
The Probiotic Bacillus hwajinpoensis Colonizes the Digestive System of Crassostrea gigas Larvae and Protects Them from Vibrio alginolyticus Infection

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
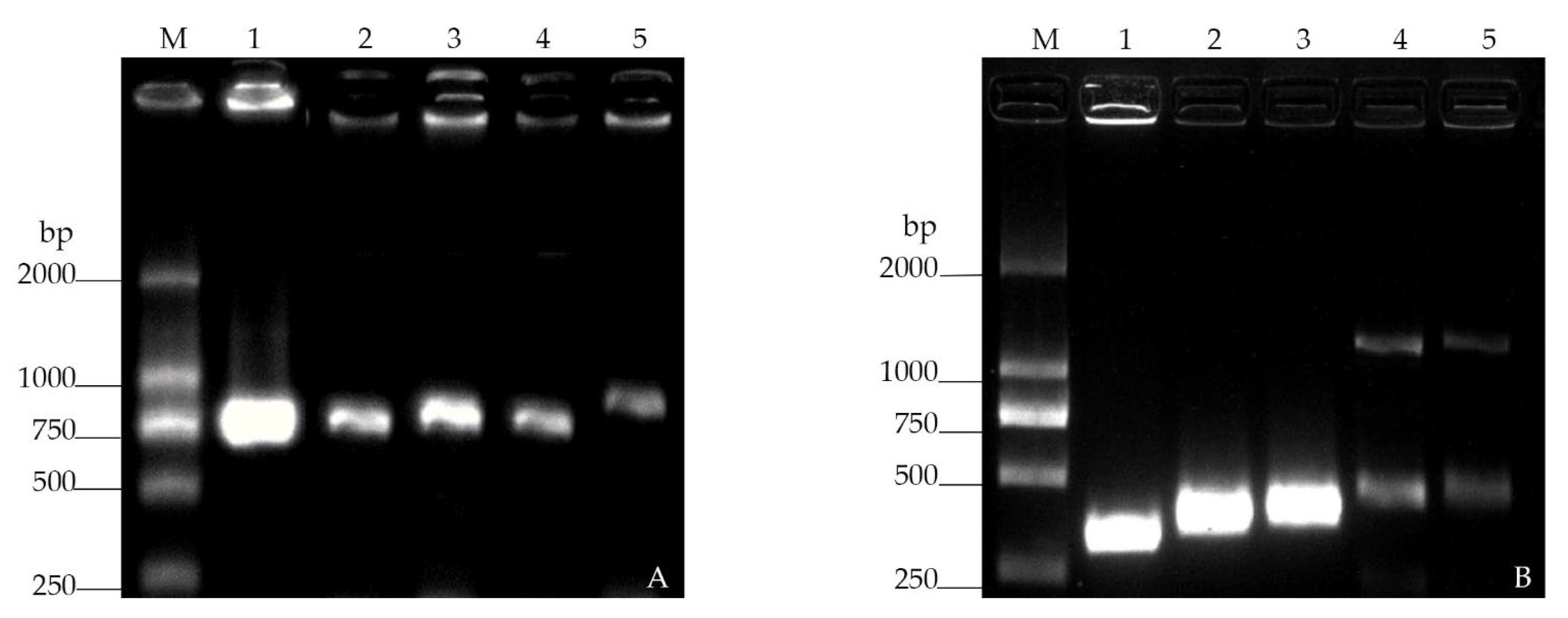
2.2. Construction of Plasmids, GFP/RFP-Tagged Strains
2.3. Bacterial Conjugation and Selection
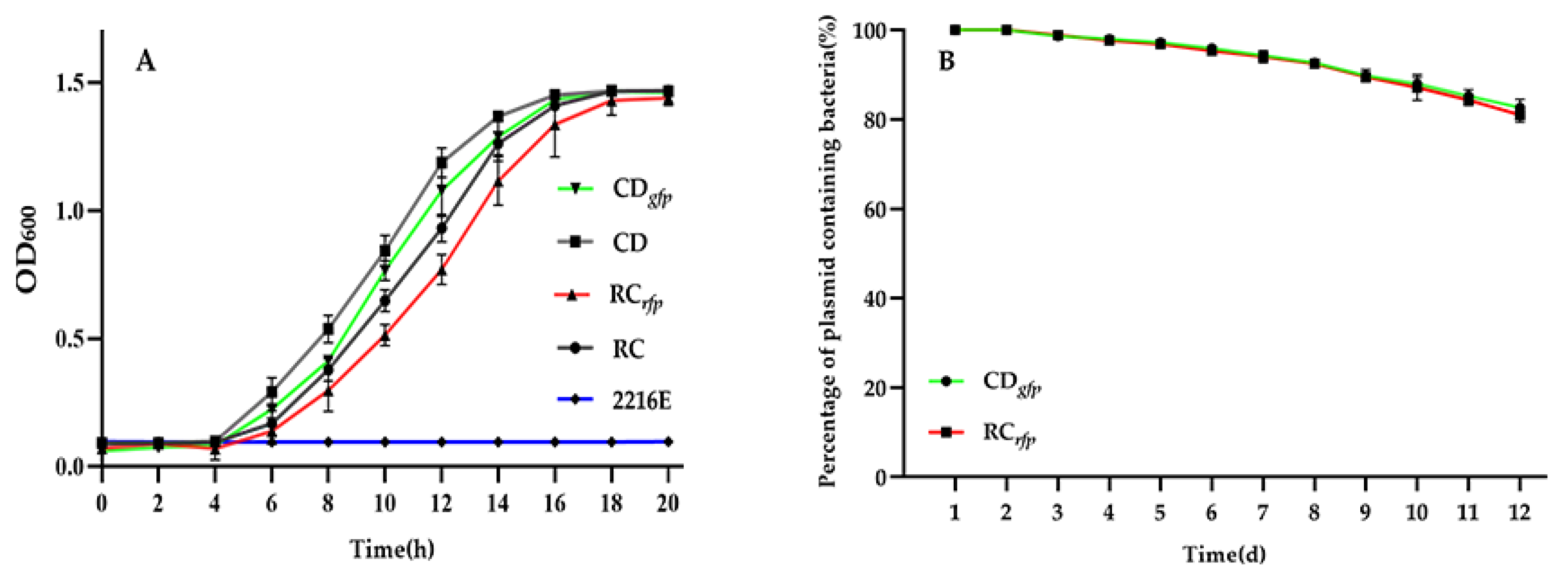
2.4. Measurement of Bacterial Growth
2.5. Measurement of Bacterial Fluorescence Stability
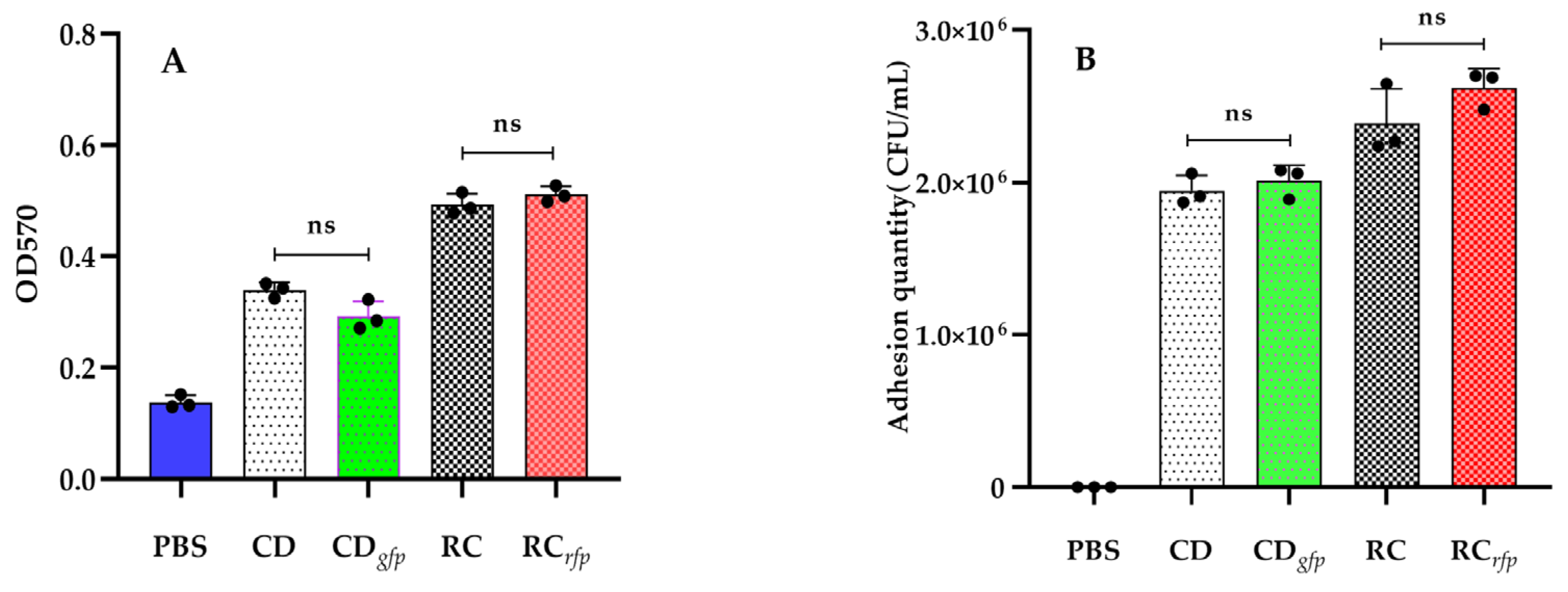
2.6. Determination of Bacterial Biofilm Formation Capability
2.7. Determination of Bacterial In Vitro Adhesion Capability
2.8. Colonization Process and Probiotic Effects
3. Results
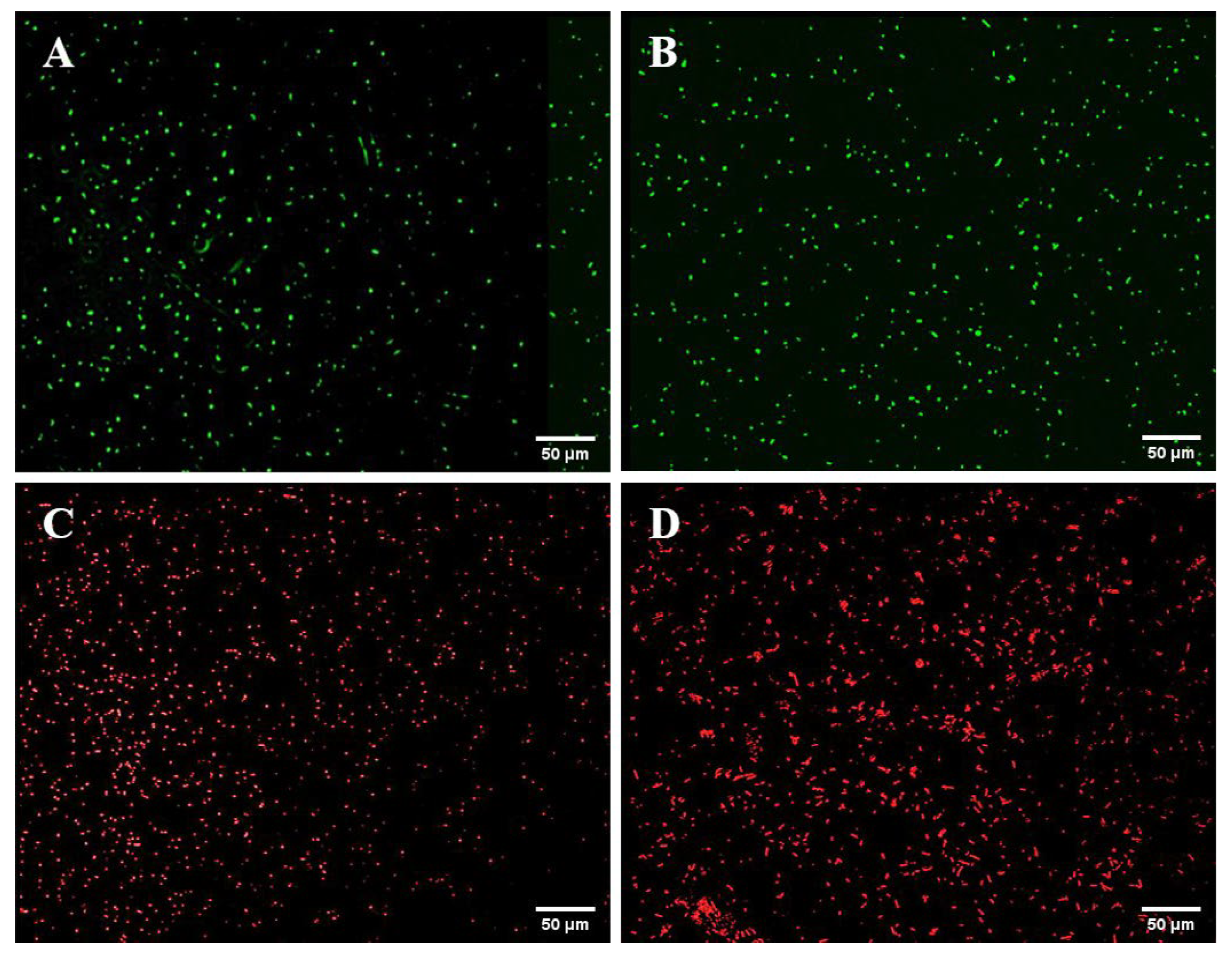
3.1. Construction of Fluorescent Strains
3.2. Growth Capability and Fluorescence Stability of the Fluorescent Strains
3.3. Biofilm Formation and In Vitro Adhesion of the Fluorescent Strains
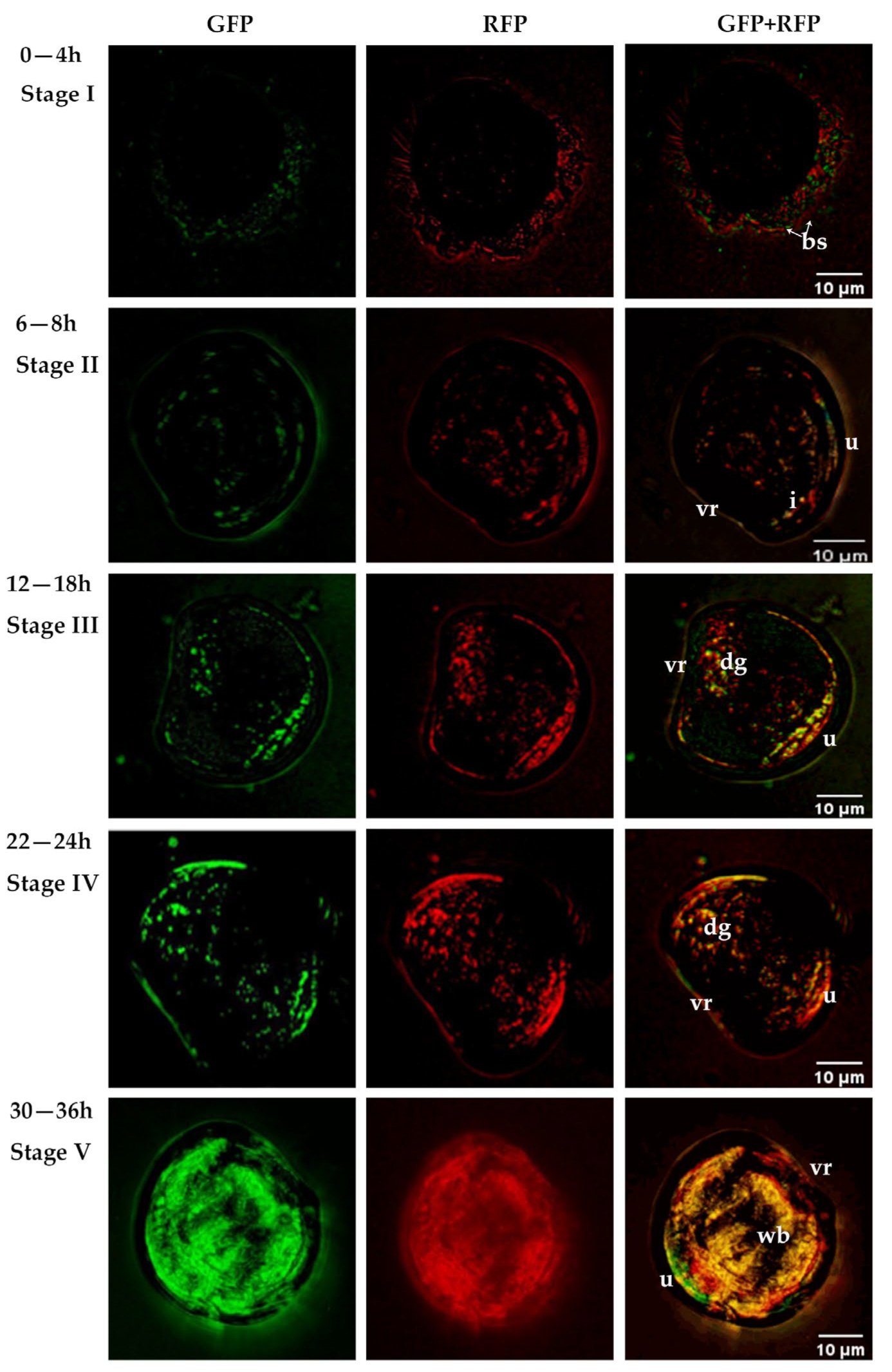
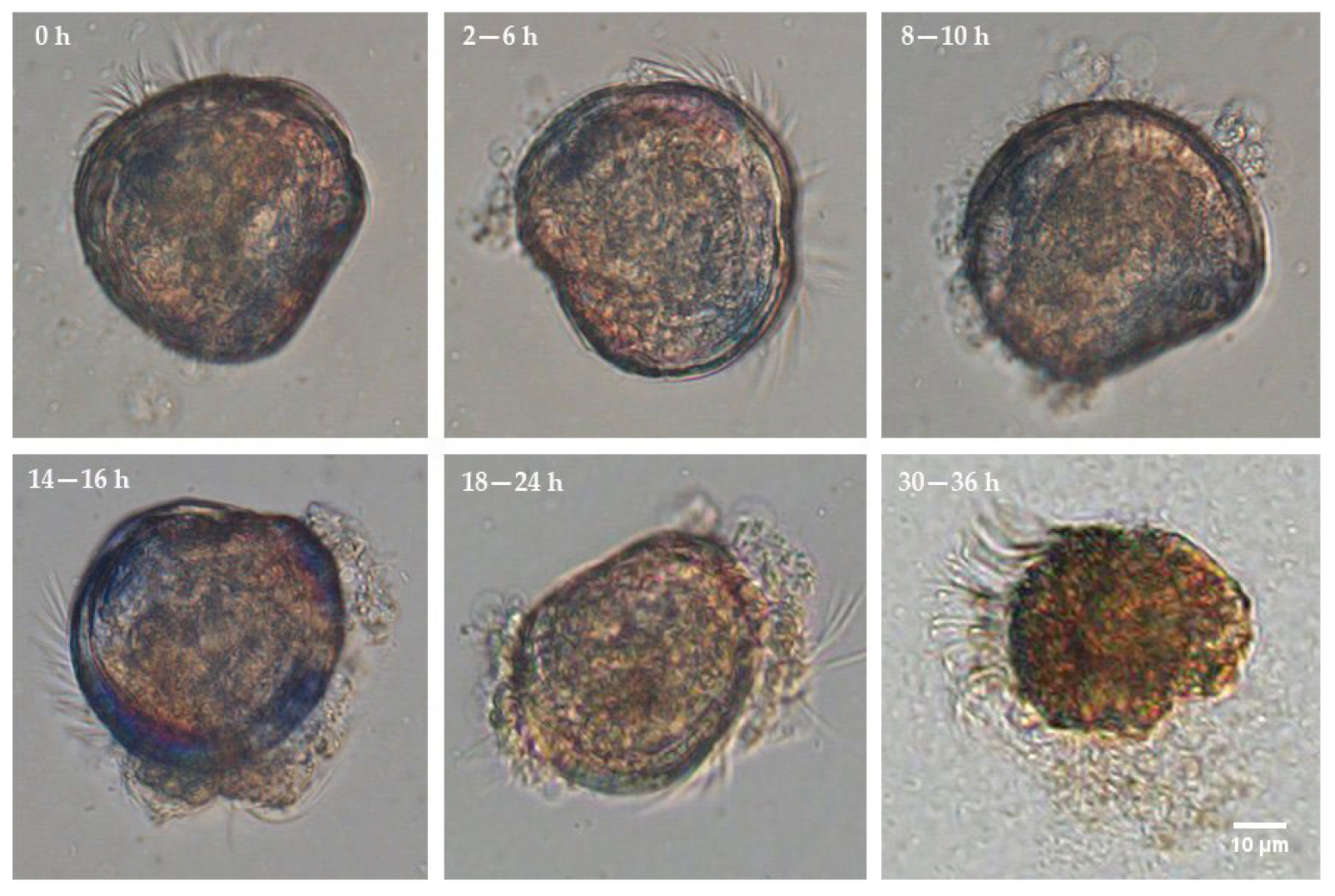
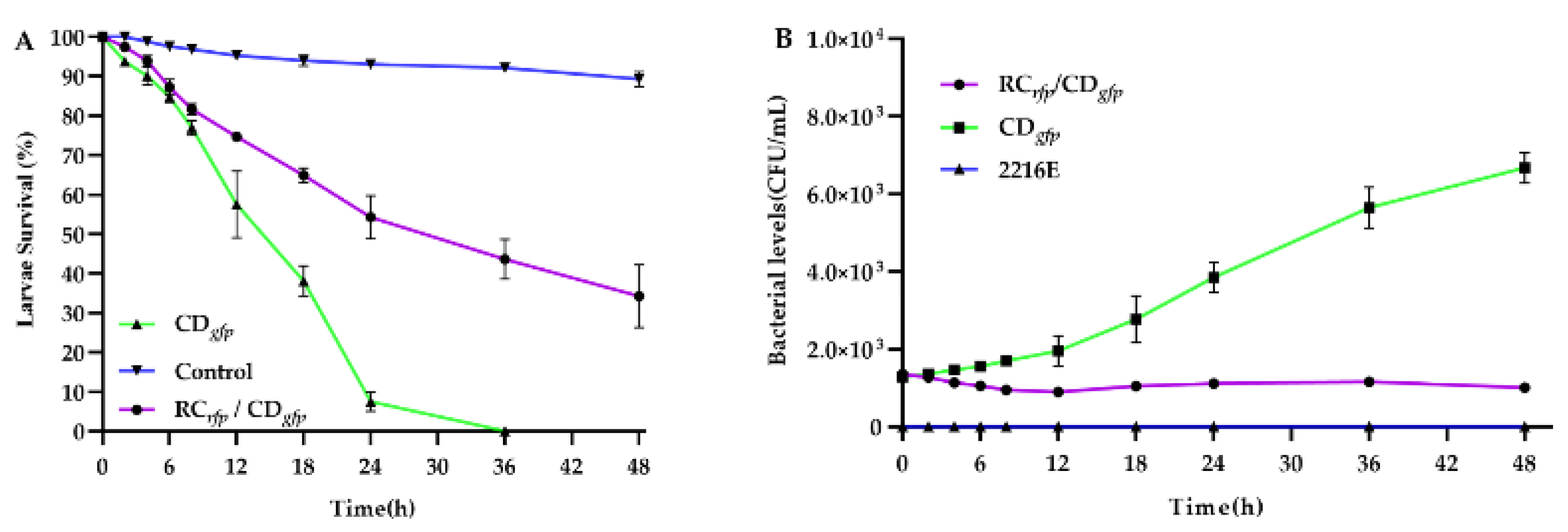
3.4. Colonisation Process and Effect on Survival of Fluorescently Tagged Strains in C. gigas D-Sharped Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bayne, B. Biology of Oysters; Academic Press: London, UK, 2017; Volume 41. [Google Scholar]
- Guo, X.M. Use and exchange of genetic resources in molluscan aquaculture. Rev. Aquacult. 2009, 1, 251–259. [Google Scholar] [CrossRef]
- China Agriculture Research System-Mollusk. Developemnt report of oyster industry in China. China Fish. 2021, 6, 20–31. [Google Scholar]
- Zhang, G.F.; Li, L.; Que, H.Y. An evolution of oyster mariculture industry in China: New knowledge, variety and product. Oceanol. Limnol. Sin. 2020, 51, 740–749. [Google Scholar]
- Jiang, R.H.; Mu, Y.T. Analysis of agglomeration characteristics of Chinese oyster breeding industry. Chin. Fish. Econ. 2021, 39, 55–61. [Google Scholar]
- Zhang, X.; Huang, B.-W.; Zheng, Y.-D.; Xin, L.-S.; Chen, W.-B.; Yu, T.; Li, C.; Wang, C.-M.; Bai, C.-M. Identification and Characterization of Infectious Pathogens Associated with Mass Mortalities of Pacific Oyster (Crassostrea gigas) Cultured in Northern China. Biology 2023, 12, 759. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Ye, J.; Xue, Q.; Liu, S.; Xu, H.; Liu, M.; Lin, Z. Changes in Bacterial Communities of Kumamoto oyster Larvae During Their Early Development and Following Vibrio Infection Resulting in a Mass Mortality Event. Mar. Biotechnol. 