The researchers made the conclusion that thiostrepton is a novel TLR7-9 inhibitor, suggesting the potential therapeutic applications of thiostrepton on immunologic disorders elicited by inappropriate activation of TLR7-9 [127].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Immunomodulation effects of bacteriocins.
| Bacteriocin | Resource | Highlights | Ref. |
|---|---|---|---|
| Nisin Z | L. lactis | Inducing MCP-1, Gro-α, and IL-6 in human PBMC. In response to LPS, reducing TNF-α and inhibiting IL-6. | [99] |
| Nisin | L. lactis | Increasing T-cells, fibroblasts, and osteoblasts; increasing B cells. Increasing the expression of IL-10, FGF-2, and TGF-β genes and the synthesis of TGF-β and FGF-2. Decreasing phagocytosis and inhibiting the synthesis of IL-6. | [100,101,102,105,106,107] |
| Nisin A | L. lactis | No influence on IL-1α, IL-1β, IL-6, IL-10, IL-12p70, IL-17A, IL-23, IL-27, GMCSF, IFN-β, TNF-α, iNOS, NO production, and NETs. | [108] |
| Inducing NETs formation and ROX in human neutrophils. | [111] | ||
| BacSp222 | Staphylococcus pseudintermedius 222 | Increasing NO, iNOS in P388.D1, and RAW 264.7; increasing IL-10 and IL-27 in combination with IFN-γ. In human neutrophils, upregulating IL-8; absence of ROS production or NETs formation. | [108] |
| AS-48 | Enterococcus spp. | Decreasing NO production induced by LPS on RAW cells; absence of pro-inflammatory effects. | [110] |
| Sublancin | Bacillus subtilis 168 | Increasing CXCL1 and MCP-1; decreasing TNF-α in murine peritoneal macrophages and neutrophils. Increasing IL-1β, IL-6, TNF-alpha, nitric oxide, and phagocytosis. Increasing T-cells in the mesenteric lymph nodes. Increasing IL-1β, IL-6, and TNF-α in the spleen of immunosuppressed BALB/c mice. | [112,113,114] |
| L. plantarum F3-2 | Lactiplantibacillus plantarum | Reducing elevated levels of IL1β, IL6, TNFα, nitric oxide, and prostaglandin E2 induced by LPS in RAW264.7 cells. Increasing the expression levels of ZO-1, Occludin, and Claudin 1. | [115] |
| Acidocin A | Lactobacillus acidophilus TK9201 | Increasing IL-6, TNFα, MIG/CXCL9, MCP-1/CCL2, MCP-3/CCL7, and MIP-1β in PBMCs. | [116] |
| Ent DD14 | Enterococcus faecalis 14 | Decreasing IL-6 and IL-8. | [117] |
| Thiostrepton | Streptomyces genus | Increasing HspA1A, Hsp70, Hsp90α, or Hsp105. | [127] |
In addition to in vitro and in vivo studies, bacteriocins’ immunomodulatory activity is also being investigated through comparative genomic analysis. A study of genes from four Lactobacillus strains revealed that L. plantarum SK151 had the highest number of genes with potential immunomodulatory activity—approximately 74. In L. johnsonii PF01, 41 genes were identified that covered both immune activation and immunosuppression, compared to L. mucosae LM1 and L. fermentum SK152, which may be more effective in activating immune cells and the pro-inflammatory cascade than in suppressing it. Based on the similarities and differences between the four Lactobacillus species, the authors conclude that each strain’s immunomodulatory function should be experimentally studied and confirmed, since some genes’ activity may be strain specific and not identifiable through comparative genomics alone [128].
The immunomodulatory activity of bacteriocins is determined by their structural features, and the presence of N-formyl methionine contributes to the manifestation of pro-inflammatory activity. At the same time, the presence of N-formyl methionine is not essential for microbicide activity.
The effect of bacteriocins on immunocompetent cells is directly dependent on concentration. Using the example of bacteriocin nisin, a change in the activity profile from neutral to pro-inflammatory with increasing concentration was shown.
The variety of effects of bacteriocins on immune cells depends not only on the structure and concentration, but also on the context of their application. For instance, bacteriocin nisin in the absence of additional stimuli did not affect IL-8 and TNFα production, while in the presence of LPS, TNFα decreased and IL-8 increased. At the same time, the combined effect of nisin and IFNγ did not affect TNFα production.
Most bacteriocins, including nisin, lose their activity under the action of stomach and intestinal enzymes. However, it turned out that oral administration of bacteriocin nisin had an effect on the ratio of T- and B-cells in the spleen and lymph nodes of mice. The explanation could be that bacteriocin began to act already in the oral cavity, affecting immune organs.
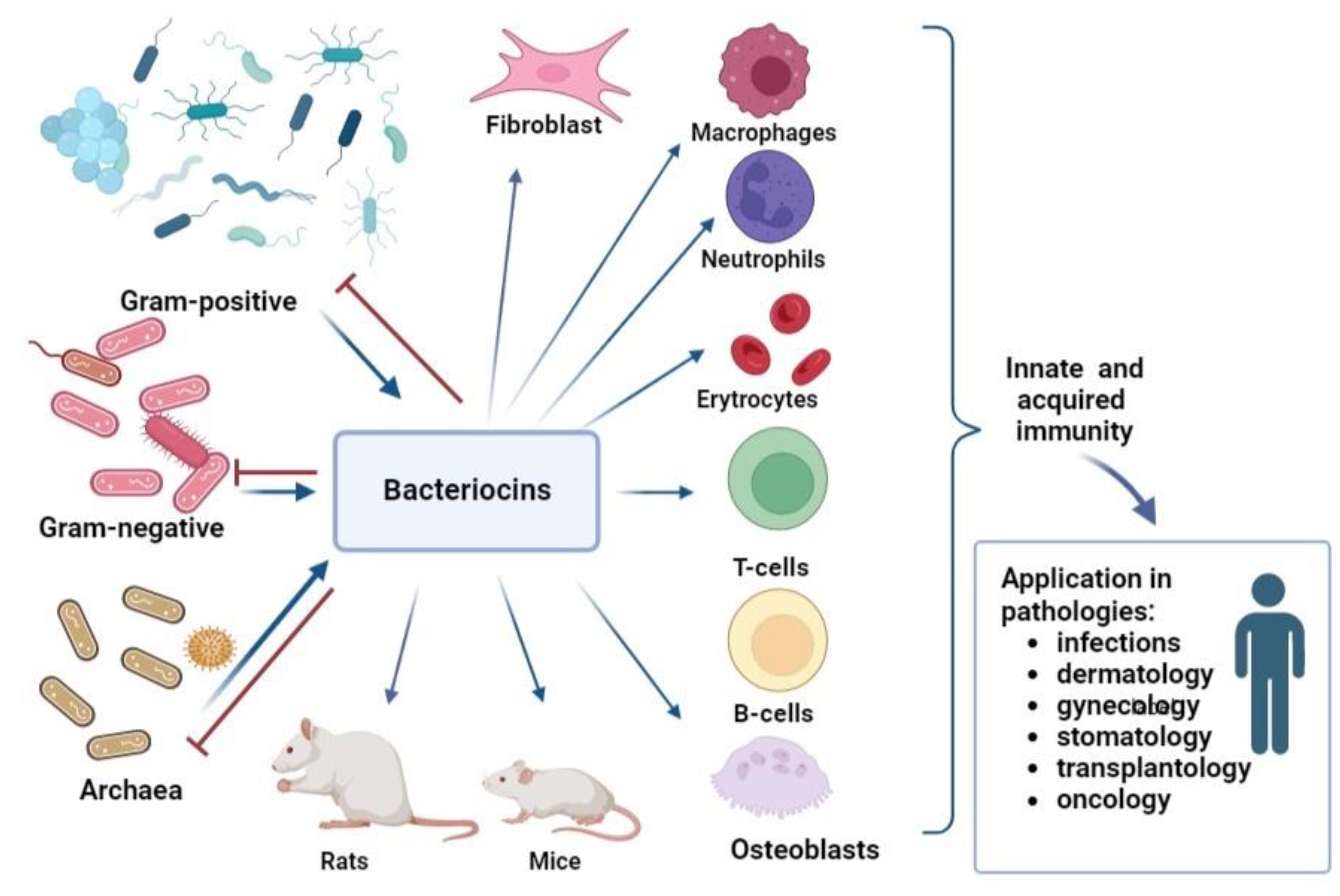
Thus, the analysis of immunomodulatory ability showed that bacteriocins have an influence on innate immunity. For instance, some of bacteriocins can induce the production of reactive oxygen species, nitric oxide, phagocytosis, and NETs formation; others do not affect and even inhibit inflammation. The ability of bacteriocins to increase the number of T-lymphocytes, simultaneously decreasing B-lymphocytes and inducing cytokine and chemokine production, makes it possible to modulate acquired immunity too (Figure 3). Moreover, it is obvious that bacteriocins can regulate inflammation, inducing pro-inflammatory factors during infection or immunosuppression and suppressing pro-inflammatory cytokines in conditions of excessive inflammation.
