
Salmonella Infection in Pigs: Disease, Prevalence, and a Link between Swine and Human Health


Istituto Zooprofilattico Sperimentale della Lombardia e dell’Emilia-Romagna (IZSLER), 25124 Brescia, Italy
*
Author to whom correspondence should be addressed.
Pathogens 2023, 12(10), 1267; https://doi.org/10.3390/pathogens12101267
Submission received: 15 September 2023
/
Revised: 12 October 2023
/
Accepted: 20 October 2023
/
Published: 21 October 2023
(This article belongs to the Special Issue Swine Bacterial Pathogens from a One Health Perspective)
Abstract
:Salmonella is one of the most spread foodborne pathogens worldwide, and Salmonella infections in humans still represent a global health burden. The main source of Salmonella infections in humans is represented by contaminated animal-derived foodstuffs, with pork products being one of the most important players. Salmonella infection in swine is critical not only because it is one of the main causes of economic losses in the pork industry, but also because pigs can be infected by several Salmonella serovars, potentially contaminating the pig meat production chain and thus posing a significant threat to public health globally. As of now, in Europe and in the United States, swine-related Salmonella serovars, e.g., Salmonella Typhimurium and its monophasic variant Salmonella enterica subsp. enterica 1,4,[5],12:i:-, are also frequently associated with human salmonellosis cases. Moreover, multiple outbreaks have been reported in the last few decades which were triggered by the consumption of Salmonella-contaminated pig meat. Throughout the years, changes and evolution across the pork industry may have acted as triggers for new issues and obstacles hindering Salmonella control along the food chain. Gathered evidence reinforces the importance of coordinating control measures and harmonizing monitoring programs for the efficient control of Salmonella in swine. This is necessary in order to manage outbreaks of clinical disease in pigs and also to protect pork consumers by controlling Salmonella subclinical carriage and shedding. This review provides an update on Salmonella infection in pigs, with insights on Salmonella ecology, focusing mainly on Salmonella Choleraesuis, S. Typhimurium, and S. 1,4,[5],12:i:-, and their correlation to human salmonellosis cases. An update on surveillance methods for epidemiological purposes of Salmonella infection in pigs and humans, in a “One Health” approach, will also be reported.
1. Introduction
Salmonella is one of the main intestinal pathogens in swine [1], and the infection in pigs is a cause for concern for two main reasons. The first is the threat that Salmonella-contaminated pork products pose to human health. The second is about animal health, with regard to clinical disease in pigs; mainly, septicemic salmonellosis associated with Salmonella Choleraesuis and enterocolitis associated with S. Typhimurium and its monophasic variant (S. 1,4,[5],12:i:-), which cause significant economic losses due to increased mortality, growth retardation [2,3,4], and cost of treatment [5].
Non-typhoidal Salmonella (NTS), together with Campylobacter spp., is one of the leading causes of foodborne disease at a global level [6], and the primary reservoir of NTS is the intestinal tract of warm-blooded animals, specifically livestock animals destined for food production, with the potential to lead to contaminated food products [7,8]. Eating contaminated food, especially foodstuffs of animal origin, is believed to be the major transmission source of Salmonella infections to humans, with a high burden on health systems internationally [7,9]. Human salmonellosis is mainly attributed to contaminated food consumption, such as poultry, eggs, pork, and beef, as well as fresh products [10]. Pork products are one of the main animal-derived foodstuffs involved in Salmonella transmission to humans [11,12]. It has been estimated that, every year, around 80.3 million human Salmonellosis cases have a foodborne origin [9]. In the European Union (EU), Salmonella is the second-most common source of foodborne infections in humans after Campylobacter since 2005 [13,14]. In the United States (US), according to the Centers for Disease Control and Prevention (CDC), illnesses caused by Salmonella are estimated to be around 1.35 million, with 26,500 hospitalizations and 420 deaths each year, with contaminated food acting as the main source of infections [15]. It is also estimated that foodborne salmonellosis costs the US $2.7 billion per year, with pork producers losing approximately $100 million [16]. According to recent data, as of January 2020, there were approximately 677.6 million pigs worldwide [17]. The pig production system has undergone great changes in the past century, transitioning from small herds to large holding facilities housing a great number of animals [5,18]. The exponential growth of intensive farming and food production practices could have led to new issues regarding the management and control of swine salmonellosis [11,19]. Moreover, available data on the incidence of Salmonella infection in pigs are scarce and often represent only the tip of the iceberg, mainly because pigs infected with Salmonella are often subclinical carriers [20]. Given that the pig production chain and, consequently, pig meat consumption have a direct impact on human health, a more collaborative approach between human and veterinary medicine (“One Health”) is crucial for preventing and minimizing potential health threats along the food chain, in order to preserve animal, human, and environmental safety [21].
The purpose of this review is to update the information on clinically relevant Salmonella serovars (S. Choleraesuis, S. Typhimurium, and its monophasic variant) responsible for infection and disease in pigs. Salmonella Derby will also be discussed, given its strong association with the pig production chain, even if it is not considered a cause of enteric disease in pigs. Moreover, the impact of these serovars on human health will also be evaluated from a “One Health” point of view.
2. Etiology
The genus Salmonella is divided into two species: S. enterica and S. bongori. Moreover, the species enterica is divided into six different subspecies: enterica (I), salamae (II), arizonae (IIIa), diarizonae (IIIb), houtenae (IV), and indica (VI). There are more than 2600 different Salmonella serovars, each characterized by a distinct antigenic formula [22,23]. While S. bongori is mainly isolated from cold-blooded animals and from environmental sources, S. enterica subspecies enterica’s main reservoir are warm-blooded animals, which harbor the pathogen in their intestinal tract [24,25]. Moreover, bacteria belonging to the genus Salmonella are well known for their capability to infect an ample range of hosts, often exhibiting different behaviors regarding their host range [26]. The majority of NTS, namely any serovar different from Salmonella Typhi or Salmonella Paratyphi A, B, and C [27], are characterized by having a broad host range, and are otherwise called “generalist” serovars, e.g., S. Typhimurium, which can infect and cause disease in several animal species, including humans, pigs, cattle, and poultry [7]. Nonetheless, there are serovars which are adapted or restricted to a single host species; for example, Salmonella Dublin is adapted to cattle [28], Salmonella Gallinarum is restricted to fowl [29], and S. Choleraesuis is adapted to pigs [30].
3. Salmonellosis in Swine
In swine, clinical salmonellosis has been mostly associated with two serovars: S. Choleraesuis, especially the Kunzendorf variant, and S. Typhimurium [30]. Clinical disease is usually characterized by systemic disease with septicemia (often associated with pneumonia), mainly caused by the S. Choleraesuis serovar; and enteric disease, characterized by diarrhea, which is typically due to S. Typhimurium and S. 1,4,[5],12:i:- [30,31]. Even though not associated with enteric disease in pigs, S. Derby will also be discussed, given its relevance to the pig production chain.
3.1. S. Choleraesuis
3.1.1. Overview
The first report of S. Choleraesuis dates back to 1886 in the US, when it was isolated by Salmon and his assistant Smith, who believed it to be the causative agent of “hog cholera” (swine fever) [32]. From a serological point of view, S. Choleraesuis shares its antigenic formula (6,7:c:1,5) with the S. Paratyphi C and Salmonella Typhisuis serovars. In addition, three S. Choleraesuis biotypes have been identified: S. Choleraesuis sensu stricto, S. Choleraesuis variant Kunzendorf, and S. Choleraesuis variant Decatur; these variants share the same antigenic formula but have different biochemical characteristics (dulcitol and mucate fermentation, H2S production) [22].
S. Choleraesuis is considered a swine-adapted serovar, but it is not host-restricted, given that it is also capable of causing invasive disease in humans [33,34,35,36,37], and while it is rarely reported in Europe, it is still frequent in Asia [13,35,37,38].
Historically, most S. Choleraesuis outbreaks in swine, characterized by septicemic forms, have been caused by the H2S-producing Kunzendorf variant [30,31,39].
In the US, S. Choleraesuis was the predominant serovar isolated from swine in the 1950s and the 1960s [31]. This predominance continued up until the mid-1990s. After 1995, there was a shift in the main serovars isolated from swine in the US, with a decrease in S. Choleraesuis prevalence and an increase in reported isolations of S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby, which are now the predominant serovars in swine [10].
In the EU, as of now, S. Choleraesuis is not widespread in pigs anymore [40], as it is isolated at low frequency in some European countries (e.g., Estonia and Romania) [13,38]. Data obtained by the 2006–2007 EFSA baseline survey, which aimed to evaluate Salmonella prevalence in slaughter pigs, highlighted that S. Choleraesuis was reported in only 4 out of 25 Member States (MSs, namely any nation that is a member of the European Union), with only 10 positive samples out of 2600 serotyped ileocecal lymph nodes [41]. Later on, S. Choleraesuis variant Kunzendorf accounted for 2.5% of all serotyped Salmonella isolates from pigs in the EU in 2013 [14]. According to the latest EFSA-ECDC report on zoonoses, out of all serotyped Salmonella isolates in pigs (from both animal and food sources), S. Choleraesuis sensu stricto and variant Kunzendorf were reported only six times each, with a very low prevalence of 0.38% [13]. Nowadays, in the EU, there are still reports about the isolation of S. Choleraesuis from pigs in countries such as Italy [42] and Serbia [43]. With regard to Northern Europe, after an absence of 13 years, S. Choleraesuis var. Kunzendorf was detected for the first time in swine herds in Denmark between 2012 and 2013, with reported outbreaks characterized by high mortality rates (20–30%) [39]. Moreover, after 40 years of absence, S. Choleraesuis was reported in 5 pig herds in Sweden in 2022 [44].
Furthermore, it is worth mentioning that wild boars are considered a natural wildlife reservoir for S. Choleraesuis, given that this serovar has been isolated multiple times from this species, with the potential for generating spillover events between wildlife and farmed animals [32,44,45]. In the EU, there has been a growing trend of S. Choleraesuis isolation from wild boars, whose number has been increasing in the past few decades [46]. Various outbreaks caused by S. Choleraesuis var. Kunzendorf have been reported in the wild boar population [47,48], as well as its isolation from sickened and killed wild boars, characterized by septicemic salmonellosis [49,50]. In Sweden, it was reported that, out of a total of 633 wild boars sampled within surveillance programs, 80 animals tested positive for S. Choleraesuis var. Kunzendorf, and all sequenced isolates clustered with those isolated from domestic pig farms in the same region [44]. This highlights the importance of implementing strict biosecurity measures to prevent and hinder the possibility of transmission events from the wild boar population to domestic pigs [50].
3.1.2. Clinical Signs and Gross Lesions
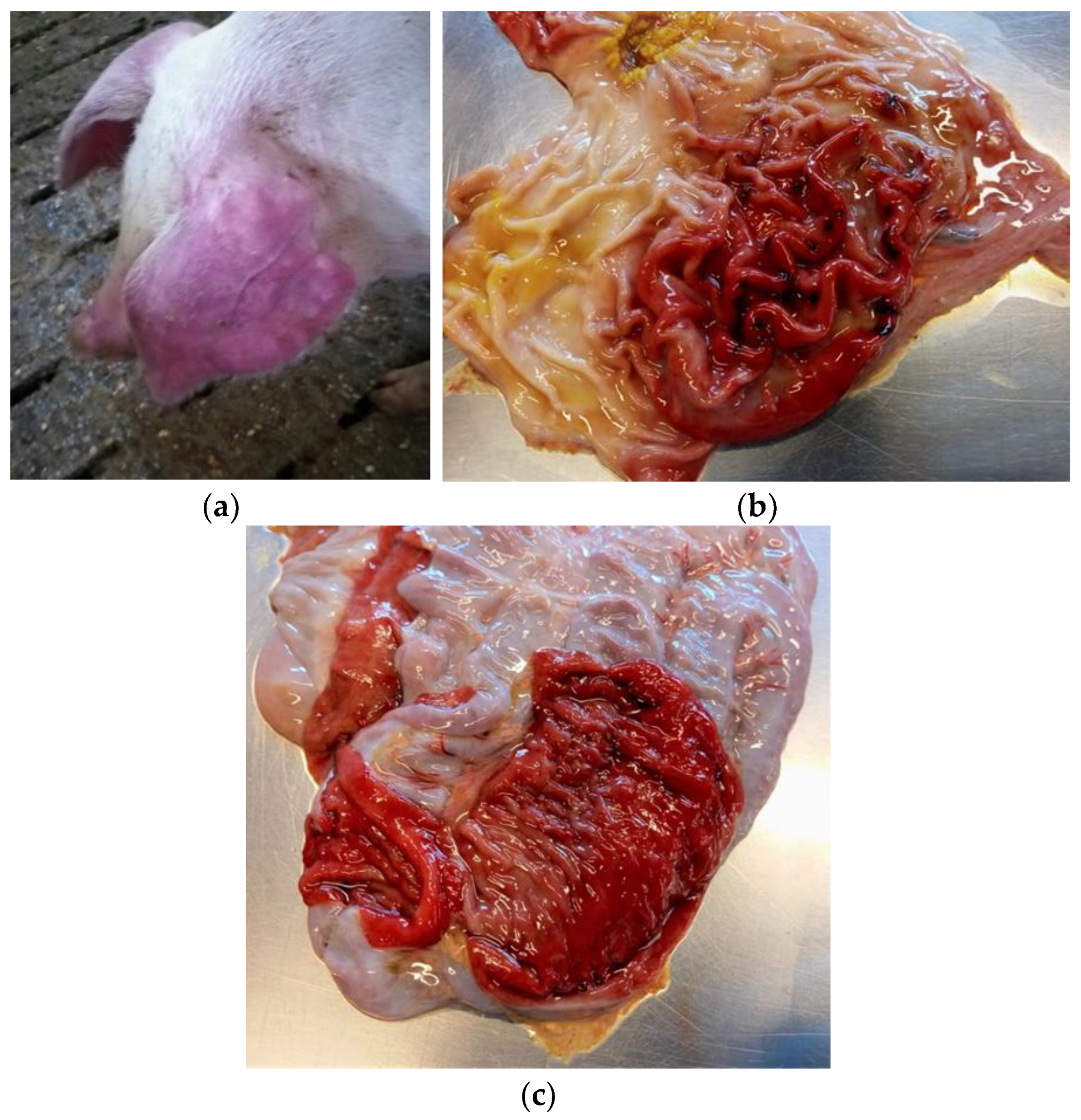
In pigs infected with S. Choleraesuis, typical signs of acute disease are cyanosis and dyspnea, with red-blue discoloration affecting mainly the ears, the chest, and the abdominal area, but also feet, belly, and ventral neck, sometimes associated with enterocolitis [31,43,51,52] (Figure 1a).
Among other common clinical manifestations, there are pyrexia, inappetence, drowsiness, moist cough, and labored breathing, while diarrhea is less pronounced or absent. Usually, clinical signs tend to arise 24–36 hours after infection, while gastrointestinal signs typically appear after 4–5 days [31]. During most outbreaks, morbidity tends to be variable, while mortality is generally high [31].
