
Proline Dehydrogenase and Pyrroline 5 Carboxylate Dehydrogenase from Mycobacterium tuberculosis: Evidence for Substrate Channeling
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cloning, Expression, and Purification of PruB and PruA
2.3. Measurement of PruB Enzymatic Activity
2.4. Synthesis of DL-Pyrroline-5-Carboxylic Acid [(DL)-P5C]
2.5. Measurement of PruA Enzymatic Activity
2.6. Coupled PruB-PruA Reaction
2.7. P5C Trapping Assays
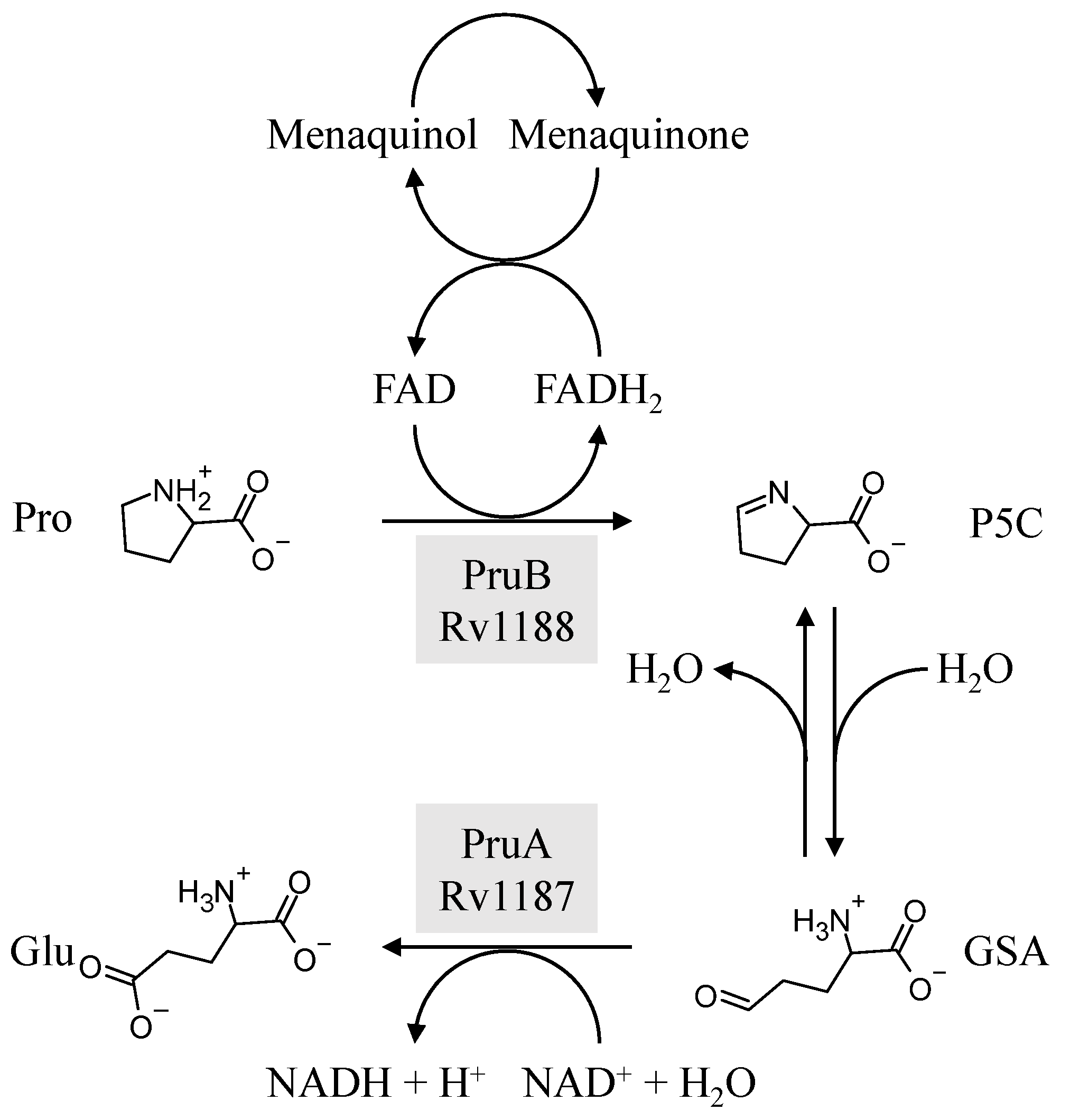
3. Results and Discussion
3.1. Expression and Purification of PruB and PruA
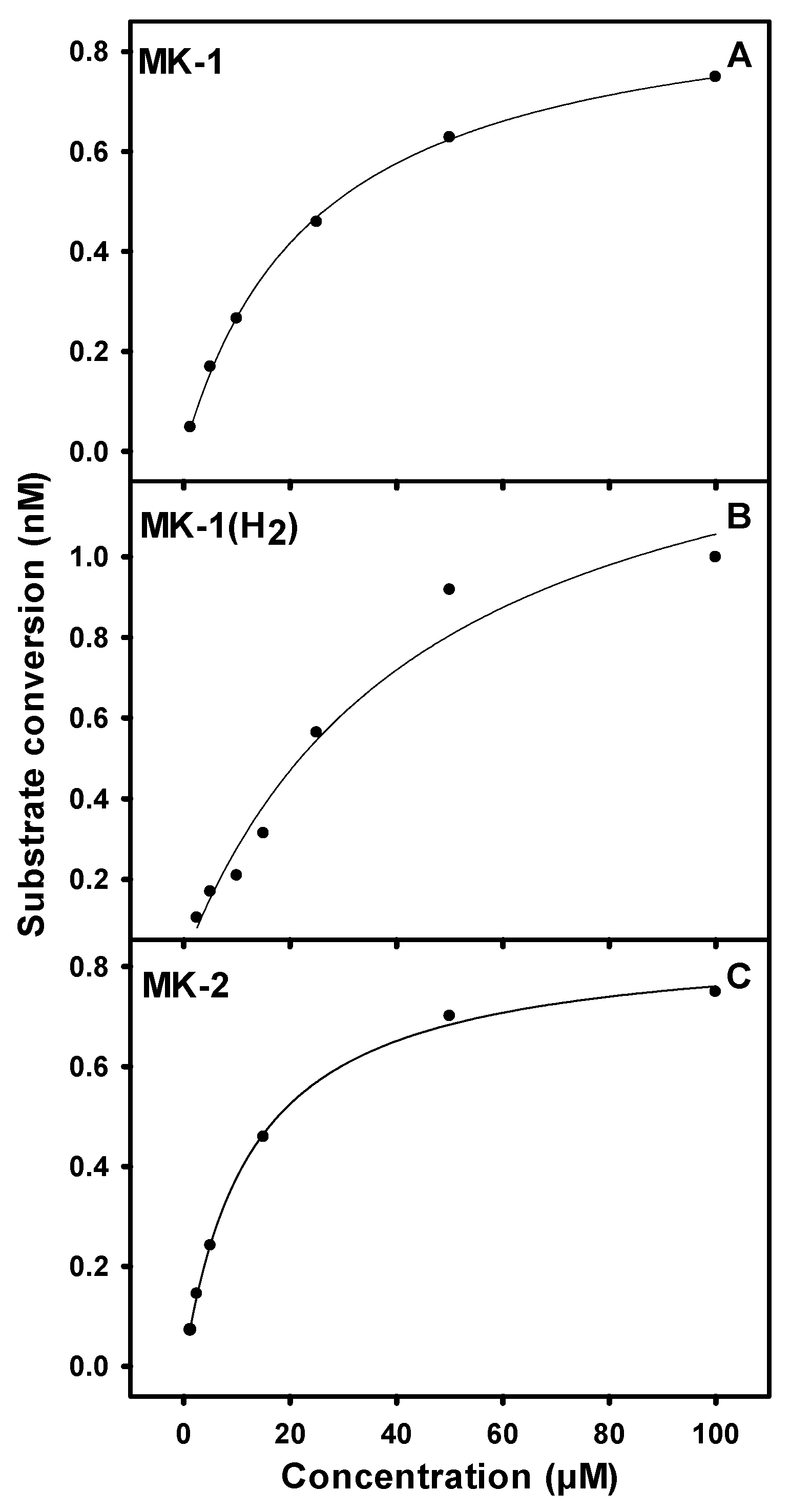
3.2. Biochemical Characterization of PruB
3.3. Kinetic Parameters for PruB
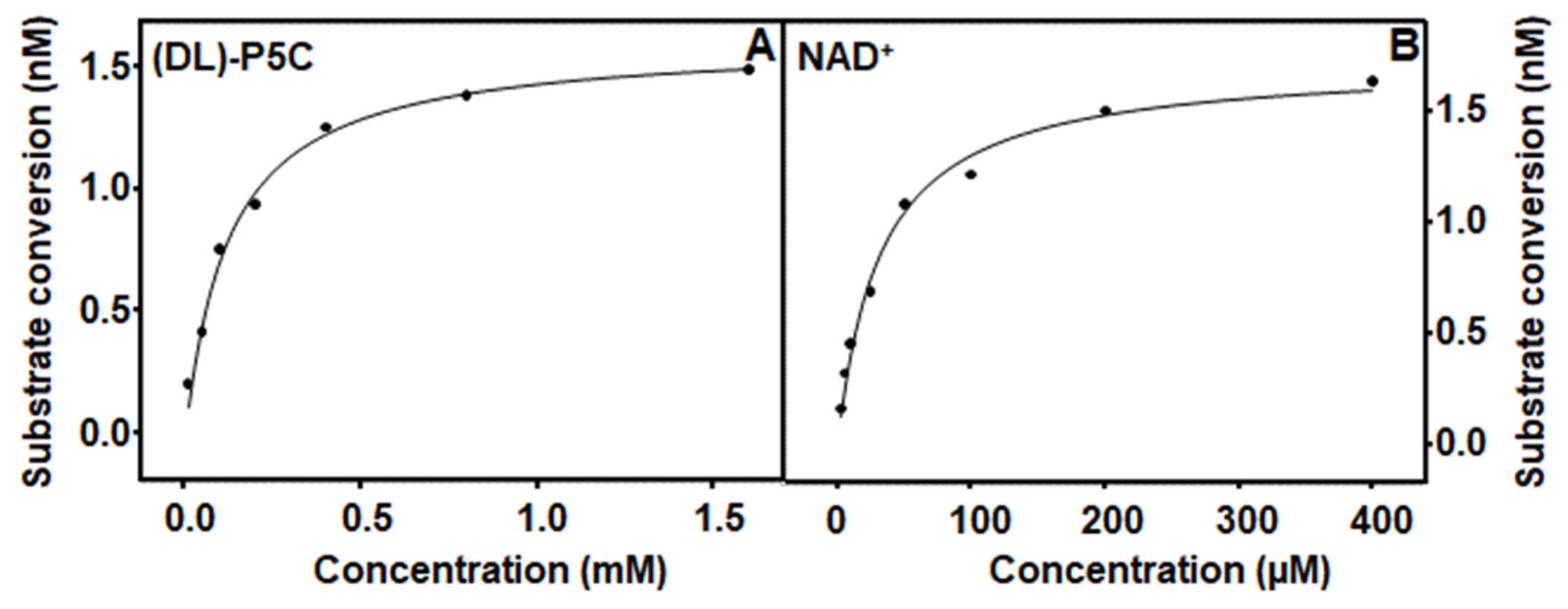
3.4. Characterization and Kinetic Parameters of PruA
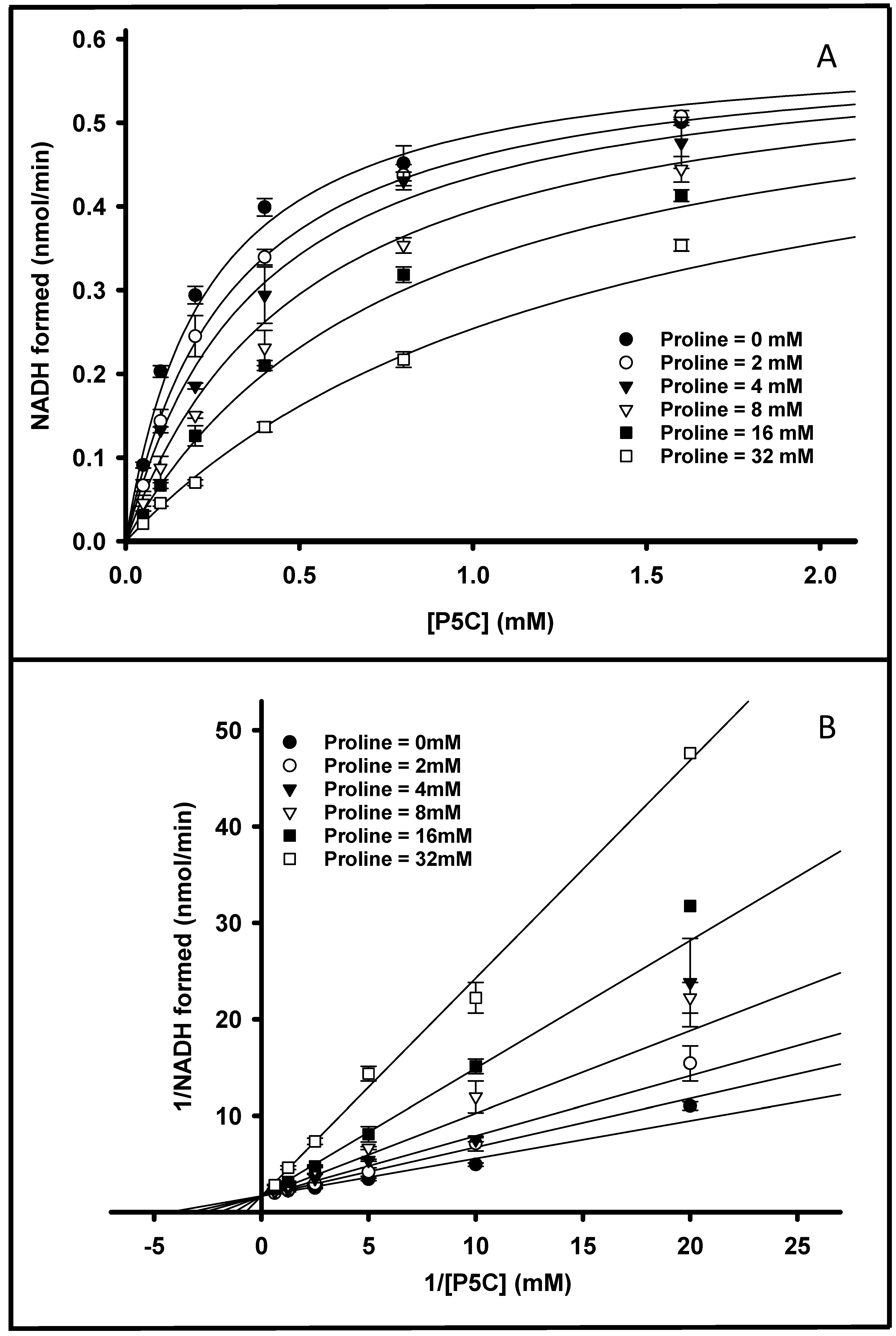
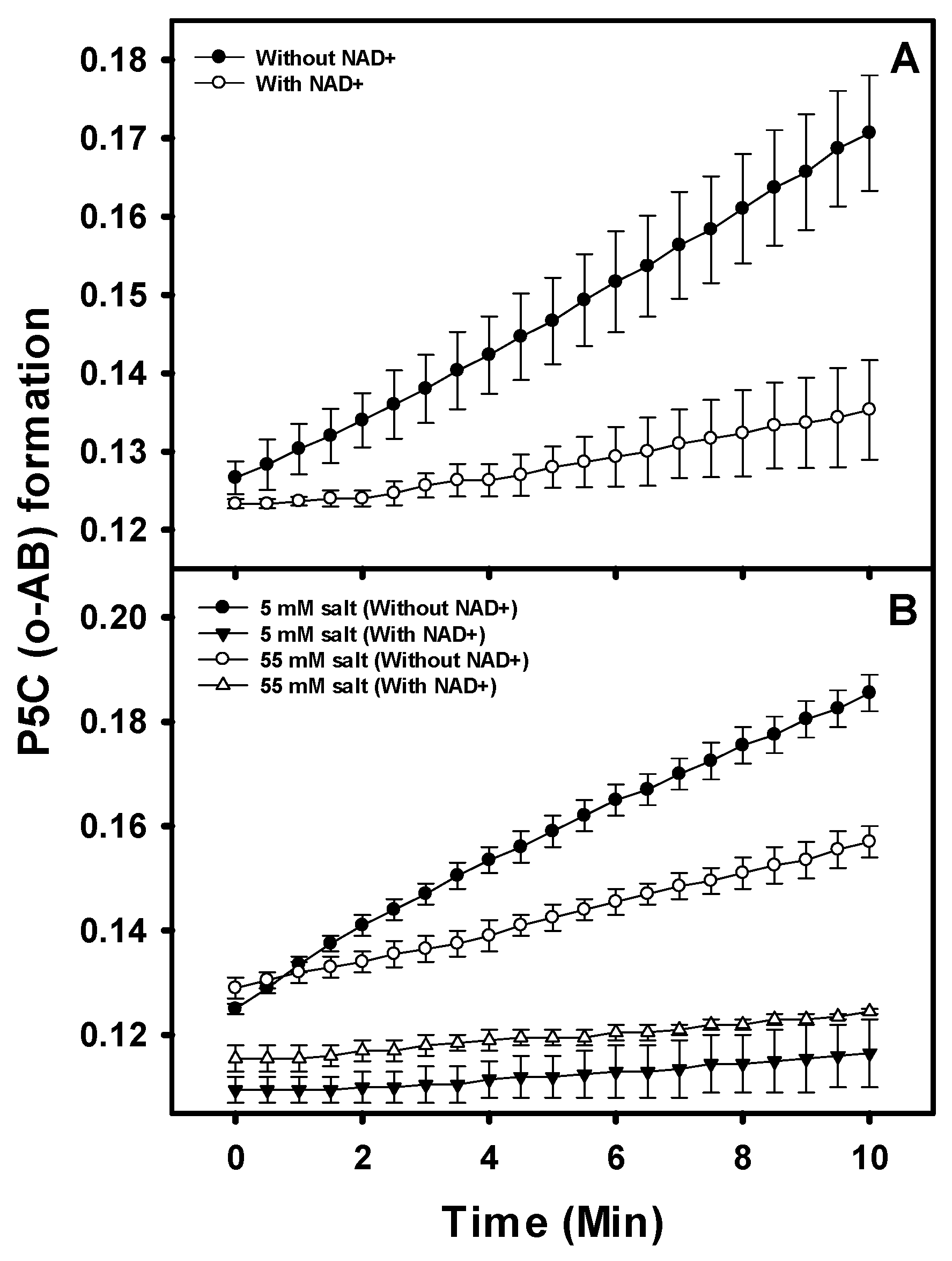
3.5. PruB-PruA Channeling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berney, M.; Weimar, M.R.; Heikal, A.; Cook, G.M. Regulation of proline metabolism in mycobacteria and its role in carbon metabolism under hypoxia. Mol. Microbiol. 2012, 84, 664–681. [Google Scholar] [CrossRef] [PubMed]
- Rachman, H.; Strong, M.; Ulrichs, T.; Grode, L.; Schuchhardt, J.; Mollenkopf, H.; Kosmiadi, G.A.; Eisenberg, D.; Kaufmann, S.H. Unique transcriptome signature of Mycobacterium tuberculosis in pulmonary tuberculosis. Infect. Immun. 2006, 74, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Karakousis, P.C.; Yoshimatsu, T.; Lamichhane, G.; Woolwine, S.C.; Nuermberger, E.L.; Grosset, J.; Bishai, W.R. Dormancy phenotype displayed by extracellular Mycobacterium tuberculosis within artificial granulomas in mice. J. Exp. Med. 2004, 200, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Berney, M.; Cook, G.M. Unique flexibility in energy metabolism allows mycobacteria to combat starvation and hypoxia. PLoS ONE 2010, 5, e8614. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.E.; Gawronski, J.D.; Dejesus, M.A.; Ioerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011, 7, e1002251. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Ioerger, T.R.; Huttenhower, C.; Long, J.E.; Sassetti, C.M.; Sacchettini, J.C.; Rubin, E.J. Global assessment of genomic regions required for growth in Mycobacterium tuberculosis. PLoS Pathog. 2012, 8, e1002946. [Google Scholar] [CrossRef] [PubMed]
- Lagautriere, T.; Bashiri, G.; Paterson, N.G.; Berney, M.; Cook, G.M.; Baker, E.N. Characterization of the proline-utilization pathway in Mycobacterium tuberculosis through structural and functional studies. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 968–980. [Google Scholar] [CrossRef]
- Serrano, H.; Blanchard, J.S. Kinetic and isotopic characterization of L-proline dehydrogenase from Mycobacterium tuberculosis. Biochemistry 2013, 52, 5009–5015. [Google Scholar] [CrossRef] [PubMed]
- Phang, J.M.; Donald, S.P.; Pandhare, J.; Liu, Y. The metabolism of proline, a stress substrate, modulates carcinogenic pathways. Amino Acids 2008, 35, 681–690. [Google Scholar] [CrossRef]
- Tanner, J.J. Structural biology of proline catabolism. Amino Acids 2008, 35, 719–730. [Google Scholar] [CrossRef]