An important factor that should be taken into account when using bacteriocins as antimicrobial agents is the release of the contents of bacterial cells during their destruction. A large number of biologically active compounds will affect immunocompetent cells, including through innate immunity receptors. The activation of innate immune receptors triggers intracellular pathways and alters immune response.
5. Bioavailability and Safety of Bacteriocins
Since the discovery of bacteriocins, researchers have mainly focused on determining their antimicrobial activity. However, bacteriocins can affect not only microbial communities but also eukaryotic cells. To use bacteriocins clinically as antimicrobial drugs, it is necessary to study their clinical efficacy, bioavailability and safety. To prevent side effects, it is also necessary to understand the mechanisms of action on tissues and organs when applied topically, as well as on immunocompetent cells when applied systemically.
To assess oral bioavailability, it is important to evaluate trans-epithelial transport and the effect of proteolytic enzymes in the gastrointestinal tract on maintaining the biological activity of bacteriocins. Nisin is a standard of comparison in the analysis of other bacteriocins due to its well-studied mechanism of action [129].
To analyze the possibility of migration of bacteriocins through epithelial cells of the gastrointestinal tract and vascular endothelial cells, the transport of fluorescently labeled bacteriocins—nisin, plantaricin 423, and bacST4SA—through colon adenocarcinoma cells (Caco-2) and human umbilical vein endothelial cells (HUVEC) was determined in vitro. It was found that after 3 h, 75% of nisin, 85% of plantaricin 423, and 82% of bacST4SA had migrated through the monolayer of Caco-2 cells. Over the same time period, 88% of nisin, 93% of plantaricin 423, and 91% of bacST4SA migrated through the HUVEC monolayer. It should be noted that the viability of both cell types remained unchanged when exposed to 50 μM of nisin, plantaricin 423, or bacST4SA. However, the effect of human plasma on the activity of bacteriocins depended on the structure and concentration of bacteriocins, and among the tested compounds, nisin was less stable. This study is one of the first to provide evidence that bacteriocins can cross the intestinal–hematological barrier [130].
When administered orally, it is crucial to evaluate the preservation of bacteriocin activity against gastrointestinal enzymes. Research conducted on laboratory and farm animals has shown that proteolytic enzymes in the stomach and small intestine, such as pepsin, trypsin, and chymotrypsin, cleave and inactivate many bacteriocins [131,132,133]. Bacteriocins without post-translational modifications are particularly sensitive to intestinal proteases, resulting in decreased antimicrobial activity when taken orally. An in vitro dynamic model examining the biostability of pediocin PA-1 under upper gastrointestinal conditions found that pediocin remained stable in the stomach but degraded completely when exposed to conditions equivalent to those found in the small intestine [134].
Bacteriocins with post-translational modifications are generally more resistant to proteases [135,136], but intestinal proteases can still inactivate and digest nisin A [137,138]. Initial research on the stability of microcin J25 by Pomares et al. (2009) showed that microcin J25 was resistant to digestion by proteolytic enzymes in the stomach and intestinal contents [139]. However, subsequent research conducted by Naimi et al. (2018) investigated the degradation of microcin J25 using both dynamic and static digestion models associated with antibacterial assays, LC-MS/MS, and molecular network analysis. The study found that while microcin J25 is remarkably stable under extreme conditions due to its lasso topology, it is degraded by the action of the pancreatic protease elastase and loses its antimicrobial activity [140]. To preserve the biological activity of bacteriocins, researchers are developing structural modifications using bioengineering methods, as well as delivery methods to the small and large intestines where bacteriocins act [41,42,44].
When bacteriocins are topically applied as part of hydrogels and nanoparticles, they can significantly increase the effectiveness of therapy in wound healing and combating infections of human and animal mucous membranes [104,141,142,143]. Bacteriocins, such as nisin, are also applied to titanium-based alloys, such as Ti6Al4V and its ultra-low interstitial version Ti6Al4V-ELI, which are commonly used for medical implants in orthopedics. Problems associated with infection, acute and chronic inflammation, osteolysis, and implant loosening and failure can be solved by applying bacteriocins to orthopedic materials [144]. Various solvents are being investigated to increase the solubility of bacteriocins. For example, an analysis of the effectiveness of nisin in promoting apoptosis of MG-63 osteosarcoma cells found that it was most effective in 0.05% acetic acid at concentrations of 800 μg/mL or higher, whereas in DMSO, and methanol at 0.05%, more than 90% of cells remained viable [145].
Safety studies have shown that bacteriocins have low toxicity and hemolytic activity. For example, the cytotoxicity of nisin is several times higher than the minimum inhibitory concentration [146,147]. In particular, an MTT-based cytotoxicity assay demonstrated nisin’s A cytotoxicity against human T-lymphoma Jurkat cells, Molt-4 cells, and freshly cultured human lymphocytes at concentrations greater than 200 µM (IC (50) 225 µM) [111].
Turicin H, which has inhibitory activity against B. cereus, Bt Cry-B, wild-type E. faecium, and E. faecium ATCC, but not Bt Cry-B/pThurH and Bt Cry-B/pThurHΔThnA, did not have hemolytic activity at a concentration of 20 µg/mL, which is four times higher than the minimum inhibitory concentration (MIC). This suggests the potentially safe use of turincin H as an antibacterial peptide for medical use [148]. While most bacteriocins are non-toxic to eukaryotic cells, enterococcal cytolysin has been found to be toxic [149]. Bacteriocin Ba49 from Bacillus subtilis subsp. spizizenii strain Ba49 present on the onion Allium cepa, showed low toxicity in three mammalian cell lines (HEK 293T, RAW 264.7, and L929) at concentrations several times higher than the MIC [150].