Typical lesions induced by S. Choleraesuis infection in swine are splenomegaly, gastritis, gastric mucosa infarction and gastric erosions (Figure 1b), swollen and enlarged mesenteric and gastro-hepatic lymph nodes, lung congestion and pneumonia, random white foci of necrosis in the liver, and enterocolitis (Figure 1c) [30,31].
The main microscopic lesions are characterized by scattered foci of coagulative necrosis with neutrophils and histiocytes infiltrates in the liver. Similar lesions can be detected in the spleen and in lymph nodes. Fibrinoid thrombi can be observed in the venules of cyanotic skin, gastric mucosa, kidneys (affecting glomerular capillaries), and pulmonary vessels [30]. The lungs can show interstitial pneumonia or suppurative bronchopneumonia. Segmental necrotizing vasculitis with perivascular histiocytic infiltrates, sometimes with localized necrotizing encephalitis, is uncommonly observed [30].
3.2. S. Typhimurium and Its Monophasic Variant S. 1,4,[5],12:i:-
3.2.1. Overview
Even though a broad range of Salmonella serovars can infect pigs, only a restricted number of serovars are considered a primary source of disease, with S. Typhimurium and S. 1,4,[5],12:i:- being among the most important ones [53]. S. 1,4,[5],12:i:- is a relatively recent serovar, having been isolated from poultry in the late 1980s in Portugal [54]. S. 1,4,[5],12:i:- is characterized by the lack of expression of the phase 2 flagellar antigen (H2), encoded by locus fljB [55]. This could be due to different mutations (point mutations included) or to complete or partial deletion in the fljB locus or in neighboring genes [56]. According to the White–Kaufmann–Le Minor scheme, when it was first isolated, the strain could either be a monophasic variant, S. Typhimurium, S. Lagos, or an uncharacterized serotype [22,57]. Further investigations, which involved methods such as pulsed-field gel electrophoresis (PFGE), phage typing, plasmid profiling, and multilocus VNTR analysis (MLVA), allowed researchers to determine that there was indeed a close genetic relatedness between S. 1,4,[5],12:i:- and S. Typhimurium [57,58,59]. Therefore, S. 1,4,[5],12:i:- is now widely accepted as a S. Typhimurium variant [60]. After its discovery, S. 1,4,[5],12:i:- has become one of the most widespread Salmonella serovars worldwide and one of the most reported serovars from human salmonellosis cases [56,60,61]. Probably, S. 1,4,[5],12:i:- possesses competitive and evolutionary advantages compared to other biphasic strains, and this has contributed to its increasing predominance worldwide; nevertheless, further studies are needed to investigate the reasons that allow the successful spread of monophasic strains [53,62,63,64].
An Italian study reported data about the isolation of S. Typhimurium and S. 1,4,[5],12:i:- in 1359 pig farms in Northern Italy, where clinical enteric forms or mortality occurred. S. Typhimurium and S. 1,4,[5],12:i:- represented 12.23% and 6.18% of the isolated serovars, respectively. In this study, the association between the isolated serovars and the presence of clinical signs showed a stronger correlation with S. Typhimurium compared to S. 1,4,[5],12:i:- [65].
It was suggested that, probably, the different antigenicity and pathogenicity of S. 1,4,[5],12:i:- make the infection harder to recognize and control [65]. Furthermore, when compared to S. Typhimurium, a number of factors (including prophage involvement and antigenic changes) can cause a reduced immune response in pigs to S. 1,4,[5],12:i:- [62].
In an experimental challenge study, three groups of piglets were inoculated with S. Typhimurium, S. Derby, and S. 1,4,[5],12:i:-. Despite the fact that all groups displayed diarrhea, no fever was detected in the group challenged with S. 1,4,[5],12:i:-, and only piglets challenged with S. 1,4,[5],12:i:- shed Salmonella continuously throughout the trial, with higher excretion levels than the group challenged with S. Typhimurium [66]. These data allow for hypothesizing a competitive and selective advantage of S. 1,4,[5],12:i:- over S. Typhimurium, which may be aided by microevolution affecting antigenicity, pathogenicity, and transmission [67]. Because of its high transmission ability, this serovar has become a global public and animal health hazard [56].
3.2.2. Clinical Signs and Gross Lesions
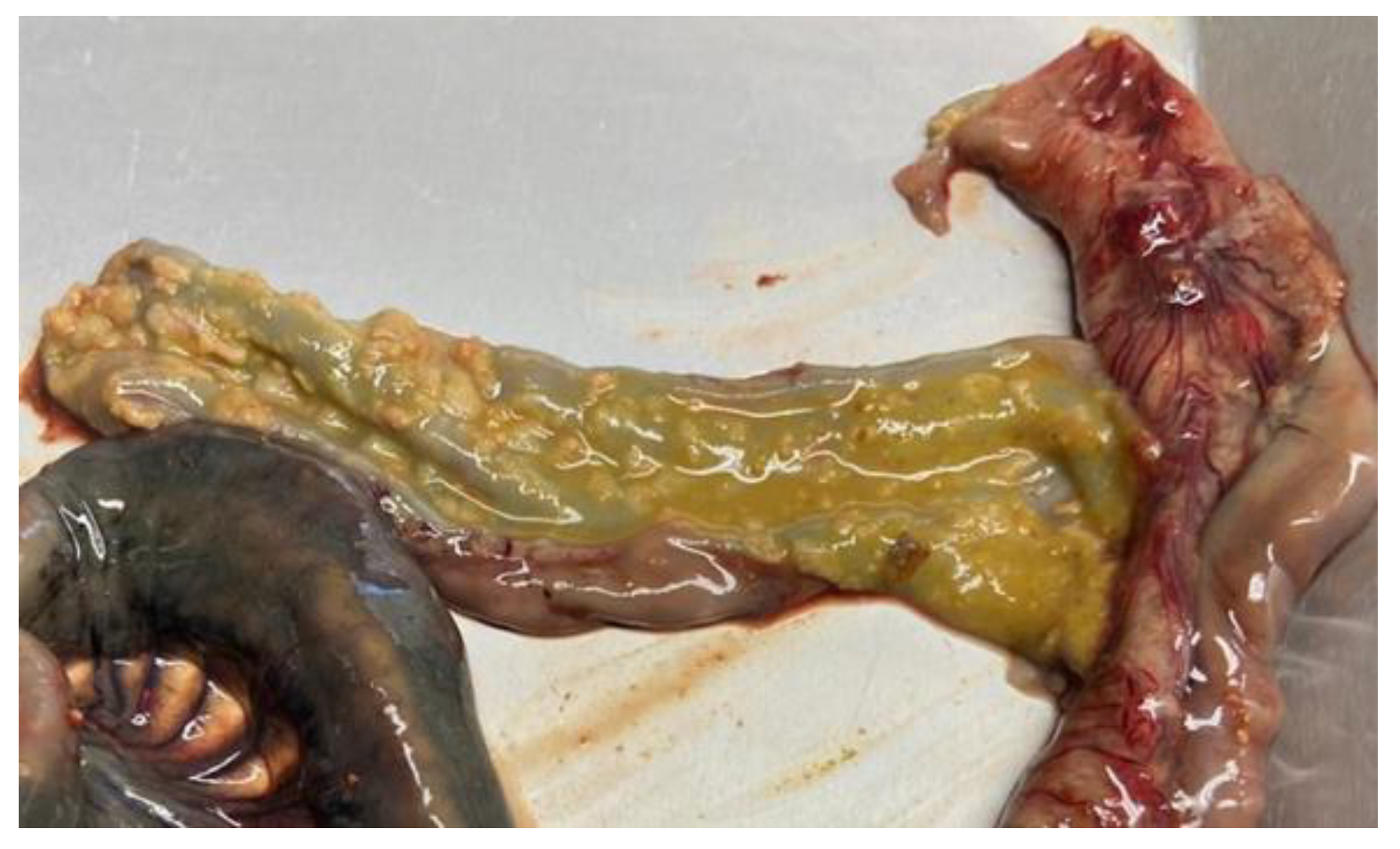
Salmonella infections in pigs are frequently asymptomatic, even though fecal shedding can occur continuously or intermittently, even over long periods of time, regardless of the presence of clinical signs. In symptomatic pigs, indistinguishable clinical signs can be observed after infection with S. Typhimurium and S. 1,4,[5],12:i:- [68], which are characterized by yellow diarrhea (rarely containing blood), dehydration, decreased feed intake, fever, inanition, and wasting. Debilitating conditions, such as poor hygiene and viral infections that induce immunosuppression, e.g., porcine reproductive and respiratory virus (PRRSV) and porcine circovirus 2 (PCV2), can trigger the onset of disease. Mortality is usually low and due to hypokalemia and dehydration, often occurring after several days of diarrhea [68]. After complete clinical recovery, a percentage of pigs can act as carriers and intermittent shedders for at least 5 months. The persistence of the organism is particularly frequent in submandibular and ileocolic lymph nodes, tonsils, and in the large intestine [68]; in addition to that, stressful conditions for the animals, e.g., commingling, transport, diet change, and lairage can increase shedding [69,70]. A few pigs may remain chronically wasted, and some may show obstipation and marked distension of the abdomen. This condition is described as a consequence of rectal strictures due to defective healing, with fibrosis, of ulcerative proctitis caused by S. Typhimurium [30]. Necrotic enterotyphlocolitis is the most consistent gross lesion in pigs suffering from S. Typhimurium. The lesions are mainly recorded in the ileum, cecum, and spiral colon, with the formation of diphtheritic membranes on the mucosal surface and roughened mucosa with a granular appearance. Systemic dissemination and septicemia are rare [68] (Figure 2).
Multifocal or coalescing mucosal erosions and ulcers may be detected, associated with adherent grey-yellow fibrino-necrotic material, while mesenteric lymph nodes are markedly enlarged and congested [30].
3.3. S. Derby
As opposed to S. Choleraesuis and S. Typhimurium, S. Derby is not known to be a cause of significant enteric disease in pigs, despite being frequently isolated from swine [53]. During a study in which pigs were challenged with S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby, no visible gross lesions were observed in necropsied pigs infected with S. Derby, as opposed to those infected with S. Typhimurium and S. 1,4,[5],12:i:-. With regard to histological lesions (mainly in the cecum and spiral colon), higher scores were registered for piglets infected with S. Typhimurium and S. 1,4,[5],12:i:- compared to those challenged with S. Derby [53]. Given this, S. Derby is known for its ability to cause durable asymptomatic infections in swine [71]. However, in another study in which piglets were inoculated with S. Derby, the animals showed diarrhea and fever [66]. As a matter of fact, it has been demonstrated that piglets experimentally challenged with S. Derby shed the pathogen in the feces at higher levels compared to those challenged with S. Typhimurium [72]. Taken together, these data could explain the wide diffusion of this serovar in pigs and why S. Derby is among the most reported serovars linked to the pork industry [73].
4. Salmonella Choleraesuis, S. Typhimurium, and S. 1,4,[5],12:i:- Infection in Swine: Carrier State and Environmental Persistence
Among the main Salmonella transmission sources on farm grounds, there are the introduction of infected animals [74], contaminated feed [75,76,77], and pests, such as rodents, birds, and flies, which can act as Salmonella vectors [31,78,79]. However, asymptomatic Salmonella carriers are one of the main infection sources, with the fecal–oral route being the main way of Salmonella transmission among pigs [76]. There has also been evidence of the role played by the upper respiratory tract as an equally important route of Salmonella infection in swine [80].
4.1. S. Choleraesuis
A study involving experimental infection in pigs with S. Choleraesuis evidenced that the pathogen was prevalently recovered from ileocolic lymph nodes, tonsils, lungs, colon, and cecal content [81]. Following oral transmission, tonsils become rapidly contaminated with Salmonella, which can then be found in oropharyngeal secretions; this could allow for nose-to-nose transmission [30]. In a study in which pigs were challenged intranasally with S. Choleraesuis, the bacterium could persist in the tonsils for up to 19 weeks post-infection. These data led researchers to hypothesize that tonsils could be a crucial site for S. Choleraesuis maintenance in swine herds [82]. In addition, S. Choleraesuis spread and persistence at herd level is strongly influenced by the carrier state recovered animals can acquire, shedding the bacteria through their feces for extended periods of time [51]. It was also demonstrated that, in pigs challenged with 108 CFU/mL of S. Choleraesuis, the carrier state could be maintained for up to 12 weeks, regardless of the inoculation route [82]. An experimental infection of neonatal piglets with S. Choleraesuis was performed to evaluate the pattern of long-term shedding, which revealed continuous shedding for up to 85 days after infection [83]. Given that piglets infected with S. Choleraesuis rarely exhibit clinical signs, shedding by subclinically infected piglets could play a role in the spread of Salmonella on farm grounds [83]. In addition to that, the carrier state, as well as the frequency and duration of shedding, are not influenced by the use of antibiotics in pigs. This differs from what is described in human enteric salmonellosis, where the carrier state is prolonged by the use of antibiotics [84,85,86,87,88,89].
The main routes of S. Choleraesuis transmission in swine herds are believed to be the horizontal transfer from carrier or diseased pigs to healthy individuals [83], and the facilities which were previously contaminated by this serovar [77]. In contrast, it is rare to find S. Choleraesuis in feed or in animals other than pigs [30,31]. Additionally, it has been proven that S. Choleraesuis is capable of surviving in wet swine feces for 3 months after shedding, while in dry feces, the bacterium survives for at least 13 months. Hence, S. Cholerasuis is capable of remaining viable and infective in the environment for extended periods of time [90]. Furthermore, S. Choleraesuis can remain dormant in swine herds and then be activated by other diseases, such as coinfection with PRRSV or PCV2, which induce an immunosuppressed condition in affected pigs [48].
4.2. S. Typhimurium and S. 1,4,[5],12:i:-
After infection with S. Typhimurium and its monophasic variant, the majority of pigs recover completely, while a lower portion acquire the carrier state, shedding the pathogen intermittently for up to 5 months [30]. After challenge, S. Typhimurium has been shown to persist in low numbers in swine for up to 28 weeks; moreover, after oral experimental challenge in pigs, S. Typhimurium showed a marked tropism for the tonsils [74,81,82]. The highest excretion rates were recorded at 2 weeks post-infection in a study in which 42-day-old piglets were orally challenged with 109 CFU of S. Typhimurium; after that, shedding decreased and became intermittent. Moreover, this study evidenced that long-term S. Typhimurium persistence in swine is restricted to the tonsils, the gastrointestinal tract, gut-associated lymphatic tissue, and mesenteric lymph nodes [91].
Another study evaluated S. Typhimurium prevalence on a pig farm, characterized by recurrent infections with this serovar. The authors detected the same S. Typhimurium PFGE profile from fecal samples isolated from farm grounds, waste slurry, agricultural soil spread with Salmonella-positive animal waste, and also from samples isolated from asymptomatic carrier pigs. These findings highlighted the ability of S. Typhimurium to persist in the environment, particularly in agricultural soil, for 14 days after spreading contaminated slurry. This persistence could fuel a cycle of continuous reinfection in herds. As a result, adequate and effective waste management practices are critical to contrast the long-term survival of Salmonella on farm grounds [92]. In addition, S. Typhimurium long-term survival in pig slurry was also reported (34 days in artificially contaminated slurry) [93]. It was also reported that Salmonella can survive for up to 50 months in the environment (e.g., slurry and dust) [94]. In addition to that, it has been demonstrated that S. Typhimurium can survive for up to 5 weeks in soil and up to 7 weeks in pig shelter huts, further leading to environmental contamination [95].
