The Mayan Tropical Rainforest: An Uncharted Reservoir of Tritrophic Host-Fruit Fly-Parasitoid Interactions

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
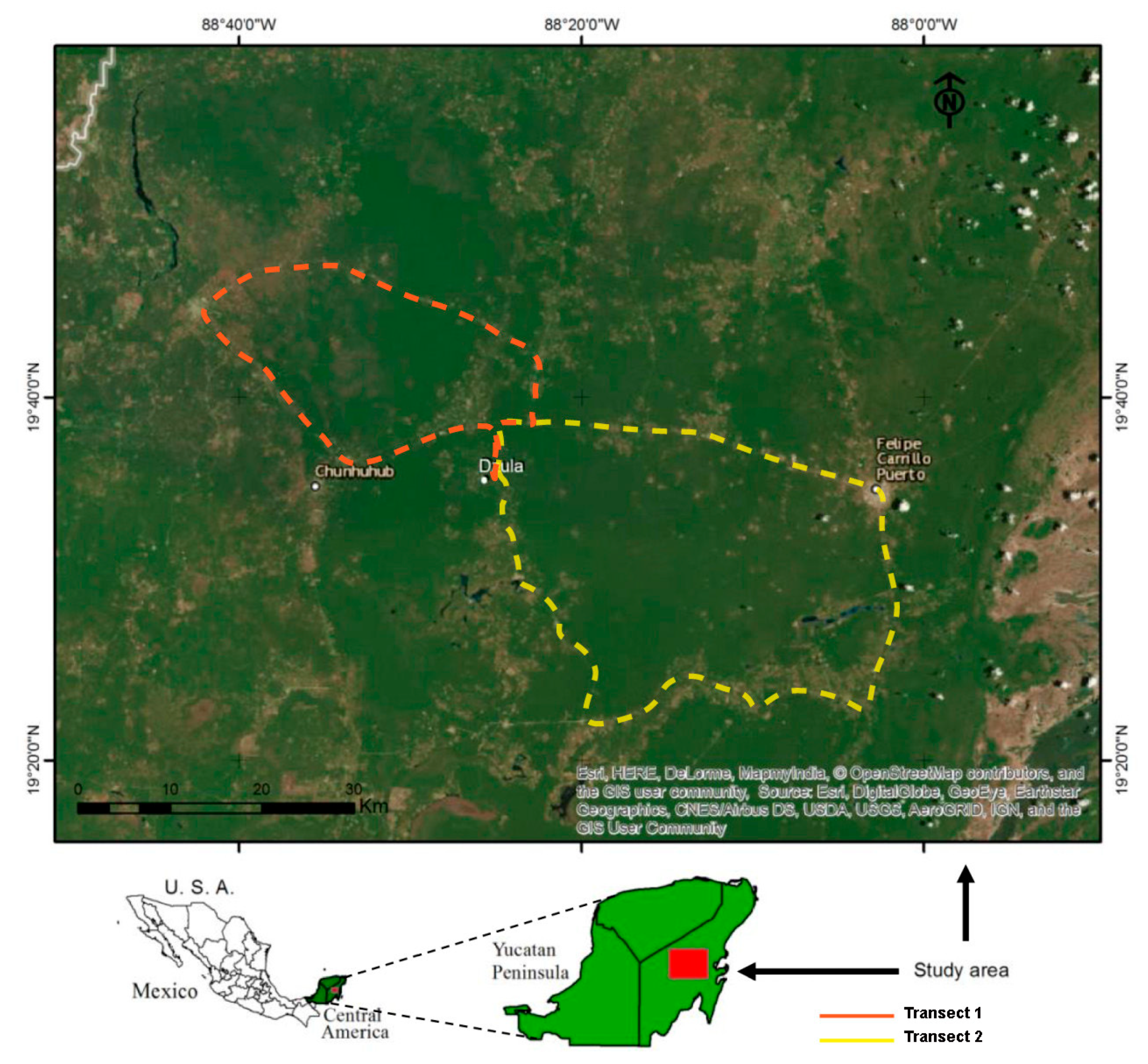
2.1. Study Site
2.2. Collection and Processing of Fruit Samples
2.3. Data Analyses
3. Results
3.1. Fruit Fly-Host Plant Interactions
3.2. Fruit Infestation and Parasitism Rates
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Becerra, J.X. On the factors that promote the diversity of herbivorous insects and plants in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 6098–6103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, P.W.; Denno, R.F.; Eubanks, M.D.; Finke, D.L.; Kaplan, I. Insect Ecology: Behavior, Populations and Communities; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Keck, C.B. Relation of oviposition punctures of the Mediterranean fruit fly to the premature dropping of citrus fruits. J. Econ. Entomol. 1934, 28, 908–914. [Google Scholar] [CrossRef]
- Stephenson, A.G. Flower and fruit abortion: Proximate causes and ultimate functions. Annu. Rev. Ecol. Syst. 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Drew, R.A.I. Amino acid increases in fruit infested by fruit flies of the family Tephritidae. Zool. J. Linn. Soc. 1988, 93, 107–112. [Google Scholar] [CrossRef]
- Pimm, S.; Russell, G.; Gittleman, J.; Brooks, T. The future of Biodiversity. Science 1995, 269, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Norrbom, A.L.; Korytkowski, C.A.; Zucchi, R.A.; Uramoto, K.; Venable, G.L.; McCormick, J.; Dallwitz, M.J. Anastrepha and Toxotrypana: Descriptions, Illustrations, and Interactive Keys; Version: 16th October 2018; 2012; Dallwitz, Paine, and Zurcher 1995, 2000, USDA; Available online: http://delta-intkey.com (accessed on 6 December 2018).