The bacteriocin AS-48, produced by Enterococcus faecalis and active against a number of Gram-positive bacteria, including Mycobacterium tuberculosis, did not show any cytotoxicity against macrophage cell lines THP-1, MHS, and J774.2 at concentrations close to its MIC [151]. The low cytotoxicity of AS-48, the absence of lymphocyte proliferation in vivo after skin sensitization in mice, and the lack of toxicity in a murine model support the consideration of the broad spectrum antimicrobial peptide AS-48 as a promising therapeutic agent for the control of a vast array of microbial infections, in particular, those involved in skin and soft tissue diseases [150]. Therefore, it is necessary to study the toxicity of bacteriocins to develop drugs based on them, as well as the methods and duration of application. It is also important to establish potential resistance to bacteriocins with repeated use [152].
6. Conclusions
Bacteriocins are not limited to acting on bacteria alone, as they exhibit activity against viruses, fungi, and parasites, and they also have an immunomodulatory effect on eukaryotic cells.
The limited number of scientific studies on the effects of bacteriocins on human immunocompetent cells restricts their potential use in medicine. However, the discovered properties of bacteriocins to modulate immunocompetent cells reveal their potential as immunomodulators. Furthermore, their cytotoxic activity against certain tumors presents opportunities for their use in complex therapy for oncology.
Multiple types of bacterial metabolites (quorum sensing molecules, pigments, antibiotics, etc.) have been proven to have immunomodulatory effects. The additional capabilities of bacteriocins to defeat pathogenic bacteria and concomitantly limit their inflammatory reactions provide support for applications of bacteriocins as therapeutic agents. Inflammation regulation by bacteriocins can be used in the treatment of various pathologies
Compared to other drugs, bacteriocins offer several advantages, such as high activity in the nano-molar range, low toxicity, and stability at low pH and high temperatures, as well as specific mechanisms of action. Additionally, probiotics can produce bacteriocins, which can be regulated in the gastrointestinal tract to prevent the spread of intestinal infections. This is particularly important in medical hospitals, where intestinal infections are a serious problem.
Research on both broad and narrow-spectrum bacteriocins can serve as a platform for developing complex and personalized therapies. Obtaining bacteriocins involves the use of prokaryotic systems and bioengineering methods, which significantly reduces production costs compared to eukaryotic antimicrobial peptides.
The widespread use of bacteriocins in the food industry, their low toxicity, and the presence of compounds with broad and narrow specificity make them a promising candidate for medical use. Therefore, researchers must pay attention to the immunomodulatory properties of bacteriocins to find opportunities for their medical use.
To determine the further directions of medical applications of bacteriocins, detailed studies on their effects on target cells and immunomodulatory activities on various populations of immunocompetent cells are necessary. Full-scale studies on the immunomodulatory activity of bacteriocins can expand the arsenal of drugs and optimize preventive and therapeutic strategies for precise medicine.
Funding
This paper has been supported by the RUDN University Strategic Academic Leadership Program.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.; Kering, K.; Wairimu, C.; Onsare, R.; Mbae, C. Antimicrobial Resistance Rates and Surveillance in Sub-Saharan Africa: Where Are We Now? Infect. Drug Resist. 2022, 15, 3589–3609. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Aqib, A.I.; Muzammil, I.; Majeed, N.; Bhutta, Z.A.; Kulyar, M.F.-E.; Fatima, M.; Zaheer, C.-N.F.; Muneer, A.; Murtaza, M.; et al. MRSA compendium of epidemiology, transmission, pathophysiology, treatment, and prevention within one health framework. Front. Microbiol. 2023, 13, 1067284. [Google Scholar] [CrossRef]
- WHO. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report. 2022. Available online: https://www.who.int/publications/i/item/9789240062702 (accessed on 19 February 2023).
- de Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, A.; Kurittu, P.; Al-Mustapha, A.I.; Heljanko, V.; Johansson, V.; Thakali, O.; Mishra, S.K.; Lehto, K.-M.; Lipponen, A.; Oikarinen, S.; et al. Wastewater surveillance of antibiotic-resistant bacterial pathogens: A systematic review. Front. Microbiol. 2022, 13, 977106. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Vu, H.P.; Nguyen, L.N.; Wang, Q.; Djordjevic, S.P.; Donner, E.; Yin, H.; Nghiem, L.D. Monitoring antibiotic resistance genes in wastewater treatment: Current strategies and future challenges. Sci. Total. Environ. 2021, 783, 146964. [Google Scholar] [CrossRef]
- Alam, M.-U.; Ferdous, S.; Ercumen, A.; Lin, A.; Kamal, A.; Luies, S.K.; Sharior, F.; Khan, R.; Rahman, Z.; Parvez, S.M.; et al. Effective Treatment Strategies for the Removal of Antibiotic-Resistant Bacteria, Antibiotic-Resistance Genes, and Antibiotic Residues in the Effluent From Wastewater Treatment Plants Receiving Municipal, Hospital, and Domestic Wastewater: Protocol for a Systematic Review. JMIR Res. Protoc. 2021, 10, e33365. [Google Scholar] [CrossRef]
- Meier, H.; Spinner, K.; Crump, L.; Kuenzli, E.; Schuepbach, G.; Zinsstag, J. State of Knowledge on the Acquisition, Diversity, Interspecies Attribution and Spread of Antimicrobial Resistance between Humans, Animals and the Environment: A Systematic Review. Antibiotics 2022, 12, 73. [Google Scholar] [CrossRef]
- Xiao, R.; Huang, D.; Du, L.; Song, B.; Yin, L.; Chen, Y.; Gao, L.; Li, R.; Huang, H.; Zeng, G. Antibiotic resistance in soil-plant systems: A review of the source, dissemination, influence factors, and potential exposure risks. Sci. Total. Environ. 2023, 869, 161855. [Google Scholar] [CrossRef]
- Kim, J.Y.; Suh, J.W.; Kang, J.S.; Kim, S.B.; Yoon, Y.K.; Sohn, J.W. Gram-Negative Bacteria’s Outer Membrane Vesicles. Infect. Chemother. 2023, 55, 557902. [Google Scholar] [CrossRef]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.; Cork, S.C.; Ronksley, P.E.; Barkema, H.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- WHO. Stop Using Antibiotics in Healthy Animals to Prevent the Spread of Antibiotic Resistance. 7 November 2017. Available online: https://www.who.int/news/item/07-11-2017-stop-using-antibiotics-in-healthy-animals-to-prevent-the-spread-of-antibiotic-resistance (accessed on 19 February 2023).
