
ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes
by
 , ,
, ,
Ricardo Franco-Duarte
1,2,*,† ,
,
Isabel Fernandes
1,2,†,
Vladislav Gulis
3,
Fernanda Cássio
1,2 and
Cláudia Pascoal
1,2 1
CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, 4710-057 Braga, Portugal
2
Institute of Science and Innovation for Bio-Sustainability (IB-S), University of Minho, 4710-057 Braga, Portugal
3
Department of Biology, Coastal Carolina University, Conway, SC 29528-6054, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Microorganisms 2022, 10(8), 1569; https://doi.org/10.3390/microorganisms10081569
Submission received: 12 July 2022
/
Revised: 1 August 2022
/
Accepted: 2 August 2022
/
Published: 4 August 2022
(This article belongs to the Section Environmental Microbiology)
Abstract
:Aquatic hyphomycetes are key microbial decomposers of allochthonous organic matter in freshwater ecosystems. Although their importance in carbon flow and food webs in streams is widely recognized, there are still gaps in our understanding of their molecular diversity and distribution patterns. Our study utilized the growing database of ITS rDNA barcodes of aquatic hyphomycetes (1252 sequences) and aimed to (i) produce new barcodes for some lesser-known taxa; (ii) clarify the taxonomic placement of some taxa at the class or order level, based on molecular data; and (iii) provide insights into the biogeographical origins of some taxa. This study increased the number of aquatic hyphomycete species with available ITS barcodes from 119 (out of ~300 species described) to 136. Phylogenetically, the 136 species were distributed between 2 phyla, 6 classes, and 10 orders of fungi. Future studies should strive to increase the database of ITS sequences, especially focusing on species with unclear phylogenetic relationships (incertae sedis) and with few sequences available. The geographical distribution of species with available ITS sequences included 50 countries from five continents, but 6 countries had more than 20 species associated, showing a bias toward the northern hemisphere, likely due to sampling bias.
1. Introduction
Aquatic hyphomycetes are the major microbial decomposers of plant litter in streams [1,2]. They play a pivotal role in these ecosystems by driving carbon and nutrient cycling and channeling energy to higher trophic levels [3], thereby contributing to the functioning of freshwater ecosystems. Aquatic hyphomycetes comprise over 300 species of fungi [4,5] with a worldwide distribution [6]. However, the occurrence of individual species is likely to depend on latitude and/or altitude [7,8,9] and to be influenced by physical characteristics of streams and rivers, as well as water chemistry [10,11,12]. A majority of aquatic hyphomycetes belong to the phylum Ascomycota [5,13]. A large number of species are in the class Leotiomycetes, while others are distributed among Sordariomycetes, Dothideomycetes, Orbiliomycetes, and Pezizomycetes [14,15,16,17]. However, the taxonomic positioning of many species of aquatic hyphomycetes remains undefined due to the lack of either teleomorph observations or molecular data.
Conidial morphology still plays a large role in the taxonomy of aquatic hyphomycetes [18,19], with many species producing stauroconidia (mostly tetraradiate spores), variously branched spores, or scolecoconidia (sigmoid, variously curved, or substraight spores). Along with spore shapes, the details of conidiogenesis are also traditionally used in systematics [20,21]. However, the conidial morphologies of aquatic hyphomycetes are believed to have evolved convergently as independent adaptations to similar environmental pressures in different phylogenetic lineages of fungi, making the conidial shape an unreliable indicator of phylogenetic relationships [16]. For some years now, great efforts in fungal taxonomy and systematics have focused on comparisons of nucleotide sequences of select genes instead of, or in addition to, phenotypic characters [22]. DNA sequences are increasingly used to investigate anamorph/teleomorph connections and phylogenetic relationships among fungal taxa [23,24,25,26]. In addition, molecular barcodes, including ITS rDNA sequence data [27], are invaluable in studies dealing with analyses of fungal community structure from environmental samples [9,28,29]. Compared to morphology, molecular data provide considerably more information for phylogenetic analyses and therefore have improved resolving power. For instance, molecular data showed that several genera of aquatic hyphomycetes are polyphyletic [17,18,30], helped to connect anamorphs to teleomorphs [26], and suggested that many aquatic hyphomycetes have relatives of terrestrial origin [18].
In an attempt to better understand phylogenetic relationships among aquatic hyphomycetes, Duarte et al. [31] found that only 26% of all described species had an internal transcribed spacer (ITS) rDNA barcode available. Although there has been an effort to sequence different loci (including ITS) from isolates of aquatic hyphomycetes since then [18,30,32,33], a large-scale comprehensive analysis of available ITS barcodes of aquatic hyphomycetes is still missing. Here, we analyzed sequence data from the ITS rDNA (most widely used DNA barcode for fungi [27]) from all species of aquatic hyphomycetes available in GenBank (119 species) and also included new sequences generated in this study in our labs from 53 pure cultures (41 species, with 17 of them sequenced for the first time). The objectives of our study were to (i) provide new barcodes of aquatic hyphomycetes, (ii) clarify the placement of some taxa at the class- or order-level based on molecular data, and (iii) discuss the biogeographical distribution of aquatic hyphomycetes with ITS barcodes.
2. Materials and Methods
2.1. Dataset Compilation
We assembled a dataset by compiling available information on aquatic hyphomycetes from the literature and public databases, using as query the terms “aquatic hyphomycetes”, “Ingoldian fungi”, and “Ingoldian hyphomycetes”. Species’ accepted name and synonyms and the information on the teleomorph and anamorph connections from three databases—Mycobank (https://www.mycobank.org/; accessed on 1 January 2021), Index Fungorum (www.indexfungorum.org; accessed on 1 January 2021), and NCBI Taxonomy (https://www.ncbi.nlm.nih.gov/taxonomy; accessed on 1 January 2021)—were compiled (Supplementary Data S1). In addition, ITS rDNA sequences were downloaded from GenBank (www.ncbi.nlm.nih.gov/genbank/; accessed on 1 January 2021), retrieving only sequences that were obtained from pure cultures (metagenomic and environmental studies were not considered).
2.2. New ITS Barcodes
Fungi from environmental samples (submerged decaying plant litter or stream foam) were isolated according to Pascoal et al. [34] and Descals [35]. Pure cultures were grown at 15 °C on 1% malt extract agar for approximately 15–20 days before DNA extraction. DNA extractions were performed by using the DNeasy PowerSoil Kit or DNeasy UltraClean Microbial Kit (Qiagen) according to the manufacturer’s instructions. PCR amplifications were performed according to Duarte et al. [36] or Baschien et al. [18], targeting the entire fungal ITS1-5.8S-ITS2 region of the rDNA by using primer pairs ITS1F/ITS4 [37,38] or SR6R/LR1 [39]. PCR products were checked on an agarose gel to confirm the presence of the desired band and then purified by using the PureLink PCR purification Kit (Invitrogen, Walthamm, MA, USA) or ExoSAP-IT PCR product cleanup reagent (Applied Biosystems, Walthamm, MA, USA) according to the manufacturer’s instructions. DNA concentrations were checked with a NanoDrop spectrophotometer (ND-1000, Thermo Fisher Scientific, Walthamm, MA, USA). The amplicons were sequenced in both directions either at StabVida (Oeiras, Portugal) or Eurofins Genomics (Louisville, KY, USA).
