
Are We Ready to Get Rid of the Terms “Chalimus” and ”Preadult” in the Caligid (Crustacea: Copepoda: Caligidae) Life Cycle Nomenclature?


Abstract
:1. Background of This Review
2. Materials and Methods
3. Early Studies on Caligid Ontogeny
4. Advanced Research on Caligid Life Cycles
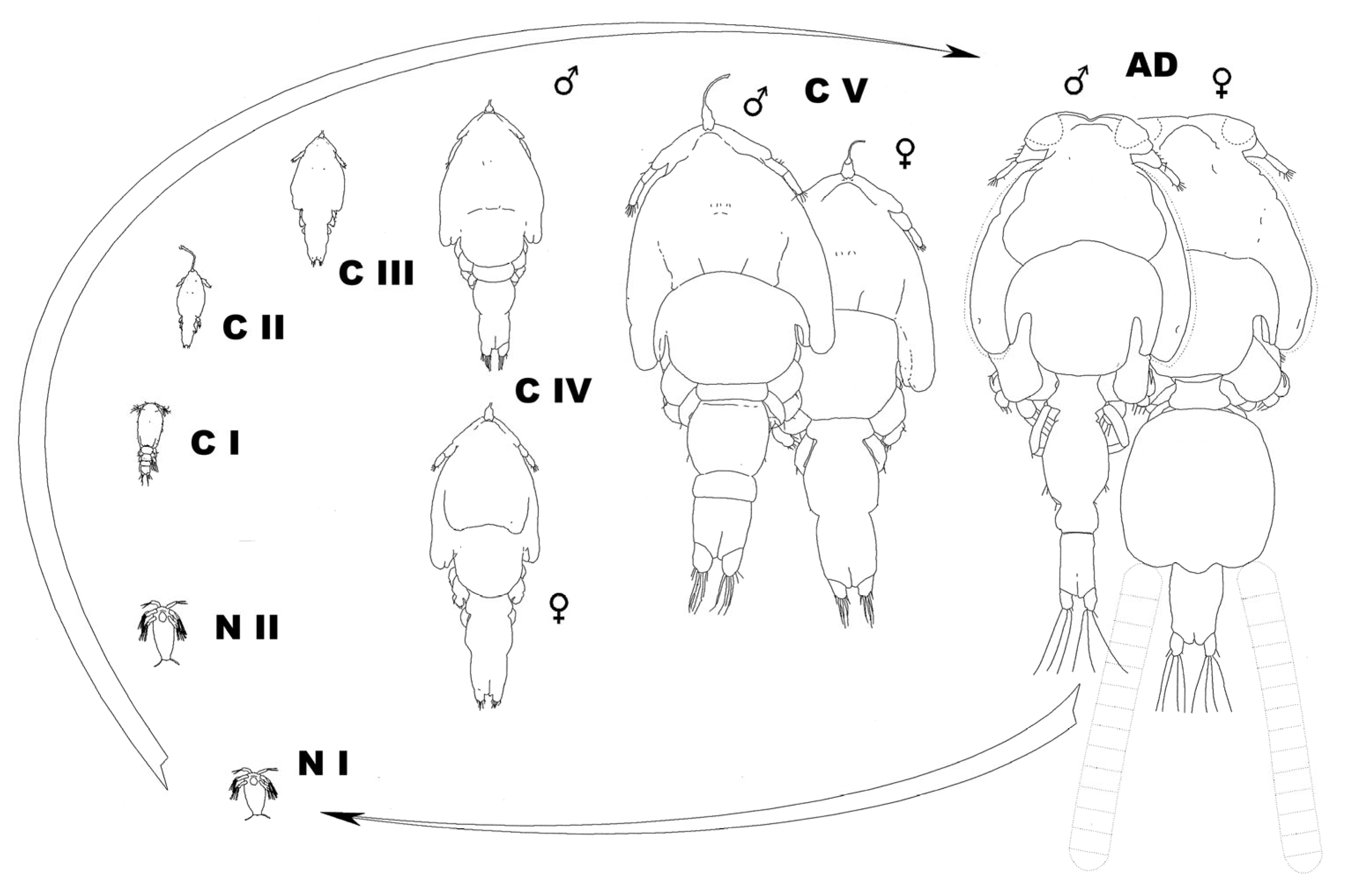
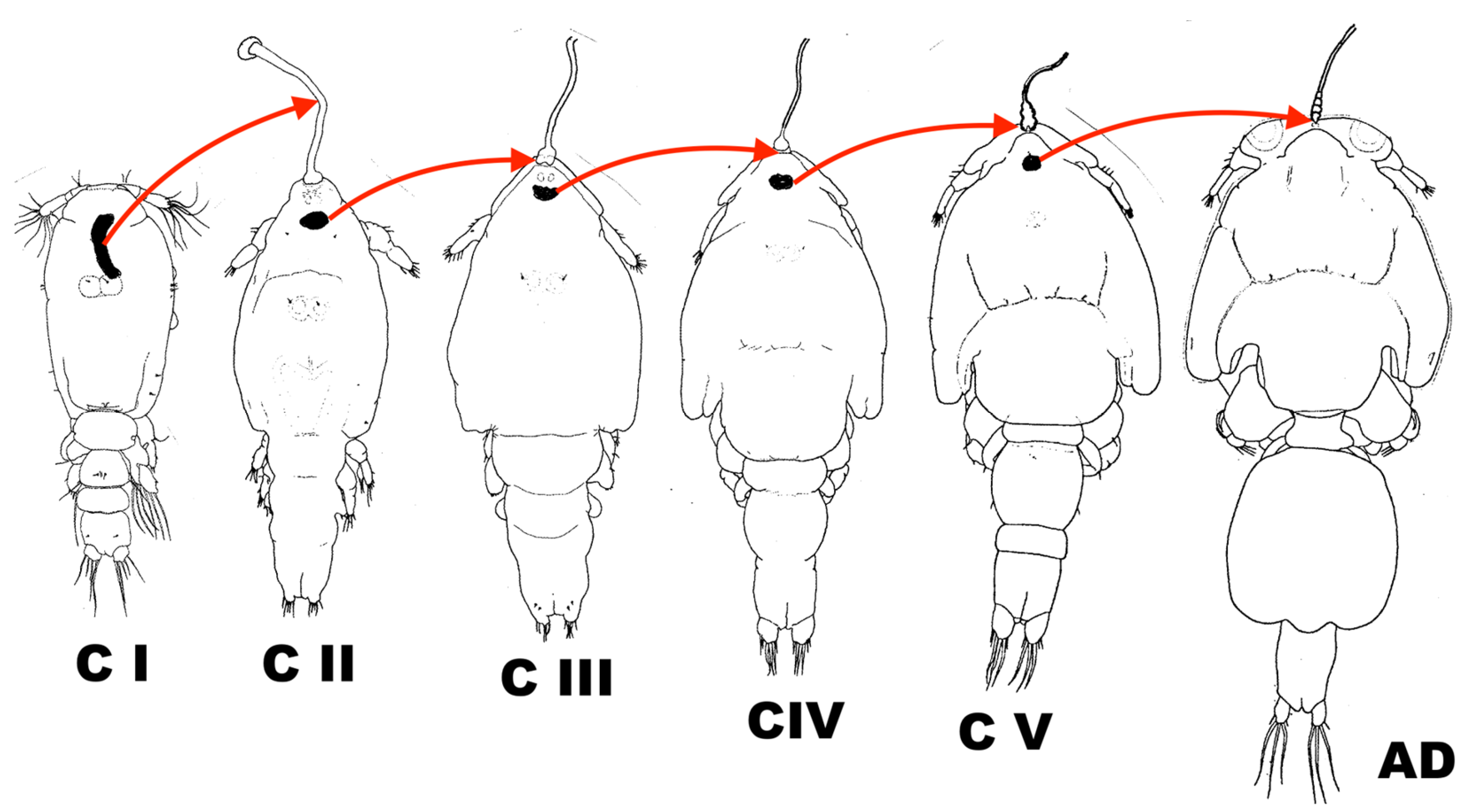
5. Reports on Individual Developmental Stages
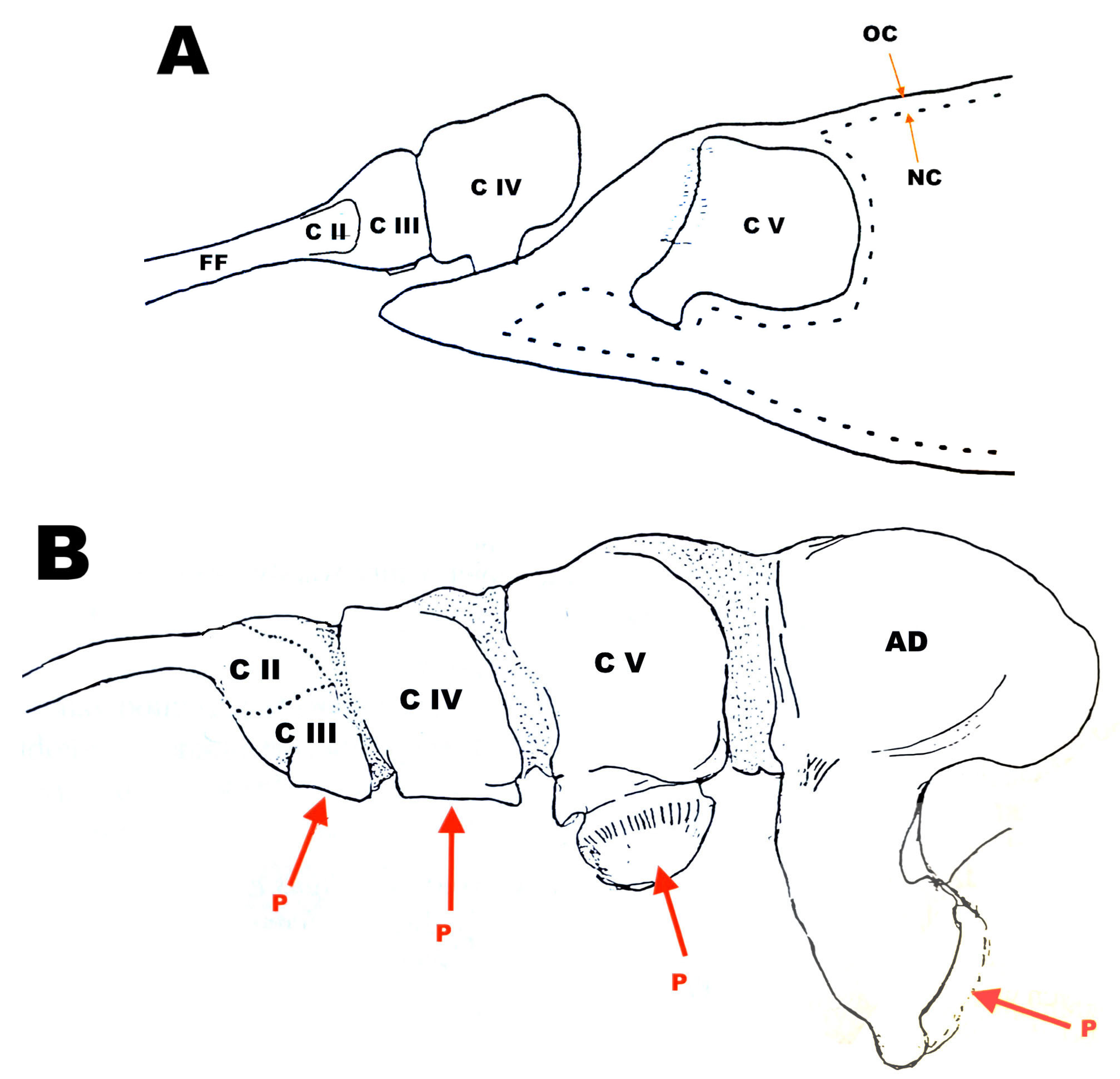
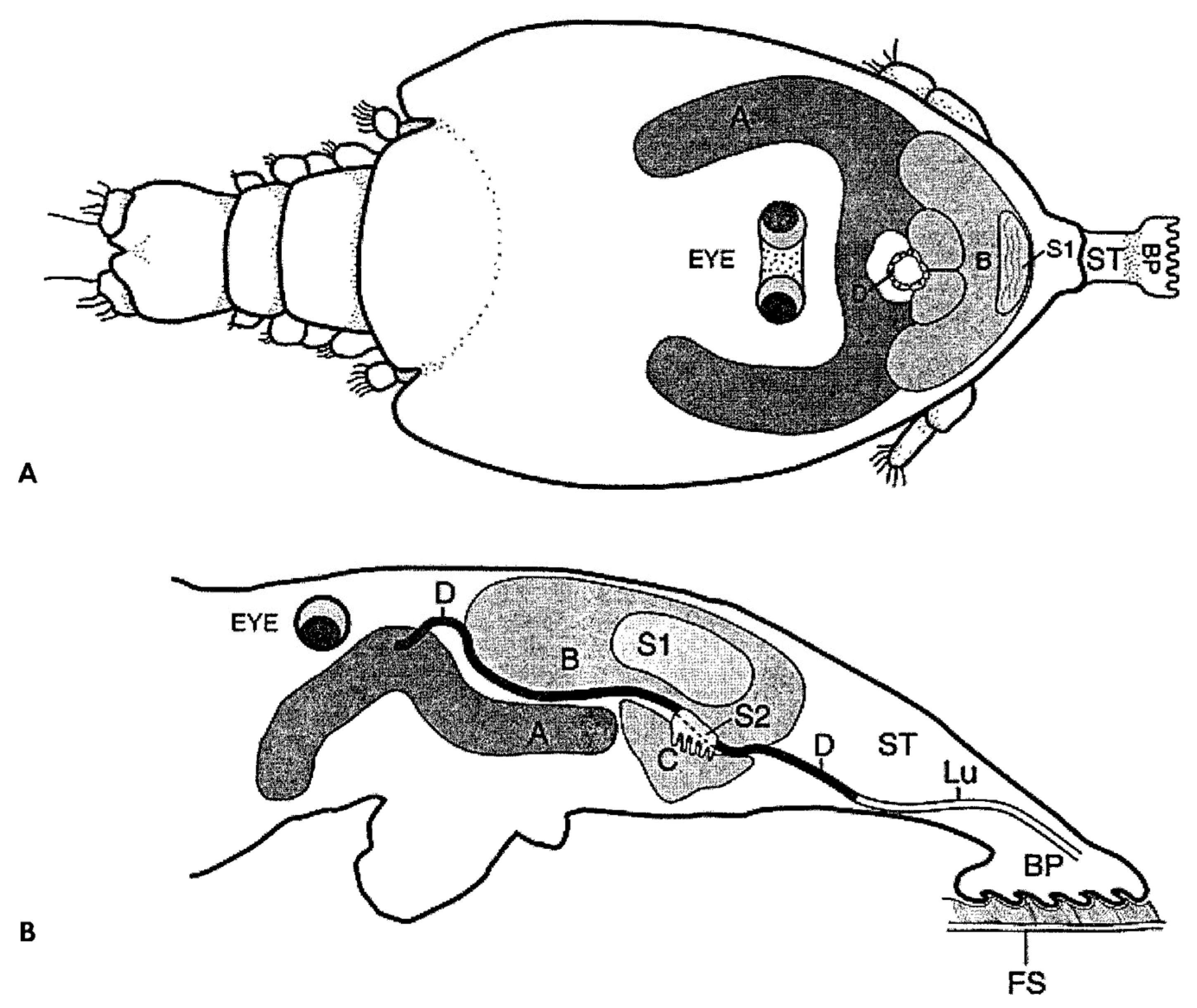
6. Frontal Filament in Species of Caligus
7. Frontal Filament in Species of Lepeophtheirus
8. “Copepodid Attached by the Frontal Filament”?
- ○
- The copepodid finds a suitable place on the surface of the host and firmly attaches there using the claws of the antennae and the maxillipeds;
- ○