- Eskafi, F.M.; Cunningham, R.T. Host plants of fruit flies (Diptera: Tephritidae) of economic importance in Guatemala. Fla. Entomol. 1987, 70, 116–123. [Google Scholar] [CrossRef]
- Jiron, L.F.; Hedström, I. Occurrence of fruit flies of the genera Anastrepha and Ceratitis (Diptera: Tephritidae), and their host plant availability in Costa Rica. Fla. Entomol. 1988, 71, 62–73. [Google Scholar] [CrossRef]
- Borge, M.N.R.; Basedow, T. A survey on the occurrence and flight periods of fruit fly species (Diptera: Tephritidae) in a fruit growing area in southwest Nicaragua, 1994/95. Bull. Entomol. Res. 1997, 87, 405–412. [Google Scholar] [CrossRef]
- Uramoto, K.; Martins, D.S.; Zucchi, R.A. Fruit flies (Diptera, Tephritidae) and their associations with native host plants in a remnant area of the highly endangered Atlantic Rain Forest in the State of Espírito Santo, Brazil. Bull. Entomol. Res. 2008, 98, 457–466. [Google Scholar] [CrossRef]
- Silva, J.G.; Dutra, V.S.; Santos, M.S.; Silva, N.M.; Vidal, D.B.; Nink, R.A.; Guimaraes, J.A.; Araujo, E.L. Diversity of Anastrepha spp. (Diptera: Tephritidae) and associated braconid parasitoids from native and exotic hosts in Southeastern Bahia, Brazil. Environ. Entomol. 2010, 39, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.R.; Norrbom, A.L. Tephritoid flies (Diptera, Tephritoidea) and their plant hosts from the state of Santa Catarina in southern Brazil. Fla. Entomol. 2011, 94, 151–157. [Google Scholar] [CrossRef]
- Jesus-Barros, C.R.; Adaime, R.; Oliveira, M.N.; Silva, W.R.; Costa-Neto, S.V.; Souza-Filho, M.F. Anastrepha (Diptera: Tephritidae) species, their hosts and parasitoids (Hymenoptera: Braconidae) in five municipalities of the state of Amapá, Brazil. Fla. Entomol. 2012, 95, 694–705. [Google Scholar] [CrossRef]
- Norrbom, A.L.; Castillo-Meza, A.L.; García-Chávez, J.H.; Aluja, M.; Rull, J. A new species of Anastrepha (Diptera: Tephritidae) from Euphorbia tehuacana (Euphorbiaceae) in Mexico. Zootaxa 2014, 3780, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Clavijo, P.A.R.; Norrbom, A.L.; Peñaranda, E.A.; Diaz, P.A.; Benitez, M.C.; Gallego, J.; Cruz, M.I.; Montes, J.M.; Rodriguez, E.J.; Steck, G.J.; et al. New records of Anastrepha (Diptera: Tephritidae) primarily from Colombia. Zootaxa 2018, 4390, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ortiz, V.; Pérez-Alonso, R. The natural host plants of Anastrepha (Diptera: Tephritidae) in a tropical rain forest of Mexico. Fla. Entomol. 1993, 70, 447–460. [Google Scholar] [CrossRef]
- Aluja, M.; Rull, J.; Sivinski, J.; Norrbom, A.L.; Wharton, R.A.; Macías-Ordóñez, R.; Díaz-Fleischer, F.; López, M. Fruit flies of the genus Anastrepha (Diptera: Tephritidae) and associated native parasitoids (Hymenoptera) in the tropical rainforest biosphere reserve of Montes Azules, Chiapas, Mexico. Environ. Entomol. 2003, 32, 1377–1385. [Google Scholar] [CrossRef]
- Deus, E.G.; Pinheiro, L.S.; Lima, C.R.; Sousa, M.D.S.M.; Guimarāes, J.A.; Strikis, P.C.; Adaime, R. Wild hosts of frugivorous dipterans (Tephritidae and Lonchaeidae) and associated parasitoids in the Brazilian Amazon. Fla. Entomol. 2013, 96, 1621–1625. [Google Scholar] [CrossRef]
- Almeida, R.R.; Cruz, K.R.; Sousa, M.S.M.; Costa-Neto, S.V.; Jesus Barros, C.R.; Lima, A.L.; Adaime, R. Frugivorous flies (Diptera: Tephritidae, Lonchaeidae) associated with fruit production on Ilha de Santana, Brazilian Amazon. Fla. Entomol. 2016, 99, 426–436. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Aluja, M. Listado de especies del género neotropical Anastrepha (Diptera: Tephritidae), con notas sobre su distribución y plantas hospederas. Folia Entomol. Mex. 1993, 88, 89–105. [Google Scholar]
- Hernández-Ortiz, V.; Canal, N.A.; Salas, J.O.T.; Ruíz-Hurtado, F.M.; Dzul-Cauich, J.F. Taxonomy and phenotypic relationships of the Anastrepha fraterculus complex in the Mesoamerican and Pacific Neotropical dominions (Diptera, Tephritidae). ZooKeys 2015, 540, 95–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Ortiz, V. Diversidad y biogeografía del género Anastrepha en México. In Moscas de la Fruta en Latinoamérica (Diptera: Tephritidae): Diversidad, Biología y Manejo; S y G, Ed.; Distrito Federal, Mexico, 2007; pp. 53–76. [Google Scholar]