Isolates are maintained in culture collections, in particular at the Centre of Molecular and Environmental Biology (CBMA) of the University of Minho, Portugal; at the Czech Collection of Microorganisms (CCM), Czech Republic; and at the Coastal Carolina University, USA. The geographical origin and the substrate of all fungal isolates are given in Table 1.
2.3. Phylogenetic Analysis
Consensus sequences of ITS region were obtained with BioEdit software, version 7.2.5 (Hall TA, Raleigh, USA) [40], and were deposited in GenBank, under the accession numbers shown in Table 1. To analyze the phylogenetic relationships of aquatic hyphomycetes, we used the assembled dataset of 1252 sequences and aligned them by using the multiple sequence alignment algorithm FFT-NS2 implemented in MAFFT software version 7 (Katoh K) [41,42]. Maximum likelihood (ML) phylogeny was inferred by using IQ-TREE based on the best-fitting model (SYM + I + G4), according to the Bayesian Information Criterion (BIC), after testing 88 DNA models [43,44], with automated model finder [45] and the bootstrap algorithm UFBoot [46]. Branch support was assessed with bootstrap analysis (1000 replicates) [47]. Dissimilarity between DNA sequences assessed within and between genera was calculated by using MEGA-X software [48]. The ITS sequence of Mucor hiemalis (type strain CBS 201.65, GenBank accession number NR_152948.1) was used as the outgroup to root the trees. Phylograms were pruned, formatted, and colored in iTOL [49,50].
3. Results and Discussion
We compiled a dataset summarizing data regarding described species of aquatic hyphomycetes (Supplementary Data S1). Our dataset combines taxonomic data for a total of 323 species, in particular, the accepted name, synonyms, basionym, teleomorph, and anamorph, as well as the taxonomic placement (phylum, class, order, and family). The results showed a high taxonomic diversity of aquatic hyphomycetes that were distributed among two phyla (Ascomycota and Basidiomycota), 8 classes, 16 orders, 22 families, and 124 genera. We searched GenBank for the 323 species of aquatic hyphomycetes, and of those, ITS rDNA sequences were found for 119 species and 1073 strains, with a total of 1198 sequences since some strains had more than one sequence deposited, as they were sequenced by different researchers (Supplementary Data S2). Only sequences of aquatic hyphomycetes identified to the species level were considered (i.e., data for specimens identified to the genus level were ignored, e.g., Flagellospora sp.). We expanded this database by including new ITS rDNA sequences generated in this study from 53 pure cultures of 41 species of aquatic hyphomycetes, with 17 of them belonging to species for which no ITS data were yet available (Table 1). The 1252 barcodes considered in our study (136 species and 1127 strains) have an average size of 504 base pairs, with some isolates having longer barcodes (up to 975 base pairs), due to the existence of long inserts in the ITS region, especially among Dothideomycetes (data not shown).
Our database represents the largest number of taxa and barcodes compiled so far, much greater than those from other studies, e.g., 19 species and 94 isolates [51]; 7 species and 21 strains [52]; 6 species and 130 isolates [36]; and 75 species [18]. In addition, we generated 53 new ITS barcodes (Table 1). Our results increased the percentage of species of aquatic hyphomycetes with an available ITS barcode from 37% (119 species out of 323) to 42% (136 species out of 323). The latter number still appears to be low, emphasizing the need to generate more barcodes to better understand genetic diversity and to facilitate advances in molecular fungal ecology [53,54]. At the same time, it is likely that some of the 323 species names in our database will be eventually synonymized with others when new molecular evidence becomes available in the future. Thus, we believe that we likely covered at least 50% of the known diversity of aquatic hyphomycetes.
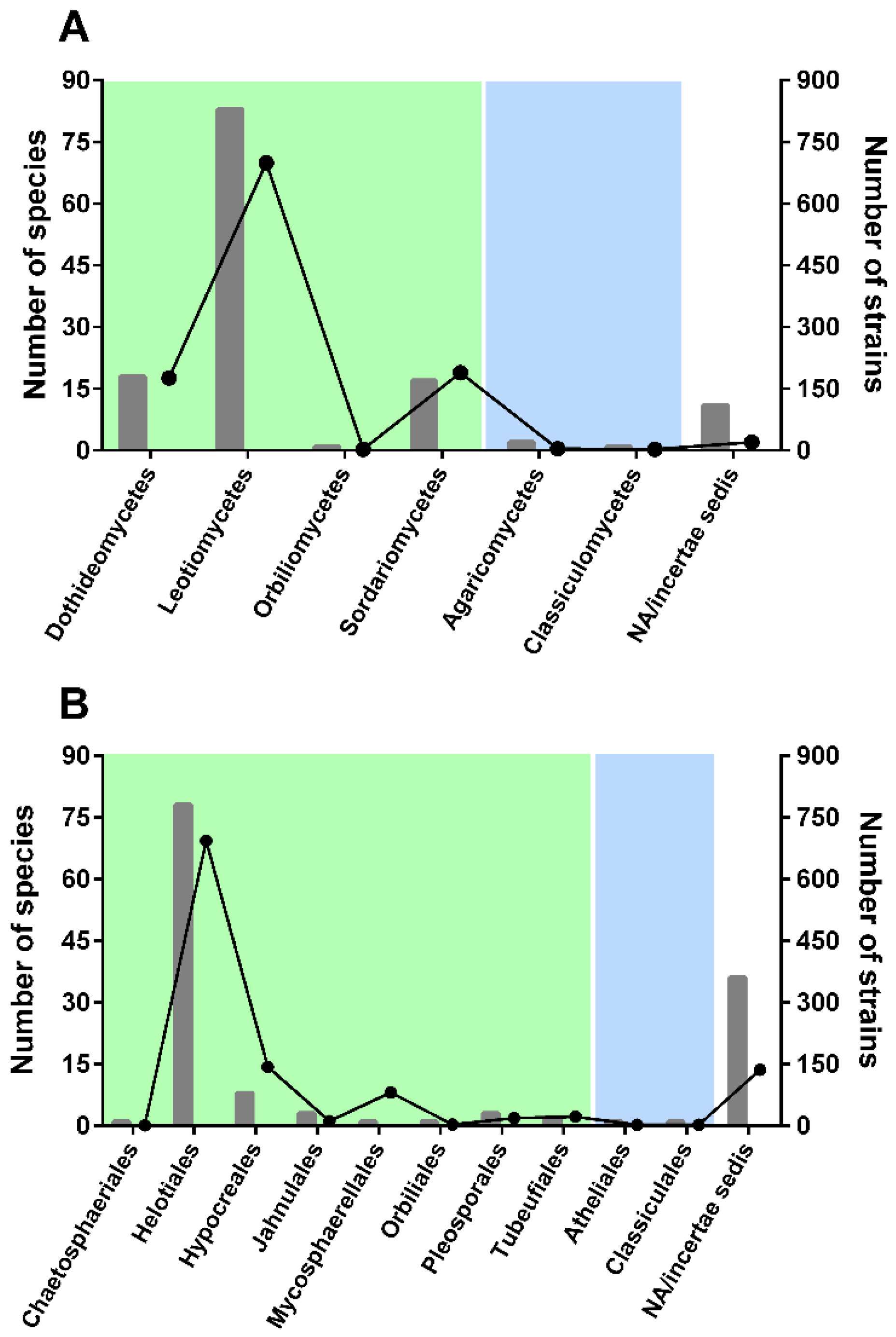
As expected, a much larger number of aquatic hyphomycetes showed an affinity to ascomycetes than to basidiomycetes (Figure 1).