- Serwecińska, L. Antimicrobials and Antibiotic-Resistant Bacteria: A Risk to the Environment and to Public Health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- van Staa, T.P.; Palin, V.; Li, Y.; Welfare, W.; Felton, T.W.; Dark, P.; Ashcroft, D.M. The effectiveness of frequent antibiotic use in reducing the risk of infection-related hospital admissions: Results from two large population-based cohorts. BMC Med. 2020, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Sharma, S. Role of alternatives to antibiotics in mitigating the antimicrobial resistance crisis. Indian J. Med. Res. 2022, 156, 464–477. [Google Scholar]
- Ye, J.; Chen, X. Current Promising Strategies against Antibiotic-Resistant Bacterial Infections. Antibiotics 2022, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, S.; Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry of Russian Academy of Sciences; Khaitov, R.; National Research Center—Institute of Immunology of the Federal Medico-Biological Agency. Glucosaminylmuramyldipeptide—GMDP: Effect on mucosal immunity (on the issue of immunotherapy and immunoprophylaxis). Immunologiya 2020, 41, 174–183. [Google Scholar] [CrossRef]
- Rechkina, E.A.; Denisova, G.F.; Masalova, O.V.; Lideman, L.F.; Denisov, D.A.; Lesnova, E.I.; Ataullakhanov, R.I.; Gur’Ianova, S.V.; Kushch, A. Epitope mapping of antigenic determinants of hepatitis C virus proteins by phage display. Mol. Biologiia 2006, 40, 357–368. (In Russian) [Google Scholar]
- Konorev, M.R.; Guryanova, S.V.; Tyshevich, E.N.; Pavlyukov, R.A.; Borisova, O.Y. Advisable including glucosaminylmuramyldipeptide in Helicobacter pylori therapy: Experience of ten-year investigation. Rudn. J. Med. 2020, 24, 269–282. [Google Scholar] [CrossRef]
- L’vov, V.L.; Gur’yanova, S.V.; Rodionov, A.V.; Gorshkova, R.P. Structure of the repeating unit of the O-specific polysaccha-ride of the lipopolysaccharide of yersinia kristensenii strain 490 (O:12,25). Carbohydr. Res. 1992, 228, 415–422. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- L’Vov, V.L.; Gur’Ianova, S.V.; Rodionov, A.V.; Dmitriev, B.A.; Shashkov, A.S.; Ignatenko, A.V.; Gorshkova, R.P.; Ovodov, I.S. The structure of a repetitive unit of the glycerolphosphate- containing O-specific polysaccharide chain from Yersinia kristensenii strain 103 (0:12,26) lipopolysaccharide. Bioorg. Khim. 1990, 16, 379–389. [Google Scholar] [PubMed]
- Tataurshchikova, N. OM-85: Personalized approach to the treatment of acute respiratory infections in children. Quest. Pract. Pediatr. 2020, 15, 61–68. [Google Scholar] [CrossRef]
- Ayswaria, R.; Vijayan, J.; Nathan, V.K. Antimicrobial peptides derived from microalgae for combating antibiotic resistance: Current status and prospects. Cell Biochem. Funct. 2023, 41, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Cooper, I.I. A review of the potential for bacteriophages to effect antibiofilm activity, using selected examples. J. Appl. Microbiol. 2022, 134, lxac056. [Google Scholar] [CrossRef]
- Tataurshchikova, N.S.; Sidorovich, I.G. The cytokine status as a criterion for the efficacy of intranasal aerosol therapy with the use of a cycloferon solution in the patients presenting with allergic rhinosinusitis. Vestn. Otorinolaringol. 2012, 3, 79–82. [Google Scholar]
- Al-Awsi, G.R.L.; Alameri, A.A.; Al-Dhalimy, A.M.B.; Gabr, G.A.; Kianfar, E. Application of nano-antibiotics in the diagnosis and treatment of infectious diseases. Braz. J. Biol. 2023, 84, e264946. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Ovchinnikova, T.V. Immunomodulatory and Allergenic Properties of Antimicrobial Peptides. Int. J. Mol. Sci. 2022, 23, 2499. [Google Scholar] [CrossRef]
- Reeves, P. The Bacteriocins. Bacteriol. Rev. 1965, 29, 24–45. [Google Scholar] [CrossRef]
- Gratia, A. “Sur un remarquable exemple d’antagonisme entre deux souches de coilbacille” [On a remarkable example of antagonism between two strains of coilbacille]. Compt. Rend. Soc. Biol. 1925, 93, 1040–1042. NAID 10027104803. (In French) [Google Scholar]
- Fredericq, P. Colicines et Autres Bacteriocines [Colicines and Other Bacteriocins]. Ergeb. Mikrobiol. Immun. Exp. Ther. 1963, 37, 114–161. PMID: 14324288. (In French) [Google Scholar] [PubMed]
- Cotter, P.D.; Ross, R.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Genet. 2012, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Hill, C.; Ross, R.P. Food microbiology: Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777. [Google Scholar] [CrossRef]
- Wilaipun, P.; Zendo, T.; Sangjindavong, M.; Nitisinprasert, S.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. The two-synergistic peptide bacteriocin produced by Enterococcus faecium NKR- 5-3 isolated from Thai fermented fish (Plara). Sci. Asia 2004, 30, 115–122. [Google Scholar] [CrossRef]
- Fatima, D.; Mebrouk, K. Characterization and determination of the factors affecting anti-listerial bacteriocins from Lactobacillus plantarum and Pediococcus pentosaceus isolated from dairy milk products. Afr. J. Food Sci. 2013, 7, 35–44. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Darbandi, A.; Asadi, A.; Ari, M.M.; Ohadi, E.; Talebi, M.; Zadeh, M.H.; Emamie, A.D.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and potential use as antimicrobials. J. Clin. Lab. Anal. 2021, 36, e24093. [Google Scholar] [CrossRef]
- Benech, R.-O.; Kheadr, E.E.; Lacroix, C.; Fliss, I. Antibacterial Activities of Nisin Z Encapsulated in Liposomes or Produced In Situ by Mixed Culture during Cheddar Cheese Ripening. Appl. Environ. Microbiol. 2002, 68, 5607–5619. [Google Scholar] [CrossRef] [Green Version]
- Chandrakasan, G.; Rodríguez-Hernández, A.-I.; López-Cuellar, M.D.R.; Palma-Rodríguez, H.-M.; Chavarría-Hernández, N. Bacteriocin encapsulation for food and pharmaceutical applications: Advances in the past 20 years. Biotechnol. Lett. 2019, 41, 453–469. [Google Scholar] [CrossRef]
- Habib, W.; Sakr, A. Development and human in vivo evaluation of a colonic drug delivery system. Pharm. Ind. 1999, 61, 1145–1149. [Google Scholar]
- Gough, R.; Cabrera Rubio, R.; O’Connor, P.M.; Crispie, F.; Brodkorb, A.; Miao, S.; Hill, C.; Ross, R.P.; Cotter, P.D.; Nilaweera, K.N.; et al. Oral delivery of nisin in resistant starch based matrices alter the gut micro- biota in mice. Front Microbiol. 2018, 9, 1186. [Google Scholar] [CrossRef] [Green Version]
- Rollema, H.S.; Kuipers, O.P.; Both, P.; De Vos, W.M.; Siezen, R.J. Improvement of solubility and stability of the antimicrobial peptide nisin by protein engineering. Appl. Environ. Microbiol. 1995, 61, 2873–2878. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Blake, T.; Mathur, H.; O’Connor, P.M.; Cotter, P.D.; Ross, R.P.; Hill, C. Bioengineering Nisin to overcome the Nisin Resistance Protein. Mol. Microbiol. 2019, 111, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Duquesne, S.; Destoumieux-Garz’on, D.; Peduzzi, J.; Rebuffat, S. Microcins, gene-encoded antibacterial peptides from enterobacteria. Nat. Prod. Rep. 2007, 24, 708–734. [Google Scholar] [CrossRef] [PubMed]
