Characteristics of Immunoglobulin M Type Antibodies of Different Origins from the Immunologic and Clinical Viewpoints and Their Application in Controlling Antibody-Mediated Allograft Rejection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Molecular Pathophysiology
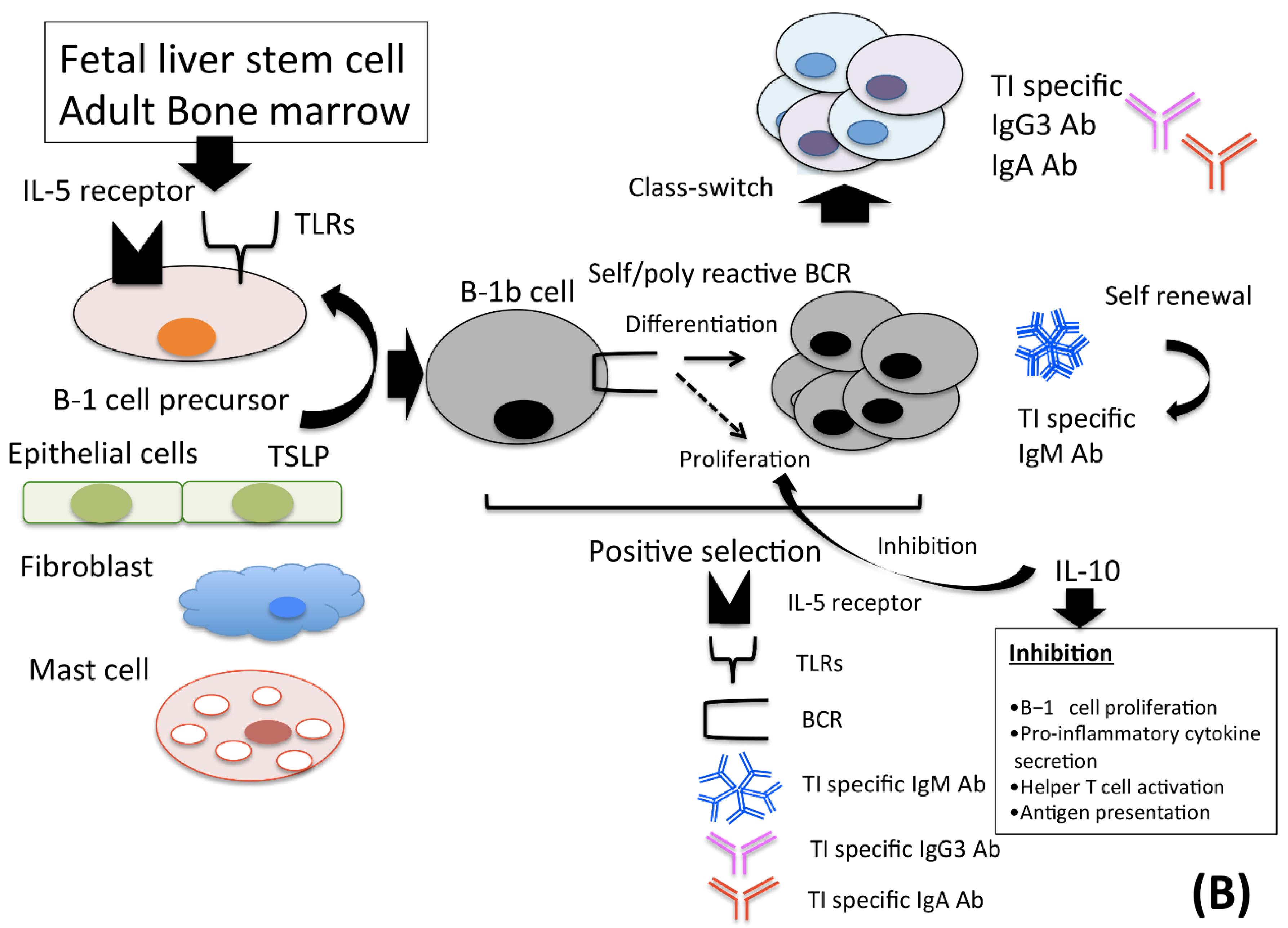
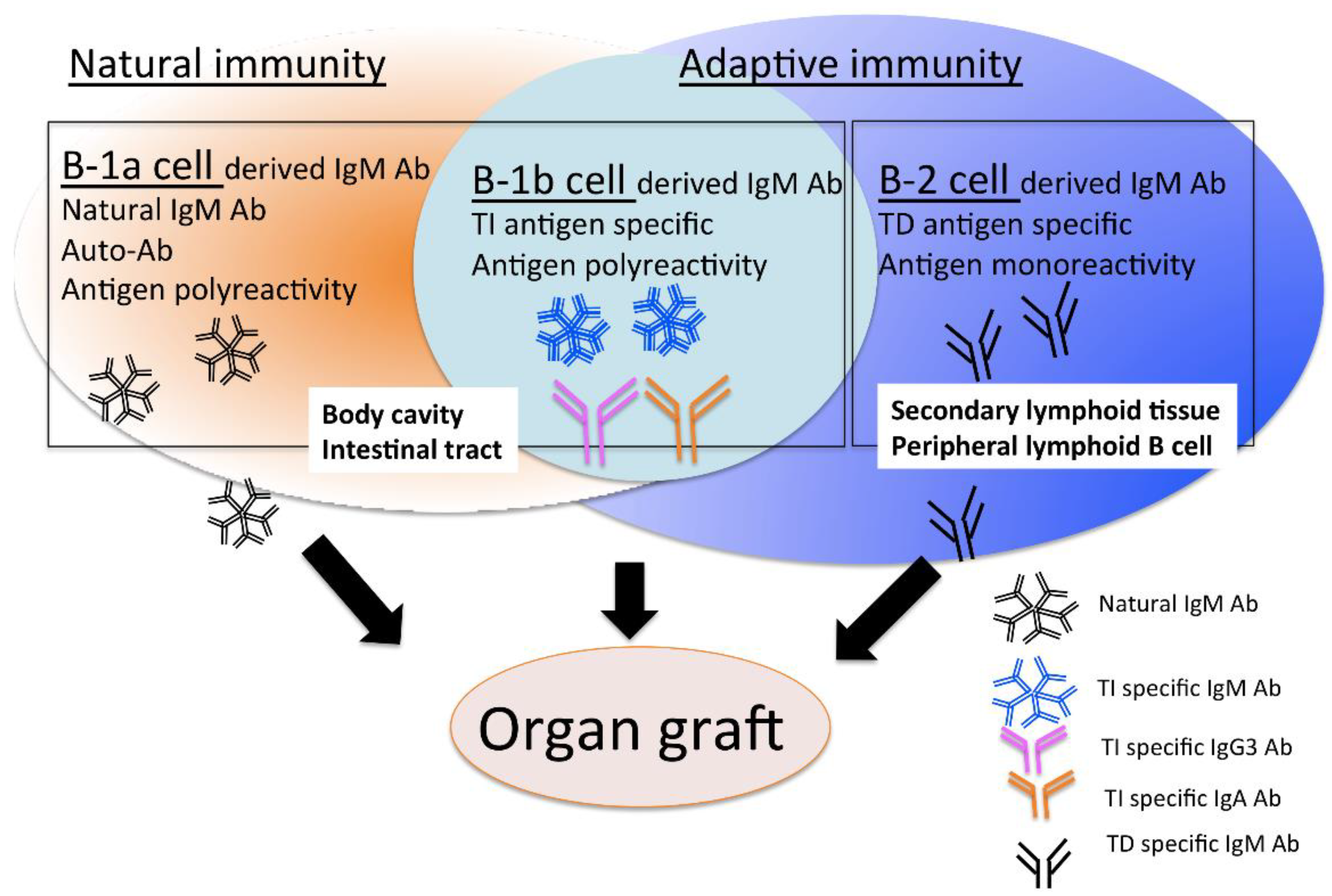
2.1. B-1 Cells
2.2. B-2 Cells
3. The Role of IgM Antibodies in Pathology
3.1. B-1 Cell-Derived Natural IgM Antibodies
3.2. The Role of B-2 Cell-Derived IgM Antibodies in Pathology
3.3. Involvement of IgM Antibodies in Complement Activation
3.4. Involvement of IgM Antibodies in Immune Responses via the IgM Receptor (FcµR)
4. Significance of IgM Antibodies in Various Disease States
4.1. Infection
4.2. Autoimmunity
4.3. Cancer
4.4. Clinical Significance of IgM-Based Therapies against Inflammation and Infection
5. The Role of IgM Antibodies against Donor-Specific HLA in the Field of Transplantation
5.1. Conventional Policy and Problems
5.2. Effect of IgM DSA on Transplanted Organ
5.3. Challenging Problems
6. ABO Blood Group-Incompatible Antibodies in the Field of Transplantation
6.1. Kidney Transplants
6.1.1. Development of Immunosuppressive Therapy for ABO-Incompatible Kidney Transplants
6.1.2. Current Outcomes of ABO-Incompatible Kidney Transplants
6.2. Liver Transplants
6.2.1. Development of Immunosuppressive Therapy for ABO-Incompatible Liver Transplants
6.2.2. Current Outcomes of ABO-Incompatible Liver Transplants
6.3. The Role of IgM Antibodies in ABO-Incompatible Transplant Survival
6.4. Challenging Problems in ABO-Incompatible Transplantation
7. Significance of Complement Activation in the Field of Transplantation
8. Clinical Significance of IgM-Based Therapies in the Field of Transplantation
9. The Possibilities of IgM Antibody Manufacturing for AMR Control
9.1. B-1 Cell-Derived Natural IgM Antibodies
9.2. B-2 Cell-Derived IgM Antibody
9.3. FcµR
9.4. Current Status of the Clinical Application of IgM Antibodies of Different Origins as a Standard Diagnostic and Therapeutic Entity
10. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- De Silva, N.S.; Klein, U. Dynamics of B cells in germinal centres. Nat. Rev. Immunol. 2015, 15, 137–148. [Google Scholar] [CrossRef] [Green Version]
- McHeyzer-Williams, L.J.; Malherb, L.P.; McHeyzer-Williams, M.G. Helper T cell-regulated B cells immunity. Curr. Top. Microbiol. Immunol. 2006, 311, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Marrari, M.; Duquesnoy, R.J. Detection of donor-specific HLA antibodies before and after removal of a rejected kidney transplant. Transpl. Immunol. 2010, 22, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Visentin, J.; Guidicelli, G.; Couzi, L.; Merville, P.; Lee, J.H.; Di Primo, C.; Taupin, J.L. Deciphering IgM interference in IgG anti-HLA antibody detection with flow beads assays. Hum. Immunol. 2016, 77, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Everly, M.J.; Rebellato, L.M.; Haisch, C.E.; Briley, K.P.; Bolin, P.; Kendrick, W.T.; Kendrick, S.A.; Morgan, C.; Maldonado, A.Q.; Harland, R.C.; et al. Impact of IgM and IgG3 anti-HLA alloantibodies in primary renal allograft recipients. Transplantation 2014, 97, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.; Andreou, A.; Briggs, D.; Krishnan, N.; Higgins, R.; Mitchell, D.; Barber, T.; Daga, S. Clinical Relevance of Donor-Specific IgM Antibodies in HLA Incompatible Renal Transplantation: A Retrospective Single-Center Study. Clin. Transpl. 2016, 32, 173–179. [Google Scholar] [PubMed]
- WarnerIdoia, P.; Danny, G.; Erin, Y.; AaronIng, D.; Arsenault, K.; Nelson, K. Are IgM donor-specific hla antibodies (DSA) clinically relevant? Hum. Immunol. 2020, 76, 138. [Google Scholar] [CrossRef]
- Küppers, R.; Klein, U.; Hansmann, M.L.; Rajewsky, K. Cellular origin of human B-cell lymphomas. N. Engl. J. Med. 1999, 341, 1520–1529. [Google Scholar] [CrossRef]
- Moras, B.L.E.; Teraasaki, P.I.; Marino-Vázquez, L.A.; Lee, J.H. Natural human leukocyte antigen antibodies found in non alloimmunized healthy males. Transplantation 2008, 86, 1111–1115. [Google Scholar] [CrossRef]
- Pozsonyi, E.; György, B.; Berki, T.; Bánlaki, Z.; Buzás, E.; Rajczy, K.; Hossó, A.; Prohászka, Z.; Szilágyi, A.; Cervenak, L.; et al. HLA-association of serum levels of natural antibodies. Mol. Immunol. 2009, 46, 1416–1423. [Google Scholar] [CrossRef]
- Urlacher, A.; Tongio, M.M.; Pasquali, J.L. IgM anti-idiotypes that block anti-HLA antibodies: Naturally occurring or immune antibodies? Clin. Exp. Immunol. 1991, 83, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Kosmoliatss, V.; Bradley, J.A.; Peacock, S.; Chaudhry, A.N.; Taylor, C.J. Detection of immunoglobulin G human leukocyte antigen-specific alloantibodies in renal transplant patients using single-antigen-beads is compromised by the presence of immunoglobulin M human leukocyte antigen-specific alloantibodies. Transplantation 2009, 87, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, U.S.; Bennet, W. ABO-Incompatible Liver Transplantation-A Review of the Historical Background and Results. Int. Rev. Immunol. 2019, 38, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Kim, J.M. Immunologic strategies and outcomes in ABO-incompatible living donor liver transplantation. Clin. Mol. Hepatol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, M.; Kawachi, S.; Obara, H.; Shinoda, M.; Hibi, T.; Kitagawa, Y.; Wakabayashi, G.; Shimazu, M.; Kitajima, M. Current progress in ABO-incompatible liver transplantation. J. Clin. Investig. 2010, 40, 943–949. [Google Scholar] [CrossRef]