In Ascomycota, the highest number of species was found to belong to Leotiomycetes (83 species, Figure 1A), with ~700 strains with available ITS sequences (56% of the total sequences considered in our study). Very few species were found to belong to Basidiomycota classes: Agaricomycetes and Classiculomycetes, with only two and one species attributed, respectively. In terms of orders (Figure 1B), Helotiales accounted for the highest number of species and strains—78 and 693, respectively. Notably, a considerable number of species (36 species; 26% of all species considered) could not be attributed to any order (incertae sedis). Clearly, additional barcodes should be generated for less represented taxa to better understand genetic diversity and placement of some species.
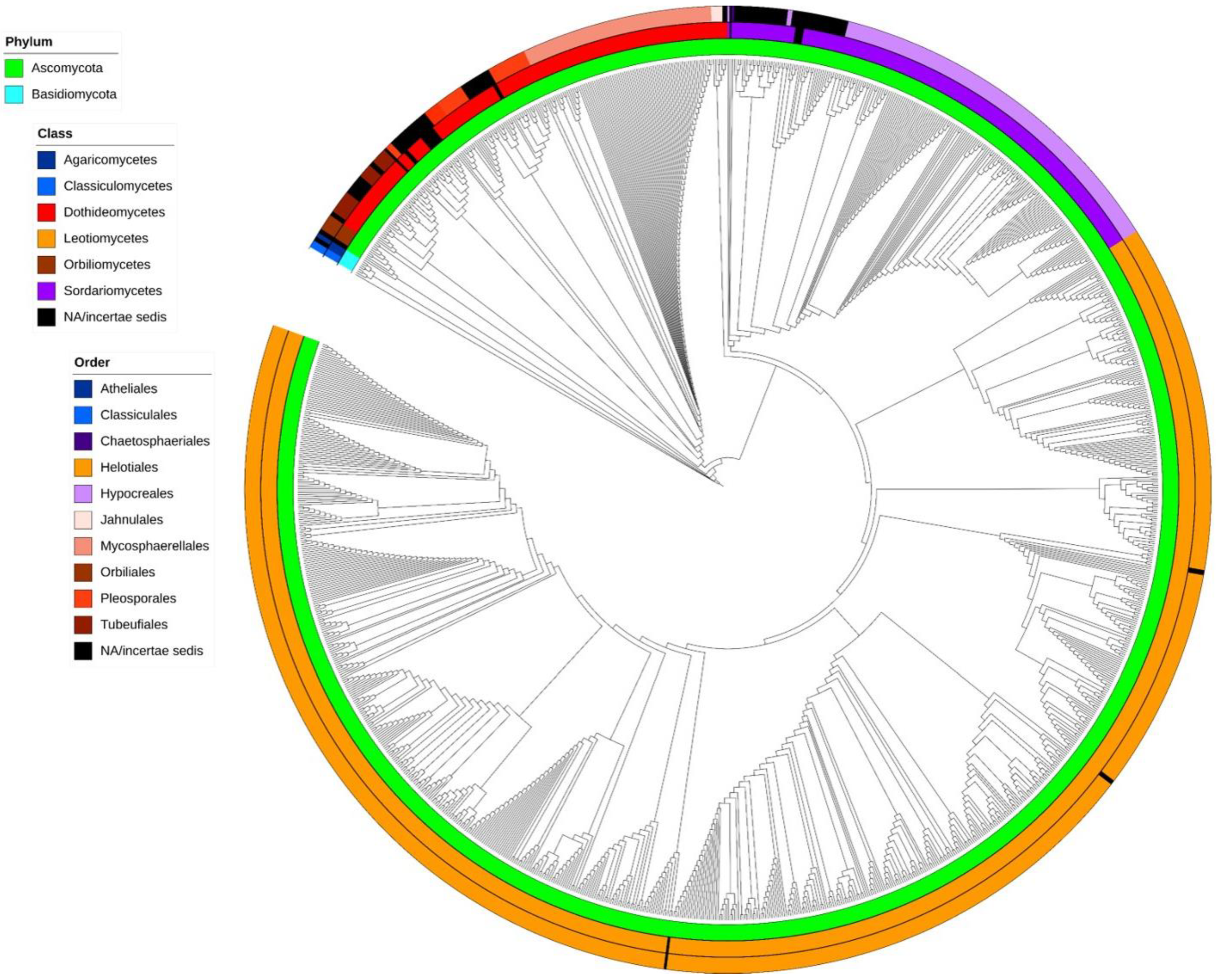
A cladogram showing phylogenetic relationships of aquatic hyphomycetes, based on ITS barcodes, is shown in Figure 2 (phylogram in traditional rectangular format is available as Supplementary Data S3). The proposed cladogram divides the 136 species between two phyla, namely Ascomycota (Figure 2, green) and Basidiomycota (Figure 2, blue), and displays the separation of 6 classes and 10 orders into well-defined clades. While the general topology of the tree based on ITS rDNA sequences of aquatic hyphomycetes was reasonable, some species and genera were problematic or not properly resolved. For example, in case of Tetracladium, even though the genus represents a well-defined clade, no clear separation among the seven species was evident. Similar results were previously reported by using sequences of 18S rDNA [55], 28S rDNA [56], and ITS+28S [18] regions. The five species of the genus Lemonniera also clustered in a well-defined clade, but, as for Tetracladium, no clear separation among the species was evident. Interestingly, our results positioned all five species within Leotiomycetes, Helotiales. This contrasts with previous results based on 28S region, where L. pseudofloscula was positioned within Dothideomycetes, Pleosporales [16]. Moreover, the genus Fontanospora was split into four groups: one group with F. eccentrica only; a second with F. fusiramosa only; a third group with F. alternibrachiata; and a final one with a mix of F. fusiramosa, F. eccentrica, and Articulospora tetracladia (the latter isolate was most probably misidentified, since all the other isolates of A. tetracladia clustered together). Fontanospora was previously reported to be polyphyletic based on analysis of 28S rDNA [17]. Isolates of Filosporella versimorpha (2) and F. fistucella (5) are intermingled on a tree, and the same pattern was observed for isolates of Alatospora flagellata (2), A. acuminata (27), and Flagellospora leucorhynchos (1), suggesting that using just ITS rDNA sequences is not sufficient to resolve their phylogenetic relationships. Anguillospora crassa separated into two distinct groups, with Tricladium obesum and Anguillospora furtiva being phylogenetically close; all of these species belong to a recently described family, Tricladiaceae [30]. In our analysis, isolates of Tumularia aquatica are separated into two groups within Dothideomycetes. One group clustered with Colispora cavincola, C. elongata, Clavariopsis aquatica, and Tumularia tuberculata. The other group (two isolates) formed a separate clade that was distant from the previous one. This may suggest a misidentification of the strains and highlights the importance of using ex-type strains with available DNA barcodes to help in the identification of problematic isolates [57]. Strains identified as Speiropsis pedatospora were also separated into two groups. One group (including ex-type culture) was clustered with Speiropsis scopiformis within Dothidiomycetes and close to the order Tubeufiales. In a recent study based on ITS + 28S sequences, S. pedatospora was positioned in the family Weisneriomycetaceae as a sister group to Tubeufiales [58]. The second group contains likely misidentified isolates (SS2229 and SS2236) and is placed in Jahnulales, Dothideomycetes [59,60,61]. ITS region does not seem suitable to resolve the phylogeny of Wiesneriomyces laurinus, since isolates of this species were split into three groups with other species in between (Speiropsis spp., Phalangispora nawawii, and P. constricta). The analysis of both 18S and 28S regions seems to have better resolving power for Wiesneriomyces laurinus [62]. We also found problems with a few isolates that did not group together with the remaining isolates of the same species, possibly due to misidentification: Amniculicola longissima WPRHD03, Neonectria lugdunensis NRRL-20592, Flagellospora curvula 30-67, Anguillospora furtiva NBRC-103659, Varicosporium elodeae AU-CRYP05, and Articulospora tetracladia CCM F-12313.