- Drider, D.; Rebuffat, S. Prokaryotic Antimicrobial Peptides: From Genes to Applications; Springer Science & Business Media: Berlin/Heidelburg, Germany, 2011. [Google Scholar]
- Hammami, R.; Fernandez, B.; Lacroix, C.; Fliss, I. Anti-infective properties of bacteriocins: An update. Cell. Mol. Life Sci. 2013, 70, 2947–2967. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.; Cotter, P.; Ross, R.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B.; Sc, Y.; Ch, L.; Ct, S.; Jy, F. Faculty Opinions recommendation of Antibacterial activities of bacteriocins: Application in foods and pharmaceuticals. Front Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Riley, M.A.; Wertz, J.E. Bacteriocins evolution, ecology, and application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef] [Green Version]
- Shand, R.F.; Leyva, K.J. Archaeal antimicrobials: An undiscovered country. In Archaea: New Models for Prokaryotic Biology; Caister Academic Press: Norfolk, UK, 2008; 248p. [Google Scholar]
- Besse, A.; Peduzzi, J.; Rebuffat, S.; Carré-Mlouka, A. Antimicrobial peptides and proteins in the face of extremes: Lessons from archaeocins. Biochimie 2015, 118, 344–355. [Google Scholar] [CrossRef]
- Oman, T.J.; van der Donk, W.A. Insights into the mode of action of the two-peptide lantibiotic haloduracin. ACS Chem. Biol. 2009, 4, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Hanchi, H.; Hammami, R.; Gingras, H.; Kourda, R.; Bergeron, M.G.; Ben Hamida, J.; Ouellette, M.; Fliss, I. Inhibition of MRSA and of Clostridium difficile by durancin 61A: Synergy with bacteriocins and antibiotics. Futur. Microbiol. 2017, 12, 205–212. [Google Scholar] [CrossRef]
- Cui, Z.; Chen, Z.-H.; Zhang, Q.; Gribova, V.V.; Filaretov, V.F.; Huang, D.-S. RMSCNN: A Random Multi-Scale Convolutional Neural Network for Marine Microbial Bacteriocins Identification. IEEE/ACM Trans. Comput. Biol. Bioinform. 2021, 19, 3663–3672. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-Origin Lactic Acid Bacteria May Exhibit Probiotic Properties: Review. BioMed Res. Int. 2018, 2018, 5063185. [Google Scholar] [CrossRef] [Green Version]
- Verso, L.L.; Lessard, M.; Talbot, G.; Fernandez, B.; Fliss, I. Isolation and Selection of Potential Probiotic Bacteria from the Pig Gastrointestinal Tract. Probiotics Antimicrob. Proteins 2018, 10, 299–312. [Google Scholar] [CrossRef]
- Ryan, K.; Jayaraman, T.; Daly, P.; Canchaya, C.; Curran, S.; Fang, F.; Quigley, E.; O’toole, P. Isolation of lactobacilli with probiotic properties from the human stomach. Lett. Appl. Microbiol. 2008, 47, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef] [PubMed]
- Gautam, N.; Sharma, N. Purification and characterization of bacteriocin produced by strain of Lactobacillus brevis MTCC 7539. Indian J. Biochem. Biophys. 2009, 46, 337–341. [Google Scholar] [PubMed]
- Knorr, D. Technology aspects related to microorganisms in functional foods. Trends Food Sci. Technol. 1998, 9, 295–306. [Google Scholar] [CrossRef]
- Karpinski, T.; Szkaradkiewicz, A.K. Characteristic of Bacteriocines and Their Application. Pol. J. Microbiol. 2013, 62, 223–235. [Google Scholar] [CrossRef]
- Choi, S.; Baek, M.-G.; Chung, M.-J.; Lim, S.; Yi, H. Distribution of bacteriocin genes in the lineages of Lactiplantibacillus plantarum. Sci. Rep. 2021, 11, 20063. [Google Scholar] [CrossRef]
- Collins, F.W.J.; O’Connor, P.M.; O’Sullivan, O.; Gómez-Sala, B.; Rea, M.C.; Hill, C.; Ross, R.P. Bacteriocin Gene-Trait matching across the complete Lactobacillus Pan-genome. Sci. Rep. 2017, 7, 3481. [Google Scholar] [CrossRef] [Green Version]
- Chérier, D.; Patin, D.; Blanot, D.; Touzé, T.; Barreteau, H. The Biology of Colicin M and Its Orthologs. Antibiotics 2021, 10, 1109. [Google Scholar] [CrossRef]
- De Jong, A.; Van Hijum, S.A.F.T.; Bijlsma, J.J.E.; Kok, J.; Kuipers, O.P. BAGEL: A web-based bacteriocin genome mining tool. Nucleic Acids Res. 2006, 34, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Spangler, R.; Zhang, S.P.; Krueger, J.; Zubay, G. Colicin synthesis and cell death. J. Bacteriol. 1985, 163, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Freire Bastos, M.d.C.; Coelho, M.L.V.; da Silva Santos, O.C. Resistance to bacteriocins produced by Gram-positive bacteria. Microbiol 2015, 161, 683–700. [Google Scholar] [CrossRef] [Green Version]
- Bountra, K.; Hagelueken, G.; Choudhury, H.G.; Corradi, V.; El Omari, K.; Wagner, A.; Mathavan, I.; Zirah, S.; Wahlgren, W.Y.; Tieleman, D.P.; et al. Structural basis for antibacterial peptide selfimmunity by the bacterial ABC transporter McjD. EMBO J. 2017, 36, 3062–3079. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and Leaderless Bacteriocins: Biosynthesis, Mode of Action, Applications, and Prospects. Front. Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef] [Green Version]
- Žgur-Bertok, D. Regulating colicin synthesis to cope with stress and lethality of colicin production. Biochem. Soc. Trans. 2012, 40, 1507–1511. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Moia, G.d.S.; Silva, A.; Baraúna, R.A.; Veras, A.A.d.O. BADASS: BActeriocin-Diversity ASsessment Software. BMC Bioinform. 2023, 24, 24. [Google Scholar] [CrossRef]
- Hammami, R.; Zouhir, A.; Cetal, L.L. BACTIBASE second release: A database and tool platform for bacteriocin characteriza-tion. BMC Microbiol 2010, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- The sbv IMPROVER Project Team (in Alphabetical Order); Boué, S.; Fields, B.; Hoeng, J.; Park, J.; Peitsch, M.C.; Schlage, W.K.; Talikka, M.; Binenbaum, I.; Bondarenko, V.; et al. Enhancement of COPD biological networks using a web-based collaboration interface. F1000Research 2015, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, S.; Guryanova, A. sbv IMPROVER: Modern Approach to Systems Biology. Methods Mol. Biol. 2017, 1613, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Cai, J.; Zhang, B.; Wang, Y.; Wong, D.F.; Siu, S.W.I. Recent Progress in the Discovery and Design of Antimicrobial Peptides Using Traditional Machine Learning and Deep Learning. Antibiotics 2022, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Andrès, E. Cationic antimicrobial peptides in clinical development, with special focus on thanatin and heliomicin. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 31, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Panina, I.S.; Balandin, S.V.; Tsarev, A.V.; Chugunov, A.O.; Tagaev, A.A.; Finkina, E.I.; Antoshina, D.V.; Sheremeteva, E.V.; Paramonov, A.S.; Rickmeyer, J.; et al. Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity. Int. J. Mol. Sci. 2023, 24, 1332. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Bacteriocins. In Encyclopedia of Food and Health; Caballero, B., Ed.; Elsivier Science: Amsterdam, The Netherlands, 2016; pp. 312–319. [Google Scholar]
