
Anatomy and Ultrastructure of Galls Induced by Neuroterus quercusbaccarum (Hymenoptera: Cynipidae) on Oak Leaves (Quercus robur)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Gall Collection
2.2. Cytochemical Investigations in Light (LM) and Fluorescence (FM) Microscopy
2.3. Scanning Electron Microscopy (SEM)
2.4. Transmission Electron Microscopy (TEM)
3. Results
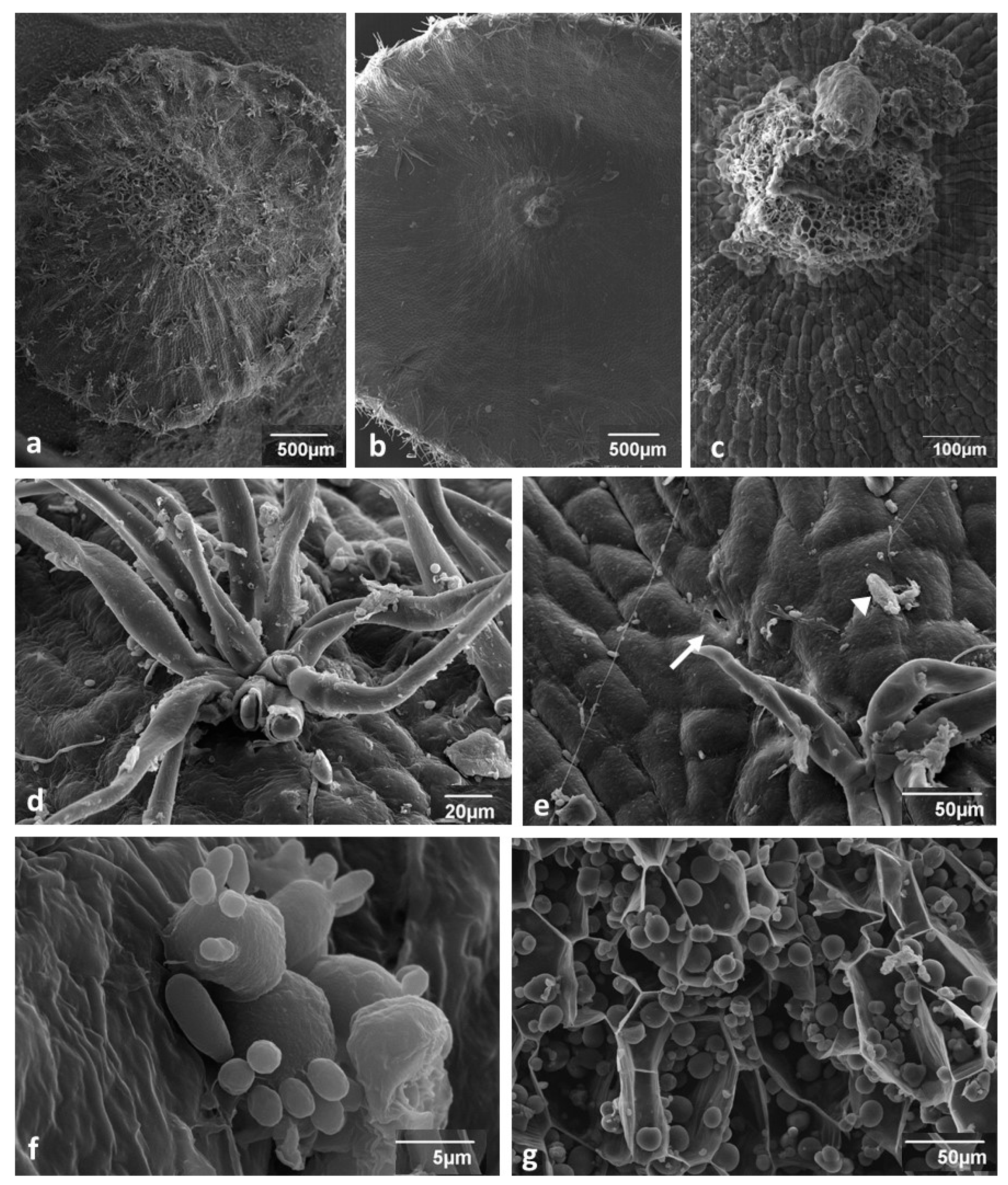
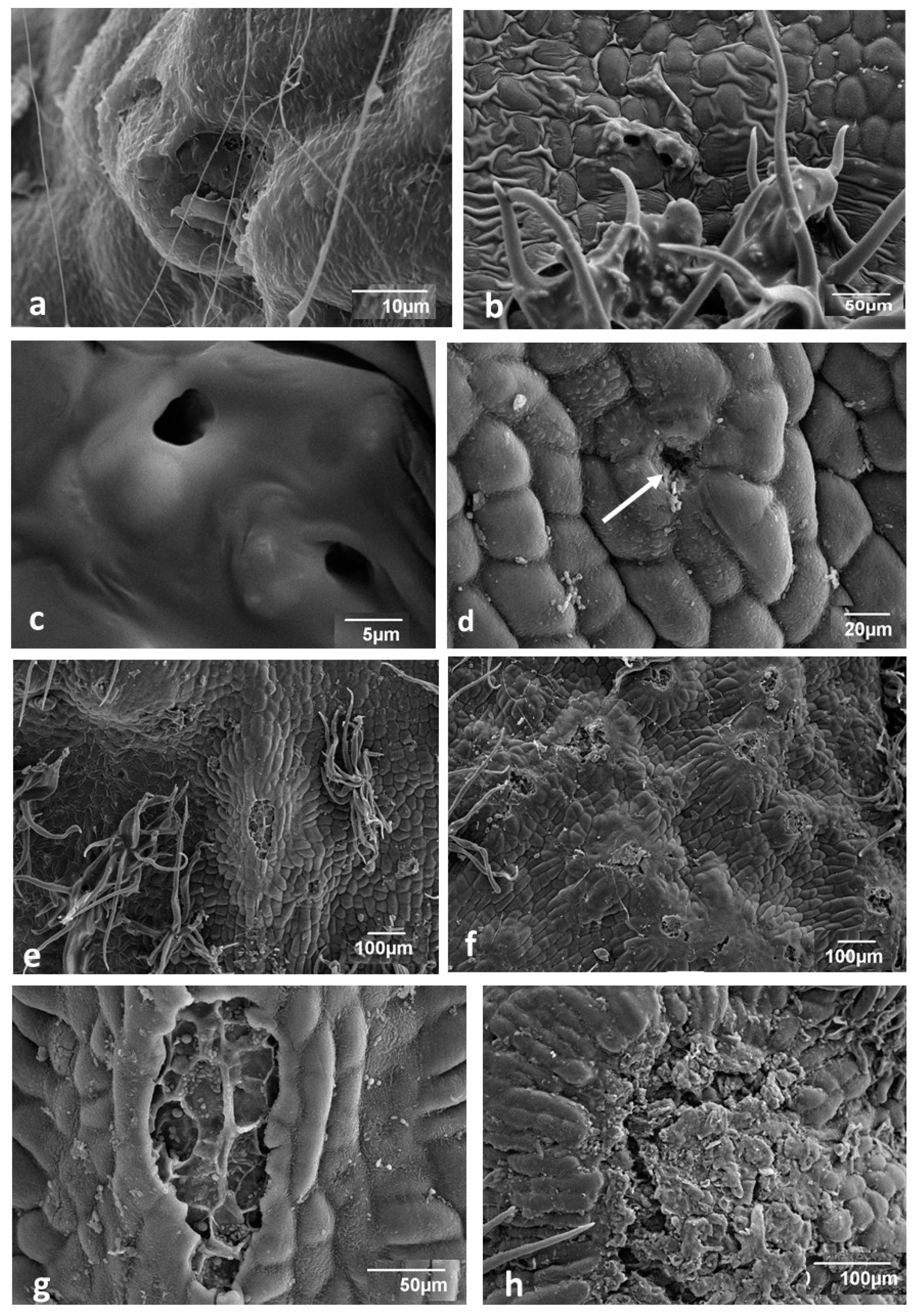
3.1. Young Gall Collected in July—SEM Observations
3.2. Changes in Gall Surface during Development—SEM Observations
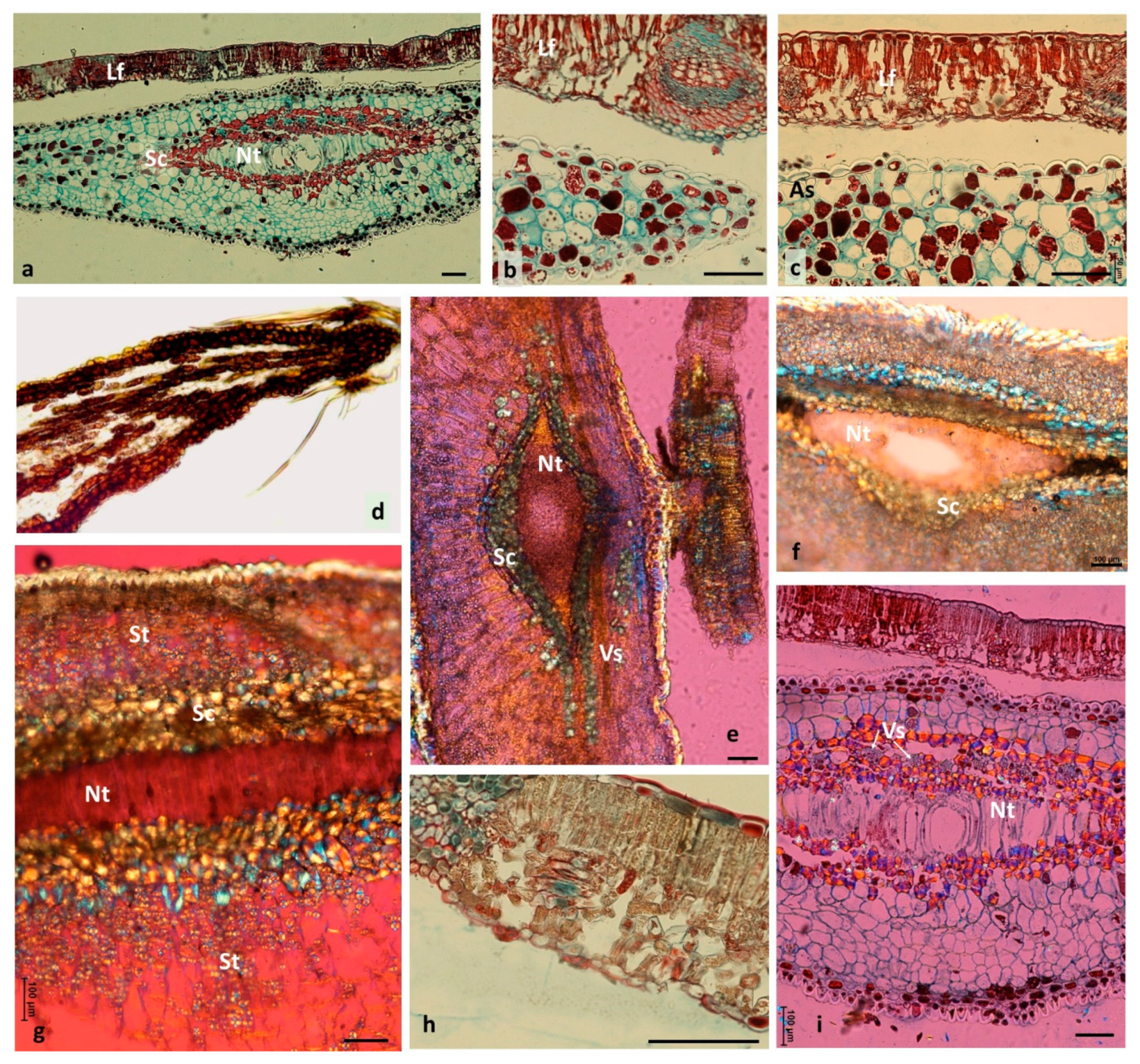
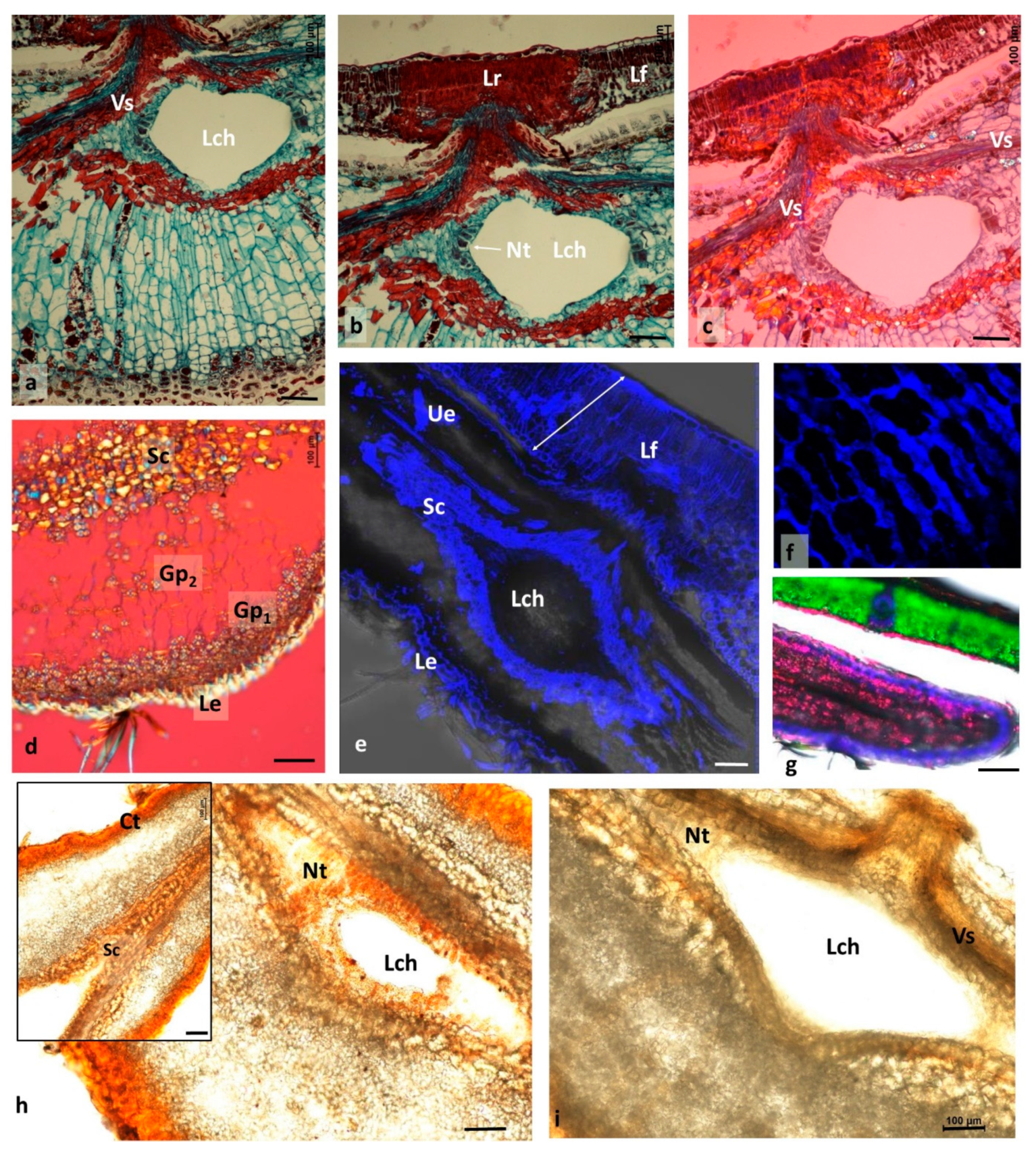
3.3. Changes Accompanying Gall Maturation (FM, LM—Polarized Light)
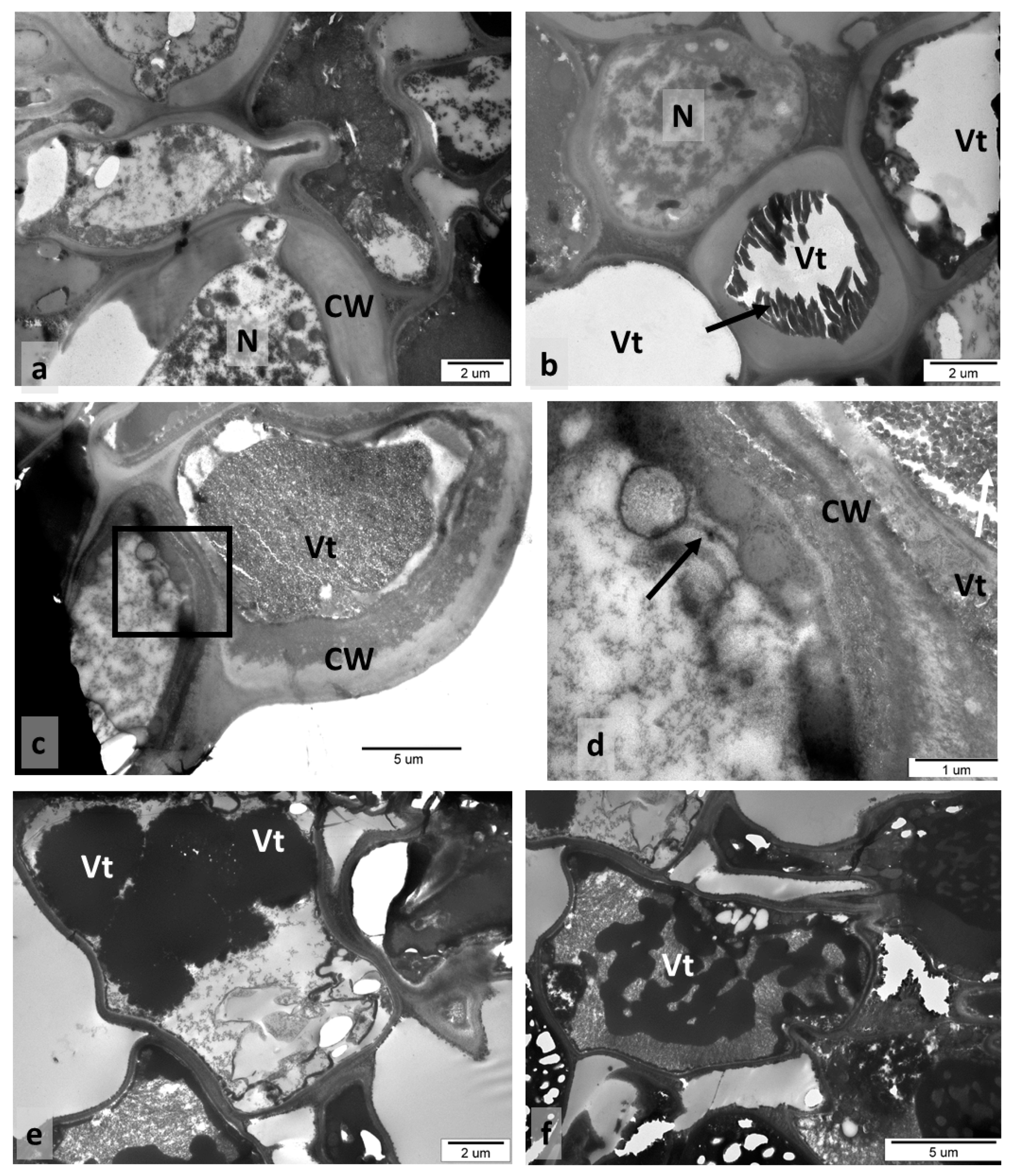
3.4. Older Galls Collected in Autumn—TEM Observations
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maresquelle, H.J.; Meyer, J. Physiologie et morphogenése des galles d’origine animale (zoocécidies). Handb. Pflanzephysiol. 1965, 15, 280–329. [Google Scholar]
- Mani, M.S. Ecology of Plant Galls, 1st ed.; Monographiae Biologicae; Dr W. Junk Publishers: Hague, The Netherlands, 1964; p. 434. [Google Scholar]
- Meyer, J.; Maresquelle, H.J. Anatomie des Galles; Handbuch der Pflanzenanatomie; Gebrüder Borntraeger: Stuttgart, Germany, 1983; p. 662. [Google Scholar]
