The Destructive Fungal Pathogen Botrytis cinerea—Insights from Genes Studied with Mutant Analysis



1
Michael Smith Laboratories, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
2
Department of Botany, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to the review.
Pathogens 2020, 9(11), 923; https://doi.org/10.3390/pathogens9110923
Submission received: 8 October 2020
/
Revised: 1 November 2020
/
Accepted: 4 November 2020
/
Published: 7 November 2020
(This article belongs to the Collection Reviews for "Plant Pathogens")
Abstract
:Botrytis cinerea is one of the most destructive fungal pathogens affecting numerous plant hosts, including many important crop species. As a molecularly under-studied organism, its genome was only sequenced at the beginning of this century and it was recently updated with improved gene annotation and completeness. In this review, we summarize key molecular studies on B. cinerea developmental and pathogenesis processes, specifically on genes studied comprehensively with mutant analysis. Analyses of these studies have unveiled key genes in the biological processes of this pathogen, including hyphal growth, sclerotial formation, conidiation, pathogenicity and melanization. In addition, our synthesis has uncovered gaps in the present knowledge regarding development and virulence mechanisms. We hope this review will serve to enhance the knowledge of the biological mechanisms behind this notorious fungal pathogen.
1. Introduction
Ascomycete Botrytis cinerea is a fungal pathogen responsible for gray (or grey) mold diseases. It has a broad host range, affecting many important agricultural crops. First described by Christiaan Hendrik Persoon in 1794 [1], it is considered a species within Botrytis, as it is the major pathogen within the genus [2]. Botryotinia spp. and Botrytis spp. previously caused taxonomy confusion because they induce similar disease symptoms as Sclerotinia spp. In 1949, Gregory confirmed that Botryotinia fuckeliana was the apothecial (sexual) stage of B. cinerea, establishing Botryotinia spp. and Botrytis spp. as anamorphs and teleomorphs of the same fungus [3].
B. cinerea belongs to the Sclerotiniaceae family of the class Leotiomycetes [4]. The Botrytis genus as to date comprises of 32 species [5]. Members of Botrytis genus are generally necrotrophic pathogens; they induce host cell death and lysis to access cellular nutrients. An exception to this is B. deweyae, which grows almost asymptomatically within Hemerocallis host species as an endophyte [6]. Botrytis species usually have a narrow host range, many are considered specialist pathogens that are restricted to infecting a single species or a small group of related species [7]. However, B. cinerea and B. pseudocinerea are generalists, with B. cinerea causing grey mold diseases in over 200 plants species [8]. Furthermore, B. cinerea is considered of higher agricultural and scientific importance due to its tendency of developing fungicide resistance [9].
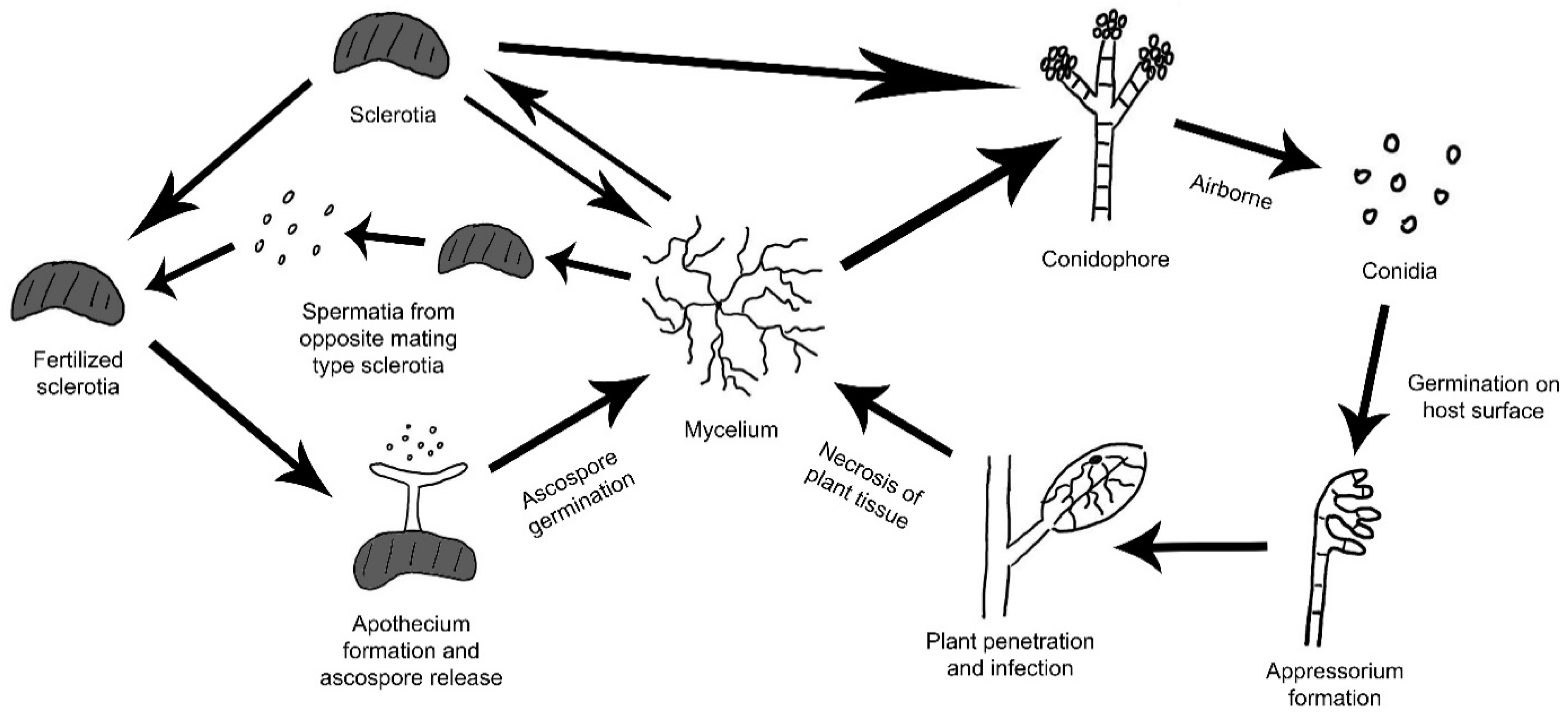
B. cinerea is a highly successful pathogen due to its flexible infection modes, high reproductive output, wide host range and ability to survive for extended periods as conidia and/or small hardened mycelia masses called sclerotia [8]. It is primarily airborne as asexual conidia spores from mature conidiophores serve as major means of transmission (Figure 1). Mycelia from sclerotia and other infected tissues as well as seeds serve as alternative inoculum. In addition to its asexual cycle, sclerotia of B. cinerea can undergo a heterothallic sexual cycle to form apothecia, which release ascospores [8,9]. Production of ascospores from apothecia involves the fertilization of sclerotia with microconidia from an opposite mating type. However, the B. cinerea sexual cycle is rarely observed in nature [8].
Invasion of host plants by B. cinerea can start from damaged tissues or natural openings, allowing the fungus to establish infection [10]. Initial infected tissue often results in limited damage. However, massive conidia production enables fast subsequent spreading. For fruiting plants, the sites of infection are usually the floral organs and the fungus has diverse means of infecting different species. In grapes, conidia infect and spread in the receptacle area [11]. In raspberries and strawberries, the stigmatic fluid serves as adhesive and nutrient medium for the airborne conidia to germinate and develop hyphae [12]. Mycelia can then grow along pathway normally taken by pollen grains, thereby entering the carpel and invading the undeveloped ovaries.
After initial infection, B. cinerea enters a short phase where it exists as a biotroph within the plant [8]. Later on, it enters an aggressive necrotrophic phase, which is proposed to be triggered by biochemical changes such as an increase in volatile organic compounds, sugar and nitrogen contents in the ripening host tissues [13,14]. During this stage, the fungus secretes virulence factors such as oxalic acid (OA), cell wall degrading enzymes (CWDEs) and analogues of plant hormones to disrupt host metabolism, immune system and cellular structure [15,16]. Effects of virulence factors on hosts are characterized by fruit decay, resulting in softening of the flesh and a browning, leathery skin [17]. B. cinerea itself undergoes rapid growth of mycelia on plant surfaces and forms massive grey conidia masses [8]. At the same time, adjacent plant tissue may become infected, allowing for the spread of grey mold across the whole plant and to plants nearby. Upon killing the host tissue, B. cinerea can continue to grow saprophytically within the plant remains in the form of sclerotia and mycelia. The sclerotia of B. cinerea are highly resistant to environmental changes. They can remain viable in soil for up to 360 days, likely due to their melanized surface, β-glucan matrix and intracellular nutrient reserves such as protein, glycogen, polyphosphate and lipids [18,19].
Hosts of B. cinerea include over 500 species of mostly dicotyledons and some monocotyledons [8], many are of economic importance. Most severely affected are agricultural crops include vegetables (e.g., cucumber, tomato, zucchini) and fruit bearing plants (e.g., strawberry, grape, raspberry) [10,20]. It is estimated that B. cinerea causes a $10 to 100 billion of produce loss annually worldwide [21]. As an example, in Florida, BFR (botrytis fruit rot) contributed to a 36% decline in strawberry harvest from 2007 to 2016, which resulted in a net production value loss of $250 million annually [22]. Due to the highly destructive nature of B. cinerea, it was ranked second on a list of fungal pathogens of scientific and economic importance [23].
As with other fungal pathogens, the most common method of controlling B. cinerea spread is by chemical means; approximately 8% of the global fungicide market is used to control this pathogen [24]. However, fungicide usage is harmful to both the environment and human health [25]. Worse still, fungicide resistance in B. cinerea can develop quickly in the field. For example, resistance to dicarboximides arose after being used to prevent grey mold. Nowadays, the effectiveness of dicarboximides has severely decreased and it is only effective when used in combination with other fungicides [26]. As an alternative to chemical control, biological control via antagonistic yeasts has been used to inhibit the onset of grey mold post-harvesting, although the effectiveness of such method is often inconsistent. Cultural control is another viable method of minimizing fruit rot damage. Removal of excessive shoots and leaves reduces the formation and spread of sclerotia and/or conidia [27]. However, cultural control is often unrealistic for large scale commercial farming.
As with most major crop pathogens, the use of resistant cultivars is the most environmentally friendly and socially accepted control method. Some strong heritable host resistance sources against B. cinerea have been found, for example, in Vitis spp. (grapevine species). However, these resistant cultivars carry undesirable commercial traits such as thicker skins and increased epidermal waxes [28]. Similar to other necrotrophic pathogens, resistance against B. cinerea is mostly multigenic. In addition, fungal pathogen resistance genes appear to be negatively correlated with fungicide resistance genes, hindering efficient breeding [28].
In this review, we will focus on recent molecular findings from genes of B. cinerea that have been studied by mutant analysis. A brief summary on its genome will be discussed first. The B. cinerea genome has been sequenced recently and only been available for the last two decades [9]. It was updated in 2016 with improved coverage and annotation [29]. We hope this review aids in a comprehensive understanding of the molecular mechanisms of the biological processes behind this widespread airborne pathogen.
2. The features of B. cinerea Genome
2.1. Genomic Sequences
The B. cinerea strain B05.10 genome, originally isolated from grapes in California USA [30], was first sequenced in 2005 by Broad Institute and Syngenta Biotechnology using Sanger technology. Later Arachne assembled the genome in 2011 with 4.5x genomic coverage [9]. The most recent revision in 2016 produced a near-complete and gapless genome sequence using third-generation sequencing methods with data acquired by Illumina and PacBio, resulting in an increased 90x coverage [29]. The total B. cinerea genome was estimated to be 41.2 Mb in size (BioProject Accession number: PRJNA15632). It comprises of 18 chromosomes with an average 42.75% GC content [29]. After the removal of repetitive sequences, 10,701 protein-coding genes were predicted using Augustus and manual curation [29]. Another reference genome available is the B. cinerea strain T4 with a genome size of 41.6 Mb and 10,427 protein-coding genes [31]. However, we will be primarily focusing on the strain B05.10 genome analysis as it is the most frequently used strain in B. cinerea mutant analysis studies. Genes with strain-specific effects are specified.
Initial analysis of the B. cinerea strain B05.10 genome sequenced in 2005 unveiled numerous pathogenicity-related genes [9], including key genes involved in reactive oxygen species (ROS) generation and tolerance, peptidase secretion, OA biosynthesis and genes encoding CWDEs. Furthermore, significant number of genes involved in secondary metabolism pathways were identified. B. cinerea genome contains 43 essential secondary metabolites (SM) enzyme-encoding genes, 24 of which are B. cinerea specific. As a result, B. cinerea can potentially produce over 40 different SM, including sesquiterpene botrydial and polyketide botcinic acid phytotoxins [32]. Interestingly, an intein element encoding a homing endonuclease (HEG) was found within the Prp8 gene in B. cinerea strain B05.10, which might have been acquired by horizontal gene transfer [33]. During meiosis with intein +/- heterozygotes, the HEG induces gene conversion between intein-carrying and empty Prp8 isolates, whereby the HEG copies the intein into the empty allele regardless of independent assortment. Later annotation of the carbohydrate-active enzymes (CAZymes) revealed a wide diversity of 229 different CAZymes, likely responsible for the decomposition of pectin, hemicellulose and cellulose in plant cell walls [34]. The large variety of CAZymes implies their critical roles in defining the broad host range of B. cinerea.
In addition to whole genome sequencing of B. cinerea strain B05.10 to a gapless near-finish state in 2016, an optical map and a genetic linkage map were constructed to minimize assembly errors [29]. This gapless genome has become the standardized B. cinerea genome due to the vast improvements in accuracy and completion over other genome versions. It also features the first sequencing of mini-chromosomes 17 and 18, containing 18 and 14 genes respectively [29]. In addition, 40 regions on chromosomes with high reshuffling rates were identified, suggesting a high frequency of recombination in B. cinerea during sexual reproduction.
In 2015, Atwell et al. sequenced the genomes of 13 B. cinerea field isolates collected from Germany, United Kingdom, California and South Africa to determine genetic variation within the species [35]. Consensus sequences in genomes were aligned and alleles variants were annotated with pairwise single nucleotide polymorphisms (SNPs) analysis. Conceivably, the most common mutations found were insertions, deletions and point mutations; an average of 28 polymorphisms per kb were found within the B. cinerea population. Intriguingly, the genomic diversity within B. cinerea seems higher relative to that in other fungal species. Further analysis on genomic variation revealed a significant number of recombinant breakpoints, suggesting extensive whole genome recombination among B. cinerea strains during meiosis. Lab propagated B. cinerea isolates were also sequenced by Atwell et al. Contrary to the mutation accumulation hypothesis [36], SNPs differences across generations cannot be attributed to elevated mutation rates [35]. This suggests that the broad host range and rapid fungicide resistance development in B. cinerea are likely a result of the high genetic diversity and recombinant shuffling within the B. cinerea population rather than spontaneous mutations. The sexual cycle of the pathogen likely occurs more prevalently in nature than under laboratory conditions, contributing to its versatile adaptivity.
2.2. Transcriptomic and Secretomic Analysis
The availability of the whole genome sequences has allowed transcriptomic and secretomic analysis of B. cinerea, permitting future investigations into biological mechanisms. Changes in gene expression between stages of conidia development were identified using microarray focusing on the initial 15 h of infection; specifically stages of dormancy, pregermination, postgermination, appressoria development and early mycelia growth [37]. These data provided evidence that a large alteration in gene expression during conidial germination aids in host cell invasion by germ tube outgrowth and appressorium differentiation.
In recent years, RNA-Seq has become a popular and flexible method to identify genes potentially associated with certain biological processes. Many transcriptomic analyses were carried out to identify contributing factors of virulence in B. cinerea. For example, a recent co-transcriptomic study on plant-necrotroph interactions between the host Arabidopsis thaliana and pathogen B. cinerea [38] discovered 10 novel pathogen co-expression networks (GCNs) encoding potential virulence factors, including proteins involved in SM secretion, copper acquisition and detoxification. The co-transcriptome data also revealed that B. cinerea GCNs are negatively associated with plant immunity and positively associated with plant photosynthetic potential. These newly identified GCNs provide a future research avenue for reverse genetic analysis to determine the molecular mechanisms of plant-host interactions and pathogenesis in B. cinerea.
Secretomics is another useful method for identifying key genes by analyzing secreted proteins and their secretion pathways. One of the previous secretomic researches on B. cinerea predicted 34.38% of all gene products to be extracellular proteins secreted by the classic endoplasmic reticulum (ER)-Golgi secretion pathway [39]. Types of secreted proteins include CAZymes, proteases, proteins activating host plant hypersensitive response (HR) and other enzymes involved in oxidative burst, OA biosynthesis. Environmental conditions such as nutrients, ambient pH and metal ions influences protein secretion levels and composition [40]. Combining secretomics and transcriptomics datasets, these newly identified GCNs and secreted proteins provide a rich reservoir of candidates for reverse genetic analysis to determine the molecular mechanisms of plant-host interactions and pathogenesis in B. cinerea.