- Cotter, P.D. An ‘Upp’-turn in bacteriocin receptor identification. Mol. Microbiol. 2014, 92, 1159–1163. [Google Scholar] [CrossRef]
- Kouwen, T.R.H.M.; Trip, E.N.; Denham, E.L.; Sibbald, M.J.J.B.; Dubois, J.-Y.F.; van Dijl, J.M. The Large Mechanosensitive Channel MscL Determines Bacterial Susceptibility to the Bacteriocin Sublancin 168. Antimicrob. Agents Chemother. 2009, 53, 4702–4711. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Tarr, A.W.; Penfold, C.N. Colicin import into E. coli cells: A model system for insights into the import mechanisms of bacteriocins. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2014, 1843, 1717–1731. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Khait, R.; Harmalkar, A.; Pham, P.; Webby, M.N.; Housden, N.G.; Elliston, E.; Hopper, J.T.S.; Mohammed, S.; Robinson, C.V.; Gray, J.J.; et al. Colicin-Mediated Transport of DNA through the Iron Transporter FepA. mBio 2021, 12, e0178721. [Google Scholar] [CrossRef]
- Bieler, S.; Silva, F.; Soto, C.; Belin, D. Bactericidal Activity of both Secreted and Nonsecreted Microcin E492 Requires the Mannose Permease. J. Bacteriol. 2006, 188, 7049–7061. [Google Scholar] [CrossRef] [Green Version]
- James, R.; Lazdunski, C.; Pattus, F. Bacteriocins, Microcins and Lantibiotics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 65. [Google Scholar]
- Chiumento, S.; Roblin, C.; Kieffer-Jaquinod, S.; Tachon, S.; Leprètre, C.; Basset, C.; Aditiyarini, D.; Olleik, H.; Nicoletti, C.; Bornet, O.; et al. Ruminococcin C, a promising antibiotic produced by a human gut symbiont. Sci. Adv. 2019, 5, eaaw9969. [Google Scholar] [CrossRef] [Green Version]
- Hatakka, K.; Saxelin, M. Probiotics in intestinal and non-intestinal infectious diseases—Clinical evidence. Curr. Pharm. Design. 2008, 14, 1351–1367. [Google Scholar] [CrossRef] [PubMed]
- Katla, T.; Naterstad, K.; Vancanneyt, M.; Swings, J.; Axelsson, L. Differences in Susceptibility of Listeria monocytogenes Strains to Sakacin P, Sakacin A, Pediocin PA-1, and Nisin. Appl. Environ. Microbiol. 2003, 69, 4431–4437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, B.; Guinane, C.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Assessing the Contributions of the LiaS Histidine Kinase to the Innate Resistance of Listeria monocytogenes to Nisin, Cephalosporins, and Disinfectants. Appl. Environ. Microbiol. 2012, 78, 2923–2929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, B.; Curtis, N.; Cotter, P.D.; Hill, C.; Ross, R.P. The ABC Transporter AnrAB Contributes to the Innate Resistance of Listeria monocytogenes to Nisin, Bacitracin, and Various β-Lactam Antibiotics. Antimicrob. Agents Chemother. 2010, 54, 4416–4423. [Google Scholar] [CrossRef] [Green Version]
- Rasch, M.; Knochel, S. Variations in tolerance of Listeria monocytogenes to nisin, pediocin PA-1 and bavaricin A. Lett. Appl. Microbiol. 1998, 27, 275–278. [Google Scholar] [CrossRef]
- Nes, I.F.; Gabrielsen, C.; Brede, D.A.; Diep, D.B. Novel developments in bacteriocins from lactic acid bacteria. In Biotechnology of Lactic Acid Bacteria: Novel Applications, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 80–99. [Google Scholar]
- Sedgley, C.M.; Clewell, D.B.; Flannagan, S.E. Plasmid pAMS1-Encoded, Bacteriocin-Related “Siblicide” in Enterococcus faecalis. J. Bacteriol. 2009, 191, 3183–3188. [Google Scholar] [CrossRef] [Green Version]
- Martãnez, B.; Rodrãguez, A. Antimicrobial susceptibility of nisin resistant Listeria monocytogenes of dairy origin. FEMS Microbiol. Lett. 2005, 252, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Maaß, S.; Bartel, J.; Mücke, P.-A.; Schlüter, R.; Sura, T.; Zaschke-Kriesche, J.; Smits, S.H.J.; Becher, D. Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin. Cells 2021, 10, 372. [Google Scholar] [CrossRef]
- Lohans, C.T.; Vederas, J.C. Development of Class IIa Bacteriocins as Therapeutic Agents. Int. J. Microbiol. 2012, 2012, 386410. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of innate immunity by a bacterial secreted peptide: Lantibiotic nisin Z is selectively immunomodulatory. J. Endotoxin Res. 2013, 19, 315–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pablo, M.A.; Gaforio, J.J.; Gallego, A.M.; Ortega, E.; Gã¡Lvez, A.M.; Lã³Pez, G.A.d.C. Evaluation of immunomodulatory effects of nisin-containing diets on mice. FEMS Immunol. Med. Microbiol. 1999, 24, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.M.; Ateia, I.; Paulus, J.R.; Liu, H.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Antimicrobial nisin acts against saliva derived multi-species biofilms without cytotoxicity to human oral cells. Front. Microbiol. 2015, 6, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.C.; Schroeder, H.E. Pathogenesis of inflammatory periodontal disease. A summary of current work. Lab. Investig. 1976, 34, 235. [Google Scholar] [PubMed]
- Radaic, A.; Brody, H.; Contreras, F.; Hajfathalian, M.; Lucido, L.; Kamarajan, P.; Kapila, Y.L. Nisin and Nisin Probiotic Disrupt Oral Pathogenic Biofilms and Restore Their Microbiome Composition towards Healthy Control Levels in a Peri-Implantitis Setting. Microorganisms 2022, 10, 1336. [Google Scholar] [CrossRef]
- Jia, Z.; He, M.; Wang, C.; Chen, A.; Zhang, X.; Xu, J.; Fu, H.; Liu, B. Nisin reduces uterine inflammation in rats by modulating concentrations of pro and antiinflammatory cytokines. Am. J. Reprod. Immunol. 2019, 81, e13096. [Google Scholar] [CrossRef]
- Gao, L.; Kuraji, R.; Zhang, M.J.; Martinez, A.; Radaic, A.; Kamarajan, P.; Le, C.; Zhan, L.; Ye, C.; Rangé, H.; et al. Nisin probiotic prevents inflammatory bone loss while promoting reparative proliferation and a healthy microbiome. NPJ Biofilms Microbiomes 2022, 8, 45. [Google Scholar] [CrossRef]
- Karimi, M.; Maghsoud, Z.; Halabian, R. Effect of Preconditioned Mesenchymal Stem Cells with Nisin Prebiotic on the Expression of Wound Healing Factors Such as TGF-β1, FGF-2, IL-1, IL-6, and IL-10. Regen. Eng. Transl. Med. 2021, 7, 30–40. [Google Scholar] [CrossRef]
- Małaczewska, J.; Kaczorek-Łukowska, E.; Wójcik, R.; Rękawek, W.; Siwicki, A.K. In vitro immunomodulatory effect of nisin on porcine leucocytes. J. Anim. Physiol. Anim. Nutr. 2019, 103, 882–893. [Google Scholar] [CrossRef]