2023, 25, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Ye, J.; Liu, S.; Chang, G.; Xu, H.; Lin, Z.; Xue, Q. Bacterial Community Dynamics in Kumamoto oyster Crassostrea sikamea Hatchery During Larval Development. Front. Microbiol. 2022, 13, 933941. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Liu, S.; Dai, W.; He, L.; Xu, H.; Zhang, H.; Xue, Q. Characterization of Vibrio mediterranei Isolates as Causative Agents of Vibriosis in Marine Bivalves. Microbiol. Spectr. 2023, 11, e0492322. [Google Scholar] [CrossRef]
- Song, L.-S. An early warning system for diseases during mollusc mariculture: Exploration and utilization. J. Dalian Ocean Univ. 2020, 35, 1–9. [Google Scholar]
- Sainz-Hernandez, J.C.; Maeda-Martinez, A.N. Sources of Vibrio bacteria in mollusc hatcheries and control methods: A case study. Aquac. Res. 2005, 36, 1611–1618. [Google Scholar] [CrossRef]
- Batista, F.M.; Hatfield, R.; Powell, A.; Baker-Austin, C.; Lowther, J.; Turner, A.D. Methodological advances in the detection of biotoxins and pathogens affecting production and consumption of bivalve molluscs in a changing environment. Curr. Opin. Biotechnol. 2023, 80, 102896. [Google Scholar] [CrossRef] [PubMed]
- Dubert, J.; Barja, J.L.; Romalde, J.L. New Insights into Pathogenic Vibrios Affecting Bivalves in Hatcheries: Present and Future Prospects. Front. Microbiol. 2017, 8, 762. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.P.; Watson, M.A.; Needleman, D.S.; Church, K.M.; Hase, C.C. Mortalities of Eastern and Pacific oyster Larvae caused by the pathogens Vibrio coralliilyticus and Vibrio tubiashii. Appl. Environ. Microbiol. 2015, 81, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Tubiash, H.S.; Chanley, P.E.; Leifson, E. Bacillary necrosis, a disease of larval and juvenile bivalve mollusks. I. Etiology and epizootiology. J. Bacteriol. 1965, 90, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Sugumar, G.; Nakai, T.; Hirata, Y.; Matsubara, D.; Muroga, K. Vibrio splendidus biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea gigas larvae. Dis. Aquat. Org. 1998, 33, 111–118. [Google Scholar] [CrossRef]
- Genard, B.; Miner, P.; Nicolas, J.L.; Moraga, D.; Boudry, P.; Pernet, F.; Tremblay, R. Integrative Study of Physiological Changes Associated with Bacterial Infection in Pacific Oyster Larvae. PLoS ONE 2013, 8, e64534. [Google Scholar] [CrossRef]