In another study, in which swine feces were artificially inoculated with both S. Typhimurium and S. 1,4,[5],12:i:-, bacteria were able to survive for 88 days [96]. Moreover, in a study in which pigs were exposed to low environmental doses of S. 1,4,[5],12:i:-, the excretion rate reached 104 CFU/g in feces [97]. For S. Typhimurium, both low, persistent carriers, as well as super shedders, have been documented [16,98]. Taken together, these findings confirm that sick animals’ shedding of Salmonella can contaminate the environment, continuously fueling a cycle of reinfection in newly introduced animals [90].
5. Prevention and Control of Salmonella Infection in Pig Farms and at Slaughter
Humoral immunity to Salmonella infections has a limited effect because for much of the infection cycle, the organism stays within body cells, shielded from antibody action [99]. However, there is a strong humoral response to natural infection, including secretory IgA responses that may be effective in preventing the initial invasion of the mucosa [100]. Cell-mediated immunity (CMI), characterized by a T-helper1 (Th1) lymphokine profile associated with the activation of macrophages and cytotoxic lymphocytes, appears to be a critical part of effective anti-Salmonella immunity [101]. In general, for particular strains, such as host-adapted S. Choleraesuis, it is often necessary to control the outbreaks of clinical disease, which have an important economic impact in terms of mortality and antibiotic consumption. Vaccination can be an effective tool to control Salmonella infections at the farm level. This approach has substantial differences when the goal of vaccination is to protect the consumers of pig products by controlling subclinical carriage and shedding [99]. In this latter case, vaccination should guarantee:
- The control of a broad variety of strains and serovars;
- The reduction in tissue colonization and/or shedding at the time of slaughter;
- No adverse effect on serological monitoring for Salmonella infection where this is employed before or at the time of slaughter, as performed in Denmark and other European countries to categorize the level of Salmonella infection in pig herds [102].
It has been demonstrated that vaccination against S. Choleraesuis cross-protected pigs against other strains, such as S. Typhimurium [103,104] and S. Derby [105], showing a certain degree of cross-protection between serovars [106]. Despite this, it is generally accepted that serovar-specific vaccines are more likely to be effective, as antibodies induced by different Salmonella serovars show only a low level of cross-protection [106].
In experimental challenge and field studies, inactivated vaccines with appropriate administration protocols and adjuvants showed protective effects against antigenically similar strains [99], whereas live vaccines should be able to cross-protect vaccinated animals against different Salmonella serogroups. The use of an autogenous, inactivated vaccine prepared from outbreak strains is a rapid intervention that may be effective, in concert with other control measures, if a licensed commercial vaccine is not available [107,108]. Alborali and colleagues (2017) showed that the combination of S. Typhimurium- and S. Choleraesuis-attenuated and inactivated vaccines, respectively, is effective against challenge infection with S. Choleraesuis. As, in field conditions, the simultaneous infection with more than one serovar is common, these data suggest that the development of a new effective vaccine with this strategy could be promising in tackling the effects of Salmonella infection [109].
Vaccination is widely accepted to play a role in reducing Salmonella prevalence in pigs, and it may become an adjunct to on-farm control [110], by preventing Salmonella from colonizing the gut and, as a result, reducing subsequent shedding and the development of the carrier state [111]. Moura and colleagues (2021) showed that vaccinating pigs twice (with an interval of 21 days) with a commercial inactivated vaccine containing strains of S. Choleraesuis, Pasteurella multocida, and S. Typhimurium, and then performing an oral challenge with 108 CFU of S. Typhimurium, partially protected the animals, reducing Salmonella excretion in feces and the colonization of organs [112]. The strategy combining maternal and pre-weaned piglets’ vaccination is needed for better protection against challenge in the post-weaning period [99]. Nonetheless, vaccination alone is not sufficient to eliminate the infection; a combination of different strategies such as biosecurity measures, cleaning and disinfection procedures, and feeding practices is required in order to reduce Salmonella prevalence in pigs.
An all-in/all-out production system can prevent cross-contamination between production cycles by allowing thorough cleaning and disinfection and, consequently, reducing the potential of Salmonella exposure and infection in subsequent batches [113]. This is of huge importance, considering that Salmonella can persist in the environment for several months to years [92,94]; furthermore, the farm environment can act as a Salmonella reservoir because of inadequate disinfection. In addition, Salmonella intestinal infection is positively correlated with ambient temperature (between 35 °C and 37 °C), as warmer temperatures enable rapid replication [114].
To limit Salmonella persistence and spread in farms and lairage environments, appropriate cleaning and disinfection regimes must be implemented. A significant reduction in the prevalence of Salmonella in pigs in appropriately cleaned and disinfected buildings was demonstrated, as well as the correct use of boot dips [115]. As an example, Gradel and colleagues (2004) reported that the preferred class of disinfectants appears to be peroxygen-based products [116]. Other disinfectants effective against Salmonella are glutaraldehyde, quaternary ammonium compounds (QACs), iodine-based compounds, and chlorocresols [78]. Disinfectants’ effectiveness may be compromised by the presence of organic matter or by over-dilution if used before the surfaces are completely dried [116]. The drying of pens greatly reduces the probability of detecting Salmonella, and its complete elimination was reported to be achieved 24 hours after cleaning with detergents and a chlorocresol-based disinfectant [117]. Smooth surfaces are less likely to have a high level of residual contamination than rough ones [118]. This is important, especially because concrete is a material that is widely used in pig farrowing accommodations and its rough surface could harbor higher numbers of bacteria. It has been reported that Salmonella contamination is very quickly transferred to roads, standing water, transport trucks, and other mobile equipment [119], thus making vehicles’ disinfection a crucial part of any rigorous biosecurity practices [120]. Fecal contamination of feed, drinkers, or farm equipment by rodents, wild birds, insects, and pets (dogs and cats) can represent a possible way for the introduction and transmission of Salmonella to pigs [121,122]. In particular, rodents can efficiently contribute to spreading Salmonella as they are very efficient vectors and amplifiers of the pathogen [115]. Rodenticides and biosecurity procedures, as well as the disposal of dead animal and feed remains, should be used in combination for efficient pest control [122].
Furthermore, the feed form is a very important risk factor. Wet feed is preferred to dry feed [123], while dry meal feed is preferred over pelleted feed, as long as the feed particle size of the meal is not too small [124]. Given that pellets have to be made from very finely ground ingredients to maintain their integrity, this reduces the transit time through the digestive tract and therefore does not achieve a low protective intestinal pH. On the contrary, coarsely ground meal decreases the survival of Salmonella during stomach passage because of slower gastric transit and lower gastric pH [125], thus improving productivity in pigs and delaying exposure to Salmonella [126]. Heating treatment of feed, performed at 93 °C for 90 seconds with 15% moisture, may eliminate Salmonella [127], even if the contamination level and possible post-treatment contamination are critical factors.
Manure management is important in order to reduce the risk of introduction and spread of Salmonella, as well as other pigs’ infectious agents. For this reason, manure treatments should be implemented in order to reduce or kill Salmonella. Anaerobic digestion, composting, and separation technologies are used for this purpose [128].
All these mitigation strategies should be coupled with appropriate measures to reduce the risk of carcass contamination by Salmonella at slaughter [129]. Pigs entering the slaughter line are first stunned, killed, and then exsanguinated. After that, a series of treatments, e.g., scalding, dehairing, singeing, and polishing, are performed on carcasses in order to lower microbiological contamination [130]. All of these steps should be performed in accordance with good manufacturing practices (GMPs) and following strict cleaning protocols and hygiene practices [130]; for example, it has been demonstrated that scalding water should always be kept between 60–62 °C to keep the water free from Salmonella contamination [131]. The aforementioned stages are followed by evisceration, which is one of the crucial stages in the slaughtering process; the accidental leakage of intestinal content due to perforation is one of the leading causes of Salmonella carcass contamination [130]. Moreover, slaughterhouse personnel’s hygiene and utensil cleaning and sanitation are necessary to avoid permanent contamination [130]; for these reasons, personnel should be properly trained about correct working procedures. In addition, proper carcass splitting, decontamination, and chilling procedures should be performed to at least reduce, if not eliminate, bacterial contamination [131]. The control of transportation and refrigeration temperature is crucial in hindering Salmonella outgrowth during food storage at retail level [131]. Lastly, correct food handling practices are to be adopted, coupled with high hygienical standards and proper cooking temperatures, in order to avoid possible cross-contamination and to lower the risk of infection in humans [131].
6. Salmonella in Swine Farms and in the Pig Production Chain
6.1. Salmonella Prevalence in Swine Farms
The EFSA baseline survey conducted in 2008, in which pooled fecal samples were analyzed, reported an overall Salmonella prevalence of 31.8% and of 33.3% for pigs’ breeding and production holdings, respectively, in the EU. It was also reported that S. Typhimurium had a prevalence of 25.4% and of 20.1% in breeding and production holdings [132]. In more recent years, S. 1,4,[5],12:i:- and S. Typhimurium were among the most isolated serovars from pigs in the EU (Table 1) [13,133,134,135].
Furthermore, a retrospective analysis of Salmonella serovars isolated from pigs between 1994 and 2010 in Great Britain (GB) highlighted the predominance of both S. Typhimurium and S. Derby in the pig population; in addition, S. Typhimurium was the most reported serovar every year. Nevertheless, a decreasing trend in S. Typhimurium isolation was observed during this study, while at the same time, an increasing trend was registered for S. 1,4,[5],12:i:-, which accounted for 25% of all isolates in 2010, making it the second-most isolated serovar from pigs that year [62]. In the 2017–2021 time period, S. 1,4,[5],12:i:- and S. Typhimurium accounted, together, for more than 70% of all isolations from pigs in livestock productions in the United Kingdom [136].
With regard to the US, S. 1,4,[5],12:i:- was the most isolated serovar from swine in 2016, followed by S. Typhimurium and S. Derby (33%, 15%, and 9.6%, respectively), considering both clinical and non-clinical cases [137]. In addition, S. 1,4,[5],12:i:- has become the predominant serovar isolated from swine samples analyzed by veterinary diagnostic laboratories in the US since 2014 [64].
6.2. Salmonella Prevalence at Slaughterhouse
In a study comparing the on-farm prevalence of Salmonella infections in pigs to prevalence at slaughter, it was reported that on-farm prevalence often seemed to be lower, in part because of latent, undetectable carriers [138], which led to an underestimated on-farm prevalence [139]. Furthermore, Salmonella could be shed only after carrier animals had left the farm, mainly due to stress related to transport, commingling with different animals, and lairage [140], even after a short-term exposure [73,130]. In addition, the longer the time pigs spent in lairage, the higher the risk of Salmonella infection [141]. Lairage duration has been positively associated with Salmonella detection in pigs’ lymph nodes; it was reported that when the lairage time exceeded 12 hours, pigs were most likely to be infected with Salmonella from the lairage environment compared to pigs held in lairage for approximately 1–3 hours (16.7% versus 11.1%) [142].
Animals’ transport to the slaughterhouse, as well as transport time, together with stress induced by handling, feed withdrawal, and commingling, could exacerbate Salmonella shedding by infected animals, and simultaneously increase infection probability in healthy pigs [20,130,143]. In addition, both holding pens and trucks that are contaminated with Salmonella before and after transportation increase the possibility of infections [140,144].
At slaughter, pigs infected with Salmonella can carry the microorganism on the skin, in the oral cavity, in feces, and in lymph nodes [20,141]; this could potentially lead to carcass cross-contamination during slaughtering stages [129,145]. Furthermore, during slaughter, pig meat could be contaminated with Salmonella because of incorrect evisceration practices or when hygiene measures are not properly followed, which could involve the accidental leakage of cecal content, mainly feces. In turn, this could lead to carcass contamination [130,145,146]. A Dutch study estimated that approximately 55–90% of carcass contamination takes place during the evisceration process [145]. It has also been demonstrated that pigs coming from farms with a high Salmonella fecal positivity had higher rates of carcass contamination at slaughter [10], and that Salmonella isolation from ileocecal lymph nodes is believed to accurately reflect on-farm prevalence [73,130,147].
The 2006–2007 EFSA survey, which was conducted with the aim of evaluating Salmonella prevalence in slaughter pigs in the EU, reported a prevalence of 10.3% in lymph nodes (data from 24 MSs) and of 8.3% for carcass swabs (data from 13 MSs) [41]. This survey also ascertained a great deal of variability among different MSs [148]. Furthermore, it is worth pointing out that in the EU, there are still no harmonized programs for Salmonella control in the pig production chain, even though some MSs apply their own monitoring plans [13]. For example, Denmark, Estonia, Finland, Germany, Norway, and Sweden are applying Salmonella control programs in the swine population [149]. According to more recent data from European Competent Authorities (CAs), in the 2017–2021 period, the number of Salmonella-positive pig carcasses was lower (1.7–3.6%) than that reported in the EFSA survey for the 2006–2007 period. Data from food business operators’ (FBOp) self-monitoring were lower compared to data from CAs (Table 2).
In 2019, an abattoir-based Salmonella prevalence study performed in GB found an overall prevalence of 32.2% from cecal samples, in which S. 1,4,[5],12:i:- was the most common serovar (36.6%); moreover, S. 1,4,[5],12:i:- and S. Typhimurium, together, accounted for 41.1% of all isolates, further highlighting the close relationship between these serovars and swine [151]. In addition to this, an Irish report, in which pooled lymph nodes and cecal material were analyzed, evidenced a high Salmonella prevalence at slaughter (pooled cecal content—55.5%, ileocecal lymph nodes—31.7%, carcass swabs—11.5%), with S. Typhimurium and S. 1,4,[5],12:i:- accounting for 65% of all the isolates. This is consistent with data reporting that these two serovars are among the most circulating ones in swine across Europe [147].
In the US, with regard to the Salmonella positivity rate in market hogs’ and sows’ cecal content at slaughter, data coming from USDA-FSIS evidenced a Salmonella positivity rate ranging from 34.7% to 49.7% in market hogs and from 50% to 67.9% in sows from 2013 to 2021; S. Derby, S. Typhimurium, and S. 1,4,[5],12:i:- were among the most frequently reported serovars [152,153].
Compared to the EU, other countries are characterized by a higher Salmonella prevalence in swine at slaughter [154]. In 2011, a Chinese study assessing Salmonella prevalence at pig slaughterhouses in three districts of Henan Province reported a prevalence of 29.2% (considering both carcass surface swabs and lymph nodes), in which S. Typhimurium was the most isolated serovar (28.6%), followed by S. Derby (27.1%) [155]. Later on, in 2016, an overall Salmonella prevalence of 22.9% was registered in slaughterhouses in Wuhan Province after the analysis of rectal swabs, carcass swabs, and pork samples [156].