- Śmiałek, J.; Bzowska, M.; Hinz, A.; Mężyk-Kopeć, R.; Sołtys, K.; Mak, P. Bacteriocin BacSp222 and Its Succinylated Forms Exhibit Proinflammatory Activities Toward Innate Immune Cells. J. Inflamm. Res. 2022, 15, 4601–4621. [Google Scholar] [CrossRef]
- Śmiałek, J.; Nowakowski, M.; Bzowska, M.; Bocheńska, O.; Wlizło, A.; Kozik, A.; Dubin, G.; Mak, P. Structure, Biosynthesis, and Biological Activity of Succinylated Forms of Bacteriocin BacSp222. Int. J. Mol. Sci. 2021, 22, 6256. [Google Scholar] [CrossRef] [PubMed]
- Cebrian, R.; Rodriguez-Cabezas, M.E.; Martín-Escolano, R.; Rubiño, S.; Barros, M.G.; Montalbán-López, M.; Rosales, M.J.; Sánchez-Moreno, M.; Valdivia, E.; Martínez-Bueno, M.; et al. Preclinical studies of toxicity and safety of the AS-48 bacteriocin. J. Adv. Res. 2019, 20, 129–139. [Google Scholar] [CrossRef]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory efficacy of nisin-a bacterial lantibiotic peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.; Yang, G.; Tao, L. Sublancin protects against methicillin-resistant Staphylococcus aureus infection by the combined modulation of innate immune response and microbiota. Peptides 2021, 141, 170533. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ye, Q.; Wang, K.; Zeng, X.; Huang, S.; Yu, H.; Ge, Q.; Qi, D.; Qiao, S. Enhancement of Macrophage Function by the Antimicrobial Peptide Sublancin Protects Mice from Methicillin-Resistant Staphylococcus aureus. J. Immunol. Res. 2019, 2019, 3979352. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Huang, S.; Ye, Q.; Zeng, X.; Yu, H.; Qi, D.; Qiao, S. Prevention of Cyclophosphamide-Induced Immunosuppression in Mice with the Antimicrobial Peptide Sublancin. J. Immunol. Res. 2018, 2018, 4353580. [Google Scholar] [CrossRef] [Green Version]
- Bu, Y.; Liu, Y.; Liu, Y.; Wang, S.; Liu, Q.; Hao, H.; Yi, H. Screening and Probiotic Potential Evaluation of Bacteriocin-Producing Lactiplantibacillus plantarum In Vitro. Foods 2022, 11, 1575. [Google Scholar] [CrossRef]
- Antoshina, D.V.; Balandin, S.V.; Bogdanov, I.V.; Vershinina, M.A.; Sheremeteva, E.V.; Toropygin, I.Y.; Finkina, E.I.; Ovchinnikova, T.V. Antimicrobial Activity and Immunomodulatory Properties of Acidocin A, the Pediocin-like Bacteriocin with the Non-Canonical Structure. Membranes 2022, 12, 1253. [Google Scholar] [CrossRef]
- Teiar, R.; Pérez-Ramos, A.; Zgheib, H.; Cudennec, B.; Belguesmia, Y.; Drider, D. Anti-adhesion and Anti-inflammatory Potential of the Leaderless Class IIb Bacteriocin Enterocin DD14. Probiotics Antimicrob. Proteins 2022, 14, 613–619. [Google Scholar] [CrossRef]
- Donovick, R.; Pagano, J.F.; Stout, H.A.; Weinstein, M.J. Thiostrepton, a new antibi-otic. I. In vitro studies. Antibiot Annu. 1955, 3, 554–559. [Google Scholar]
- Chiu, M.L.; Folcher, M.; Griffin, P.; Holt, T.; Klatt, T.; Thompson, C.J. Characterization of the covalent binding of thiostrepton to a thiostrepton-induced protein from Streptomyces lividans. Biochemistry 1996, 35, 2332–2341. [Google Scholar] [CrossRef] [PubMed]
- Halasi, M.; Zhao, H.; Dahari, H.; Bhat, U.G.; Gonzalez, E.B.; Lyubimo, A.V.; Tonetti, D.A.; Gartel, A.L. Thiazole antibiotics against breast cancer. Cell Cycle 2010, 9, 1214–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, S.; Lamore, S.D.; Cabello, C.M.; Lesson, J.L.; Munoz-Rodriguez, J.L.; Wondrak, G.T. Thiostrepton is an inducer of oxidative and proteotoxic stress that impairs viability of human melanoma cells but not primary melanocytes. Biochem. Pharmacol. 2012, 83, 1229–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newick, K.; Cunniff, B.; Preston, K.; Held, P.; Arbiser, J.; Pass, H.; Mossman, B.; Shukla, A.; Heintz, N. Peroxiredoxin 3 is a re-dox-dependent target of thiostrepton in malignant mesothelioma cells. PLoS ONE 2012, 7, e39404. [Google Scholar] [CrossRef] [Green Version]
- Hegde, N.S.; Sanders, D.A.; Rodriguez, R.; Balasubramanian, S. The transcription factor FOXM1 is a cellular target of the natural product thiostrepton. Nat. Chem. 2011, 3, 725–731. [Google Scholar] [CrossRef]
- Bhat, U.G.; Halasi, M.; Gartel, A.L. FoxM1 is a general target for proteasome in-hibitors. PLoS ONE 2009, 4, e6593. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C. The bacterial thiopeptide thiostrepton. An update of its mode of action, pharmacological properties and applications. Eur. J. Pharmacol. 2022, 914, 174661. [Google Scholar] [CrossRef]
- Sandu, C.; Wetie, A.G.N.; Darie, C.C.; Steller, H. Thiostrepton, a Natural Compound That Triggers Heat Shock Response and Apoptosis in Human Cancer Cells: A Proteomics Investigation. Adv. Exp. Med. Biol. 2014, 806, 443–451. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Yeh, D.-W.; Lu, C.-H.; Liu, Y.-L.; Huang, L.-R.; Kao, C.-Y.; Chen, H.-Y.; Huang, C.-Y.F.; Chang, C.-H.; Luo, Y.; et al. Identification of Thiostrepton as a Novel Inhibitor for Psoriasis-like Inflammation Induced by TLR7–9. J. Immunol. 2015, 195, 3912–3921. [Google Scholar] [CrossRef] [Green Version]
- Salvador, P.B.U.; Dalmacio, L.M.M.; Kim, S.H.; Kang, D.-K.; Balolong, M.P. Immunomodulatory potential of four candidate probiotic Lactobacillus strains from plant and animal origin using comparative genomic analysis. Access Microbiol. 2021, 3, 000299. [Google Scholar] [CrossRef]
- Paiva, A.D.; de Oliveira, M.D.; de Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 2012, 158, 2851–2858. [Google Scholar] [CrossRef]
- Dreyer, L.; Smith, C.; Deane, S.M.; Dicks, L.M.T.; van Staden, A.D. Migration of Bacteriocins across Gastrointestinal Epithelial and Vascular Endothelial Cells, as Determined Using In Vitro Simulations. Sci. Rep. 2019, 9, 11481. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications. Microb. Physiol. 2007, 13, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, B.; Le Lay, C.; Jean, J.; Fliss, I. Growth, acid production and bacteriocin production by probiotic candidates under simulated colonic conditions. J. Appl. Microbiol. 2013, 114, 877–885. [Google Scholar] [CrossRef]
- Kheadr, E.; Zihler, A.; Dabour, N.; Lacroix, C.; Le Blay, G.; Fliss, I. Study of the physicochemical and biological stability of pediocin PA-1 in the upper gastrointestinal tract conditions using a dynamic in vitro model. J. Appl. Microbiol. 2010, 109, 54–64. [Google Scholar] [CrossRef]
- Birri, D.J.; Brede, D.A.; Nes, I.F. Salivaricin D, a Novel Intrinsically Trypsin-Resistant Lantibiotic from Streptococcus salivarius 5M6c Isolated from a Healthy Infant. Appl. Environ. Microbiol. 2012, 78, 402–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.M.; Jung, Y.-G.; Jin, Y.-Y.; Jayabalan, R.; Yang, S.H.; Suh, J.W. Bacteriocins as food preservatives: Challenges and emerging horizons. Crit. Rev. Food Sci. Nutr. 2018, 58, 2743–2767. [Google Scholar] [CrossRef]