- Through intensive scraping by some appendages it removes a portion of the host’s epithelium, thereby exposing a suitable place for permanent attachment;
- ○
- Molting starts with a rupture of the copepod’s larval cuticle in the frontal area;
- ○
- The cuticular pocket containing fully formed frontal filament, previously visible within the cephalothorax, evaginates to become a frontal extremity;
- ○
- The filament is released and its proximal end becomes firmly attached to the tip of the frontal extremity;
- ○
- The distal end of the filament is glued to the previously exposed part of the host’s body;
- ○
- After the hardening of the filament has assured that the attachment to the host is secure, the organism continues its molting process;
- ○
- The next stage—copepodid II (chalimus 1)—withdraws from the copepodid I exuvium and hangs from the host, attached only by the frontal filament. The exuvium itself is discarded.
9. The Concept of ”Preadult”
10. Recapitulation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WoRMS Editorial Board. World Register of Marine Species. 2022. at VLIZ (Vlaams Instituut voor de Zee = Flanders Marine Institute, Oostende, Belgium). Available online: https://www.marinespecies.org (accessed on 29 November 2022).
- Piasecki, W.; Avenant-Oldewage, A. Diseases caused by Crustacea. In Fish Diseases; Eiras, J., Segner, H., Wahli, T., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2008; pp. 1115–1200. [Google Scholar]
- Froese, R.; Pauly, D. (Eds.) FishBase. Version 08/2022. 2022. Available online: https://www.fishbase.org (accessed on 29 November 2022).
