
In Vitro Effect of the Cell-Free Supernatant of the Lactobacillus casei Strain IMAU60214 against the Different Pathogenic Properties of Diarrheagenic Escherichia coli
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Coaggregation Assay
+ OD 600 nm specific pathogen/2] × 100
2.3. Preparation of Cell-Free Supernatant (CSF) of L. casei IMAU60214
2.4. Evaluation of the Antimicrobial Activity of CSF against the Diarrheagenic E. coli (DEC) Strains
2.5. Biofilm Formation of Diarrheagenic Strains (DEC)
2.6. Evaluation of the Anti-Biofilm Formation Activity of the CSF of L. casei IMAU60214 against the DEC Diarrheagenic Strain
2.6.1. Effect of CSF from L. casei IMAU60214 on the Biofilm Formation of the DEC Strains
2.6.2. Effect of the CSF of L. casei IMAU60214 on the Dispersion of Biofilms of the DEC Strains
2.6.3. Evaluation of the Anti-Biofilm Activity and Biofilm Dispersion of the DEC Strains after Proteolytic Enzymatic Treatment with the Catalase and Proteinase K of the CSF of L. casei IMAU60214
2.7. Culture of the HT-29 Human Intestinal Epithelium Cells
2.8. Infection Assay of the HT-29 Epithelial Cells with the DEC Strains
2.9. Cellular Cytotoxicity Assay in HT-29 the Human Epithelial Cells Post Infection with the Escherichia coli DEC Strain
2.10. Evaluation of the Effect of the CSF from L. casei IMAU60214 on the Co-Culture of HT-29 Human Epithelial Cells Infected with the Bacterial Cells of the DEC 49766 EHEC and EDL933 Strains
2.11. Statistical Analysis
3. Results
3.1. Coaggregation of L. casei IMAU60214 with the DEC Strains
3.2. Antimicrobial Activity of the Cell-Free Supernatant (CSF) of L. casei IMAU60214 against Diarrheagenic E. coli
3.3. Inhibitory Effect of the CFS from L. casei IMAU60214 on the Biofilm Formation of the DEC Strains
3.4. Effect of the CFS from L. casei IMAU60214 on the Formation and Dispersion of DEC Biofilms
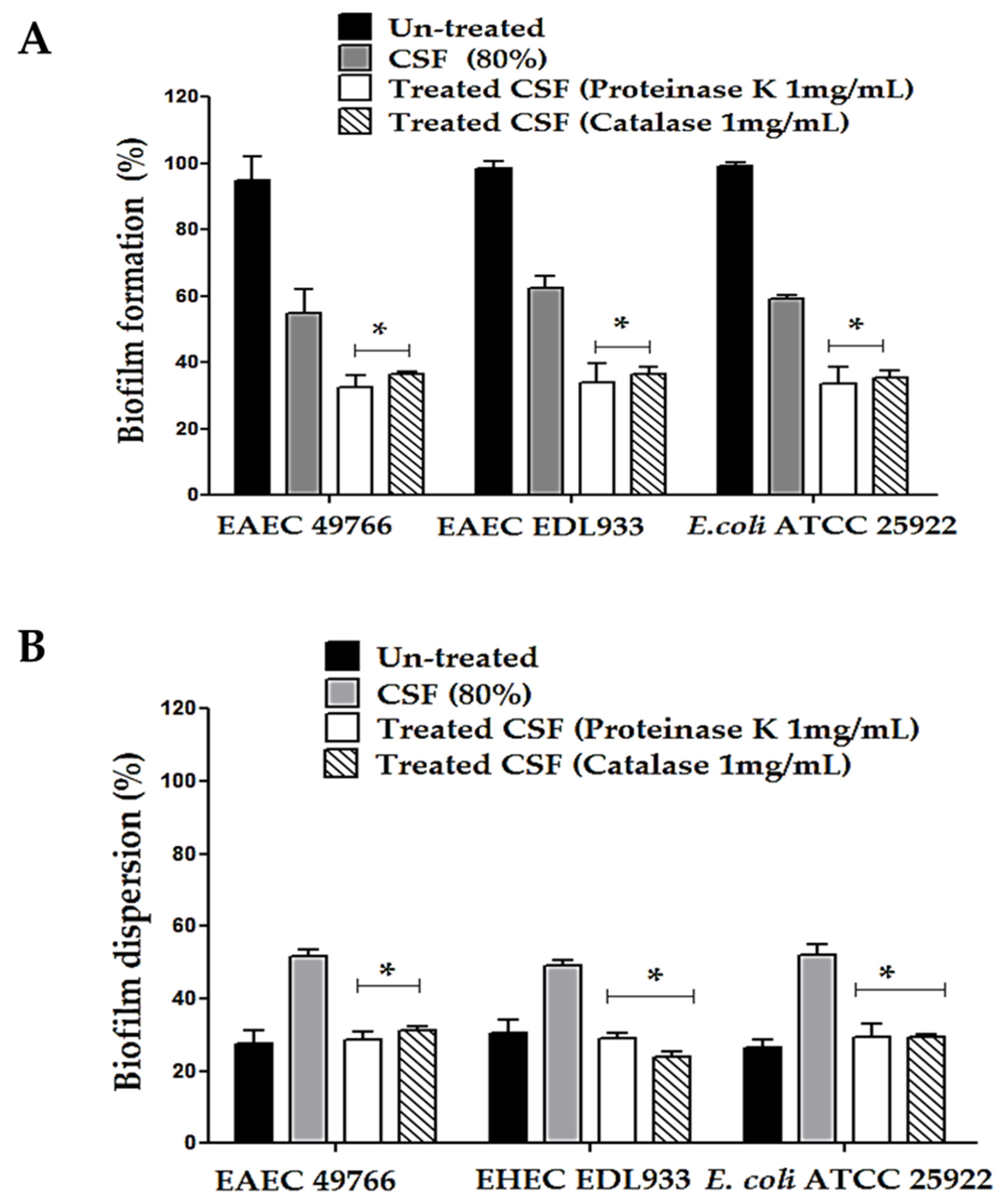