- Becker, D.F.; Thomas, E.A. Redox properties of the PutA protein from Escherichia coli and the influence of the flavin redox state on PutA-DNA interactions. Biochemistry 2001, 40, 4714–4721. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.C.; Huang, Y.W.; Hung, H.J.; Ho, C.T.; Wu, M.L. Delta1-pyrroline-5-carboxylic acid formed by proline dehydrogenase from the Bacillus subtilis ssp. natto expressed in Escherichia coli as a precursor for 2-acetyl-1-pyrroline. J. Agric. Food Chem. 2007, 55, 5097–5102. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, E.; Ohshima, N.; Takahashi, H.; Kuroishi, C.; Yokoyama, S.; Tahirov, T.H. Crystal structure of Thermus thermophilus delta1-pyrroline-5-carboxylate dehydrogenase. J. Mol. Biol. 2006, 362, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Moxley, M.A.; Sanyal, N.; Krishnan, N.; Tanner, J.J.; Becker, D.F. Evidence for hysteretic substrate channeling in the proline dehydrogenase and Delta1-pyrroline-5-carboxylate dehydrogenase coupled reaction of proline utilization A (PutA). J. Biol. Chem. 2014, 289, 3639–3651. [Google Scholar] [CrossRef] [PubMed]
- Surber, M.W.; Maloy, S. The PutA protein of Salmonella typhimurium catalyzes the two steps of proline degradation via a leaky channel. Arch. Biochem. Biophys. 1998, 354, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Farrant, R.D.; Walker, V.; Mills, G.A.; Mellor, J.M.; Langley, G.J. Pyridoxal phosphate de-activation by pyrroline-5-carboxylic acid. Increased risk of vitamin B6 deficiency and seizures in hyperprolinemia type II. J. Biol. Chem. 2001, 276, 15107–15116. [Google Scholar] [CrossRef]
- Bearne, S.L.; Wolfenden, R. Glutamate gamma-semialdehyde as a natural transition state analogue inhibitor of Escherichia coli glucosamine-6-phosphate synthase. Biochemistry 1995, 34, 11515–11520. [Google Scholar] [CrossRef]