- Raman, A. Insect-plant interaction: The gall factor. In All Flesh is Grass: Plant-Animal Interactions; Seckbach, J., Dubinsky, Z., Eds.; Springer: Heidelberg, Germany, 2010; pp. 119–146. [Google Scholar]
- Raman, A. Morphogenesis of insect-induced plant galls: Facts and questions. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 517–533. [Google Scholar] [CrossRef]
- Giron, D.; Huguet, E.; Stone, G.N.; Body, M. Insect-induced effects on plants. J. Insect Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Kopcewicz, J.; Lewak, S. (Eds.) Fizjologia Roślin; Wydawnictwo Naukowe PWN: Warsaw, Poland, 2015; p. 210. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 1998. [Google Scholar]
- Harper, L.J.; Schönrogge, K.; Lim, K.Y.; Francis, P.; Lichtenstein, C.P. Cynipid galls: Insect-induced modifications of plant development create novel plant organs. Plant Cell Environ. 2004, 27, 327–335. [Google Scholar] [CrossRef]
- Hearn, J.; Blaxter, M.; Schönrogge, K.; Nieves-Aldrey, J.-L.; Pujade-Villar, J.; Huguet, E.; Drezen, J.-M.; Shorthouse, J.D.; Stone, G.N. Genomic dissection of an extended phenotype: Oak galling by a cynipid gall wasp. PLoS Genet. 2019, 15, e1008398. [Google Scholar] [CrossRef] [Green Version]
- Schönrogge, K.; Harper, L.J.; Lichtenstein, C.P. The protein content of tissues in cynipid galls (Hymenoptera: Cynipidae): Similarities between cynipid galls and seeds. Plant Cell Environ. 2000, 23, 215–222. [Google Scholar] [CrossRef]