6.3. Salmonella Prevalence in Pig Meat
In recent years, S. 1,4,[5],12:i:-, S. Typhimurium, and S. Derby have been the most reported serovars out of serotyped isolates from pig meat in the EU (Table 3) [133,134,135]. For example, S. 1,4,[5],12:i:-, S. Derby, and S. Typhimurium accounted for, respectively, 26.6%, 21.3%, and 14% of serotyped Salmonella isolated from pig meat in the EU in 2019 (Table 3), further underlying the tight connection between these three serovars and the pig production chain.
In the US, according to NARMS, in the 2013–2021 period, Salmonella prevalence in retail pork samples ranged from 0.8% to 4.5% [152].
According to a Chinese report evaluating Salmonella prevalence in food samples collected from 2011 to 2014, the vast majority of S. 1,4,[5],12:i:- isolates (84.6%) came from beef and pig meat products considered together [55].
To summarize, the main serovars responsible for infection in pigs in Europe, in the US, and in China are S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby. Available data highlight the existing correlation between these serovars and the pig production chain. Taken together, the collected information highlights the urgency of strengthening and improving Salmonella control measures worldwide, with the aim of reducing Salmonella prevalence in pigs and in pig-derived foodstuffs.
7. Salmonella Serovars Associated with Human Infections and Correlation to Pork Products
In the EU, in the years 2017–2021, S. Typhimurium and S. 1,4,[5],12:i:- were, respectively, the second- and the third-most isolated serovar from human salmonellosis cases, after S. Enteritidis, while S. Derby was the fifth-most isolated serovar from humans, albeit with a lower prevalence (Table 4) [13].
The most isolated serovars from pigs (S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby) are also among the most reported serovars isolated from human salmonellosis cases in the EU in recent years, although S. Derby is reported with a lower prevalence as opposed to S. Typhimurium and S. 1,4,[5],12:i:- (Table 4). In addition to these data, S. Typhimurium was the second main serovar involved in human outbreaks (9%), followed by S. 1,4,[5],12:i:- (1.6%) in 2018 in the EU [134]. S. Derby high prevalence in swine and relatively lower prevalence in humans could be explained by the lack of some virulence-associated genes [7]. Furthermore, in the EU, in 2020, pork products were the foodstuffs most involved in human salmonellosis outbreaks after eggs and egg-derived products. S. Typhimurium was and still is mainly related to poultry and pigs, while S. 1,4,[5],12:i:- and S. Derby are primarily linked to swine [150].
In the US, a total of 902 foodborne outbreaks were registered in 2015. Salmonella was the second-most prevalent cause of illness, being responsible for 34% of the outbreaks and for 39% of all the illnesses. Pork products were among the top three main food categories involved; S. Enteritidis was the most isolated serovar (35%), followed by S. 1,4,[5],12:i:- (10%) [157]. Since 2011, according to reports made by the laboratory-based Enteric Disease Surveillance system, the S. 1,4,[5],12:i:- human cases incidence rate has increased by 580% in the US. This considerable rise could be explained in part by the increased awareness toward this serovar [158]. In 2016, a total of 46,623 human salmonellosis cases were recorded, with both S. Typhimurium (9.8%) and S. 1,4,[5],12:i:- (4.7%) among the five most isolated serovars in the US (third and fifth, respectively) [158].
Moreover, the role played by pigs in the transmission of Salmonella to humans via contaminated food products has been documented, and several studies have highlighted that S. 1,4,[5],12:i:- and S. Typhimurium strains isolated from pork were responsible for foodborne outbreaks in humans (Table 5) [13,135,150].
Taking S. Choleraesuis into consideration, to the best of our knowledge, there is a paucity of information about foodborne outbreaks caused by this serovar, mainly because of its low incidence in humans, given that this serovar is swine-adapted. Nevertheless, in 2021, in Northern Italy, according to data from our institution, five S. Choleraesuis human cases showed a high degree of genomic similarity with one isolate coming from swine in the Emilia-Romagna region (unpublished data).
In conclusion, in order to control Salmonella along the pig production chain, the provided data strengthen the need to apply rigorous hygiene policies throughout the entire production system, coupled with surveillance systems implementing an interdisciplinary, holistic approach, in accordance with the One Health paradigm [159].

{kind=link}
{kind=link}
Table 5.
Example of pork product-derived Salmonella foodborne outbreaks (2004–2021), associated with S. 1,4,[5],12:i:- and S. Typhimurium in the EU and in the US. Modified from Campos et al., 2019 [11].
Table 5.
Example of pork product-derived Salmonella foodborne outbreaks (2004–2021), associated with S. 1,4,[5],12:i:- and S. Typhimurium in the EU and in the US. Modified from Campos et al., 2019 [11].
| Serovar | Year(s) | Country(ies) | Human Cases | Infection Source | Reference |
|---|---|---|---|---|---|
| 2006 | Luxembourg | 133 | Pork meat | [160] | |
| S. 1,4,[5],12:i:- | 2010 | France | 69 | Dried pork sausage | [161] |
| 2011 | Italy | 16 | Cooked pork product | [162] | |
| 2011 | France | 337 | Dried pork sausage | [163] | |
| 2011 | Spain | 38 | Dried pork sausage | [164] | |
| 2013 | Italy | NS | Pork salami | [165] | |
| 2013 | Germany | 61 | Minced pork | [166] | |
| 2015 | USA | 188 | Pork meat | [167] | |
| 2017 | Greece | 37 | Pork meat | [168] | |
| 2018 | England | 15 | Pork | [169] | |
| 2018–2019 | Denmark | 49 | Raw pork sausage | [170] | |
| 2020–2021 | France | 11 | Dried pork sausages | [171] | |
| 2021 | USA | 34 | Italian-style salami sticks | [172] | |
| S. Typhimurium | 2004 | Italy | 63 | Pork salami | [173] |
| 2005 | Denmark | 26 | Pork products | [174] | |
| 2008 | Denmark, Norway, Sweden | 37, 10, 4 | Danish pork meat, minced meat | [175] | |
| 2008 | Denmark | 1054 | Pork products | [176] | |
| 2010 | Italy | 30 | Pork salami | [177] | |
| 2010 | Italy | 5 | Pork salami | [177] | |
| 2010 | Denmark | 20 | Pork salami | [178] | |
| 2010 | Denmark | 172 | Pork products | [179] | |
| 2011 | Denmark | 22 | Smoked pork tenderloin | [180] | |
| 2011 | England | 51 | Pork products | [181] | |
| 2011 | Spain | 8 | Dried pork sausage | [164] | |
| 2018 | England | 28 | Pork | [169] | |
| 2021 | USA | 26 | Italian-style meats | [182] |
8. Surveillance of Foodborne Salmonellosis Outbreaks
Foodborne outbreaks due to Salmonella spp. have a strong impact on public health, causing illness with high hospitalization rates and significant socio-economic costs [183]. Taking this into consideration, laboratory-based surveillance is one of the pillars in the field of infectious disease monitoring [184]. Typically, it is carried out by public health and clinical laboratories. These laboratories collect and analyze samples from patients who are suspected to be ill with foodborne diseases. Then, positive cases are reported to public health authorities and strains are sent to reference laboratories for further testing [185,186]. Therefore, laboratory-based surveillance is a crucial part of monitoring foodborne diseases at a worldwide level, with the aim of collecting data on zoonoses trends and to detect and confirm outbreaks [185,187]. As previously stated, most salmonellosis cases in humans are associated with the consumption of contaminated animal-derived food products, such as pig meat, although in most cases, the origin of the infection is difficult to identify. The assignment of possible infection sources needs methods for targeted pathogen monitoring and rapid cluster detection, in order to reduce the burden posed by this disease and to limit the spread of infections [188].
Prior to the advent of whole genome sequencing (WGS), traditional bacterial typing techniques such as serotyping, phage typing, PFGE, MLVA, and MLST (multilocus sequence typing) were the standard typing tools for several foodborne bacterial pathogens [189]. Even though these techniques are still employed all over the world, they lack the ability to accurately discriminate between isolates during outbreaks [189,190]. In recent years, WGS has emerged as a powerful tool for genotyping pathogenic strains, including Salmonella, showing greater sensitivity and specificity compared to traditional methods [191,192]. WGS allows for an increased ability to discriminate reliably between isolates from the same species that are related, thus enhancing outbreak detection and discrimination power [193]. As a result, WGS is regarded as an “all-in-one” test, since data that would normally be obtained by a combination of traditional typing techniques can be retrieved in silico from sequencing data [194]. WGS has become and continues to become a key feature of public health surveillance systems [195,196]. Public health laboratories are implementing WGS approaches to better track and manage Salmonella foodborne-derived outbreaks [197] and, as of lately, many public health laboratories have successfully transitioned to an integrative WGS surveillance system of clinically relevant pathogens, such as PulseNet in the US [195]. To conclude, WGS has made it possible not only to perform better discrimination between outbreak-related and sporadic isolates but also to connect sporadic human cases to specific foodstuffs or animal sources; moreover, it allows us to identify contamination points across the food chain, which enables interventions during contaminated product trace-back analysis and eventual product recalls [189]. Nevertheless, it is important to acknowledge that to perform meaningful and accurate epidemiological surveillance, it is crucial to combine genomic evidence with clinical and epidemiological data [196]. Moreover, to prompt better inter-laboratory coordination and communication, standardized international protocols and workflows must be implemented [198,199], in order to improve and to guarantee microbial food safety and consumer protection. This translates to a transdisciplinary and multisectoral “One Health” approach, aimed at managing the threat posed by foodborne diseases, with the collaboration of experts comprising human, animal, and environmental health [200].
9. Conclusions
Salmonellosis and Salmonella infections in pigs are a problem for both animal and human health. Swine salmonellosis causes economic losses due to its lethality rate, growth retardation, and the need for antimicrobial usage. Salmonellosis also has an important impact on human health, as it is one of the most frequently reported foodborne diseases in EU countries. As reported, pork products are one of the main sources responsible for human salmonellosis. As of now, S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby are the most prevalent serovars linked to the pork production chain at a worldwide level. For this reason, it is important to improve knowledge about the role of carrier pigs, the epidemiology of the infection, the on-farm risk factors, the distribution of Salmonella serovars among pigs, and contamination routes at slaughter.
Different risk factors can influence Salmonella prevalence at the farm level, and its reduction needs different measures to be combined. Mitigation strategies should be adopted or reinforced, mainly regarding on-farm biosecurity, uncontaminated feed provision, quarantine of newly introduced animals, and implementation of cleaning and disinfecting practices. Vaccinating animals could contribute to decreasing Salmonella prevalence at slaughter, and procedures to prevent or minimize cross-contamination should be reinforced at the retail level. Measures at the farm level should be associated with strategies to reduce the risk of animal infection and carcass contamination in the stages immediately preceding slaughter. Transportation practices and holding at slaughter are crucial in influencing the prevalence of pigs positive for Salmonella entering the slaughter chain. In particular, proper hygienic practices showed a huge importance in reducing the level of contamination of carcasses at slaughter, reducing the persistence of Salmonella in the slaughter environment, and preventing the subsequent spread of Salmonella to pig carcasses.
Measures for reducing prevalence must be associated with effective Salmonella monitoring along the food chain, based on laboratory surveillance, in order to connect sporadic human cases to specific foodstuffs or animal sources; moreover, it allows us to identify contamination points across the food chain that enable interventions during contaminated product trace-back analysis and product recalls. Hence, more powerful control measures must be applied within an integrative and global “One Health” surveillance system, coordinating different stakeholders such as farmers, veterinarians working on animal health and food safety, and regulatory agencies.
Author Contributions
Conceptualization, A.L. and L.S.; data curation, L.S.; writing—original draft preparation, L.S., G.R. and A.L.; writing—review and editing, L.S., G.R., A.P., C.C. and A.L.; visualization, L.S., G.R., A.P., C.C. and A.L.; supervision, A.L and G.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
We would like to thank Stefano Pongolini, Risk Analysis and Genomic Epidemiology Unit, Istituto Zooprofilattico Sperimentale della Lombardia e dell’Emilia-Romagna (IZSLER), for his kind assistance and for his revision of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellido-Carreras, N.; Argüello, H.; Zaldívar-López, S.; Jiménez-Marín, Á.; Martins, R.P.; Arce, C.; Morera, L.; Carvajal, A.; Garrido, J.J. Salmonella Typhimurium Infection along the Porcine Gastrointestinal Tract and Associated Lymphoid Tissues. Vet. Pathol. 2019, 56, 681–690. [Google Scholar] [CrossRef]
- Jenkins , N.L.; Turner, J.L.; Dritz, S.S.; Durham, S.K.; Minton, J.E. Changes in circulating insulin-like growth factor-I, insulin-like growth factor binding proteins, and leptin in weaned pigs infected with Salmonella enterica serovar Typhimurium. Domest. Anim. Endocrinol. 2004, 26, 49–60. [Google Scholar] [CrossRef]
- Farzan, A.; Friendship, R.M. A clinical field trial to evaluate the efficacy of vaccination in controlling Salmonella infection and the association of Salmonella-shedding and weight gain in pigs. Can. J. Vet. Res. 2010, 74, 258–263. [Google Scholar]
- Gil Molino, M.; García, A.; Zurita, S.G.; Martín-Cano, F.E.; García-Jiménez, W.; Risco, D.; Rey, J.; Fernández-Llario, P.; Quesada, A. Spread of Antimicrobial Resistance by Salmonella enterica Serovar Choleraesuis between Close Domestic and Wild Environments. Antibiotics 2020, 9, 750. [Google Scholar] [CrossRef]
- Ojha, S.; Kostrzynska, M. Approaches for Reducing Salmonella in Pork Production. J. Food Prot. 2007, 70, 2676–2694. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States–major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-analysis. Appl. Env. Microbiol. 2019, 85, 591. [Google Scholar] [CrossRef] [PubMed]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- Foley, S.L.; Lynne, A.M.; Nayak, R. Salmonella challenges: Prevalence in swine and poultry and potential pathogenicity of such isolates1,2. J. Anim. Sci. 2008, 86, E149–E162. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.; Mourão, J.; Peixe, L.; Antunes, P. Non-typhoidal Salmonella in the Pig Production Chain: A Comprehensive Analysis of Its Impact on Human Health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef]
- Rodrigues, G.L.; Panzenhagen, P.; Ferrari, R.G.; Paschoalin, V.M.F.; Conte-Junior, C.A. Antimicrobial Resistance in Nontyphoidal Salmonella Isolates from Human and Swine Sources in Brazil: A Systematic Review of the Past Three Decades. Microb. Drug Resist. 2020, 26, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, 7666. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2013. EFSA J. 2015, 13, 3991. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Available online: https://www.cdc.gov/salmonella/index.html (accessed on 15 June 2022).