- Nomura, M.; Nomura, N.; Newcomb, E.W.; Lukyanov, Y.; Tamasdan, C.; Zagzag, D. Geldanamycin induces mitotic catastrophe and subsequent apoptosis in human glioma cells. J. Cell. Physiol. 2004, 201, 374–384. [Google Scholar] [CrossRef]
- Marcotte, E.M.; Pellegrini, M.; Ng, H.L.; Rice, D.W.; Yeates, T.O.; Eisenberg, D. Detecting protein function and protein-protein interactions from genome sequences. Science 1999, 285, 751–753. [Google Scholar] [CrossRef]
- Enright, A.J.; Iliopoulos, I.; Kyrpides, N.C.; Ouzounis, C.A. Protein interaction maps for complete genomes based on gene fusion events. Nature 1999, 402, 86–90. [Google Scholar] [CrossRef]
- Upadhyay, A.; Kumar, S.; Rooker, S.A.; Koehn, J.T.; Crans, D.C.; McNeil, M.R.; Lott, J.S.; Crick, D.C. Mycobacterial MenJ: An oxidoreductase involved in menaquinone biosynthesis. ACS Chem. Biol. 2018, 13, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Koehn, J.T.; Beuning, C.N.; Peters, B.J.; Dellinger, S.K.; Van Cleave, C.; Crick, D.C.; Crans, D.C. Investigating substrate snalogues for mycobacterial MenJ: Truncated and partially saturated menaquinones. Biochemistry 2019, 58, 1596–1615. [Google Scholar] [CrossRef]
- Koehn, J.T.; Crick, D.C.; Crans, D.C. Synthesis and characterization of partially and fully saturated menaquinone derivatives. ACS Omega 2018, 3, 14889–14901. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Koehn, J.T.; Gonzalez-Juarrero, M.; Crans, D.C.; Crick, D.C. Mycobacterium tuberculosis survival in J774A.1 cells is dependent on MenJ moonlighting activity, not its enzymatic activity. ACS Infect. Dis. 2020, 6, 2661–2671. [Google Scholar] [CrossRef] [PubMed]