- Carrejo, N.S.; Gonzalez, R. Parasitoids reared from species of Anastrepha (Diptera: Tephritidae) in Valle del Cauca, Colombia. Fla. Entomol. 1999, 82, 113–118. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Delfín-González, H.; Escalante-Tio, A.; Manrique-Saide, P. Hymenopteran parasitoids of Anastrepha fruit flies (Diptera: Tephritidae) reared from different hosts in Yucatan, Mexico. Fla. Entomol. 2006, 89, 508–515. [Google Scholar] [CrossRef]
- Clausen, C.P.; Clancy, D.W.; Chock, Q.C. Biological Control of the Oriental Fruit Fly (Dacus dorsalis Hendel) and Other Fruit Flies in Hawaii; USDA Tech. Bull.: Washington, DC, USA, 1965; Volume 1322, p. 102.
- Wharton, R.A. Classical Biological Control of Fruit-Infesting Tephritidae. In World Crop Pests, Fruit Flies: Their Biology, Natural Enemies and Control; Robinson, A.S., Hooper, G., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; Volume 3B, pp. 303–313. [Google Scholar]
- Leonel, F.L., Jr.; Zucchi, R.A.; Wharton, R.A. Distribution and tephritid hosts (Diptera) of braconid parasitoids (Hymenoptera) in Brazil. Int. J. Pest Manag. 1995, 41, 208–213. [Google Scholar] [CrossRef]
- Wharton, R.A.; Gilstrap, F.E.; Rhode, R.H.; Fischel-m, M.; Hart, W.G. Hymenopterous egg-pupal and larval-pupal parasitoids of Ceratitis capitata and Anastrepha spp. (Dip.: Tephritidae) in Costa Rica. Entomophaga 1981, 26, 285–290. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Pérez-Alonso, R.; Wharton, R.A. Native parasitoids associated with the genus Anastrepha (Diptera: Tephritidae) in Los Tuxtlas, Veracruz, Mexico. Entomophaga 1994, 39, 171–178. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M.; López, M. Spatial and temporal distributions of parasitoids of Mexican Anastrepha species (Diptera: Tephritidae) within the canopies of fruit trees. Ann. Entomol. Soc. Am. 1997, 90, 604–618. [Google Scholar] [CrossRef]
- López, M.; Aluja, M.; Sivinski, J. Hymenopterous larval–pupal and pupal parasitoids of Anastrepha flies (Diptera: Tephritidae) in Mexico. Biol. Cont. 1999, 15, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Ovruski, S.; Aluja, M.; Sivinski, J.; Wharton, R. Hymenopteran parasitoids on fruit-infesting Tephritidae (Diptera) in Latin America and the southern United States: Diversity, distribution, taxonomic status and their use in fruit fly biological control. Integr. Pest Mgmt. Rev. 2000, 5, 81–107. [Google Scholar] [CrossRef]
- Montoya, P.; Ayala, A.; López, P.; Cancino, J.; Cabrera, H.; Cruz, J.; Martinez, A.M.; Figueroa, I.; Liedo, P. Natural parasitism in fruit fly (Diptera: Tephritidae) populations in disturbed areas adjacent to commercial mango orchards in Chiapas and Veracruz, Mexico. Environ. Entomol. 2016, 45, 328–337. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Manrique-Saide, P.; Delfín- González, H.; Novelo-Rincón, I. First report of Anastrepha compressa in Mexico and new records for other Anastrepha species in the Yucatan Peninsula (Diptera: Tephritidae). Fla. Entomol. 2002, 85, 389–391. [Google Scholar] [CrossRef]
- Aluja, M.; Piñero, J.; López, M.; Ruíz, C.; Zúñiga, A.; Piedra, E.; Díaz-Fleisher, F.; Sivinski, J. New host plant and distribution records in Mexico for Anastrepha spp., Toxotrypana curvicauda Gerstacker, Rhagoletis zoqui Bush, Rhagoletis sp., and Hexachaeta sp. (Diptera: Tephritidae). Proc. Entomol. Soc. Wash. 2000, 102, 802–815. [Google Scholar]
- Rzedowski, J. Vegetacion de Mexico; Limusa: Distrito Federal, Mexico, 1978; p. 432. [Google Scholar]
- Miranda, F.D.P.; Hernández, X.E. Los tipos de vegetación de México y su clasificación. Bol. Soc. Bot. México 1963, 28, 29–179. [Google Scholar] [CrossRef]
- Wharton, R.A.; Marsh, P.M.; Sharkey, M.J. Manual of the New World genera of the family Braconidae (Hymenoptera). Spec. Publ. Int. Soc. Hymenopterists 1997, 1, 459. [Google Scholar]
- Tropicos.org. Missouri Botanical Garden. Available online: http://www.tropicos.org (accessed on 6 December 2018).