- Sliva, M.D.; Shorthouse, J.D. Comparison of the development of stem galls induced by Aulacidea hieracii (Hymenoptera: Cynipidae) on hawkweed and by Diplolepis spinosa (Hymenoptera: Cynipidae) on rose. Can. J. Bot. 2011, 84, 1052–1074. [Google Scholar] [CrossRef]
- Bronner, R. The role of nutritive cells in the nutrition of Cynipids and Cecidomyiidae. In Biology of Insect Induced Galls; Shorthouse, J.D., Rohfritsch, O., Eds.; Oxford University Press: New York, NY, USA, 1992; pp. 118–140. [Google Scholar]
- Bronner, R. Propriétés lytiques des oeufs de Biorhiza pallida OL. Acad. Sci. Paris. CR Ser. D 1973, 276, 189–192. [Google Scholar]
- Madden, J.; Stone, C. Induction and formation of pouch and emergence galls in Eucalyptus pulchella leaves. Aust. J. Bot. 1984, 32, 33–42. [Google Scholar] [CrossRef]
- Barnewell, E.C.; de Clerck-Floate, R.A. A preliminary histological investigation of gall induction in an unconventional galling system. Arthropod-Plant Interact. 2012, 6, 449–459. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Tanaka, H.; Hasegawa, M.; Tokuda, M.; Asami, T.; Suzuki, Y. Phytohormones and willow gall induction by a gall-inducing sawfly. New Phytol. 2012, 196, 586–595. [Google Scholar] [CrossRef]
- Bartlett, L.; Connor, E.F. Exogenous phytohormones and the induction of plant galls by insects. Arthropod-Plant Interact. 2014, 8, 339–348. [Google Scholar] [CrossRef]