- Noens, E.E.; Williams, C.; Anandhakrishnan, M.; Poulsen, C.; Ehebauer, M.T.; Wilmanns, M. Improved mycobacterial protein production using a Mycobacterium smegmatis groEL1DeltaC expression strain. BMC Biotechnol. 2011, 11, 27. [Google Scholar] [CrossRef]
- Kumar, S.; Mittal, E.; Deore, S.; Kumar, A.; Rahman, A.; Krishnasastry, M.V. Mycobacterial tlyA gene product is localized to the cell-wall without signal sequence. Front. Cell. Infect. Microbiol. 2015, 5, 60. [Google Scholar] [CrossRef]
- Nurul Islam, M.; Hitchings, R.; Kumar, S.; Fontes, F.L.; Lott, J.S.; Kruh-Garcia, N.A.; Crick, D.C. Mechanism of fluorinated anthranilate-induced growth inhibition in Mycobacterium tuberculosis. ACS Infect. Dis. 2019, 5, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Williams, I.; Frank, L. Improved chemical synthesis and enzymatic assay of delta-1-pyrroline-5-carboxylic acid. Anal. Biochem. 1975, 64, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Mezl, V.A.; Knox, W.E. Properties and analysis of a stable derivative of pyrroline-5-carboxylic acid for use in metabolic studies. Anal. Biochem. 1976, 74, 430–440. [Google Scholar] [CrossRef]
- Dawson, R.M.; Quinn, P.J. Protein-lipid interactions studies using monolayers of phospholipid and radioactive cytochrome c. J. Physiol. 1969, 200, 20P. [Google Scholar]
- Moxley, M.A.; Tanner, J.J.; Becker, D.F. Steady-state kinetic mechanism of the proline:ubiquinone oxidoreductase activity of proline utilization A (PutA) from Escherichia coli. Arch. Biochem. Biophys. 2011, 516, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Arentson, B.W.; Luo, M.; Pemberton, T.A.; Tanner, J.J.; Becker, D.F. Kinetic and structural characterization of tunnel-perturbing mutants in Bradyrhizobium japonicum proline utilization A. Biochemistry 2014, 53, 5150–5161. [Google Scholar] [CrossRef]