- Kabata, Z. Parasitic Copepoda of British Fishes; No. 152; Ray Society: London, UK, 1979. [Google Scholar]
- Dudley, P.L. Aspects of general body shape and development in Copepoda. Syllogeus 1986, 58, 7–25. [Google Scholar]
- Huys, R.; Boxshall, G.A. (Eds.) Copepod Evolution; No. 159; Ray Society: London, UK, 1991. [Google Scholar]
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; No. 166; Ray Society: London, UK, 2004. [Google Scholar]
- Kabata, Z. Copepoda (Crustacea) parasitic on fishes: Problems and perspectives. Adv. Parasitol. 1981, 19, 1–71. [Google Scholar]
- Anstensrud, M. Moulting and mating in Lepeophtheirus pectoralis (Copepoda, Caligidae). J. Mar. Biol. Assoc. UK 1990, 70, 269–281. [Google Scholar] [CrossRef]
- Ferrari, F.D.; Dahms, H.U. Post-Embryonic Development of the Copepoda; Crustacean Monographs 8; Brill: Leiden, The Netherlands, 2007. [Google Scholar]
- Venmathi Maran, B.A.; Moon, S.Y.; Ohtsuka, S.; Soh, H.Y.; Myoung, J.G.; Oh, S.-Y.; Iglikowska, A.; Boxshall, G.A. The caligid life cycle: New evidence from Lepeophtheirus elegans reconciles the cycles of Caligus and Lepeophtheirus. Parasite 2013, 20, e15. [Google Scholar] [CrossRef] [Green Version]
- Hamre, L.A.; Eichner, C.; Caipang, C.M.A.; Dalvin, S.T.; Bron, J.E. The salmon louse Lepeophtheirus salmonis (Copepoda: Caligidae) life cycle has only two chalimus stages. PLoS ONE 2013, 8, e73539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmeister, H. Beschreibung einiger neuen oder weniger bekannten Schmarotzerkrebse: Nebst allgemeinen Betrachtungen über die Gruppe, welcher sie angehören. Acad. Caes. Leop. Nova Acta. 1835, 17, 269–336. [Google Scholar] [CrossRef]
- Krøyer, H. Om Snyltekrebsene, isaer med Hensyn til den danske Fauna. Naturhist. Tidsskr. 1837, 2, 172–208. [Google Scholar]
- Milne-Edwards, H. Histoire Naturelle des Crustacés: Comprenant L’anatomie, la Physiologie et la Classification de ces Animaux; Tome Troisième; Librairie Encyclopedique de Roret: Paris, France, 1840. [Google Scholar] [CrossRef]
- Goodsir, H.D.S. On a new genus and six new species of Crustacea with observations on the development of the egg, and on the metamorphosis of Caligus, Carcinus and Pagurus. Edinb. New Phil. J. 1842, 33, 174–192. [Google Scholar]
- Baird, W. The Natural History of British Entomostraca; No. 17; Ray Society: London, UK, 1850. [Google Scholar] [CrossRef]
- Müller, F. Eine Beobachtung über die Beziehung der Gattungen Caligus und Chalimus. Arch. Naturgesch. 1852, 18, 91–92. [Google Scholar]
- Hesse, E. Mémoire sur les moyens à l’aide desquels certaines Crustacés parasites assurent la conservation de leurs espèces. Ann. Sci. Nat. Zool. Botan. Anatom. Phys. 1858, 4, 120–125. [Google Scholar]
- Von Nordmann, A. Neue Beiträge zur Kenntnis parasitischer Copepoden. Bull. Soc. Imp. Natur. Moscou 1864, 37, 461–520. [Google Scholar]
- Gadd, P. Parasit-Copepoder i Finland. Master’s Thesis, University of Helsinki, Helsinki, Finland, 1901. [Google Scholar]
- Scott, A., VI. Lepeophtheirus and Lernaea; LMBC Memoirs: London, UK, 1901. [Google Scholar]
- Wilson, C.B. North American parasitic copepods belonging to the family Caligidae—Part I. The Caliginae. Proc. U. S. Natl. Mus. 1905, 28, 479–672. [Google Scholar] [CrossRef]
- Gurney, R. British Fresh-Water Copepoda; No. 3; Ray Society: London, UK, 1933. [Google Scholar]
- Russell, F.S. LXX.—A new species of Caligus from Egypt, Caligus pageti, sp. n. Ann. Mag. Nat. Hist. 1925, 9, 611–618. [Google Scholar] [CrossRef]
- Argilas, A. Un Copépode parasite de Mugil auratus Risso, nouveau pour l’Algerie: Caligus pageti Russel. Bull. Trav. Sta. Aquic. Alger. 1931, 2, 93–106. [Google Scholar]
- Gurney, R. The development of certain parasitic Copepoda of the families Caligidae and Clavellidae. Proc. Zool. Soc. Lond. 1934, 105, 177–217. [Google Scholar] [CrossRef]
- White, H.C. Life history of Lepeophtheirus salmonis. J. Fish. Res. Board Can. 1942, 6, 24–29. [Google Scholar] [CrossRef]
- Heegaard, P. Contribution to the phylogeny of the arthropods; Copepoda. Spolia Zool. Mus. Haun. 1947, 8, 1–227. [Google Scholar]
- Kabata, Z. Developmental stages of Caligus clemensi (Copepoda: Caligidae). J. Fish. Res. Board. Can. 1972, 29, 1571–1593. [Google Scholar] [CrossRef]
- Kozikowska, Z. Skorupiaki pasożytnicze (Crustacea parasitica) Polski. Część I. Pasożyty ryb wód ujściowych Odry. Zool. Polon. 1957, 8, 217–270. [Google Scholar]
- Lewis, A.G. Life history of the caligid copepod Lepeophtheirus dissimulatus Wilson, 1905 (Crustacea: Caligoida). Pac. Sci. 1963, 17, 195–242. [Google Scholar]
- Ohtsuka, S.; Takami, I.; Venmathi Maran, B.A.; Ogawa, K.; Shimono, T.; Fujita, Y.; Asakawa, M.; Boxshall, G.A. Developmental stages and growth of Pseudocaligus fugu Yamaguti, 1936 (Copepoda: Siphonostomatoida: Caligidae) host-specific to puffer. J. Nat. Hist. 2009, 43, 1779–1804. [Google Scholar] [CrossRef]
- Hwa, T.-K. Studies on the life history of a fish louse. Acta Zool. Sin. 1965, 17, 48–57. (In Chinese) [Google Scholar]
- Izawa, K. Life history of Caligus spinosus Yamaguti, 1939 obtained from cultured yellow tail, Seriola quinqueradiata T. & S. (Crustacea: Caligoida). Rep. Fac. Fish. Prefect. Univ. Mie 1969, 9, 127–157. [Google Scholar]