- Takahashi, K.; Saito, K.; Takahara, S.; Okuyama, A.; Tanabe, K.; Toma, H.; Uchida, K.; Hasegawa, A.; Yoshimura, N.; Kamiryo, Y. Japanese ABO-Incompatible Kidney Transplantation Committee. Excellent long-term outcome of ABO-incompatible living donor kidney transplantation in Japan. Am. J. Transpl. 2004, 4, 1089–1096. [Google Scholar] [CrossRef]
- Montgomery, J.R.; Berger, J.C.; Warren, D.S.; James, N.T.; Montgomery, R.A.; Segev, D.L. Outcomes of ABO-incompatible kidney transplantation in the United States. Transplantation 2012, 93, 603–639. [Google Scholar] [CrossRef] [Green Version]
- Chow, K.V.; Flint, S.M.; Shen, A.; Landgren, A.; Finlay, M.; Murugasu, A.; Masterson, R.; Hughes, P.; Cohney, S.J. Histological and Extended Clinical Outcomes After ABO-Incompatible Renal Transplantation Without Splenectomy or Rituximab. Transplantation 2017, 101, 1433–1440. [Google Scholar] [CrossRef]
- Masterson, R.; Hughes, P.; Walker, R.G.; Hogan, C.; Haeusler, M.; Robertson, A.R.; Millar, R.; Suh, N.; Cohney, S. ABO incompatible renal transplantation without antibody removal using conventional immunosuppression alone. J. Am. J. Transpl. 2014, 14, 2807–2813. [Google Scholar] [CrossRef]
- Hanto, D.W.; Fecteau, A.H.; Alonso, M.H.; Valente, J.F.; Whiting, J.F. ABO-incompatible liver transplantation with no immunological graft losses using total plasma exchange, splenectomy, and quadruple immunosuppression: Evidence for accommodation. Liver. Transpl. 2003, 9, 22–30. [Google Scholar] [CrossRef]
- Lee, S.D.; Kim, S.H.; Kong, S.Y.; Kim, Y.K.; Lee, S.A.; Park, S.J. ABO-incompatible living donor liver transplantation without graft local infusion and splenectomy. HPB 2014, 16, 807–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.C.; Kim, S.H.; Park, S.J. Outcomes after liver transplantation in accordance with ABO compatibility: A systematic review and meta-analysis. World J. Gastroenterol. 2017, 23, 6516–6533. [Google Scholar] [CrossRef] [PubMed]
- Segev, D.L.; Simpkins, C.E.; Warren, D.S.; King, K.E.; Shirey, R.S.; Maley, W.R.; Melancon, J.K.; Cooper, M.; Kozlowski, T.; Montgomery, R.A. ABO Incompatible High-Titer Renal Transplantation without Splenectomy or Anti-CD20 Treatment. Am. J. Transpl. 2005, 5, 2570–2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, W.D.; Grande, J.P.; Ninova, D.; Nath, K.A.; Platt, J.L.; Gloor, J.M.; Stegall, M.D. Accommodation in ABO-Incompatible Kidney Allografts, a Novel Mechanism of Self-Protection Against Antibody-Mediated Injury. Am. J. Transpl. 2003, 3, 952–960. [Google Scholar] [CrossRef]
- Castro, L.; Yarur, A.; Segovia, R.; Ponce, R.; Uribe, M.; Ferrario, M.; Buckel, E.; Iturriaga, H. [Accomodation process: ABO-incompatible liver transplantation. Report of one case]. Rev. Med. Chil. 2009, 137, 1061–1065. [Google Scholar]
- Zhong, X.; Gao, W.; Degauque, N.; Bai, C.; Lu, Y.; Kenny, J.; Oukka, M.; Strom, T.B.; Rothstein, T.L. Reciprocal generation of Th1/Th17 and T(reg) cells by B1 and B2 B cells. Eur. J. Immunol. 2007, 37, 2400–2404. [Google Scholar] [CrossRef]
- Haas, K.M.; Poe, J.C.; Steeber, D.A.; Tedder, T.F. B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity 2005, 23, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Nicole, B. B-1 Cell Heterogeneity and the Regulation of Natural and Antigen-Induced IgM Production. Front. Immunol. 2016, 9, 324. [Google Scholar] [CrossRef] [Green Version]
- Vosshenrich, C.A.; Cumano, A.; Müller, W.; Di Santo, J.P.; Vieira, P. Pre-B cell receptor expression is necessary for thymic stromal lymphopoietin responsiveness in the bone marrow but not in the liver environment. Proc. Natl. Acad. Sci. USA 2004, 101, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Kouro, T.; Takatsu, K. IL-5- and eosinophil-mediated inflammation: From discovery to therapy. Int. Immunol. 2009, 12, 1303–1309. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Baumgarth, N. Natural IgM and the Development of B Cell-Mediated Autoimmune Diseases. Crit. Rev. Immunol. 2016, 36, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Baumgarth, N.; Herman, O.C.; Jager, G.C.; Brown, L.E.; Herzenberg, L.A.; Chen, J. B-1 and B-2 cell-derived immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J. Exp. Med. 2000, 192, 271–280. [Google Scholar] [CrossRef]