- Sanyal, N.; Arentson, B.W.; Luo, M.; Tanner, J.J.; Becker, D.F. First evidence for substrate channeling between proline catabolic enzymes: A validation of domain fusion analysis for predicting protein-protein interactions. J. Biol. Chem. 2015, 290, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Holden, H.M.; Raushel, F.M. Channeling of substrates and intermediates in enzyme-catalyzed reactions. Annu. Rev. Biochem. 2001, 70, 149–180. [Google Scholar] [CrossRef] [PubMed]
- Srere, P.A.; Ovadi, J. Enzyme-enzyme interactions and their metabolic role. FEBS Lett. 1990, 268, 360–364. [Google Scholar] [CrossRef]
- Ishikawa, M.; Tsuchiya, D.; Oyama, T.; Tsunaka, Y.; Morikawa, K. Structural basis for channelling mechanism of a fatty acid beta-oxidation multienzyme complex. EMBO J. 2004, 23, 2745–2754. [Google Scholar] [CrossRef]
- Meyer, F.M.; Gerwig, J.; Hammer, E.; Herzberg, C.; Commichau, F.M.; Völker, U.; Stülke, J. Physical interactions between tricarboxylic acid cycle enzymes in Bacillus subtilis: Evidence for a metabolon. Metab. Eng. 2011, 13, 18–27. [Google Scholar] [CrossRef]
- Rudolph, J.; Stubbe, J. Investigation of the mechanism of phosphoribosylamine transfer from glutamine phosphoribosylpyrophosphate amidotransferase to glycinamide ribonucleotide synthetase. Biochemistry 1995, 34, 2241–2250. [Google Scholar] [CrossRef]
