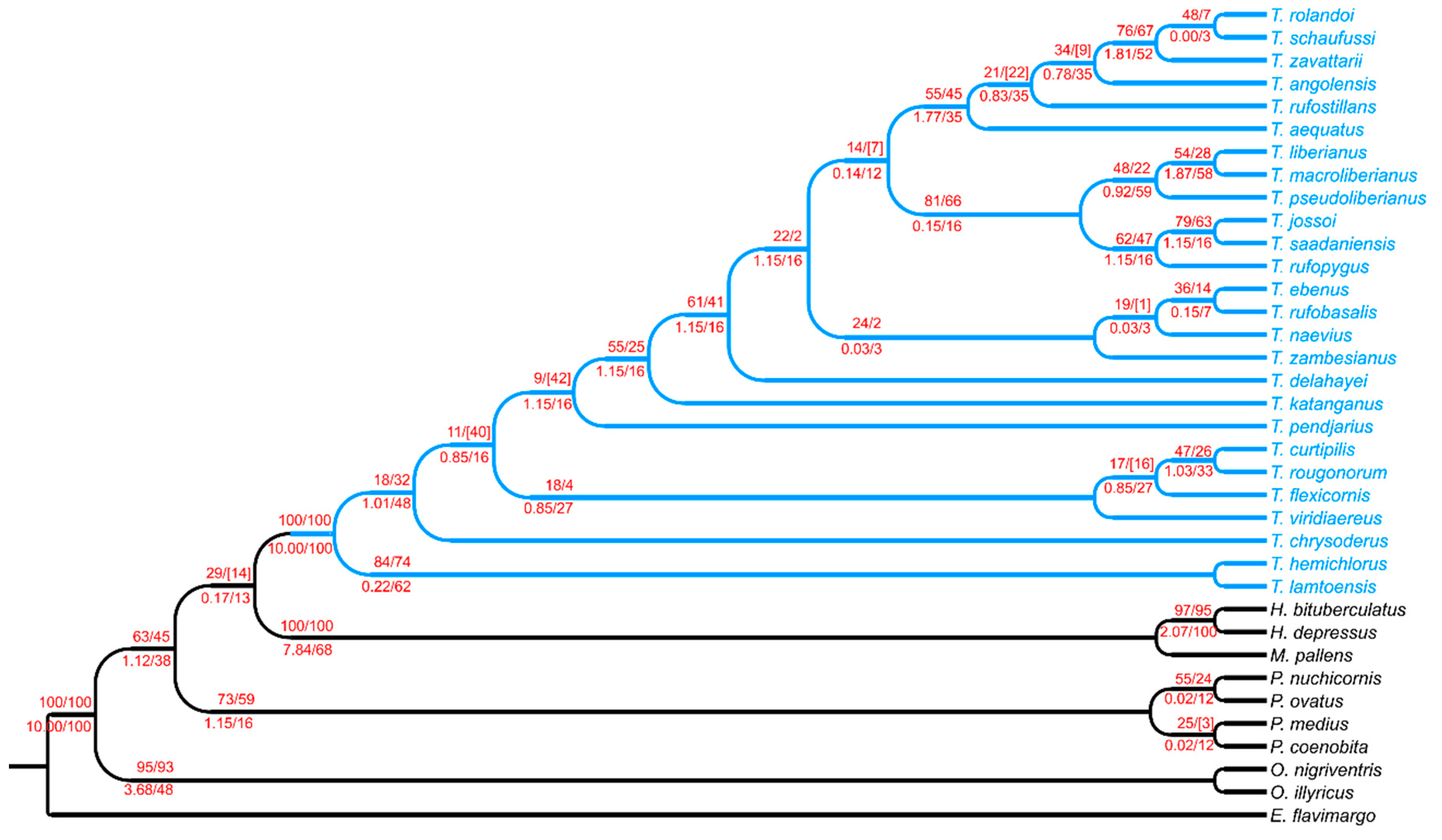
3.4.2. The Tiaronthophagus Species
At present 26 species are included in the genus Tiaronthophagus:
T. aequatus (Péringuey, 1901) n.comb.
T. angolensis n.sp.
T. chrysoderus (d’Orbigny, 1905a) n.comb.
T. curtipilis (d’Orbigny, 1905b) n.comb.
T. delahayei (Josso, 2011) n.comb.
T. ebenus (Péringuey, 1888) n.comb.
T. flexicornis (d’Orbigny, 1902) n.comb.
T. hemichlorus (d’Orbigny, 1915) n.comb.
T. jossoi n.sp.
T. katanganus n.sp.
T. lamtoensis (Cambefort, 1984) n.comb.
T. liberianus (Lansberge, 1883) n.comb.
T. macroliberianus (Moretto, 2010) n.comb.
T. naevius (d’Orbigny, 1913) n.comb.
T. pendjarius (Josso and Prévost, 2006) n.comb.
T. pseudoliberianus (Moretto, 2010) n.comb.
T. rolandoi n.sp.
T. rougonorum (Cambefort, 1984) n.comb.
T. rufobasalis (Fairmaire, 1887) n.comb.
T. rufopygus (Frey, 1957) n.comb.
T. rufostillans (d’Orbigny, 1907) n.comb.
T. saadaniensis n.sp.
T. schaufussi (Harold, 1867) n.comb.
T. viridiaereus (d’Orbigny, 1908) n.comb.
T. zambesianus n.sp.
T. zavattarii (Müller, 1939) n.comb.
Onthophagus aequatus Péringuey, 1901: 205.
Type material. Lectotype. ♂ (here designated) ‘Salisbury Mashonaland Feb 1899 G. A. K. Marshall in carrion <partly handwritten>’ ‘224 <handwritten>’ ‘Onthophagus aequatus ♂ P. type <handwritten>’ ‘Type SAM/Ent 2752’ ‘Lectotype Onthophagus aequatus Peringuey, 1901 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus aequatus (Peringuey, 1901) Roggero, Moretto, Barbero, Palestrini 2019’ (SAMC). Paralectotypes. Not yet traced.
Other material. BOTSWANA. 10 km NE Martins Drift (PMOC). MALAWI. Blantyre, Michiru Mt. Park (EBCT). TANZANIA. Chimala Escarpment (PMOC); Morogoro Reg., Mikesse Hills, 420 m (PMOC); Mts Uluguru (PMOC). ZAMBIA. Central Province, 25 km NE Lilemone 1250 m (PMOC); 60 km NE Serenje, Bwale (JFJC); Kafue NP, Kacheleko wildlife outpost (JFJC); Kafue NP, Tateyoyo (JFJC); Lower Zambesi NP, 3 km Mukamba gate (JFJC); Lusaka Province, Lower Zambesi NP (JFJC); Mfwanta 1355 m (JFJC); NW Province, 30 km N Lubungu, NE Kafue NP (JFJC); Lusaka (PMOC).
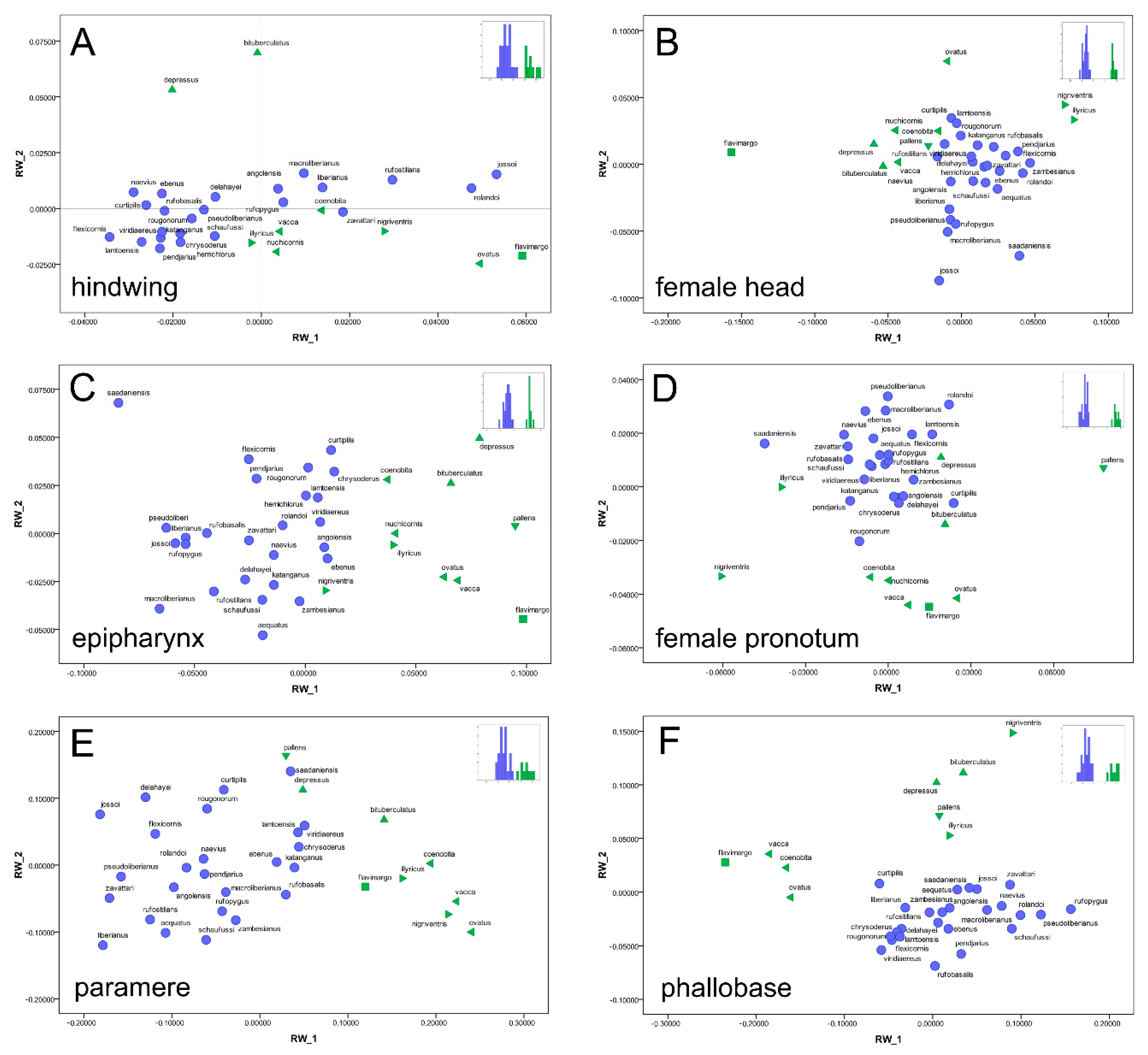
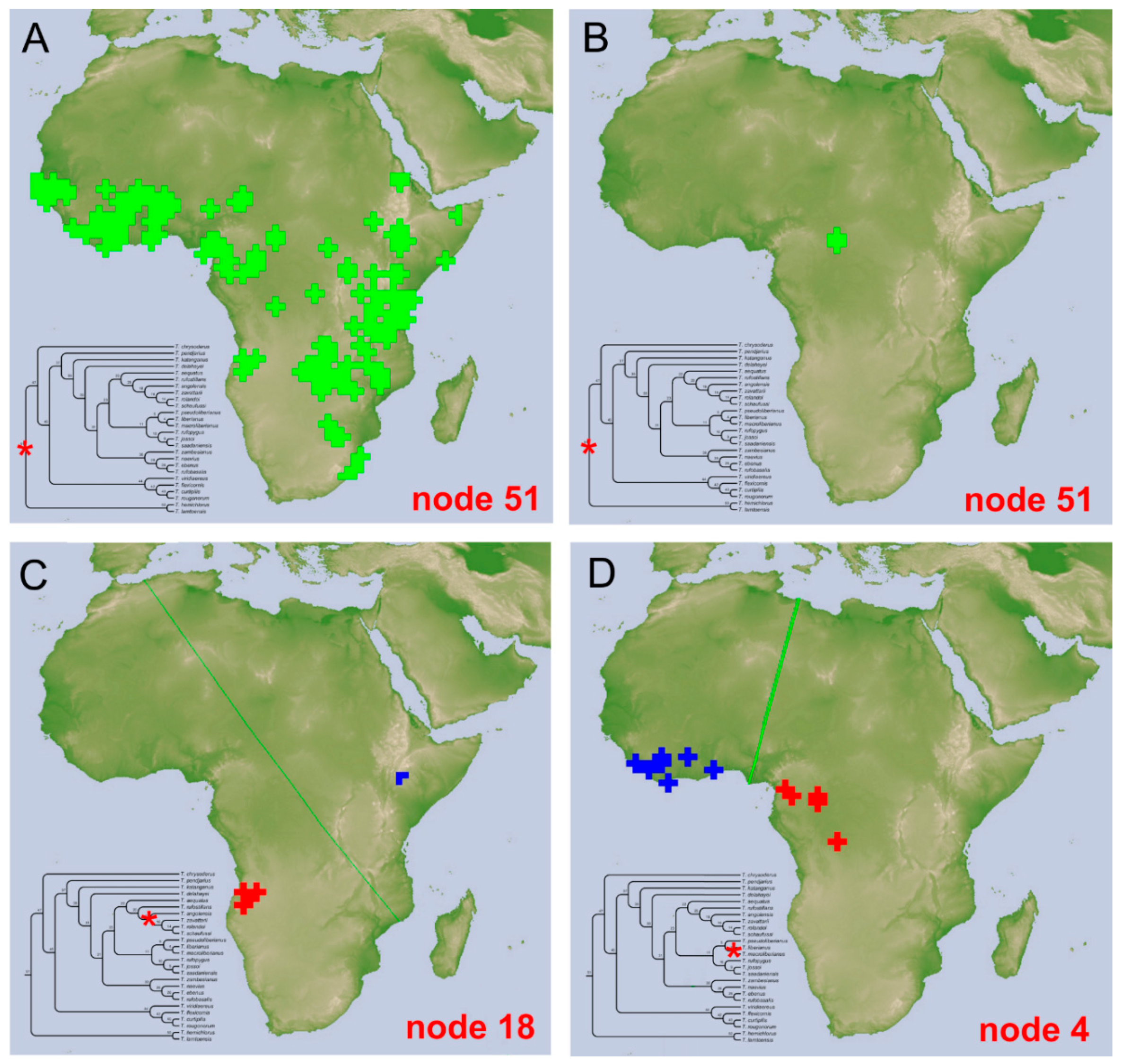
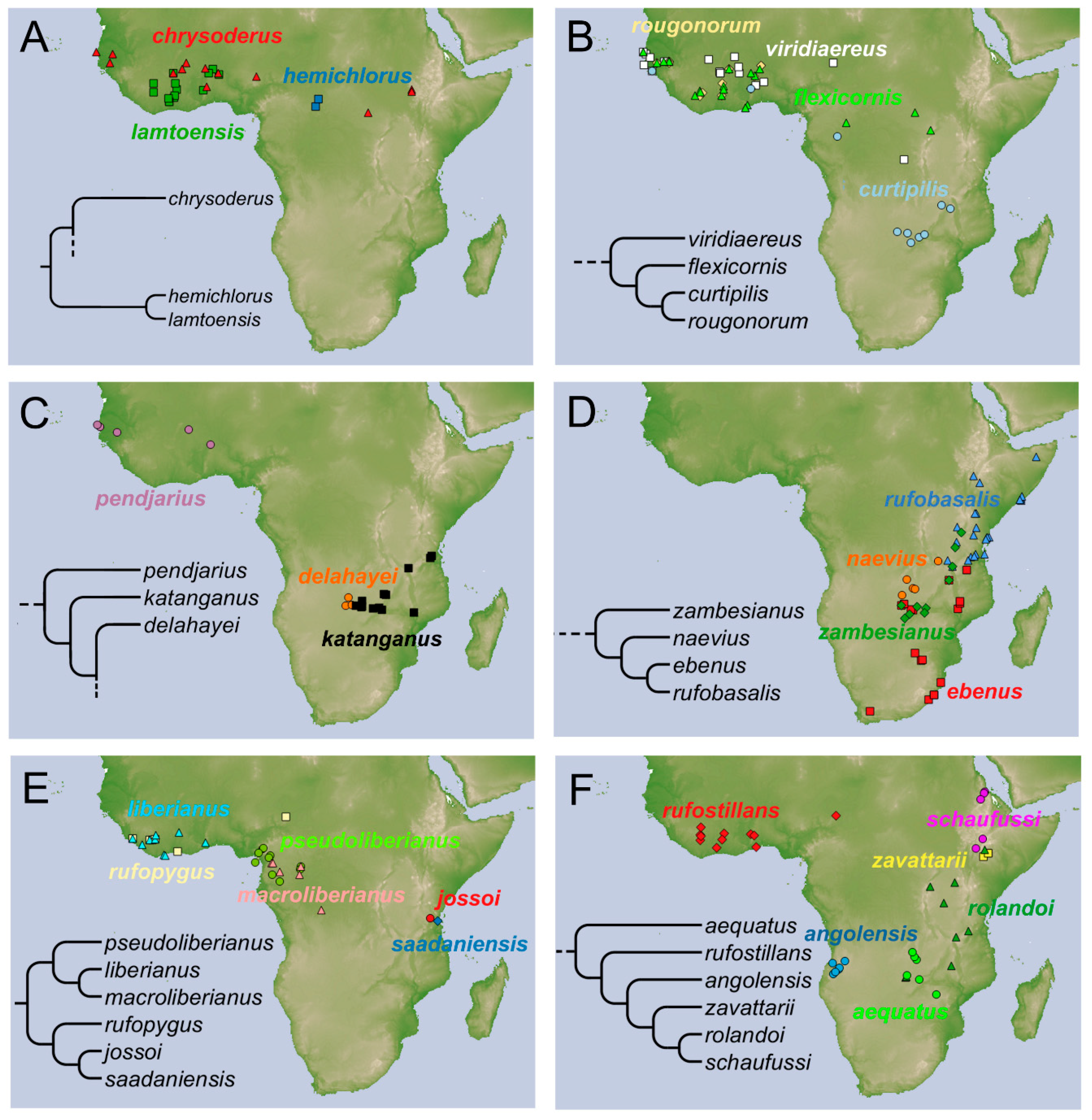
Distribution. The species is known from East and South Central Africa (
Figure 7F), extending from Tanzania, Malawi, and Zambia, until the
locus typicus in Zimbabwe.
Notes. In the SAMC a single specimen is housed, labelled as “type”. It is possible that other specimens had originally been included in the type series, since, in the original description, by Péringuey it is written “length 10–10 1/2 mm”, which likely means that the author had more than one specimen. Furthermore, in the last sentence of the description, there is written “In both this.... the punctures or the granules... are occasionally less numerous in the second interval”, meaning that there was at least another type specimen in the Péringuey collection. Thus, here we can account only for the lectotype, while any paralectotype has not yet been found.