- Voth, D.R. Life history of the caligid copepod Lepeophtheirus hospitalis Fraser, 1920 (Crustacea: Caligoida). Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1972. [Google Scholar]
- Boxshall, G.A. The developmental stages of Lepeophtheirus pectoralis (Müller, 1776) (Copepoda: Caligidae). J. Nat. Hist. 1974, 8, 681–700. [Google Scholar] [CrossRef]
- Caillet, C. Biologie Comparée de Caligus minimus Otto, 1848 et de Clavellodes Macrotrachelus (Brian, 1906), Copépods Parasites de Poissons Marins. Ph.D. Thesis, Université de Montpellier II (Université des Sciences et Techniques du Languedoc), Montpellier, France, 1979. [Google Scholar]
- Wootten, R.; Smith, J.W.; Needham, E.A. Aspects of the biology of the parasitic copepods Lepeophtheirus salmonis and Caligus elongatus on farmed salmonids, and their treatment. Proc. R. Soc. B 1982, 81, 185–197. [Google Scholar] [CrossRef]
- Ben Hassine, O.K. Les Copépodes Parasites de Poissons Mugilidae en Méditerranée Occidentale (Côtes Françaises et Tunisiennes); Morphologie, Bio-Écologie, Cycles Évolutifs. Ph.D. Thesis, Université de Montpellier II (Université des Sciences et Techniques du Languedoc), Montpellier, France, 1983. [Google Scholar]
- Costello, M.J. The global economic cost of sea lice to the salmonid farming industry. J. Fish Dis. 2009, 32, 115–118. [Google Scholar] [CrossRef]
- Bravo, S.; Erranz, F.; Silva, M.T. Comparison under controlled conditions of the life cycle of Lepeophtheirus mugiloidis and Caligus rogercresseyi, parasites of the Patagonian blenny Eleginops maclovinus. Aquac. Res. 2021, 52, 4198–4204. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Defaye, D. (Eds.) Pathogens of Wild and Farmed Fish: Sea Lice; Ellis Horwood: Chichester, UK, 1993. [Google Scholar]
- Hogans, W.E.; Trudeau, D.J. Preliminary studies on the biology of sea lice, Caligus elongatus, Caligus curtus and Lepeophtheirus salmonis (Copepoda: Caligoida) parasitic on cage-cultured salmonids in the Lower Bay of Fundy. Can. Tech. Rep. Fish. Aquat. Sci. 1989, 1715, 14. [Google Scholar]
- Piasecki, W.; MacKinnon, B.M. Changes in structure of the frontal filament in sequential developmental stages of Caligus elongatus von Nordmann, 1832 (Crustacea, Copepoda, Siphonostomatoida). Can. J. Zool. 1993, 71, 889–895. [Google Scholar] [CrossRef]
- Ohtsuka, S.; Nawata, M.; Nishida, Y.; Nitta, M.; Hirano, K.; Adachi, K.; Kondo, Y.; Venmathi Maran, B.A.; Suárez-Morales, E. Discovery of the fish host of the ‘planktonic’ caligid Caligus undulatus Shen & Li, 1959 (Crustacea: Copepoda: Siphonostomatoida). Biodivers. Data. J. 2020, 8, e52271. [Google Scholar] [CrossRef]
- Schram, T.A. 1990. Morten Anstensrud; 30 December 1957–27 March 1990. J. Crustac. Biol. 1990, 10, 751–752. [Google Scholar]
- Bron, J.; Sommerville, C.; Jones, M.; Rae, G.H. The settlement and attachment of early stages of the salmon louse, Lepeophtheirus salmonis (Copepoda: Caligidae) on the salmon host, Salmo salar. J. Zool. 1991, 224, 201–212. [Google Scholar] [CrossRef]
- Johnson, S.C.; Albright, L.J. The developmental stages of Lepeophtheirus salmonis (Krøyer, 1837) (Copepoda: Caligidae). Can. J. Zool. 1991, 69, 929–950. [Google Scholar] [CrossRef]
- Johnson, S.C.; Albright, L.J. Development, growth, and survival of Lepeophtheirus salmonis (Copepoda: Caligidae) under laboratory conditions. J. Mar. Biol. Assoc. UK 1991, 71, 425–436. [Google Scholar] [CrossRef]
- Ogawa, K. Caligus longipedis infection of cultured striped jack, Pseudocaranx dentex (Teleostei: Carangidae) in Japan. Gyobyo Kenkyu 1992, 27, 191–205. [Google Scholar] [CrossRef]
- Lin, C.-L.; Ho, J.-S. Life history of Caligus epidemicus Hewitt parasitic on tilapia (Oreochromis mossambicus) cultured in brackish water. In Pathogens of Wild and Farmed Fish: Sea Lice; Boxshall, G.A., Defaye, D., Eds.; Ellis Horwood: Chichester, UK, 1993; pp. 5–15. [Google Scholar]
- Kim, I.-H. Developmental stages of Caligus punctatus Shiino, 1955 (Copepoda: Caligidae). In Pathogens of Wild and Farmed Fish: Sea Lice; Boxshall, G.A., Defaye, D., Eds.; Ellis Horwood: Chichester, UK, 1993; pp. 16–29. [Google Scholar]
- Schram, T.A. Supplementary descriptions of the developmental stages of Lepeophtheirus salmonis (Krøyer, 1837) (Copepoda: Caligidae). In Pathogens of Wild and Farmed Fish: Sea Lice; Boxshall, G.A., Defaye, D., Eds.; Ellis Horwood: Chichester, UK, 1993; pp. 30–47. [Google Scholar]
- Pike, A.W.; Mordue (Luntz), A.J.; Ritchie, G. The development of Caligus elongatus Nordmann from hatching to copepodid in relation to temperature. In Pathogens of Wild and Farmed Fish: Sea Lice; Boxshall, G.A., Defaye, D., Eds.; Ellis Horwood: Chichester, UK, 1993; pp. 51–60. [Google Scholar]
- Pike, A.W.; MacKenzie, K.; Rowand, A. Ultrastructure of the frontal filament in chalimus larvae of Caligus elongatus and Lepeophtheirus salmonis from Atlantic salmon, Salmo salar. In Pathogens of Wild and Farmed Fish: Sea Lice; Boxshall, G.A., Defaye, D., Eds.; Ellis Horwood: Chichester, UK, 1993; pp. 99–113. [Google Scholar]
- Lin, C.-L.; Ho, J.-S.; Chen, S.-N. Developmental stages of Caligus epidemicus Hewitt, a copepod parasite of tilapia cultured in brackish water. J. Nat. His. 1996, 30, 661–684. [Google Scholar] [CrossRef]
- Øines, Ø.; Heuch, P.A. Identification of sea louse species of the genus Caligus using mtDNA. J. Mar. Biol. Assoc. UK 2005, 85, 73–79. [Google Scholar] [CrossRef]