- Zhao, H.; French, J.B.; Fang, Y.; Benkovic, S.J. The purinosome, a multi-protein complex involved in the de novo biosynthesis of purines in humans. Chem. Commun. 2013, 49, 4444–4452. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Arentson, B.W.; Becker, D.F.; Tanner, J.J. Structures of the PutA peripheral membrane flavoenzyme reveal a dynamic substrate-channeling tunnel and the quinone-binding site. Proc. Natl. Acad. Sci. USA 2014, 111, 3389–3394. [Google Scholar] [CrossRef]
- Moxley, M.A.; Becker, D.F. Rapid reaction kinetics of proline dehydrogenase in the multifunctional proline utilization A protein. Biochemistry 2012, 51, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Monteoliva, M.I.; Rizzi, Y.S.; Cecchini, N.M.; Hajirezaei, M.R.; Alvarez, M.E. Context of action of proline dehydrogenase (ProDH) in the hypersensitive response of Arabidopsis. BMC Plant. Biol. 2014, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Park, C. Bacterial responses to glyoxal and methylglyoxal: Reactive electrophilic species. Int. J. Mol. Sci. 2017, 18, 169. [Google Scholar] [CrossRef]
- Lai, S.W.T.; Lopez Gonzalez, E.J.; Zoukari, T.; Ki, P.; Shuck, S.C. Methylglyoxal and its adducts: Induction, repair, and association with disease. Chem. Res. Toxicol. 2022, 35, 1720–1746. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.C.; Teng, C.S.; Chang, J.L.; Chuang, H.S.; Ho, C.T.; Wu, M.L. Biosynthetic mechanism of 2-acetyl-1-pyrroline and its relationship with delta1-pyrroline-5-carboxylic acid and methylglyoxal in aromatic rice (Oryza sativa L.) callus. J. Agric. Food Chem. 2008, 56, 7399–7404. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipoquinone | Structure |
|---|---|
| UQ-1 |  |
| UQ-2 |  |