- Jarvis, B.; Mahoney, R. Inactivation of Nisin by Alpha-Chymotrypsin. J. Dairy Sci. 1969, 52, 1448–1450. [Google Scholar] [CrossRef]
- Gough, R.; O’Connor, P.M.; Rea, M.C.; Gómez-Sala, B.; Miao, S.; Hill, C.; Brodkorb, A. Simulated gastrointestinal digestion of nisin and interaction between nisin and bile. LWT 2017, 86, 530–537. [Google Scholar] [CrossRef] [Green Version]
- Pomares, M.F.; Salomón, R.A.; Pavlova, O.; Severinov, K.; Farías, R.; Vincent, P.A. Potential Applicability of Chymotrypsin-Susceptible Microcin J25 Derivatives to Food Preservation. Appl. Environ. Microbiol. 2009, 75, 5734–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naimi, S.; Zirah, S.; Hammami, R.; Fernandez, B.; Rebuffat, S.; Fliss, I. Fate and Biological Activity of the Antimicrobial Lasso Peptide Microcin J25 under Gastrointestinal Tract Conditions. Front. Microbiol. 2018, 9, 1764. [Google Scholar] [CrossRef] [PubMed]
- Soleimanpour, M.; Mirhaji, S.S.; Jafari, S.; Derakhshankhah, H.; Mamashli, F.; Nedaei, H.; Karimi, M.R.; Motasadizadeh, H.; Fatahi, Y.; Ghasemi, A.; et al. Designing a new alginate-fibrinogen biomaterial composite hydrogel for wound healing. Sci. Rep. 2022, 12, 7213. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Pourakbari, B.; Bahador, A. Contribution of antimicrobial photo-sonodynamic therapy in wound healing: An in vivo effect of curcumin-nisin-based poly (L-lactic acid) nanoparticle on Acinetobacter baumannii biofilms. BMC Microbiol. 2022, 22, 28. [Google Scholar] [CrossRef]
- Cunha, E.; Tavares, L.; Oliveira, M. Revisiting Periodontal Disease in Dogs: How to Manage This New Old Problem? Antibiotics 2022, 11, 1729. [Google Scholar] [CrossRef]
- Lallukka, M.; Gamna, F.; Gobbo, V.A.; Prato, M.; Najmi, Z.; Cochis, A.; Rimondini, L.; Ferraris, S.; Spriano, S. Surface Functionalization of Ti6Al4V-ELI Alloy with Antimicrobial Peptide Nisin. Nanomaterials 2022, 12, 4332. [Google Scholar] [CrossRef]
- Azmi, M.F.; Al Khateeb, A.; Ab Rahim, S.; Froemming, G.R.A.; Omar, E. Effect of different solvents on nisin ZP potential as anticancer agent against MG-63 osteosarcoma cells. Asia Pac. J. Mol. Biol. Biotechnol. 2022, 30, 43–54. [Google Scholar] [CrossRef]
- Shin, J.; Gwak, J.; Kamarajan, P.; Fenno, J.; Rickard, A.; Kapila, Y. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [Green Version]
- Thanjavur, N.; Sangubotla, R.; Lakshmi, B.A.; Rayi, R.; Mekala, C.D.; Reddy, A.S.; Viswanath, B. Evaluating the antimicrobial and apoptogenic properties of bacteriocin (nisin) produced by Lactococcus lactis. Process. Biochem. 2022, 122, 76–86. [Google Scholar] [CrossRef]
- Ortiz-Rodríguez, T.; Mendoza-Acosta, F.; Martínez-Zavala, S.A.; Salcedo-Hernández, R.; Casados-Vázquez, L.E.; Bideshi, D.K.; Barboza-Corona, J.E. Thurincin H Is a Nonhemolytic Bacteriocin of Bacillus thuringiensis with Potential for Applied Use. Probiotics Antimicrob. Proteins 2022, 6, 77–84. [Google Scholar] [CrossRef]
- Cox, P.S.C.A.M.S.G.C.R.; Coburn, P.S.; Gilmore, M.S. Enterococcal Cytolysin: A Novel Two Component Peptide System that Serves as a Bacterial Defense against Eukaryotic and Prokaryotic Cells. Curr. Protein Pept. Sci. 2005, 6, 77–84. [Google Scholar] [CrossRef]
- Taggar, R.; Jangra, M.; Dwivedi, A.; Bansal, K.; Patil, P.B.; Bhattacharyya, M.S.; Nandanwar, H.; Sahoo, D.K. Bacteriocin isolated from the natural inhabitant of Allium cepa against Staphylococcus aureus. World J. Microbiol. Biotechnol. 2021, 37, 20. [Google Scholar] [CrossRef]
- Aguilar-Pérez, C.; Gracia, B.; Rodrigues, L.; Vitoria, A.; Cebrián, R.; Deboosère, N.; Song, O.-R.; Brodin, P.; Maqueda, M.; Aínsa, J.A. Synergy between Circular Bacteriocin AS-48 and Ethambutol against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62, e00359-18. [Google Scholar] [CrossRef] [Green Version]
- Behrens, H.M.; Six, A.; Walker, D.; Kleanthous, C. The therapeutic potential of bacteriocins as protein antibiotics. Emerg. Top. Life Sci. 2017, 1, 65–74. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Bacteriocins are produced by bacteria living in water, soil, and in numerous organisms. Commensal microorganisms colonizing human skin and mucous and bacteria that enter human organisms from the environment also produce bacteriocins. Bacteriocins produced by human microbiota and by bacteria entering the human body from the environment affect human immunity by influencing on various populations of immune cells.
Figure 1.
Bacteriocins are produced by bacteria living in water, soil, and in numerous organisms. Commensal microorganisms colonizing human skin and mucous and bacteria that enter human organisms from the environment also produce bacteriocins. Bacteriocins produced by human microbiota and by bacteria entering the human body from the environment affect human immunity by influencing on various populations of immune cells.

Figure 2.
Investigation of the immunomodulatory activity of bacteriocins in vivo, ex vivo, and in vitro.
Figure 2.
Investigation of the immunomodulatory activity of bacteriocins in vivo, ex vivo, and in vitro.

Figure 3.
Bacteriocins resources, activities, and perspective of application. Bacteriocins derived from archaea, Gram-positive, and Gram-negative bacteria, may limit the viability of related strains by various means (indicated by red lines). In vivo studies in rodent models, as well as ex vivo and in vitro studies on various immune cells and cell lines, have shown the ability of bacteriocins to activate various subpopulations of immunocompetent cells, demonstrating an immunomodulatory ability (indicated by blue arrows).
Figure 3.
Bacteriocins resources, activities, and perspective of application. Bacteriocins derived from archaea, Gram-positive, and Gram-negative bacteria, may limit the viability of related strains by various means (indicated by red lines). In vivo studies in rodent models, as well as ex vivo and in vitro studies on various immune cells and cell lines, have shown the ability of bacteriocins to activate various subpopulations of immunocompetent cells, demonstrating an immunomodulatory ability (indicated by blue arrows).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guryanova, S.V. Immunomodulation, Bioavailability and Safety of Bacteriocins. Life 2023, 13, 1521. https://doi.org/10.3390/life13071521
AMA Style
Guryanova SV. Immunomodulation, Bioavailability and Safety of Bacteriocins. Life. 2023; 13(7):1521. https://doi.org/10.3390/life13071521
Chicago/Turabian StyleGuryanova, Svetlana V. 2023. "Immunomodulation, Bioavailability and Safety of Bacteriocins" Life 13, no. 7: 1521. https://doi.org/10.3390/life13071521
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.