- Egan, S.P.; Hood, G.R.; Martinson, E.O.; Ott, J.R. Cynipid gall wasps. Curr. Biol. 2018, 28, R1370–R1374. [Google Scholar] [CrossRef] [Green Version]
- Stone, G.N.; Schonrogge, K.; Atkinson, R.J.; Bellido, D.; Pujade-Villar, J. The population biology of oak gall wasps (Hymenoptera: Cynipidae). Annu. Rev. Entomol. 2002, 47, 633–668. [Google Scholar] [CrossRef] [Green Version]
- Cornell, H.V. The secondary chemistry and complex morphology of galls formed by the Cynipinae (Hymenoptera): Why and how? Am. Midl. Nat. 1983, 110, 225. [Google Scholar] [CrossRef]
- Cambier, S.; Ginis, O.; Moreau, S.J.M.; Gayral, P.; Hearn, J.; Stone, G.N.; Giron, D.; Huguet, E.; Drezen, J.-M. gall wasp transcriptomes unravel potential effectors involved in molecular dialogues with oak and rose. Front. Physiol. 2019, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Jankiewicz, L.S.; Dyki, B.; Machlańska, A.; Dubert, F. Oak leaf galls: Neuroterus numismalis and Cynips quercusfolii, their structure and ultrastructure. Acta Soc. Bot. Pol. 2017, 86. [Google Scholar] [CrossRef] [Green Version]
- Gätjens-Boniche, O. The mechanism of plant gall induction: Revealing clues, facts and consequences in a cross-kingdom complex integration. Rev. Biol. Trop. 2019, 67, 1359–1382. [Google Scholar] [CrossRef]
- Wrzesińska, D. Foliofagi Tworzące Wyrośla na Quercus robur (L.); Rozprawy; Wydawnictwa Uczelniane Uniwersytetu Technologiczno-Przyrodniczego: Bydgoszcz, Poland, 2013; Volume 167. (In Polish) [Google Scholar]