Type material. Holotype. ♂ ‘ANGOLA, Huila, 3.5 km SW Negola, 14°08′53″ S, 14°28′16″ E 29.xi.2012 open forest, human shit P. Schüle leg. #10’ ‘Tiaronthophagus angolensis n.sp. holotype’ (BMNH). Paratypes. 7♂ 5♀ ‘ANGOLA, 8.xii.2012, Huila Prov. 3.5 km SW Negola, 14°28′16″ S, 14°08′53″ E, P. Schule leg. #22’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ 2♀ ‘ANGOLA, prov. Huila, 10 km SW Cacula, 4–6.xi.2011 P. Schule leg.’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ ‘5.xii.2012 env. Aldeia Cambala (S. Cacula) Huambo prov. Angola S. Rojkoff réc.’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ ‘6-xii-2012 PL 75 km N Cacula 13°26′192″ S, 15°22′755″ E, Huíla prov. Angola S. Rojkoff det.’ ‘Onthophagus cfr schaufussi Harold M. Dierkens det. 2013’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ 1♀ ‘6-xii-2012 PL 75 km N Cacula 13°26′192″ S, 15°22′755″ E, Huíla prov. Angola S. Rojkoff det.’ ‘Tiaronthophagus angolensis n.sp. paratype’ (MDCL); 2♂ 1♀ ‘29.xi.2012 PL 2 km S Negola 14°28′16″ E, 14°8′53″ S, 9.xii.2012 Huíla prov. Angola S. Rojkoff rec.’ ‘Tiaronthophagus angolensis n.sp. paratype’ (JFJC); 1♂ ‘ANGOLA, 19.xi.13, Malanje Prov. 20 km SE Cacandala, 1109 m 9°48′873″ S, 16°36′398″ E, P. Schule leg. #13’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 17♂ 7♀ ‘ANGOLA, Huila 3.5 km SW Negola 14°8′53″ S, 14°28′16″ E, 8.xii.2012, P. Schule leg. #22’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♀ ‘ANGOLA, Huila, 3.5 km SW Negola, 14°8′53″ S, 14°28′16″ E, 29.xi.2012 open forest, human shit P. Schüle leg. #10’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♀ ‘ANGOLA Huila, 75 km NE Caconda, 5-xii-2012, 13°26′19″ S, 15°22′55″ E, P. Schüle leg. #20/21’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ 1♀ ‘ANGOLA, 11–12.xi.2013, Huila Prov. 3.5 km SW Negola, 14°28′16″ S, 14°8′53″ E, P. Schüle leg. #4’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ ‘13.xi.13, Huíla Prov., near Nova Monção, 1489 m 13°30′292″ S 15°12′746″ E, P. Schule leg. WP 6 5 ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC); 1♂ 12.xii.2015, Huila prov, 3 km SW Nova Monção, 13°26.612 S, 5°21.881 E. G.Werner leg. ‘Tiaronthophagus angolensis n.sp. paratype’ (KWC): 1♂ 6♀ ‘Huila prov. 3.5 km SW Negola, 14°08′53″ S, 14°28′16″ E, 29.xi.2012 open forest, human shit P. Schule leg. #10’ ‘Tiaronthophagus angolensis n.sp. paratype’ (PMOC).
Other material. No other material is known at present.
Etymology. The species was named after the locality of the type material collection.
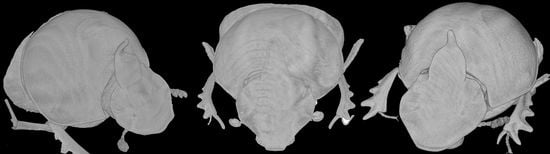
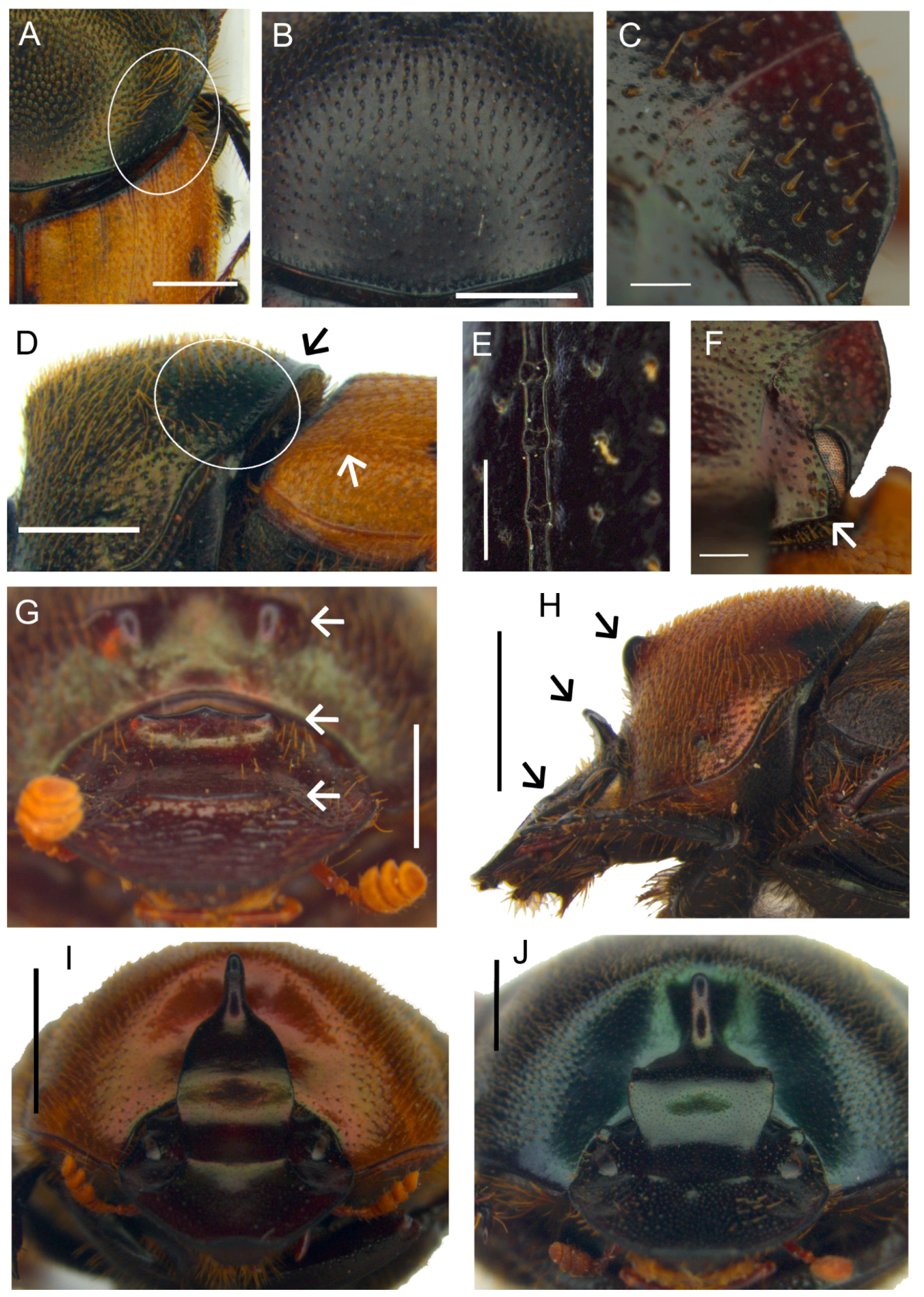
Description. Male (
Figure 8A,C): length 10–13 mm. Body black, mat, surface covered by short and thick testaceous pubescence, longer on sides and legs. Head surface smooth, with many small points, and some larger ones near the genae suturae; frontal carina barely visible in major males, rounded, and evident in minor males; genae rounded, not much developed; vertex carina in major male lamina shaped, elongate, with a digitiform expansion apically, in minor male reduced to a short expansion truncated at apex. Antennae yellowish brown. Pronotum ovalar, enlarged, covered with granulate setigerous points, granules large and rounded, base with a row of large points, anteriorly a large, concave area in major males, and two triangular protuberances on disc in minor males. Elytra striae geminate, interstriae covered by setigerous points that are regularly spaced on the surface. Pygidium black, mat with dense, superficial puncture. Legs black with long, testaceous setae.