- Kim, H.J.; Giri, S.S.; Kim, S.G.; Kim, S.W.; Kwon, J.; Lee, S.B.; Park, S.C. Isolation and Characterization of Two Bacteriophages and Their Preventive Effects against Pathogenic Vibrio coralliilyticus Causing Mortality of Pacific Oyster (Crassostrea gigas) Larvae. Microorganisms 2020, 8, 926. [Google Scholar] [CrossRef]
- Takyi, E.; LaPorte, J.; Sohn, S.; Stevick, R.J.; Witkop, E.M.; Gregg, L.S.; Chesler-Poole, A.; Small, J.; White, M.M.; Giray, C.; et al. Development and evaluation of a formulation of probiont Phaeobacter inhibens S4 for the management of vibriosis in bivalve hatcheries. Aquac. Fish Fish. 2023, 3, 256–267. [Google Scholar] [CrossRef]
- Fontana, L.; Bermudez-Brito, M.; Plaza-Diaz, J.; Munoz-Quezada, S.; Gil, A. Sources, isolation, characterisation and evaluation of probiotics. Brit. J. Nutr. 2013, 109, S35–S50. [Google Scholar] [CrossRef]
- Kouhounde, S.; Adéoti, K.; Mounir, M.; Giusti, A.; Refinetti, P.; Otu, A.; Effa, E.; Ebenso, B.; Adetimirin, V.O.; Barceló, J.M.; et al. Applications of Probiotic-Based Multi-Components to Human, Animal and Ecosystem Health: Concepts, Methodologies, and Action Mechanisms. Microorganisms 2022, 10, 1700. [Google Scholar] [CrossRef]
- Prado, S.; Romalde, J.L.; Barja, J.L. Review of probiotics for use in bivalve hatcheries. Vet. Microbiol. 2010, 145, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture 2008, 274, 1–14. [Google Scholar] [CrossRef]
- Ringø, E. Probiotic Strains for Shellfish Aquaculture. Aquac. Fish. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Zhao, W.; Dao, C.; Karim, M.; Gomez-Chiarri, M.; Rowley, D.; Nelson, D.R. Contributions of tropodithietic acid and biofilm formation to the probiotic activity of Phaeobacter inhibens. BMC Microbiol. 2016, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.; Zhao, W.J.; Rowley, D.; Nelson, D.; Gomez-Chiarri, M. Probiotic Strains for Shellfish Aquaculture: Protection of Eastern Oyster, Crassostrea Virginica, Larvae and Juveniles against Bacterial Challenge. J. Shellfish. Res. 2013, 32, 401–408. [Google Scholar] [CrossRef]
- Zhao, W.; Yuan, T.; Piva, C.; Spinard, E.J.; Schuttert, C.W.; Rowley, D.C.; Nelson, D.R. The Probiotic Bacterium Phaeobacter inhibens Downregulates Virulence Factor Transcription in the Shellfish Pathogen Vibrio coralliilyticus by N-Acyl Homoserine Lactone Production. Appl. Environ. Microbiol. 2019, 85, e01545-18. [Google Scholar] [CrossRef]