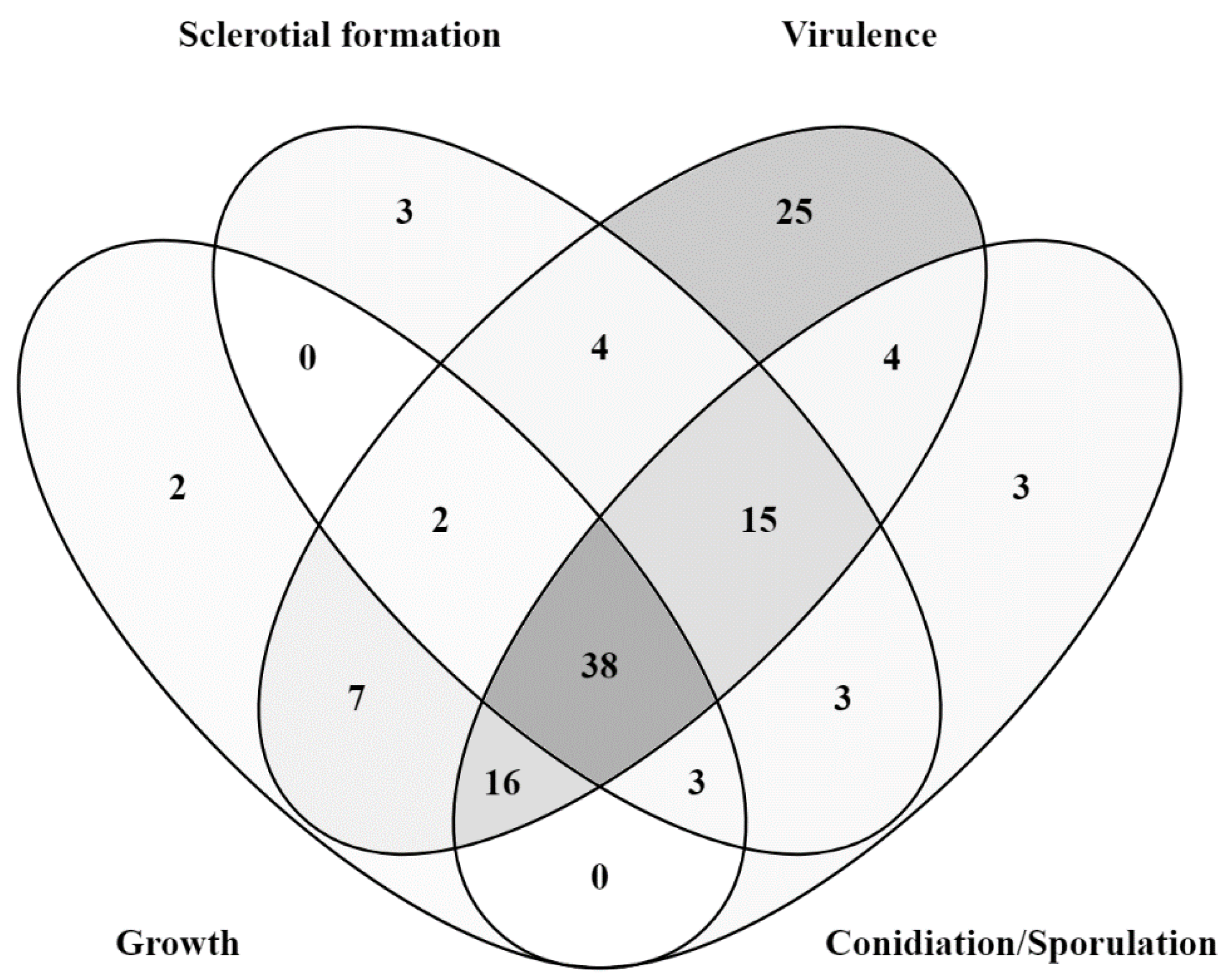
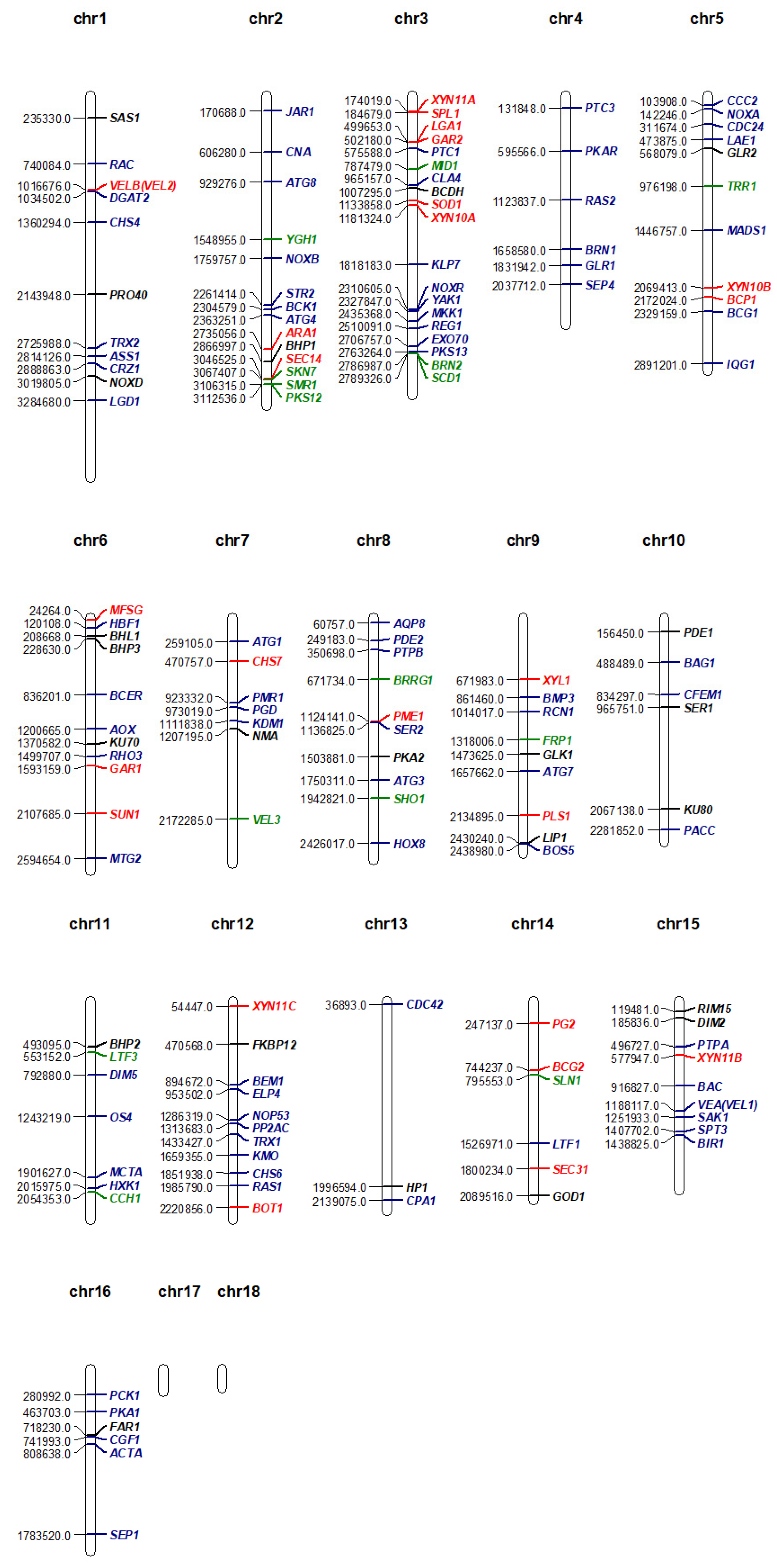
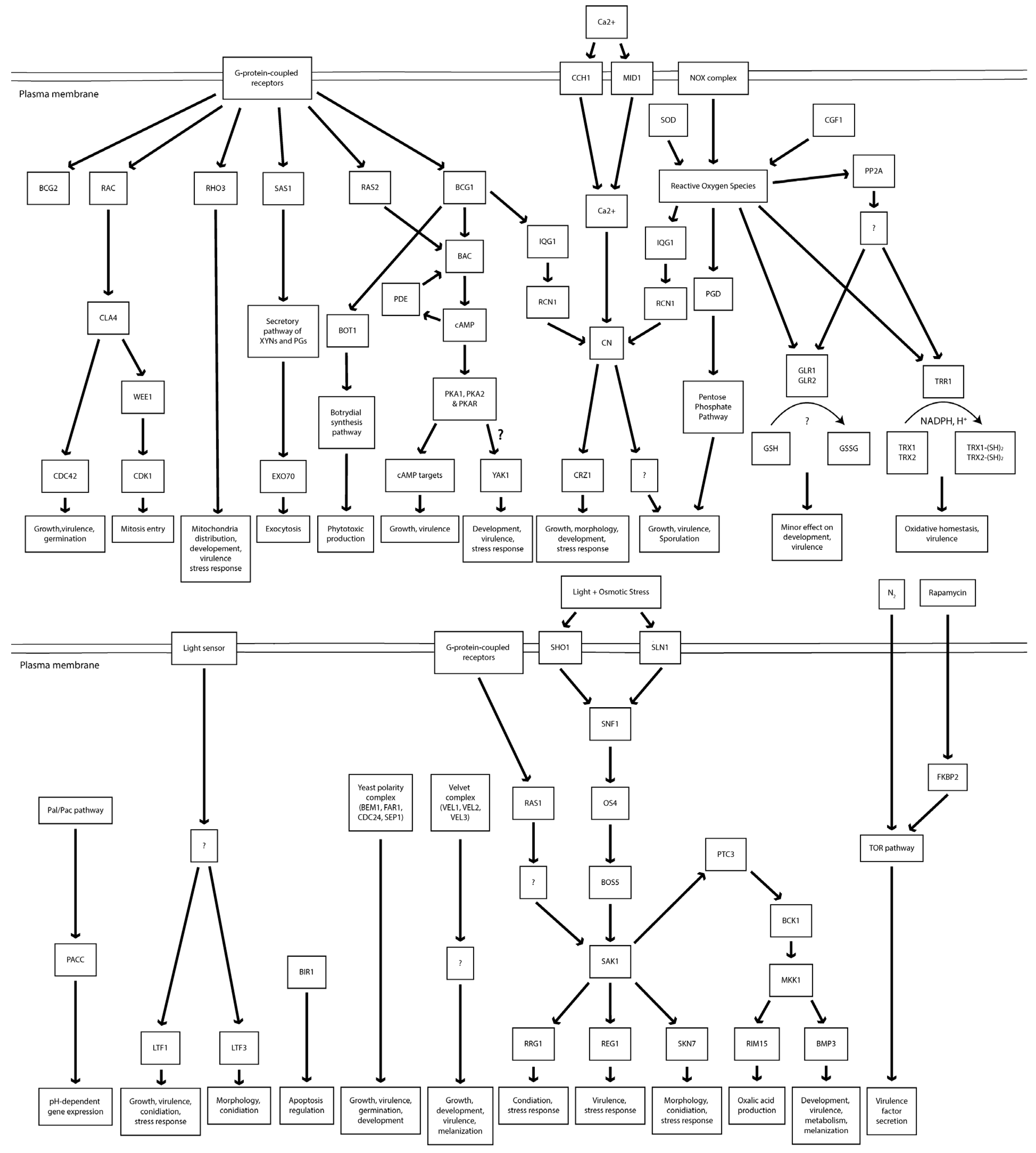
Mutant analysis is essential to establish a causal relationship between a gene and the affiliated biological process, beyond the association seen in omics analysis. Therefore, the rest of this review will provide an overview of all B. cinerea genes studied to date with mutant analysis methods (Table 1). A Venn diagram of the encoded proteins has been provided for readers’ overview (Figure 2). It is clear to see from the diagram that the majority of genes studied so far are either virulence-specific or associated with both virulence and development. A chromosomal map is constructed to detect possible clustering of virulence-associated genes (Figure 3). However, no significant gene clustering can be observed, likely because B. cinerea is still an understudied organism regardless of its economic importance. We also include a cell signaling diagram summarizing major signaling pathways discussed in the review (Figure 4).
3. Molecular Dissection of B. cinerea Biology
There are many gene nomenclature variations used by Botrytis researchers. To avoid confusion, we will be adopting the most commonly used gene/mutant/protein nomenclature. For example, ABC1 protein is encoded by italicized wild-type (WT) gene ABC1, while the mutant is designated by lowercase abc1. Exceptions will be indicated. It should be kept in mind of the different source strains of B. cinerea and plant hosts used in different studies (Table 1), which may contribute to some discrepancies in observed mutant phenotypes of the same genes.
3.1. Hyphal Growth and Virulence
Over the last two decades, accessibility to the B. cinerea whole genome sequence and advancements in mutant analysis techniques have greatly improved molecular studies of this fungus [29]. Most of the growth-related genes studied so far are highly conserved genes. Therefore, it is not surprising that these mutants often exhibit defects in sclerotial formation and/or conidiation (Table 1). They will be discussed in following sections. In this section, only genes specifically affecting hyphal growth and virulence but not differentiation, will be discussed. As mycelial growth precedes formation of structures such as appressoria, sclerotia and conidiophores, mutants with reduced growth likely would manifest diminished virulence as well. However, there are exceptions such as mutants sho1 and sln1, which exhibit severely defective growth, sclerotial formation and conidiation but normal virulence (To be discussed later).
The single and double knockout mutants of highly conserved calcium channel proteins copper chaperone (CCH1) and mating pheromone-induced death (MID1) led to reduced vegetative growth under low environmental calcium conditions [41]. Other functions such as differentiation and pathogenicity were not affected. The aforementioned channel proteins facilitate intracellular calcium homeostasis through the Ca2+/Calcineurin (CN) signaling system. Although the similar phenotypes between cch1 and mid1 mutants suggested a complex formation by CCH1 and MID1, no experimental evidence of any interactions was found in B. cinerea. In mutants of other fungal species, growth under low calcium conditions is always impacted, along with virulence and development also being affected to varying degrees [42,43]. This suggests that CCH1 and MID1 play alternative roles in later stages of signaling pathways unlike in other fungi but they are universally essential to maintain calcium homeostasis for optimal growth.
BCG1 and BCG2 encode B. cinerea G alpha (Gα) subunits, which are components of heterotrimeric GTP-binding proteins [44]. They transduce environmental signals to activate many signaling cascades such as the well-known cyclic adenosine monophosphate (cAMP) pathway and Ca2+/CN signaling system. Previous studies indicated that Gα proteins are critical for conidiation and appressorium formation [45]. Deletion of BCG1 in B. cinerea caused a decrease in growth rate and compacted colonies. Curiously, exogenous application of cAMP rescued the colony morphology, suggesting BCG1 is a direct or indirect activator of adenylyl cyclase and the cAMP-dependent pathway (discussed below) [44]. Although bcg1 mutants were able to initially colonize and penetrate host tissues, further host invasion and spread did not occur. The bcg1 mutants also failed to secrete proteases, suggesting BCG1 contributes to secretion of hydrolytic enzymes. BCG2 belongs to subfamily II of fungal Gα proteins, whose members are not well studied [44]. Unlike bcg1, bcg2 mutants undergo normal growth and infection but with slower lesion expansion. Though further research is still needed to fully understand the signaling pathway and effects of BCG2, these data suggest that BCG1 and BCG2 may control different signal transduction pathways.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of genes from B. cinerea that have been studied using mutant analysis.
| Gene Code (New) | Fungal Strains | Mutant Name | Gene Full Name | Mutant Type | Mutant Phenotypes | Host Species | Other Functions of Encoded Protein | Reference | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hyphal Growth | Sclerotial Formation | Oxalate Production | Virulence | Compound Appressoria Formation (Penetration) | Conidiation/Sporulation | Induce Host HR/Resistance | Secretion Signal | ||||||||
| Bcin01g00550 | B05.10 strain | sas1 | secretion-related Rab/GTPase gene | deletion | + | + | NA | + | NA | + | NA | No | Protein secretion, sporulation | [46] | |
| Bcin01g02000 | B05.10 strain | rac | Small GTPases | deletion | + | + | NA | + | NA | + | NA | No | Polar growth, reproduction | [47] | |
| Bcin01g02730 | B05.10 strain | vel2 | velvet-like gene | deletion | + | + | + | + | NA | + | NA | No | Light response, acidification | [48] | |
| Bcin01g02730 | 38B1 strain | velB | velvet-like gene | deletion | + | + | NA | + | - | + | NA | No | Negative role in asexual development and melanin biosynthesis | [49] | |
| Bcin01g02790 | Chickpea isolate from fields of Govind Ballabh Pant University | dgat2 | diacylglycerol O-acyl transferase 2 | T-DNA, deletion | - | + | + | + | NA | + | NA | No | Penetration and consequently virulence | [50] | |
| Bcin01g03790 | Bd90 strain | chs4 | chitin synthases | deletion | - | - | NA | - | NA | - | NA | No | [51] | ||
| Bcin01g06080 | 38B1 strain | pro40 | scaffold protein | deletion | NA | NA | + | NA | NA | NA | NA | No | [52] | ||
| Bcin01g07770 | B05.10 strain | trx2 | thioredoxin | deletion | - | NA | NA | - | - | - | NA | No | Resist to oxidative stress; strx1/trx2 double mutant has retarded growth as trr1 | [53] | |
| Bcin01g08050 | B05.10 strain | ass1 | argininosuccinate synthase | deletion | + | NA | NA | + | NA | NA | NA | No | Production of L-arginine | [54] | |
| Bcin01g08230 | B05.10 strain | crz1 | calcineurin-Responsive Zinc Finger Transcription Factor | deletion | + | + | NA | + | NA | + | NA | No | Acts downstream of calcineurin but not the only target of calcineurin | [55] | |
| Bcin01g08690 | B05.10 strain | noxD | component of the NADPH oxidase complex | deletion | NA | + | NA | + | - | + | NA | No | Interact with NOXA | [56] | |
| Bcin01g09450 | B05.10 strain | lgd1 | galactonate dehydratase gene | deletion | - | NA | NA | + | NA | NA | defence-related genes were not induced | No | Arabidopsis thaliana and Nicotiana benthamiana, not Solanum lycopersicum | D-galacturonic acid catabolism | [57] |
| Bcin02g00190 | B05.10 strain | jar1 | Histone 3 Lysine 4 (H3K4) demethylation | deletion | - | + | NA | + | + | + | NA | No | Oxidative and low-oxygen stress adaptation | [58] | |
| Bcin02g01540 | B05.10 strain | cnA | catalytic subunit of calcineurin | deletion | + | + | NA | + | NA | + | NA | No | [59] | ||
| Bcin02g02570 | B05.10 strain | atg8 | autophagy-related gene | deletion | + | + | NA | + | NA | + | NA | No | Interact with ATG4, lipid droplet metabolism | [60] | |
| Bcin02g04360 | B05.10 strain | ygh1 | alpha/beta hydrolases | deletion (heterokaryotic) | + | + | NA | - | NA | + | NA | No | Required for the formation of the key intermediate T4HN | [61] | |
| Bcin02g04930 | B05.10 strain | noxB | NADPH oxidases | deletion | - | + | - | + | + | - | NA | No | Penetration | [62] | |
| Bcin02g06470 | B05.10 strain | str2 | cystathionine γ-synthase | deletion | + | + | NA | + | NA | + | NA | No | Response to various stresses | [63] | |
| Bcin02g06590 | 38B1 strain | bck1 | MAPK cascade | deletion | + | NA | - | + | + | + | NA | No | Melanin biosynthesis | [52] | |
| Bcin02g06770 | B05.10 strain | atg4 | cysteine protease | deletion | + | + | NA | + | + | + | NA | No | Autophagy | [64] | |
| Bcin02g07700 | IK2018/B05.10 | ara1 | α-1,5-L-endo-arabinanase | deletion | - | NA | NA | + | NA | NA | NA | Yes | Arabidopsis thaliana | Host dependent, secondary lesion formation during infection | [65] |
| Bcin02g07970 | B05.10 strain | bhp1 | hydrophobin encoding gene | deletion | - | - | NA | - | - | - | NA | Yes | Development of apothecia | [66,67] | |
| Bcin02g08570 | B05.10 strain | sec14 | protein secretion related gene | deletion | - | NA | NA | + | NA | - | NA | No | Protein secretion | [68] | |
| Bcin02g08650 | 38B1 strain | skn7 | response regulator in the high-osmolarity glycerol pathway | deletion | - | + | NA | - | NA | + | NA | No | Regulation of vegetative differentiation and in the response to various stresses | [69] | |
| Bcin02g08760 | B05.10 strain | smr1 | sclerotial melanogenesis-regulatory gene | deletion | NA | + | NA | NA | NA | NA | NA | No | Sclerotial melanogenesis | [70] | |
| Bcin02g08770 | B05.10 strain | pks12 | polyketide synthase | deletion | - | + | NA | - | NA | - | NA | No | [61] | ||
| Bcin03g00500 | B05.10 strain | spl1 | cerato-platanin family protein | deletion | - | NA | NA | + | NA | NA | + | Yes | a variety of hosts | HR and PR gene induction, BAK1 required | [71] |
| Bcin03g01490 | B05.10 strain | lga1 | keto-3-deoxy-L-galactonate aldolase gene | deletion | - | NA | NA | + | NA | NA | defence-related genes were not induced | No | Arabidopsis thaliana and Nicotiana benthamiana, not Solanum lycopersicum | D-galacturonic acid catabolism | [57] |
| Bcin03g01500 | B05.10 strain | gar2 | galacturonate reductase genes | deletion | - | NA | NA | + | NA | NA | defence-related genes were not induced | No | Arabidopsis thaliana and Nicotiana benthamiana, not Solanum lycopersicum | D-galacturonic acid catabolism | [57] |
| Bcin03g01720 | 38B1 strain | ptc1 | Type 2C Ser/Thr phosphatases | deletion | + | + | NA | + | NA | + | NA | No | Melanin biosynthesis, regulation of multiple stress tolerance and virulence | [72] | |
| Bcin03g02380 | B05.10 strain | mid1 | calcium channel protein | deletion | + | - | NA | - | - | - | NA | No | Vegetative growth under conditions of low extracellular calcium | [41] | |
| Bcin03g02930 | B05.10 strain | cla4 | Rac effectors | deletion | + | NA | NA | + | NA | + | NA | No | Cell cycle regulating processes downstream of RAC | [73] | |
| Bcin03g03060 | B05.10 strain | bcdh | UDP-glucose-4,6-dehydratase | deletion | - | - | NA | - | NA | - | NA | No | Production of rhamnose-containing glycan | [74] | |
| Bcin03g03390 | B05.10 strain | sod1 | Cu-Zn-superoxide dismutase | deletion | NA | NA | NA | + | NA | NA | NA | No | Phaseolus vulgaris | [75] | |
| Bcin03g05410 | collected from chickpea feld of Govind Ballabh Pant University | klp7 | kinesin | T-DNA | + | NA | + | + | + | + | NA | No | Reduced activities of polygalacturonase (PG) and pectin methyl esterases (PME) | [76] | |
| Bcin03g06840 | B05.10 strain | noxR | regulatory subunit of the Nox complex | deletion | - | + | - | + | + | - | NA | No | Activation of both NOX enzymes | [62] | |
| Bcin03g06910 | B05.10 strain | yak1 | dual-specificity tyrosine phosphorylation-regulated protein kinase | deletion | - | + | NA | + | + | + | NA | No | Adaptation to oxidative stress and triadimefon | [77] | |
| Bcin03g07190 | 38B1 strain | mkk1 | MAPK kinase | deletion | + | NA | + | + | + | + | NA | No | Melanin biosynthesis, negatively regulates oxalic acid biosynthesis | [52] | |
| Bcin03g07420 | B05.10 strain | reg1 | ortholog of the F. oxysporum transcriptional regulator FoSge1 | deletion | - | - | NA | + | - | + | NA | No | Phaseolus vulgaris | Toxin production | [78] |
| Bcin03g07900 | B05.10 strain | exo70 | exocyst subunit gene | deletion | + | + | NA | + | NA | + | NA | No | [79] | ||
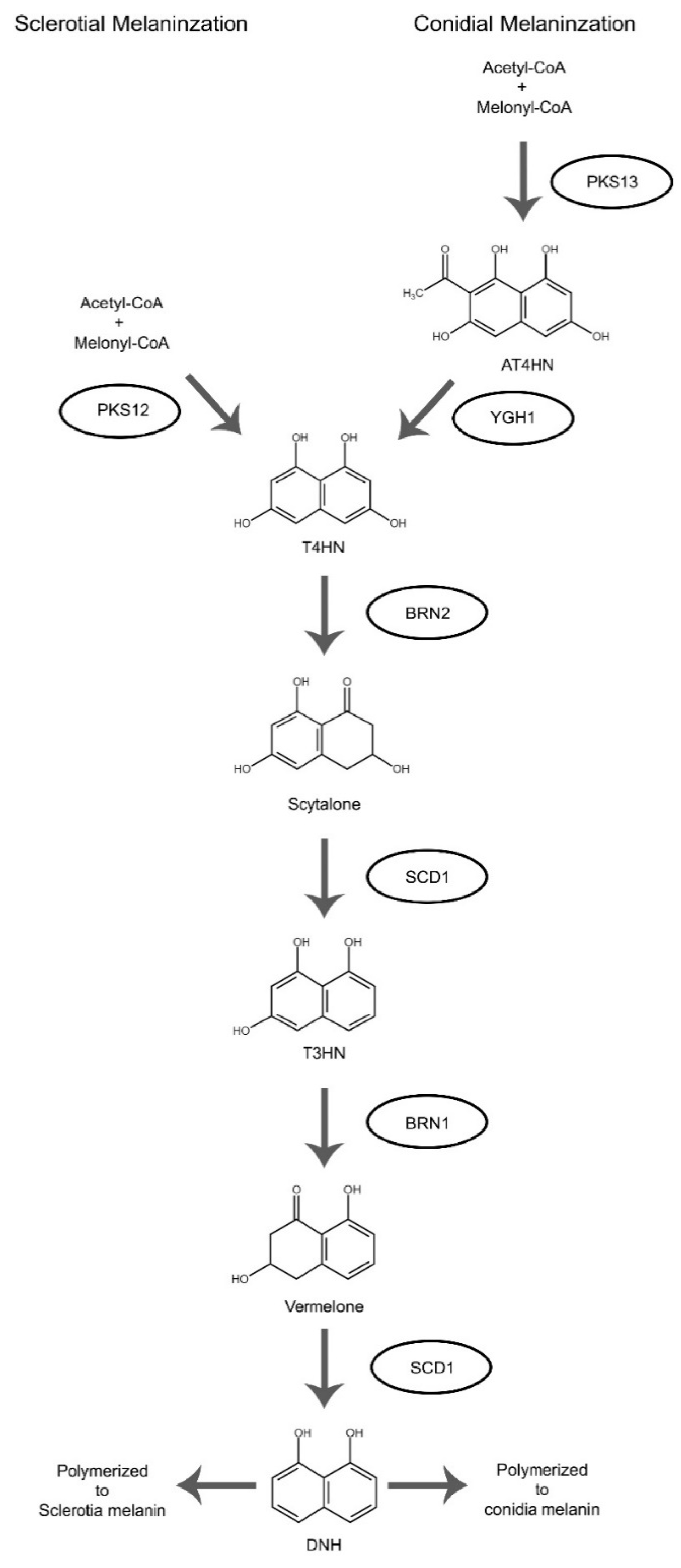
| Bcin03g08050 | B05.10 strain | pks13 | polyketide synthase | deletion | + | + | + | + | NA | + | NA | No | Melanin synthesis, mutant shows enhanced growth rate and virulence, white sclerotia | [80] | |
| Bcin03g08100 | B05.10 strain | brn2 | tetrahydroxynaphthalene (THN) reductases | deletion | - | + | NA | - | NA | + | NA | No | [61] | ||
| Bcin03g08110 | B05.10 strain | scd1 | scytalone dehydratases | deletion | - | + | NA | - | NA | + | NA | No | [61] | ||
| Bcin04g00340 | 38B1 strain | ptc3 | Type 2C Ser/Thr phosphatases | deletion | + | + | NA | + | NA | + | NA | No | Melanin biosynthesis, regulation of multiple stress tolerance and virulence | [72] | |
| Bcin04g01630 | B05.10 strain | pkaR | regulatory regulatory subunit of cAMP-dependent protein kinase | deletion | + | - | - | + | NA | - | NA | No | [81] | ||
| Bcin04g03140 | B05.10 strain | ras2 | fungal-specific Ras GTPase | deletion | + | - | - | + | NA | - | NA | No | Conidial germination | [81] | |
| Bcin04g04800 | B05.10 strain | brn1 | tetrahydroxynaphthalene (THN) reductases | deletion | + | + | + | + | NA | + | NA | No | Melanin synthesis, mutant shows enhanced growth rate and virulence, orange sclerotia | [80] | |
| Bcin04g05300 | B05.10 strain | glr1 | GSH reductase | deletion | - | NA | NA | + | + | NA | NA | No | Conidia germination | [53] | |
| Bcin04g05920 | B05.10 strain | sep4 | septin gene | deletion | - | + | NA | + | + | + | NA | No | Melanin and chitin accumulation in hyphal tips | [82] | |
| Bcin05g00240 | HYOGO11 | ccc2 | copper-transporting ATPase | deletion | - | + | NA | + | + | + | NA | No | Melanization | [83] | |
| Bcin05g00350 | B05.10 strain | noxA | NADPH oxidases | deletion | - | + | - | + | - | - | NA | No | Colonize the host tissue | [62] | |
| Bcin05g00760 | B05.10 strain | cdc24 | GEF (guanine nucleotide exchange factor) | deletion (heterokaryotic) | + | NA | NA | + | + | + | NA | No | [84] | ||
| Bcin05g01210 | B05.10 strain | lae1 | putative interaction partner of BcVEL1 | deletion | - | + | + | + | NA | + | NA | No | Putative interaction partner of VEL1 | [53] | |
| Bcin05g01430 | B05.10 strain | glr2 | GSH reductase | deletion | - | NA | NA | - | - | NA | NA | No | [53] | ||
| Bcin05g02680 | B05.10 strain | trr1 | thioredoxin reductase | deletion | + | NA | NA | + | - | - | NA | No | Resist to oxidative stress | [53] | |
| Bcin05g04030 | B05.10 strain | mads1 | MADS-box transcription factor | deletion | + | + | NA | + | NA | + | NA | No | Regulates the expression of light-responsive genes | [68] | |
| Bcin05g06320 | T4 strain | bcp1 | Cyclophilin A | deletion | - | NA | NA | + | NA | - | NA | No | [85] | ||
| Bcin05g06770 | B05.10 strain | bcg1 | Gαi subunits (I) | deletion | + | NA | NA | + | - | NA | NA | No | Protease secretion | [44] | |
| Bcin05g08290 | B05.10 strain | iqg1 | fungal homolog of the RasGAP scaffold protein IQGAP | deletion | _ | + | NA | + | + | + | NA | No | Resistance against oxidative and membrane stress | [86] | |
| Bcin06g00026 | B05.10 strain | mfsG | Major Facilitator Superfamily transporter | deletion | NA | NA | NA | + | NA | NA | NA | No | Increases tolerance to glucosinolates | [87] | |
| Bcin06g00240 | B05.10 strain | hbf1 | hyphal branching-related factor 1 | T-DNA, deletion | - | + | NA | + | + | - | NA | No | Hyphal branching | [88] | |
| Bcin06g00450 | B05.10 strain | bhl1 | Botrytis hydrophobin-like gene | deletion | - | - | NA | - | - | - | NA | Yes | [67] | ||
| Bcin06g00510 | B05.10 strain | bhp3 | hydrophobin encoding gene | deletion | - | - | NA | - | - | - | NA | Yes | Development of apothecia | [66,67] | |
| Bcin06g02380 | B05.10 strain | bcer | UDP-4-keto-6-deoxyglucose-3,5-epimerase/-4-reductas | deletion | + | + | NA | + | NA | + | NA | No | Production of rhamnose-containing glycan | [74] | |