Female (
Figure 8B,D): length 11–12 mm. Body black and mat as in male. Head surface wrinkled with some few large points near the genal suturae on clypeus, smooth with many small simple points on the remaining surface; frontal carina rounded, evident; genae rounded, not developed; vertex carina tricuspid, large and low, rectilinear from above view. Pronotum granulation as in male, with two evident conical tubercles on disc. Elytra and legs as in male. Pygidium is shorter than in male, but the same colour and puncture.
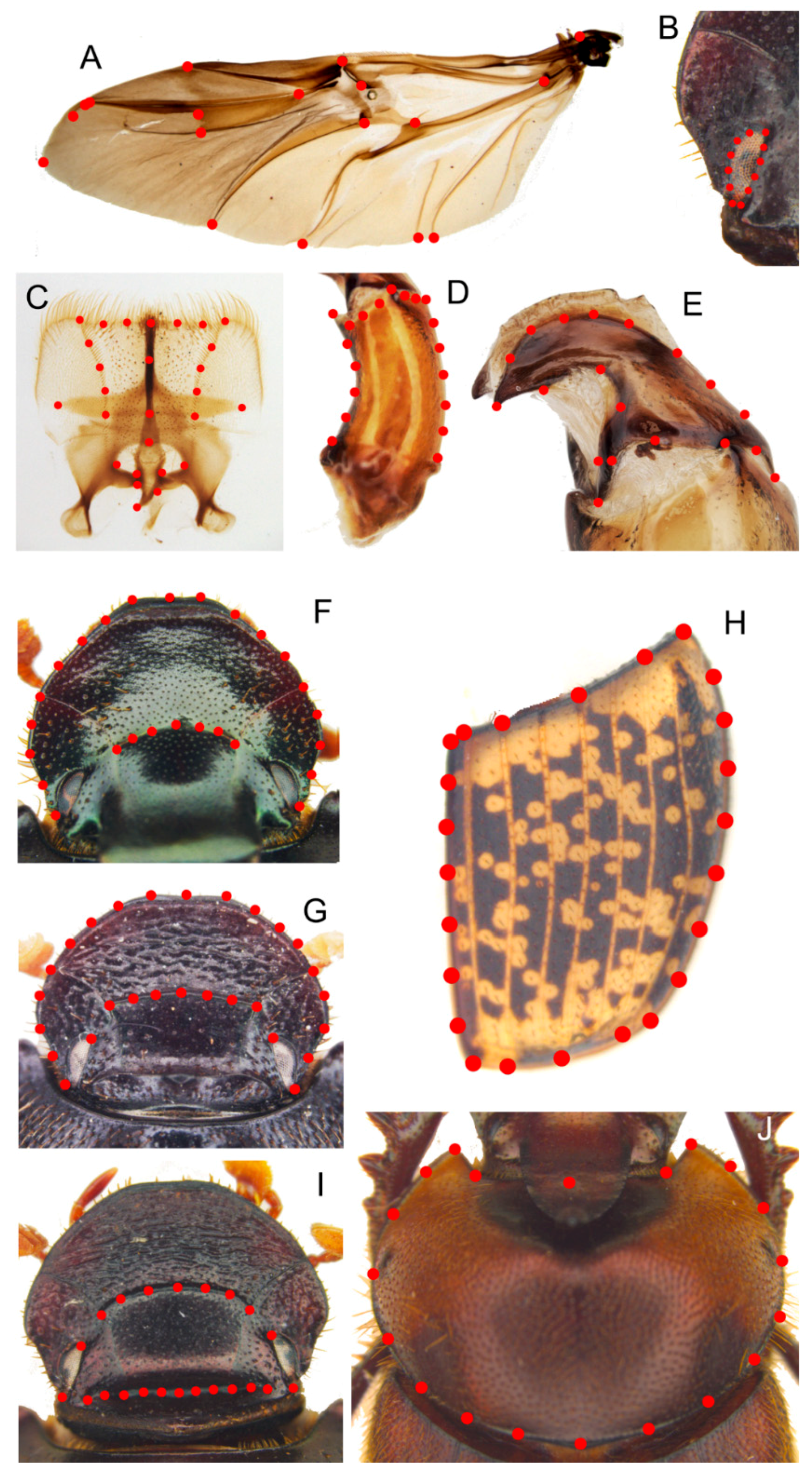
Epipharynx (
Figure 8E) fore margin slightly concave, with evenly long setae, chaetopariae angular, with setae very short in basal half; anterior epitorma rod-shaped, thin, well sclerotized; the zygum is constituted by long setae, proplegmatium triangular, low, the sides thick and rounded; apotormae present, thin, rod-like; plegmatic area present, ovalar; pternotormae short, thick and rectilinear; laeotorma and dexiotorma symmetrical, short; crepis only slightly asymmetrical, being pointed at apex.
Male genitalia (
Figure 8F,G,I): phallobase cylindrical, with a rectangular expansion ventrally; parameres symmetrical, tapering to apex, arched, the base diameter smaller than phallobase; endophallus with a large denticulate area, and an evident raspula being constituted by long, thin setae; lamella copulatrix present, constituted by two well-sclerotized parts (
Figure 8I); accessory lamellae present, well-developed (
Figure 8G).
Female genitalia (
Figure 8H): vagina domed, membranous; infundibular wall reversed U-shaped, apically squared; well-sclerotized infundibulum question mark that is shaped toward the receptaculum seminis, rectilinear in central portion, and upward turned toward the ovarioles; receptaculum seminis not much expanded, lobate, tapering to apex carrying a small, rounded laminal claw, the desclerotized area small and near the apex.
Distribution. The species is known only from Angola (
Figure 7F).
Onthophagus chrysoderus d’Orbigny, 1905a: 490.
Type material. Holotype. ♀ (by monotypy), ‘Sénégal’ ‘chrysoderus n.sp. D’Orbigny’ ‘Holotype Onthophagus chrysoderus d’Orbigny, 1905 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus chrysoderus (d’Orbigny, 1905) Roggero, Moretto, Barbero, Palestrini 2019’ (BMNH).
Other material. BENIN. North Eastern, E of Kandi, around Saa (PMOC). BURKINA FASO. Bobo Diulasso, Farako Ba (PMOC); Mogtedo, piste de Bomboré (PMOC); Passoré 8 km SW Yako 320 m (PMOC); Sanguié, Forêt de Sorobouli, 270 m (PMOC). DEMOCRATIC REPUBLIC OF CONGO. Haut Uele env., Doruma (NHMW). ETHIOPIA. Dembi (= Dembidolo), Illubabor (PMOC); Gambela (PMOC). GUINEA. Région Kindia, Mt Gangan, 500 m (BMNH). GUINEA BISSAU. Coli (Quebo), Tombali (CAS). NIGERIA. north Lama Burra Game Reserve 630 m. (PMOC). SENEGAL. Kolda, Mahon Forêt de Bakor (PMOC); Sebikotane (MNHN). TOGO. Sokodé (NHMW).
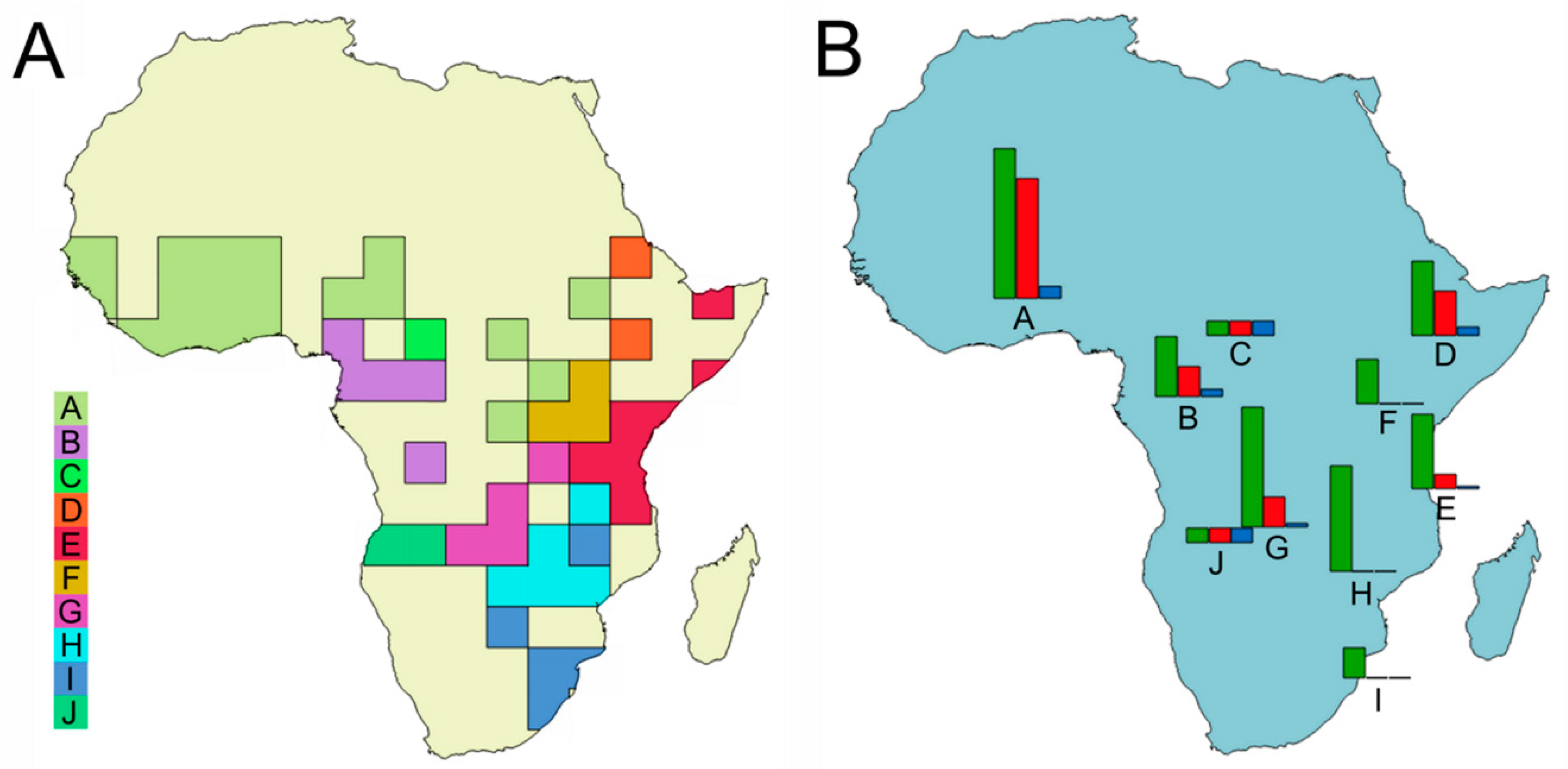
Distribution. The species is known from the Western and Central Africa (
Figure 7A).
Onthophagus curtipilis d’Orbigny, 1905b: 440.
= Onthophagus altidorsis d’Orbigny, 1905b: 443. Josso and Moretto, 2018: 11.
Type material. Holotype. ♀ (by monotypy) ‘Guinea Portoghese Bolama vi–xii.1899. L.Fea’ ‘Typus’ ‘curtipilis n.sp. D’Orb. <handwritten by d’Orbigny>’ ‘curtipilis d’Orb. <handwritten>’ ‘Holotypus Onthophagus curtipilis d’Orbigny, 1905’ ‘Mus. Civ. Genova’ ‘Tiaronthophagus curtipilis (d’Orbigny, 1905) Roggero, Moretto, Barbero, Palestrini 2019’ (MSNG).