- Modak, T.H.; Gomez-Chiarri, M. Contrasting Immunomodulatory Effects of Probiotic and Pathogenic Bacteria on Eastern Oyster, Crassostrea Virginica, Larvae. Vaccine 2020, 8, 588. [Google Scholar] [CrossRef]
- Lim, H.J.; Kapareiko, D.; Schott, E.J.; Hanif, A.; Wikfors, G.H. Isolation and Evaluation of New Probiotic Bacteria for use in Shellfish Hatcheries: I. Isolation and Screening for Bioactivity. J. Shellfish. Res. 2011, 30, 609–615. [Google Scholar] [CrossRef]
- Kapareiko, D.; Lim, H.J.; Schott, E.J.; Hanif, A.; Wikfors, G.H. Isolation and Evaluation of New Probiotic Bacteria for use in Shellfish Hatcheries: II. Effects of a Vibrio sp. Probiotic Candidate Upon Survival of Oyster Larvae (Crassostrea virginica) in Pilot-Scale Trials. J. Shellfish. Res. 2011, 30, 617–625. [Google Scholar] [CrossRef]
- Sandlund, N.; Torkildsen, L.; Magnesen, T.; Mortensen, S.; Bergh, Ø. Immunohistochemistry of great scallop Pecten maximus larvae experimentally challenged with pathogenic bacteria. Dis. Aquat. Organ. 2006, 69, 163–173. [Google Scholar] [CrossRef]
- Chudakov, D.M.; Matz, M.V.; Lukyanov, S.; Lukyanov, K.A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol. Rev. 2010, 90, 1103–1163. [Google Scholar] [CrossRef] [PubMed]
- Sawabe, T.; Fukui, Y.; Stabb, E.V. Simple conjugation and outgrowth procedures for tagging vibrios with GFP, and factors affecting the stable expression of the gfp tag. Lett. Appl. Microbiol. 2006, 43, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Dubert, J.; Nelson, D.R.; Spinard, E.J.; Kessner, L.; Gomez-Chiarri, M.; Costa, F.; Prado, S.; Barja, J.L. Following the infection process of vibriosis in Manila clam (Ruditapes philippinarum) larvae through GFP-tagged pathogenic Vibrio species. J. Invertebr. Pathol. 2016, 133, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R.; Miranda, C.D.; Romero, J.; Barja, J.L.; Dubert, J. Isolation and Pathogenic Characterization of Vibrio bivalvicida Associated with a Massive Larval Mortality Event in a Commercial Hatchery of Scallop Argopecten purpuratus in Chile. Front. Microbiol. 2019, 10, 855. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Mbewe, N.; De Bels, L.; Couck, L.; Van Stappen, G.; Van den Broeck, W.; Nevejan, N. Pathogenesis of experimental vibriosis in blue mussel (Mytilus edulis) larvae based on accurate positioning of GFP-tagged Vibrio strains and histopathological and ultrastructural changes of the host. Aquaculture 2021, 535, 736347. [Google Scholar] [CrossRef]