The ITS rDNA region is considered the primary fungal barcode region (e.g., the Consortium for the Barcode of Life 2007 [27,63]), showing better results for species-level identification than other markers [64], and is widely used to analyze the structure of fungal communities from environmental samples [9,28,29]. Even though our study based on ITS barcodes allowed us to clarify the taxonomic placement of many taxa (Figure 2), in line with the previous use in species- or genus-level phylogenies [64,65], results should be treated with caution. Whenever possible, multiple loci (e.g., 28S, beta-tubulin, RPB1, TEF1a, MCM7, etc.) should be used to obtain robust phylogenetic hypotheses including aquatic hyphomycetes, especially considering higher-level taxa. The ITS region, when combined with other molecular markers, showed promising results for taxonomic groups higher than the species or genus level [66,67]. Our results allowed us to shed light on the taxonomic placement of several species whose phylogenetic affinities were previously unclear (black color code in Figure 2). (i) Campylospora and Lunulospora species are placed within the order Hypocreales in Sordariomycetes (Figure 2 and Supplementary Data S3) with high robustness (bootstraps ≥ 99%). (ii) Tumularia aquatica (excluding the two ambiguous sequences) and Tumularia tuberculata both group within Dothidiomycetes, and close to species of the order Pleosporales. (iii) Three species (Goniopila monticola, Culicidospora aquatica, and C. gravida) with no clear phylogenetic affinity or with contradictory classification among databases (Mycobank vs. Index Fungorum vs. NCBI) were positioned within the order Helotiales in Leotiomycetes. C. gravida was attributed to Helotiales based on the ITS barcode produced for the first time in our study. More sequences from C. aquatica and C. gravida are needed to increase robustness of these observations. (iv) Speiropsis scopiformis, Phalangispora nawawii, and Phalangispora constricta are within Dothidiomycetes, but further analyses are needed to define their position in terms of order. Other species were attributed to classes, but because only one sequence of each species is available, these observations need to be confirmed in the future. (v) Retiarius bovicornutus and Isthmotricladia gombakiensis are clustered close to Dactylellina appendiculata within Orbiliomycetes. (vi) Heliscella stellata is close to Stenocladiella neglecta and Isthmomyces lanceatus in Dothidiomycetes. (vii) Lateriramulosa uni-inflata, Colispora cavincola, and C. elongata are also placed in Dothidiomycetes.
The highest average evolutionary divergence for all ITS sequences of aquatic hyphomycetes was found between genera Stenocladiella and Classicula (0.647; Supplementary Data S4). Actually, S. neglecta is an ascomycete (Dothidiomycetes) while C. sinensis is a basidiomycete (Classiculomycetes). Regarding the average evolutionary divergence within genus (between two or more species of the same genus), the highest divergence was observed for Mycofalcella (0.22) and the lowest for Aquanectria (0.0081) and Variocladium (0.0085). The genus Mycofalcella comprises two species: M. calcarata, which was recently repositioned in the family Tricladiaceae (Helotiales, Leotiomycetes) [30], and M. iqbalii, which is also connected to Tricladiaceae (Helotiales, Leotiomycetes), according to Mycobank and Index Fungorum. However, in our analysis, M. iqbalii is positioned within Dothideomycetes, which explains the high estimate of evolutionary divergence. Future analysis using additional markers is needed to confirm the position of M. iqbalii. Aquanectria (Hypocreales, Sordariomycetes) is a recent genus erected to accommodate two species (A. penicillioides and A. submersa) previously in the genera Flagellospora (as F. penicillioides) and Heliscus (as H. submersus), respectively [68]. The genus now includes five more species described based on multilocus phylogenetic analyses [69,70], but none of the new species is considered to be aquatic hyphomycetes. Variocladium contains two species, V. giganteum and V. rangiferinum (Helotiales, Leotiomycetes) [17,71]. Interestingly, the average evolutionary divergence within genera Aquanectria (0.0081) and Variocladium (0.0085) was lower than that of the species Vibrissea flavovirens (0.0086) (Supplementary Data S4). Thus, Aquanectria and Variocladium illustrate a situation of quick morphological diversification and/or slow molecular evolution of ITS region.
In our study, 46 species were represented by just a single sequence each. This may influence conclusions about the taxonomic placement of these particular species due to possible misidentification of isolates or inaccurate sequences deposited in public collections. We encourage continuous efforts to isolate and produce new DNA barcodes of aquatic fungi. In addition, a larger number of DNA sequences from strains isolated from different geographical regions are needed to provide an opportunity to address other questions such as haplotype analysis [72,73] in aquatic hyphomycetes.