- Knetter, S.M.; Bearson, S.M.; Huang, T.; Kurkiewicz, D.; Schroyen, M.; Nettleton, D.; Berman, D.; Cohen, V.; Lunney, J.K.; Ramer-Tait, A.E.; et al. Salmonella enterica serovar Typhimurium-infected pigs with different shedding levels exhibit distinct clinical, peripheral cytokine and transcriptomic immune response phenotypes. Innate Immun. 2015, 21, 227–241. [Google Scholar] [CrossRef]
- Asghari, A.; Sadrebazzaz, A.; Shamsi, L.; Shams, M. Global prevalence, subtypes distribution, zoonotic potential, and associated risk factors of Blastocystis sp. in domestic pigs (Sus domesticus) and wild boars (Sus scrofa): A systematic review and meta-analysis. Microb. Pathog. 2021, 160, 105183. [Google Scholar] [CrossRef]
- Alarcón, L.V.; Allepuz, A.; Mateu, E. Biosecurity in pig farms: A review. Porc. Health Manag. 2021, 7, 5. [Google Scholar] [CrossRef]
- Delsart, M.; Pol, F.; Dufour, B.; Rose, N.; Fablet, C. Pig Farming in Alternative Systems: Strengths and Challenges in Terms of Animal Welfare, Biosecurity, Animal Health and Pork Safety. Agriculture 2020, 10, 261. [Google Scholar] [CrossRef]
- Lo Fo Wong, D.M.A.; Hald, T.; Van Der Wolf, P.J.; Swanenburg, M. Epidemiology and control measures for Salmonella in pigs and pork. Livest. Prod. Sci. 2002, 76, 215–222. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Jeggo, M. The One Health Approach—Why Is It So Important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef]
- Grimont, P.A.D.; Weill, F.-X. Antigentic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella Institut: Pasteur, Paris, 2007. Available online: https://www.pasteur.fr/sites/default/files/veng_0.pdf (accessed on 20 June 2022).
- Achtman, M.; Wain, J.; Weill, F.; Nair, S.; Zhou, Z.; Sangal, V.; Krauland, M.G.; Hale, J.L.; Harbottle, H.; Uesbeck, A.; et al. Multilocus Sequence Typing as a Replacement for Serotyping in Salmonella enterica. PLoS Pathog. 2012, 8, e1002776. [Google Scholar] [CrossRef] [PubMed]
- Fookes, M.; Schroeder, G.N.; Langridge, G.C.; Blondel, C.J.; Mammina, C.; Connor, T.R.; Seth-Smith, H.; Vernikos, G.S.; Robinson, K.S.; Sanders, M.; et al. Salmonella bongori provides insights into the evolution of the Salmonellae. PLoS Pathog. 2011, 7, e1002191. [Google Scholar] [CrossRef] [PubMed]
- Gay, N.; Le Hello, S.; Weill, F.; de Thoisy, B.; Berger, F. Salmonella serotypes in reptiles and humans, French Guiana. Vet. Microb. 2014, 170, 167–171. [Google Scholar] [CrossRef]
- Baumler, A.J.; Tsolis, R.M.; Ficht, T.A.; Adams, L.G. Evolution of host adaptation in Salmonella enterica. Infect. Immun. 1998, 66, 4579–4587. [Google Scholar] [CrossRef] [PubMed]
- Bjork, K.E.; Fields, V.; Garber, L.P.; Kopral, C.A. Factors Associated with Salmonella Prevalence in U.S. Swine Grower–Finisher Operations, 2012. Foodborne Pathog. Dis. 2018, 15, 489–497. [Google Scholar] [CrossRef]
- Holschbach, C.L.; Peek, S.F. Salmonella in Dairy Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Eswarappa, S.M.; Janice, J.; Balasundaram, S.V.; Dixit, N.M.; Chakravortty, D. Host-specificity of Salmonella enterica serovar Gallinarum: Insights from comparative genomics. Infect. Genet. Evol. 2009, 9, 468–473. [Google Scholar] [CrossRef]
- Griffith, R.W.; Carlson, S.A.; Krull, A.C. Salmonellosis. In Disease of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 912–925. [Google Scholar]
- Fedorka-Cray, P.J.; Gray, J.T.; Wray, C. Salmonella Infections in Pigs. In Salmonella in Domestic Animals; Wray, C., Wray, A., Eds.; CABI Publishing: New York, NY, USA, 2000; pp. 191–208. [Google Scholar]
- Leekitcharoenphon, P.; Sørensen, G.; Löfström, C.; Battisti, A.; Szabo, I.; Wasyl, D.; Slowey, R.; Zhao, S.; Brisabois, A.; Kornschober, C.; et al. Cross-Border Transmission of Salmonella Choleraesuis var. Kunzendorf in European Pigs and Wild Boar: Infection, Genetics, and Evolution. Front. Microbiol. 2019, 10, 179. [Google Scholar] [CrossRef]
- Meneguzzi, M.; Pissetti, C.; Rebelatto, R.; Trachsel, J.; Kuchiishi, S.S.; Reis, A.T.; Guedes, R.M.C.; Leão, J.A.; Reichen, C.; Kich, J.D. Re-Emergence of Salmonellosis in Hog Farms: Outbreak and Bacteriological Characterization. Microorganisms 2021, 9, 947. [Google Scholar] [CrossRef]
- Sugimoto, R.; Suzuki, H.; Nei, T.; Tashiro, A.; Washio, Y.; Sonobe, K.; Nakamura, Y.; Wakayama, N.; Inai, S.; Izumiya, H. Neck abscess due to Salmonella Choleraesuis: Case study and literature review. JMM Case Rep. 2017, 4, e005109. [Google Scholar] [CrossRef]
- Sriyapai, P.; Pulsrikarn, C.; Chansiri, K.; Nyamniyom, A.; Sriyapai, T. Molecular Characterization of Cephalosporin and Fluoroquinolone Resistant Salmonella Choleraesuis Isolated from Patients with Systemic Salmonellosis in Thailand. Antibiotics 2021, 10, 844. [Google Scholar] [CrossRef]
- Luk-in, S.; Chatsuwan, T.; Pulsrikarn, C.; Bangtrakulnonth, A.; Rirerm, U.; Kulwichit, W. High prevalence of ceftriaxone resistance among invasive Salmonella enterica serotype Choleraesuis isolates in Thailand: The emergence and increase of CTX-M-55 in ciprofloxacin-resistant S. Choleraesuis isolates. Int. J. Med. Microbiol. 2018, 308, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Hou, J.; Lin, H.; Wu, C.; Chu, C. Differences in immune responses of pigs vaccinated with Salmonella Typhimurium and S. Choleraesuis strains and challenged with S. Choleraesuis. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 41–47. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control). The Euro-pean Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar] [CrossRef]
- Pedersen, K.; Sørensen, G.; Löfström, C.; Leekitcharoenphon, P.; Nielsen, B.; Wingstrand, A.; Aarestrup, F.M.; Hendriksen, R.S.; Baggesen, D.L. Reappearance of Salmonella serovar Choleraesuis var. Kunzendorf in Danish pig herds. Vet. Microbiol. 2015, 176, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, S. Salmonella in the pork production chain and its impact on human health in the European Union. Epidemiol. Infect. 2017, 145, 1513–1526. [Google Scholar] [CrossRef]
- Report of the Task Force on Zoonoses Data Collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, Part A. EFSA J. 2008, 6, 135r. [CrossRef]
- Enter-Vet Report Dati 2019. Available online: https://www.izsvenezie.it/documenti/temi/salmonellosi/enter-vet/entervet-report-2019.pdf (accessed on 28 July 2023).
- Savic, B.; Zdravkovic, N.; Radanovic, O.; Jezdimirovic, N.; Kureljusic, B.; Stevancevic, O. A Salmonella enterica subspecies enterica serovar Choleraesuis outbreak in weaned piglets in Serbia: Clinical signs, pathologic changes, and microbiologic features. J. Vet. Diagn. Investig. 2021, 33, 993–996. [Google Scholar] [CrossRef]
- Ernholm, L.; Sternberg-Lewerin, S.; Ågren, E.; Ståhl, K.; Hultén, C. First Detection of Salmonella enterica Serovar Choleraesuis in Free Ranging European Wild Boar in Sweden. Pathogens 2022, 11, 723. [Google Scholar] [CrossRef]
- Papić, B.; Kušar, D.; Mićunović, J.; Vidrih, Š.; Pirš, M.; Ocepek, M.; Avberšek, J. Genomic insights into Salmonella Choleraesuis var. Kunzendorf outbreak reveal possible interspecies transmission. Vet. Microbiol. 2021, 263, 109282. [Google Scholar] [CrossRef]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Gil Molino, M.; Risco Pérez, D.; Gonçalves Blanco, P.; Fernandez Llario, P.; Quesada Molina, A.; García Sánchez, A.; Cuesta Gerveno, J.M.; Gómez Gordo, L.; Martín Cano, F.E.; Pérez Martínez, R.; et al. Outbreaks of antimicrobial resistant Salmonella Choleraesuis in wild boars piglets from central-western Spain. Transbound. Emerg. Dis. 2018, 66, 225–233. [Google Scholar] [CrossRef]
- Longo, A.; Losasso, C.; Vitulano, F.; Mastrorilli, E.; Turchetto, S.; Petrin, S.; Mantovani, C.; Dalla Pozza, M.C.; Ramon, E.; Conedera, G.; et al. Insight into an outbreak of Salmonella Choleraesuis var. Kunzendorf in wild boars. Vet. Microbiol. 2019, 238, 108423. [Google Scholar] [CrossRef]
- Methner, U.; Heller, M.; Bocklisch, H. Salmonella enterica subspecies enterica serovar Choleraesuis in a wild boar population in Germany. Eur. J. Wildl. Res. 2010, 56, 493–502. [Google Scholar] [CrossRef]
- Methner, U.; Merbach, S.; Peters, M. Salmonella enterica subspecies enterica serovar Choleraesuis in a German wild boar population: Occurrence and characterisation. Acta Vet. Scand. 2018, 60, 65. [Google Scholar] [CrossRef]
- Baskerville, A. Pneumonia of pigs: A review. N. Z. Vet. J. 1981, 29, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Hyland, K.A.; Brown, D.R.; Murtaugh, M.P. Salmonella enterica serovar Choleraesuis infection of the porcine jejunal Peyer’s patch rapidly induces IL-1β and IL-8 expression. Vet. Immunol. Immunopathol. 2006, 109, 1–11. [Google Scholar] [CrossRef]
- Naberhaus, S.A.; Krull, A.C.; Arruda, B.L.; Arruda, P.; Sahin, O.; Schwartz, K.J.; Burrough, E.R.; Magstadt, D.R.; Matias Ferreyra, F.; Gatto, I.R.H.; et al. Pathogenicity and Competitive Fitness of Salmonella enterica Serovar 4,[5],12:i:- Compared to Salmonella Typhimurium and Salmonella Derby in Swine. Front. Vet. Sci. 2020, 6, 502. [Google Scholar] [CrossRef]
- Machado, J.; Bernardo, F. Prevalence of Salmonella in chicken carcasses in Portugal. J. Appl. Bacteriol. 1990, 69, 477–480. [Google Scholar] [CrossRef]
- Yang, X.; Wu, Q.; Zhang, J.; Huang, J.; Guo, W.; Cai, S. Prevalence and Characterization of Monophasic Salmonella Serovar 1,4,[5],12:i:- of Food Origin in China. PLoS ONE. 2015, 10, e0137967. [Google Scholar] [CrossRef]
- Sun, H.; Wan, Y.; Du, P.; Bai, L. The Epidemiology of Monophasic Salmonella Typhimurium. Foodborne Pathog. Dis. 2020, 17, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Echeita, M.A.; Herrera, S.; Usera, M.A. Atypical, fljB -Negative Salmonella enterica subsp. enterica Strain of Serovar 4,5,12:i:− Appears To Be a Monophasic Variant of Serovar Typhimurium. J. Clin. Microbiol. 2001, 39, 2981–2983. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, E.; Zapata, D.; Tello, M.; Mejia, W.; Frias, N.; Garcia Pena, F.J.; Mateu, E.M.; Torre, E. Several Salmonella enterica subsp. enterica Serotype 4,5,12:i:− Phage Types Isolated from Swine Samples Originate from Serotype Typhimurium DT U302. J. Clin. Microbiol. 2003, 41, 2395–2400. [Google Scholar] [CrossRef]
- Hauser, E.; Tietze, E.; Helmuth, R.; Junker, E.; Blank, K.; Prager, R.; Rabsch, W.; Appel, B.; Fruth, A.; Malorny, B. Pork Contaminated with Salmonella enterica Serovar 4,[5],12:i:−, an Emerging Health Risk for Humans. Appl. Environ. Microbiol. 2010, 76, 4601–4610. [Google Scholar] [CrossRef]
- Switt, A.I.M.; Soyer, Y.; Warnick, L.D.; Wiedmann, M. Emergence, Distribution, and Molecular and Phenotypic Characteristics of Salmonella enterica Serotype 4,5,12:i. Foodborne Pathog. Dis. 2009, 6, 407–415. [Google Scholar] [CrossRef]
- Elnekave, E.; Hong, S.; Mather, A.E.; Boxrud, D.; Taylor, A.J.; Lappi, V.; Johnson, T.J.; Vannucci, F.; Davies, P.; Hedberg, C.; et al. Salmonella enterica Serotype 4,[5],12:i:- in Swine in the United States Midwest: An Emerging Multidrug-Resistant Clade. Clin. Infect. Dis. 2018, 66, 877–885. [Google Scholar] [CrossRef]
- Mueller-Doblies, D.; Speed, K.; Davies, R.H. A retrospective analysis of Salmonella serovars isolated from pigs in Great Britain between 1994 and 2010. Prev. Vet. Med. 2013, 110, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.L.; Burrough, E.R.; Schwartz, K.J. Salmonella enterica I 4,[5],12:i:- Associated with Lesions Typical of Swine Enteric Salmonellosis. Emerg. Infect. Dis. 2019, 25, 1377–1379. [Google Scholar] [CrossRef]
- Naberhaus, S.A.; Krull, A.C.; Bradner, L.K.; Harmon, K.M.; Arruda, P.; Arruda, B.L.; Sahin, O.; Burrough, E.R.; Schwartz, K.J.; Kreuder, A.J. Emergence of Salmonella enterica serovar 4,[5],12:i:- as the primary serovar identified from swine clinical samples and development of a multiplex real-time PCR for improved Salmonella serovar-level identification. J. Vet. Diagn. Investig. 2019, 31, 818–827. [Google Scholar] [CrossRef]
- D’Incau, M.; Salogni, C.; Giovannini, S.; Ruggeri, J.; Scali, F.; Tonni, M.; Formenti, N.; Guarneri, F.; Pasquali, P.; Alborali, G.L. Occurrence of Salmonella Typhimurium and its monophasic variant (4, [5],12:i:-) in healthy and clinically ill pigs in northern Italy. Porc. Health Manag. 2021, 7, 34. [Google Scholar] [CrossRef]
- Cevallos-Almeida, M.; Martin, L.; Houdayer, C.; Rose, V.; Guionnet, J.; Paboeuf, F.; Denis, M.; Kerouanton, A. Experimental infection of pigs by Salmonella Derby, S. Typhimurium and monophasic variant of S. Typhimurium: Comparison of colonization and serology. Vet. Microbiol. 2019, 231, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Petrovska, L.; Mather, A.E.; AbuOun, M.; Branchu, P.; Harris, S.R.; Connor, T.; Hopkins, K.L.; Underwood, A.; Lettini, A.A.; Page, A.; et al. Microevolution of monophasic Salmonella Typhimurium during epidemic, United Kingdom, 2005–2010. Emerg. Infect. Dis. 2016, 22, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Plattner, B.L.; Hostetter, J.M. The alimentary system. In Jubb, Kennedy, and Palmer’s Pathology of Domestic Animals, 6th ed.; Grant Maxie, M., Ed.; Elsevier: St. Louis, MO, USA, 2016; Volume 2, pp. 1–257. [Google Scholar]
- Massacci, F.R.; Morelli, A.; Cucco, L.; Castinel, A.; Ortenzi, R.; Tofani, S.; Pezzotti, G.; Estellé, J.; Paniccià, M.; Magistrali, C.F. Transport to the Slaughterhouse Affects the Salmonella Shedding and Modifies the Fecal Microbiota of Finishing Pigs. Animals 2020, 10, 676. [Google Scholar] [CrossRef] [PubMed]
- Bernad-Roche, M.; Casanova-Higes, A.; Marín-Alcalá, C.M.; Mainar-Jaime, R.C. Salmonella Shedding in Slaughter Pigs and the Use of Esterified Formic Acid in the Drinking Water as a Potential Abattoir-Based Mitigation Measure. Animals 2022, 12, 1620. [Google Scholar] [CrossRef]
- Yuan, X.; Xue, H.; Xu, X.; Jiao, X.; Pan, Z.; Zhang, Y. Closely related Salmonella Derby strains triggered distinct gut microbiota alteration. Gut Pathog. 2022, 14, 6. [Google Scholar] [CrossRef]
- Matiasovic, J.; Stepanova, H.; Kudlackova, H.; Havlickova, H.; Sisak, F.; Rychlik, I.; Chlebova, K.; Leva, L.; Osvaldova, A.; Gebauer, J.; et al. Immune response of pigs to Salmonella enterica serovar Derby and Typhimurium infections. Vet. Microbiol. 2014, 170, 284–290. [Google Scholar] [CrossRef]
- Roasto, M.; Bonardi, S.; Mäesaar, M.; Alban, L.; Gomes-Neves, E.; Vieira-Pinto, M.; Vågsholm, I.; Elias, T.; Lindegaard, L.L.; Blagojevic, B. Salmonella enterica prevalence, serotype diversity, antimicrobial resistance and control in the European pork production chain. Trends Food. Sci. Technol. 2023, 131, 210–219. [Google Scholar] [CrossRef]