- Dai, F.; Zhuang, Q.; Zhao, X.; Shao, Y.; Guo, M.; Lv, Z.; Li, C.; Han, Q.; Zhang, W. Green fluorescent protein-tagged Vibrio splendidus for monitoring bacterial infection in the sea cucumber Apostichopus japonicus. Aquaculture 2020, 523, 735169. [Google Scholar] [CrossRef]
- Aboubaker, M.H.; Sabrié, J.; Huet, M.; Koken, M. Establishment of stable GFP-tagged Vibrio aestuarianus strains for the analysis of bacterial infection-dynamics in the Pacific oyster, Crassostrea gigas. Vet. Microbiol. 2013, 164, 392–398. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef]
- Dai, F.; Li, Y.; Shao, Y.; Li, C.; Zhang, W. FliC of Vibrio splendidus-related strain involved in adhesion to Apostichopus japonicus. Microb. Pathog. 2020, 149, 104503. [Google Scholar] [CrossRef]
- Parizzi, S.Q.F.; Andrade, N.J.d.; Silva, C.A.d.S.; Soares, N.d.F.F.; Silva, E.A.M.d. Bacterial adherence to different inert surfaces evaluated by epifluorescence microscopy and plate count method. Braz. Arch. Biol. Technol. 2004, 47, 77–83. [Google Scholar] [CrossRef]
- Prado, S.; Dubert, J.; Barja, J.L. Characterization of pathogenic vibrios isolated from bivalve hatcheries in Galicia, NW Atlantic coast of Spain. Description of Vibrio tubiashii subsp. europaeus [corrected] subsp. nov. Syst. Appl. Microbiol. 2015, 38, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Guo, X.; Scarpa, J. Induction and Establishment of Tetraploid Oyster Breeding Stocks for Triploid Oyster Production. In UF/IFAS Extension FA215; Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2019; pp. 1–8. [Google Scholar]
- Colsoul, B.; Boudry, P.; Perez-Paralle, M.L.; Cetinic, A.B.; Hugh-Jones, T.; Arzul, I.; Merou, N.; Wegner, K.M.; Peter, C.; Merk, V.; et al. Sustainable large-scale production of European flat oyster (Ostrea edulis) seed for ecological restoration and aquaculture: A review. Rev. Aquac. 2021, 13, 1423–1468. [Google Scholar] [CrossRef]
- Helm, M.M.; Bourne, N. Hatchery Culture of Bivalves: A Practical Manual; FAO Fisheries Department: Rome, Italy, 2004. [Google Scholar]
- Mou, M.J. The Development of Oyster Breeding Industry in Shandong Province under the New Situation. J. Aquac. 2020, 41, 73–74+77. [Google Scholar]
- Sui, W.J.; Wang, H.X.; Liu, B.Z. Analysis of bacterial community and identification of pathogenic bacteria in the diseased clam larvae of Meretrix petechialis. J. Fish. China 2023, 47, 069413. [Google Scholar]