- Garcí Ramírez, M.D.J.; Antonio Hernández, E.; Vargas Magaña, J.J.; Valencia Gutiérrez, M.D.C.; Chi Ruiz, J.C.; Placensia Valerio, Y. First host plant record for Anastrepha ampliata Hernández-Ortiz, 1990 (Diptera: Tephritidae). Biocyt Biol. Cienc. Tecnol. 2018, 11, 789–791. [Google Scholar]
- Hernaández-Ortiz, V. El Género Anastrepha Schiner en México (Diptera: Tephritidae). Taxonomía, Distribución y sus Plantas Huéspedes; Publicacón No. 33; Instituto de Ecología: Xalapa, Mexico, 1992; p. 162. [Google Scholar]
- Antonio Hernández, E.; Ramírez, M.D.J.G.; Lara, D.F. New records of the genus Anastrepha Schiner, 1868 (Diptera: Tephritidae) in the Isthmus of Tehuantepec, Oaxaca, Mexico. Biocyt Biol. Cienc. Tecnol. 2018, 11, 824–833. [Google Scholar]
- Sosa-Armenta, J.M.; López-Martínez, V.; Villegas-Torres, Ó.G.; Juárez-López, P.; Burgos-Solorio, A. Dinámica poblacional de moscas de la fruta en Quintana Roo, México. Southwest Entomol. 2017, 42, 275–282. [Google Scholar] [CrossRef]
- Sosa-Armenta, J.M.; López-Martínez, V.; Alia-Tejacal, I.; García-Jiménez, D.; Guillen-Sánchez, D.; Delfín-González, H. Hosts of five Anastrepha species (Diptera: Tephritidae) in the state of Quintana Roo, Mexico. Fla. Entomol. 2015, 98, 1000–1002. [Google Scholar] [CrossRef] [Green Version]
- Araujo, M.R.; Uramoto, K.; Ferreira, E.N.L.; Mesquita Filho, W.; Walder, J.M.M.; Savaris, M.; Zucchi, R.A. Fruit Fly (Diptera: Tephritidae) diversity and host relationships in diverse environments estimated with two sampling methods. Environ. Entomol. 2018, 48, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Plummer, C.C.; McPhail, M. The yellow chapote, a native host of the Mexican fruit fly. USDA Tech. Bull. 1941, 775, 1–12. [Google Scholar]
- Bush, G.L. The cytotaxonomy of the larvae of some Mexican fruit flies in the genus Anastrepha (Tephritidae, Diptera). Psyche 1962, 69, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.B. Mexican fruit fly (Diptera: Tephritidae) and the phenology of its native host plant yellow chapote (Rutaceae) in Mexico. J. Entomol. Sci. 1999, 47, 1–16. [Google Scholar] [CrossRef]
- Salter, E.A. Flora Nicaragüense. In Arboles y Arbustos Más Notables y el Uso de Sus Maderas y Otros Productos; Imprenta La Salle: Bluefield, Nicaragua, 1947; p. 280. [Google Scholar]
- Pennington, T.D. Flora neotropica. Monograph 52. Sapotaceae; New York Botanical Garden for the Organization for Flora Neotropica: New York, NY, USA, 1990; p. 770. [Google Scholar]
- Barbeau, G. Frutas Tropicales en Nicaragua. Edit. Ciencias Sociales. Managua, Nicaragua. 1990, p. 397. Available online: https://agris.fao.org/agris-search/search.do?recordID=NI2006000072 (accessed on 1 May 2020).