Synonymy type material. O. altidorsis. ♂ (holotype by monotypy) ‘Guinea Portoghese Bolama vi–xii.1899. L.Fea’ ‘Typus’ ‘altidorsis n.sp. D’Orb. <handwritten by d’Orbigny>’ ‘altidorsis d’Orb. <handwritten>’ ‘Holotypus Onthophagus altidorsis d’Orbigny, 1905’ ‘Mus. Civ. Genova’ ‘Tiaronthophagus curtipilis (d’Orbigny, 1905) Roggero, Moretto, Barbero, Palestrini 2019’ (MSNG).
Other material. BURKINA FASO. Sanguié, Boromo, Ft. de Sorobouli; Ouagadougou, la forêt; Bobodiolasso, Farako-Bâ (PMOC). CAMEROON. N.W. Province, Bankim (PMOC). CENTRAL AFRICAN REPUBLIC. Ombella-Mpoko Prov., 20 km N.W. Yaloke (PMOC). GHANA. Kyabobo N.P.; Great Accra Region, Shai Hills (PMOC). GUINEA BISSAU. Coli (Quebo), Tombali (CAS). IVORY COAST. Comoé, Zamou; P.N. du Mt. Sangbé; Korhogo, Koko; Touba, Biémasso; Odienné, Massif du Denguélé (PMOC). MALAWI. Northern, Vwaza Marsh Reserve, Rumphi district, 1150 m (JFJC). REPUBLIC OF THE CONGO. Odzala N. P. (JFJC). SENEGAL. Tambacounda, Niokolo-Koba N.P.; Kolda, Mahon, Forêt de Bakor; Kédougou, 5 km S. Ségou. (PMOC). TANZANIA. Mwanza region, Geita (JFJC); Morogoro Reg., Mbarika Mts. (Ikongwa) (PMOC). TOGO. Fazao (NHMW). ZAMBIA. 17 km N Choma I. Bruce-Miller farm (JFJC); Kafue N. P., Kacheleko wildlife outpost (JFJC); Kafue river east (JFJC); Lower Zambesi N. P., 3 km Mukamba gate (JFJC); Southern, Mwense Muchinga 3620Ft. (JFJC); Western, 14 km SW Kaoma, 1180 m (JFJC).
Distribution. The species is widespread from Guinea Bissau (the locus typicus) to Togo and Zambia (
Figure 7B).
Notes. The species
T. curtipilis was described by a single female specimen, while
O. altidorsis was described by a single male specimen from the same locality [
42].
Onthophagus delahayei Josso, 2011: 4.
Type material. Holotype. ♂ ‘Zambie 18 km W Kaoma 14°52′0.3″ S, 24°38′29.4″ E, 9.xii-2008 JF Josso R Minetti leg’ ‘Exc. humains’ ‘holotype Onthophagus delahayei n.sp. J-F Josso det 2010’ ‘Tiaronthophagus delahayei (Josso, 2011) Roggero, Moretto, Barbero, Palestrini 2019’ (JFJC). Paratypes. 6♂ 6♀ ‘Zambie 18 km W Kaoma 14°52′0.3″ S, 24°38′29.4″ E, 9.xii-2008 JF Josso R Minetti leg’ ‘Exc. humains’ ‘paratype Onthophagus delahayei n.sp. J-F Josso det 2010’ ‘Tiaronthophagus delahayei (Josso, 2011) Roggero, Moretto, Barbero, Palestrini 2019’ (JFJC); 12♂ 9♀ ‘Zambie 18 km W Kaoma 14°52′0.3″ S, 24°38′29.4″ E, 14–15.xii-2009 Josso Juhel Minetti leg’ ‘Exc humains’ ‘paratype Onthophagus delahayei n. sp J-F Josso det 2010’ ‘Tiaronthophagus delahayei (Josso, 2011) Roggero, Moretto, Barbero, Palestrini 2019’ (JFJC) (PMOC) (PPRC) (IRSNB) (RMIC); 1♂ 1♀ ‘Zambie (E Mongu) piste de Ndanda 15°7′25.4″ S, 23°41′23.1″ E, 10-XII-2008 JF Josso R Minetti leg’ ‘Exc humains’ ‘Tiaronthophagus delahayei (Josso, 2011) Roggero, Moretto, Barbero, Palestrini 2019’ (JFJC); 4♂ 2♀ ‘Zambie 3 km NW Kandunda 1120 m 14°50′19″ S, 24°38′23″ E, 8-XII-2009 M. François leg.’ ‘Exc. humains’ ‘Tiaronthophagus delahayei (Josso, 2011) Roggero, Moretto, Barbero, Palestrini 2019’ (MFRC).
Other material. ZAMBIA. North Western province, 60 km N Kabompo (PMOC).
Distribution. At present the species is known only from Zambia (
Figure 7C).
Notes. The species was originally included in 16th group, which was based on the features of the pronotum posterior margin that is not wholly re-bordered on sides. However, the author also stated that the species is different from any other one of the group.
Onthophagus ebenus Péringuey, 1888: 97.
= Onthophagus natalicus d’Orbigny, 1902: 177. D’Orbigny, 1903: 296.
= Onthophagus bituber d’Orbigny, 1904: 289, n.syn.
Type material. Lectotype. ♂ (here designated) ‘Durban Natal <handwritten by Péringuey>‘ ‘Type SAM/Ent 2753b’ ‘Onthophagus ebenus Pe. type ♂ <handwritten by Péringuey>‘ ‘Lectotype Onthophagus ebenus Peringuey, 1888 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (SAMC). Paralectotypes. ♀ ‘Onthophagus ebenus Pe. type <handwritten by Péringuey>‘ ‘Kipibethe 1884 <handwritten by Péringuey>’ ‘Type SAM/Ent 6008’ ‘Imaged LAS 4.9 SAMC 2017’ ‘Paralectotype Onthophagus ebenus Peringuey, 1888 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (SAMC); ♀ ‘Durban Natal <handwritten by Péringuey>‘ ‘Type SAM/Ent 2753a’ ‘Onthophagus ebenus Pe. type ♀ <handwritten by Péringuey>‘ ‘Imaged LAS 4.9 SAMC 2017’ ‘Paralectotype Onthophagus ebenus Peringuey, 1888 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (SAMC).
Synonymy type material. O. natalicus. ♂ (lectotype) ‘Natal Dr Martin’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); (2 paralectotypes) ‘Natal Dr. Martin’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); (3 paralectotypes) ‘Durban, Natal’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); (paralectotypes) Transvaal, Makapan near Pietersburg (MNHN). O. bituber. ♀ (holotype by monotypy), “Botoka Feb. 98. <handwritten>” “bituber n.sp. d’Orb. <handwritten>” “d’Orbigny vidit 1904” “holotype” ‘Holotype Onthophagus bituber d’Orbigny, 1904 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus ebenus (Peringuey, 1888) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Other material. BOTSWANA. 10 km NE Martins Drift (PMOC). MALAWI. Liwonde Nat. Park (JFJC); Northern, Vwaza Marsh Reserve, Rumphi district, 1150 m (JFJC); Mzimba district, Vwanza Marsh Game res. 1150 m (JFJC); Blantyre Michiru Mt Park (PMOC); Southern, gorges de la Mpatamaga (JFJC). SOUTH AFRICA. KwaZulu Natal, 3 km S Nyamande Kwa Gudlumfula Mts. 350 m (JFJC) (MDCL); KwaZulu Natal, 10 km N Hluhluwe (PMOC); KwaZulu Natal, Durban (PMOC); KwaZulu-Natal, 4.5 km NW Gudlucingu Umzimkulu river (JFJC); Limpopo, Makopane 1395 m Shikwaru lodge (PMOC); Eastern Cape, 3 km N Qhaka, 138 m near Lusikisiki (JFJC). TANZANIA. Morogoro region, 360 m, Mbarika Mts, (Ikongwa) between Kichangari and Idunda (PMOC). ZAMBIA. Kafue NP, Chunga (JFJC); Kafue NP, Mayukuyuku Camp (JFJC); Magoye (PMOC); Kafue env. (PMOC); Kafue (MDCL); Southern province, Kafue, Chiron du Mts (MDCL); Southern province, 11 km E Mukuni, 960 m (MDCL); Southern province, Mosi Oa Tunya NP (JFJC).
Distribution.Onthophagus ebenus was described by Vaal River (Mpumalanga), as quoted by Péringuey, while
Onthophagus natalicus was described from Natal, Durban, Transvaal, and Makapan near Pietersburg. Besides, in the SAMC there are three specimens labelled as ‘
Onthophagus ebenus type’. The species is widespread in Southeastern Africa (
Figure 7D) from Tanzania (Morogoro region) to South Africa (KwaZulu Natal).
Notes. The synonymy of T. ebenus and O. natalicus was confirmed by the survey of the typical material of the latter species. The comparison of the type material of O. ebenus and O. bituber, shows that the specimens belong to a single species. Thus, we propose the new synonymy. The type locality of T. ebenus was defined according to the ICZN (articles 74E, 76.2).
Onthophagus flexicornis d’Orbigny, 1902: 178.
= Onthophagus atroaereus d’Orbigny, 1908: 164 n.syn.
Type material.Lectotype. ♂ (here designated) ‘Accra <written by hand> Ex. Musaeo H.W. Bates1892’ ‘flexicornis n.sp. D’Orb. <d’Orbigny handwritten> ‘d’Orbigny Ontho. Afr. 1902’ ‘Muséum Paris 1952 Coll. R.Oberthür’ ‘LECTOTYPE’ ‘Lectotype Onthophagus flexicornis d’Orbigny, 1902 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus flexicornis (d’Orbigny, 1902) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN). Paralectotypes. 1♀’Accra W. Afr. <written by hand> Ex. Musaeo H.W.Bates 1892’ ‘d’Orbigny Ontho. Afr. 1902’ ‘Muséum Paris 1952 Coll. R.Oberthür’ ‘PARALECTOTYPE’ ‘Paralectotype Onthophagus flexicornis d’Orbigny, 1902 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus flexicornis (d’Orbigny, 1902) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); 1♀ ‘Accra <written by hand> Ex. Musaeo H.W.Bates 1892’ ‘Muséum Paris Coll. H. d’Orbigny 1915’ ‘PARALECTOTYPE’ ‘Paralectotype Onthophagus flexicornis d’Orbigny, 1902 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus flexicornis (d’Orbigny, 1902) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Synonymy type material. O. atroaereus. ♀ (holotype by monotypy) ‘Guinée Portug. Chime 1906 Rio Geba G.Favarel’ ‘atroaereus n.sp. D’Orb. <d’Orbigny handwritten>‘ ‘d’Orbigny Ontho. Afr. 1908’ ‘Muséum Paris 1952 Coll. R.Oberthür’ ‘HOLOTYPE’ ‘Tiaronthophagus flexicornis (d’Orbigny, 1902) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Other material. BENIN. Northern, Parc Pendjari, Tanoungou (JFJC); Parc Pendjari, vers hotel Pendjari (JFJC). BURKINA FASO. Kompienga Pama, 230 m (PMOC); Sanguié Boromo forêt de Sorobouli 270 m (PMOC); Park W, 7km Koudou (vers Kondjo) (JFJC). CENTRAL AFRICAN REPUBLIC Bozo (PMOC); North, Parc de la Sangha (MHNL). DEMOCRATIC REPUBLIC OF CONGO. Haut-Uele, Moto (MNHN); Haut-Uele env. Doruma (NHMW). GHANA. Great Accra Region, Hills north end, 125 m (PMOC). GUINEA. Nimba (MNHN). IVORY COAST. Zamou (PMOC); Comoé, Zamou (PMOC). Korhogo Koko 347 m (PMOC); PN Comoé (MHNL); Touba (PMOC); Touba, Biémasso-Dolla (PMOC); Touba, Dolla (PMOC); Youho (MHNL). SENEGAL. Niokolo Koba NP, Niokolo (PMOC); Kolda Mahon, Forêt de Bakor (PMOC); Niokolo-Koba NP, poste de Niokolo 65m (PMOC); Niokolo-Koba NP poste de Siminti (PMOC); Sangalkam (MNHN); Sebikotane (MNHN); Tambacounda Botou, 52 m (PMOC).
Distribution. The species is widespread in Western Africa until the Democratic Republic of Congo (
Figure 7B).
Notes. The comparison of the type material of the two species, in addition with series of recently collected material shows that all of the specimens belong to a single species. Thus, we propose the new synonymy.
Onthophagus hemichlorus (d’Orbigny, 1915: 392.
Type material. Holotype. ♂ (by monotypy) ‘haut Chari Fort-Crampel coll. Felsche’ ‘Tiaronthophagus hemichlorus (d’Orbigny, 1915) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN, not yet traced).
Other material. CENTRAL AFRICAN REPUBLIC. Kemo prov., 45 km N Sibut, 550 m (JFJC).
Distribution. Described from Haut-Chari, Fort-Crampel (now, Kaga-Bandoro) in Central African Republic (
Figure 7A).