| Bcin06g03440 | B05.10 strain | aox | alternative oxidase | deletion | - | + | NA | + | NA | + | NA | No | Adaptation to environmental stress | [89] | |
| Bcin06g03990 | B05.10 strain | ku70 | inhibitor of NHEJ | deletion | - | NA | NA | - | NA | - | NA | No | Ku deficiencies improved HR efficiency | [90] | |
| Bcin06g04390 | B05.10 strain | rho3 | small GTPases of the Rho family | deletion | + | + | NA | + | + | + | NA | No | [91] | ||
| Bcin06g04660 | B05.10 strain | gar1 | galacturonate reductase genes | deletion | - | NA | NA | + | NA | NA | defence-related genes were not induced | No | Arabidopsis thaliana and Nicotiana benthamiana, not Solanum lycopersicum | D-galacturonic acid catabolism | [57] |
| Bcin06g06040 | B05.10 strain | sun1 | Group-I SUN family of proteins | deletion | - | + | NA | + | NA | + | NA | Yes | Production of reproductive structures and adhesion to plant surface | [92] | |
| Bcin06g07300 | B05.10 strain | mtg2 | Obg protein | deletion | + | + | NA | + | NA | + | NA | No | Asexual development, environmental stress response | [30] | |
| Bcin07g00720 | B05.10 strain | atg1 | autophagy-related gene | deletion | + | + | NA | + | + | + | NA | No | Lipid metabolism | [93] | |
| Bcin07g01300 | Bd90 strain | chs7 | chitin synthases | deletion | - | - | NA | + | NA | - | NA | No | Phaseolus vulgaris, ecotype Col-0 of Arabidopsis thaliana | Virulence depends on host plants | [51] |
| Bcin07g02480 | B05.10 strain | pmr1 | P-type Ca2+/Mn2+-ATPase | deletion | + | + | NA | + | NA | + | NA | No | Solanum lycopersicum leaves and fruit and Malus domestica fruit | Biofilm formation | [94] |
| Bcin07g02610 | B05.10 strain | pgd | 6-phosphogluconate dehydrogenase | deletion | + | NA | NA | + | NA | + | NA | No | Influenced by NOX | [95] | |
| Bcin07g03050 | B05.10 strain | kdm1 | histone 3 lysine 36 (H3K36)-specific demethylas | T-DNA, deletion | + | + | NA | + | NA | + | NA | No | Stress responses and photomorphogenesis | [96] | |
| Bcin07g03340 | B05.10 strain | nma | high-temperature requirement (HtrA) family of serine proteases | deletion | - | NA | NA | - | NA | - | NA | No | Pro-apoptotic activity | [97] | |
| Bcin07g05880 | B05.10 strain | vel3 | velvet-like gene | deletion | - | - | - | - | NA | + | NA | No | Light response, acidification | [48] | |
| Bcin08g00120 | B05.10 strain | aqp8 | aquaporin 8 | deletion | + | + | NA | + | + | + | NA | No | Pigment metabolism | [98] | |
| Bcin08g00550 | B05.10 strain | pde2 | phosphodiesterase | deletion | + | + | NA | + | NA | + | NA | No | Camp signaling pathway | [99] | |
| Bcin08g00850 | 38B1 strain | ptpB | putative protein tyrosine phosphatase (PTP) gene | deletion | + | + | NA | + | NA | + | NA | No | Negative role in melanin biosynthesis; PTPA and PTPB have opposite functions in conidiation | [100] | |
| Bcin08g01740 | 38B1 strain | brrg1 | putative response regulator protein | deletion | - | NA | NA | - | NA | + | NA | No | Sensitivity to fungicides and osmotic stress | [101] | |
| Bcin08g02970 | Bd90 strain | pme1 | pectin methylesterase | deletion | - | - | NA | + | NA | - | NA | Yes | [102] | ||
| Bcin08g02990 | B05.10 strain | ser2 | subtilisin-like protease 2 | deletion | + | + | NA | + | + | + | NA | Yes | [103] | ||
| Bcin08g03910 | B05.10 strain | pka2 | catalytic subunit of cAMP-dependent protein kinase | deletion | - | - | - | - | NA | - | NA | No | [81] | ||
| Bcin08g04530 | B05.10 strain | atg3 | ubiquitin-like (UBL) protein-activating enzymes | deletion | + | + | NA | + | NA | + | NA | No | Autophagy | [104] | |
| Bcin08g05150 | B05.10 strain | sho1 | biosensors of HOG pathway | deletion | + | + | NA | - | NA | + | NA | No | Redundant for sln1 mutant | [105] | |
| Bcin08g06620 | B05.10/T4strain | hox8 | homeobox transcription factor encoding gene | deletion | + | - | NA | + | + | + | NA | No | [106] | ||
| Bcin09g01800 | B05.10 strain | xyl1 | Xylanase | deletion | - | NA | NA | + | NA | NA | + | Yes | Trigger PTI | [107] | |
| Bcin09g02390 | B05.10 strain | bmp3 | cell wall integrity MAPK | deletion | + | + | NA | + | + | + | NA | No | Melanin biosynthesis | [108] | |
| Bcin09g02820 | B05.10 strain | rcn1 | calcipressin | deletion | + | - | NA | + | NA | - | NA | No | Positive modulator of CNA | [59] | |
| Bcin09g03710 | B05.10 strain | frp1 | FRP1 F-box gene | deletion | - | + | NA | - | NA | - | NA | No | Sexual reproduction | [109] | |
| Bcin09g04170 | B05.10 strain | glk1 | glucokinase | deletion | - | NA | NA | - | NA | - | NA | No | [110] | ||
| Bcin09g04730 | B05.10 strain | atg7 | ubiquitin-like (UBL) protein-activating enzymes | deletion | + | + | NA | + | NA | + | NA | No | Autophagy | [104] | |
| Bcin09g06130 | B05.10 strain | pls1 | tetraspanin | deletion | - | - | - | + | + | - | NA | No | Penetration; sexual development | [111] | |
| Bcin09g06880 | B05.10 strain | lip1 | lipase gene | deletion | NA | NA | NA | - | NA | NA | NA | Yes | Catabolite repression | [112] | |
| Bcin09g06900 | 38B1 strain | bos5 | mitogen-activated protein kinase kinase | deletion | + | NA | NA | + | - | + | NA | No | Adaptation to iprodione and ionic stress | [113] | |
| Bcin10g00450 | B05.10 strain | pde1 | phosphodiesterase | deletion | - | - | NA | - | NA | - | NA | No | Camp signaling pathway, enhance PDE2 function | [99] | |
| Bcin10g01250 | B05.10 strain | bag1 | Bcl-2 associated athanogene | deletion | + | + | NA | + | + | + | NA | No | Hyphal melanization, response to multiple abiotic stresses and UPR pathway | [114] | |
| Bcin10g02180 | B05.10 strain | cfem1 | CFEM protein with putative GPIanchored site | deletion | - | - | NA | + | NA | + | NA | Yes | Stress tolerance | [115] | |
| Bcin10g02530 | B05.10 strain | ser1 | subtilisin-like protease 1 | deletion | - | - | NA | - | - | - | NA | Yes | [103] | ||
| Bcin10g05490 | B05.10 strain | ku80 | inhibitor of NHEJ | deletion | - | NA | NA | - | NA | - | NA | No | Ku deficiencies improved HR efficiency | [90] | |
| Bcin10g05950 | B05.10 strain | pacC | PacC transcription factor | deletion | + | + | + | + | NA | + | + | No | Production of reactive oxygen species; enzyme secretion | [116] | |
| Bcin11g01450 | B05.10 strain | bhp2 | hydrophobin encoding gene | deletion | - | - | NA | - | - | - | NA | Yes | Development of apothecia | [66,67] | |
| Bcin11g01720 | B05.10 strain | ltf3 | putative C2H2 transcription factor | deletion | - | NA | NA | - | NA | + | NA | No | [78] | ||
| Bcin11g02360 | B05.10 strain | dim5 | Histone H3 Lysine 9 Methyltransferase | deletion | + | + | NA | + | NA | + | NA | No | [117] | ||
| Bcin11g03560 | 38B1 strain | os4 | mitogen-activated protein kinase kinase kinase gene | deletion | + | NA | NA | + | - | + | NA | No | Adaption to hyperosmotic and oxidative stresses | [118] | |
| Bcin11g05350 | B05.10 strain | mctA | putative monocarboxylate transporter | deletion | - | + | NA | + | NA | + | NA | No | Pyruvate uptake | [119] | |
| Bcin11g05700 | B05.10 strain | hxk1 | hexokinase | deletion | + | NA | NA | + | NA | + | NA | No | fruit | Sugar metabolism | [110] |
| Bcin11g05810 | B05.10 strain | cch1 | calcium channel protein | deletion | + | - | NA | - | - | - | NA | No | Vegetative growth under conditions of low extracellular calcium | [41] | |
| Bcin12g01360 | B05.10 strain | fkbp12 | FK506-binding protein | deletion | - | NA | NA | - | NA | - | NA | No | Sulfur repression of the synthesis of a secreted serine protease | [120] | |
| Bcin12g02530 | B05.10 strain | bem1 | scaffold protein | deletion | - | NA | NA | + | + | + | NA | No | Part of a polarity complex involving the GEF CDC24 | [84] | |
| Bcin12g02750 | B05.10 strain | elp4 | elongator complex protein | deletion | + | + | NA | + | NA | NA | NA | No | Mycelia differentiation, melanization, various environmental stress response | [121] | |
| Bcin12g03770 | B05.10 strain | nop53 | pre-rRNA processing factor | deletion | + | + | NA | + | + | + | NA | No | Oxidative and osmotic stress adaptation | [122] | |
| Bcin12g03880 | B05.10 strain | pp2ac | a catalytic subunit of a PP2A serine/threonine protein phosphatase | T-DNA, RNAi | + | + | NA | + | NA | - | NA | No | Resistance to H2O2 | [123] | |
| Bcin12g04280 | B05.10 strain | trx1 | thioredoxin | deletion | - | NA | NA | + | - | - | NA | No | Resist to oxdative stress; bstrx1bctrx2 double mutant has retarded growth as bctrr1 | [53] | |
| Bcin12g04900 | BC22 strain | kmo | kynurenine 3-monooxygenase (KMO) | T-DNA | + | + | + | + | NA | + | NA | No | Cell wall degrading enzymes activity | [124] | |
| Bcin12g05360 | Bd90 strain | chs6 | chitin synthases | deletion (heterokaryotic ) | + | - | NA | + | NA | + | NA | No | Sexual cycle | [51] | |
| Bcin12g05760 | B05.10 strain | ras1 | Small GTPases | deletion | + | + | NA | + | NA | + | NA | No | Polar growth, reproduction | [47] | |
| Bcin12g06380 | T4 strain | bot1 | P450 monooxygenase | deletion | NA | NA | NA | + | NA | NA | NA | No | Strain-specific virulence factor | [125] | |
| Bcin13g00090 | B05.10 strain | cdc42 | small GTPase | deletion | + | + | NA | + | + | + | NA | No | Conidial germination and nuclear distribution | [126] | |
| Bcin13g05340 | B05.10 strain | hp1 | heterochromatin protein 1 | deletion | - | - | NA | - | NA | - | NA | No | [117] | ||
| Bcin13g05610 | B05.10 strain | cpa1 | capping protein (CP) subunit | deletion | + | + | NA | + | + | + | NA | No | Conidial germination; interact with CPB1 | [127] | |
| Bcin14g00610 | B05.10 strain | pg2 | endopolygalacturonase enzyme | deletion | NA | NA | NA | + | NA | NA | NA | Yes | Solanum lycopersicum and Vicia faba | [128] | |
| Bcin14g01730 | B05.10 strain | bcg2 | group II of Gα subunits | deletion | - | NA | NA | + | - | NA | NA | No | [44] | ||
| Bcin14g01870 | B05.10 strain | sln1 | biosensors of high-osmolarity glycerol(HOG) pathway | deletion | + | + | NA | - | NA | + | NA | No | Redundant for shn1 mutant | [105] | |
| Bcin14g03930 | B05.10 strain | ltf1 | light-responsive transcription factor 1 | deletion | + | NA | NA | + | + | + | NA | No | ROS homoeostasis, light-dependent differentiation | [129] | |
| Bcin14g04650 | B05.10 strain | sec31 | protein secretion related gene | deletion | - | NA | NA | + | NA | - | NA | No | Protein secretion | [68] | |
| Bcin14g05500 | B05.10 strain | god1 | putative glucose oxidase gene | deletion | NA | NA | NA | - | NA | NA | NA | Yes | Phaseolus vulgaris | [75] | |
| Bcin15g00280 | 38B1 strain | rim15 | Per-Arnt-Sim (PAS) kinase | deletion | + | NA | + | - | NA | NA | NA | No | [52] | ||
| Bcin15g00450 | B05.10 strain | dim2 | DNA methyltransferase | deletion | - | - | NA | - | NA | - | NA | No | [117] | ||
| Bcin15g01330 | 38B1 strain | ptpA | putative protein tyrosine phosphatase (PTP) gene | deletion | + | + | NA | + | NA | + | NA | No | Negative role in melanin biosynthesis; bcptpa and bcptpb have opposite functions in conidiation | [100] | |
| Bcin15g02590 | B05.10 strain | bac | adenylate cyclase | deletion | + | NA | NA | + | NA | + | NA | No | [130] | ||
| Bcin15g03390 | 38B1 strain | veA | velvet-like gene | deletion | - | + | NA | + | - | + | NA | No | Negative role in asexual development and melanin biosynthesis | [49] | |
| Bcin15g03390 | B05.10 strain | vel1 | VELVET Gene | deletion | NA | + | + | + | NA | + | NA | No | [131] | ||
| Bcin15g03580 | B05.10 strain | sak1 | Hog-type stress-activated MAPK | deletion | NA | + | NA | + | NA | + | NA | No | Early stages of infection; regulation of secondary metabolism | [132] | |
| Bcin15g04040 | B05.10 strain | spt3 | SPT3 subunit of a Spt-Ada-Gcn5-acetyltransferase | T-DNA, deletion | + | + | NA | + | NA | + | NA | No | Resistance to H2O2 | [123] | |
| Bcin15g04140 | B05.10 strain | bir1 | baculovirus IAP (inhibitor of apoptosis protein) repeat | partial knockout | + | NA | NA | + | NA | + | NA | No | Anti-apoptotic activity | [97] | |
| Bcin16g00630 | B05.10 strain | pck1 | phosphoenolpyruvatecarboxykinase gene | T-DNA, deletion | - | + | NA | + | + | + | NA | No | [133] | ||
| Bcin16g01130 | B05.10 strain | pka1 | catalytic subunit of cAMP-dependent protein kinase | deletion | + | - | - | + | NA | - | NA | No | [81] | ||
| Bcin16g01780 | B05.10 strain | far1 | scaffold protein | deletion | - | NA | NA | - | - | - | NA | No | No obvious phenotypes | [84] | |
| Bcin16g01820 | B05.10 strain | cgf1 | conidial germination-associated factor 1 | deletion | - | - | NA | + | + | + | NA | No | ROS production, osmotic and oxidative stress adaptation | [134] | |
| Bcin16g02020 | B05.10 strain | actA | actin protein | deletion | + | NA | NA | + | NA | + | NA | No | Hyphae structure | [135] | |
| Bcin16g04910 | B05.10 strain | sep1 | formin | T-DNA | + | NA | NA | + | + | + | NA | No | Septum formation and polarized growth | [84] | |
| Bcin03g03480 | B05.10 strain | xyn10A | xylanases of family GH10 | RNAi | - | - | NA | + | NA | - | NA | Yes | [136] | ||
| Bcin05g06020 | xyn10B | xylanases of family GH10 | Yes | ||||||||||||
| Bcin03g00480 | xyn11A | xylanases of family GH11 | Yes | ||||||||||||
| Bcin15g01600 | xyn11B | xylanases of family GH11 | Yes | ||||||||||||
| Bcin12g00090 | xyn11C | xylanases of family GH11 | Yes | ||||||||||||
The gene code is according to ASM185786v1 published in 2017. ‘+’ in the table represents that the phenotype of mutant is altered as compared to the corresponding WT strain, while ‘-’ indicates the mutant phenotype is unchanged. NA means not assessed. The secretion signals were found using ‘SignalP-5.0′. Unless specified, all deletions were generated with homologous recombination. The names of host species used for pathogenicity test of mutants are: Arabidopsis thaliana (Arabidopsis), Nicotiana benthamiana (tobacco), Solanum lycopersicum (tomato), Phaseolus vulgaris (French bean), Malus domestica (Apple), Vicia faba (Broad bean). The ecotype of A. thaliana mentioned in the table is Columbia-0.
The cAMP-dependent pathway requires protein kinase A (PKA) in order to phosphorylate downstream proteins under high cAMP concentration. PKA1 and PKA2 are catalytic subunits of PKA in B. cinerea [81]. Curiously, pka1 mutants displayed retarded growth and virulence, while pka2 mutants had no obvious phenotype change. Further studies on PKA activity indicated that PKA1 is the predominant catalytic subunit, as only pka1 mutants have a detectable PKA activity decrease. PKAR is the regulatory subunit of PKA. Conceivably, deletion of PKAR displayed similar phenotypes and PKA activity as pka1 mutants [81], suggesting that PKAR is essential for normal PKA1 function. However, the exact molecular mechanisms of PKAR and PKA1 interactions are not well established. Other studies in Neurospora crassa and Cryphonectria parasitica have suggested that PKA protein subunits regulate partner subunits transcription and degradation [137,138]. In B. cinerea, PKAR may likewise stabilizes PKA1 and is essential for maintaining normal PKA1 cellular concentration.
Deletion of argininosuccinate synthase gene ASS1 resulted in a decrease in growth rate under insufficient extracellular L-arginine concentration [54]. A general decrease in virulence was also observed in most host tissues but infection still occurred in tissues containing high amount of L-arginine (e.g., grapes). However, lesion expansion was slowed down over time before halting as free L-arginine was consumed, suggesting that vegetative growth and initiation of pathogenic phase in B. cinerea are dependent on free arginine concentration.
3.2. Sclerotia Development
Sclerotia naturally form within dying host tissue. During early spring when warm and humid conditions occur, sclerotia can rapidly initiate conidiophores development and production [8]. Both mycelia and conidia can serve as inoculum to initiate new infection. When fertilized with microconidia during sexual cycle, apothecia can emerge from sclerotia and upon maturation, release ascospores (Figure 1). The sclerotia are typically pigmented black from 1,8-dihydroxynaphthalene (DNH) melanin, which aids in sclerotial survival in unfavorable environments [8]. Here we first discuss genes affecting only sclerotial structure and formation. The associated mutants developed defective sclerotia and often exhibit greatly reduced sclerotial survivability. Melanization associated genes, on the other hand, will be discussed later.