- Chauhan, A.; Singh, R. Probiotics in aquaculture: A promising emerging alternative approach. Symbiosis 2019, 77, 99–113. [Google Scholar] [CrossRef]
- Fernandes, S.; Kerkar, S. Bacterial probiotics over antibiotics: A boon to aquaculture. In Advances in Biological Science Research—A Practical Approach; Meena, S.N., Naik, M.M., Eds.; Academic Press: Salt Lake City, UT, USA, 2019; pp. 215–232. [Google Scholar]
- Sharifuzzaman, S.M.; Austin, B. Probiotics for Disease Control in Aquaculture. In Diagnosis and Control of Diseases of Fish and Shellfish; Austin, B., Newaj-Fyzul, A., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 2017; pp. 189–222. [Google Scholar]
- Sohn, S.; Lundgren, K.M.; Tammi, K.; Karim, M.; Smolowitz, R.; Nelson, D.R.; Rowley, D.C.; Gómez-Chiarri, M. Probiotic Strains for Disease Management in Hatchery Larviculture of the Eastern Oyster Crassostrea virginica. J. Shellfish Res. 2016, 35, 307–317. [Google Scholar] [CrossRef]
- Kesarcodi-Watson, A.; Miner, P.; Nicolas, J.-L.; Asmani, K.; Robert, R. Pathogenic threats and probiotic use in larviculture of the scallop, Pecten maximus. Aquac. Res. 2016, 47, 1221–1230. [Google Scholar] [CrossRef]
- Abasolo-Pacheco, F.; Campa-Córdova, Á.I.; Mazón-Suástegui, J.M.; Tovar-Ramírez, D.; Araya, R.; Saucedo, P.E. Enhancing growth and resistance to Vibrio alginolyticus disease in catarina scallop (Argopecten ventricosus) with Bacillus and Lactobacillus probiotic strains during early development. Aquac. Res. 2017, 48, 4597–4607. [Google Scholar] [CrossRef]
- Kesarcodi-Watson, A.; Miner, P.; Nicolas, J.-L.; Robert, R. Protective effect of four potential probiotics against pathogen-challenge of the larvae of three bivalves: Pacific oyster (Crassostrea gigas), flat oyster (Ostrea edulis) and scallop (Pecten maximus). Aquaculture 2012, 344–349, 29–34. [Google Scholar] [CrossRef]
- Zheng, Y.D. Screening of Probiotic bacteria in Shellfish and Their Effect on Bacterial Communities and Larval Survival; Ocean University of China: Qingdao, China, 2023. [Google Scholar]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Two pathogens of Greenshell™ mussel larvae, Perna canaliculus: Vibrio splendidus and a V. coralliilyticus/neptunius-like isolate. J. Fish. Dis. 2009, 32, 499–507. [Google Scholar] [CrossRef]
- Urtubia, R.; Miranda, C.D.; Rodríguez, S.; Dubert, J.; Barja, J.L.; Rojas, R. First Report, Characterization and Pathogenicity of Vibrio chagasii Isolated from Diseased Reared Larvae of Chilean Scallop, Argopecten purpuratus (Lamarck, 1819). Pathogens 2023, 12, 183. [Google Scholar] [CrossRef]