- Popi, A.F.; Longo-Maugéri, I.M.; Mariano, M. An Overview of B-1 Cells as Antigen-Presenting Cells. Front. Immunol. 2016, 7, 138. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Coligan, J.E.; Morse, H.C., 3rd. Emerging Functions of Natural IgM and Its Fc Receptor FCMR in Immune Homeostasis. Front. Immunol. 2016, 17, 99. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Zhurbenko, N.; Quach, T.D.; Hopkins, T.J.; Rothstein, T.L.; Hernandez, A.M. Human B-1 Cells and B-1 Cell Antibodies Change With Advancing Age. Front. Immunol. 2019, 10, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindhava, V.J.; Bondada, S. Multiple regulatory mechanisms control B-1 B cell activation. Front. Immunol. 2012, 3, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeney, A.J. Predominance of the prototypic T15 anti-phosphorylcholine junctional sequence in neonatal pre-B cells. J. Immunol. 1991, 147, 4343–4350. [Google Scholar]
- Eskdale, J.; Kube, D.; Tesch, H.; Gallagher, G. Mapping of the human IL10 gene and further characterization of the 5’ flanking sequence. Immunogenetics 1997, 46, 120–128. [Google Scholar] [CrossRef]
- Kessler, B.; Rinchai, D.; Kewcharoenwong, C.; Nithichanon, A.; Biggart, R.; Hawrylowicz, C.M.; Bancroft, G.J.; Lertmemongkolchai, G. Interleukin 10 inhibits pro-inflammatory cytokine responses and killing of Burkholderia pseudomallei. Sci. Rep. 2017, 7, 42791. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.G.; Ng, C.H.; Woehl, B.; Sutherland, A.P.; Huo, J.; Xu, S. BAFF costimulation of toll-like receptor-activated B-1 cells. Eur. J. Immunol. 2006, 36, 1837–1846. [Google Scholar] [CrossRef]
- Kantor, A.B.; Herzenberg, L.A. Origin of murine B cell lineages. Ann. Rev. Immunol. 1993, 11, 501–538. [Google Scholar] [CrossRef] [PubMed]
- Firmino-Cruz, L.; Ramos, T.D.; da Fonseca-Martins, A.M.; Oliveira-Maciel, D.; Oliveira-Silva, G.; Dos Santos, J.S.; Cavazzoni, C.; Morrot, A.; Gomes, D.C.O.; Vale, A.M.; et al. B-1 lymphocytes are able to produce IL-10, but is not pathogenic during Leishmania (Leishmania) amazonensis infection. Immunobiology 2020, 225, 151857. [Google Scholar] [CrossRef] [PubMed]
- Alhakeem, S.S.; Sindhava, V.J.; McKenna, M.K.; Gachuki, B.W.; Byrd, J.C.; Muthusamy, N.; Bondada, S. Role of B cell receptor signaling in IL-10 production by normal and malignant B-1 cells. Ann. N. Y. Acad. Sci. 2015, 1362, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holodick, N.E.; Rothstein, T.L. Atypical response of B-1 cells to BCR ligation: A speculative model. Front. Immunol. 2013, 4, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, J.; Huang, H. The development and function of thymic B cells. Cell Mol. Life Sci. 2015, 72, 265720142663. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.; Ode, Y.; Zhou, M.; Ochani, M.; Holodick, N.E.; Rothstein, T.L.; Wang, P. B-1a cells protect mice from sepsis-induced acute lung injury. Mol. Med. 2018, 24, 26. [Google Scholar] [CrossRef] [Green Version]
- Aziz, M.; Holodick, N.E.; Rothstein, T.L.; Wang, P. The role of B-1 cells in inflammation. Immunol. Res. 2015, 63, 153–166. [Google Scholar] [CrossRef]
- Aziz, M.; Holodick, N.E.; Rothstein, T.L.; Wang, P. B-1a Cells Protect Mice from Sepsis: Critical Role of CREB. J. Immunol. 2017, 199, 750–760. [Google Scholar] [CrossRef]
- Kobayashi, T.; Oishi, K.; Okamura, A.; Maeda, S.; Komuro, A.; Hamaguchi, Y.; Fujimoto, M.; Takehara, K.; Matsushita, T. Regulatory B1a Cells Suppress Melanoma Tumor Immunity via IL-10 Production and Inhibiting T Helper Type 1 Cytokine Production in Tumor-Infiltrating CD8+ T Cells. J. Investig. Dermatol. 2019, 139, 1535–1544.e1. [Google Scholar] [CrossRef]
- Grönwall, C.; Vas, J.; Silverman, G.J. Protective Roles of Natural IgM Antibodies. Front. Immunol. 2012, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Dalal, R.; Cao, C.D.; Postoak, J.L.; Yang, G.; Zhang, Q.; Wang, Z.; Lal, H.; Van Kaer, L. IL-10-producing B cells are enriched in murine pericardial adipose tissues and ameliorate the outcome of acute myocardial infarction. Proc. Natl. Acad. Sci. USA 2019, 116, 21673–21684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, L.H.; Li, K.P.; Chu, K.H.; Chiang, B.L. A B-1a cell subset induces Foxp3(-) T cells with regulatory activity through an IL-10-independent pathway. Cell Mol. Immunol. 2015, 12, 354–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Liu, Y.; Li, T.; Wang, P.; Mo, X.; Lv, P.; Ge, Q.; Ma, D.; Han, W. Cmtm7 knockout inhibits B-1a cell development at the transitional (TrB-1a) stage. Int. Immunol. 2019, 31, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Margry, B.; Kersemakers, S.C.; Hoek, A.; Arkesteijn, G.J.; Wieland, W.H.; van Eden, W.; Broere, F. Activated peritoneal cavity B-1a cells possess regulatory B cell properties. PLoS ONE 2014, 9, e88869. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Sinha, D.; Biswas, R.; Biswas, T. Pathogen-associated porin turns IL-10 competent B-1a cells toward proinflammatory cytokine response. Immunobiology 2016, 221, 1369–1373. [Google Scholar] [CrossRef]
- Tung, J.W.; Mrazek, M.D.; Yang, Y.; Herzenberg, L.A.; Herzenberg, L.A. Phenotypically distinct B cell development pathways map to the three B cell lineages in the mouse. Proc. Natl. Acad. Sci. USA 2006, 103, 6293–6298. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.; Kearney, J.F. B1 cells: Similarities and differences with other B cell subsets. Curr. Opin. Immunol. 2001, 13, 195–201. [Google Scholar] [CrossRef]