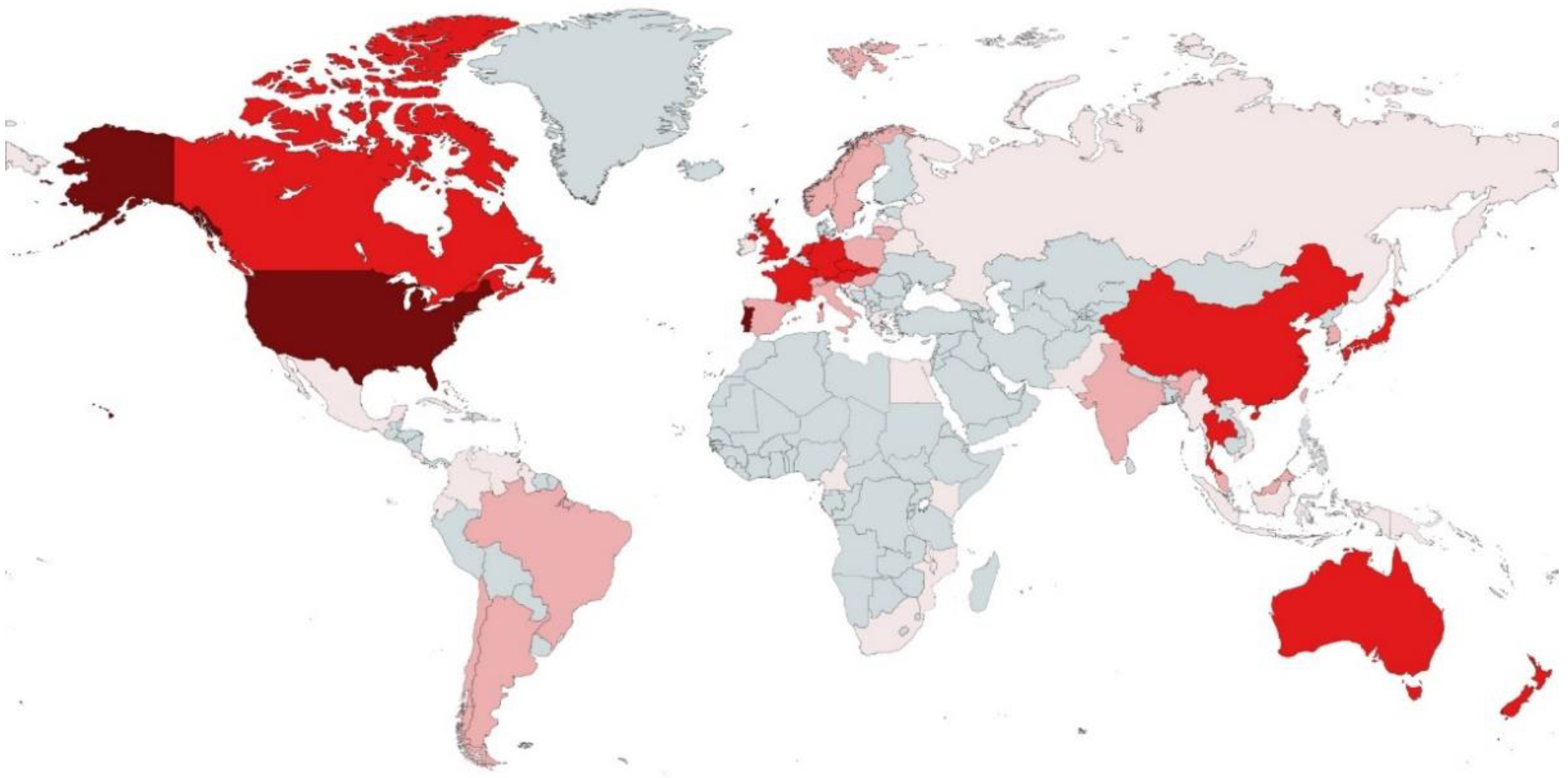
The geographical origin of all species with available ITS sequences included a total of 50 countries from five continents (Figure 3 and Supplementary Data S2).
North American and European countries had the highest number of species of aquatic hyphomycetes with available ITS barcodes: Portugal (34 species), United States of America (33 species), United Kingdom (29 species), Germany (28 species), France (22 species), and Czech Republic (21 species). These findings suggest that a larger effort isolating and barcoding aquatic hyphomycetes is still needed, especially in some parts of the world without any representation, such as in many countries of Africa and some in Asia. Clearly, the number of species reported here for different countries does not necessarily indicate the level of biodiversity but rather reflects collecting efforts or the existing expertise in the individual countries.
4. Conclusions
This study used the largest dataset of ITS rDNA barcodes (1252) of aquatic hyphomycetes to advance our understanding of phylogenetic relationships among these fungi and their biogeographical origins. Our data showed that 136 species of aquatic hyphomycetes were distributed between the fungal phyla Ascomycota and Basidiomycota, in 6 classes and 10 orders. We generated new barcodes for 17 species and clarified the taxonomic positions of some genera and species, which were previously classified as incertae sedis. Future studies should strive to increase the database of ITS sequences, especially focusing on species with unclear phylogenetic relationships (incertae sedis). A greater effort in regard to sampling, isolating, and sequencing aquatic hyphomycetes from geographically less explored regions is crucial, particularly from Africa and certain Asian countries. It would also be useful to explore extreme habitats (e.g., intermittent streams, polar regions, and deserts). In addition to sequencing rDNA loci, a multilocus approach including structural gene analysis or comparison of entire genomes might help to provide new insights into fungal classification [22,74]. Whole-genome sequencing and annotation will also facilitate the study of the phylogeography of aquatic hyphomycetes, while environmental metagenomics will help to unravel patterns of their distribution in aquatic ecosystems, including those affected by anthropogenic impacts. These modern approaches that allow for species detection in the absence of reproductive structures may open new avenues to fungal conservation [75].
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/microorganisms10081569/s1. Supplementary Data S1: List of species of aquatic hyphomycetes. Supplementary Data S2: Species of aquatic hyphomycetes with available ITS rDNA barcodes considered in this study. Supplementary Data S3: Phylogram of aquatic hyphomycetes based on 1252 ITS rDNA barcodes. Supplementary Data S4: Evolutionary divergence within and between genera.
Author Contributions
All authors contributed to the study conception and design. Data collection and analysis were performed by R.F.-D., I.F. and V.G. Funding acquisition was performed by F.C., C.P. and V.G. The first draft of the manuscript was written by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the project STREAMECO—Biodiversity and ecosystem functioning under climate change: from the gene to the stream: PTDC/CTA-AMB/31245/2017 funded by the Portuguese Foundation for Science and Technology (FCT) and by the “Contrato-Programa” UIDB/04050/2020 funded by national funds through the FCT I.P. Additional support from the National Science Foundation (NSF DEB-1655797) to V.G. is gratefully acknowledged.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Accession numbers for DNA sequences obtained for the first time in this study are listed in Table 1. Accession numbers for DNA sequences obtained from GenBank are listed in Supplementary Data S2.