Botrytis-hydrophobin BHP1, BHP2 and BHP3 are small, nonpolar proteins responsible for coating fungal surfaces with hydrophobic layers. They serve many different functions in other fungi [66]. In B. cinerea, single mutants of BHP1, BHP2 or BHP3 did not have any aberrant phenotypes compared to WT. Double mutant bhp1 bhp3 and triple mutant had an easily wettable sclerotia and suffered from compromised structural rigidity in humid conditions, suggesting BHP1 and BHP3 perform redundant functions in sclerotia development. Apothecia produced by fertilized sclerotia of double bhp1 bhp2 and triple mutant resulted in swelling of apothecium structure and collapse upon outgrowth, indicating BHP1 and BHP2 play overlapping roles for such developmental process. When a hydrophobin-like protein BHL1 was also knocked out, however, no phenotypic changes from WT were observed [67]. It remains unknown what the role of BHL1 is and whether it shares redundancy with other BHPs in B. cinerea.
FRP1 (F-box protein required for pathogenicity) is a protein that is part of an SKP1, Cullin1 and F-box (SCF) complex E3 ligase, predicted to be involved in the ubiquitination of its substrate protein [139]. Its close homologs in Fusarium oxysporum are required for virulence and non-sugar carbon metabolism [140]. FRP1 deletion in B. cinerea resulted in enhanced growth on simple sugars and inability to produce apothecia [109], indicating a function of FRP1 in suppressing growth and it is essential for apothecium development in B. cinerea. Sclerotium formation was delayed and the sclerotia formed were smaller, heavily pigmented and textured roughly. The frp1 mutant phenotype variations across fungal species suggests that FRP1 may target different proteins in varied species, which will be interesting to examine in the future.
The following discussion will focus on genes affecting virulence, sclerotium formation and conidiation. In fungi, NADPH oxidases (NOX) homologs are involved in differentiation processes and ROS generation [62]. Single and double mutants of noxA and noxB led to failed formation of sclerotia and a reduction in conidiation and virulence [56,62]. Double noxA noxB mutant exhibited greater phenotypic changes than the sum of individual noxA and noxB changes, suggesting that NOXA and NOXB have overlapping biological functions. Virulence effects of NOXA are on lesion spread while NOXB affects host tissue penetration more. Interestingly, no change in ROS production and secretion was observed in either single or double mutants, indicating that NADPH oxidases in B. cinerea do not contribute significantly to the oxidative burst mechanism and are mainly involved in differentiation processes. The mutant noxR, with a defective regulatory NOX subunit, displayed similar phenotypes as double mutant noxA noxB [62,95], suggesting that NOXR is required for normal NOXA and NOXB function. In addition, a recent study revealed a new NADPH oxidases subunit, NOXD, whose mutant exhibited identical phenotype as noxA [56]. Localization and immunoprecipitation experiments suggested that NOXA requires interaction with NOXD for normal function and NOXD may be an activator or regulator of NOXA. Possible functions of NOXD include stabilizing NOXA or assisting in binding regulatory subunits such as NOXR.
IQ motif-containing GTPase-activating protein IQG1 serves as a linkage protein for multiple intracellular components in B. cinerea, including mitogen-activated protein kinase (MAPK), Ca2+/CN and the NOX complex. IQG1 homologs in mammalian cells are involved in a wide range of central signaling pathways. iqg1 mutant is avirulent and produces significantly less conidia but more sclerotia [86]. The mutant phenotype is similar to those of noxA and noxD mutants, suggesting that IQG1 is required for normal NOXA and NOXD function. IQG1 deletion hampers MAPK and Ca2+/CN pathway activation, suggesting IQG1 is a key component in linking multiple signaling systems in B. cinerea. IQG1 may act as a bridge resolving the lack of evidence for direct interactions between NOXA and NOXR. However, no supporting data have been available so far.
Protein kinases are critical to intracellular functions because of their ability to phosphorylate components of signal transduction systems to transduce signals [141]. Not surprisingly, protein kinases were found to be involved in pathogenicity and developmental processes. Mutation of the dual-specificity tyrosine kinase Yet Another Kinase (YAK1) resulted in fewer conidia and sclerotia production [77]. Penetration ability and H2O2 resistance were also compromised. MAPK Snf1 Activating Kinase (SAK1) yeast homologs are involved in the oxidative stress response via the high-osmolarity glycerol (HOG) pathway [132]. Interestingly, unlike other fungal SAK1 homologs [142], B. cinerea SAK1 does not appear to be significantly involved in light-dependent development, as sak1 conidiophore and sclerotial formation were unaffected when grown in light and dark conditions respectively. sak1 mutant also exhibited defective conidiation and loss of virulence. Virulence loss stems from loss of appressorium development and secondary metabolism of major phytotoxins. Therefore, the SAK1 cascade in B. cinerea seems to be a general regulator of metabolism and development rather than a specific stress response regulator.
The transmembrane osmosensors synthetic high osmolarity-sensitive protein 1 (SHO1) and synthetic lethal of N-end rule (SLN1) sense osmotic stress and serve as upstream biosensors of SAK1. Single and double mutants both exhibited growth, virulence, sporulation and sclerotium formation defects [105]. Interestingly, conidia size was significantly reduced in these mutants. sln1 mutant failed to form any sclerotia but it produced a large amount of conidia regardless of lighting conditions. sho1 sln1 double mutant grown in both light and dark exhibited reduced virulence and increased sclerotia production; however, sclerotia formed were much smaller. It was suggested SHO1 and SLN1 are involved in the light-dependent differentiation process. In particularly, SLN1 appears to be a major component in this process, as sln1 failed to detect dark conditions and form sclerotia. However, SHO1 is also involved to a lesser degree, as sho1 sln1 double mutant shifted from favoring sclerotia formation to conidiation. Future investigation into SHO1 and SLN1 interactions with light-dependent differentiation associated genes such as light-responsive transcription factors may reveal how SHO1 and SLN1 are involved in this process.
Nutrient accessibility is a key element for pathogen survival and host invasion. Conceivably, pathogenicity and growth of B. cinerea would logically decrease once nutrients are diminished. Phosphoenolpyruvate carboxykinase PCK1 allows for the generation of glucose in the absence of carbohydrates [133]. pck1 deletion mutant displayed reduced conidiation, more but smaller sclerotia formation and delayed conidial and sclerotial germination. Also, the mutant conidia lost their ability of host penetration, which may partly explain its impaired pathogenicity. Besides, the mutant phenotype could be rescued by exogenous treatment of glucose. Thus, blocking of PCK1 activity may serve as a method to decrease B. cinerea spread and pathogenicity in agricultural crops.
Histone demethylation is a common way of regulating gene expression. Studies on dimethyladenosine transferase DIM5 and lysine(K)-specific histone demethylase KDM1 indicated their varying effects on pathogenicity and development, which will be discussed in later sections. Herein, deletion of one Histone 3 Lysine 4 (H3K4) demethylase JAR1 (JARID1) significantly suppresses conidiation, appressorium formation and virulence [58]. Curiously, sclerotia formation was promoted. A reduction in host-related stress adaptation and ROS production was thought to contribute to decreased virulence. Exogenous application of fructose rescued the appressorium defect, suggesting that loss of JAR1 leads to reduced uptake and utilization of fructose. Septin SEP4 plays an essential role in fungal development [82]. GFP-tagging of SEP4 revealed that absence of JAR1 prevented SEP4 from assembly within hyphae, suggesting JAR1 is required for proper SEP4 assembly and fungal tissue differentiation. Sclerotia formation may have increased to conserve energy because of absence of nutrient uptake and pathogenicity.
Synthesis of cell membrane constituents is crucial for cellular growth and development. Diacylglycerol O-acyl transferase 2 (DGAT2) catalyzes the final step of triacylglycerol (TAG) synthesis [143]. In fungi, TAG plays a significant role of maintaining lipid homeostasis and signal transduction. Deletion of DGAT2 resulted in reduced sporulation and failure to form sclerotia due to lack of TAG [50]. As TAG comprises a high proportion of sclerotia membrane lipids, TAG shortage in the mutant is suspected to have limited sclerotia formation. Mutant virulence was also negatively impacted by low host penetration and reduced OA secretion. Hyphae formed from dgat2 had swollen tips, preventing successful penetration of host tissue. The diminished activity of pH-dependent exogenous hydrolases was suspected to be diminished due to a lack of OA. Although TAG seems to affect OA levels in B. cinerea, it is unclear how DGAT2 influences OA biosynthesis and secretion.
The alterative oxidative pathway serves as a substitute electron transport chain (ETC) pathway to bypass the typical cytochrome ETC pathway. It uses alternative oxidase (AOX) as the terminal oxidase and AOX was proposed to aid in regulating intracellular ROS [89,144]. Conidiation and lesion expansion were decreased in the aox mutants and the mutants produced a larger number of smaller sclerotia. As expected, aox mutants accumulated more ROS and exhibited greater oxidative stress sensitivity. Thus, the alterative oxidative pathway in B. cinerea significantly affects both development and pathogenicity.
The B. cinerea specific hyphal branching-related factor gene HBF1 is a recent discovery whose function is still widely unknown. Mutant hbf1 suffered from significantly altered conidia morphology, hyphal branching, sclerotium formation and virulence [88]. The virulence loss was suspected to be caused by reduction in appressorium numbers and delay of appressorium host penetration. HBF1 expression was upregulated during growth and early invasion phase. The mutant phenotypes and transcriptomic data indicate that HBF1 plays an essential role in early developmental and pathogenic stages. However, there are insufficient studies to determine the exact function of HBF1. Further research into the localization and functional domains of HBF1 would be necessary.
3.3. Signaling Events Leading to Conidiation
Conidia sporulation, which is the release of asexual spores from conidiophores, serves as the major inoculum for B. cinerea [8]. Conidiophores are developed from sclerotia and mycelia in early spring. Air currents caused by temperature fluctuations can lead to conidia release. Studies have indicated that conidia formation is light-dependent and light is necessary to stimulate sporulation. In this section, we will be mainly discussing genes affecting conidiation, growth and pathogenicity. As conidiation requires large amounts of nutrients acquired from host tissue invasion, nutrient availability is inheritably linked with fungal growth [145,146]. It is therefore not surprising that genes affecting conidiation would also affect growth, which likely indirectly affects virulence.
The cAMP signaling pathway is essential for environmental signal perception and transduction. It is proved to be involved in pathogenesis and differentiation in many fungal species. In this process, cellular cAMP level has to be regulated to ensure normal signaling. At the very beginning, cAMP synthesis from ATP is catalyzed by botrytis adenylate cyclase (BAC). When BAC was knocked out, the mutant exhibited reduced vegetative growth and lesion expansion [130]. Interestingly, leaves inoculated with bac mutants failed to develop any conidia, suggesting that BAC is essential for sporulation. However, low cAMP levels were still detected in bac mutants, indicating that other similar enzymes such as guanylate cyclase might be able to partly compensate for the loss in BAC function.
Small GTPases are GTP-binding hydrolases enzymes which regulate a wide range of cellular process based on GTPase phosphorylation state. Deletion of RAS-GTPase RAS2 resulted in a delayed conidia germination, pathogenicity phase as well as reduced growth [81]. Interestingly, exogenous application of cAMP rescued the mutant to WT phenotype, suggesting that RAS2 is connected to the cAMP pathway probably by activating BAC.
MAP kinase cascades are conserved signaling modules for eukaryotic biology. The botrytis osmosensor BOS5 and OS4 are upstream histidine kinase components of the high osmolarity glycerol (HOG) pathway that significantly influences development, virulence and stress resistance in B. cinerea. Deletion of the MAPKK and MAPKKK-encoding genes BOS5 [113] OS4 [118] in B. cinerea resulted in significantly impaired hyphal growth and increased sensitivity to fungicides and ionic stress. Both mutants were unable to form any conidia or infect plant leaves. Furthermore, cucumbers inoculated with bos5 and os4 spores only produced sparse mycelium fibers without conidiation. The downstream HOG pathway component SAK1 exhibited reduced phosphorylation in bos5 and os4 mutants, supporting roles of BOS5 and OS4 in proper HOG pathway signaling.
Transcriptional regulators are essential components of signal transduction to control gene expression. The response regulator RRG1 is the putative regulator of HOG pathway in B. cinerea [101]. rrg1 deletion mutant failed to form conidia and had increased fungicide and ionic stress sensitivity but their virulence was not affected. The decreased virulence in mutants of upstream HOG pathway components such as BOS5 and SAK1 suggests that there are other regulators controlling pathogenicity. Regulator 1 (REG1) belongs to a novel class of fungal transcription regulators involved in pathogenicity and morphology [147]. The knockout mutant exhibited reduced expression of oxidative stress response genes and virulence. reg1 mycelium could penetrate host tissue but was unable to produce lesions. It is suspected that the inability to cause lesions is due to the loss of phytotoxin production. Analysis of sak1 mutant revealed that REG1 expression is dependent on SAK1 and might have contributed to the virulence loss in sak1. Suppressor of Kre Null 7 (SKN7) is another downstream transcription factor in the HOG pathway [69]. SKN7 homologs are associated with fungal development and various stress adaption. In consistency, deletion of SKN7 in B. cinerea resulted in fewer sclerotia, no conidia and increased sensitivity to H2O2.
Light-responsive transcription factor 1 (LTF1) has a significant role in regulating gene expression against the detrimental effects of light [129]. Although deletion of LTF1 decreased virulence and growth, it caused excessive but precocious conidiation. Virulence and growth could be restored by antioxidant application, indicating that the virulence and growth loss could be due to ROS accumulation. On the other hand, overexpression of LTF1 caused B. cinerea to produce more aerial mycelia and less conidia. Similarly, deletion of LTF3 promoted conidiation under all light conditions but no mature conidia could be produced [78]. ltf3 conidiophores would develop hyphae and branch off into secondary conidiophores, producing a mycelia-like structure. Conversely, overexpression of LTF3 resulted in significant suppression of conidiation development. These results indicate that both LTF1 and LTF3 are general repressors of conidia development and LTF1 suppresses it through regulating ROS homeostasis while LTF3 does so via unknown means. It is likely the two LTFs are functionally overlapping, as the double mutant ltf1 ltf3 exhibited additive mutant phenotypes [78].
Homeobox transcription factor genes (HOX) encode highly conserved master developmental regulators known to play major roles in fungal growth and differentiation. The B. cinerea genome contains 9 HOX genes. So far, only HOX8 has been studied in detail [106]. Mutants of HOX8 exhibited slow vegetative growth, reduced infection efficiency and disease progression. As HOX8 is expressed at higher level in conidia, it is not surprising that conidiogenesis was strongly affected by HOX8 deletion and hox8 mutants produced a small number of deformed conidia. It is suspected that misshaped conidia had contributed to the loss of infectivity. The hox8 phenotypes are quite unique among known B. cinerea mutants, indicating HOX8 may be part of an unknown signaling pathway.
The pentose phosphate pathway (PPP) is a highly conserved metabolic pathway playing a major role in NADPH synthesis. The initial reaction is catalyzed by the enzyme 6-phosphogluconate dehydrogenase (PGD). First discovered as a downregulated protein in noxR mutants, PGD appeared to be a major development and virulence factor regulated by the NOX complex. pgd mutants exhibited impaired virulence, growth and sporulation [95]. It is unclear how sporulation and virulence are affected but reduction in NADPH synthesis is believed to influence pathogenicity through affecting conidiophore and appressorium formation.
Hexose kinases play key roles in sugar catabolism by phosphorylating glucose and fructose. Hexokinase hxk1 mutants displayed severe growth retardation, impaired lesion formation and conidiation [110]. hxk1 conidia exhibited failed germ tubes elongation, significantly affecting the invasion ability of the mutants. The loss of sugar phosphorylation likely disrupted sugar metabolism, leading to hxk1 mutant phenotypes. On the contrary, deletion of glucokinase GLK1 had no detectable phenotypic difference from WT [110]. As enzymatic activity analysis indicated that GLK1 and HXK1 both significantly contribute to phosphorylating glucose, GLK1 likely has other redundant enzymes in B. cinerea.
3.4. Infection and Pathogenicity Mechanisms
B. cinerea mostly initiates infection through conidia spores landing on host plants. They germinate and form appressoria used for host tissue penetration [8]. Evading host immunity and production of molecules leading to host cell death are key invasion strategies in B. cinerea. Genes discussed in this section are highly specific and only affect discrete parts of the pathogenic process. We will first focus on genes essential during the pre-penetration stage of B. cinerea, where germinating conidia must adhere to plant surfaces and avoid elimination by host defenses.
Conidia of B. cinerea initially passively attach to plant surfaces via hydrophobic forces [148]. In later germination stages, an adhesive extracellular matrix (ECM) composed of carbohydrates and proteins is secreted. Secretomic analysis identified β-glucosidase Sad1 Unc-84 Domain Protein (SUN1) as a major component of B. cinerea adhesive matrix [149,150]. SUN protein family members are ascomycete specific and classified by a cysteine-rich SUN domain. Disruption of SUN1 significantly decreased conidia and mycelia adhesion to host surfaces and interfered with reproductive structure formation mostly through decreased ECM [92]. The reduced adherence contributed to the lower capacity of initiating a successful infection of sun1 mutants, thus exhibiting largely reduced symptoms in the host plants. Interestingly, sclerotia formed by sun1 mutants were significantly increased while conidiophore formation was halved, suggesting the contributions of β-glucosidase to fungal development as well. In addition to the reduced ECM adhesion, the change in conidiation significantly contributed to reduced dispersal of conidia.
Host cell wall and antifungal SM secretions are key components of plant immunity against microbial pathogens. B. cinerea needs to evade plant defenses for successful invasion. Apoptosis-like programmed cell death (PCD) in fungi can be induced by plant-secreted antifungal SMs. Thus, deactivation of antifungal SMs is necessary to prevent apoptosis. In B. cinerea, antifungal SMs are inhibited by baculovirus inhibitor of apoptosis protein (IAP) repeat BIR1 and its regulator protein nuclear mediator of apoptosis (NMA) [97]. BIR1 overexpression led to increased lesion expansion and reduced apoptosis markers. On the other hand, bir1 knockdown mutant showed decreased virulence and increased PCD. Further fluorescence imaging showed that BIR1 was strongly expressed during the first 24 h post-inoculation, indicating that BIR1 has a central anti-apoptosis role in B. cinerea during initial infection stages. nma deletion mutants exhibited higher hyphal growth and reduced apoptosis, while NMA overexpression led to increased PCD with no change in growth rate. However, both NMA deletion and overexpression strains caused slightly milder symptoms in the host. These indicate that NMA has a pro-apoptosis function in B. cinerea but it does not act as a major regulator of apoptosis and has minor effects on pathogenicity.
Intracellular accumulation of toxic compounds such as phytoanticipins and phytoalexins can severely impact pathogen survival on host plants. Active efflux of fungitoxic compounds allows fungi to gain resistance against SMs, antibiotics and fungicides. However, active efflux transporters are not well studied in plant pathogens. The Major Facilitator Superfamily transporter, MFSG plays an essential role in B. cinerea resistance to isothiocyanates (ITCs) [87]. MFSG deletion resulted in a significant virulence reduction and decreased survivability in planta, indicating that MFSG is a key component of resistance against antifungal compounds in B. cinerea. mfsg mutants exhibited varying degrees of sensitivity to products of phytoalexin glucosinolate hydrolysis, suggesting that MFSG may also export other antifungal compounds, the identities of which are unknown.
Plants also utilize ROS secretion to disrupt cellular processes in pathogens for defense. Redox systems have a central role in maintaining oxidative homeostasis in fungi. Two major redox systems in B. cinerea are the thioredoxin and glutathione (GSH) systems. Although thioredoxin and GSH systems participate in both enzymatic and antioxidative processes, we will be mainly focusing on their antioxidative properties. Thioredoxin system includes two components, thioredoxins and thioredoxin reductase. Two thioredoxins encoding genes TRX1 and TRX2 and one thioredoxin reductase encoding gene TRR1 were identified [53]. As expected, knockout mutants trx1 and trr1 exhibited increased oxidative stress sensitivity and impaired virulence. trr1 also showed enhanced H2O2 accumulation, in which oxidative stress from excess ROS caused decreased vegetative growth rate. In contrast, trx2 had no phenotypic changes, indicating a major role of TRX1. Deletion of glutathione reductase gene GLR1 only resulted in slightly decreased virulence and appressorium formation, while glr2 did not have any phenotypic changes [53]. This indicates that the GSH is not a major redox system in B. cinerea. Conceivably, studies on enzymatic activity revealed overlapping functions between the thioredoxin and GSH systems [53], whereby thioredoxin contributes to the majority of antioxidative processes in B. cinerea while GSH is a redundant and minor redox system.
CWDEs are essential in B. cinerea as they assist penetration through the plant cell walls and the breakdown of host tissue post-infection. Pectin methylesterase 1 (PME1) is secreted early in the invasion phase to hydrolyze pectin, a major plant cell wall component [102]. pme1 deletion mutant exhibited a four-fold reduction in PME activity. Consequently, mutant pathogenicity was considerably decreased. PME activity was not totally eradicated in pme1, indicating the presence of other PMEs. Investigating these PMEs is essential for fully understanding the mechanisms of PMEs in B. cinerea virulence.
The sugar side chains of pectin arabinan are depolymerized by endo-arabinasanse ARA1. ARA1 seems to be the sole arabinasanse in B. cinerea, as indicated by knockout mutants ara1 being deficient in arabinan-degrading activity [65]. ara1 exhibited decreased growth on medium with arabinan as the only carbon sources and severely decreased lesion expansion on only A. thaliana. No virulence alteration was observed in other tested hosts including Nicotiana benthamiana and tomato plants. It was proposed that the high arabinan content in A. thaliana serves as physical barriers to impede the spread of ara1. This suggests that ARA1 only plays a major pathogenicity role in plants with cell walls rich in arabinan. In comparison, deletion of the endopolygalacturonase PG2 gene resulted in various degrees of host-dependent delay in lesion expansion rate [128]. It is suggested that difference in host cell wall composition might be the cause of variation in pg2 virulence in different plants.