- Hsu, M.C.; Toellner, K.M.; Vinuesa, C.G.; Maclennan, I.C. B cell clones that sustain long-term plasmablast growth in T-independent extrafollicular antibody responses. Proc. Natl. Acad. Sci. USA 2006, 103, 5905–5910. [Google Scholar] [CrossRef] [Green Version]
- Obukhanych, T.V.; Nussenzweig, M.C. T-independent type II immune responses generate memory B cells. J. Exp. Med. 2006, 203, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Alugupalli, K.R. A distinct role for B1b lymphocytes in T cell-independent immunity. Curr. Top. Microbiol. Immunol. 2008, 319, 105–130. [Google Scholar] [CrossRef]
- Foote, J.B.; Kearney, J.F. Generation of B cell memory to the bacterial polysaccharide alpha-1,3 dextran. J. Immunol. 2009, 183, 6359–6368. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.M. Programmed cell death 1 suppresses B-1b cell expansion and long-lived IgG production in response to T cell-independent type 2 antigens. J. Immunol. 2011, 187, 5183–5195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, H.M.; Bender, T.P.; McNamara, C.A. B cell subsets in atherosclerosis. Front. Immunol. 2012, 3, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, A.F.; Flores-Langarica, A.; Bobat, S.; Medina, C.C.; Cook, C.N.; Ross, E.A.; Lopez-Macias, C.; Henderson, I.R. B1b cells recognize protective antigens after natural infection and vaccination. Front. Immunol. 2014, 5, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikakulapu, P.; Hu, D.; Yin, C.; Mohanta, S.K.; Bontha, S.V.; Peng, L.; Beer, M.; Weber, C.; McNamara, C.A.; Grassia, G.; et al. Artery Tertiary Lymphoid Organs Control Multilayered Territorialized Atherosclerosis B-Cell Responses in Aged ApoE-/-Mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1174–1185. [Google Scholar] [CrossRef] [Green Version]
- Mussalem, J.S.; Squaiella-Baptistão, C.C.; Teixeira, D.; Yendo, T.M.; Thies, F.G.; Popi, A.F.; Mariano, M.; Longo-Maugéri, I. Adjuvant effect of killed Propionibacterium acnes on mouse peritoneal B-1 lymphocytes and their early phagocyte differentiation. PLoS ONE 2012, 7, e33955. [Google Scholar] [CrossRef] [Green Version]
- Roco, J.A.; Mesin, L.; Binder, S.C.; Nefzger, C.; Gonzalez-Figueroa, P.; Canete, P.F.; Ellyard, J.; Shen, Q.; Robert, P.A.; Cappello, J.; et al. Class-Switch Recombination Occurs Infrequently in Germinal Centers. Immunity 2019, 51, 337–350. [Google Scholar] [CrossRef]
- Seifert, M.; Przekopowitz, M.; Taudien, S.; Lollies, A.; Ronge, V.; Drees, B.; Lindemann, M.; Hillen, U.; Engler, H.; Singer, B.B.; et al. Functional capacities of human IgM memory B cells in early inflammatory responses and secondary germinal center reactions. Proc. Natl. Acad. Sci. USA 2015, 112, E546–E555. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Sarwal, M.M. Unraveling the Role of Allo-Antibodies and Transplant Injury. Front. Immunol. 2016, 7, 432. [Google Scholar] [CrossRef] [Green Version]
- Jourdan, M.; Caraux, A.; De Vos, J.; Fiol, G.; Larroque, M.; Cognot, C.; Bret, C.; Duperray, C.; Hose, D.; Klein, B. An in vitro model of differentiation of memory B cells into plasmablasts and plasma cells including detailed phenotypic and molecular characterization. Blood 2009, 114, 5173–5181. [Google Scholar] [CrossRef] [Green Version]
- Zotos, D.; Coquet, J.M.; Zhang, Y.; Light, A.; D’Costa, K.; Kallies, A.; Corcoran, L.M.; Godfrey, D.I.; Toellner, K.M.; Smyth, M.J.; et al. IL-21 regulates germinal center B cell differentiation and proliferation through a B cell-intrinsic mechanism. J. Exp. Med. 2010, 207, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, H.; Zeng, Q.; Liang, Y.; Xiao, C.; Xie, S.; Xu, X. Correlation between TLR9 Expression and Cytokine Secretion in the Clinical Diagnosis of Systemic Lupus Erythematosus. Mediat. Inflamm. 2015, 2015, 710720. [Google Scholar] [CrossRef] [PubMed]
- Herlands, R.A.; Christensen, S.R.; Sweet, R.A.; Hershberg, U.; Shlomchik, M.J. T cell-independent and toll-like receptor-dependent antigen-driven activation of autoreactive B cells. Immunity 2008, 29, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rookhuizen, D.C.; DeFranco, A.L. Toll-like receptor 9 signaling acts on multiple elements of the germinal center to enhance antibody responses. Proc. Natl. Acad. Sci. USA 2014, 111, E3224–E3233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhamnani, R.D.; Nunes-Santos, C.J.; Bergerson, J.; Rosenzweig, S.D. Class-Switch Recombination (CSR)/Hyper-IgM (HIGM) Syndromes and Phosphoinositide 3-Kinase (PI3K) Defects. Front. Immunol. 2018, 9, 2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guthmiller, J.J.; Graham, A.C.; Zander, R.A.; Pope, R.L.; Butler, N.S. Cutting Edge: IL-10 Is Essential for the Generation of Germinal Center B Cell Responses and Anti-Plasmodium Humoral Immunity. J. Immunol. 2017, 198, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Toellner, K.M. Cognate interactions: Extrafollicular IL-4 drives germinal-center reactions, a new role for an old cytokine. Eur. J. Immunol. 2014, 44, 1917–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, I.; Kageyama, R.; Monticelli, L.; Johnston, R.J.; Ditoro, D.; Hansen, K.; Barnett, B.; Crotty, S. Germinal center T follicular helper cell IL-4 production is dependent on signaling lymphocytic activation molecule receptor (CD150). J. Immunol. 2010, 185, 190–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onuora, S. Autoimmunity: IFNγ signals control germinal centre formation. Nat. Rev. Rheumatol. 2016, 12, 312. [Google Scholar] [CrossRef]