- Redfern, M. Plant Galls; Harper Collins Publishers: London, UK, 2011. [Google Scholar]
- Hough, J.S. Studies on the common spangle gall of oak I. The developmental history. New Phytol. 1953, 52, 149–177. [Google Scholar] [CrossRef]
- Kot, I.; Sempruch, C.; Rubinowska, K.; Michałek, W. Effect of Neuroterus quercusbaccarum (L.) galls on physiological and biochemical response of Quercus roburleaves. Bull. Entomol. Res. 2020, 110, 34–43. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Isaias, R.M.S.; Fernandes, G.W.; Ferreira, B.G.; Carneiro, R.G.S.; Fuzaro, L. Manipulation of host plant cells and tissues by gall-inducing insects and adaptive strategies used by different feeding guilds. J. Insect Physiol. 2016, 84, 103–113. [Google Scholar] [CrossRef]
- Kovácsné Koncz, N.; Szabó, L.; Máthé, C.; Jámbrik, K.; Mikóné Hamvas, M. Histological study of quercus galls of Neuroterus quercusbaccarum (Linnaeus 1758) (Hymenoptera: Cynipidae). Acta Biol. Szeged. 2011, 55, 247–253. [Google Scholar]
- Gerlach, D. Zarys Mikrotechniki Botanicznej; Powszechne Wydawnictwo Rolnicze i Leśne (PWRiL): Warsaw, Poland, 1972. [Google Scholar]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999; p. 322. [Google Scholar]
- Müller-Taubenberger, A.; Ishikawa-Ankerhold, H.C. Fluorescent reporters and methods to analyse fluorescent signals. Singer Protoc. 2013, 983, 93–112. [Google Scholar]