Notes. The species was described from a single specimen [
46] after the publication of the Synopsis [
2] and included in the 24th species-group. The species is given, as strictly related to
T. schaufussi and
T. ebenus.
Type material. Holotype. ♂ ‘Tanzanie, VII.2005 Nguu Mts., Tamota Fst. Local coll. > R. Minetti’ ‘Tiaronthophagus jossoi n.sp. holotype’ (BMNH). Paratypes. 3♂, 3♀ ‘Tanzanie, VII.2005 Nguu Mts., Tamota Fst. Local coll. > R. Minetti’ ‘Tiaronthophagus jossoi n.sp. paratype’ (PMOC); 5♂ ‘Morogoro Region Udzungwa National Park campsite 2, forêt, altitude 390 m., 7°50.743′ S, 36°52.584′ E, ‘Tiaronthophagus jossoi n.sp. paratype’ (PMOC).
Other material. No other material is known at present.
Etymology. The species was named after our colleague, the French entomologist J.F. Josso.
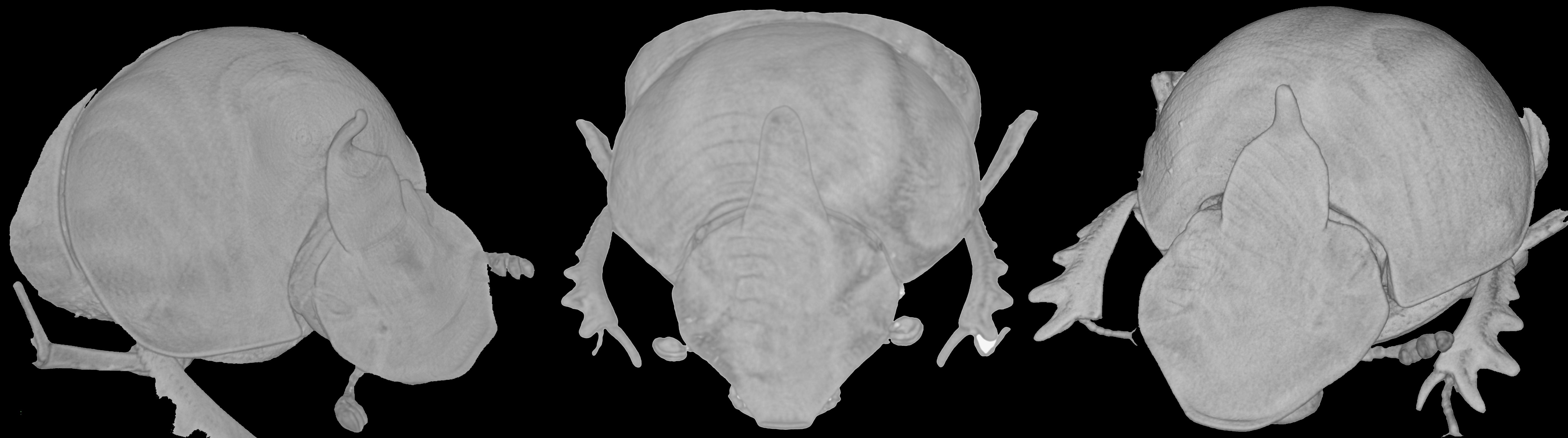
Description. Male (
Figure 9A,C): length 11–14 mm. Body dark reddish brown with some symmetrical testaceous patches on elytra, relatively flat, mat, densely granulate, with a long testaceous pubescence on the whole surface. Head pentagonal, fore margin of the clypeus upward-turned, surface that is covered by many very thick large points, which are fewer and mixed to smaller ones in minor males, genae not much developed, squared; frontal carina being barely visible in major males, almost rectilinear; vertex carina in major male modified into a long, flat rectangular lamina with apical sides elongate and sharp, digitiform expansion that is not much developed, rectilinear, while in minor male is a small triangular expansion. Eyes slightly rounded and medium sized. Antennae yellowish brown. Pronotum with small dense granules, anterior angles with a testaceous longitudinal patch, base with a dense row of large points. Elytra rounded, wider than pronotum, with two symmetrical patches on the third and 5–6th interstriae at an anterior margin near the humeral callus and a larger one at posterior margin of elytra. Pygidium greenish brown, with a thick, long yellow pubescence, rounded small relatively dense granules on whole surface. Legs reddish brown with long, thick testaceous setae.
Female (
Figure 9B,D): length 12–14 mm. Body reddish brown as in male. Head pentagonal, but more rounded, and less developed anteriorly than in males; the whole surface being covered by a rough granulation, with many large points; frontal carina evident, elevated, almost rectilinear; genae not much expanded, rounded; vertex carina small, short, triangular shaped. Pronotum with two symmetrically large expansions, triangular lamina shaped on side view, encircling a smooth, concave narrow area without pubescence. Elytra and legs as in male. Pygidium is shorter than in male, but the same colour and puncture.
Epipharynx (
Figure 9E) fore margin rectangular shaped, acropariae with a thick long pubescence, zygum constituted by a thick tuft of setae; anterior epitorma thin but well-sclerotized, basal part enlarged, drop shaped, proplegmatium triangular-shaped, lowered, sides thick and rounded; apotormae present, triangular-shaped; plegmatic area ovalar, small; pternotormae thick, short, downturned; laeotorma and dexiotorma short, thin, symmetrical; crepis well developed, apex left-turned; chaetopariea rectilinear, the setae longer in the distal half, then very small; haptomerum with short thick setae that are mixed to very short, thinner ones.
Male genitalia (
Figure 9F,G,I): phallobase short and large, with a rectangular expansion anteriorly; parameres with triangular apices; endophallus raspula that are constituted by a fringe of very long thin setae, lamella copulatrix present, saddle shaped (
Figure 9I); accessory lamellae present, well-developed.
Female genitalia (
Figure 9H): vagina dome-shaped, with two spherical expansions that are covered by thick and short setae on sides; infundibular wall with a semiovalar sclerotization; infundibulum question mark shaped, the part toward the ovarioles upward turned; receptaculum seminis lobate, tapering to the apex, the small sclerotized area near the apex.
Distribution. The species is known at present from Tanzania (
Figure 7E).
Type material. Holotype. ♂ ‘R. D. Congo, Katanga, Mwera i/ii.2001 coll. T. Bouyer’ ‘Onthophagus aequatus Peringuey J.L Nicolas det. 2001 <handwritten>’ ‘aequatus Per. <handwritten>’ ‘Tiaronthophagus katanganus n.sp. holotype’ (BMNH). Paratypes. 2♀ ‘R. D. Congo, Katanga, Mwera i/ii.2001 coll. T. Bouyer’ ‘Tiaronthophagus katanganus n.sp. paratype’ (PMOC); 1♂ ‘R. D. Congo Katanga Kasumbalesa 13.xii.01 coll. T. Bouyer’ ‘Tiaronthophagus katanganus n.sp. paratype’ (JFJC); 1♀ ‘R. D. Congo Katanga Lubumbashi 20-xi/13-xii-2002 coll. T. Bouyer’ ‘Tiaronthophagus katanganus n.sp. paratype’ (PMOC); 1♂ 2♀ ‘Zambie, 18.xii.2006 Lusaka C.diGennaro leg.’ ‘Tiaronthophagus katanganus n.sp. paratype’ (PMOC); 1♂ ‘Zambie, 14.xii.2006 Lusaka C. di Gennaro leg.’ ‘Tiaronthophagus katanganus n.sp. paratype’ (PMOC); Zambia 11.xi.2002 50–150 km S Kasempa Werner leg.’ ‘Tiaronthophagus katanganus n.sp. paratype’ (PMOC); 2♀ ‘Zambie NW Prov. 30km N Lubungo NE Kafue NP 10-11xii2012 14°11′ S, 26°29′ E, 3775Ft leg. Josso Juhel Minetti’ ‘lumiere’ ‘Tiaronthophagus katanganus n.sp. paratype’ (JFJC); 2♀ ‘22-xi-2003 Zambia N W prov.’ ‘Mufumbwe to Kasempa Werner et SMRZ’ ‘Onthophagus rufobasalis Fairmaire M. Dierkens det. 04’ ‘Tiaronthophagus katanganus n.sp. paratype’ (MDCL); 1♂ ‘Zambie Monze 7-10-xi-2010 J-F Josso leg’ ‘Tiaronthophagus katanganus n.sp. paratype’ (JFJC).
Other material. No other material is known at present.
Etymology. The species was named after the collection locality of the holotype.
Description. Male (
Figure 10A,C): length 10–11 mm. Body blackish brown, which is covered by a short testaceous pubescence, with two large testaceous patches at base and apex of elytra, extending from elytral sutura to side margin. Head surface smooth covered by dense small points mixed to larger ones near the genal suturae; frontal carina sometimes almost inapparent in major males, slightly curved, well-developed in minor males; genae not much developed, rounded; vertex carina lamina-shaped, slightly rounded on sides with the digitiform apex relatively short and rectilinear in major male, short expansion with the superior edge rounded large one third of the width of the head base in minor male. Medium sized, slightly rounded eyes. Antennae yellowish brown. Pronotum that is covered by dense granulate setigerous points, the granules large, and rounded on disc, and larger, thicker, and ovalar on sides; in major males, a large, smooth concave area in anterior part; base with very large points very close. Elytra testaceous and blackish brown, the interstriae covered by regularly spaced setigerous points with small granules. Pygidium black, with large, thick points. Legs blackish brown with short, thick testaceous setae.
Female (
Figure 10B,D): length 10–11 mm. Body as in male. Head rounded, clypeus surface rough, with some large points that are near the genal carinae; genae small, rounded, with many large points; frontal carina arched, well-developed, and bulging; vertex carina large, almost reaching the eyes, thick not much elevated, rectilinear; the area between the carinae smooth, concave, with only few small simple points. Pronotum with setigerous granulate points as in male, and two well-developed, very close, symmetrical, anteriorly rounded, and flat expansions on the disc. Elytra and legs as in male. Pygidium shorter than in male, but same colour and puncture.
Epipharynx (
Figure 10E): fore margin rectilinear with evenly long, thick setae; zygum constituted by a tuft of long setae; chaetopariae shorter on the proximal half, longer near acropariae; anterior epitorma thin but well-sclerotized, enlarged at base; proplegmatium triangular shaped, low, tapering on sides; apotormae present, rod-shaped, thin; pternotormae short, thick, downturned; laeotorma and dexiotorma symmetrical, short; crepis asymmetrical, well-developed, left-turned; haptomerum covered by many short thin setae, with few being longer on chaetopedia.
Male genitalia (
Figure 10F,G,I): phallobase cylindrical, with an anterior rectangular expansion; parameres symmetrical, arched, tapering to the sharp apex; endophallus with a evident raspula with long, thin setae, lamella copulatrix (
Figure 10I) present, which is constituted by two distinct parts; accessory lamellae present, well-developed.
Female genitalia (
Figure 10H): vagina membranaceous, dome-shaped with a squared, quite elongate infundibular wall; infundibulum question-mark shaped, rectilinear, thick, distal part to ovarioles upward turned; receptaculum seminis not much expanded, lobate, tapering to apex carrying a small, rounded laminal claw, the desclerotized area small and near the apex.
Distribution. At present, the species is only known from a circumscribed area between the Democratic Republic of Congo and Zambia (
Figure 7C).
Onthophagus lamtoensis Cambefort, 1984: 7.
Type material. Holotype. ♂ ‘P139 <Côte d’Ivoire, Lamto, 5-6.7.1980, Piste du Grand Sud, EH, Y.Cambefort leg.’ ‘holotype’ ‘Tiaronthophagus lamtoensis (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN). Paratypes. 550 specimens from Lamto, Abokouamékro, Ouango Fitini, vii.1979-vii.1981 ‘Tiaronthophagus lamtoensis (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN)
Other material. BENIN. North Eastern, E of Kandi, 4 km W of Bensekou (PMOC); Northern, Parc Pendjari vers Tanoungou (JFJC). BURKINA FASO. Bobo Diulasso, Farako Ba (PMOC); Park W, 7 km Koudou (JFJC). GHANA. Kyabobo NP, near Laboum outpost (PMOC). IVORY COAST. Zamou (PMOC) (MNHN); Haut Nzi (MNHL); Korhogo Koko 347 m (PMOC); Odienné (PMOC); Touba Biémasso-Dola (PMOC); Zamou (Comoé) (MNHN).
Distribution. The species shows a Western African distribution (
Figure 7A).
Notes. The species was included in the 24th group [
47] and it was considered very close to
T. ebenus.
Onthophagus liberianus Lansberge, 1883: 15.
Type material. Holotype. ♀ (by monotypy) ‘Liberia Sala & Büttikofer leg.’ ‘Holotype Onthophagus liberianus Lansberge, 1883 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus liberianus (Lansberge, 1883) Roggero, Moretto, Barbero, Palestrini 2019’ (RMNH).