- Domeier, P.P.; Chodisetti, S.B.; Soni, C.; Schell, S.L.; Elias, M.J.; Wong, E.B.; Cooper, T.K.; Kitamura, D.; Rahman, Z.S. IFN-γ receptor and STAT1 signaling in B cells are central to spontaneous germinal center formation and autoimmunity. J. Exp. Med. 2016, 213, 715–732. [Google Scholar] [CrossRef] [Green Version]
- Perez-Shibayama, C.; Gil-Cruz, C.; Pastelin-Palacios, R.; Cervantes-Barragan, L.; Hisaki, E.; Chai, Q.; Onder, L.; Scandella, E.; Regen, T.; Waisman, A.; et al. IFN-γ-producing CD4+ T cells promote generation of protective germinal center-derived IgM+ B cell memory against Salmonella Typhi. J. Immunol. 2014, 192, 5192–5200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takatsuka, S.; Yamada, H.; Haniuda, K.; Saruwatari, H.; Ichihashi, M.; Renauld, J.C.; Kitamura, D. IL-9 receptor signaling in memory B cells regulates humoral recall responses. Nat. Immunol. 2018, 19, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Cresswell, P. Assembly, transport, and function of MHC class II molecules. Ann. Rev. Immunol. 1994, 12, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Ni, Z.; Liu, X.; Feng, S.; Dong, X.; Shi, X.; Zhai, J.; Mai, S.; Jiang, J.; Wang, Z.; et al. Beyond T cells: Understanding the role of PD-1/PD-L1 in Tumor-Associated Macrophages. J. Immunol. Res. 2019, 2019, 1919082. [Google Scholar] [CrossRef] [Green Version]
- Podojil, J.R.; Miller, S.D. Targeting the B7 family of co-stimulatory molecules: Successes and challenges. BioDrugs 2013, 27, 1–13. [Google Scholar] [CrossRef]
- Yao, S.; Zhu, Y.; Zhu, G.; Augustine, M.; Zheng, L.; Goode, D.J.; Broadwater, M.; Ruff, W.; Flies, S.; Xu, H.; et al. B7-h2 is a costimulatory ligand for CD28 in human. Immunity 2011, 34, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Pentcheva-Hoang, T.; Egen, J.G.; Wojnoonski, K.; Allison, J.P. B7-1 and B7-2 selectively recruit CTLA-4 and CD28 to the immunological synapse. Immunity 2004, 21, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Nagamatsu, T.; Barrier, B.F.; Schust, D.J. The regulation of T-cell cytokine production by ICOS-B7H2 interactions at the human fetomaternal interface. Immunol. Cell Biol. 2011, 89, 417–425. [Google Scholar] [CrossRef]
- Lobo, P.I. Role of Natural Autoantibodies and Natural IgM Anti-Leucocyte Autoantibodies in Health and Disease. Front. Immunol. 2016, 7, 198. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notley, C.A.; Brown, M.A.; Wright, G.P.; Ehrenstein, M.R. Natural IgM Is Required for Suppression of Inflammatory Arthritis by Apoptotic Cells. J. Immunol. 2011, 186, 4967–4972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, K.; Jin, J.; Atkinson, C.; Alawieh, A.; Qiao, F.; Lei, B.; Chavin, K.D.; He, S.; Tomlinson, S. Natural immunoglobulin M initiates an inflammatory response important for both hepatic ischemia reperfusion injury and regeneration in mice. Hepatology 2018, 67, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Morbach, H.O.; Eichhorn, E.M.; Liese, J.G.; Girschick, H.J. Reference values for B cell subpopulations from infancy to adulthood. Clin. Exp. Immunol. 2010, 162, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Imamura, R.; Takahara, S. Evaluation of Antigen-Specific IgM and IgG Production during an In Vitro Peripheral Blood Mononuclear Cell Culture Assay. Front. Immunol. 2017, 8, 794. [Google Scholar] [CrossRef] [Green Version]
- Lúcia, M.; Luque, S.; Crespo, E.; Melilli, E.; Cruzado, J.M.; Martorell, J.; Jarque, M.; Gil-Vernet, S.; Manonelles, A.; Grinyó, J.M.; et al. Preformed circulating HLA-specific memory B cells predict high risk of humoral rejection in kidney transplantation. Kidney Int. 2015, 88, 874–887. [Google Scholar] [CrossRef] [Green Version]
- Browne, E.P. Toll-like receptor 7 controls the anti-retroviral germinal center response. PLoS Pathog. 2011, 7, e1002293. [Google Scholar] [CrossRef] [Green Version]
- Desnues, B.; Macedo, A.B.; Roussel-Queval, A.; Bonnardel, J.; Henri, S.; Demaria, O.; Alexopoulou, L. TLR8 on dendritic cells and TLR9 on B cells restrain TLR7-mediated spontaneous autoimmunity in C57BL/6 mice. Proc. Natl. Acad. Sci. USA 2014, 111, 1497–1502. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Rogers, J.A.; Lavingia, B.; Stastny, P. Peripheral blood B cells producing donor-specific HLA antibodies in vitro. Hum. Immunol. 2009, 70, 29–34. [Google Scholar] [CrossRef]