- Fedorka-Cray, P.J.; Whipp, S.C.; Isaacson, R.E.; Nord, N.; Lager, K. Transmission of Salmonella typhimurium to swine. Vet. Microbiol. 1994, 41, 333–344. [Google Scholar] [CrossRef]
- Fedorka-Cray, P.J.; Hogg, A.; Gray, J.T.; Lorenzen, K.; Velasquez, J.; Von Behren, P. Feed and feed trucks as sources of Salmonella contamination in swine. J. Swine Health Prod. 1997, 5, 189–193. [Google Scholar]
- Boyen, F.; Haesebrouck, F.; Maes, D.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef]
- Davies, P.R.; Scott Hurd, H.; Funk, J.A.; Fedorka-Cray, P.J.; Jones, F.T. The Role of Contaminated Feed in the Epidemiology and Control of Salmonella enterica in Pork Production. Foodborne Pathog. Dis. 2004, 1, 22–215. [Google Scholar] [CrossRef] [PubMed]
- De Lucia, A.; Ostanello, F. On-farm risk factors associated with Salmonella in pig herds. Large Anim. Rev. 2020, 26, 133–140. [Google Scholar]
- Fosse, J.; Seegers, H.; Magras, C. Prevalence and Risk Factors for Bacterial Food-Borne Zoonotic Hazards in Slaughter Pigs: A Review. Zoonoses Public Health 2009, 56, 429–454. [Google Scholar] [CrossRef]
- Fedorka-Cray, P.J.; Kelley, L.C.; Stabel, T.J.; Gray, J.T.; Laufer, J.A. Alternate routes of invasion may affect pathogenesis of Salmonella typhimurium in swine. Infect. Immun. 1995, 63, 2658–2664. [Google Scholar] [CrossRef]
- Gray, J.T.; Fedorka-Cray, P.J.; Stabel, T.J.; Kramer, T.T. Natural transmission of Salmonella choleraesuis in swine. Appl. Environ. Microbiol. 1996, 62, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.T.; Fedorka-Cray, P.J.; Stabel, T.J.; Ackermann, M.R. Influence of inoculation route on the carrier state of Salmonella choleraesuis in swine. Vet. Microbiol. 1995, 47, 43–59. [Google Scholar] [CrossRef]
- Anderson, R.C.; Genovese, K.J.; Harvey, R.B.; Stanker, L.H.; DeLoach, J.R.; Nisbet, D.J. Assessment of the long-term shedding pattern of Salmonella serovar choleraesuis following experimental infection of neonatal piglets. J. Vet. Diagn. Investig. 2000, 12, 257–260. [Google Scholar] [CrossRef]
- Aserkoff, B.; Bennett, J.V. Effect of Antibiotic Therapy in Acute Salmonellosis on the Fecal Excretion of Salmonellae. N. Engl. J. Med. 1969, 281, 636–640. [Google Scholar] [CrossRef]
- Dixon, J.M.S. Effect of Antibiotic Treatment on Duration of Excretion of Salmonella Typhimurium by Children. BMJ 1965, 2, 1343–1345. [Google Scholar] [CrossRef]
- Gutzmann, F.; Layton, H.; Simkins, K.; Jarolmen, H. Influence of antibiotic-supplemented feed on occurrence and persistence of Salmonella typhimurium in experimentally infected swine. Am. J. Vet. Res. 1976, 37, 649–655. [Google Scholar]
- Jacks, T.M.; Frazier, E.; Judith, F.R.; Olson, G. Effect of efrotomycin in feed on the quantity, duration, and prevalence of shedding and antibacterial susceptibility of Salmonella typhimurium in experimentally infected swine. Am. J. Vet. Res. 1988, 49, 1832–1835. [Google Scholar]
- Jones, F.T.; Langlois, B.E.; Cromwell, G.L.; Hays, V.W. Effect of Feeding Chlortetracycline or Virginiamycin on Shedding of Salmonellae from Experimentally-Infected Swine. J. Anim. Sci. 1983, 57, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Wilcock, B.; Olander, H. Influence of oral antibiotic feeding on the duration and severity of clinical disease, growth performance, and pattern of shedding in swine inoculated with Salmonella typhimurium. J. Am. Vet. Med. Assoc. 1978, 172, 472–477. [Google Scholar]
- Gray, J.T.; Fedorka-Cray, P.J. Survival and Infectivity of Salmonella Choleraesuis in Swine Feces. J. Food Prot. 2001, 64, 945–949. [Google Scholar] [CrossRef]
- Scherer, K.; Szabo, I.; Rosler, U.; Appel, B.; Hensel, A.; Nockler, K. Time Course of Infection with Salmonella Typhimurium and Its Influence on Fecal Shedding, Distribution in Inner Organs, and Antibody Response in Fattening Pigs. J. Food Prot. 2008, 71, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Baloda, S.B.; Christensen, L.; Trajcevska, S. Persistence of a Salmonella enterica Serovar Typhimurium DT12 Clone in a Piggery and in Agricultural Soil Amended with Salmonella -Contaminated Slurry. Appl. Environ. Microbiol. 2001, 67, 2859–2862. [Google Scholar] [CrossRef]
- Mannion, C.; Lynch, P.B.; Egan, J.; Leonard, F.C. Seasonal effects on the survival characteristics of Salmonella Typhimurium and Salmonella Derby in pig slurry during storage. J. Appl. Microbiol. 2007, 103, 1386–1392. [Google Scholar] [CrossRef]
- van der Wolf, P.; Meijerink, M.; Libbrecht, E.; Tacken, G.; Gijsen, E.; Lillie-Jaschniski, K.; Schüller, V. Salmonella Typhimurium environmental reduction in a farrow-to-finish pig herd using a live attenuated Salmonella Typhimurium vaccine. Porc. Health Manag. 2021, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.N.; Dalsgaard, A.; Stockmarr, A.; Nielsen, E.M.; Baggesen, D.L. Survival and Transmission of Salmonella enterica Serovar Typhimurium in an Outdoor Organic Pig Farming Environment. Appl. Environ. Microbiol. 2006, 72, 1833–1842. [Google Scholar] [CrossRef]
- Rajtak, U.; Boland, F.; Leonard, N.; Bolton, D.; Fanning, S. Roles of Diet and the Acid Tolerance Response in Survival of Common Salmonella Serotypes in Feces of Finishing Pigs. Appl. Environ. Microbiol. 2012, 78, 110–119. [Google Scholar] [CrossRef]
- Lynch, H.; Argüello, H.; Walia, K.; Lawlor, P.G.; Duffy, G.; Gardiner, G.E.; Leonard, F.C. Evaluation of an Alternative Experimental Infection Method, Which Closely Mimics the Natural Route of Transmission of Monophasic Salmonella Typhimurium in Pigs. Foodborne Pathog. Dis. 2017, 14, 23–28. [Google Scholar] [CrossRef]
- Berriman, A.D.C.; Clancy, D.; Clough, H.E.; Christley, R.M. Semi-stochastic models for Salmonella infection within finishing pig units in the UK. Math. Biosci. 2013, 245, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Wales, A.D.; Davies, R.H. Salmonella Vaccination in Pigs: A Review. Zoonoses Public Health 2017, 64, 1–13. [Google Scholar] [CrossRef] [PubMed]
- McSorley, S.J.; Cookson, B.T.; Jenkins, M.K. Characterization of CD4+ T Cell Responses During Natural Infection with Salmonella typhimurium. J. Immunol. 2000, 164, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P. Advances in swine immunology help move vaccine technology forward. Vet. Immunol. Immunopathol. 2014, 159, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Alban, L.; Stege, H.; Dahl, J. The new classification system for slaughter-pig herds in the Danish Salmonella surveillance-and-control program. Prev. Vet. Med. 2002, 53, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Charles, S.D.; Abraham, A.S.; Trigo, E.T.; Jones, G.F.; Settje, T.E. Reduced shedding and clinical signs of Salmonella Typhimurium in nursery pigs vaccinated with a Salmonella Choleraesuis vaccine. J. Swine Health Prod. 2000, 8, 107–112. [Google Scholar]
- Maes, D.; Gibson, K.; Trigo, E.; Saszak, A.; Grass, J.; Carlson, A.; Blaha, T. Evaluation of cross-protection afforded by a Salmonella Choleraesuis vaccine against Salmonella infections in pigs under field conditions. Berl. Munch. Tierarztl. Wochenschr. 2001, 114, 339–341. [Google Scholar]
- Groninga, K.J.; Springer, E.; Braunschmidt, M.; Pankratz, D. Salmonella derby cross-protection study. Vet. Ther. 2000, 1, 59–62. [Google Scholar]
- Foss, D.L.; Agin, T.S.; Bade, D.; Dearwester, D.A.; Jolie, R.; Keich, R.L.; Lohse, R.M.; Reed, M.; Rosey, E.L.; Schneider, P.A.; et al. Protective immunity to Salmonella enterica is partially serogroup specific. Vet. Immunol. Immunopathol. 2013, 155, 76–86. [Google Scholar] [CrossRef]
- Barrow, P.A.; Methner, U. Salmonella in Domestic Animals, 2nd ed.; CABI: Oxfordshire, UK, 2013. [Google Scholar]
- Roesler, U.; Heller, P.; Waldmann, K.H.; Truyen, U.; Hensel, A. Immunization of Sows in an Integrated Pig-breeding Herd using a Homologous Inactivated Salmonella Vaccine Decreases the Prevalence of Salmonella typhimurium Infection in the Offspring. J. Vet. Med. 2006, 53, 224–228. [Google Scholar] [CrossRef]
- Alborali, G.L.; Ruggeri, J.; Pesciaroli, M.; Martinelli, N.; Chirullo, B.; Ammendola, S.; Battistoni, A.; Ossiprandi, M.C.; Corradi, A.; Pasquali, P. Prime-boost vaccination with attenuated Salmonella Typhimurium ΔznuABC and inactivated Salmonella Choleraesuis is protective against Salmonella Choleraesuis challenge infection in piglets. BMC Vet. Res. 2017, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Denagamage, T.N.; O’Connor, A.M.; Sargeant, J.M.; Rajic, A.; McKean, J.D. Efficacy of vaccination to reduce Salmonella prevalence in live and slaughtered swine: A systematic review of literature from 1979 to 2007. Foodborne Pathog. Dis. 2007, 4, 539–549. [Google Scholar] [CrossRef]
- Haesebrouck, F.; Pasmans, F.; Chiers, K.; Maes, D.; Ducatelle, R.; Decostere, A. Efficacy of vaccines against bacterial diseases in swine: What can we expect? Vet. Microbiol. 2004, 100, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves De Oliveira Moura, A.; Gomes Da Silva, D.; Turco, H.; Thainara, V.C.; Sanches, G.Y.; Storino, H.; Meiroz De Souza Almeida, M.; Lopes Mechler-Dreibi, I.P.; Rabelo, K.; Sonalio, L.; et al. Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs. Microorganisms 2021, 9, 1163. [Google Scholar] [CrossRef]
- Gotter, V.; Klein, G.; Koesters, S.; Kreienbrock, L.; Blaha, T.; Campe, A. Main risk factors for Salmonella-infections in pigs in north-western Germany. Prev. Vet. Med. 2012, 106, 301–307. [Google Scholar] [CrossRef]
- Akil, L.; Ahmad, H.A.; Reddy, R.S. Effects of Climate Change on Salmonella Infections. Foodborne Pathog. Dis. 2014, 11, 974–980. [Google Scholar] [CrossRef]
- Martelli, F.; Lambert, M.; Butt, P.; Cheney, T.; Tatone, F.A.; Callaby, R.; Rabie, A.; Gosling, R.J.; Fordon, S.; Crocker, G.; et al. Evaluation of an enhanced cleaning and disinfection protocol in Salmonella contaminated pig holdings in the United Kingdom. PLoS ONE 2017, 12, e0178897. [Google Scholar] [CrossRef] [PubMed]
- Gradel, K.O.; Sayers, A.R.; Davies, R.H. Surface disinfection tests with Salmonella and a putative indicator bacterium, mimicking worst-case scenarios in poultry houses. Poult. Sci. 2004, 83, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Walia, K.; Argüello, H.; Lynch, H.; Grant, J.; Leonard, F.C.; Lawlor, P.G.; Gardiner, G.E.; Duffy, G. The efficacy of different cleaning and disinfection procedures to reduce Salmonella and Enterobacteriaceae in the lairage environment of a pig abattoir. Int. J. Food Microbiol. 2017, 246, 64–71. [Google Scholar] [CrossRef]
- Madec, F.; Humbert, F.; Salvat, G.; Maris, P. Measurement of the Residual Contamination of Post-Weaning Facilities for Pigs and Related Risk Factors. J. Vet. Med. 1999, 46, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Wales, A.D.; McLaren, I.M.; Bedford, S.; Carrique-Mas, J.J.; Cook, A.J.C.; Davies, R.H. Longitudinal survey of the occurrence of Salmonella in pigs and the environment of nucleus breeder and multiplier pig herds in England. Vet. Rec. 2009, 165, 648–657. [Google Scholar] [CrossRef]
- Twomey, D.F.; Miller, A.J.; Snow, L.C.; Armstrong, J.D.; Davies, R.H.; Williamson, S.M.; Featherstone, C.A.; Reichel, R.; Cook, A.J.C. Association between biosecurity and Salmonella species prevalence on English pig farms. Vet. Rec. 2010, 166, 722–724. [Google Scholar] [CrossRef]
- Zheng, D.M.; Bonde, M.; Sørensen, J.T. Associations between the proportion of Salmonella seropositive slaughter pigs and the presence of herd level risk factors for introduction and transmission of Salmonella in 34 Danish organic, outdoor (non-organic) and indoor finishing-pig farms. Livest. Sci. 2007, 106, 189–199. [Google Scholar] [CrossRef]
- Zamora-Sanabria, R.; Alvarado, A.M. Preharvest Salmonella Risk Contamination and the Control Strategies. In Current Topics in Salmonella and Salmonellosis; IntechOpen: London, UK, 2017; pp. 193–213. [Google Scholar]
- Belœil, P.A.; Fravalo, P.; Fablet, C.; Jolly, J.P.; Eveno, E.; Hascoet, Y.; Chauvin, C.; Salvat, G.; Madec, F. Risk factors for Salmonella enterica subsp. enterica shedding by market-age pigs in French farrow-to-finish herds. Prev. Vet. Med. 2004, 63, 103–120. [Google Scholar] [CrossRef]
- Lo Fo Wong, D.M.A.; Dahl, J.; Stege, H.; van der Wolf, P.J.; Leontides, L.; von Altrock, A.; Thorberg, B.M. Herd-level risk factors for subclinical Salmonella infection in European finishing-pig herds. Prev. Vet. Med. 2004, 62, 253–266. [Google Scholar] [CrossRef]
- Kjӕrsgaard, H.D.; Jørgensen, L.; Wachmann, H.; Dahl, J. Effect on Salmonella prevalence by feeding sows meal or pelleted feed. In Proceedings of the 4th International Symposium on Epidemiology and Control of Salmonella and other Foodborne Pathogens in Pork (Salinpork 2001), Leipzig, Germany, 2–5 September 2001; pp. 115–117. [Google Scholar]
- Roldan-Henao, M.; Dalsgaard, A.; Cardona-Castro, N.; Restrepo-Rivera, L.; Veloza-Angulo, L.C.; Alban, L. Pilot study of the productivity and Salmonella seroprevalence in pigs administered organic acids. Front. Vet. Sci. 2023, 10, 1123137. [Google Scholar] [CrossRef]
- Callegari, M.A.; Dalto, D.B.; da Silva, C.A. A Review of Prevention and Control Methods of Salmonella species in Swine Production and the Role of Dietary Non-Nutritional Additives. Asian J. Anim. Vet. Adv. 2015, 10, 803–829. [Google Scholar] [CrossRef]
- Ziemer, C.J.; Bonner, J.M.; Cole, D.; Vinjé, J.; Constantini, V.; Goyal, S.; Gramer, M.; Mackie, R.; Meng, X.J.; Myers, G.; et al. Fate and transport of zoonotic, bacterial, viral, and parasitic pathogens during swine manure treatment, storage, and land application. J. Anim. Sci. 2010, 88, 84. [Google Scholar] [CrossRef] [PubMed]
- De Busser, E.V.; De Zutter, L.; Dewulf, J.; Houf, K.; Maes, D. Salmonella control in live pigs and at slaughter. Vet. J. 2013, 196, 20–27. [Google Scholar] [CrossRef]
- Arguello, H.; Álvarez-Ordoñez, A.; Carvajal, A.; Rubio, P.; Prieto, M. Role of Slaughtering in Salmonella Spreading and Control in Pork Production. J. Food Prot. 2013, 76, 899–911. [Google Scholar] [CrossRef]
- FSIS Guideline to Control Salmonella in Swine Slaughter and Pork Processing Establishments. June 2023. FSIS-GD-2023-0003. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/documents/FSIS-GD-2023-0003.pdf (accessed on 9 October 2023).