| MK-1 |  |
| MK-1(H2) |  |
| MK-2 |  |
| MK-4 |  |
| Substrate | Km | Vmax (pmol) | Kcat (S−1) | Kcat/Km (S−1 M−1) |
|---|---|---|---|---|
| UQ-1 | 9.7 ± 1.4 μM | 1.6 ± 0.39 | 2.3 | 2.3 × 105 |
| MK-1 | 23 ± 3.2 μM | 1.9 ± 0.04 | 2.8 | 1.2 × 106 |
| MK-1(H2) | 46 ± 13 μM | 3.2 ± 0.21 | 4.6 | 1.0 × 106 |
| MK-2 | 12 ± 0.66 μM | 1.8 ± 0.01 | 2.6 | 2.1 ×106 |
| Proline (saturating UQ-1) | 2.5 ± 0.3 mM | 1.2 ± 0.03 | 1.7 | 6.9 × 102 |
| Proline (saturating MK-1) | 14 ± 4.1 mM | nd | nd | nd |
| Substrate | Km (µM) | Vmax (pmol) | Kcat (S−1) | Kcat/Km (S−1 M−1) |
|---|---|---|---|---|
| NAD+ | 33 ± 4.3 | 0.57 ± 0.06 | 1.1 | 3.4 × 105 |
| P5C | 120 ± 0.02 | 0.54 ± 0.05 | 1.1 | 8.5 × 103 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Sega, S.; Lynn-Barbe, J.K.; Harris, D.L.; Koehn, J.T.; Crans, D.C.; Crick, D.C. Proline Dehydrogenase and Pyrroline 5 Carboxylate Dehydrogenase from Mycobacterium tuberculosis: Evidence for Substrate Channeling. Pathogens 2023, 12, 1171. https://doi.org/10.3390/pathogens12091171
Kumar S, Sega S, Lynn-Barbe JK, Harris DL, Koehn JT, Crans DC, Crick DC. Proline Dehydrogenase and Pyrroline 5 Carboxylate Dehydrogenase from Mycobacterium tuberculosis: Evidence for Substrate Channeling. Pathogens. 2023; 12(9):1171. https://doi.org/10.3390/pathogens12091171
Chicago/Turabian StyleKumar, Santosh, Steven Sega, Jamie K. Lynn-Barbe, Dannika L. Harris, Jordan T. Koehn, Debbie C. Crans, and Dean C. Crick. 2023. "Proline Dehydrogenase and Pyrroline 5 Carboxylate Dehydrogenase from Mycobacterium tuberculosis: Evidence for Substrate Channeling" Pathogens 12, no. 9: 1171. https://doi.org/10.3390/pathogens12091171