- Bernays, E.A.; Chapman, R.F. Behavior: The Process of Host-Plant Selection. In Host-Plant Selection by Phytophagous Insects; Bernays, E.A., Chapman, R.F., Eds.; Chapman & Hall: New York, NY, USA, 1994; pp. 95–165. [Google Scholar]
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. The geographic range: Size, shape, boundaries, and internal structure. Annu. Rev. Ecol. Evol. Syst. 1996, 27, 597–623. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Ballou, J.D.; Dudash, M.R.; Eldridge, M.D.B.; Fenster, C.B.; Lacy, R.C.; Mendelson, J.R.; Porton, I.J.; Ralls, K.; Ryder, O.A. Implications of different species concepts for conserving biodiversity. Biol. Conserv. 2012, 153, 25–31. [Google Scholar] [CrossRef]
- Medianero, E.; Korytkowski, C.A.; Campo, C.; De León, C. Hymenoptera parasitoids associated with Anastrepha (Diptera: Tephritidae) at Cerro Jefe and Altos de Pacora, Panama. Rev. Colomb. Entomol. 2006, 32, 136–139. [Google Scholar]
- Leyva, J.L.; Browning, H.W.; Gilstrap, F.E. Effect of host fruit species, size, and color on parasitization of Anastrepha ludens (Diptera: Tephritidae) by Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 1991, 20, 1469–1474. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M. The roles of parasitoid foraging for hosts, food and mates in the augmentative control of Tephritidae. Insects 2012, 3, 668–691. [Google Scholar] [CrossRef] [Green Version]
- Sivinski, J.M.; Calkins, C.O.; Baranowski, R.; Harris, D.; Brambila, J.; Diaz, J.; Burns, R.E.; Holler, T.; Dodson, G. Suppression of a Caribbean fruit fly (Anastrepha suspensa (Loew) Diptera: Tephritidae) population through augmented releases of the parasitoid Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Cont. 1996, 6, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Montoya, P.; López, P.; Cruz, J.; Lopez, F.; Cadena, C.; Cancino, J.; Liedo, P. Effect of Diachasmimorpha longicaudata releases on the native parasitoid guild attacking Anastrepha spp. larvae in disturbed zones of Chiapas, Mexico. BioControl 2017, 62, 581–593. [Google Scholar] [CrossRef]
- Harbi, A.; Beitia, F.; Ferrara, F.; Chermiti, B.; Sabater-Muñoz, B. Functional response of Diachasmimorpha longicaudata (Ashmead) over Ceratitis capitata (Wiedemann): Influence of temperature, fruit location and host density. Crop. Prot. 2018, 109, 115–122. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Family | Scientific Name | Mayan Local Name | Sample Weight (Kg) | Infested Fruit Yes/No |
|---|---|---|---|---|
| Anacardiaceae | Metopium brownei (Jacq.) | Chechen | 0.265 | N |
| Spondias purpurea L. | Ciruela | 1.100 | Y | |
| Spondias mombin L. | Jobo | 1.62 | Y | |
| Mangifera indica L. | Mango | 4.58 | Y | |
| Annonaceae | Annona globiflora Schlecht. | Anona | 0.270 | N |
| Annona scleroderma Saff. | Chujun op | 0.680 | N | |
| Mosannona depressa (Baill.) Chatrou | Elemuy | 0.950 | N | |
| Araliaceae | Dendropanax arboreous (L.) Decne & Planch. | Sakchaca | 0.1620 | N |
| Bixaceae | Cochlospermum vitifolium (Willd.) Spreng. | Chuun | 0.870 | N |
| Boraginaceae | Ehretia tinifolia L. | Beek | 0.282 | N |
| Cordia dodecandra DC. | Ciricote | 5.310 | N | |
| Burseraceae | Bursera simaruba (L.) Sarg. | Chaca Rojo | 0.176 | N |
| Cannabaceae | Celtis iguanaea (Jacq.) Sarg. | Muk | 0.475 | N |
| Capparaceae | Crataeva tapia L. | Kookche | 3.320 | N |
| Caricaceae | Carica papaya L. | Chichput | 1.500 | Y |
| Ebeneaceae | Diospyros anisandra S.F. Blake | Kabche | 0.300 | N |
| Euphorbiaceae | Croton arboreus Millsp. | Perescuch | 0.310 | N |
| Gymnanthes lucida Sw. | Yaiti | 1.200 | N | |
| Fabaceae | Caesalpinia platyloba S. Watson | Chacteviga | 0.235 | N |
| Platymiscium yucatanum Standl. | Granadillo | 0.340 | N | |