Endo-β-1,4-xylanases (XYNs) digest xylan, the hemicellulose component of the plant cell wall. Most fungal xylanases belong to two major families, glycosyl hydrolase families 10 (GH10) and 11 (GH11) [136]. XYN10A and XYN10B belong to the GH10 family while XYN11A, XYN11B and XYN11C belong to the GH11 family. Disrupting XYN11A resulted in 30% decrease in endoxylanase activity and severe virulence defects in lesion formation and expansion [151]. Strains with simultaneous silencing of all five XYN genes exhibited slight reduction in growth rates and lesion growth [136]. XYN activity in the RNAi mutant was reduced to the same level as xyn11A mutants. Expression analysis of the RNAi mutant indicated that XYN11A and XYN11C genes were overexpressed. This could be the result of homologous gene compensation. Further studies involving deletion of individual XYN genes would be necessary to understand the function of XYN gene family in B. cinerea. XYL1 is another xylanase in B. cinerea belonging to the SGNH (Ser-Gly-Asn-His motif) hydrolase subfamily. Deletion xyl1 mutants exhibited significant reduction in virulence [107]. Interestingly, XYL1 protein with impaired xylanase function was still able to induce plant cell death, suggesting that the cell death induced by XYL1 is separated from its xylanase activity. Further experiments on plant immunity indicated that XYL1 induces plant cell death by acting as pathogen-associated molecular patterns (PAMP) and conferring PAMP-triggered responses (PTI) in host plants.
The lipase gene (LIP1) encodes a lipolytic serine esterase, which is commonly known as lipase. Lipases are secreted by pathogenic fungi and used to degrade the outer wax layer of the cuticle [112]. As expected, deletion of LIP1 resulted in significant loss of lipase activity. However, lip1 retained full WT-like virulence. Although not all lipase secretion was eliminated in lip1, it appears that lipases do not play major pathogenic roles in B. cinerea.
B. cinerea utilizes the D-galacturonic acid (DGA) catabolic pathway to convert DGA, a monosaccharide abundant in pectin polysaccharide cell walls into pyruvate and L-glyceraldehyde. The pathway involves two nonhomologous galacturonate reductase GAR1 and GAR2, L-galactonate dehydratase LGD1 and 2-keto-3-deoxy-L-galactonate aldolase LGA1 [57]. All single knockout mutants exhibited strongly reduced virulence on DGA rich plants such as A. thaliana and N. benthamiana. A WT-like pathogenicity was seen on plants with low DGA content like tomato. The DGA catabolic intermediates appeared to inhibit B. cinerea growth, as mutants with disrupted DGA catabolic pathways exhibited reduced growth rates when incubated with DGA catabolic intermediates. It is unclear if the DGA catabolic pathway contributes to virulence as a CWDE or aids in fungal growth as an alternative nutrient source.
Monocarboxylate transporter MCTA is a major pyruvate importer in B. cinerea and is essential to access nutrients from degraded plant cell walls [119]. Deletion of MCTA resulted in decreased sclerotia formation, virulence and growth on acetate or pyruvate carbon media. However, conidiation did increase two-folds. Currently, further investigations are needed to explain the role of MCTA in the virulence of B. cinerea. Research on the fungal pH-responsive transcription factor PacC [116] suggested that B. cinerea necrosis activity is induced by low cellular pH levels, which can occur by importing acidic compounds such as pyruvate into fungal cells.
Conidial germination is essential for B. cinerea colonization. Germ tubes grown from germinating conidia develop into appressoria upon maturation. Chitin, an essential structural component of fungal cell walls, is synthesized by chitin synthases (CHSs). The functions of CHS4, CHS6 and CHS7 were previously investigated [51]. Disruption of CHS6 resulted in significant decrease in hyphal growth, conidiation and germination in heterokaryotic strains. A strong virulence reduction was also observed. No chs6 homokaryotic strains were isolated, indicating that CHS6 may be an essential enzyme for B. cinerea. chs4 and chs7 mutants had no changes in CHS activity, indicating their redundancy in chitin biosynthesis. Interestingly, the pathogenicity of chs7 mutants only decreased in A. thaliana among all the tested plant species, suggesting that CHS7 may act as a host-specific virulence factor.
Cytoskeleton is a dynamic protein network composed of microtubules, microfilaments and intermediate filaments. Microfilaments consists of polymerized actin subunits. In filamentous fungi, actin (ACT) is essential for hyphal growth and enzyme secretion. Deletion of a conserved actin gene ACTA in B. cinerea resulted in reduced growth, sporulation and virulence [135]. Proteomic analysis of actA revealed a decreased secretion of 11 CWDEs including the aforementioned XYNs. Actin is essential for normal development and pathogenicity in B. cinerea, especially for virulence factor secretion. Kinesins are microtubules-based motor proteins crucial for intracellular transportation and cell division. The kinesin-like protein KLP7 belongs to a novel kinesin subfamily exclusively found in fungi [76]. As kinesins are linked with fungal growth and development, it is not surprising that disruption of KLP7 resulted in observable defects in hyphal growth, sporulation and conidium germination. The hyphae of klp7 were irregular and stunted, which may lower the penetration ability of the mutant. Analysis of the known virulence factors indicated severely decreased secretion of OA, polygalacturonase (PG) and PMEs. Thus, KLP7 may be involved in transportation of secretory vesicles containing these virulence factors.
Identifying growth direction is essential for proper hyphal growth, reproductive structure formation and especially in host tissue penetration for fungal pathogens. Deletion mutants of orthologous components in the yeast polarity complex were generated, including the scaffold proteins bud emergence protein 1 (BEM1) and factor arrest protein FAR1, the guanine nucleotide exchange factor cell division control protein 24 (CDC24) and septin SEP1 [84]. BEM1 is a major scaffold protein in the polarity complex and the deletion mutants of bem1 exhibited significantly reduced conidiation and sclerotia formation. The reduction in colonization and virulence of bem1 could be because appressoria structure and germination rates were disrupted in the mutants. In other fungi, CDC24 is known to be critical for fungal growth. Heterokaryotic cdc24 strains of B. cinerea were used in this study as no homokaryotic strains could be isolated. The cdc24 mutants mainly exhibited growth retardation, conidium deformation and host penetration defects. Most conidia from bem1 and cdc24 mutants could not germinate and the few germinated ones gradually stopped growing. These results indicate that BEM1 and CDC24 significantly contribute to conidium germination, hyphal growth and host penetration. SEP1 plays a role in formation of septum, cell wall structures essential for fungal cell division. Deletion of SEP1 resulted in decreased growth and inability to form reproductive structures such as conidiophores and sclerotia. The reduced virulence was also observed; however, it is unclear of the reasons for pathogenicity loss. far1 mutant had no observable phenotypic changes, suggesting it plays a minor role in the B. cinerea polarity complex. Determining specific interactions between CDC24 and SEP1 with BEM1 may yield further knowledge about major signaling pathways in B. cinerea, as BEM1 significantly affects multiple cellular processes.
Following hyphal growth, the germ tube tips become swollen and form compound appressoria for host tissue penetration. Fungal appressorium formation requires the transmembrane protein tetraspanin. In ascomycetes, the punchless 1 (PLS1) tetraspanin protein is commonly associated with virulence [111]. Deletion of PLS1 resulted in female sterility and reduced virulence in B. cinerea. Interestingly, appressoria-like swollen hyphal tips were still observed in pls1 deletion mutants but they were unable to penetrate host tissue. This suggests that PLS1 may indirectly initiate penetration by interactions with proteins associated with appressorium development such as BHP1, BHP2, JAR2 or HBF1.
After penetration, B. cinerea secretes numerous virulence factors to induce host cell damage and necrosis. Generally, these factors include ROS and fungal toxins. The two H2O2 generating enzymes in B. cinerea are Cu-Zn-superoxide dismutase SOD1 and glucose oxidase GOD1 [75]. sod1 deletion mutants exhibited significantly retarded lesion development, indicating that SOD1 is a key gene in H2O2 generation. However, god1 mutants had no observable phenotypic change and GOD1 expression level in WT was very low. Thus, GOD1 may be a minor H2O2 generation enzyme in B. cinerea.
The fungal glucose methanol choline (GMC) oxidoreductase conidial germination-associated factor 1 (CGF1) is a novel virulence factor associated with extracellular H2O2 production in basidiomycetes. B. cinerea cgf1 deletion mutants exhibited impaired conidia production and germination and drastically reduced formation of invasive germ tubes and appressia [134]. Further analysis revealed that CGF1 mediates virulence by promoting endogenous ROS production, which in turn simulates infection processes such as appressorium formation. Interestingly, secretion signals in CGF1 were found via bioinformatics analysis, suggesting that CGF1 may be a secreted protein and have extracellular functions. In order to further understand functions of CGF1, more studies focusing on identifying subcellular localization and functional domains of CGF1 are necessary.
The phytotoxin botrydial is secreted only by B. cinerea, which induces necrotic lesions in hosts. The biosynthesis pathway of botrydial is still unknown. So far, only the P450 monooxygenase BOT1 has been identified as part of this synthesis pathway [125]. As expected, bot1 deletion mutants no longer produced botrydial and were significantly less pathogenic. Interestingly, BOT1 expression appeared to be regulated by a signaling cascade including BCG1 (detailed in earlier section) and suppression of BOT1 may be a major cause of virulence loss in bcg1.
Regulator of IME2 (RIM15) is a Per-Arnt-Sim (PAS) kinase downstream of Mitogen-activated protein Kinase-Kinase (MKK1) and the cell wall integrity (CWI) pathway [52]. RIM15 deletion mutants resulted in slight decreased growth rates but significantly increased OA production and extracellular protease activity. The significantly increased enzymatic activity is likely due to the secreted hydrolases being optimal under acidic conditions with extra OA. Interestingly, virulence of rim15 was not significantly different from WT, suggesting that acidification of host tissue is not a major contributor to B. cinerea pathogenicity as in other fungi.
Fungal pathogens exploit plant defense systems by inducing a HR in host plants. In doing so, a rapid necrosis of plant tissue surrounding infected areas would occur. The snod-prot-like protein SPL1, a member of the cerato-platanin family, is one of the most abundant proteins in the B. cinerea secretome [71]. Knockout spl1 mutants exhibited weaker HR and decreased lesion growth. Expression of B. cinerea SPL1 gene into Pichia pastoris yeast cells resulted in purified protein able to trigger HR in infected tissue, indicating SPL1 is a major fungal phytotoxin.
Proteins with CFEM domains (common in several fungal-specific eight-Cys-containing domain of extracellular membrane proteins) are fungal-specific extracellular membrane proteins associated with virulence in pathogenic fungi [152]. In B. cinerea, CFEM1 induces chlorosis in host leaves [115]. Deletion of CFEM1 significantly decreased pathogenicity, conidiation and increased sensitivity to osmotic and oxidative stresses. In addition to lack of chlorosis induction, the increased sensitivity to H2O2 may have contributed to virulence loss in the cfem1 mutant. Currently, the exact epitopes of CFEM1 that triggers the chlorosis pathway in host plants are unknown. Contrary to the initial assumption of being membrane-bound by a glycophosphatidylinositol (GPI) anchor, secretion signal peptide was found in CFEM1, suggesting it may be a secreted protein. Discovering the epitopes of CFEM1 and figuring out whether CFEM1 is membrane-bound or secreted would be critical to understanding the pathogenic mechanisms of CFEM1.
In filamentous fungi, the target of rapamycin (TOR) pathway links nitrogen sensing to cellular growth and development. It can be suppressed with rapamycin [120]. The FK506-binding protein FKBP12 is required for normal rapamycin disruption of the TOR pathway. As expected, deletion of FKBP12 resulted in rapamycin-resistant mutant strains, indicating that B. cinerea FKBP12 is functionally orthologous to other fungal FKBP12s in function. Interestingly, fkbp12 deletion had varying effects on virulence in different B. cinerea strains. In strain B05.10, pathogenicity was unaffected on tomato and grape but exhibited improved colonization rates on apple and cucumber fruits. However, in strain T4, fkbp12 exhibited reduced lesion expansion on tomato and bean leaves. It was suggested that FKBP12 inactivation could affect virulence factor secretion such as CDWEs and phytotoxins to various degrees in different strains.
3.5. Generalist Genes–Mutants with Defects in Multiple Aspects of B. Cinerea Biology, Including Growth, Virulence and Sclerotia or Conidia Development
In this section, we will be discussing master contributors to B. cinerea biology. These transcription factors and components of signaling systems regulate a broad range of downstream factors. Thus, it is not surprising that their disruptions would yield multiple developmental and virulence defects.
The Ca2+/CN signaling pathway is a critical fungal system required for normal virulence, hyphal growth and morphology in pathogenic fungi. CN belongs to the serine/threonine-protein phosphatase 2B (PP2B) family. Deletion of the catalytic subunit of calcineurin (CNA) in B. cinerea resulted in avirulent mutants with significantly reduced growth rates and inability to form conidia [59]. Few sclerotia were produced only after long incubation in dark conditions. Interestingly, the color of cna colonies changed over weeks from dark to light-colored, suggesting a loss of melanization. However, it remains unclear whether the change in melanization significantly contributed to other mutant phenotypes such as virulence. Furthermore, CN activity is regulated by regulators of CN (RCN1). Knockout mutant rcn1 exhibited retarded hyphal growth similar to cna and WT strains treated with CN inhibitors, indicating RCN1 functions as a positive growth regulator [59]. This was further supported by the fact that mutations in RCN1 phosphorylation sites resulted in deactivation of CN-promoting activity.
Cyclophilins are ubiquitous in all organisms and they are known for forming complexes with the drug cyclosporin A (CsA) to inhibit CN function. Like CN, cyclophilins are also cellular components involved in fungal morphogenesis and pathogenesis but their mechanism is unclear. In B. cinerea, cyclophilin A is encoded by BCP1 and deletion of the gene resulted in decelerated lesion expansion in the host [85]. In the study, three cyclopilian-dependant genes (CPD) were identified by cDNA macroarray analysis, suggesting BCP1 plays a B. cinerea specific role in promoting CPD expression. Interestingly, CN inhibition with CsA resulted in different phenotypes, including abnormal hyphal morphology and failure to form appressoria. These differences indicate that CN and cyclophilin A affect distinct aspects of morphogenesis and virulence in B. cinerea.
The CN-Responsive Zinc Finger 1 (CRZ1) is a transcription factor downstream of CN in B. cinerea. Deletion crz1 mutants showed altered hyphal morphology and a strong reduction in growth, conidiation and sclerotia formation [55]. crz1 also displayed increased sensitivity to oxidative (H2O2) and osmotic (Li+ and Ca2+) stress. crz1 penetration ability was severely impaired, indicated by delayed necrosis symptoms on intact leaves and WT-like pathogenicity on pre-wounded leaves. Treatment of crz1 conidia with CN inhibitors resulted in further additive growth defects to crz1, indicating that crz1 is not the sole target of CN. Gene expression analysis revealed that the expression of most of CRZ1-dependent genes were downregulated by CRZ1 deletion, indicating that CRZ1 acts as a transcriptional activator. However, it is uncertain whether CRZ1 directly regulates expression of these genes. Functional study of these genes is paramount to determine the effects of CRZ1 on global gene expression regulation.
PP2A is a serine/threonine protein phosphatase 2A essential for fungal growth and pathogenicity and it plays similarly critical roles as CN in signaling pathways including fungal growth, differentiation and metabolism. B. cinerea heterokaryotic PP2A deletion and PP2A silenced strains were generated, as homokaryotic PP2A deletion strains could not be isolated [123]. pp2A mutants exhibited severely defective growth and complete failure in sclerotia formation and virulence. The growth rate of PP2A deletion and silenced strains under oxidative stress was further decreased, indicating an increased sensitivity to H2O2-induced oxidative stress in the mutants. Similar change in oxidative stress sensitivity to H2O2 was observed in mutants of redox system genes such as noxB, noxR, trx1 and trr1, suggesting that PP2A may act in conjugation with these pathways to regulate ROS production.
After activation of the cAMP signaling pathway, the cAMP cascade recovers by hydrolyzation of cAMP to AMP via phosphodiesterases (PDEs). Two PDEs, a low-affinity PDE1 and a high-affinity PDE2, have been described in filamentous fungi. Deletion mutant pde1 exhibited WT-like phenotypes, while pde2 displayed severe growth, sclerotium formation defects and decreased invasive growth [99]. pde2 was also unable to sense light and surfaces for conidiation and germination. Intriguingly, intracellular cAMP levels in the pde2 were slightly lower compared to WT. It is proposed that unknown linkages between PDE2, BAC and PKA1 may be the cause of such anomaly, as BAC and PKA1 regulates cAMP levels and they had altered activity in pde2. Overexpression of PDE1 in pde2 rescued the WT-like phenotype. Therefore, PDE2 is a major PDE in B. cinerea, while PDE1 is a minor PDE with redundant functions.
Signaling components are connected by small GTPases, a group of hydrolase enzymes which transform from inactive to active state by binding to GTP. Small GTPases are classified by the conserved G domain, where GTP binding occurs. The Ras small GTPase family (first discovered in rat sarcoma) is the most studied and is known for being involved in fungal development and pathogenicity. Deletion of RAS1 gene in B. cinerea resulted in stunted hyphal growth, avirulence and inability to produce conidia [47]. Phosphorylation assays indicated that SAK1 was not phosphorylated in ras1, suggesting that RAS1 may act upstream of the SAK1 pathway.
Another small GTPase family is the RAS homologous (Rho) family, known to plays roles in cytoskeleton development and cell cycle. A prominent Rho signaling cascade is composed of the upstream RAS-related C3 (RAC), the downstream p21-activated kinase CLn activity dependent (CLA4) and effector cell division control protein 42 (CDC42). As expected, single deletion mutants rac, cla4 and cdc42 all exhibited the same phenotypes of decreased hyphal growth, defective sclerotium formation and inability to invade host tissue [47,73,126]. In both rac and cla4, the cell cycle was affected as mutants were unable to undergo mitosis to form conidia. Interestingly, that cdc42 mutants did produce misshaped conidia but with germination defects, suggesting CDC42 plays essential roles in B. cinerea growth, virulence and germination but not in conidia formation. Analysis with phosphorylation assays suggested that CLA4 may be linked to cyclin-dependent kinase CDK1, a regulator of mitosis entry, as phosphorylated CDK1 levels dropped significantly in cla4. It is suspected that CLA4 is linked to CDK1 by the signaling component WEE1 in a similar manner to other fungi [153]. Future analysis of WEE1 would be necessary to confirm its role as a linkage protein between CLA4 and CDK1.
RHO3 is another member of the Rho GTPase family and it plays varying roles in cell growth, division and protein secretion in filamentous fungi by influencing actin localization. rho3 deletion mutant showed decreased growth, conidiation, appressorium formation and virulence but enhanced sclerotium formation [91]. Besides, more ROS generation was observed in the mutant hyphal tips. Mitochondria staining experiments revealed an absence of mitochondria distribution in hyphal tips of rho3. As a result, not only did the loss of hyphal mitochondria accumulation affect germ tube and appressorium development due to reduced energy production but also virulence by attenuating intercellular ROS.
The Rab (Ras-related in brain) is a small GTPase family involved in the secretory pathway mechanisms of vesicle docking and fusion. Disruption of the Rab GTPase SAS1 (something about silencing) resulted in decreased growth, conidiation and lesion expansion in addition to failed sclerotium formation and abnormal hyphal morphology [46]. Analysis of CWDE activity indicated secretion of PGs and XYNs was significantly suppressed while gene expression was not affected. The mutants were able to penetrate and establish in host tissue but they failed to cause further symptoms. The reduction in fungal growth and development was suspected to be due to the central role SAS1 plays in vesicle secretion containing synthases and hydrolases, essential enzymes for cell wall growth, to the fungal cell wall. In summary, the small GTPase SAS1 contributes to both B. cinerea development and pathogenicity by its involvement in enzymes secretion.
The MTG2 gene encodes mitochondrial GTPase 2, a member of the Obg GTPase family (SpoOB-associated GTP-binding proteins), which is critical for mitochondrial ribosome assembly in yeast. Deletion of MTG2 gene resulted in slower growth rates and reduced conidium formation, germination and sclerotium formation in B. cinerea [30]. Interestingly, mtg2 mutant also displayed increased sensitivity to cell wall disruption, oxidative and osmotic stress. The loss of mitochondrial ribosomes in mtg2 may have caused the disruption of development, pathogenicity and environmental stress intolerances.
Transcription factors regulate gene expression by promoting or suppressing RNA polymerase II binding to DNA. So far, only a few transcription factors have been studied in B. cinerea. The pH-responsive transcription factor PACC, the terminal component of the Pal/Pac pathway, is of particular high interest, as it is essential for the necrotrophic activities of B. cinerea [116]. The fungal-specific Pal/Pac pathway regulates pH-dependent gene expression, ensuring that extracellular enzymes are only produced at suitable environmental pH. pacC deletion mutant inoculated on host tissues with neutral pH (bean and cucumbers) exhibited significant reduction in growth and virulence with no conidiophore or sclerotium formation. Secretomic analysis of pacC revealed a decrease in OA, ROS and CWDE secretion, suggesting the change in virulence is due to reduction in virulence factors secretion. Interestingly, pacC mutants grown on host tissue with acidic pH such as fruits of apple (pH 4.0), cherry tomato (pH 4.7) and grape (pH 3.8) plants exhibited a WT-like phenotype, demonstrating the importance of initial environmental acidification to induce the onset of B. cinerea pathogenicity. This is unlike rim15 mutants, where post-infection acidification is unnecessary for virulence of B. cinerea.
The ubiquitous and highly conserved MADS-Box transcription factors have been demonstrated to be essential for fungal development and virulence but the regulatory mechanisms of MADS are not well understood. Deletion of the myocyte enhancer factor (MEF2)-type MADS1 gene in B. cinerea resulted in a mutant with slower vegetative and lesion growth and produced larger conidia [68]. Interestingly, conidiation instead of sclerotia formation was induced in dark conditions, indicating MADS1 deletion disrupted the expression of light-responsive genes. It was suggested the loss of MADS1 caused an increase in photoreceptor-encoding gene expression and therefore light sensitivity. Analysis of mads1 proteome revealed the secretion proteins SEC14 and SEC31 as potential targets of MADS1. sec14 and sec31 deletion mutants both exhibited reduced lesion expansion and extracellular protein secretion, indicating they are essential for B. cinerea pathogenicity [68]. Studies on subcellular localization and secretomics of sec14 and sec31 would be necessary to understand the full roles of these secretory proteins.
The subunit of the Spt-Ada-Gcn5-acetyltransferase (SAGA) transcriptional regulator Suppressor of Ty (SPT3) was previously found to regulate growth and virulence in Candida albicans and F. oxysporum [154,155]. In addition to losing conidiation, sclerotia formation and some pathogenic properties, SPT3 knockout mutants exhibited increased sensitivity to fungicides, oxidative stress and cell wall disrupting compounds [123]. Interestingly, spt3 mutants grown without light showed accelerated growth rates, suggesting SPT3 may regulate expression of some light-dependent growth genes. In sum, the SAGA transcriptional regulator in B. cinerea regulates development, virulence and stress-induced gene expression.