- Analysis of the baseline survey on the prevalence of Salmonella in holdings with breeding pigs in the EU, 2008—Part A: Salmonella prevalence estimates. EFSA J. 2009, 7, 93. [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar] [CrossRef]
- Salmonella in Animals and Feed in Great Britain. 2022. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1120012/salmonella-animals-feed-gb-2021-v.2__003_.pdf (accessed on 20 October 2022).
- Centers for Disease Control and Prevention (CDC). National Center for Emerging and Zoonotic Infectious Diseases. Salmonella Serotypes Isolated from Animals and Related Sources. Division of Foodborne, Waterborne, and Environmental Diseases. Available online: https://www.cdc.gov/nationalsurveillance/pdfs/salmonella-serotypes-isolated-animals-and-related-sources-508.pdf (accessed on 15 June 2022).
- Hurd, H.S.; McKean, J.D.; Griffith, R.D.; Rostagno, M.H. Estimation of the Salmonella enterica prevalence in finishing swine. Epidemiol. Infect. 2004, 132, 127–135. [Google Scholar] [CrossRef]
- Visscher, C.F.; Klein, G.; Verspohl, J.; Beyerbach, M.; Stratmann-Selke, J.; Kamphues, J. Serodiversity and serological as well as cultural distribution of Salmonella on farms and in abattoirs in Lower Saxony, Germany. Int. J. Food Microbiol. 2011, 146, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Hurd, H.S.; McKean, J.D.; Griffith, R.W.; Wesley, I.V.; Rostagno, M.H. Salmonella enterica Infections in Market Swine with and without Transport and Holding. Appl. Env. Microbiol. 2002, 68, 2376–2381. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, D.M.; Suárez, M.C. Salmonella spp. in the pork supply chain: A risk approach. Rev. Colomb. Cienc. Pecu. 2014, 27, 65–75. [Google Scholar]
- Bonardi, S.; Alpigiani, I.; Bruini, I.; Barilli, E.; Brindani, F.; Morganti, M.; Cavallini, P.; Bolzoni, L.; Pongolini, S. Detection of Salmonella enterica in pigs at slaughter and comparison with human isolates in Italy. Int. J. Food Microbiol. 2016, 218, 44–50. [Google Scholar] [CrossRef]
- Bolton, D.J.; Ivory, C.; McDowell, D. A study of Salmonella in pigs from birth to carcass: Serotypes, genotypes, antibiotic resistance and virulence profiles. Int. J. Food Microbiol. 2013, 160, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Mannion, C.; Fanning, J.; McLernon, J.; Lendrum, L.; Gutierrez, M.; Duggan, S.; Egan, J. The role of transport, lairage and slaughter processes in the dissemination of Salmonella spp. in pigs in Ireland. Food Res. Int. 2012, 45, 871–879. [Google Scholar] [CrossRef]
- Berends, B.R.; Van Knapen, F.; Snijders, J.M.; Mossel, D.A. Identification and quantification of risk factors regarding Salmonella spp. on pork carcasses. Int. J. Food Microbiol. 1997, 36, 199–206. [Google Scholar] [CrossRef]
- Wilson, C.N.; Pulford, C.V.; Akoko, J.; Perez Sepulveda, B.; Predeus, A.V.; Bevington, J.; Duncan, P.; Hall, N.; Wigley, P.; Feasey, N.; et al. Salmonella identified in pigs in Kenya and Malawi reveals the potential for zoonotic transmission in emerging pork markets. PLoS Negl. Trop. Dis. 2020, 14, e0008796. [Google Scholar] [CrossRef]
- Deane, A.; Murphy, D.; Leonard, F.C.; Byrne, W.; Clegg, T.; Madigan, G.; Griffin, M.; Egan, J.; Prendergast, D.M. Prevalence of Salmonella spp. in slaughter pigs and carcasses in Irish abattoirs and their antimicrobial resistance. Ir. Vet. J. 2022, 75, 4. [Google Scholar] [CrossRef] [PubMed]
- Report of the Task Force on Zoonoses Data Collection on the Analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, Part B. EFSA J. 2008, 206, 1–111. [CrossRef]
- Bonardi, S.; Blagojevic, B.; Belluco, S.; Roasto, M.; Gomes-Neves, E.; Vågsholm, I. Food chain information in the European pork industry: Where are we? Trends Food Sci.Technol. 2021, 118, 833–839. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 6971. [Google Scholar] [CrossRef]
- Martelli, F.; Oastler, C.; Barker, A.; Jackson, G.; Smith, R.P.; Davies, R. Abattoir-based study of Salmonella prevalence in pigs at slaughter in Great Britain. Epidemiol. Infect. 2021, 149, E218. [Google Scholar] [CrossRef]
- NARMS Now: Integrated Data. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/narms-now-integrated-data (accessed on 28 August 2023).
- Bearson, S.M.D. Salmonella in Swine: Prevalence, Multidrug Resistance, and Vaccination Strategies. Annu. Rev. Anim. Biosci. 2022, 10, 373–393. [Google Scholar] [CrossRef]
- Davies, P.R. Intensive Swine Production and Pork Safety. Foodborne Pathog. Dis. 2011, 8, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Lan, R.; Zhang, X.; Cui, S.; Xu, J.; Guo, Y.; Li, F.; Zhang, D. Prevalence of Salmonella Isolates from Chicken and Pig Slaughterhouses and Emergence of Ciprofloxacin and Cefotaxime Co-Resistant S. enterica Serovar Indiana in Henan, China. PLoS ONE 2015, 10, e0144532. [Google Scholar] [CrossRef] [PubMed]
- Jiu, Y.; Meng, X.; Hong, X.; Huang, Q.; Wang, C.; Chen, Z.; Zhao, L.; Liu, X.; Lu, Y.; Li, S. Prevalence and Characterization of Salmonella in Three Typical Commercial Pig Abattoirs in Wuhan, China. Foodborne Pathog. Dis. 2020, 17, 62–627. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Surveillance for Foodborne Disease Outbreaks, United States, 2015, Annual Report. Atlanta, Georgia: US Department of Health and Human Services, CDC. 2017. Available online: https://www.cdc.gov/foodsafety/pdfs/2015foodborneoutbreaks_508.pdf (accessed on 10 August 2022).
- Centers for Disease Control and Prevention (CDC). National Salmonella Surveillance Annual Report, 2016. Atlanta, Georgia: US Department of Health and Human Services, CDC. 2018. Available online: https://www.cdc.gov/nationalsurveillance/pdfs/2016-Salmonella-report-508.pdf (accessed on 10 August 2022).
- Qian, J.; Wu, Z.; Zhu, Y.; Liu, C. One Health: A holistic approach for food safety in livestock. Sci. One Health 2022, 1, 100015. [Google Scholar] [CrossRef]
- Mossong, J.; Marques, P.; Ragimbeau, C.; Huberty-Krau, P.; Losch, S.; Meyer, G.; Moris, G.; Strottner, C.; Rabsch, W.; Schneider, F. Outbreaks of monophasic Salmonella enterica serovar 4,[5],12:i:- in Luxembourg, 2006. Eurosurveillance 2007, 12, 719. [Google Scholar] [CrossRef]
- Bone, A.; Noel, H.; Le Hello, S.; Pihier, N.; Danan, C.; Raguenaud, M.E.; Salah, S.; Bellali, H.; Vaillant, V.; Weill, F.X. Nationwide outbreak of Salmonella enterica serotype 4,12:i:-infections in France, linked to dried pork sausage, March-May 2010. Eurosurveillance 2010, 24, 19592. [Google Scholar] [CrossRef]
- Lettini, A.A.; Saccardin, C.; Ramon, E.; Longo, A.; Cortini, E.; Dalla Pozza, M.C.; Barco, L.; Guerra, B.; Luzzi, I.; Ricci, A. Characterization of an unusual Salmonella phage type DT7a and report of a foodborne outbreak of salmonellosis. Int. J. Food Microbiol. 2014, 189, 11–17. [Google Scholar] [CrossRef]
- Gossner, C.M.; Cauteren, D.V.; Hello, S.L.; Weill, F.X.; Terrien, E.; Tessier, S.; Janin, C.; Brisabois, A.; Dusch, V.; Vaillant, V.; et al. Nationwide outbreak of Salmonella enterica serotype 4,[5],12:i:- infection associated with consumption of dried pork sausage, France, November to December 2011. Eurosurveillance 2012, 17, 20071. [Google Scholar] [CrossRef]
- Arnedo-Pena, A.; Sabater-Vidal, S.; Herrera-León, S.; Bellido-Blasco, J.B.; Silvestre-Silvestre, E.; Meseguer-Ferrer, N.; Yague-Muñoz, A.; Gil-Fortuño, M.; Romeu-García, A.; Moreno-Muñoz, R. An outbreak of monophasic and biphasic Salmonella Typhimurium, and Salmonella Derby associated with the consumption of dried pork sausage in Castellon (Spain). Enferm. Infecc. Microbiol. Clin. 2015, 34, 544–550. [Google Scholar] [CrossRef]
- Morganti, M.; Bolzoni, L.; Scaltriti, E.; Casadei, G.; Carra, E.; Rossi, L.; Gherardi, P.; Faccini, F.; Arrigoni, N.; Sacchi, A.R.; et al. Rise and fall of outbreak-specific clone inside endemic pulsotype of Salmonella 4,[5],12:i:-; insights from high-resolution molecular surveillance in Emilia-Romagna, Italy, 2012 to 2015. Eurosurveillance 2017, 23, 17–00375. [Google Scholar] [CrossRef]
- Alt, K.; Simon, S.; Helmeke, C.; Kohlstock, C.; Prager, R.; Tietze, E.; Rabsch, W.; Karagiannis, I.; Werber, D.; Frank, C.; et al. Outbreak of uncommon O4 non-agglutinating Salmonella Typhimurium linked to minced pork, Saxony-Anhalt, Germany, January to April 2013. PLoS ONE. 2015, 10, e0128349. [Google Scholar] [CrossRef]
- Multistate Outbreak of Multidrug-Resistant Salmonella I 4,[5],12:i:- and Salmonella Infantis Infections Linked to Pork (Final Update). Available online: https://www.cdc.gov/salmonella/pork-08-15/index.html (accessed on 14 October 2022).
- Mandilara, G.; Sideroglou, T.; Chrysostomou, A.; Rentifis, I.; Papadopoulos, T.; Polemis, M.; Tzani, M.; Tryfinopoulou, K.; Mellou, K. The Rising Burden of Salmonellosis Caused by Monophasic Salmonella Typhimurium (1,4,[5],12:i:-) in Greece and New Food Vehicles. Antibiotics 2021, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- UK Health Security Agency. Non-typhoidal Salmonella data 2009 to 2018. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1026149/salmonella-annual-report-2018.pdf (accessed on 9 September 2022).