| Piscidia piscipula (L.) Sarg. | Jabin | 0.410 | N | |
| Swartzia cubensis (Britton & Wills) Standl. | Katalox | 0.790 | N | |
| Caesalpinia gaumeri (Britton & Rose) Greenm. | Kitamche | 0.550 | N | |
| Lysiloma latisiliquum (L.) Benth. | Tzalam | 0.300 | N | |
| Lonchocarpus yucatanensis Pittier | Xuul | 0.260 | N | |
| Lauraceae | Nectandra salicifolia (H.B.K.) Nees. | Sakelemuy | 1.650 | N |
| Malpighiaceae | Bunchosia swartziana Griseb. | Sipche | 0.615 | N |
| Malvaceae | Hampea trilobata Standl. | Jool | 0.520 | N |
| Luehea candida (DC.) Mart. | Kaskaat | 0.960 | N | |
| Pseudobombax ellipticum (Kunth) Dugand | Amapola | 0.700 | N | |
| Ceiba petandra (L.) Gaerth. | Yaaxche | 1.300 | N | |
| Menispermaceae | Hyperbaena winzerlingii Standl. | Kekenche | 0.173 | N |
| Mimosaceae | Acacia milleriana Standl. | Chimay | 0.150 | N |
| Moraceae | Ficus pertusa L.f. | Juunkiix | 1.100 | N |
| Brosimum alicastrum Sw. | Ramon | 4.820 | N | |
| Myrtaceae | Psidium sartorianum (O. Berg) Nied. | Guayabillo | 0.885 | Y |
| Myrcianthes fragrans (Sw.) Mc Vaugh | Kojkann | 0.312 | N | |
| Eugenia biflora (L.) DC. | Pichiche | 0.500 | N | |
| Psidium guajava L. | Guayaba | 1.225 | Y | |
| Opiliaceae | Agonandra macrocarpa L. O. Williams | Napche | 1.765 | N |
| Passifloraceae | Passiflora foetida L. | Poochil | 0.150 | Y |
| Passiflora serratifolia L. | Maracuya del monte | 0.560 | Y | |
| Passiflora yucatanensis Killip | Yaax pooch | 2.400 | Y | |
| Polygonaceae | Coccoloba acapulcensis Standl. | Boob/Toyub | 0.220 | N |
| Putranjivaceae | Drypetes lateriflora (Sw.) Krug & Urb. | Ejuleb | 1.910 | Y |
| Rhamnaceae | Krugiodendrom ferraum (Vahl) Urb. | Chintoc | 0.100 | N |
| Rubiaceae | Cosmocalyx spectabilis Standl. | Chactecook | 0.164 | N |
| Randia truncata Greenm. & C.H.Thomps. | Kaakalche | 0.400 | N | |
| Exostema mexicanum A Gray | Sabasche | 0.395 | N | |
| Guettarda combsii Urb. | Tastab | 0.270 | N | |
| Morinda citrifolia L. | Noni | 3.500 | N | |
| Rutaceae | Citrus aurantium L. | Naranja agria | 6.270 | Y |
| Esenbeckia pentaphylla (Macfad.) Griseb. | Narnaha che | 2.630 | N | |
| Citrus sinensis (L.) Osbek | Naranja dulce | 3.310 | Y | |
| Casimiroa microcarpa Lundell | Yuuy | 7.300 | Y | |
| Salicaceae | Laetia thamnia L. | Chauche | 3.141 | Y |
| Casearia corymbosa Kunth | Ixiimche | 0.424 | N | |
| Zuelania guidonia (Sw.) Britton & Millsp. | Tamay | 5.672 | Y | |
| Sapindaceae | Blomia prisca (Standl.) Lundell | Tzol | 4.900 | Y |
| Cupania belizensis Standl. | Sal poom | 1.200 | N | |
| Thouinia paucidentata Radlk. | Kanchunup | 0.136 | N | |
| Melicoccus bijugatus Jacq. | Guaya | 1.200 | N | |
| Matayba oppositifolia (A. Rich.) Britton | Ikche | 0.370 | N | |
| Allophylus camptostachys Radlk. | Kanchunup | 0.783 | N | |
| Talisia oliviformis (Kunth) Radlk. | Wayum | 1.380 | N | |
| Sapotaceae | Manilkara zapota (L.) Van Royen | Chicozapote | 4.200 | Y |
| Chrysophyllum cainito L. | Cayumito | 2.800 | Y | |
| Chrysophyllum mexicanum Brandegee ex Standl. | Chique | 0.690 | Y | |
| Pouteria campechiana (Kunth) Baehni | Kaniste | 9.393 | Y | |
| Pouteria sapota (Jacq.) H. E. Moore and Stearn | Hazz | 3.500 | Y | |
| Sideroxylon capiri subsp. tempisque (Pittier) T.D. Penn. | Subul | 6.554 | Y | |
| Sideroxylon foetidissimum subsp. gaumeri (Pittier) T.D. Penn. | Tsiimimche | 0.800 | N | |
| Pouteria glomerata (Miq.) Radlk. | Zapote del pueblo | 9.935 | Y | |
| Pouteria reticulata (Engl.) Eyma | Zapotillo | 5.220 | Y | |
| Simaroubaceae | Simarouba glauca DC. | Paasac | 0.360 | N |
| Verbenaceae | Vitex gaumeri Greenm. | Yaxnic | 9.096 | Y |
| Host Family | Host Scientific Name | Fruit Fly Species | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anacardiaceae | Mangifera indica L. | A. obliqua | · | · | ||||||||||