Other material. BENIN. Lama (MDCL). GUINEA Nzérékore Ft. Classée de Diecké (PMOC). IVORY COAST. Sassandra, Pauly-Brousse (PMOC) (MDCL); W Sassandra (PMOC) (MDCL); Man, Mt. Tonkoui 1200 m (PMOC) (MDCL); San-Pédro, Monogaga (PMOC); Touba, Biémasso 441 m (PMOC); Comoé, Zamou (PMOC). SIERRA LEONE. 120 m Tiwai island, Moa river (PMOC). TOGO. 10 km N Kpalimé reg. de Missahohe, ft. de Demetui (EBCT).
Distribution. The species has a Western African distribution (
Figure 7E).
Onthophagus macroliberianus Moretto, 2010: 465.
Type material. Holotype. ♂ ‘Centrafrique Bayanga 16.III–6.IV.96 <handwritten>’ ‘pieges excr. humains <handwritten>’ ‘P. Moretto leg. <handwritten>’ ‘Photo description Moretto 2010 <handwritten>’ ‘Holotype Onthophagus macroliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus macroliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC). Paratypes. 2♀ ‘Cameroun, Obout, ii.2007 local coll. >D. Moore’ ‘Paratype Onthophagus macroliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus macroliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 1♀ ‘Cameroun Centre, XI.1996 Mt. Kala M. Desfontaine leg.’ ‘Paratype Onthophagus macroliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus macroliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 1♀ ‘C Dewalsche Ipamu Kasai’ ‘Paratype Onthophagus macroliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus macroliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); ‘forêt 40 km S/O2-3-83 <handwritten>’ ‘Allotype Onthophagus macroliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus macroliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC).
Other material. REPUBLIC OF CONGO. Mayoko, 28km NE of Pointe Noire, 663m (PMOC).
Distribution. The species is located in the Central Africa (
Figure 7E), and is rare in collections.
Onthophagus naevius d’Orbigny, 1913: 403.
Type material. Holotype. ♀ (by monotypy) ‘150–200 miles W.of Kambove. 3500–4500 ft. 23.10.07’ ‘Neave Coll. 1907-230.’ ‘Type’ ‘naevius n.sp. d’Orb. <handwritten by d’Orbigny>‘ ‘Holotype Onthophagus naevius d’Orbigny, 1913 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus naevius (d’Orbigny, 1913) Roggero, Moretto, Barbero, Palestrini 2019’ (BMNH).
Other material. TANZANIA. Rukwa Region, Mbizi Mts. Fst. Res., 2252 m (PMOC). ZAMBIA. 50–150 km S Kasempa (PMOC); NorthWestern prov. 80 km W Chingola (PMOC); North Eastern prov. Chimfunsi Wildlife reserve (PMOC).
Distribution. The species is found in Central Africa (
Figure 7D). It is extremely rare in collections.
Onthophagus pendjarius Josso and Prévost, 2006: 4.
Type material. Holotype. ♂ ‘N BENIN Parc Pendjari vers hotel Pendjari 1-3-vi-2005 Josso Juhel Monfort leg.’ ‘lumiere’ ‘holotype Onthophagus pendjarius n.sp. Josso et Prévost det. 2006’ ‘Tiaronthophagus pendjarius (Josso and Prévost, 2006) Roggero, Moretto, Barbero, Palestrini 2019’ (JFJC). Paratypes. 1♀ ‘N BENIN Parc Pendjari vers hotel Pendjari 1-3-vi-2005 Josso Juhel Monfort leg.’ ‘lumiere’ ‘paratype Onthophagus pendjarius n.sp. Josso et Prévost det. 2006’ ‘Tiaronthophagus pendjarius (Josso and Prévost, 2006) Roggero, Moretto, Barbero, Palestrini 2019’ (JFJC); 2♂ ‘Burkina Faso, 8.viii.2005 Loroum, Toulfé, 330 m, 13°54′16″ N, 1°54′19″ W, O, piège excréments humains, P. Moretto leg.’ ‘paratype’ ‘Onthophagus pendjarius n.sp. Josso Prévost det. ix.2006’ ‘Tiaronthophagus pendjarius (Josso and Prévost, 2006) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC).
Other material. SENEGAL. Fatick Diouroup (PMOC); Gouloumbou (PMOC); Sebikotane (MNHN).
Distribution. The species was collected from Western Africa (
Figure 7C).
Notes. Although the species was formerly included in the 16th group [
50], the similarities with the 24th and 28th groups were already highlighted in the original description.
Onthophagus pseudoliberianus Moretto, 2010: 466.
Type material. Holotype. ♂ ‘Congo (Gabon) Ogooué, Lambaréné, 1911–1912–1913 Ellenberger’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN). Paratypes. 1♂ ‘Cameroun Bafia 600m i.1991 Ly? <handwritten>’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘World Scarab Database WSD00257398’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 1♀ ‘Gabon Woleu-Ntem Oyem ix.2000 >P. Oremans’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘World Scarab Database WSD00257416’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); Congo (Gabon), Ogooué, Lambaréné, 1911–1912–1913 Ellenberger ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); 7♂ 2♀ ‘Centrafrique Bayanga 16.iii 6.iv.96 <handwritten>’ ‘pieges iules <handwritten>’ ‘P. Moretto leg. <handwritten>’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 5♂ 6♀ ‘Centrafrique Bayanga, 16.iii 6.iv.96 ‘pieges iules P. Moretto leg.’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 4♂ ‘Cameroun, N W Province Région de Bamenda, 1–3.vii.2008 Local. coll Voma eg.’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 3♂ 6♀ ‘Cameroun, N W Province Région de Bamenda, Bali, 5–16.vii.2006 Local. coll Jomia leg.’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 4♂ 2♀ ‘Cameroun N W Province Région de Bamenda, Bali-Nyonga 28.viii-1.ix.2006. Local. coll Jomia leg.’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 1♀ ‘O. liberianus… J.L. Nicolas det.1994 <handwritten>’ ‘Cameroun N Sanaga Ndom ii.93 J.C. Thibaud leg. <handwritten>’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); 2♂ ‘Africa Cameroon SWP Bali 28-vii-05 #19 V.L. Voma Human Feces Cornell Voma Korup NP Exped’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Paratype Onthophagus pseudoliberianus n.sp. P. Moretto det. 2009’ ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); Gabon Woleu Ntem Oyem ix.2000 P. Oremans (PMOC); Cameroun, mount Cameroun, Bafia 600m i.1991 ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC); Cameroun Bafia 600m i.91 ‘Tiaronthophagus pseudoliberianus (Moretto, 2010) Roggero, Moretto, Barbero, Palestrini 2019’ (PMOC).
Other material. CAMEROON. W region, Lebialem division, Wabane sub.div., Folepi village (PMOC); NW province, Region de Bamenda Bali (PMOC). CENTRAL AFRICAN REPUBLIC. N.P. Ndoki Lac (PMOC). EQUATORIAL GUINEA. Island Fernando Poo (now, Bioko), Musola. 500–800 m.s.m. (MSNG). GABON. E Ogoouè, Ivindo province, Makokou SW Ivindo N.P., Ipassa forest research station (PMOC).
Distribution. The species is known from the Central Western Africa (
Figure 7E).
Type material. Holotype. ♂ ‘Tanzania, Manyara, 1660 m, Ngorongoro N.W. 3°19′39 S, 35°36′19 E, 25.II–14.III.2008 T. & M. Ströhle leg.’ ‘Tiaronthophagus rolandoi n.sp. holotype’ (BMNH). Paratypes 1♂ 1♂ ‘Tanzanie, Mbeya Prov. Usangu Flats 1139m savane à épineux, 21.ii, 2004 R. Minetti leg.’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (PMOC); 1♀ ‘Tanzanie Mwanza reg. Geita iv-2011. Minetti leg.’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 2♀ ‘Tanzania, Manyara, 1660m, Ngorongoro N.W. 3°19′39 S, 35°36′19 E, 25.II–14.III.2008 T. & M. Ströhle leg. ‘Tiaronthophagus rolandoi n.sp. paratype’ (PMOC); 1♀ ‘Tanzanie 21.III-10.IV.2007. Manyara, Ngorongoro, NW. Karatu 1660m 3°19′39 S, 35°36′19″ E, Ströhle leg. ‘Tiaronthophagus rolandoi n.sp. paratype’ (PMOC); 1♂ 1♀ ‘N Malawi Chiwenga 1600 m 10–12.xii.2006 JossoJuhelMonfort leg.’ ‘poisson pourri’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 1♂ ‘N Malawi Chiwenga 1600 m 10–12.xii.2006 JossoJuhelMonfort leg.’ ‘cadavre iule’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 1♀ ‘Uganda Busia I.90 B. Wandora leg. <handwritten>’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (PMOC); 1♂ ‘Uganda 5.v.2009 Queen Elizabeth NP P. Malec lgt.’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 1♂ 1♀ ‘Uganda Queen Elizabeth NP 0°15’ S/30°00’ E ii.1996–iii.1997 leg. A. Hoffmann’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 1♂ ‘acquis en iii-2003 Kenya’ ‘Onthophagus rufobasalis Fairmaire J.L. Nicolas det. 03’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (MDCL); 1♂ 1♀ ‘4.XII.2012 PL. 1075m 6 km N Chunga 14°59′39″ S, 26°01′11″ E, Zambie Central Prov. S. Rojkoff réc.’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (PMOC); 1♂ 8-xii-205 40 km E Kapiri Mposhi Central Province Zambia S. Rojkoff & K. Werner rec.’ ‘Onthophagus cf rufobasalis Fairmaire M. Dierkens det. 2014’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (MDCL); 1♂ ‘Zambie Kafue NP Chunga 15°02,362’ S, 25°59,437’ E, 11–12-xii-2009 Josso Juhel Minetti leg.’ ‘exc. humains’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 3♂ ‘Zambie Kafue NP Mayukuyuku Camp 13-xii-2009 Josso Juhel Minetti leg.’ ‘lumiere’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC); 1♂ 2♀ ‘Zambie NW Prov. 30km N Lubungu NE Kafue NP. 10-11xii2012 14°11’ S, 26°29’ E, 3775Ft. leg. Josso Juhel Minetti’ ‘lumiere’ ‘Tiaronthophagus rolandoi n.sp. paratype’ (JFJC).
Other material. ETHIOPIA. Oromia 11.5 km S of Kibre Mengist 2200 m (MECI).
Etymology. The species was named after our colleague, the Italian ornithologist Antonio Rolando.
Description. Male (
Figure 11A,C): length 10–12 mm. Body greenish/bluish black, being covered by a short testaceous pubescence, with two large testaceous patches at the base and apex of elytra, basal one extending from elytral sutura to side margin, the apical one rounded and then placed on interstriae 1–4. Head surface smooth and covered by small points mixed to few larger ones; frontal carina largely triangular, sometimes being almost inapparent in major males, curved, more evident in minor males; genae not developed, slightly rounded; vertex carina lamina-shaped, rectangular with the digitiform apex relatively long and arched in major male, and shaped as a short, narrow protuberance with the superior edge that is rounded in minor male. Medium sized, slightly rounded eyes. Antennae yellowish brown. Pronotum covered by dense granulate setigerous points, large, ovalar granules becoming larger and thicker (almost embricate) on the sides; in major males a large, smooth, concave area in anterior part; pronotum base with a tight row of very large points. Elytra testaceous and black with greenish/bluish hue, the first two interstriae without granules, almost smooth, the others covered by sparse, regularly spaced setigerous points with small granules. Pygidium black, covered by deep, dense puncture, with large, simple points mixed to smaller ones. Legs blackish brown with short, thick testaceous setae.
Female (
Figure 11B,D): length 11–12 mm. Body as in male. Head surface rough on clypeus with some large points near the genal suturae, smooth with small sparse points on the remaining parts; genae not developed, slightly rounded, covered by many large points; frontal carina rounded, elevated, thick; vertex carina thick, rectilinear, large, reaching the eyes, area between the carinae smooth with relatively dense, small points being simple. Pronotum granulation as in male, two well-developed, conical protuberances slightly diverging, flat, and rounded from dorsal view. Elytra and legs as in male. Pygidium shorter than in male, but the same colour and puncture.
Epipharynx (
Figure 11E) anterior margin slightly concave, without central notch, acropariae that is constituted by very long, thin setae; zygum constituted by a tuft of long thick setae; proplegmatium triangular shaped, lowered; anterior epitorma thin, rod-shaped, but enlarged in the proximal third; plegmatic area ovalar lowered; chaetopariae rectilinear, proximal half with very short setae, longer on distal half; apotormae present, thin, rod-shaped, short; pternotormae short and thick; crepis asymmetrical, left-turned, sharp at apex; laeotorma and dexiotorma symmetrical, short, and thick; haptomerum with very short and thick setae.
Male genitalia (
Figure 11F,G,I): phallobase cylindrical, short, with an anterior rectangular expansion; parameres symmetrical, arched, tapering to the sharp apex; endophallus with a evident raspula constituted by very long, thin setae, lamella copulatrix (
Figure 11I) present, well-sclerotized, constituted by two distinct parts; accessory lamellae present, well-developed.