- Øines, Ø.; Heuch, P.A. Caligus elongatus Nordmann genotypes on wild and farmed fish. J. Fish Dis. 2007, 30, 81–91. [Google Scholar] [CrossRef]
- Øines, Ø.; Simonsen, J.H.; Knutsen, J.A.; Heuch, P.A. Host preference of Caligus elongatus Nordmann in the laboratory and its implications for Atlantic cod aquaculture. J. Fish Dis. 2006, 29, 167–174. [Google Scholar] [CrossRef]
- Øines, Ø.; Schram, T. Intra- or inter-specific difference in genotypes of Caligus elongatus Nordmann 1832. Acta Parasitol. 2008, 53, 93–105. [Google Scholar] [CrossRef]
- Piasecki, W.; MacKinnon, B.M. Life cycle of a sea louse, Caligus elongatus von Nordmann, 1832 (Copepoda, Siphonostomatoida, Caligidae). Can. J. Zool. 1995, 73, 74–82. [Google Scholar] [CrossRef]
- Piasecki, W. The developmental stages of Caligus elongatus von Nordmann, 1832 (Copepoda: Caligidae). Can. J. Zool. 1996, 74, 1459–1478. [Google Scholar] [CrossRef]
- Piasecki, W. Cykl Rozwojowy Caligus elongatus von Nordmann, 1832 (Crustacea, Coepoda, Siphonostomatoida); Rozprawy No. 165; Akademia Rolnicza w Szczecinie: Szczecin, Poland, 1995; 69p. [Google Scholar]
- Lin, C.-L.; Ho, J.-S.; Chen, S.N. Development of Caligus multispinosus Shen, a caligid copepod parasitic on the black sea bream (Acanthopagrus schlegeli) cultured in Taiwan. J. Nat. Hist. 1997, 31, 1483–1500. [Google Scholar] [CrossRef]
- Ho, J.-S.; Lin, C.-L.; Chen, S.-N. Species of Caligus Müller, 1785 (Copepoda: Caligidae) parasitic on marine fishes of Taiwan. Syst. Parasitol. 2000, 46, 159–179. [Google Scholar] [CrossRef]
- Gonzalez-Alanis, P.; Wright, G.M.; Johnson, S.C.; Burka, J.F. Frontal filament morphogenesis in the salmon louse Lepeophtheirus salmonis. J. Parasitol. 2001, 87, 561–574. [Google Scholar] [CrossRef] [Green Version]
- González, L.; Carvajal, J. Life cycle of Caligus rogercresseyi, (Copepoda: Caligidae) parasite of Chilean reared salmonids. Aquaculture 2003, 220, 101–117. [Google Scholar] [CrossRef]
- Ho, J.-S.; Lin, C.-L. Sea Lice of Taiwan (Copepoda: Siphonostomatoida: Caligidae); Sueichan Press: Keelung, Taiwan, 2004. [Google Scholar]
- Madinabeitia, I.; Nagasawa, K. Chalimus stages of Caligus latigenitalis (Copepoda: Caligidae) parasitic on blackhead seabream from Japanese waters, with discussion of terminology used for developmental stages of caligids. J. Parasitol. 2011, 97, 221–236. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Terms Used in Bionomenclature: The Naming of Organisms (and Plant Communities). Global Biodiversity Information Facility, Copenhagen Publication. 2010. Available online: http:/www.gbif.org/document/80577 (accessed on 29 November 2022).
- Eichner, C.; Hamre, L.A.; Nilsen, F. Instar growth and molt increments in Lepeophtheirus salmonis (Copepoda: Caligidae) chalimus larvae. Parasitol. Int. 2015, 64, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Khoa, T.N.D.; Mazelan, S.; Muda, S.; Shaharom-Harrison, F. The life cycle of Caligus minimus on seabass (Lates calcarifer) from floating cage culture. Thalassas 2019, 35, 77–85. [Google Scholar] [CrossRef]
- Hamre, L.A.; Bui, S.; Oppedal, F.; Skern-Mauritzen, R.; Dalvin, S. Development of the salmon louse Lepeophtheirus salmonis parasitic stages in temperatures ranging from 3 to 24 °C. Aquac. Environ. Interact. 2019, 11, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Hemmingsen, W.; MacKenzie, K.; Sagerup, K.; Remen, M.; Bloch-Hansen, K.; Imsland, A.K.D. Caligus elongatus and other sea lice of the genus Caligus as parasites of farmed salmonids: A review. Aquaculture 2020, 522, e735160. [Google Scholar] [CrossRef]
- Jeong, J.; McEwan, G.F.; Arriagada, G.; Gallardo-Escárate, C.; Revie, C.W. Quantifying key parameters related to the life cycle of Caligus rogercresseyi. J. Fish Dis. 2022, 45, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Markewitsch, A.P. Les Crustacés parasites des poissons de la Mer Caspiènne. Bull. l’Inst. Océanograph. Monaco 1933, 638, 1–27. [Google Scholar]
- Bere, R. Parasitic copepods from Gulf of Mexico fish. Amer. Midl. Nat. 1936, 17, 577–625. [Google Scholar] [CrossRef]
- Gnanamuthu, C.P. Sex differences in the chalimus and adult forms of Caligus polycanthi sp. nov. (Crustacea: Copepoda) parasitic on Balistes maculatus from Madras. Rec. Indian Mus. 1950, 47, 159–170. [Google Scholar] [CrossRef]
- Kaj, J. Caligus lacustris Stp. et Ltk. Materiały do znajomości pasożytniczych widłonogów Polski. Pol. Arch. Hydrobiol. 1953, 1, 45–48. [Google Scholar]
- Hewitt, G.C. Two species of Caligus (Copepoda, Caligidae) from Australian waters, with a description of some developmental stages. Pac. Sci. 1971, 25, 145–164. [Google Scholar]
- Lopez, G. Redescription and ontogeny of Lepeophtheirus kareii Yamaguti, 1936 (Copepoda, Caligoida). Crustaceana 1976, 31, 203–207. [Google Scholar] [CrossRef]
- Johannessen, A. early stages of Lepeophtheirus salmonis (Copepoda, Caligidae). Sarsia 1978, 63, 169–176. [Google Scholar] [CrossRef]
- Izawa, K. The copepodid and two chalimus stages of Caligus latigenitalis Shiino, 1954 (Copepoda, Siphonostomatoida, Caligidae), parasitic on Japanese black sea bream, Acanthopagrus schlegeli. Contrib. Biol. Lab. Kyoto Univ. 2004, 29, 329–341. [Google Scholar]