| Spondias purpurea L. | A. obliqua | · | · | |||||||||||
| Spondias mombin L. | A. obliqua | · | · | · | ||||||||||
| Caricaceae | Carica papaya L. | A. curvicauda | · | · | · | · | · | |||||||
| Myrtaceae | Psidium guajava L. | A. striata | · | · | · | · | · | |||||||
| Psidium guajava | A. fraterculus | · | · | · | · | · | ||||||||
| Psidium sartorianum (O. Berg) Nied. | A. fraterculus | · | · | · | · | · | ||||||||
| Putranjivaceae | *Drypetes lateriflora (Sw.) Krug & Urb. | A. fraterculus | · | · | · | · | · | · | ||||||
| Sapindaceae | *Blomia prisca (Standl.) Lundell | A. fraterculus | · | · | ||||||||||
| Passifloraceae | *Passiflora yucatanensis Killip | A. chiclayae | · | · | · | · | · | |||||||
| *Passiflora serratifolia L. | A. chiclayae | · | · | · | · | |||||||||
| *Passiflora foetida L. | A. chiclayae | · | · | · | ||||||||||
| Rutaceae | Citrus aurantium L. | A. ludens | · | · | · | · | · | · | ||||||
| Citrus sinensis (L.) Osbek | A. ludens | · | · | · | · | |||||||||
| *Casimiroa microcarpa Lundell | A. ludens | · | · | · | ||||||||||
| Salicaceae | *Laetia thamnia L. | A. zuelaniae | · | · | · | |||||||||
| Zuelania guidonia (Sw.) Britton & Millsp. | A. zuelaniae | · | · | · | ||||||||||
| Sapotaceae | Chrysophyllum mexicanum Brandegee ex Standl. | A. serpentina | · | · | · | |||||||||
| Chrysophyllum cainito L. | A. serpentina | · | · | · | ||||||||||
| Manilkara zapota (L.) Van Royen | A. serpentina | · | · | · | · | · | · | · | · | · | · | · | · | |
| Pouteria campechiana Baehni | A. serpentina | · | · | · | · | · | · | · | · | · | ||||
| Pouteria glomerata (Miq.) Radlk. | A. serpentina | · | · | · | · | · | · | · | ||||||
| *Pouteria reticulata (Engl.) Eyma | A. serpentina | · | · | · | · | · | · | |||||||
| Pouteria sapota (Jacq.) H.E. Moore and Stearn. | A. serpentina | · | · | |||||||||||
| Sideroxylon capiri subsp. tempisque (Pittier) T.D. Penn. | A. serpentina | · | · | · | · | |||||||||
| Pouteria campechiana Baehni | A. hamata | · | · | · | · | · | · | · | · | · | ||||
| *Pouteria reticulata | Anastrepha sp. 1 | · | · | · | · | · | · | |||||||
| Verbenaceae | *Vitex gaumeri Greenm. | A. ampliata | · | · | · | · | · | · |
| Host Family | Host Scientific Name | Fruit Fly Species | Recovered Pupae | Number of Pupae/Kg of Fruit | Sex Ratio (F/M) | Emergence % |
|---|---|---|---|---|---|---|
| Anacardiaceae | Mangifera indica | A. obliqua (Macquart) | 55 | 1.20 | 28/19 | 85.45 |
| Spondias mombin | A. obliqua | 280 | 17.28 | 73/61 | 47.86 | |
| Spondias purpurea | A. obliqua | 106 | 9.64 | 37/49 | 81.13 | |
| Caricaceae | Carica papaya | A. curvicauda (Gerstaecker) | 83 | 5.53 | 31/29 | 72.29 |
| Myrtaceae | Psidium guajava | A. striata Schiner | 92 | 7.51 | 4/5 | 9.78 |
| A. fraterculus (Wiedemann) | -- | 19/16 | 38.04 | |||
| Psidium sartorianum | A. fraterculus | 21 | 2.37 | 14/3 | 80.95 | |
| Putranjivaceae | Drypetes lateriflora | A. fraterculus | 142 | 7.43 | 43/47 | 63.38 |
| Sapindaceae | Blomia prisca | A. fraterculus | 720 | 14.69 | 358/302 | 91.67 |
| Passifloraceae | Passiflora foetida | A. chiclayae Greene | 9 | 6.00 | 4/3 | 77.78 |
| Passiflora serratifolia | A. chiclayae | 37 | 6.61 | 21/15 | 97.3 | |
| Passiflora yucatanensis | A. chiclayae | 5 | 0.21 | 3/1 | 80.0 | |
| Rutaceae | Citrus aurantium | A. ludens (Loew) | 278 | 4.43 | 128/123 | 90.29 |
| Citrus sinensis | A. ludens | 68 | 2.05 | 31/25 | 82.35 | |
| Casimiroa microcarpa | A. ludens | 383 | 5.24 | 191/162 | 92.17 | |
| Salicaceae | Laetia thamnia | A. zuelaniae Stone | 199 | 6.34 | 89/76 | 82.91 |
| Zuelania guidonia | A. zuelaniae | 180 | 3.17 | 61/52 | 62.78 | |
| Sapotaceae | Chrysophyllum cainito | A. serpentina (Wiedemann) | 64 | 2.28 | 36/19 | 85.94 |