- Cabello, A.E.; Espejo, R.T.; Romero, J. Tracing Vibrio parahaemolyticus in oysters (Tiostrea chilensis) using a Green Fluorescent Protein tag. J. Exp. Mar. Biol. Ecol. 2005, 327, 157–166. [Google Scholar] [CrossRef]
- Travers, M.A.; Barbou, A.; Le Goïc, N.; Huchette, S.; Paillard, C.; Koken, M. Construction of a stable GFP-tagged Vibrio harveyi strain for bacterial dynamics analysis of abalone infection. Fems Microbiol. Lett. 2008, 289, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Rekecki, A.; Gunasekara, R.A.; Dierckens, K.; Laureau, S.; Boon, N.; Favoreel, H.; Cornelissen, M.; Sorgeloos, P.; Ducatelle, R.; Bossier, P.; et al. Bacterial host interaction of GFP-labelled Vibrio anguillarum HI-610 with gnotobiotic sea bass, Dicentrarchus labrax (L.), larvae. J. Fish. Dis. 2012, 35, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Allison, D.G.; Sattenstall, M.A. The influence of green fluorescent protein incorporation on bacterial physiology: A note of caution. J. Appl. Microbiol. 2007, 103, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.S.; Zhang, S.; Eggermont, M.; Bruto, M.; Roux, F.L.; Defoirdt, T. The impact of the multichannel quorum sensing systems of Vibrio tasmaniensis and Vibrio crassostreae on virulence towards blue mussel (Mytilus edulis) larvae. Aquaculture 2022, 547, 737414. [Google Scholar] [CrossRef]
- Campbell, A.H.; Meritt, D.W.; Franklin, R.B.; Boone, E.L.; Nicely, C.T.; Brown, B.L. Effects of age and composition of field-produced biofilms on oyster larval setting. Biofouling 2011, 27, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Stevick, R.J.; Sohn, S.; Modak, T.H.; Nelson, D.R.; Rowley, D.C.; Tammi, K.; Smolowitz, R.; Lundgren, K.M.; Post, A.F.; Gomez-Chiarri, M. Bacterial Community Dynamics in an Oyster Hatchery in Response to Probiotic Treatment. Front. Microbiol. 2019, 10, 1060. [Google Scholar] [CrossRef]
- Lidsky, P.V.; Lukyanov, K.A.; Misra, T.; Handke, B.; Mishin, A.S.; Lehner, C.F. A genetically encoded fluorescent probe for imaging of oxygenation gradients in living Drosophila. Development 2018, 145, dev156257. [Google Scholar] [CrossRef]
- Schmitt, P.; Duperthuy, M.; Montagnani, C.; Bachère, E.; Destoumieux-Garzón, D. Immune responses in the Pacific oyster Crassostrea gigas: An overview with focus on summer mortalities. In Oysters: Physiology, Ecological Distribution and Mortality; Qin, J., Ed.; Nova Science Pub Inc: Hauppauge, NY, USA, 2011; p. 311. [Google Scholar]