Acknowledgments
Authors are grateful to Ludmila Marvanová and Monika Laichmanová for providing fungal cultures from the Czech Collection of Microorganisms (CCM).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pascoal, C.; Cássio, F. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river. Appl. Environ. Microbiol. 2004, 70, 5266–5273. [Google Scholar] [CrossRef] [Green Version]
- Gessner, M.; Gulis, V.; Kuehn, K.; Chauvet, E.K.S. Fungal decomposers of plant litter in aquatic ecosystems. In The Mycota: Environmental and Microbial Relationships; Kubicek, C., Druzhinina, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 301–321. [Google Scholar]
- Graça, M.A.S.; Canhoto, C. Leaf litter processing in low order streams. Limnetica 2006, 25, 1–10. [Google Scholar] [CrossRef]
- Descals, E. Ingoldian Fungi: Some field and laboratory techniques. Bolletí Soc. Història Nat. Balear. 1997, 40, 169–222. [Google Scholar]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Duarte, S.; Bärlocher, F.; Pascoal, C.; Cássio, F. Biogeography of aquatic hyphomycetes: Current knowledge and future perspectives. Fungal Ecol. 2016, 19, 169–181. [Google Scholar] [CrossRef]
- Chauvet, E. Aquatic Hyphomycete Distribution in South-Western France. J. Biogeogr. 1991, 18, 699. [Google Scholar] [CrossRef]
- Jabiol, J.; Bruder, A.; Gessner, M.O.; Makkonen, M.; McKie, B.G.; Peeters, E.T.H.M.; Vos, V.C.A.; Chauvet, E. Diversity patterns of leaf-associated aquatic hyphomycetes along a broad latitudinal gradient. Fungal Ecol. 2013, 6, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Seena, S.; Bärlocher, F.; Sobral, O.; Gessner, M.O.; Dudgeon, D.; McKie, B.G.; Chauvet, E.; Boyero, L.; Ferreira, V.; Frainer, A.; et al. Biodiversity of leaf litter fungi in streams along a latitudinal gradient. Sci. Total Environ. 2019, 661, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Cássio, F.; Pascoal, C. Environmental drivers are more important for structuring fungal decomposer communities than the geographic distance between streams. Limnetica 2017, 36, 491–506. [Google Scholar] [CrossRef]
- Ferreira, V.; Gulis, V.; Pascoal, C.; Graça, M.A.S. Stream pollution and fungi. In Freshwater Fungi and Fungal-Like Organisms; Jones, E.B.G., Hyde, K.D., Pang, K.L., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 389–412. [Google Scholar]
- Gulis, V.; Su, R.; Kuehn, K.A. Fungal decomposers in freshwater environments. In Advances in Environmental Microbiology. Vol. 7: The Structure and Function of Aquatic Microbial Communities; Hurst, C.J., Ed.; Springer: Cham, Switzerland, 2019; pp. 121–155. [Google Scholar]
- Barlocher, F. The Ecology of Aquatic Hyphomycetes; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Belliveau, M.J.R.; Bärlocher, F. Molecular evidence confirms multiple origins of aquatic hyphomycetes. Mycol. Res. 2005, 109, 1407–1417. [Google Scholar] [CrossRef]
- Baschien; Marvanová, L.; Szewzyk, U. Phylogeny of selected aquatic hyphomycetes based on morphological and molecular data. Nov. Hedwigia 2006, 83, 311–352. [Google Scholar] [CrossRef]
- Campbell, J.; Shearer, C.; Marvanová, L. Evolutionary relationships among aquatic anamorphs and teleomorphs: Lemonniera, Margaritispora, and Goniopila. Mycol. Res. 2006, 110, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.; Marvanová, L.; Gulis, V. Evolutionary relationships between aquatic anamorphs and teleomorphs: Tricladium and Varicosporium. Mycol. Res. 2009, 113, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Baschien, C.; Tsui, C.K.M.; Gulis, V.; Szewzyk, U.; Marvanová, L. The molecular phylogeny of aquatic hyphomycetes with affinity to the Leotiomycetes. Fungal Biol. 2013, 117, 660–672. [Google Scholar] [CrossRef]
- Gulis, V.; Marvanová, L.; Descals, E. An illustrated key to the common temperate species of aquatic hyphomycetes. In Methods to Study Litter Decomposition: A Practical Guide; Graça, M.A.S., Bärlocher, F., Gessner, M.O., Eds.; Springer: Cham, Switzerland, 2020; pp. 223–239. [Google Scholar]
- Alexopoulos, C.; Mims, C.; Blackwell, M. Introductory Mycology, 4th ed.; John Wiley and Sons: New York, NY, USA, 1996; ISBN 0-471-52229-5. [Google Scholar]
- Marvanová, L. Freshwater hyphomycetes: A survey with remarks on tropical taxa. In Tropical Mycology; Janardhanan, K., Rajendran, C., Natarajan, K., Hawksworth, D., Eds.; Science Publisher Inc.: Enfield, UK, 1997; pp. 169–226. [Google Scholar]
- Johnston, P.R.; Quijada, L.; Smith, C.A.; Baral, H.O.; Hosoya, T.; Baschien, C.; Pärtel, K.; Zhuang, W.Y.; Haelewaters, D.; Park, D.; et al. A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus 2019, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bruns, T.D.; White, T.J.; Taylor, J.W. Fungal molecular systematics. Annu. Rev. Ecol. Syst. 1991, 22, 525–564. [Google Scholar] [CrossRef]
- Berbee, M.; Taylor, J. Fungal molecular evolution: Gene trees and geologic time. In Systematics and Evolution; Springer: Berlin/Heidelberg, Germany, 2001; pp. 229–245. [Google Scholar]
- Taylor, J. A contemporary view of the holomorph: Nucleic acid sequence and computer databases are changing fungal classification. In The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, D., Taylor, J., Eds.; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Sati, S.C.; Pathak, R. Anamorph (asexual stage) Teleomorph (sexual stage) Connections in Aquatic hyphomycetes. Int. J. Plant Reprod. Biol. 2016, 8, 65–74. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Bärlocher, F.; Trabulo, J.; Cássio, F.; Pascoal, C. Stream-dwelling fungal decomposer communities along a gradient of eutrophication unraveled by 454 pyrosequencing. Fungal Divers. 2014, 70, 127–148. [Google Scholar] [CrossRef]
- Fernandes, I.; Pereira, A.; Trabulo, J.; Pascoal, C.; Cássio, F.; Duarte, S. Microscopy- or DNA-based analyses: Which methodology gives a truer picture of stream-dwelling decomposer fungal diversity? Fungal Ecol. 2015, 18, 130–134. [Google Scholar] [CrossRef]
- Johnston, P.R.; Baschien, C. Tricladiaceae fam. nov. (Helotiales, Leotiomycetes). Fungal Syst. Evol. 2020, 6, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Duarte, S.; Barlocher, F.; Cássio, F.; Pascoal, C. Current status of DNA barcoding of aquatic hyphomycetes. Sydowia 2014, 66, 191–202. [Google Scholar]
- Duarte, S.; Batista, D.; Bärlocher, F.; Cássio, F.; Pascoal, C. Some new DNA barcodes of aquatic hyphomycete species. Mycoscience 2015, 56, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Seena, S.; Marvanová, L.; Letourneau, A.; Bärlocher, F. Articulospora—Phylogeny vs morphology. Fungal Biol. 2018, 122, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, C.; Marvanová, L.; Cássio, F. Aquatic hyphomycete diversity in streams of Northwest Portugal. Fungal Divers. 2005, 19, 109–128. [Google Scholar]
- Descals, E. Techniques for handling Ingoldian fungi. In Methods to Study Litter Decomposition—A Practical Guide; Springer: Berlin/Heidelberg, Germany, 2005; pp. 129–141. [Google Scholar] [CrossRef]