- Helmuth, I.G.; Espenhain, L.; Ethelberg, S.; Jensen, T.; Kjeldgaard, J.; Litrup, E.; Schjørring, S.; Müller, L. An outbreak of monophasic Salmonella Typhimurium associated with raw pork sausage and other pork products, Denmark 2018–19. Epidemiol Infect 2019, 147, E315. [Google Scholar] [CrossRef] [PubMed]
- Pardos de la Gandara, M.; Fournet, N.; Bonifait, L.; Lefèvre, S.; Chemaly, M.; Grastilleur, C.; Cadel-Six, S.; Fach, P.; Pignault, A.; Brisabois, A.; et al. Countrywide multi-serotype outbreak of Salmonella Bovismorbificans ST142 and monophasic Salmonella Typhimurium ST34 associated with dried pork sausages in France, September 2020 to January 2021. Eurosurveillance 2023, 28, 2200123. [Google Scholar] [CrossRef]
- CDC: Salmonella Outbreak Linked to Salami Sticks. Available online: https://www.cdc.gov/salmonella/i45-10-21/index.html (accessed on 14 October 2022).
- Luzzi, I.; Galetta, P.; Massari, M.; Rizzo, C.; Dionisi, A.M.; Filetici, E.; Cawthorne, A.; Tozzi, A.; Argentieri, M.; Bilei, S.; et al. An Easter outbreak of Salmonella Typhimurium DT 104A associated with traditional pork salami in Italy. Eurosurveillance 2007, 12, 11–12. [Google Scholar] [CrossRef]
- Torpdahl, M.; Sorensen, G.; Ethelberg, S.; Sandø, G.; Gammelgård, K.; Porsbo, L.J. A regional outbreak of S. Typhimurium in Denmark and identification of the source using MLVA typing. Eurosurveillance 2006, 11, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Bruun, T.; Sørensen, G.; Forshell, L.P.; Jensen, T.; Nygard, K.; Kapperud, G.; Lindstedt, B.A.; Berglund, T.; Wingstrand, A.; Petersen, R.F. An outbreak of Salmonella Typhimurium infections in Denmark, Norway and Sweden, 2008. Eurosurveillance 2009, 14, 19147. [Google Scholar] [CrossRef]
- Ethelberg, S.; Wingstrand, A.; Jensen, T.; Sørensen, G.; Muller, L.; Lisby, M.; Nielsen, E.M.; Mølbak, K. Large outbreaks of Salmonella Typhimurium infection in Denmark in 2008. Eurosurveillance 2008, 13, 19023. [Google Scholar] [CrossRef]
- Andreoli, G.; Merla, C.; Valle, C.D.; Corpus, F.; Morganti, M.; D’Incau, M.; Colmegna, S.; Marone, P.; Fabbi, M.; Barco, L.; et al. Foodborne Salmonellosis in Italy: Characterization of Salmonella enterica Serovar Typhimurium and Monophasic Variant 4,[5],12:i− Isolated from Salami and Human Patients. J. Food Prot. 2017, 80, 632–639. [Google Scholar] [CrossRef]
- Kuhn, K.G.; Torpdahl, M.; Frank, C.; Sigsgaard, K.; Ethelberg, S. An outbreak of Salmonella Typhimurium traced back to salami, Denmark, April to June 2010. Eurosurveillance 2011, 16, 19863. [Google Scholar] [CrossRef]
- Kuhn, K.G.; Sørensen, G.; Torpdahl, M.; Kjeldsen, M.K.; Jensen, T.; Gubbels, S.; Bjerager, G.O.; Wingstrand, A.; Porsbo, L.J.; Ethelberg, S. A long-lasting outbreak of Salmonella Typhimurium U323 associated with several pork products, Denmark, 2010. Epidemiol. Infect. 2013, 141, 260–268. [Google Scholar] [CrossRef]
- Wójcik, O.P.; Kjelsø, C.; Kuhn, K.G.; Müller, L.; Jensen, T.; Kjeldsen, M.K.; Ethelberg, S. Salmonella Typhimurium outbreak associated with smoked pork tenderloin in Denmark, January to March 2011. Scand. J. Infect. Dis. 2012, 44, 903–908. [Google Scholar] [CrossRef]
- Paranthaman, K.; Haroon, S.; Latif, S.; Vinnyey, N.; de Souza, V.; Welfare, W.; Tahir, M.; Cooke, E.; Stone, K.; Lane, C.; et al. Emergence of a Multidrug-Resistant (ASSuTTm) Strain of Salmonella enterica Serovar Typhimurium DT120 in England in 2011 and the Use of Multiple-Locus Variable-Number Tandem-Repeat Analysis in Supporting Outbreak Investigations. Foodborne Pathog. Dis. 2013, 10, 850–855. [Google Scholar] [CrossRef]
- CDC: Salmonella Outbreaks Linked to Italian-Style Meats. Available online: https://www.cdc.gov/salmonella/italian-style-meat-08-21/details.html (accessed on 14 October 2022).
- Uelze, L.; Becker, N.; Borowiak, M.; Busch, U.; Dangel, A.; Deneke, C.; Fischer, J.; Flieger, A.; Hepner, S.; Huber, I.; et al. Toward an Integrated Genome-Based Surveillance of Salmonella enterica in Germany. Front. Microbiol. 2021, 12, 626941. [Google Scholar] [CrossRef]
- Niesters, H.G.; Rossen, J.W.; van der Avoort, H.; Baas, D.; Benschop, K.; Claas, E.C.; Kroneman, A.; van Maarseveen, N.; Pas, S.; van Pelt, W.; et al. Laboratory-based surveillance in the molecular era: The TYPENED model, a joint data-sharing platform for clinical and public health laboratories. Eurosurveillance 2013, 18, 20387. [Google Scholar] [CrossRef]
- Ford, L.; Miller, M.; Cawthorne, A.; Fearnley, E.; Kirk, M. Approaches to the Surveillance of Foodborne Disease: A Review of the Evidence. Foodborne Pathog. Dis. 2015, 12, 927–936. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Foodborne Disease Outbreaks: Guidelines for Investigation and Control. World Health Organization. 2008. Available online: https://apps.who.int/iris/handle/10665/43771 (accessed on 20 February 2023).
- Deng, X.; Ran, L.; Wu, S.; Ke, B.; He, D.; Yang, X.; Zhang, Y.; Ke, C.; Klena, J.D.; Yan, M.; et al. Laboratory-Based Surveillance of Non-typhoidal Salmonella Infections in Guangdong Province, China. Foodborne Pathog. Dis. 2012, 9, 35–312. [Google Scholar] [CrossRef] [PubMed]
- Chui, L.; Ferrato, C.; Li, V.; Christianson, S. Comparison of Molecular and In Silico Salmonella Serotyping for Salmonella Surveillance. Microorganisms 2021, 9, 955. [Google Scholar] [CrossRef]
- Deng, X.; den Bakker, H.C.; Hendriksen, R.S. Genomic Epidemiology: Whole-Genome-Sequencing-Powered Surveillance and Outbreak Investigation of Foodborne Bacterial Pathogens. Annu. Rev. Food Sci. Technol. 2016, 7, 353–374. [Google Scholar] [CrossRef] [PubMed]
- Moran-Gilad, J. Whole genome sequencing (WGS) for food-borne pathogen surveillance and control—Taking the pulse. Eurosurveillance 2017, 22, 30547. [Google Scholar] [CrossRef]
- den Bakker, H.C.; Allard, M.W.; Bopp, D.; Brown, E.W.; Fontana, J.; Iqbal, Z.; Kinney, A.; Limberger, R.; Musser, K.A.; Shudt, M.; et al. Rapid Whole-Genome Sequencing for Surveillance of Salmonella enterica Serovar Enteritidis. Emerg. Infect. Dis. 2014, 20, 1306–1314. [Google Scholar] [CrossRef]
- Expert Opinion on the Introduction of Next-Generation Typing Methods for Food- and Waterborne Diseases in the EU and EEA. Available online: https://www.ecdc.europa.eu/sites/default/files/media/en/publications/Publications/food-and-waterborne-diseases-next-generation-typing-methods.pdf (accessed on 11 October 2023).
- Quainoo, S.; Coolen, J.P.M.; Hijum, S.A.F.T.V.; Huynen, M.A.; Melchers, W.J.G.; Schaik, W.V.; Wertheim, H.F.L. Whole-Genome Sequencing of Bacterial Pathogens: The Future of Nosocomial Outbreak Analysis. Clin. Microbiol. Rev. 2017, 30, 1015–1063. [Google Scholar] [CrossRef] [PubMed]
- Atxaerandio-Landa, A.; Arrieta-Gisasola, A.; Laorden, L.; Bikandi, J.; Garaizar, J.; Martinez-Malaxetxebarria, I.; Martinez-Ballesteros, I. A Practical Bioinformatics Workflow for Routine Analysis of Bacterial WGS Data. Microorganisms 2022, 10, 2364. [Google Scholar] [CrossRef]
- Turcotte, M.R.; Smith, J.T.; Li, J.; Zhang, X.; Wolfe, K.L.; Gao, F.; Benton, C.S.; Andam, C.P. Genome characteristics of clinical Salmonella enterica population from a state public health laboratory, New Hampshire, USA, 2017–2020. BMC Genom. 2022, 23, 537. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Expert Opinion on Whole Genome Sequencing for Public Health Surveillance; ECDC: Stockholm, Sweden, 2016; Available online: https://data.europa.eu/doi/10.2900/12442 (accessed on 8 November 2022).
- Robertson, J.; Yoshida, C.; Kruczkiewicz, P.; Nadon, C.; Nichani, A.; Taboada, E.N.; Nash, J.H.E. Comprehensive assessment of the quality of Salmonella whole genome sequence data available in public sequence databases using the Salmonella in silico Typing Resource (SISTR). Microb. Genom. 2018, 4, e000151. [Google Scholar] [CrossRef] [PubMed]
- Nadon, C.; Van Walle, I.; Gerner-Smidt, P.; Campos, J.; Chinen, I.; Concepcion-Acevedo, J.; Gilpin, B.; Smith, A.M.; Kam, K.M.; Perez, E.; et al. PulseNet International: Vision for the implementation of whole genome sequencing (WGS) for global food-borne disease surveillance. Eurosurveillance 2017, 22, 30544. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Whole genome sequencing for foodborne disease surveillance: Landscape paper. World Health Organization. Licence: CC BY-NC-SA 3.0 IGO. 2018. Available online: https://apps.who.int/iris/handle/10665/272430 (accessed on 8 November 2022).
- Prioritizing Zoonotic Diseases for Multisectoral One Health Collaboration in Colombia. Available online: https://www.cdc.gov/onehealth/pdfs/colombia-508.pdf (accessed on 10 October 2023).
Figure 1.
(a) Pig affected by salmonellosis (S. Choleraesuis). Cyanosis, with obvious red-blue discoloration of the ears. (b) Pig affected by salmonellosis (S. Choleraesuis). Severe gastritis and ulcers are observed in the gastric mucosa. (c) Pig affected by salmonellosis (S. Choleraesuis). Severe colitis with particularly reddened colon mucosa.
Figure 1.
(a) Pig affected by salmonellosis (S. Choleraesuis). Cyanosis, with obvious red-blue discoloration of the ears. (b) Pig affected by salmonellosis (S. Choleraesuis). Severe gastritis and ulcers are observed in the gastric mucosa. (c) Pig affected by salmonellosis (S. Choleraesuis). Severe colitis with particularly reddened colon mucosa.

Figure 2.
Pig affected by salmonellosis (S. Typhimurium). Necrotic enterotyphlocolitis with diphtheritic membrane on the mucosal surface.
Figure 2.
Pig affected by salmonellosis (S. Typhimurium). Necrotic enterotyphlocolitis with diphtheritic membrane on the mucosal surface.

Table 1.
Prevalence of S. 1,4,[5],12:i:- and of S. Typhimurium in serotyped Salmonella isolates from pigs in 2017–2019 and in 2021 in the EU.
Table 1.
Prevalence of S. 1,4,[5],12:i:- and of S. Typhimurium in serotyped Salmonella isolates from pigs in 2017–2019 and in 2021 in the EU.
| Year | S. 1,4,[5],12:i:- | S. Typhimurium | Reference |
|---|---|---|---|
| 2017 | 37.4% | 20.6% | [133] |
| 2018 | 25.7% | 12.4% | [134] |
| 2019 | 28.8% | 12.7% | [135] |
| 2021 | 28.2% | 15.3% | [13] |
Data not available for 2020.
Table 2.
Pigs’ carcasses testing positive for Salmonella (before chilling and after dressing) in the EU from 2017 to 2021; data coming from CAs and FBOp.
Table 2.
Pigs’ carcasses testing positive for Salmonella (before chilling and after dressing) in the EU from 2017 to 2021; data coming from CAs and FBOp.
| Year | CAs | FBOp | Reference |
|---|---|---|---|
| 2017 | 2.15% | 1.85% | [133] |
| 2018 | 2.69% | 1.57% | [134] |
| 2019 | 3.15% | 1.51% | [135] |
| 2020 | 3.60% | 1.70% | [150] |
| 2021 | 1.70% | 1.40% | [13] |
Table 3.
Percentages of the most reported Salmonella serovars out of serotyped isolates from pig meat in the EU from 2017 to 2019.
Table 3.
Percentages of the most reported Salmonella serovars out of serotyped isolates from pig meat in the EU from 2017 to 2019.
| Year | S. 1,4,[5],12:i:- | S. Typhimurium | S. Derby | Reference |
|---|---|---|---|---|
| 2017 | 22.0% | 27.0% | NS* | [133] |
| 2018 | 12.4% | 13.9% | 20.6% | [134] |
| 2019 | 26.6% | 14.0% | 21.3% | [135] |
NS*, not specified.
Table 4.
Salmonellosis-confirmed human cases in the EU per year from 2017 to 2021, together with the prevalence of S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby associated with human cases.
Table 4.
Salmonellosis-confirmed human cases in the EU per year from 2017 to 2021, together with the prevalence of S. Typhimurium, S. 1,4,[5],12:i:-, and S. Derby associated with human cases.
| Year | Human Cases | S. Typhimurium | S. 1,4,[5],12:i:- | S. Derby | References |
|---|---|---|---|---|---|
| 2017 | 91,662 | 13.4% | 8.0% | 0.8% | [133] |
| 2018 | 91,858 | 13.0% | 8.1% | 0.9% | [134] |
| 2019 | 87,923 | 11.9% | 8.2% | 0.9% | [135] |
| 2020 | 52,702 | 12.4% | 11.1% | 1.2% | [150] |
| 2021 | 60,050 | 11.4% | 8.8% | 0.9% | [13] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Soliani, L.; Rugna, G.; Prosperi, A.; Chiapponi, C.; Luppi, A. Salmonella Infection in Pigs: Disease, Prevalence, and a Link between Swine and Human Health. Pathogens 2023, 12, 1267. https://doi.org/10.3390/pathogens12101267
AMA Style
Soliani L, Rugna G, Prosperi A, Chiapponi C, Luppi A. Salmonella Infection in Pigs: Disease, Prevalence, and a Link between Swine and Human Health. Pathogens. 2023; 12(10):1267. https://doi.org/10.3390/pathogens12101267
Chicago/Turabian StyleSoliani, Laura, Gianluca Rugna, Alice Prosperi, Chiara Chiapponi, and Andrea Luppi. 2023. "Salmonella Infection in Pigs: Disease, Prevalence, and a Link between Swine and Human Health" Pathogens 12, no. 10: 1267. https://doi.org/10.3390/pathogens12101267
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.