- Elston, R.; Leibovitz, L. Pathogenesis of Experimental Vibriosis in Larval American Oysters, Crassostrea virginica. Can. J. Fish. Aquat. Sci. 1980, 37, 964–978. [Google Scholar] [CrossRef]
- Chabrillón, M.; Rico, R.M.; Arijo, S.; Díaz-Rosales, P.; Balebona, M.C.; Moriñigo, M.A. Interactions of microorganisms isolated from gilthead sea bream, Sparus aurata L., on Vibrio harveyi, a pathogen of farmed Senegalese sole, Solea senegalensis (Kaup). J. Fish. Dis. 2005, 28, 531–537. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) * | Enzyme Site | Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| 27-F | AGAGTTTGATCCTGGCTCAG | none | 1517 | 55 |
| 1492-R | TACGACTTAACCCCAATCGC | |||
| PTrc-F |  | Bgl I | 48 | 56 |
| PTrc-R |  | |||
| GFP-F |  | Nco I | 738 | 60 |
| GFP-R |  | Xho I | ||
| RFP-F |  | Nco I | 715 | 58 |
| RFP-R |  | Xho I | ||
| RFP-HF | CGACTTCTTTAAGCAGTCCTTCC | none | 336 | 55 |
| RFP-HR | ATCTTGAGGTTCTTAGCGGGTTT |
| Antibiotics | Strains | ||
|---|---|---|---|
| V. alginolyticus CD | B. hwajinpoensis RC | E. coli S17-1λpir/pET28a-Ptrc-T7-GFP/RFP | |
| Penicillin | + | − | ++ |
| Ampicillin | − | +++ | +++ |
| Kanamycin | +++ | ++ | − |
| Gentamicin | +++ | ++++ | +++ |
| Erythromycin | +++ | +++ | − |
| Ciprofloxacin | +++ | − | +++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.-D.; Huang, B.-W.; Zhang, X.; Liu, C.-F.; Xin, L.-S.; Wang, C.-M.; Bai, C.-M. The Probiotic Bacillus hwajinpoensis Colonizes the Digestive System of Crassostrea gigas Larvae and Protects Them from Vibrio alginolyticus Infection. Microorganisms 2023, 11, 2918. https://doi.org/10.3390/microorganisms11122918
Zheng Y-D, Huang B-W, Zhang X, Liu C-F, Xin L-S, Wang C-M, Bai C-M. The Probiotic Bacillus hwajinpoensis Colonizes the Digestive System of Crassostrea gigas Larvae and Protects Them from Vibrio alginolyticus Infection. Microorganisms. 2023; 11(12):2918. https://doi.org/10.3390/microorganisms11122918
Chicago/Turabian StyleZheng, Yu-Dong, Bo-Wen Huang, Xiang Zhang, Chen-Feng Liu, Lu-Sheng Xin, Chong-Ming Wang, and Chang-Ming Bai. 2023. "The Probiotic Bacillus hwajinpoensis Colonizes the Digestive System of Crassostrea gigas Larvae and Protects Them from Vibrio alginolyticus Infection" Microorganisms 11, no. 12: 2918. https://doi.org/10.3390/microorganisms11122918