- Imamura, R.; Matsuda, Y.; Tsutahara, K.; Nonomura, N.; Takahara, S. Impact of Immunoglobulin M-Type Donor-Specific Human Leukocyte Antigen-Antibody Levels in Supernatants from Cultured Peripheral Blood Mononuclear Cells as Predictors of Antibody-Mediated Rejection. Pathogens 2020, 9, 733. [Google Scholar] [CrossRef]
- Sörman, A.; Zhang, L.; Ding, Z.; Heyman, B. How antibodies use complement to regulate antibody responses. Mol. Immunol. 2014, 61, 79–88. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Kläsener, K.; Zürn, C.; Castillo, P.A.; Brust-Mascher, I.; Imai, D.M.; Bevins, C.L.; Reardon, C.; Reth, M.; Baumgarth, N. The IgM receptor FcμR limits tonic BCR signaling by regulating expression of the IgM BCR. Nat. Immunol. 2017, 18, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Elsner, R.A.; Baumgarth, N. Natural IgM Prevents Autoimmunity by Enforcing B Cell Central Tolerance Induction. J. Immunol. 2015, 194, 1489–1502. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Xiong, E.; Hong, R.; Lu, Q.; Ohno, H.; Wang, J.Y. Role of the IgM Fc Receptor in Immunity and Tolerance. Front. Immunol. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- Landry, M.L. Immunoglobulin M for Acute infection: True or False? Clin. Vaccine Immunol. 2016, 23, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yi, Y.; Luo, X.; Xiong, N.; Liu, Y.; Li, S.; Sun, R.; Wang, Y.; Hu, B.; Chen, W.; et al. Development and clinical application of a rapid IgM-IgG combined antibody test for SARS-CoV-2 infection diagnosis. J. Med. Virol. 2020. Online ahead of print. [Google Scholar] [CrossRef]
- Rubens Costa Lima, J.; Rouquayrol, M.Z.; Monteiro Callado, M.R.; Florindo Guedes, M.I.; Pessoa, C. Interpretation of the presence of IgM and IgG antibodies in a rapid test for dengue: Analysis of dengue antibody prevalence in Fortaleza City in the 20th year of the epidemic. Rev. Soc. Bras. Med. Trop 2012, 45, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Keiserman, B.; Ronchetti, M.R.; Monticielo, O.A.; Keiserman, M.W.; Staub, H.L. Concomitance of IgM and IgG anti-dsDNA Antibodies Does Not Appear to Associate to Active Lupus Nephritis. Open Rheumatol. J. 2013, 7, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Saraux, A.; Bendaoud, B.; Dueymes, M.; Le Goff, P.; Youinou, P. The functional affinity of IgM rheumatoid factor is related to the disease duration in patients with rheumatoid arthritis. Ann. Rheum Dis. 1997, 56, 126–129. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Zaragoza, M.; Hernández-Ávila, R.; Viedma-Rodríguez, R.; Arenas-Aranda, D.; Ostoa-Saloma, P. Natural and adaptive IgM antibodies in the recognition of tumor-associated antigens of breast cancer (Review). Oncol. Rep. 2015, 34, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Galeotti, C.; Kaveri, S.V.; Bayry, J. IVIG-mediated effector functions in autoimmune and inflammatorydiseases. Int. Immunol. 2017, 29, 491–498. [Google Scholar] [CrossRef]
- Vassilev, T.; Mihaylova, N.; Voynova, E.; Nikolova, M.; Kazatchkine, M.; Kaveri, S. IgM-enriched human intravenous immunoglobulin suppresses T lymphocyte functions in vitro and delays the activation of T lymphocytes in hu-SCID mice. Clin. Exp. Immunol. 2006, 145, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Rieben, R.; Roos, A.; Muizert, Y.; Tinguely, C.; Gerritsen, A.F.; Daha, M.R. Immunoglobulin M-enriched human intravenous immunoglobulin prevents complement activation in vitro and in vivo in a rat model of acute inflammation. Blood 1999, 93, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Ates, I.; Dogan, N.; Aksoy, M.; Halıcı, Z.; Gündogdu, C.; Keles, M.S. The protective effects of IgM-enriched immunoglobulin and erythropoietin on the lung and small intestine tissues of rats with induced sepsis: Biochemical and histopathological evaluation. Pharm. Biol. 2015, 53, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wei, X.; Lv, H.; Li, Y.; Li, P.; Chen, Z.; Liu, G. The clinical efficacy of intravenous IgM-enriched immunoglobulin(pentaglobin) in sepsis or septic shock: A meta-analysis with trialsequential analysis. Ann. Intens. Care 2019, 9, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norrby-Teglund, A.; Haque, K.N.; Hammarström, L. Intravenous polyclonal IgM-enriched immunoglobulin therapy in sepsis: A review of clinical efficacy in relation to microbiological aetiology and severity of sepsis. J. Intern. Med. 2006, 260, 509–516. [Google Scholar] [CrossRef]
- Radbruch, A.; Muehlinghaus, G.; Luger, E.O.; Inamine, A.; Smith, K.G.; Dörner, T.; Hiepe, F. Competence and competition: The challenge of becoming a long-lived plasma cell. Nat. Rev. Immunol. 2006, 6, 41–50. [Google Scholar] [CrossRef]
- Khaled, Y.; Solh, M.M.; Ward, D.; Chang, C.C. Plasma Cell CD20 Expression After Autologous Stem Cell Transplantation in Multiple Myeloma: Primary Aberrant Expression or Receptor up-Regulation. Blood 2012, 120, 4538. [Google Scholar] [CrossRef]
- Toyoda, M.; Shin, B.H.; Ge, S.; Mirocha, J.; Thomas, D.; Chu, M.; Rodriguez, E.; Chao, C.; Petrosyan, A.; Galera, O.A.; et al. Impact of Desensitization on Antiviral Immunity in HLA-Sensitized Kidney Transplant Recipients. J. Immunol. Res. 2017, 2017, 5672523. [Google Scholar] [CrossRef]