| Chrysophyllum mexicanum | A. serpentina | 12 | 1.74 | 5/6 | 91.67 | |
| Manilkara zapota | A. serpentina | 342 | 8.14 | 110/129 | 71.13 | |
| Pouteria campechiana | A. serpentina | 117 | 1.25 | 64/50 | 97.44 | |
| Pouteria glomerata | A. serpentina | 126 | 1.27 | 60/57 | 92.86 | |
| Pouteria sapota | A. serpentina | 70 | 2.00 | 24/35 | 84.29 | |
| Pouteria reticulata | A. serpentina | 738 | 19.17 | 185/168 | 47.83 | |
| Sideroxylon capiri subsp. tempisque | A. serpentina | 1195 | 18.23 | 605/433 | 86.86 | |
| Pouteria campechiana | A. hamata (Loew) | 172 | 1.83 | 95/68 | 94.77 | |
| Pouteria reticulata | Anastrepha sp. 1 | 265 | 6.88 | 102/116 | 82.26 | |
| Verbenaceae | Vitex gaumeri | A. ampliata Hernández-Ortiz | 633 | 6.96 | 197/216 | 65.24 |
| Family | Host Plant Scientific Name | Anastrepha Species | Recovered Fruit Fly Pupae | Parasitoid Species | Parasitoid Sex Ratio (F/M) | Total No. Parasitoids | % Parasitism |
|---|---|---|---|---|---|---|---|
| Anacardiaceae | Mangifera indica | A. obliqua | 55 | Doryctobracon areolatus (Szépligeti) | 1/2 | 3 | 5.45 |
| Spondias mombin | 280 | D. areolatus | 38/25 | ||||
| Utetes anastrephae (Viereck) | 15/21 | 99 | 35.36 | ||||
| Spondias purpurea | 106 | D. areolatus | 10/8 | 18 | 16.98 | ||
| Myrtaceae | Psidium guajava | A. fraterculus | 92 | D. areolatus | 8/5 | ||
| Aganaspis pelleranoi (Brethes) | 7/2 | ||||||
| Odontosema anastrephae Borgmeier | 2/1 | 25 | 27.17 | ||||
| Putranjivaceae | Drypetes lateriflora | 142 | D. areolatus | 5/3 | |||
| U. anastrephae | 3/4 | ||||||
| Opius hirtus (Fisher) | 2/1 | ||||||
| A. pelleranoi | 8/12 | 38 | 26.76 | ||||
| Sapindaceae | Blomia prisca | 720 | D. areolatus | 27/30 | |||
| U. anastrephae | 12/11 | ||||||
| O. hirtus | 7/4 | 91 | 12.64 | ||||
| Salicaceae | Laetia thamnia | A. zuelaniae | 199 | D. areolatus | 13/17 | ||
| O. hirtus | 2/0 | 32 | 16.08 | ||||
| Zuelania guidonia | 180 | D. areolatus | 12/7 | ||||
| A. pelleranoi | 4/3 | 26 | 14.44 | ||||
| Sapotaceae | Manilkara zapota | A. serpentina | 342 | D. areolatus | 16/17 | ||
| O. hirtus | 4/2 | ||||||
| A. pelleranoi | 22/15 | ||||||
| O. anastrephae | 3/1 | 80 | 23.81 | ||||
| Pouteria reticulata | 738 | D. areolatus | 29/31 | ||||
| U. anastrephae | 48/38 | ||||||
| O. hirtus | 28/21 | ||||||
| A. pelleranoi | 37/21 | 263 | 35.54 | ||||
| Sideroxylon capiri subsp. tempisque | 1195 | D. areolatus | 35/39 | ||||
| O. hirtus | 11/4 | ||||||
| A. pelleranoi | 1/2 | 112 | 9.37 | ||||
| Sapotaceae | Pouteria campechiana | A. hamata | 172 | Doryctobracon zeteki Musebeck | 5/1 | 6 | 3.49 |
| Pouteria reticulata | Anastrepha sp. | 265 | D. areolatus | 7/12 | 7.17 | ||
| Verbenaceae | Vitex gaumeri | A. ampliata | 633 | D. areolatus | 58/51 | ||
| U. anastrephae | 15/11 | ||||||
| O. hirtus | 27/25 | 187 | 29.54 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Ortega, M.; Díaz-Fleischer, F.; Piñero, J.C.; Valdez-Lazalde, J.R.; Hernández-Ortiz, M.; Hernández-Ortiz, V. The Mayan Tropical Rainforest: An Uncharted Reservoir of Tritrophic Host-Fruit Fly-Parasitoid Interactions. Insects 2020, 11, 495. https://doi.org/10.3390/insects11080495
López-Ortega M, Díaz-Fleischer F, Piñero JC, Valdez-Lazalde JR, Hernández-Ortiz M, Hernández-Ortiz V. The Mayan Tropical Rainforest: An Uncharted Reservoir of Tritrophic Host-Fruit Fly-Parasitoid Interactions. Insects. 2020; 11(8):495. https://doi.org/10.3390/insects11080495
Chicago/Turabian StyleLópez-Ortega, Maurilio, Francisco Díaz-Fleischer, Jaime C. Piñero, José René Valdez-Lazalde, Manuel Hernández-Ortiz, and Vicente Hernández-Ortiz. 2020. "The Mayan Tropical Rainforest: An Uncharted Reservoir of Tritrophic Host-Fruit Fly-Parasitoid Interactions" Insects 11, no. 8: 495. https://doi.org/10.3390/insects11080495