- Duarte, S.; Seena, S.; Bärlocher, F.; Cássio, F.; Pascoal, C. Preliminary Insights into the Phylogeography of Six Aquatic Hyphomycete Species. PLoS ONE 2012, 7, e45289. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Inorg. Chem. Front. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Seena, S.; Pascoal, C.; Marvanová, L.; Cássio, F. DNA barcoding of fungi: A case study using ITS sequences for identifying aquatic hyphomycete species. Fungal Divers 2010, 44, 77–87. [Google Scholar] [CrossRef]
- Letourneau, A.; Seena, S.; Marvanová, L.; Bärlocher, F. Potential use of barcoding to identify aquatic hyphomycetes. Fungal Divers 2010, 40, 51–64. [Google Scholar] [CrossRef]
- Barlocher, F. Molecular approaches promise a deeper and broader understanding of the evolutionary ecology of aquatic hyphomycetes. J. N. Am. Benthol. Soc. 2010, 29, 1027–1041. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Nikolcheva, L.G.; Bärlocher, F. Phylogeny of Tetracladium based on 18S rDNA. Czech Mycol. 2002, 53, 285–295. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, X.; Wu, W.; Hao, Y.; Su, Y.; Cai, L.; Xiang, M.; Liu, X. Psychrophilic fungi from the world’s roof. Persoonia Mol. Phylogeny Evol. Fungi 2015, 34, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Pratibha, J.; Bhat, D.J.; Prabhugaonkar, A. Molecular phylogeny of Speiropsis pedatospora. Mycosphere 2016, 7, 679–686. [Google Scholar] [CrossRef]
- Prihatini, R.; Boonyuen, N.; Sivichai, S. Phylogenetic Evidence that Two Submerged-Habitat Fungal Species, Speiropsis pedatospora and Xylomyces chlamydosporus, Belong to the Order Jahnulales Insertae Sedis Dothideomycetes. Microbiol. Indones. 2008, 2, 136–140. [Google Scholar] [CrossRef]
- Suetrong, S.; Boonyuen, N.; Pang, K.L.; Ueapattanakit, J.; Klaysuban, A.; Sri-Indrasutdhi, V.; Sivichai, S.; Jones, E.B.G. A taxonomic revision and phylogenetic reconstruction of the Jahnulales (Dothideomycetes), and the new family Manglicolaceae. Fungal Divers 2011, 51, 163–188. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes; Springer: Dordrecht, The Netherlands, 2020; Volume 105, ISBN 1322502000463. [Google Scholar]
- Suetrong, S.; Rungjindamai, N.; Sommai, S.; Rung-Areerate, P.; Sommrithipol, S.; Gareth Jones, E.B. Wiesneriomyces a new lineage of Dothideomycetes (Ascomycota) basal to Tubeufiales. Phytotaxa 2014, 176, 283–297. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. Barcoding: Bold: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekpinar, A.D.; Kalmer, A. Utility of various molecular markers in fungal identification and phylogeny. Nov. Hedwigia 2019, 109, 187–224. [Google Scholar] [CrossRef]
- Harrower, E.; Ammirati, J.F.; Cappuccino, A.A.; Ceska, O.; Kranabetter, J.M.; Kroeger, P.; Lim, S.R.; Taylor, T.; Berbee, M.L. Cortinarius species diversity in British Columbia and molecular phylogenetic comparison with European specimen sequences. Botany 2011, 89, 799–810. [Google Scholar] [CrossRef]
- Ge, Z.W.; Jacobs, A.; Vellinga, E.C.; Sysouphanthong, P.; van der Walt, R.; Lavorato, C.; An, Y.F.; Yang, Z.L. A multi-gene phylogeny of Chlorophyllum (Agaricaceae, Basidiomycota): New species, new combination and infrageneric classification. MycoKeys 2018, 32, 65–90. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-K.; Hyde, K.D.; Bhat, D.J.; Wen, T.-C. Novel Taxa within Nectriaceae: Cosmosporella gen. nov. and Aquanectria sp. nov. from Freshwater Habitats in China. Cryptogam. Mycol. 2018, 39, 169–192. [Google Scholar] [CrossRef]
- Gordillo, A.; Decock, C. Multigene phylogenetic and morphological evidence for seven new species of Aquanectria and Gliocladiopsis (Ascomycota, Hypocreales) from tropical areas. Mycologia 2019, 111, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Descals, E.; Marvanová, L.; Webster, J. New taxa and combinations of aquatic hyphomycetes. Can. J. Bot. 1998, 76, 1647–1659. [Google Scholar] [CrossRef]
- Wirtz, N.; Printzen, C.; Lumbsch, H.T. Using haplotype networks, estimation of gene flow and phenotypic characters to understand species delimitation in fungi of a predominantly Antarctic Usnea group (Ascomycota, Parmeliaceae). Org. Divers. Evol. 2012, 12, 17–37. [Google Scholar] [CrossRef]
- Chiva, S.; Garrido-Benavent, I.; Moya, P.; Molins, A.; Barreno, E. How did terricolous fungi originate in the Mediterranean region? A case study with a gypsicolous lichenized species. J. Biogeogr. 2019, 46, 515–525. [Google Scholar] [CrossRef]
- Li, Y.; Steenwyk, J.L.; Chang, Y.; Wang, Y.; James, T.Y.; Stajich, J.E.; Spatafora, J.W.; Groenewald, M.; Dunn, C.W.; Hittinger, C.T.; et al. A genome-scale phylogeny of the kingdom Fungi. Curr. Biol. 2021, 31, 1653–1665. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Seena, S. Fungi in Freshwaters: Prioritising Aquatic Hyphomycetes in Conservation Goals. Water 2022, 14, 605. [Google Scholar] [CrossRef]
Figure 1.
Distribution of aquatic hyphomycete species (bars) and strains (lines) among classes (A) and orders (B) of ascomycetes (green) and basidiomycetes (blue).
Figure 1.
Distribution of aquatic hyphomycete species (bars) and strains (lines) among classes (A) and orders (B) of ascomycetes (green) and basidiomycetes (blue).

Figure 2.
Cladogram showing phylogenetic affinities of aquatic hyphomycetes based on 1252 ITS rDNA barcodes. Circle sections represent taxonomic divisions: inner circle—phyla, middle circle—classes, and outer circle—orders.
Figure 2.
Cladogram showing phylogenetic affinities of aquatic hyphomycetes based on 1252 ITS rDNA barcodes. Circle sections represent taxonomic divisions: inner circle—phyla, middle circle—classes, and outer circle—orders.

Figure 3.
World map showing the countries of origin for strains/sequences of aquatic hyphomycetes used in this study. Colors indicate the number of different species obtained from each country: ![Microorganisms 10 01569 i001]() —more than 30;
—more than 30; ![Microorganisms 10 01569 i002]() —from 11 to 29;
—from 11 to 29; ![Microorganisms 10 01569 i003]() —from 3 to 10;
—from 3 to 10; ![Microorganisms 10 01569 i004]() —less than 3 species;
—less than 3 species; ![Microorganisms 10 01569 i005]() —zero species.
—zero species.
 —more than 30;
—more than 30;  —from 11 to 29;
—from 11 to 29;  —from 3 to 10;
—from 3 to 10;  —less than 3 species;
—less than 3 species;  —zero species.
—zero species.
Figure 3.
World map showing the countries of origin for strains/sequences of aquatic hyphomycetes used in this study. Colors indicate the number of different species obtained from each country: ![Microorganisms 10 01569 i001]() —more than 30;
—more than 30; ![Microorganisms 10 01569 i002]() —from 11 to 29;
—from 11 to 29; ![Microorganisms 10 01569 i003]() —from 3 to 10;
—from 3 to 10; ![Microorganisms 10 01569 i004]() —less than 3 species;
—less than 3 species; ![Microorganisms 10 01569 i005]() —zero species.
—zero species.
—more than 30; —from 11 to 29; —from 3 to 10; —less than 3 species; —zero species.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Sources and accession numbers of aquatic hyphomycete isolates sequenced in this study. Species with new ITS barcodes generated for the first time in this study are highlighted in bold.