Female genitalia (
Figure 11H): vagina membranaceous, dome-shaped with a symmetrical, squared, short infundibular wall, with a rounded notch at base, and a well-sclerotized expansion; infundibulum question-mark shaped, rectilinear, thick, the distal part to ovarioles upward turned; receptaculum seminis not much expanded, lobate, tapering to the apex carrying a rounded laminal claw, the large desclerotized area that is placed in the central third.
Distribution. The species was collected from Eastern Africa (
Figure 7F).
Onthophagus rougonorum Cambefort, 1984: 8.
Type material.Holotype. ♂ ‘Ouango Fitini, iii.1981’ ‘Tiaronthophagus rougonorum (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN). Paratypes. 55 specimens ‘Ouango Fitini, iii.1981’ ‘Tiaronthophagus rougonorum (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); 58 specimens ‘Ouango Fitini, v.1981’ ‘Tiaronthophagus rougonorum (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); six specimens ‘Ouango Fitini, vii.1981’ ‘Tiaronthophagus rougonorum (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); four specimens ‘Niger Tapoa, vii–viii.1976 D. Rougon’ ‘Tiaronthophagus rougonorum (Cambefort, 1984) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Other material. BENIN. Tanguieta (Atakore) (PMOC). BURKINA FASO. Sanguié, Boromo, Forêt de Sorobouli, 270 m (PMOC); Sanguié, Forêt de Sorobouli, 270 m (PMOC). IVORY COAST. Korhogo, Koko (PMOC); Korhogo (PMOC); Man, Mt. Tonkoui (PMOC); PN du Mt. Sangbé (PMOC). SENEGAL. Kolda, Mahon Forêt de Bakor (PMOC); Niokolo Koba NP, Niokolo (PMOC); Niokolo Koba NP, poste de Niokolo, 65 m (PMOC); Niokolo Koba NP, Siminti (PMOC); PN Niokolo Koba, lisière Ft. galerie, Mt. Assirik, 144 m (PMOC); PN Niokolo Koba, savane arborée, Mt. Assirik, 144 m (PMOC). TOGO. Fazao, PN du Fazao (PMOC).
Distribution. The species was collected from Western Africa (
Figure 7B).
Notes. The species was included in 28th group [
47], and related to
T. flexicornis.
Onthophagus rufobasalis Fairmaire, 1887: 113.
= Onthophagus heynei Lansberge, 1887: 108. D’Orbigny, 1902: 176.
Type material. Lectotype. ♂ (here designated) ‘Guelidi <handwritten>‘ ‘Onthophagus rufobasalis Fm. guelidi <handwritten>‘ rufobasalis Fairm. <handwritten>‘ ‘Museum Paris coll. L. Fairmaire 1906’ ‘Syntype’ ‘Syntype Onthophagus rufobasalis Fairmaire, 1887’ ‘MNHN EC8641’ ‘Lectotype Onthophagus rufobasalis Fairmaire, 1887 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus rufobasalis (Fairmaire, 1887) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN). Paralectotype. ♀ Guelidi Revoil <handwritten>‘ ‘ex Typis’ ‘Museum Paris coll. L. Fairmaire 1906’ ‘Syntype’ ‘Syntype Onthophagus rufobasalis Fairmaire, 1887’ ‘MNHN EC8642’ ‘Paralectotype Onthophagus rufobasalis Fairmaire, 1887 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus rufobasalis (Fairmaire, 1887) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Synonymy type material. O. heynei ♂ (lectotype) ‘E. Heyne Somalis Africa <handwritten by Lansberge>’ ‘Onthophagus Heynei Lansbge <handwritten by d’Orbigny>’ ‘RMNH.INS 1104278’ ‘Tiaronthophagus rufobasalis (Fairmaire, 1887) Roggero, Moretto, Barbero, Palestrini 2019’ (RMNH). 1♀ (paralectotype) ‘E. Heyne Somalis Africa <handwritten by Lansberge>’ ‘Onthophagus Heynei Lansberge ♀’ ‘RMNH.INS 1104277 <handwritten by Lansberge>’ ‘Tiaronthophagus rufobasalis (Fairmaire, 1887) Roggero, Moretto, Barbero, Palestrini 2019’ (RMNH); 1♂ (paralectotype) ‘Ex-Musaeo Van Lansberge’ ‘Heynei. Lansbe. Type Somali’ ‘Muséum Paris ex. Coll. R. Oberthür’ ‘Syntype’ ‘Syntype Onthophagus heynei Lansberge, 1887’ ‘MNHN EC8643’ ‘Tiaronthophagus rufobasalis (Fairmaire, 1887) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Other material. ETHIOPIA. 2 km SE Key Afer, Gamu Gofa (PMOC); Southern, Turmi 920 m Mango-Lodge (PMOC); Hamer or., Turmi near 950 m (MDCL). KENYA. Mombasa Diani (EBCT); Meru District, Gatunga (PMOC); Meru district, Materi (Mitunguu) mt. 800 (PMOC); Mombasa Diani beach (PMOC); 30 km S Mombasa (JFJC); 5 km N Malindi (EBCT); km 65 S Mombasa (EBCT); Mombasa (JFJC); Afr. Or. Shimo la Tewa (NHMW); Kibwezi (NHMW); Hola (MDL) (PMOC). SOMALIA. Benadir Afgoi (Mogadiscio) (NHMW); ‘Somalia italiana’ Salambà (MSNG); Benadir Mogadiscio (MSNG); ‘Somalia It.’ O. Giuba Balet Amin (MSNG); ‘Somalia italiana mer.’ Villaggio Duca degli Abruzzi (now, Giohar) (MSNG); Balad (PMOC) (EBCT); Mogadiscio (MCST); umg. Mogadiscio (PMOC). TANZANIA. Trockenvald b. Mtotohovu, ‘DOA’ (in Tanga district) (ZMHB); 35 km E Singida (JFJC); ‘Afr.-Tanzanien’ Ngorongoro res. Serengeti (JFJC); Dar es Salaam campus université (JFJC); Dar es Salaam (PMOC); Iringa prov. Chimala 1000 m (PMOC); Morogoro region, Mikumi (PMOC); Iringa Province, Mkimbizi Mts. (PMOC); Morogoro region, Mazimbu area (PMOC); Morogoro region, Mikesse Hills, 420 m (PMOC); Uluguru Mts. (PMOC); Morogoro region, 360 m, Mbarika Mts. (Ikongwa) between Kichangari and Idunda (PMOC); Morogoro region, Udzungwa N.P. forêt, 390 m (PMOC); Taga region, savane de Kisangiro, 750 m (PMOC); Rufiji river, 15 km W Ikwiriri (MDCL): Mt. Kilimanjaro (PMOC).
Distribution. The species is known from Eastern Africa (
Figure 7D).
Notes. The synonymy of T. rufobasalis and O. heynei was confirmed by the survey of the typical material of these species.
Onthophagus rufopygus Frey, 1957: 688.
Type material. Holotype. ♂ ‘Kumba 20.xi.1955’ ‘♂’ ‘H. Typus <handwritten> Onthophagus <print> rufo—♂ <handwritten> pygus m <handwritten> det. G. Frey, 1957 <print>’ ‘Exped. Mus. G. Frey Nigeria-Kamerun Bechyne 1955-56’ ‘holotype’ ‘Tiaronthophagus rufopygus (Frey, 1957) Roggero, Moretto, Barbero, Palestrini 2019’ (NHMB). Paratypes. 2♂ ‘Kumba 20.xi.1955’ ‘♂’ ‘P. Typus Onthophagus rufopygus ♂ <handwritten> det. G. Frey, 1957 <print>’ ‘Exped. Mus. G. Frey Nigeria-Kamerun Bechyne 1955-56’ ‘paratype’ ‘Tiaronthophagus rufopygus (Frey, 1957) Roggero, Moretto, Barbero, Palestrini 2019’ (NHMB). Allotypes. 1♂ and 1♀ ‘Kumba 20.xi.1955’ ‘A. Typus Onthophagus rufopygus <handwritten> det. G. Frey, 1957 <print>’ ‘Exped. Mus. G. Frey Nigeria-Kamerun Bechyne 1955–56’ ‘allotype’ ‘Tiaronthophagus rufopygus (Frey, 1957) Roggero, Moretto, Barbero, Palestrini 2019’ (NHMB).
Other material. IVORY COAST. Abidjan PN du Banco (PMOC); Man Mt. Tonkoui 1200 m (PMOC); Danané (PMOC); Man Mt. Tonkoui (PMOC). GUINEA. Nzerekore Forêt, Classée de Diecké (PMOC). SIERRA LEONE. 120 m Tiwai Island, Moa river (BMNH).
Distribution. Except the type material, all the recently collected material come from the Western Africa (
Figure 7E). The type material was collected by Bechyné during his 1956 trip in Nigeria and British Cameroon. The collection area of
O. rufopygus was classified as ‘rainforest’.
Notes. The species was included in 24th group [
52] although the lamina of vertex of
T. rufopygus is very different from that of other species of the same group, as
T. schaufussi.
Onthophagus rufostillans d’Orbigny, 1907: 173.
Type material. Holotype. ♀ ‘Togo Conradt’ ‘holotypus’ ‘rufostillans n.sp. d’Orb. <handwritten by d’Orbigny>’ ‘Tiaronthophagus rufostillans (d’Orbigny, 1907) Roggero, Moretto, Barbero, Palestrini 2019’ (SDEI).
Other material. CAMEROON. Extreme north, province Logone and Chari, division Wasa (PMOC). GHANA. Kyabobo National Park near Laboum Outpost (PMOC). IVORY COAST. Korhogo, Koko 347 m (PMOC); Touba (PMOC); Comoé, Zamou (PMOC); Odienne, Sud Massif du Denguélé (PMOC); Touba, Biémasso, 441 m (PMOC); Touba, Biémasso-Dolla (PMOC); Lamto (PMOC); Man, Mt. Tonkui (PMOC).
Distribution. The species has a Western Africa distribution (
Figure 7F).
Type material. Holotype. ♂ ‘Onthophagus Tanz. sp. 46 S. Pokorný det 2014’ ‘Tanzania 200m Zaraninge Coastal Forest Saadani N.P., S. Pools x–xi.94 Pitfall Trap UDSM coll.’ ‘BMNH(E) 2013-71 1310485’ ‘BMNH <handwritten>’ ‘Tiaronthophagus saadaniensis n.sp. holotype’ (BMNH). Paratypes. 11♂ and 2♀ ‘Tanzania 200m Zaraninge Coastal Forest Saadani N.P., S. Pools x–xi.94 Pitfall Trap UDSM coll.’ ‘BMNH(E) 2013-71 1310492’ ‘BMNH <handwritten>’ ‘Onthophagus Tanz. sp. 46 S. Pokorný det 2014’ ‘Tiaronthophagus saadaniensis n.sp. paratype’ (BMNH).
Other material. No other material is known at present.
Etymology. The species was named after the collection locality in Tanzania.
Description. Male (
Figure 12A,C): length 11–13.5 mm. Body wholly covered by a long, dense and thin pubescence light yellow; head cupreous, pronotum green, frequently with cupreous reflects; elytra brown, mat. Head surface rough, with some large points on clypeus; genae not expanded, slightly rounded, with large points; frontal carina slightly rounded, evident; vertex carina conical-shaped, sharp, very small. Antennae lamellae light yellow, the scape brown. Pronotum entirely covered by very thick, large granulate points, granules very large, ovalar, except for the fore central smooth area with two lateral elevated triangular expansions. Elytral interstriae evenly covered by very thick granulate points, the granules large, ovalar. Pygidium testaceous with a light greenish lustre, with rounded, dense points and thick, long yellow pubescence on the whole surface. Legs brown with long thick setae.
Female (
Figure 12B,D): length 11.5–14 mm. Similar to male. Pygidium shorter and darker.
Epipharynx (
Figure 12E) anterior margin largely triangular notched, acropariae evenly long; zygum reduced to few long setae; anterior epitorma very thin, rod-shaped; proplegmatium triangular, very lowered, sides rectangular, apotormae barely visible, rod-shaped; pternotormae narrow, downturned; plegmatic area ovalar, large, enlarged, crepis narrow, asymmetrical, left turned; laeotorma and dexiotorma symmetrical, short, relatively thin; haptomerum with very short, thick setae.
Male genitalia (
Figure 12F,G,I): phallobase cylindrical, short, with an anterior rectangular expansion; parameres symmetrical, almost rectilinear, tapering to the sharp apex, not vertically expanded; endophallus with a evident raspula constituted by very long, thin setae, lamella copulatrix (
Figure 12I) present, well-sclerotized; accessory lamellae present, well-developed.
Female genitalia (
Figure 12H): vagina membranaceous, dome-shaped with a symmetrical, large, triangular infundibular wall, and two sacciform, sclerotized protuberances on sides; infundibulum question-mark shaped, rectilinear, thick, distal part to ovarioles upward turned; receptaculum seminis not much expanded, tubular, apex rounded with a small laminal claw, desclerotized area that is placed in the central third.