Gene expression can be controlled by alternation of chromatin structure and DNA accessibility with histone modifications. Among varied modification, histone methylation plays pivotal roles by attaching methyl groups to lysine and arginine residues of histone 3 and 4, allowing for transcriptional activation or repression [96]. Deletion of histone 3 lysine 36 (H3K36) demethylase KDM1 gene caused B. cinerea to produce sclerotia in both light and dark conditions and affected excessive light and oxidative stress tolerance. As expected, growth was only impaired under stress conditions. KDM1 is critical for B. cinerea pathogenicity, as indicated by its mutant mycelia unable to form effective penetration structures. Thus, KDM1 is required for light-dependent development, stress resistance and formation of penetration structures (compound appressoria). Interestingly, insertion of KDM1 orthologs from other ascomycetes into kdm1 did not restore WT phenotype, indicating that the KDM1 functions may be species-specific. Histone H3 lysine 9 (H3K9) methyltransferase DIM5 (defective in methylation) deletion led to impaired hyphal growth, pathogenicity and reduced conidiophores and sclerotia production [117]. Decreased virulence may be the result of changes in expression of virulence factors. Indeed, expression assays revealed downregulation of virulence-related genes including SOD1, SPL1, VELVET complex genes and BOT genes. However, the cause of compromised development in dim5 is unknown. Since histone methylation by DIM5 regulates significant amounts of virulence factors, further investigations into DIM5 gene expression profile may yield novel virulence genes targeted by DIM5.
DNA methylation in B. cinerea is achieved by a complex containing heterochromatin protein HP1 and DNA methyltransferase DIM2. Single deletion mutants hp1 and dim2 did not have any noticeable defects, indicating DNA methylation may not play major virulence roles [117]. Further research may be necessary to understand the function of HP1 and DIM2 in B. cinerea.
The nucleolar protein NOP53 plays important roles in ribosome assembly and pre-rRNA processing in yeast. B. cinerea nop53 deletion mutant exhibited enhanced stress sensitivity, impaired virulence and defective conidia, sclerotia and infectious structures formation [122]. The virulence decrease was primarily attributed to loss of appressorium formation. However, reduction in ROS-generating enzymes expression such as NOX complex, IQG1, PLS1 and SOD1 was also observed in nop53, suggesting this may have contributed to further virulence loss. Interestingly, nop53 growth on cellulose-rich medium was impaired, indicating that NOP53 may control cellulase function.
Cell wall synthesis is essential for fungal growth and reproduction. A minor component of B. cinerea cell wall is the sugar residue rhamnose (RHA) [74]. To examine the contribution of RHA in B. cinerea biology, the RHA biosynthetic genes glucose dehydratase DH and deoxyglucose epimerase reductase ER were disrupted. As expected, single dh and er mutants and double mutant dh er had no RHA production. However, dh single and dh er double mutant displayed WT-like phenotype while er exhibited varying development and virulence defects. er suffered from decreased hyphal growth, conidiation, sclerotia formation, virulence and increased cell wall stress sensitivity. It was suggested that the production of intermediate UDP-4-keto-6-deoxy-glucose (UDP-KDG) may have accumulated and been integrated into the fungal cell wall, causing defects in er.
The Plasma Membrane ATPase Related (PMR1) gene encodes a P-type ATPase used to transport calcium and manganese into the Golgi compartment for protein glycosylation. Deletion mutant pmr1 exhibited defective growth, conidiation and lesion expansion in host tissues but increased sclerotia formation [94]. Analysis of cell wall composition revealed a significant reduction of glycoproteins with phosphomannan groups and increased chitin and glucan proportion. Interestingly, biofilm formation of pmr1 was compromised, indicating PMR1 plays essential roles in host colonization. The alteration in cell wall composition might be the cause of impaired pmr1 development. Another P-type ATPase in B. cinerea is the Copper-transporting ATPase CCC2, which is used to transport copper into the fungi for copper-containing proteins formation [83]. Deletion of CCC2 resulted in mutants that formed fewer sclerotia and conidia and developed fewer appressoria with abnormal morphology. ccc2 was completely avirulent and unable to cause lesions even in pre-wounded leaves, indicating that copper transporting ATPases are required for B. cinerea pathogenicity.
Kynurenine 3-monooxygenase (KMO) catalyzes the first step of the Kynurenine pathway, the primary synthesis pathway of the essential cofactor nicotinamide adenine dinucleotide (NAD). T-DNA insertion mutant of KMO resulted in reduced growth, failure to produce sclerotia or conidia but increased virulence [124]. An increase in CWDEs and phytotoxin secretion was observed in the mutant, suggesting that KMO may negatively regulated their activities. Surprisingly, expression of key genes in the cAMP and MAPK signaling pathways was also affected by KMO deletion, indicating that KMO is involved in these signaling pathways in an unknown manner.
Aquaporins (AQPs) are channel proteins that mediates the influx of water across membranes. In B. cinerea, AQP8 is essential for H2O2 uptake and the deletion mutant aqp8 exhibited suppressed growth, conidiation and sclerotia formation [98]. Interestingly, AQP8 deletion downregulated NOXR expression and the observed developmental defects may be caused by the low NOXR level. A loss of oxidative burst was also observed in aqp8 mutant, suggesting AQP8 may be required to induce ROS-mediated pathways through indirect interaction with ROS generating members such as RHO3 and SOD1.
Autophagy-related genes (ATG) encode proteins essential for autophagy, a conserved process of degrading intracellular molecules and organelles by engulfing and lysosome fusion. Autophagosome assembly is initiated by serine/threonine kinase ATG1, whereby substrates later targeted by ubiquitin-like (UQL) autophagy marker ATG8 [60,93]. ATG8 is post-translationally modified by cysteine protease ATG4 [64] and utilities UQL activating enzymes ATG3 and ATG7 to target substrates [104]. Single deletion of aforementioned ATG genes resulted in no autophagy or sclerotial formation and significantly retarded conidiation, hyphal growth and virulence. Growth was only suspended in deletion mutants after a few days and it is suspected to be due to waste product accumulation. Interestingly, sclerotia formation defects were rescued in atg3 and atg7 mutants by exogenous nutrient addition, indicating the defect might be the result of nutrient deficiency [104]. This is further supported by atg8 mutant grown in medium lacking nitrogen sources (MM-N) exhibiting delayed conidia germination [60]. No symptoms were observed on host tissues inoculated with any deletion mutants but necrosis appeared slightly in pre-wounded host tissues, indicating that loss of infection structure formation contributes to virulence reduction. The other causes of virulence loss remain unclear. Intriguingly, lipid droplet composition was significantly reduced in all mutant conidia, resulting in impaired conidiation. In sum, ATG genes play significant roles in B. cinerea development and pathogenicity.
The subtilisin-like serine proteases (SERs) have been shown to play significant development and virulence roles in pathogenic fungi. In a recent study, single and double mutants of B. cinerea ser1, ser2 and ser1 ser2 were generated [103]. While ser1 mutant displayed a WT-like phenotype, ser2 and ser1 ser2 had impacted growth, conidiation, sclerotia formation and virulence. Although growth of ser2 and ser1 ser2 mutants was only slightly decreased, no conidia or sclerotia production was observed. ser2 and ser1 ser2 could only infect wounded host tissues for lack of the infection cushions (compound appressoria). Based on the genetic data, SER2 is essential in B. cinerea development and pathogenicity, particularly for conidiation and sclerotial formation [156]. Subcellular localization experiments may help in determining the presence of interactions between SER1, SER2 and ATG proteins.
The exocyst complex serves a role in post-Golgi vesicle trafficking and tethers to the plasma membrane. The exocyst complex component EXO70 is essential for vesicle docking and fusion in yeast [157]. Deletion of EXO70 gene resulted in significantly decreased growth, condition and sclerotia formation in B. cinerea [79]. Microscopy of exo70 mutants revealed enlarged and scattered vesicles, indicating EXO70 is essential for normal vesicle formation and transportation. The impaired growth and decreased CWDEs secretion may have contributed to the reduced virulence of exo70. Interestingly, exocytosis was still observed in exo70, suggesting the existence of alternative exocyst complex-independent exocytotic mechanisms.
F-actin capping protein (CP) regulates actin filaments by binding to the barbed ends (fast growing ends). In B. cinerea, the CP consists of an α and β subunits heterodimer. Deletion of CP α subunit gene (CPA1) resulted in impaired growth, penetration ability and conidia germination and few conidia and sclerotia [127]. Growth was further decreased in light conditions, indicating a stress resistant role of CPA1. Investigating the interactions between CPA1 and the CP β subunit may reveal further properties of CPA1.
The STR2 gene encodes cystathionine γ-synthase, a key enzyme in methionine (Met) biosynthesis pathway responsible for converting cysteine into homocysteine [63]. str2 deletion mutant exhibited severely impaired vegetative growth, pathogenicity, differentiation and stress resistance. The decreased growth rate stems from Met deficiency, as str2 growth defect could be rescued by exogenous Met or homocysteine application. str2 produced fewer conidia and no sclerotia, indicating a role of STR2 in fungal development. The mutant also exhibited increased sensitivity to oxidative, osmotic and cell wall damaging agents, suggesting interactions between Met synthesis and regulatory pathways such as AOX or MAPK pathways. Interestingly, the str2 mutant could not even cause lesions in pre-wounded host tissues. It remains unknown what the virulence factors are affected to cause the complete loss of pathogenicity.
The heterodimer KU70/KU80 is the first key component of the nonhomologous end-joining (NHEJ) pathway, a mechanism of DNA repair [90]. ku70 and ku80 deletion mutants did not exhibit any development or virulence changes from WT strains. However, there were vastly improved genes inactivation rates in ku70 and ku80 deletion mutants, particularly for genes normally difficult to knockout in WT strains. Therefore, deleting ku70 and ku80 may assist in the success and efficiency in generating B. cinerea deletion mutants of other genes.
3.6. Melanization and Its Effects on Development and Pathogenicity
In fungi, production of dihydroxynaphthalene (DHN) melanin—a dark-colored pigment—is essential for their survival against abiotic stress such as ROS, desiccation, temperature fluctuations, UV rays and fungicides [158]. Consequently, there is strong interest in studying the biosynthesis and functions of melanin in B. cinerea. In particular, melanin is concentrated in sclerotia of B. cinerea, with smaller amounts found in the conidia and mycelia. We will first be discussing the melanin biosynthetic genes.
The DHN melanin biosynthetic pathway in B. cinerea (Figure 5) comprises of the polyketide synthase PKS12 and PKS13, tetrahydroxynaphthalene (TNH) reductase BRN1 and BRN2, scytalone dehydratase SCD1 and the hydrolase YGH1 [61]. During sclerotial melanization, the initial acetyl-CoA substrate is converted to 1,3,6,8-TNH (T4HN) by PKS12. T4HN is then catalyzed to scytalone by BRN2 and then dehydrated by SCD1 to 1,3,6-TNH (T3HN). T3HN is reduced and dehydrated by BRN1 and SCD1 sequentially to DHN, which is finally polymerized to DHN melanin by unknown enzymes. Interestingly, conidium and mycelium melanogenesis utilizes an alternative pathway of T4HN generation. In detail, acetyl-CoA is converted with PKS13 to acetyl-1,3,6,8-TNH (AT4HN) and then hydrolyzed by YGH1 to T4HN.
Deletion of the PKS12 and PKS13 genes resulted in albino sclerotia and conidia in B. cinerea, indicating complete melanogenesis disruption [61,80]. However, pks13 mutants did not exhibit any changes in development or pathogenicity. The effects of PKS12 remains unclear due to conflicting reports on pks12 mutant virulence, conidiation and hyphal growth phenotypes in the two individual studies. However, similarities between PKS13 and PKS12 structures support the idea that PKS12 is likely not involved in fungal development and virulence. As for TNH reductase BRN1 and BRN2, not only were brn1 deletion mutants deficient in melanin biosynthesis but they also exhibited reduced conidiation and increased growth and virulence [80]. This was contradicted in another study, where sclerotia from brn1 brn2 double mutants were similar to WT [61]. Although brn1 mutant analysis has conflicting results, it is likely that the effects of BRN1 deletion would be similar to other melanin biosynthesis genes and is nonessential for fungal development and pathogenicity. Deletion scd1 mutant exhibited strong reduction in melanogenesis with no effects on other cellular processes [61]. Intriguingly, heterokaryotic ygh1 deletion mutant exhibited decreased conidia formation [61], suggesting that either conidia melanization is essential for conidiation or PKS13 products (AT4HN) is detrimental to conidia formation. In fact, the latter seems more likely, as disruptions in later stages of melanin biosynthesis did not affect conidiation. In sum, studies so far indicated that melanization does not significantly affect fungal development and pathogenicity.
Studies have indicated that fungal melanin biosynthesis is controlled by MAPK pathways [159]. As mentioned in earlier sections, MAPK pathways regulate a vast number of gene expression. In B. cinerea, melanin biosynthesis is induced by the conserved CWI pathway, consisting of the cascade of BCK1 (Bypass of C Kinase), MKK1 and BMP3 (Botrytis MAPK 3) [52,108]. Single deletion mutants bck1, mkk1 and bmp3 all exhibited severely reduced pathogenicity, hyphal growth, conidiation and tolerance against cell wall and oxidative stress. bmp3 mutant also failed to produce any sclerotia. Furthermore, melanization and expression of melanogenesis genes SCD1 and PKS13 were significantly downregulated, indicating that the CWI pathway induces melanin synthesis. Although sclerotia formation was not tested in bck1 and mkk1, it is highly possible that these mutants also failed to produce sclerotia because they are upstream kinases of BMP3. Interestingly, virulence in mkk1 was not decreased relative to bck1 and bmp3 because of increased OA secretion in mkk1 [52]. Protein analysis revealed that MKK1 negatively regulates OA secretions by impeding RIM15 phosphorylation. The primary cause of defective pathogenicity appears to be from decreased appressium formation. PRO40 serves as a scaffold protein for the BCK1-MKK1-BMP3 cascade [52]. Disruption of the PRO40 gene resulted in increased acid and hydrolase sensitivity and reduced activation of components in the CWI pathway, indicating PRO40 is essential for the function of the CWI pathway. Overall, the CWI pathway appears to be essential to B. cinerea development and pathogenicity and plays key roles in inducing melanogenesis gene expression. It remains unknown how the CWI pathway connects with other MAPK pathways.
In B. cinerea, protein tyrosine phosphatase PTPA and PTPB positively regulate phosphorylation of SAK1 and BMP3 [100]. Subsequently, single deletion of PTPA and PTPB genes resulted in reduced activation of the SAK1 and CWI pathway. Both deletion mutants exhibited defective sclerotium formation and virulence and had increased osmotic and oxidative stress sensitivity. Only ptpA had compromised conidiation. Interestingly, ptpA and ptpB melanin pigment was drastically increased despite suppression of the CWI pathway, indicating PTPA and PTPB are not upstream phosphatases of the CWI pathway. PTPA and PTPB may be negative regulators of melanin biosynthesis through unknown pathways. Similarly, deletion of Type 2C Serine/Threonine phosphatases (PP2Cs) PTC1 and PTC3 genes resulted in significantly impaired growth, sclerotium formation and virulence but increased conidiation [72]. Also, ptc1 and ptc3 deletion mutants have highly pigmented mycelia compared to WT. Furthermore, in vivo phosphorylation analysis in the study revealed that PTC3 is phosphorylated by SAK1, indicating PTC3 may be indirectly manipulating melanin biosynthesis through the MAPK-associated CWI pathway.
The regulators of DHN melanin biosynthetic gene expression appear to specifically control melanization of sclerotia and conidia separately. The sclerotial melanin regulator SMR1 regulates melanization of sclerotia [70]. Analysis of black colored (BS) and orange colored (OS) sclerotia mutants, named respectively after sclerotia appearance, revealed a mutation in SMR1, which resulted in downregulation of sclerotial melanogenesis genes including PKS12, YGH1, BRN1/2, SCD1. The loss of melanin in OS resulted in lower survival rates under all conditions and increased susceptibility to mycoparasites infection [160]. However, conidiation and conidial melaninization were not affected, suggesting that SMR1 is a positive regulator of genes only for sclerotial melanization. Change in development and pathogenicity in smr1 was not tested.
Disruption of multiple broad regulators also severely affects melanization in B. cinerea. The deletion of the transcription elongator factor ELP4 resulted in a mutant with decreased growth, virulence and sensitivity to osmotic and oxidative stress [121]. No sclerotia were formed in elp4. Hyphal and conidia melanization were reduced, which was confirmed by expression analysis of downregulated PKS13 and SCD1. The change in cell wall protein and polysaccharide composition may have contributed to the reduction in growth and virulence. Another mutation affecting conidial melanization was the BAG1 gene deletion. BAG1 belongs to the diverse Bcl-2 associated athanogene (BAG) family, a group of co-chaperones which assists chaperones in their molecular functions [114]. bag1 deletion mutants exhibited enhanced hyphal and conidial melanin accumulation, decreased pathogenicity and conidiation and failure to form sclerotia. Virulence loss stemmed from reduction in penetration structure formation and conidia germination. Increased sensitivity to salt, cell wall and temperature stress was also observed in bag1. Yeast-two hybrid assays revealed BAG1 negatively regulates the unfolded protein response (UPR) and the CWI pathway. Contrary to the assumption that melanization reduces stress sensitivity, bag1 deletion mutants displayed increased environmental stress sensitivity. This may be because ELP4 and BAG1 both regulate the CWI pathway, which would have affected the integral composition of the fungal cell wall. The effects of melanin in B. cinerea remain unclear because melanization disruptions in most cases also interfere with the CWI pathway.
The VELVET complex plays various roles in the regulation of virulence, light-dependent development and melanization in fungi by acting as transcription factors for fungal development and SM gene expression. Currently, the VELVET proteins VEL1, VEL2 and VEL3 (VEL1 and VEL2 is also known as VEA and VELB respectively) have been studied in B. cinerea [48,49,131]. Deletion of VEL1 and VEL2 genes resulted in increased mycelial melanization and conidiation but reduced hyphal growth, sclerotial formation and lesion expansion. Studies on vel1 and vel2 secretome revealed that the mutants secreted significantly less citric acid and OA, which is suspected to be the primary cause of decreased pathogenicity [48,161]. The increased melanization is due to increased expression of melanin biosynthesis genes [49,131]. Deletion vel3 mutant developed both conidiophores and sclerotia in darkness, indicating the involvement of the VELVET complex in light sensing processes [48]. A VELVET-like complex is also formed by VEL1, VEL2 and methyltransferase loss of AFLR expression (LAE1). Deleting the LAE1 gene resulted in mutants with reduced pathogenicity, light independent conidiation and sclerotial formation loss [161]. Similar to VELVET mutants, melanogenesis genes expression was increased in lae1. The cause of reduced virulence is likely due to reduced expression of SM synthesis enzymes and CWDE genes in lae1. In sum, the B. cinerea VELVET complex negatively regulates melanization and is essential for light-dependent development, acid secretion and virulence factor secretion. However, the role of melanization cannot be determined from VELVET complex mutants, as the mutant phenotypes are caused by disruption of multiple processes in B. cinerea.
4. Conclusions
Over the past decade, many genes involved in the development and pathogenicity of B. cinerea have been characterized. Accordingly, pathogenicity mechanisms and virulence factors were discovered, whereby B. cinerea suppresses plant defense to establish host infection and invasion. Degradation of plant tissues is achieved via the production of acids, ROS, CWDEs and compound appressoria and inducing plant cell death.
With almost 11,000 genes encoded in its genome, only less than 150 genes have been genetically studied so far through mutant analysis. This explains our limited understanding of this infamous fungus and the lack of knowledge about the pathways of its unique biological mechanisms. Besides the traditional targeted gene disruption methods, recent innovations in genetic analysis approaches such as CRISPR genome editing [162], should be utilized to enhance studies on this important pathogen.
Due to the interconnectivity of the signaling pathways in B. cinerea, it has been challenging to identify the specific functions of virulence factors with mutant analysis. Future improvements in genetic and biochemical tools for B. cinerea specific studies may aid researchers to unveil the mysteries of this fungus. Unfortunately, because of the complex pathogenicity mechanisms of B. cinerea, generating resistant cultivars may be sophisticated without causing unwanted side effects. Likewise, the high genomic diversity makes it difficult to develop effective long-term fungicide control methods. Further collaboration between molecular pathologists and resistance breeders will be crucial to understand the pathogenic mechanisms and host resistance behind this pathogen.
Funding
This research was funded by Natural Sciences and Engineering Research Council of Canada (NSERC)-Discovery and NSERC-CREATE-PRoTECT programs.
Acknowledgments
The authors apologize for literatures not cited due to space limit. This work is supported by funds from NSERC-Discovery and CREATE-PRoTECT programs to XL. NC is partly supported by an NSERC-USRA fellowship and LT is funded in part through a scholarship from CSC (Chinese scholarship council).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Persoon, C.H. Dispositio methodica fungorum. Neues Mag. Bot. 1794, 1, 81–128. Available online: http://www.mycobank.org/BioloMICS.aspx?TableKey=14682616000000061&Rec=16179&Fields=All (accessed on 7 May 2020).