- Morath, C.; Opelz, G.; Zeier, M.; Süsal, C.J. Clinical relevance of HLA antibody monitoring after kidney transplantation. Immunol. Res. 2014, 2014, 845040. [Google Scholar] [CrossRef] [Green Version]
- Stastny, P.; Ring, S.; Lu, C.; Arenas, J.; Han, M. Role of immunoglobulin (Ig)-G and IgM antibodies against donor human leukocyte antigens in organ transplant recipients. Hum. Immunol. 2009, 70, 600–604. [Google Scholar] [CrossRef]
- Paantjens, A.W.; van de Graaf, E.A.; Kwakkel-van Erp, J.M.; Hoefnagel, T.; van Ginkel, W.G.; Fakhry, F.; van Kessel, D.A.; van den Bosch, J.M.; Otten, H.G. The Induction of IgM and IgG Antibodies against HLA or MICA after Lung Transplantation. Pulm Med. 2011, 2011, 432169. [Google Scholar] [CrossRef] [PubMed]
- Bentall, A.; Tyan, D.B.; Sequeira, F.; Everly, M.J.; Gandhi, M.J.; Cornell, L.D.; Li, H.; Henderson, N.A.; Raghavaiah, S.; Winters, J.L.; et al. Antibody-mediated rejection despite Inhibition of terminal complement. Transpl. Int. 2014, 27, 1235–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAlister, C.C.; Gao, Z.H.; McAlister, V.C.; Gupta, R.; Wright, J.R., Jr.; MacDonald, A.S.; Peltekian, K. Protective anti-donor IgM production after crossmatch positive liver–kidney transplantation. Liver Transpl. 2004, 10, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Anani, W.Q.; Pugh, T.; Gottschall, J.L.; Denomme, G.A. Complement activating ABO anti-A IgM/IgG act synergistically to cause erythrophagocytosis: Implications among minor ABO incompatible transfusions. J. Transl. Med. 2020, 18, 216. [Google Scholar] [CrossRef]
- Wang, X.D.; Liu, J.P.; Fan, Y.; Song, T.R.; Shi, Y.Y.; Li, Y.M.; Lv, Y.H.; Li, X.H.; Huang, Z.L.; Lin, T. Individualized Preconditioning for ABO-Incompatible Living-Donor Kidney Transplantation: An Initial Report of 48 Cases from China. Ann. Transpl. 2020, 25, e920224. [Google Scholar] [CrossRef]
- Koo, T.Y.; Yang, J. Current progress in ABO-incompatible kidney transplantation. Kidney. Res. Clin. Pract. 2015. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Choe, W.; Shin, S.; Kim, Y.H.; Han, D.J.; Park, S.K.; Kwon, S.W.; Ko, D.H. ABO-incompatible kidney transplantation can be successfully conducted by monitoring IgM isoagglutinin titers during desensitization. Transfusion 2020, 598–606. [Google Scholar] [CrossRef]
- Toki, D.; Ishida, H.; Setoguchi, K.; Shimizu, T.; Omoto, K.; Shirakawa, H.; Iida, S.; Horita, S.; Furusawa, M.; Ishizuka, T.; et al. Acute antibody-mediated rejection in living ABO-incompatible kidney transplantation: Long-term impact and risk factors. Am. J. Transpl. 2009, 9, 567–577. [Google Scholar] [CrossRef]
- Shin, M.; Kim, S.J. ABO Incompatible Kidney Transplantation-Current Status and Uncertainties. J. Transpl. 2011, 2011, 970421. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K. Recent findings in ABO-incompatible kidney transplantation: Classification and therapeutic strategy for acute antibody-mediated rejection due to ABO-blood-group-related antigens during the critical period preceding the establishment of accommodation. Clin. Exp. Nephrol. 2007, 11, 128–141. [Google Scholar] [CrossRef]
- Kirk, A.D.; Baldwin, W.M.; Cascalho, M.I.; Chong, A.S.; Sykes, M.; West, L.J. American society of transplantation symposium on B cells in transplantation: Harnessing humoral immunity from rodent models to clinical practice. Am. J. Transpl. 2007, 7, 1464–1670. [Google Scholar] [CrossRef] [PubMed]
- West, L.J. Targeting antibody-mediated rejection in the setting of ABO-incompatible infant heart transplantation: Graft accommodation vs. B cell tolerance. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2005, 5, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.N.; Kawai, T.; Boskovic, S.; Nadazdin, O.; Sachs, D.H.; Cosimi, A.B.; Colvin, R.B. Four stages and lack of stable accommodation in chronic alloantibody-mediated renal allograft rejection in Cynomolgus monkeys. Am. J. Transpl. 2008, 8, 1662–1672. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.T.; Platt, J.L. The role of antibodies in transplantation. Transpl. Rev. 2009, 23, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Avery, R.K.; Motter, J.D.; Jackson, K.R.; Montgomery, R.A.; Massie, A.B.; Kraus, E.S.; Marr, K.A.; Lonze, B.E.; Alachkar, N.; Holechek, M.J.; et al. Quantifying infection risks in incompatible living donor kidney transplant recipients. Am. J. Transpl. 2020. [Google Scholar] [CrossRef]
- Sharif, A.; Alachkar, N.; Bagnasco, S.; Geetha, D.; Gupta, G.; Womer, K.; Arend, L.; Racusen, L.; Montgomery, R.; Kraus, E. Incidence and outcomes of BK virus allograft nephropathy among ABO- and HLA-incompatible kidney transplant recipients. Clin. J. Am. Soc. Nephrol. 2012, 7, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Malheiro, J.; Tafulo, S.; Dias, L.; Martins, S.; Fonseca, I.; Beirão, I.; Castro-Henriques, A.; Cabrita, A. Determining donor-specific antibody C1q-binding ability improves the prediction of antibody-mediated rejection in human leucocyte antigen-incompatible kidney transplantation. Transpl. Int. 2017, 30, 347–359. [Google Scholar] [CrossRef]