- Jones, J.B. A redescription of Caligus patulus Wilson, 1937 (Copepoda: Caligidae) from a fish farm in the Philippines. Syst. Parasitol. 2004, 2, 103–116. [Google Scholar] [CrossRef]
- Montory, J.A.; Cumillaf, J.P.; Cubillos, V.M.; Paschke, K.; Urbina, M.A.; Gebauer, P. Early development of the ectoparasite Caligus rogercresseyi under combined salinity and temperature gradients. Aquaculture 2018, 486, 68–74. [Google Scholar] [CrossRef]
- Tully, O. The succession of generations and growth of the caligid copepods Caligus elongatus and Lepeophtheirus salmonis parasitizing farmed Atlantic salmon smolts (Salmo salar L.). J. Mar. Biol. Assoc. UK 1989, 69, 279–287. [Google Scholar] [CrossRef]
- Boxshall, G.A. Lepeophtheirus pectoralis (O. F. Müller, 1776): A description, a review and some comparisons with the genus Caligus Müller, 1785. J. Nat. Hist. 1974, 8, 445–468. [Google Scholar] [CrossRef]
- Jónsdóttir, H.; Bron, J.E.; Wootten, R.; Turnbull, J.F. The histopathology associated with the pre-adult and adult stages of Lepeophtheirus salmonis on the Atlantic salmon, Salmo salar L. J. Fish Dis. 1992, 15, 521–527. [Google Scholar] [CrossRef]
- Johnson, S.C. Comparison of development and growth rates of Lepeophtheirus salmonis(Copepoda: Caligidae) on naive Atlantic (Salmo salar) and Chinook (Oncorhynchus tshawytscha) salmon. In Pathogens of Wild and Farmed Fish: Sea Lice; Boxshall, G.A., Defaye, D., Eds.; Ellis Horwood: Chichester, UK, 1993; pp. 68–142. [Google Scholar]
- Bjørn, P.; Finstad, B. The development of salmon lice (Lepeophtheirus salmonis) on artificially infected post smolts of sea trout (Salmo trutta). Can. J. Zool. 1998, 76, 970–977. [Google Scholar] [CrossRef]
- Pike, A.W.; Wadsworth, S. Sealice on salmonids: Their biology and control. Adv. Parasitol. 1999, 44, 233–337. [Google Scholar] [CrossRef]
- Schram, T.A. Practical identification of pelagic sea lice larvae. J. Mar. Biol. Assoc. UK 2004, 84, 103–110. [Google Scholar] [CrossRef]
- Beamish, R.; Neville, C.M.; Sweeting, R.M.; Jones, S.R.M.; Ambers, N.; Gordon, E.K.; Hunter, K.L.; McDonald, T.E. Proposed life history strategy for the salmon louse, Lepeophtheirus salmonis in the subarctic Pacific. Aquaculture 2007, 264, 428–440. [Google Scholar] [CrossRef]
- Pedaschenko, D.D. Èmbrional’noe razvitie i metamorfoz’’ Lernaea branchialis. Die Embryonalentwicklung und Metamorphose von Lernaea branchialis. Trudý Imp. St.-Petersbg. Obshch. Estest. Zoolog. 1898, 26, 1–307. [Google Scholar]
- Piasecki, W. Life cycle of Tracheliastes maculatus Kollar, 1835 (Copepoda, Siphonostomatoida, Lernaeopodidae). Wiad. Parazytol. 1989, 35, 187–245. [Google Scholar] [PubMed]
- Piasecki, W.; Kuźmińska, E. Developmental stages of Achtheres percarum (Crustacea: Copepoda), parasitic on European perch, Perca fluviatilis (Actinopterygii: Perciformes). Acta Ichthyol. Piscat. 2007, 37, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Huys, R.; Llewellyn-Hughes, J.; Conroy-Dalton, S.; Olson, P.D.; Spinks, J.N.; Johnston, D.A. Extraordinary host switching in siphonostomatoid copepods and the demise of the Monstrilloida: Integrating molecular data, ontogeny and antennulary morphology. Mol. Phylogenetics Evol. 2007, 43, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Kakui, K.; Munakata, M. A new Sphaeronella species (Copepoda: Siphonostomatoida: Nicothoidae) parasitic on Euphilomedes sp. (Ostracoda: Myodocopa: Philomedidae) from Hokkaido, Japan, with an 18S molecular phylogeny. Syst. Parasitol. 2023, 100, 121–131. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Copepodid I | Copepodid II (Ch 1) | Copepodid III (Ch 2) | Copepodid IV (Ch 3) | Copepodid V (Ch 4) | Adult | |
|---|---|---|---|---|---|---|
| Filament | 1-lobed | 1-lobed | 1-lobed * | 2-lobed | 3-lobed | 4-lobed |
| 3rd leg | Vestigial | Vestigial | Biramous | Biramous | Biramous | Biramous |
| 4th leg | Uniramous | Uniramous | Uniramous | Uniramous | ||
| 5th leg | Vestigial | Vestigial | Vestigial | |||
| Sternal furca | Vestigial | Present | ||||
| H-suture | Vestigial? | Present | ||||
| Lunulae | Present | |||||
| Marginal membrane | Absent | Absent | Absent | Absent | Absent | Present |
| Pinnate setae on swimming legs | Present | |||||
| Post-antennary process | Present |
| Features | Copepodid I | Copepodid II (=Chalimus 1) | Copepodid III (=Chalimus 2) | Copepodid IV (=Preadult 1) | Copepodid V (=Preadult 2) | Adult |
|---|---|---|---|---|---|---|
| Body | Highly pigmented, no frontal filament | No pigmentation; short frontal filament | No pigmentation; developed frontal filament | Well-developed frontal plates | Fully developed cephalothorax | Typical caligiform cephalothorax |
| Cephalothorax | About 1.5 times longer than free posterior somites | About 2.5 to 3 times longer than free posterior somites | About 3.5 times longer than free posterior somites | Typical H-shaped suture | About 2 times longer than free posterior somites | As in preceding stage |
| Antennule (2 segments) | Proximal: 3 setae Distal: 11 setae + 2 Aesthetascs (A) | Proximal: 7 Distal: 12 + 2A | Proximal: 13; Distal: 12 + 2A | Proximal: 20; Distal: 12 + 2A | Proximal: 27 Distal: 12 + 2A | Proximal: 27 Distal: 12 + 2A |