Table 1.
Sources and accession numbers of aquatic hyphomycete isolates sequenced in this study. Species with new ITS barcodes generated for the first time in this study are highlighted in bold.
| Species Name | Strain | Isolation Country | Isolation Substrate | GenBank Accession Number |
|---|---|---|---|---|
| Alatospora acuminata Ingold | UMB-223 | Portugal | foam | OM273714 |
| Alatospora acuminata Ingold | UMB-741 | Portugal | leaves | MZ773535 |
| Alatospora acuminata Ingold | UMB-902 | Portugal | oak leaves | OM273715 |
| Alatospora pulchella Marvanová | UMB-1115 | Portugal | oak leaves | MZ773536 |
| Anguillospora crassa Ingold | UMB-217 | Portugal | foam | OM273716 |
| Anguillospora crassa Ingold | UMB-1150 | Portugal | foam | MZ773539 |
| Anguillospora crassa Ingold | VG33-1 | USA | dead submerged tree roots | OM907724 |
| Anguillospora curvula S.H. Iqbal | VG69-4 | USA | grass blades | OM907725 |
| Anguillospora filiformis Greath. | UMB-016 | Portugal | leaves | OM273717 |
| Anguillospora filiformis Greath. | UMB-225 | Portugal | leaves | MZ773533 |
| Aquanectria penicillioides (Ingold) L. Lombard and Crous | VG205-1-2 | USA | wood | OM907726 |
| Arbusculina irregularis (R.H. Petersen) Marvanová and Descals | CCM F-23687 | Canada | unknown | OM273718 |
| Arbusculina irregularis (R.H. Petersen) Marvanová and Descals | VG76-8 | USA | foam | OM906795 |
| Articulospora atra Descals | VG233-6 | USA | wood | OM907727 |
| Articulospora proliferata A. Roldán and W.J.J. van der Merwe | VG229-6 | USA | grasses | OM907728 |
| Articulospora tetracladia Ingold | UMB-712 | Portugal | foam | OK605572 |
| Articulospora tetracladia Ingold | UMB-1144 | Portugal | foam | OK605573 |
| Casaresia sphagnorum Gonz. Frag. | VG7-1 | USA | Quercus prinus leaves | OM907729 |
| Clavariana aquatica Nawawi | VG75-4 | USA | foam | OM907730 |
| Clavatospora longibrachiata (Ingold) Sv. Nilsson ex Marvanová and Sv. Nilsson | VG80-6 | USA | Tilia sp. leaves | OM907731 |
| Culicidospora gravida R.H. Petersen | VG39-4 | USA | foam | OM907732 |
| Dendrosporomyces prolifer Nawawi, J. Webster and R.A. Davey | VG258-1 | USA | foam | OM906797 |
| Dendrosporomyces prolifer Nawawi, J. Webster and R.A. Davey | VG98-3 | USA | foam | OM906796 |
| Dimorphospora foliicola Tubaki | UMB-215 | Portugal | leaves | OM273719 |
| Dimorphospora foliicola Tubaki | UMB-1119 | Portugal | oak leaves | MZ773538 |
| Filosporella exilis Gulis and Marvanová | VG211-1 | USA | grasses | OM907733 |
| Filosporella fistucella Marvanová and P.J. Fisher | UMB-007 | Portugal | water | OM273720 |
| Fontanospora alternibrachiata Dyko | VG8-4 | USA | Rhododendron maximum leaves | OM907734 |
| Geniculospora inflata (Ingold) Sv. Nilsson ex Marvanová and Sv. Nilsson | VG79-1 | USA | twigs | OM907735 |
| Heliscella stellata (Ingold and V.J. Cox) Marvanová | VG254-5 | S. Korea | Betula sp. leaves | OM907736 |
| Heliscina antennata Marvanová | VG50-2 | USA | artificial foam | OM907737 |
| Hydrocina chaetocladia Scheuer | UMB-1116 | Portugal | oak leaves | MZ773531 |
| Isthmotricladia gombakiensis Nawawi | VG113-5 | USA | foam | OM907738 |
| Lateriramulosa uni-inflata Matsush. | VG80-7 | USA | unident. dicot leaves | OM907739 |
| Lemonniera alabamensis R.C. Sinclair and Morgan-Jones | UMB-594 | Portugal | leaves | MZ773530 |
| Lemonniera aquatica De Wild. | VG66-7 | USA | sedges | OM907740 |
| Lemonniera cornuta Ranzoni | VG77-4 | USA | foam | OM907741 |
| Lemonniera pseudofloscula Dyko | VG30-2 | USA | Acer rubrum leaves | OM907742 |
| Lemonniera terrestris Tubaki | VG209-3 | USA | leaves | OM907743 |
| Mycofalcella calcarata Marvanová, Khattab and J. Webster | VG44-4 | USA | decorticated branch | OM907744 |
| Neonectria lugdunensis (Sacc. and Therry) L. Lombard and Crous | UMB-161 | Portugal | Twigs | OK605576 |
| Pleuropedium multiseptatum Marvanová and Descals | CCM F-46594 | Canada | Unknown | OM273721 |
| Pyramidospora constricta N. Singh | VG116-5 | USA | Platanus sp. leaves | OM907745 |
| Pyramidospora ramificata Miura | VG54-1 | USA | unident. dicot leaves | OM907746 |
| Tricladium curvisporum Descals | VG69-3 | USA | Grasses | OM907747 |
| Tricladium curvisporum Descals | VG242-1 | USA | Grasses | OM907748 |
| Tricladium splendens Ingold | UMB-414 | Portugal | Foam | OK605580 |
| Tricladium splendens Ingold | UMB-1117 | Portugal | oak leaves | MZ773537 |
| Tumularia tuberculata (Gönczöl) Descals and Marvanová | VG262-4 | S. Korea | Quercus sp. leaves | OM907749 |
| Tumularia tuberculata (Gönczöl) Descals and Marvanová | VG264-4 | S. Korea | Quercus sp. leaves | OM907750 |
| Varicosporium elodeae W. Kegel | UMB-878 | Portugal | Foam | OK605582 |
| Variocladium giganteum (S.H. Iqbal) Descals and Marvanová | VG43-4 | USA | Quercus sp. leaves | OM907751 |
| Variocladium rangiferinum (Descals) Descals and Marvanová | VG71-1 | USA | Sedges | OM907752 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Franco-Duarte, R.; Fernandes, I.; Gulis, V.; Cássio, F.; Pascoal, C. ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes. Microorganisms 2022, 10, 1569. https://doi.org/10.3390/microorganisms10081569
AMA Style
Franco-Duarte R, Fernandes I, Gulis V, Cássio F, Pascoal C. ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes. Microorganisms. 2022; 10(8):1569. https://doi.org/10.3390/microorganisms10081569
Chicago/Turabian StyleFranco-Duarte, Ricardo, Isabel Fernandes, Vladislav Gulis, Fernanda Cássio, and Cláudia Pascoal. 2022. "ITS rDNA Barcodes Clarify Molecular Diversity of Aquatic Hyphomycetes" Microorganisms 10, no. 8: 1569. https://doi.org/10.3390/microorganisms10081569
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.