- Cozzi, E.; Biancone, L. C1q-binding donor-specific antibody assays help define risk and prognosis in antibody-mediated rejection. Kidney Int. 2018, 94, 657–659. [Google Scholar] [CrossRef]
- Thammanichanond, D.; Mongkolsuk, T.; Rattanasiri, S.; Kantachuvesiri, S.; Worawichawong, S.; Jirasiritham, S.; Kitpoka, P. Significance of C1q-fixing donor-specific antibodies after kidney transplantation. Transpl. Proc. 2014, 46, 368–371. [Google Scholar] [CrossRef]
- Tan, E.K.; Bentall, A.; Dean, P.G.; Shaheen, M.F.; Stegall, M.D.; Schinstock, C.A. Use of Eculizumab for Active Antibody-mediated Rejection That Occurs Early Post-kidney Transplantation: A Consecutive Series of 15 Cases. Transplantation 2019, 103, 2397–2404. [Google Scholar] [CrossRef]
- Marks, W.H.; Mamode, N.; Montgomery, R.A.; Stegall, M.D.; Ratner, L.E.; Cornell, L.D.; Rowshani, A.T.; Colvin, R.B.; Dain, B.; Boice, J.A.; et al. C10-001 Study Group. Safety and efficacy of eculizumab in the prevention of antibody-mediated rejection in living-donor kidney transplant recipients requiring desensitization therapy: A randomized trial. Am. J. Transpl. 2019, 19, 2876–2888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stites, E.; Le Quintrec, M.; Thurman, J.M. The Complement System and Antibody-Mediated TransplantRejection. J. Immunol. 2015, 195, 5525–5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ius, F.; Sommer, W.; Kieneke, D.; Tudorache, I.; Kühn, C.; Avsar, M.; Siemeni, T.; Salman, J.; Erdfelder, C.; Verboom, M.; et al. IgM-Enriched Human Intravenous Immunoglobulin-Based Treatment of Patients With Early Donor Specific Anti-HLA Antibodies After Lung Transplantation. Transplantation 2016, 100, 2682–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ius, F.; Verboom, M.; Sommer, W.; Müller, C.; Hallensleben, M.; Salman, J.; Siemeni, T.; Kühn, C.; Avsar, M.; Bobylev, D.; et al. Five-Year Results of an IgA and IgM-Enriched Human Immunoglobulin G-Based Therapy for Early Anti-HLA Donor Specific Antibodies in 158 Lung-Transplanted Patients. J. Heart Lung Transpl. 2019, 41, S166. [Google Scholar] [CrossRef]
- Liu, S.; Li, S.; Zhang, Y.; Wang, Y.; Zhu, Y.; Wang, B.; Chen, Z.N. Purification of a polyclonal antibody against CD147 for ELISA using antigen-immunoaffinity chromatography. Mol. Med. Rep. 2017, 15, 4035–4040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, S.; Kong, F. Separation of antigens and antibodies by immunoaffinity chromatography. Pharm. Biol. 2012, 50, 1038–1044. [Google Scholar] [CrossRef] [Green Version]
- Lowe, D.; Higgins, R.; Zehnder, D.; Briggs, D.C. Significant IgG subclass heterogeneity in HLA-specific antibodies: Implications for pathogenicity, prognosis, and the rejection response. Hum. Immunol. 2013, 74, 666–672. [Google Scholar] [CrossRef]
- Taniguchi, M.; Rebellato, L.M.; Briley, K.P.; Haisch, C.E.; Bolin, P.; Banuelos, N.; Hopfield, J.; Terasaki, P.I.; Everly, M.J. Risk Stratification of Human Leukocyte Antigen Class II Donor Specific Antibody Positive Patients by Immunoglobulin G Subclasses. Clin. Transpl. 2015, 31, 293–301. [Google Scholar]
- Correa, I.; Ilieva, K.M.; Crescioli, S.; Lombardi, S.; Figini, M.; Cheung, A.; Spicer, J.F.; Tutt, A.N.J.; Nestle, F.O.; Karagiannis, P.; et al. Evaluation of Antigen-Conjugated Fluorescent Beads to Identify Antigen-Specific B Cells. Front. Immunol. 2018, 9, 493. [Google Scholar] [CrossRef] [Green Version]
- Rydyznski, C.; Daniels, K.A.; Karmele, E.P.; Brooks, T.R.; Mahl, S.E.; Moran, M.T.; Li, C.; Sutiwisesak, R.; Welsh, R.M.; Waggoner, S.N. Generation of cellular immune memory and B-cell immunity is impaired by natural killer cells. Nat. Commun. 2015, 6, 6375. [Google Scholar] [CrossRef] [Green Version]
- Sala, E.; Kuka, M. The Suppressive Attitude of Inflammatory Monocytes in Antiviral Antibody Responses. Viral Immunol. 2020, 33, 327–333. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuda, Y.; Hiramitsu, T.; Li, X.-k.; Watanabe, T. Characteristics of Immunoglobulin M Type Antibodies of Different Origins from the Immunologic and Clinical Viewpoints and Their Application in Controlling Antibody-Mediated Allograft Rejection. Pathogens 2021, 10, 4. https://doi.org/10.3390/pathogens10010004
Matsuda Y, Hiramitsu T, Li X-k, Watanabe T. Characteristics of Immunoglobulin M Type Antibodies of Different Origins from the Immunologic and Clinical Viewpoints and Their Application in Controlling Antibody-Mediated Allograft Rejection. Pathogens. 2021; 10(1):4. https://doi.org/10.3390/pathogens10010004
Chicago/Turabian StyleMatsuda, Yoshiko, Takahisa Hiramitsu, Xiao-kang Li, and Takeshi Watanabe. 2021. "Characteristics of Immunoglobulin M Type Antibodies of Different Origins from the Immunologic and Clinical Viewpoints and Their Application in Controlling Antibody-Mediated Allograft Rejection" Pathogens 10, no. 1: 4. https://doi.org/10.3390/pathogens10010004