| Antenna | Three-segmented; second segment with rugose process; third segment with recurved claw | Modified from that of preceding copepodid stage; third segment with curved distal claw | Middle segment with rudiment of dorsal adhesion pad | Second segment with reniform adhesion pad; terminal claw strongly curved | As in preceding stage | Terminal part forming strong, recurved claw |
| Maxilliped | Distal segment separated by partial suture, carrying terminal claw and trifid setal element | Distal segment of subchela bearing curved claw and short inner seta | Segments comprising subchela more completely fused than in preceding stage | As in preceding stage | As in preceding stage | Distal subchela with trace of suture separating short apical claw |
| Sternal furca | Absent | Absent | Present | Broad box and divergent, slightly tapering tines | As in preceding stage | As in preceding stage |
| Reference | Species | Stages Originally Found/Determined | Stages Interpreted and Comments |
|---|---|---|---|
| Russel [25] | C. pageti | N, C1, C2, Ch (“three sizes”) | N I, C I, C II, [—], [—], C V, Ad |
| Argilas [26] | C. pageti | N, C, Ch | N I, C I, [—], [—], CIV, Ad |
| Gurney [24] | C. centrodonti | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, [—], C IV, C V, Ad |
| Heegaard [29] | C. curtus | N1, N2, C 1, C2, pupa, Ch1, Ch2, Ch3, Ch4, Ch5, Ad | N I, N II, C I, C II, [—], C IV(?), C V(?), Ad |
| Kozikowska [31] | C. lacustris | C, Ch1, Ch2, Ch3, Ch4, Ad | [N I?], [N II?], C I, C II, [—], C IV(?), C V(?), Ad |
| Hwa [34] | C. orientalis | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ch5, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Izawa [35] | C. spinosus | N1, N2, C, Ch1, Ch2, Ch3, pAd1, pAd2, Ad | N I, N II, C I, C II, C III, C IV, [--], Ad |
| Kabata [30] | C. clemensi | N1, N2, C, Ch1, Ch2, Ch3, Ch4, pAd1, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Caillet [38] | C. minimus | N1, N2, C, Ch1, Ch2, Ch3, Ch4 Ch5, Ad | N I, N II, C I, [—], C III, C IV, CV, Ad |
| Ben Hassine [40] | C. pageti | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, Ad | N I, N II, C I, C II, [—], C IV, CV, Ad |
| Hogans and Trudeau [44] | C. elongatus | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, Ad | N I, N II, C I, C II, [—], C IV, CV, Ad |
| Ogawa [51] | C. longipedis | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Kim [53] | C. punctatus | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Lin et al. [57] | C. epidemicus | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ch5, Ch6, pAd, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Piasecki [63] | C. elongatus | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Lin et al. [65] | C.rotundigenitalis | N1, N2, C, Ch1, Ch2, Ch3, Ch4, pAd, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| González and Carvajal [68] | C. rogercresseyi | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Ohtsuka et al. [33] | C. fugu | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Madinabeitia and Nagasawa [70] | C. latigenitalis | C, Ch1, Ch2, Ch3, Ch4, Ad | [N I], [N II], C I, C II, C III, C IV, CV, Ad |
| Khoa et al. [73] | C. minimus | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Bravo et al. [42] | C. rogercresseyi | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| White [28] | L. salmonis | C, Ch1, Ch2, […], Ch(final), […], Ad | C I, C II, [—], C IV(?), C V(?), Ad |
| Lewis [32] | L. dissimulatus | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ch5, Ch6, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Voth [36] | L. hospitalis | N1, N2, C, Ch1, Ch2, Ch3, Ch4, Ch5, Ch6, A | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Boxshall [37] | L. pectoralis | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, pAd2, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Johnson and Albright [49] | L. salmonis | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, pAd2, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Schram [54] | L. salmonis | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, pAd2, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Venmathi Maran et al. [11] | L. elegans | N1, N2, C, Ch1, Ch2, pA1, pA2, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Hamre et al. [12] | L. salmonis | N1, N2, C, Ch1, Ch2, pA1, pA2, Ad | N I, N II, C I, C II, C III, C IV, CV, Ad |
| Bravo et al. [42] | L. mugiloidis | N1, N2, C, Ch1, Ch2, Ch3, Ch4 pAd1, pAd2, Ad | No details available. Interpretation not possible |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piasecki, W.; Venmathi Maran, B.A.; Ohtsuka, S. Are We Ready to Get Rid of the Terms “Chalimus” and ”Preadult” in the Caligid (Crustacea: Copepoda: Caligidae) Life Cycle Nomenclature? Pathogens 2023, 12, 460. https://doi.org/10.3390/pathogens12030460
Piasecki W, Venmathi Maran BA, Ohtsuka S. Are We Ready to Get Rid of the Terms “Chalimus” and ”Preadult” in the Caligid (Crustacea: Copepoda: Caligidae) Life Cycle Nomenclature? Pathogens. 2023; 12(3):460. https://doi.org/10.3390/pathogens12030460
Chicago/Turabian StylePiasecki, Wojciech, Balu Alagar Venmathi Maran, and Susumu Ohtsuka. 2023. "Are We Ready to Get Rid of the Terms “Chalimus” and ”Preadult” in the Caligid (Crustacea: Copepoda: Caligidae) Life Cycle Nomenclature?" Pathogens 12, no. 3: 460. https://doi.org/10.3390/pathogens12030460