- Mirzaei, S.; Goltapeh, E.M.; Shams-Bakhsh, M.; Safaie, N. Identification of Botrytis spp. on plants grown in Iran. J. Phytopathol. 2007, 156, 21–28. [Google Scholar] [CrossRef]
- Gregory, P. Studies on Sclerotinia and Botrytis. Trans. Br. Mycol. Soc. 1949, 32, 1–IN3. [Google Scholar] [CrossRef]
- Whetzel, H.H. A Synopsis of the genera and species of the Sclerotiniaceae, a family of stromatic inoperculate discomycetes. Mycologia 1945, 37, 648. [Google Scholar] [CrossRef]
- Zhong, S.; Zhang, J.; Zhang, G.-Z. Botrytis polyphyllae: A new botrytis species causing gray mold on Paris polyphylla. Plant Dis. 2019, 103, 1721–1727. [Google Scholar] [CrossRef] [PubMed]
- Grant-Downton, R.T.; Terhem, R.B.; Kapralov, M.V.; Mehdi, S.; Rodriguez-Enriquez, M.J.; Gurr, S.J.; Van Kan, J.A.L.; Dewey, F.M. A novel Botrytis species is associated with a newly emergent foliar disease in cultivated hemerocallis. PLoS ONE 2014, 9, e89272. [Google Scholar] [CrossRef] [PubMed]
- Staats, M.; Van Baarlen, P.; Van Kan, J.A.L. Molecular phylogeny of the plant pathogenic genus Botrytis and the evolution of host specificity. Mol. Biol. Evol. 2004, 22, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Amselem, J.; Cuomo, C.A.; Van Kan, J.A.L.; Viaud, M.; Benito, E.P.; Couloux, A.; Coutinho, P.M.; De Vries, R.P.; Dyer, P.S.; Fillinger, S.; et al. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet. 2011, 7, e1002230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, W.R. The infection of strawberry and raspberry fruits by Botrytis cinerea Fr. Ann. Appl. Biol. 1962, 50, 569–575. [Google Scholar] [CrossRef]
- Viret, O.; Keller, M.; Jaudzems, V.G.; Cole, F.M. Botrytis cinerea infection of grape flowers: Light and electron microscopical studies of infection sites. Phytopathology 2004, 94, 850–857. [Google Scholar] [CrossRef] [Green Version]
- McNicol, R.J.; Williamson, B.; Dolan, A. Infection of red raspberry styles and carpels by Botrytis cinerea and its possible role in post-harvest grey mould. Ann. Appl. Biol. 1985, 106, 49–53. [Google Scholar] [CrossRef]
- Neri, F.; Cappellin, L.; Spadoni, A.; Cameldi, I.; Alarcon, A.A.; Aprea, E.; Romano, A.; Gasperi, F.; Biasioli, F. Role of strawberry volatile organic compounds in the development of Botrytis cinerea infection. Plant Pathol. 2014, 64, 709–717. [Google Scholar] [CrossRef]
- Prusky, D.; Lichter, A. Activation of quiescent infections by postharvest pathogens during transition from the biotrophic to the necrotrophic stage. FEMS Microbiol. Lett. 2007, 268, 1–8. [Google Scholar] [CrossRef]
- Gentile, A.C. Carbohydrate metabolism and oxalic acid synthesis by Botrytis cinerea. Plant Physiol. 1954, 29, 257–261. [Google Scholar] [CrossRef]
- Sasanuma, I.; Suzuki, T. Effect of calcium on cell-wall degrading enzymes of Botrytis cinerea. Biosci. Biotechnol. Biochem. 2016, 80, 1730–1736. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.L. Postharvest fruit rots in d’Anjou pears caused by Botrytis cinerea, Potebniamyces pyri, and Sphaeropsis pyriputrescens. Plant Heal. Prog. 2006, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Araújo, A.E.; Maffia, L.A.; Mizubuti, E.S.G.; Alfenas, A.C.; De Capdeville, G.; Grossi, J.A.S. Survival of Botrytis cinerea as mycelium in rose crop debris and as sclerotia in soil. Fitopatol. Bras. 2005, 30, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Backhouse, D.; Willetts, H.J. A histochemical study of sclerotia of Botrytis cinerea and Botrytis fabae. Can. J. Microbiol. 1984, 30, 171–178. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Medina, V.; Alonso, A.; Ayllón, M.A. Mycoviruses of Botrytis cinerea isolates from different hosts. Ann. Appl. Biol. 2013, 164, 46–61. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking Host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Qushim, B.; Wu, F.; Guan, Z.; Peres, N. The economic impact of botrytis fruit rot on strawberry production in Florida. In Proceedings of the 2018 Annual Meeting, Jacksonville, FL, USA, 2–6 February 2018. [Google Scholar]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Nishimoto, R. Global trends in the crop protection industry. J. Pestic. Sci. 2019, 44, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Katan, T. Resistance to 3,5-dichlorophenyl-N-cyclic imide (’dicarboximide’) fungicides in the grey mould pathogen Botrytis cinerea on protected crops. Plant Pathol. 1982, 31, 133–141. [Google Scholar] [CrossRef]
- Gubler, W.D. Control of Botrytis bunch rot of grape with canopy management. Plant Dis. 1987, 71, 599. [Google Scholar] [CrossRef]
- Naegele, R.P. Evaluation of host resistance to Botrytis bunch rot in Vitis spp. and its correlation with Botrytis leaf spot. HortScience 2018, 53, 204–207. [Google Scholar] [CrossRef]
- Kan, J.A.L.V.; Stassen, J.H.M.; Mosbach, A.; Van Der Lee, T.A.J.; Faino, L.; Farmer, A.D.; Papasotiriou, D.G.; Zhou, S.; Seidl, M.F.; Cottam, E.; et al. A gapless genome sequence of the fungus Botrytis cinerea. Mol. Plant Pathol. 2016, 18, 75–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, W.; Zhang, Y.; Wang, J.; Lv, C.; Chen, C. BcMtg2 is required for multiple stress tolerance, vegetative development and virulence in Botrytis cinerea. Sci. Rep. 2016, 6, 28673. [Google Scholar] [CrossRef] [PubMed]
- Staats, M.; Van Kan, J.A.L. Genome update of Botrytis cinerea Strains B05.10 and T4. Eukaryot. Cell 2012, 11, 1413–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porquier, A.; Moraga, J.; Morgant, G.; Dalmais, B.; Simon, A.; Sghyer, H.; Collado, I.G.; Viaud, M. Botcinic acid biosynthesis in Botrytis cinerea relies on a subtelomeric gene cluster surrounded by relics of transposons and is regulated by the Zn2Cys6 transcription factor BcBoa13. Curr. Genet. 2019, 65, 965–980. [Google Scholar] [CrossRef]
- Bokor, A.A.; Van Kan, J.A.; Poulter, R.T. Sexual mating of Botrytis cinerea illustrates PRP8 intein HEG activity. Fungal Genet. Biol. 2010, 47, 392–398. [Google Scholar] [CrossRef]
- Blanco-Ulate, B.; Morales-Cruz, A.; Amrine, K.C.H.; Labavitch, J.M.; Powell, A.L.T.; Cantu, D. Genome-wide transcriptional profiling of Botrytis cinerea genes targeting plant cell walls during infections of different hosts. Front. Plant Sci. 2014, 5, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwell, S.; Corwin, J.A.; Soltis, N.E.; Subedy, A.; Denby, K.; Kliebenstein, D.J. Whole genome resequencing of Botrytis cinerea isolates identifies high levels of standing diversity. Front. Microbiol. 2015, 6, 996. [Google Scholar] [CrossRef] [Green Version]
- Lamb, B.C.; Mandaokar, S.; Bahsoun, B.; Grishkan, I.; Nevo, E. Differences in spontaneous mutation frequencies as a function of environmental stress in soil fungi at “Evolution Canyon,” Israel. Proc. Natl. Acad. Sci. USA 2008, 105, 5792–5796. [Google Scholar] [CrossRef] [Green Version]
- Leroch, M.; Kleber, A.; Silva, E.; Coenen, T.; Koppenhöfer, D.; Shmaryahu, A.; Valenzuela, P.D.T.; Hahn, M. Transcriptome profiling of Botrytis cinerea conidial germination reveals upregulation of infection-related genes during the prepenetration stage. Eukaryot. Cell 2013, 12, 614–626. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Corwin, J.A.; Copeland, D.; Feusier, J.; Eshbaugh, R.; Cook, D.E.; Atwell, S.; Kliebenstein, D.J. Plant-necrotroph co-transcriptome networks illuminate a metabolic battlefield. eLife 2019, 8. [Google Scholar] [CrossRef]
- Choi, J.; Park, J.; Kim, D.; Jung, K.; Kang, S.; Lee, Y.-H. Fungal secretome database: Integrated platform for annotation of fungal secretomes. BMC Genom. 2010, 11, 105. [Google Scholar] [CrossRef] [Green Version]
- González-Fernández, R.; Valero-Galván, J.; Gómez-Gálvez, F.J.; Jorrín-Novo, J.V. Unraveling the in vitro secretome of the phytopathogen Botrytis cinerea to understand the interaction with its hosts. Front. Plant Sci. 2015, 6, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harren, K.; Tudzynski, B. Cch1 and Mid1 are functionally required for vegetative growth under low-calcium conditions in the phytopathogenic ascomycete Botrytis cinerea. Eukaryot. Cell 2013, 12, 712–724. [Google Scholar] [CrossRef]
- Nguyen, Q.B.; Kadotani, N.; Kasahara, S.; Tosa, Y.; Mayama, S.; Nakayashiki, H. Systematic functional analysis of calcium-signalling proteins in the genome of the rice-blast fungus, Magnaporthe oryzae, using a high-throughput RNA-silencing system. Mol. Microbiol. 2008, 68, 1348–1365. [Google Scholar] [CrossRef]
- Bormann, J.; Tudzynski, P. Deletion of Mid1, a putative stretch-activated calcium channel in Claviceps purpurea, affects vegetative growth, cell wall synthesis and virulence. Microbiology 2009, 155, 3922–3933. [Google Scholar] [CrossRef] [Green Version]
- Gronover, C.S.; Kasulke, D.; Tudzynski, P.; Tudzynski, B. The role of G protein alpha subunits in the infection process of the gray mold fungus Botrytis cinerea. Mol. Plant-Microbe Interact. 2001, 14, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Dean, R.A. G Protein α subunit genes control growth, development, and pathogenicity of Magnaporthe grisea. Mol. Plant-Microbe Interact. 1997, 10, 1075–1086. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Qin, G.; Li, B.; Tian, S. Knocking out Bcsas1 in Botrytis cinerea impacts growth, development, and secretion of extracellular proteins, which decreases virulence. Mol. Plant-Microbe Interact. 2014, 27, 590–600. [Google Scholar] [CrossRef]
- Dub, A.M.; Kokkelink, L.; Tudzynski, B.; Tudzynski, P.; Sharon, A. Involvement of Botrytis cinerea small GTPases BcRAS1 and BcRAC in differentiation, virulence, and the cell cycle. Eukaryot. Cell 2013, 12, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Müller, N.; Leroch, M.; Schumacher, J.; Zimmer, D.; Könnel, A.; Klug, K.; Leisen, T.; Scheuring, D.; Sommer, F.K.; Mühlhaus, T.; et al. Investigations on VELVET regulatory mutants confirm the role of host tissue acidification and secretion of proteins in the pathogenesis of Botrytis cinerea. New Phytol. 2018, 219, 1062–1074. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Chen, Y.; Ma, Z. Involvement of BcVeA and BcVelB in regulating conidiation, pigmentation and virulence in Botrytis cinerea. Fungal Genet. Biol. 2013, 50, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Sharma, E.; Tayal, P.; Anand, G.; Mathur, P.; Kapoor, R. Functional analysis of diacylglycerol O-acyl transferase 2 gene to decipher its role in virulence of Botrytis cinerea. Curr. Genet. 2017, 64, 443–457. [Google Scholar] [CrossRef]
- Morcx, S.; Kunz, C.; Choquer, M.; Assié, S.; Blondet, E.; Simond-Côte, E.; Gajek, K.; Chapeland-Leclerc, F.; Expert, D.; Soulie, M. Disruption of Bcchs4, Bcchs6 or Bcchs7 chitin synthase genes in Botrytis cinerea and the essential role of class VI chitin synthase (Bcchs6). Fungal Genet. Biol. 2013, 52, 1–8. [Google Scholar] [CrossRef]
- Yin, Y.; Wu, S.; Chui, C.; Ma, T.; Jiang, H.; Hahn, M.; Ma, Z. The MAPK kinase BcMkk1 suppresses oxalic acid biosynthesis via impeding phosphorylation of BcRim15 by BcSch9 in Botrytis cinerea. PLoS Pathog. 2018, 14, e1007285. [Google Scholar] [CrossRef]
- Viefhues, A.; Heller, J.; Temme, N.; Tudzynski, P. Redox systems in Botrytis cinerea: Impact on development and virulence. Mol. Plant-Microbe Interact. 2014, 27, 858–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.M.; Van Kan, J.A.L.; Bailey, A.M.; Foster, G. Inadvertent gene silencing of argininosuccinate synthase (bcass1) in Botrytis cinerea by the pLOB1 vector system. Mol. Plant Pathol. 2010, 11, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J.; De Larrinoa, I.F.; Tudzynski, B. Calcineurin-responsive zinc finger transcription factor CRZ1 of Botrytis cinerea is required for growth, development, and full virulence on bean plants. Eukaryot. Cell 2008, 7, 584–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegmund, U.; Marschall, R.; Tudzynski, P. BcNoxD, a putative ER protein, is a new component of the NADPH oxidase complex in Botrytis cinerea. Mol. Microbiol. 2014, 95, 988–1005. [Google Scholar] [CrossRef]
- Zhang, L.; Van Kan, J.A.L. Botrytis cinereamutants deficient ind-galacturonic acid catabolism have a perturbed virulence on Nicotiana benthamiana and Arabidopsis, but not on tomato. Mol. Plant Pathol. 2012, 14, 19–29. [Google Scholar] [CrossRef]
- Hou, J.; Feng, H.; Chang, H.; Liu, Y.; Li, G.; Yang, S.; Sun, C.; Zhang, M.; Yuan, Y.; Sun, J.; et al. The H3K4 demethylase Jar1 orchestrates ROS production and expression of pathogenesis-related genes to facilitate Botrytis cinerea virulence. New Phytol. 2019, 225, 930–947. [Google Scholar] [CrossRef]
- Harren, K.; Schumacher, J.; Tudzynski, B. The Ca2+/calcineurin-dependent signaling pathway in the gray mold Botrytis cinerea: The role of calcipress in in modulating calcineurin activity. PLoS ONE 2012, 7, e41761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Liu, N.; Sang, C.; Shi, D.; Zhou, M.; Chen, C.-J.; Qin, Q.; Chen, W. The autophagy gene BcATG8 regulates the vegetative differentiation and pathogenicity of Botrytis cinerea. Appl. Environ. Microbiol. 2018, 84, e02455-17. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J. DHN melanin biosynthesis in the plant pathogenic fungus Botrytis cinerea is based on two developmentally regulated key enzyme (PKS)-encoding genes. Mol. Microbiol. 2015, 99, 729–748. [Google Scholar] [CrossRef] [Green Version]
- Segmüller, N.; Kokkelink, L.; Giesbert, S.; Odinius, D.; Van Kan, J.; Tudzynski, P. NADPH oxidases are involved in differentiation and pathogenicity in Botrytis cinerea. Mol. Plant-Microbe Interact. 2008, 21, 808–819. [Google Scholar] [CrossRef] [Green Version]
- Shao, W.; Yang, Y.; Zhang, Y.; Lv, C.; Ren, W.; Chen, C. Involvement of BcStr2 in methionine biosynthesis, vegetative differentiation, multiple stress tolerance and virulence in Botrytis cinerea. Mol. Plant Pathol. 2015, 17, 438–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Ren, W.; Li, F.; Chen, C.-J.; Ma, Z. Involvement of the cysteine protease BcAtg4 in development and virulence of Botrytis cinerea. Curr. Genet. 2018, 65, 293–300. [Google Scholar] [CrossRef]
- Nafisi, M.; Stranne, M.; Zhang, L.; Van Kan, J.A.L.; Sakuragi, Y. The endo-arabinanase BcAra1 is a novel host-specific virulence factor of the necrotic fungal phytopathogen Botrytis cinerea. Mol. Plant-Microbe Interact. 2014, 27, 781–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terhem, R.B.; Van Kan, J.A.L. Functional analysis of hydrophobin genes in sexual development of Botrytis cinerea. Fungal Genet. Biol. 2014, 71, 42–51. [Google Scholar] [CrossRef]
- Mosbach, A.; Leroch, M.; Mendgen, K.; Hahn, M. Lack of evidence for a role of hydrophobins in conferring surface hydrophobicity to conidia and hyphae of Botrytis cinerea. BMC Microbiol. 2011, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, H.; Qin, G.; He, C.; Li, B.; Tian, S. The MADS-Box transcription factor Bcmads1 is required for growth, sclerotia production and pathogenicity of Botrytis cinerea. Sci. Rep. 2016, 6, 33901. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yin, D.; Yin, Y.; Cao, Y.; Ma, Z. The response regulator BcSkn7 is required for vegetative differentiation and adaptation to oxidative and osmotic stresses in Botrytis cinerea. Mol. Plant Pathol. 2014, 16, 276–287. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, L.; Wu, M.; Chen, W.; Li, G.; Zhang, J. A single-nucleotide deletion in the transcription factor gene bcsmr1 causes sclerotial-melanogenesis deficiency in Botrytis cinerea. Front. Microbiol. 2017, 8, 2492. [Google Scholar] [CrossRef]
- Frías, M.; González, C.; Brito, N. BcSpl1, a cerato-platanin family protein, contributes to Botrytis cinerea virulence and elicits the hypersensitive response in the host. New Phytol. 2011, 192, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Jiang, J.; Mayr, C.; Hahn, M.; Ma, Z. Involvement of two type 2C protein phosphatases BcPtc1 and BcPtc3 in the regulation of multiple stress tolerance and virulence of Botrytis cinerea. Environ. Microbiol. 2013, 15, 2696–2711. [Google Scholar] [CrossRef] [PubMed]
- Minz-Dub, A.; Sharon, A. The Botrytis cinerea PAK kinase BcCla4 mediates morphogenesis, growth and cell cycle regulating processes downstream of BcRac. Mol. Microbiol. 2017, 104, 487–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Salas, O.; Bowler, K.; Oren-Young, L.; Bar-Peled, M.; Sharon, A. Genetic alteration of UDP-rhamnose metabolism in Botrytis cinerealeads to the accumulation of UDP-KDG that adversely affects development and pathogenicity. Mol. Plant Pathol. 2016, 18, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Rolke, Y.; Liu, S.; Quidde, T.; Williamson, B.; Schouten, A.; Weltring, K.-M.; Siewers, V.; Tenberge, K.B.; Tudzynski, B.; Tudzynski, P. Functional analysis of H2O2-generating systems in Botrytis cinerea: The major Cu-Zn-superoxide dismutase (BCSOD1) contributes to virulence on French bean, whereas a glucose oxidase (BCGOD1) is dispensable. Mol. Plant Pathol. 2004, 5, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Tayal, P.; Raj, S.; Sharma, E.; Kumar, M.; Dayaman, V.; Verma, N.; Jogawat, A.; Dua, M.; Kapoor, R.; Johri, A.K. A Botrytis cinerea KLP-7 kinesin acts as a virulence determinant during plant infection. Sci. Rep. 2017, 7, 10664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Zhang, J.; Hu, J.; Wang, X.; Lv, B.; Liang, W. Involvement of BcYak1 in the regulation of vegetative differentiation and adaptation to oxidative stress of Botrytis cinerea. Front. Microbiol. 2018, 9, 281. [Google Scholar] [CrossRef] [Green Version]
- Brandhoff, B.; Simon, A.; Dornieden, A.; Schumacher, J. Regulation of conidiation in Botrytis cinerea involves the light-responsive transcriptional regulators BcLTF3 and BcREG1. Curr. Genet. 2017, 63, 931–949. [Google Scholar] [CrossRef]
- Guan, W.; Feng, J.; Wang, R.; Ma, Z.; Wang, W.-X.; Wang, K.; Zhu, T. Functional analysis of the exocyst subunit BcExo70 in Botrytis cinerea. Curr. Genet. 2019, 66, 85–95. [Google Scholar] [CrossRef]
- Zhang, C.; He, Y.; Zhu, P.; Chen, L.; Wang, Y.; Ni, B.; Xu, L. Loss of bcbrn1 and bcpks13 in Botrytis cinerea not only blocks melanization but also increases vegetative growth and virulence. Mol. Plant-Microbe Interact. 2015, 28, 1091–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, J.; Kokkelink, L.; Huesmann, C.; Jimenez-Teja, D.; Collado, I.G.; Barakat, R.; Tudzynski, P.; Tudzynski, B. The cAMP-dependent signaling pathway and its role in conidial germination, growth, and virulence of the gray mold Botrytis cinerea. Mol. Plant-Microbe Interact. 2008, 21, 1443–1459. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Li, G.; Du, S.; Yang, S.; Li, X.-Q.; De Figueiredo, P.; Qin, Q.-M. The septin protein Sep4 facilitates host infection by plant fungal pathogens via mediating initiation of infection structure formation. Environ. Microbiol. 2017, 19, 1730–1749. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, Y.; Izumitsu, K.; Morita, A.; Tanaka, C. A copper-transporting ATPase BcCCC2 is necessary for pathogenicity of Botrytis cinerea. Mol. Genet. Genom. 2010, 284, 33–43. [Google Scholar] [CrossRef]
- Giesbert, S.; Siegmund, U.; Schumacher, J.; Kokkelink, L.; Tudzynski, P. Functional analysis of BcBem1 and its interaction partners in Botrytis cinerea: Impact on differentiation and virulence. PLoS ONE 2014, 9, e95172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaud, M.; Brunet-Simon, A.; Brygoo, Y.; Pradier, J.-M.; Levis, C. Cyclophilin A and calcineurin functions investigated by gene inactivation, cyclosporin A inhibition and cDNA arrays approaches in the phytopathogenic fungus Botrytis cinerea. Mol. Microbiol. 2003, 50, 1451–1465. [Google Scholar] [CrossRef] [PubMed]
- Marschall, R.; Tudzynski, P. BcIqg1, a fungal IQGAP homolog, interacts with NADPH oxidase, MAP kinase and calcium signaling proteins and regulates virulence and development in Botrytis cinerea. Mol. Microbiol. 2016, 101, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Vela-Corcía, D.; Srivastava, D.A.; Dafa-Berger, A.; Rotem, N.; Barda, O.; Levy, M. MFS transporter from Botrytis cinerea provides tolerance to glucosinolate-breakdown products and is required for pathogenicity. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, J.; Li, G.; Zhang, M.; Zhang, Y.; Wang, Y.; Hou, J.; Yang, S.; Sun, J.; Qin, Q. A novel Botrytis cinerea- specific gene BcHBF1 enhances virulence of the grey mould fungus via promoting host penetration and invasive hyphal development. Mol. Plant Pathol. 2019, 20, 731–747. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Wu, J.; Jamieson, P.A.; Zhang, C.Q. Alternative oxidase is involved in the pathogenicity, development, and oxygen stress response of Botrytis cinerea. Phytopathology 2019, 109, 1679–1688. [Google Scholar] [CrossRef]