- García-Plazaola, J.I.; Fernández-Marin, B.; Duke, S.O.; Hernandez, A.; Lopez-Arbeloa, F.; Becerril, J.M. Autofluorescence: Biological functions and technical applications. Plant Sci. 2015, 236, 136–145. [Google Scholar] [CrossRef]
- Donaldson, L. Autofluorescence in plants. Molecules 2020, 25, 2393. [Google Scholar] [CrossRef]
- Pathan, A.; Bond, J.; Gaskin, R. Sample preparation for scanning electron microscopy of plant surfaces—Horses for courses. Micron 2008, 39, 1049–1061. [Google Scholar] [CrossRef]
- Mercer, E.H.; Birback, M.S.C. Electron Microscopy; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1970; p. 117. [Google Scholar]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as electron opaque stain for electron microscopy. J. Cell Biol. 1963, 17, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, 2nd ed.; Clarendon Press: Oxford, UK, 1979; Volume 1, p. 276. [Google Scholar]
- Dyki, B.; Jankiewicz, L.S.; Staniaszek, M. Anatomical structure and surface micromorphology of tomatillo leaf and flower (Physalis ixocarpa Brot., Solanaceae). Acta Soc. Bot. Pol. 2014, 67, 181–191. [Google Scholar] [CrossRef]
- Guzicka, M.; Karolewski, P.; Giertych, M.J. Structural modification of Quercus petrea leaf caused by Cynips quercusfolii—Histological study of galls. J. Plant Interact. 2017, 12, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Nakata, P. Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Horner, H.T. Calcium oxalate crystals in plants. Bot. Rev. 1980, 46, 361–427. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Nakata, P. Calcium oxalate in plants: Formation and function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Tooulakou, G.; Giannopoulos, A.; Nikolopoulos, D.; Bresta, P.; Dotsika, E.; Orkoula, M.G.; Kontoyannis, C.G.; Fasseas, C.; Liakopoulos, G.; Klapa, M.I.; et al. Alarm photosynthesis: Calcium oxalate crystals as an internal CO2 source in plants. Plant Physiol. 2016, 171, 2577–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korth, K.L.; Doege, S.J.; Park, S.-H.; Goggin, F.L.; Wang, Q.; Gomez, S.K.; Liu, G.; Jia, L.; Nakata, P. Medicago truncatula mutants demonstrate the role of plant calcium oxalate crystals as an effective defense against chewing insects. Plant Physiol. 2006, 141, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esau, K. Plant Anatomy; John Willey & Sons: New York, NY, USA, 1967. [Google Scholar]
- Brillouet, J.-M.; Romieu, C.; Lartaud, M.; Jublanc, E.; Torregrosa, L.; Cazevieille, C. Formation of vacuolar tannin deposits in the chlorophyllous organs of Tracheophyta: From shuttles to accretions. Protoplasma 2014, 251, 1387–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleurat-Lessard, P.; Béré, E.; Lallemand, M.; Dédaldéchamp, F.; Roblin, G. Co-occurrence of tannin and tannin-less vacuoles in sensitive plants. Protoplasma 2016, 253, 821–834. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Constabel, C.P. Tannins in plant—Herbivore interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef]