Distribution. The species at present is known only from the type locality in Tanzania (
Figure 7E).
Onthophagus schaufussi Harold, 1867: 43.
= Onthophagus nutans (nec Fabricius) var. maxima Roth, 1851 (MNHN, see notes below). Harold, 1867: 44.
Type material. Lectotype. ♂ (here designated) ‘Tigré. <handwritten>‘ ‘Schaufussi T. Harold <handwritten>‘ ‘Ex-Musaeo Harold’ ‘Muséum Paris ex Coll. R. Oberthür’ ‘Syntype’ ‘Syntype Onthophagus schaufussi Harold, 1867’ ‘MNHN EC8636’ (MNHN) ‘Lectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Paralectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’. Paralectotypes. ♂ ‘Tigré. O. schaufussi Harold. ‘<handwritten>‘ ‘Ex-Musaeo Harold’ ‘Muséum Paris ex Coll. R. Oberthür’ ‘Syntype’ ‘Syntype Onthophagus schaufussi Harold, 1867’ ‘MNHN EC8637’ ‘Paralectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Paralectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); ♀ ‘Tigré <handwritten>‘ ‘Ex-Musaeo Harold’ ‘Muséum Paris ex Coll. R. Oberthür’ ‘Syntype’ ‘Syntype Onthophagus schaufussi Harold, 1867’ ‘MNHN EC8638’ ‘Paralectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Paralectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); ♂ ‘Tigré. <handwritten>‘ ‘Ex-Musaeo Harold’ ‘Muséum Paris ex Coll. R. Oberthür’ ‘Syntype’ ‘Syntype Onthophagus schaufussi Harold, 1867’ ‘MNHN EC8639’ ‘Paralectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Paralectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN); ♂ ‘Tigré. Schimper. <handwritten>‘ ‘Ex-Musaeo Harold’ ‘Muséum Paris ex Coll. R. Oberthür’ ‘Syntype’ ‘Syntype Onthophagus schaufussi Harold, 1867’ ‘MNHN EC8640’ ‘Paralectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini 2019 ‘Paralectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Synonymy type material. O. nutans var. maxima. Lectotype ♂ (here designated), ‘Abyssinia O schaufussi Typi: Har: <handwritten>’ ‘round blue chip’ ‘something <red>’ ‘Lectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini 2019‘ ‘Paralectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’ (ZSM). Paralectotypes. O. nutans var. maxima nine ex. ‘round blue chip’ ‘Paralectotype Onthophagus nutans var. maxima Roth, 1851 Roggero, Moretto, Barbero, Palestrini2019’ ‘Paralectotype Onthophagus schaufussi Harold, 1851 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus schaufussi (Harold, 1851) Roggero, Moretto, Barbero, Palestrini 2019’ (ZSM).
Other material. ERITREA. Serayè Dubarwa (EBCT) (PMOC); env. Asmara (PMOC). ETHIOPIA. Gemu-Gofa prov. near Arba Minch (PMOC) (MDCL); Oromia reg Arsi Negele Woreda 1600 m Lake Langano near Bishangari (PMOC); Asosa, Benishangui (MDCL); Turmi, 920 m (PMOC).
Distribution. The species is distributed in Eastern Africa (
Figure 7F).
Notes. Following the ICZN code (45.6.4), the subspecific rank can be assigned to
O. nutans var.
maxima Roth, and the taxon can be considered to be valid. Harold (1867: 45) stated that the type material of
O. schaufussi come from the Roth collection. The author described the species on some specimens which were formerly described [
55] as
O. nutans Fabricius var.
maxima from material collected by Schimper in Abyssinia. Ten specimens of the same material collected by Schimper and coming from Roth collection are preserved in ZSM as type material of
O. schaufussi Harold. Together, the five syntypes of O. schaufussi Harold held in Paris and the 10 syntypes held in München are the type series of
O. nutans var.
maxima Roth and of
O. schaufussi Harold, which are synonyms. At present,
O. nutans Fabricius, 1787 is regarded as a synonym of
Palaeonthophagus verticicornis (Laichtarting, 1781), thus the proposed variety is surely due to misidentification. Although the var.
maxima was described prior
O. schaufussi, it was only mentioned as a synonym of the latter name, which must be maintained following the stability criterion that is recommended by the ICZN.
Onthophagus viridiaereus d’Orbigny, 1908: 163.
Type material. Holotype. ♂ (by monotypy) ‘Guinée Port. Chime 1906 Rio Geba G. Favarel d’Orbigny 1908 coll. Oberthur’ ‘Holotype Onthophagus viridiaereus d’Orbigny, 1908 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus viridiaereus (d’Orbigny, 1908) Roggero, Moretto, Barbero, Palestrini 2019’ (MNHN).
Other material. BENIN. Ndali, 3 km W of Sontou (PMOC); Penessoulou forêt de Penelan (PMOC). BURKINA FASO. Bama, site de Samandeni, 330 m (PMOC); Bobo Diulasso (PMOC); Bobo Diulasso, Farako Ba Station (PMOC); Loroum Toulfé 330 m (PMOC); Nahouri Tiakané 340 m (PMOC); NE Oubritenga, Zithenga Tanghen (=Zitenga-Tanghin) (PMOC). CAMEROON. Extreme north (PMOC). DEMOCRATIC REPUBLIC OF CONGO. ‘Congo Belge’ Central Province of Maniema, Kindu (MNHN). GUINEA BISSAU. Binar, Oio; Coli (Quebo), Tombali (CAS). MALI. Segou (MHNL). SENEGAL. Fatick Diouroup (PMOC); Kaolak, Nioro du Rip (PMOC); Sebikotane Dakar (MHNL); Sebikotane (MNHN); south, 10 km SW Medina Gounass (PMOC); Zinguinchor 18m Boukithingo (PMOC).
Distribution. The species is known from Western Africa (
Figure 7B).
Type material. Holotype. ♂ ‘21.xi.2010 1280 m 9 km S Lusaka 15°30′11″ S, 28°15′52″ E, Zambie (Lusaka S.Rojkoff réc.’ ‘Tiaronthophagus zambesianus n.sp. holotype’ (BMNH). Paratypes. 1♂ ‘21.xi.2010 1280 m 9 km S Lusaka 15°30′11″ S, 28°15′52″ E, Zambie (Lusaka S.Rojkoff réc.’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (JFJC); 1♂ ‘Zambie Lusaka Prov. Lower Zambesi N.P. 14-16-xii-2012 15°22’ S, 29°18’ E, leg. Josso Juhel Minetti’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (JFJC); 1♀ ‘Zambia Southern province 10km E Zimba 24.-26.12.2002 A. Kudrna jr. lgt.’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (PMOC); 1♂ ‘Zambie i.98 Kafue Co. R. Minetti leg. <handwritten>’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (PMOC); 1♂ ‘Zambie, Kafue Co. i.98 R. Minetti leg.’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (PMOC); 3♂ 6♀ ‘Zambie 17 km N Choma I. Bruce–Miller farm 16°38.22’ S, 27°01.51’ E, 26xi-7xii-2013 1170 m leg. Josso Juhel Minetti’ ‘exc. humains’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (JFJC); 3♂ 2♀ ‘N Malawi Vwaza Marsh Reserv Rumphi dist. 1150 m 17-18-i1201 J-F Josso rec.’ ‘poisson pourri’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (JFJC); 1♂ 1♀ ‘Karoi Vuti env. Zimbabwe 18-xii-1998’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (JFJC); 2♀ ‘Zimbabwe bor. Karoi Vuti env. 18.12.98 lgt. Smrž’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (JFJC); 1♂ 1♀ ‘Tanzania, 4.xii.1999 Dodoma prov. near Itigi K. Werner & R. Lizler leg.’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (PMOC); 1♂ 2♀ ‘Tanzanie, Iringa prov. Chimala savane 1674 m 20.ii.2004 8°52.510’ S, 33°59.453’ E, Ph.Darge leg.’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (PMOC); 5♂ 4♀ Tanzanie, Iringa prov. Chimala, 1000 m 22.ii.2004 R. Minetti leg.’ ‘Tiaronthophagus zambesianus n.sp. paratype’ (PMOC).
Other material. No other material was examined.
Etymology. The species was named after the collection area.
Description. Male (
Figure 13A,C): length 9–11 mm. Body brownish black, covered by a short, thick, testaceous pubescence, with some small testaceous patches at base of elytra (on interstriae 1–3, and 5–6), and at apex of elytra (barely visible, on interstriae 1–3). Head surface smooth and covered by small points mixed to few larger ones; frontal carina slightly curved. sometimes almost inapparent in major males, more evident in minor males; genae not developed, slightly rounded; vertex carina lamina-shaped, squared, with sides lightly curved, digitiform apex relatively long and arched in major male, while the carina is shaped as a short, narrow rectangular protuberance with the superior edge truncated in minor male. Medium sized, slightly rounded eyes. Antennae yellowish brown. Pronotum covered by dense granulate setigerous points, with large, ovalar; in major males a smooth, concave area in anterior part; pronotum base with a tight row of evident points. Elytra testaceous and black, first two interstriae without granules, almost smooth, the others being covered by sparse, regularly spaced setigerous points with small granules. Pygidium black, with dense, large, and deep puncture. Legs blackish brown with short, thick testaceous setae.
Female (
Figure 13B,D): length 9–11 mm. Body as in male. Head surface is wrinkled on clypeus with some large points, less rough with small mixed to large points on the remaining parts; genae not developed, slightly rounded; frontal carina rounded, elevated, thick; vertex carina thick, rectangular, elevated, narrow; area between the carinae smooth with relatively dense, small points simple. Pronotum granulation as in male, two small, conical protuberances that slightly diverge. Elytra and legs as in male. Pygidium shorter than in male, but same colour and puncture.
Epipharynx (
Figure 13E): anterior margin slightly concave, without central notch, acropariae constituted by very long, thin setae, slightly tapering toward the zygum; evident zygum that is constituted by a tuft of long thick setae; proplegmatium triangular shaped, sides rectangular; anterior epitorma thin, rectilinear in distal half, slightly expanded, bottle-shaped in proximal half; plegmatic area ovalar, small, very lowered; chaetopariae only slightly sinuate, proximal half with very short setae, longer on distal half; apotormae barely visible, thin, rod-shaped, short; pternotormae short and thick; crepis asymmetrical, small, left-turned, apex blunt; laeotorma and dexiotorma symmetrical, short, and thick; haptomerum with many short and thick setae.
Male genitalia (
Figure 13F,G,I): phallobase cylindrical, short, with an anterior rectangular expansion; parameres symmetrical, arched, tapering to the sharp apex; endophallus with a greatly developed raspula constituted by very long, thin setae, lamella copulatrix (
Figure 13I) present, well-sclerotized, constituted by two distinct parts; accessory lamellae present and well-developed.
Female genitalia (
Figure 13H): vagina membranaceous, triangular-shaped with a symmetrical, bilobed, short infundibular wall, with a large, rounded notch at base; infundibulum question-mark shaped, rectilinear, thick, distal part to ovarioles upward turned; receptaculum seminis not much expanded, tapering to the apex carrying a very small expansion, large desclerotized area placed in the central third.
Distribution. The species distribution covers the Southern East Africa (
Figure 7D).
Onthophagus zavattarii Müller, 1939: 266.
Type material. Lectotype. ♂ (here designated) ‘Aresc <sic> aprile 1937’ ‘Missione Zavattari nel Borana ROI’ ‘Onthophagus rufobasalis Fairmaire <handwritten>’ ‘Onthophagus zavattari n.sp. <handwritten>’ ‘Lectotype Onthophagus zavattarii Müller, 1939 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus zavattarii (Müller, 1939) Roggero, Moretto, Barbero, Palestrini 2019’ (MCST). Paralectotypes. 2♂, 2 ♀, ‘Neghelli 22–25.x.37’ ‘Sped. Brunelli Somalia Mer. leg. Vatova’ ‘Paralectotype Onthophagus zavattarii Müller, 1939 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus zavattarii (Müller, 1939) Roggero, Moretto, Barbero, Palestrini 2019’ (2♂, 2 ♀, MCST); 1 ♂, ‘Neghelli 31.x.37’ ‘Sped. Brunelli Somalia Mer. leg. Vatova’ ‘Paralectotype Onthophagus zavattarii Müller, 1939 Roggero, Moretto, Barbero, Palestrini 2019’ ‘Tiaronthophagus zavattarii (Müller, 1939) Roggero, Moretto, Barbero, Palestrini 2019’ (MCST).
Other material. ETHIOPIA. Sidamo, 25 km E Negele 7–10.v.2016 S. Prepsl leg. (JFJC).
Distribution. The species is distributed in Ethiopia (
Figure 7F).
Notes. A marked similarity of this species with
T. rufobasalis [
57] was highlighted, although the two species can be well differentiated by some characters, such as the features of the vertex carina in the females, and the pronotum punctures (
Supporting Information Data S5).



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}