- Choquer, M.; Robin, G.; Le PãªCheur, P.; Giraud, C.; Levis, C.; Viaud, M. Ku70orKu80deficiencies in the fungus Botrytis cinerea facilitate targeting of genes that are hard to knock out in a wild-type context. FEMS Microbiol. Lett. 2008, 289, 225–232. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Li, B.; Qin, G.; Tian, S. Function of small GTPase Rho3 in regulating growth, conidiation and virulence of Botrytis cinerea. Fungal Genet. Biol. 2015, 75, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hernández, A.; González, M.; González, C.; Van Kan, J.A.L.; Brito, N. BcSUN1, a B. cinerea SUN-Family Protein, Is Involved in Virulence. Front. Microbiol. 2017, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Zhang, Z.; Shao, W.; Yang, Y.; Zhou, M.; Chen, C.-J. The autophagy-related geneBcATG1is involved in fungal development and pathogenesis in Botrytis cinerea. Mol. Plant Pathol. 2016, 18, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Plaza, V.; Lagües, Y.; Carvajal, M.; Pérez-García, L.A.; Mora-Montes, H.M.; Canessa, P.; Larrondo, L.F.; Castillo, L. bcpmr1 encodes a P-type Ca2+/Mn2+-ATPase mediating cell-wall integrity and virulence in the phytopathogen Botrytis cinerea. Fungal Genet. Biol. 2015, 76, 36–46. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Z.; He, C.; Qin, G.; Tian, S. Comparative proteomics reveals the potential targets of BcNoxR, a putative regulatory subunit of NADPH oxidase of Botrytis cinerea. Mol. Plant-Microbe Interact. 2016, 29, 990–1003. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J.; Studt, L.; Tudzynski, P. The putative H3K36 demethylase BcKDM1 affects virulence, stress responses and photomorphogenesis in Botrytis cinerea. Fungal Genet. Biol. 2019, 123, 14–24. [Google Scholar] [CrossRef]
- Shlezinger, N.; Doron, A.; Sharon, A. Apoptosis-like programmed cell death in the grey mould fungus Botrytis cinerea: Genes and their role in pathogenicity. Biochem. Soc. Trans. 2011, 39, 1493–1498. [Google Scholar] [CrossRef]
- An, B.; Li, B.; Li, H.; Zhang, Z.; Qin, G.; Tian, S. Aquaporin8 regulates cellular development and reactive oxygen species production, a critical component of virulence in Botrytis cinerea. New Phytol. 2015, 209, 1668–1680. [Google Scholar] [CrossRef]
- Harren, K.; Brandhoff, B.; Knödler, M.; Tudzynski, B. The high-affinity phosphodiesterase BcPde2 has impact on growth, differentiation and virulence of the phytopathogenic ascomycete Botrytis cinerea. PLoS ONE 2013, 8, e78525. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Yu, F.; Yin, Y.; Ma, Z. Involvement of protein tyrosine phosphatases BcPtpA and BcPtpB in regulation of vegetative development, virulence and multi-stress tolerance in Botrytis cinerea. PLoS ONE 2013, 8, e61307. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Yang, Q.; Jiang, J.; Michailides, T.J.; Ma, Z. Involvement of a putative response regulator Brrg-1 in the regulation of sporulation, sensitivity to fungicides, and osmotic stress in Botrytis cinerea. Appl. Microbiol. Biotechnol. 2010, 90, 215–226. [Google Scholar] [CrossRef]
- Valette-Collet, O.; Cimerman, A.; Reignault, P.; Levis, C.; Boccara, M. Disruption of Botrytis cinerea pectin methylesterase gene Bcpme1 reduces virulence on several host plants. Mol. Plant-Microbe Interact. 2003, 16, 360–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xie, J.; Fu, Y.; Jiāng, D.; Chen, T.; Cheng, J. The subtilisin-like protease Bcser2 Affects the sclerotial formation, conidiation and virulence of Botrytis cinerea. Int. J. Mol. Sci. 2020, 21, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Sang, C.; Shi, D.; Song, X.; Zhou, M.; Chen, C.-J. Ubiquitin-like activating enzymes BcAtg3 and BcAtg7 participate in development and pathogenesis of Botrytis cinerea. Curr. Genet. 2018, 64, 919–930. [Google Scholar] [CrossRef]
- Ren, W.; Liu, N.; Yang, Y.; Yang, Q.; Chen, C.-J.; Gao, Q. The sensor proteins BcSho1 and BcSln1 are involved in, though not essential to, vegetative differentiation, pathogenicity and osmotic stress tolerance in Botrytis cinerea. Front. Microbiol. 2019, 10, 328. [Google Scholar] [CrossRef]
- Antal, Z.; Rascle, C.; Cimerman, A.; Viaud, M.; Billon-Grand, G.; Choquer, M.; Bruel, C. The homeobox BcHOX8 gene in Botrytis cinerea regulates vegetative growth and morphology. PLoS ONE 2012, 7, e48134. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, X.; Dong, Y.; Qiu, D. The Botrytis cinerea Xylanase BcXyl1 modulates plant immunity. Front. Microbiol. 2018, 9, 2535. [Google Scholar] [CrossRef] [Green Version]
- Rui, O.; Hahn, M. The Slt2-type MAP kinase Bmp3 of Botrytis cinerea is required for normal saprotrophic growth, conidiation, plant surface sensing and host tissue colonization. Mol. Plant Pathol. 2007, 8, 173–184. [Google Scholar] [CrossRef]
- Jonkers, W.; Van Kan, J.A.L.; Tijm, P.; Lee, Y.-W.; Tudzynski, P.; Rep, M.; Michielse, C.B. The FRP1 F-box gene has different functions in sexuality, pathogenicity and metabolism in three fungal pathogens. Mol. Plant Pathol. 2011, 12, 548–563. [Google Scholar] [CrossRef]
- Rui, O.; Hahn, M. The Botrytis cinerea hexokinase, Hxk1, but not the glucokinase, Glk1, is required for normal growth and sugar metabolism, and for pathogenicity on fruits. Microbiol. 2007, 153, 2791–2802. [Google Scholar] [CrossRef] [Green Version]
- Siegmund, U.; Heller, J.; Van Kan, J.A.L.; Tudzynski, P. The NADPH oxidase complexes in Botrytis cinerea: Evidence for a close association with the ER and the Tetraspanin Pls1. PLoS ONE 2013, 8, e55879. [Google Scholar] [CrossRef]
- Reis, H.; Pfiffi, S.; Hahn, M. Molecular and functional characterization of a secreted lipase from Botrytis cinerea. Mol. Plant Pathol. 2005, 6, 257–267. [Google Scholar] [CrossRef]
- Yan, L.; Yang, Q.Q.; Sundin, G.W.; Li, H.; Ma, Z. The mitogen-activated protein kinase kinase BOS5 is involved in regulating vegetative differentiation and virulence in Botrytis cinerea. Fungal Genet. Biol. 2010, 47, 753–760. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Dickman, M.B.; Wang, Z. Cytoprotective co-chaperone BcBAG1 is a component for fungal development, virulence, and Unfolded Protein Response (UPR) of Botrytis cinerea. Front. Microbiol. 2019, 10, 685. [Google Scholar] [CrossRef]
- Zhu, W.; Wei, W.; Wu, Y.; Zhou, Y.; Peng, F.; Zhang, S.; Chen, P.; Xu, X. BcCFEM1, a CFEM domain-containing protein with putative GPI-anchored site, is involved in pathogenicity, conidial production, and stress tolerance in Botrytis cinerea. Front. Microbiol. 2017, 8, 1807. [Google Scholar] [CrossRef]
- Rascle, C.; Dieryckx, C.; Dupuy, J.-W.; Muszkieta, L.; Souibgui, E.; Droux, M.; Bruel, C.; Girard, V.; Poussereau, N. The pH regulator PacC: A host-dependent virulence factor in Botrytis cinerea. Environ. Microbiol. Rep. 2018, 10, 555–568. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Zhao, Y.; Cheng, J.; Xie, J.; Fu, Y.; Jiang, D.; Chen, T. Histone H3 lysine 9 methyltransferase DIM5 is required for the development and virulence of Botrytis cinerea. Front. Microbiol. 2016, 7, 1289. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Yan, L.; Gu, Q.; Ma, Z. The mitogen-activated protein kinase kinase kinase BcOs4 is required for vegetative differentiation and pathogenicity in Botrytis cinerea. Appl. Microbiol. Biotechnol. 2012, 96, 481–492. [Google Scholar] [CrossRef]
- Cui, Z.; Gao, N.; Wang, Q.; Ren, Y.; Wang, K.; Zhu, T. BcMctA, a putative monocarboxylate transporter, is required for pathogenicity in Botrytis cinerea. Curr. Genet. 2015, 61, 545–553. [Google Scholar] [CrossRef]
- Meléndez, H.G.; Billon-Grand, G.; Fevre, M.; Mey, G. Role of the Botrytis cinerea FKBP12 ortholog in pathogenic development and in sulfur regulation. Fungal Genet. Biol. 2009, 46, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Lv, C.; Zhang, Y.; Wang, J.; Chen, C.-J. Involvement of BcElp4 in vegetative development, various environmental stress response and virulence of Botrytis cinerea. Microb. Biotechnol. 2017, 10, 886–895. [Google Scholar] [CrossRef]
- Cao, S.-N.; Yuan, Y.; Qin, Y.; Zhang, M.-Z.; De Figueiredo, P.; Li, G.-H.; Qin, Q.-M. The pre-rRNA processing factor Nop53 regulates fungal development and pathogenesis via mediating production of reactive oxygen species. Environ. Microbiol. 2018, 20, 1531–1549. [Google Scholar] [CrossRef]
- Giesbert, S.; Schumacher, J.; Kupas, V.; Espino, J.; Segmüller, N.; Haeuser-Hahn, I.; Schreier, P.H.; Tudzynski, P. Identification of pathogenesis-associated genes by T-DNA–mediated insertional mutagenesis in Botrytis cinerea: A Type 2A phosphoprotein phosphatase and an SPT3 transcription factor have significant impact on virulence. Mol. Plant-Microbe Interact. 2012, 25, 481–495. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yuan, X.; Zang, J.; Wang, M.; Zhao, F.; Li, P.; Cao, H.; Han, J.; Xing, J.; Dong, J. The kynurenine 3-monooxygenase encoding gene, BcKMO, is involved in the growth, development, and pathogenicity of Botrytis cinerea. Front. Microbiol. 2018, 9, 1039. [Google Scholar] [CrossRef] [PubMed]
- Siewers, V.; Viaud, M.; Jimenez-Teja, D.; Collado, I.G.; Gronover, C.S.; Pradier, J.-M.; Tudzynski, B.; Tudzynski, P. Functional analysis of the cytochrome P450 monooxygenase gene bcbot1 of Botrytis cinerea indicates that botrydial is a strain-specific virulence factor. Mol. Plant-Microbe Interact. 2005, 18, 602–612. [Google Scholar] [CrossRef] [Green Version]
- Kokkelink, L.; Minz, A.; Al-Masri, M.; Giesbert, S.; Barakat, R.; Sharon, A.; Tudzynski, P. The small GTPase BcCdc42 affects nuclear division, germination and virulence of the gray mold fungus Botrytis cinerea. Fungal Genet. Biol. 2011, 48, 1012–1019. [Google Scholar] [CrossRef]
- González-Rodríguez, V.E.; Garrido, C.; Cantoral, J.M.; Schumacher, J. The F-actin capping protein is required for hyphal growth and full virulence but is dispensable for septum formation in Botrytis cinerea. Fungal Biol. 2016, 120, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Kars, I.; Krooshof, G.H.; Wagemakers, L.; Joosten, R.; Benen, J.A.; Van Kan, J.A. Necrotizing activity of five Botrytis cinerea endopolygalacturonases produced in Pichia pastoris. Plant J. 2005, 43, 213–225. [Google Scholar] [CrossRef]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Viaud, M.; Tudzynski, P. The transcription factor BcLTF1 regulates virulence and light responses in the necrotrophic plant pathogen Botrytis cinerea. PLoS Genet. 2014, 10, e1004040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimpel, A.; Gronover, C.S.; Williamson, B.; Stewart, J.A.; Tudzynski, B. The adenylate cyclase (BAC) in Botrytis cinerea is required for full pathogenicity. Mol. Plant Pathol. 2002, 3, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J.; Pradier, J.-M.; Simon, A.; Traeger, S.; Moraga, J.; Collado, I.G.; Viaud, M.; Tudzynski, B. Natural variation in the VELVET Gene bcvel1 affects virulence and light-dependent differentiation in Botrytis cinerea. PLoS ONE 2012, 7, e47840. [Google Scholar] [CrossRef] [Green Version]
- Heller, J.; Ruhnke, N.; Espino, J.J.; Massaroli, M.; Collado, I.G.; Tudzynski, P. The mitogen-activated protein kinase BcSak1 of Botrytis cinerea is required for pathogenic development and has broad regulatory functions beyond stress response. Mol. Plant-Microbe Interact. 2012, 25, 802–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-K.; Chang, H.-W.; Liu, Y.; Qin, Y.; Ding, Y.-H.; Wang, L.; Zhao, Y.; Zhang, M.-Z.; Cao, S.-N.; Li, L.-T.; et al. The key gluconeogenic gene PCK1 is crucial for virulence of Botrytis cinerea via initiating its conidial germination and host penetration. Environ. Microbiol. 2018, 20, 1794–1814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, C.; Liu, Y.; Feng, H.; Chang, H.; Cao, S.; Li, G.; Yang, S.; Hou, J.; Zhu-Salzman, K.; et al. Transcriptome analysis and functional validation reveal a novel gene, BcCGF1, that enhances fungal virulence by promoting infection-related development and host penetration. Mol. Plant Pathol. 2020, 21, 834–853. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, Z.; Qin, G.; He, C.; Li, B.; Tian, S. Actin is required for cellular development and virulence of Botrytis cinerea via the mediation of secretory proteins. mSystems 2020, 5, 00732-19. [Google Scholar] [CrossRef] [Green Version]
- García, N.; González, M.A.; González, C.; Brito, N. Simultaneous silencing of xylanase genes in Botrytis cinerea. Front. Plant Sci. 2017, 8, 2174. [Google Scholar] [CrossRef]
- Parsley, T.B.; Segers, G.C.; Nuss, D.L.; Dawe, A.L. Analysis of altered G-protein subunit accumulation in Cryphonectria parasitica reveals a third Gα homologue. Curr. Genet. 2003, 43, 24–33. [Google Scholar] [CrossRef]
- Krystofova, S.; Borkovich, K.A. The heterotrimeric G-protein subunits GNG-1 and GNB-1 Form a Gβγ dimer required for normal female fertility, asexual development, and Gα protein levels in Neurospora crassa. Eukaryot. Cell 2005, 4, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Jin, Y.; Wang, G. The role of SCF ubiquitin-ligase complex at the beginning of life. Reprod. Biol. Endocrinol. 2019, 17, 1–9. [Google Scholar] [CrossRef]
- Jonkers, W.; Rodrigues, C.D.A.; Rep, M. Impaired colonization and infection of tomato roots by the Δfrp1 mutant of Fusarium oxysporum correlates with reduced CWDE gene expression. Mol. Plant-Microbe Interact. 2009, 22, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Hunter, T. Protein kinases and phosphatases: The Yin and Yang of protein phosphorylation and signaling. Cell 1995, 80, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Armant, O.; Fischer, R. Fungi use the SakA (HogA) pathway for phytochrome-dependent light signalling. Nat. Microbiol. 2016, 1, 16019. [Google Scholar] [CrossRef]
- Yen, C.-L.E.; Stone, S.J.; Koliwad, S.; Harris, C.; Farese, R.V.F., Jr. Thematic review series: Glycerolipids.DGAT enzymes and triacylglycerol biosynthesis. J. Lipid Res. 2008, 49, 2283–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, A.E. Alternative oxidase: An inter-kingdom perspective on the function and regulation of this broadly distributed ’cyanide-resistant’ terminal oxidase. Funct. Plant Biol. 2008, 35, 535–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R.; Bingham, I. Influence of nutrition on disease development caused by fungal pathogens: Implications for plant disease control. Ann. Appl. Biol. 2007, 151, 307–324. [Google Scholar] [CrossRef]
- Michielse, C.B.; Becker, M.; Heller, J.; Moraga, J.; Collado, I.G.; Tudzynski, P.; Moraga, J. The Botrytis cinerea Reg1 protein, a putative transcriptional regulator, is required for pathogenicity, conidiogenesis, and the production of secondary metabolites. Mol. Plant-Microbe Interact. 2011, 24, 1074–1085. [Google Scholar] [CrossRef] [Green Version]
- Doss, R.P.; Potter, S.W.; Soeldner, A.H.; Christian, J.K.; Fukunaga, L.E. Adhesion of germlings of Botrytis cinerea. Appl. Environ. Microbiol. 1995, 61, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espino, J.J.; Gutiérrez-Sánchez, G.; Brito, N.; Shah, P.; Orlando, R.; González, C. The Botrytis cinerea early secretome. Proteomics 2010, 10, 3020–3034. [Google Scholar] [CrossRef] [Green Version]
- González, M.; Brito, N.; González, C. Identification of glycoproteins secreted by wild-type Botrytis cinerea and by protein O-mannosyltransferase mutants. BMC Microbiol. 2014, 14, 254. [Google Scholar] [CrossRef] [Green Version]
- Brito, N.; Espino, J.J.; González, C. The endo-β-1,4-xylanase Xyn11A is required for virulence in Botrytis cinerea. Mol. Plant-Microbe Interact. 2006, 19, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, R.D.; Kelkar, H.S.; Dean, R.A. An eight-cysteine-containing CFEM domain unique to a group of fungal membrane proteins. Trends Biochem. Sci. 2003, 28, 118–121. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, J.-J.; Ying, S.; Feng, M.-G. Wee1 and Cdc25 control morphogenesis, virulence and multistress tolerance of Beauveria bassiana by balancing cell cycle-required cyclin-dependent kinase 1 activity. Environ. Microbiol. 2014, 17, 1119–1133. [Google Scholar] [CrossRef]
- Laprade, L.; Boyartchuk, V.L.; Dietrich, W.F.; Winston, F. Spt3 plays opposite roles in filamentous growth in Saccharomyces cerevisiae and Candida albicans and is required for C. albicans virulence. Genetics 2002, 161, 509–519. [Google Scholar]
- Michielse, C.B.; Van Wijk, R.; Reijnen, L.; Cornelissen, B.J.C.; Rep, M. Insight into the molecular requirements for pathogenicity of Fusarium oxysporum f. sp. lycopersici through large-scale insertional mutagenesis. Genome Biol. 2009, 10, R4. [Google Scholar] [CrossRef] [Green Version]
- Donofrio, N.; Oh, Y.; Lundy, R.; Pan, H.; Brown, D.; Jeong, J.; Coughlan, S.; Mitchell, T.; Dean, R. Global gene expression during nitrogen starvation in the rice blast fungus, Magnaporthe grisea. Fungal Genet. Biol. 2006, 43, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Adamo, J.E.; Moskow, J.J.; Gladfelter, A.S.; Viterbo, D.; Lew, D.J.; Brennwald, P.J. Yeast Cdc42 functions at a late step in exocytosis, specifically during polarized growth of the emerging bud. J. Cell Biol. 2001, 155, 581–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doss, R.P.; Deisenhofer, J.; Von Nidda, H.-A.K.; Soeldner, A.H.; McGuire, R.P. Melanin in the extracellular matrix of germlings of Botrytis cinerea. Phytochemistry 2003, 63, 687–691. [Google Scholar] [CrossRef]
- Liu, W.; Soulié, M.-C.; Perrino, C.; Fillinger, S. The osmosensing signal transduction pathway from Botrytis cinerea regulates cell wall integrity and MAP kinase pathways control melanin biosynthesis with influence of light. Fungal Genet. Biol. 2011, 48, 377–387. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, N.; Yang, J.; Yang, L.; Wu, M.; Chen, W.; Li, G.; Zhang, J. Contrast between orange-and black-colored sclerotial isolates of Botrytis cinerea: Melanogenesis and ecological fitness. Plant. Dis. 2018, 102, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Traeger, S.; Porquier, A.; Dalmais, B.; Viaud, M.; Tudzynski, B. The VELVET complex in the gray mold fungus Botrytis cinerea: Impact of BcLAE1 on differentiation, secondary metabolism, and virulence. Mol. Plant.-Microbe Interact. 2015, 28, 659–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leisen, T.; Bietz, F.; Werner, J.; Wegner, A.; Schaffrath, U.; Scheuring, D.; Willmund, F.; Mosbach, A.; Scalliet, G.; Hahn, M. CRISPR/Cas with ribonucleoprotein complexes and transiently selected telomere vectors allows highly efficient marker-free and multiple genome editing in Botrytis cinerea. PLoS Pathog. 2020, 16, e1008326. [Google Scholar] [CrossRef]
Figure 1.
Life cycle of Botrytis cinerea.

Figure 2.
A Venn Diagram summary of B. cinerea genes studied with mutant analysis (detailed in Table 1).
Figure 2.
A Venn Diagram summary of B. cinerea genes studied with mutant analysis (detailed in Table 1).

Figure 3.
Map positions of the genes studied in the B. cinerea genome through mutant analysis. The numbers on the left of each chromosome represent the locations of these genes. Genes labelled in green are involved in development, including hyphal growth, sclerotial formation, conidiation and so forth. Genes labelled in red mainly play roles in virulence. Genes labelled in blue are involved in both while the ones labelled in black are involved in other biological processes. The chromosomal map was drawn using ‘MapChart’ software using information from Table 1.
Figure 3.
Map positions of the genes studied in the B. cinerea genome through mutant analysis. The numbers on the left of each chromosome represent the locations of these genes. Genes labelled in green are involved in development, including hyphal growth, sclerotial formation, conidiation and so forth. Genes labelled in red mainly play roles in virulence. Genes labelled in blue are involved in both while the ones labelled in black are involved in other biological processes. The chromosomal map was drawn using ‘MapChart’ software using information from Table 1.

Figure 4.
Signaling pathways in B. cinerea (detailed in Table 1).
Figure 4.
Signaling pathways in B. cinerea (detailed in Table 1).

Figure 5.
Dihydroxynaphthalene (DHN) melanin synthesis pathway in B. cinerea.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cheung, N.; Tian, L.; Liu, X.; Li, X. The Destructive Fungal Pathogen Botrytis cinerea—Insights from Genes Studied with Mutant Analysis. Pathogens 2020, 9, 923. https://doi.org/10.3390/pathogens9110923
AMA Style
Cheung N, Tian L, Liu X, Li X. The Destructive Fungal Pathogen Botrytis cinerea—Insights from Genes Studied with Mutant Analysis. Pathogens. 2020; 9(11):923. https://doi.org/10.3390/pathogens9110923
Chicago/Turabian StyleCheung, Nicholas, Lei Tian, Xueru Liu, and Xin Li. 2020. "The Destructive Fungal Pathogen Botrytis cinerea—Insights from Genes Studied with Mutant Analysis" Pathogens 9, no. 11: 923. https://doi.org/10.3390/pathogens9110923
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.