- González-Lamothe, R.; Mitchell, G.; Gattuso, M.; Diarra, M.S.; Malouin, F.; Bouarab, K. Plant antimicrobial agents and their effects on plant and human pathogens. Int. J. Mol. Sci. 2009, 10, 3400–3419. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Dhiman, S.; Mukherjee, G.D.S.; Singh, A.K. Recent trends and advancement in microbial tannase-catalysed biotransformation of tannins: A review. Int. Microbiol. 2018, 21, 175–195. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankiewicz, L.S.; Guzicka, M.; Marasek-Ciolakowska, A. Anatomy and Ultrastructure of Galls Induced by Neuroterus quercusbaccarum (Hymenoptera: Cynipidae) on Oak Leaves (Quercus robur). Insects 2021, 12, 850. https://doi.org/10.3390/insects12100850
Jankiewicz LS, Guzicka M, Marasek-Ciolakowska A. Anatomy and Ultrastructure of Galls Induced by Neuroterus quercusbaccarum (Hymenoptera: Cynipidae) on Oak Leaves (Quercus robur). Insects. 2021; 12(10):850. https://doi.org/10.3390/insects12100850
Chicago/Turabian StyleJankiewicz, Leszek Stanisław, Marzenna Guzicka, and Agnieszka Marasek-Ciolakowska. 2021. "Anatomy and Ultrastructure of Galls Induced by Neuroterus quercusbaccarum (Hymenoptera: Cynipidae) on Oak Leaves (Quercus robur)" Insects 12, no. 10: 850. https://doi.org/10.3390/insects12100850