3.5. Effect of CSF Pretreated with Catalase and Proteinase K on the Biofilm Formation and Dispersion Activity of the DEC Strains
3.6. Effect of CSF Pretreated with Catalase and Proteinase K on the Biofilm Formation and Dispersion Activity of the DEC Strains
3.7. Effect of the CSF from L. casei IMAU60214 in an In Vitro Co-Culture with Human HT-29 Epithelial Cells Infected with the DEC Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leimbach, A.; Hacker, J.; Dobrindt, U. E. coli as an all-rounder: The thin line between commensalism and pathogenicity. Curr. Top. Microbiol. Immunol. 2013, 358, 3–32. [Google Scholar]
- Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D. Extraintestinal Pathogenic Escherichia coli: A Combination of Virulence with Antibiotic Resistance. Front. Microbiol. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, F.R.; Garcia-Lozano, H.; Rodriguez, E.; Cervantes, Y.; Gómez, A.; Melo, M.; Anaya, L.; Ovalle, J.C.; Torres, J.; Diaz De Jesus, B.; et al. Diarrhea morbidity and mortality in Mexican children: Impact of rotavirus disease. Pediatr. Infect. Dis. J. 2004, 23, S149–S155. [Google Scholar] [CrossRef] [PubMed]
- Souza, Y.F.V.P.; Souza, E.V.; Azevedo, L.S.; Medeiros, R.S.; Timenetsky, M.D.C.S.T.; Luchs, A. Enteric adenovirus epidemiology from historical fecal samples in Brazil (1998–2005): Pre-rotavirus vaccine era. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2021, 94, 105007. [Google Scholar] [CrossRef]
- Shah, M.P.; Hall, A.J. Norovirus Illnesses in Children and Adolescents. Infect. Dis. Clin. N. Am. 2018, 32, 103–118. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W., 4th. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar]
- O’Neill, J. Antimicrobial Resistence; Tackling a Crisis for the Health and Wealth of Nations. 2014. Available online: https://amr-review.org/sites/default/files/AMR%20Review%20Paper%20-%20Tackling%20a%20crisis%20for%20the%20health%20and%20wealth%20of%20nations_1.pdf (accessed on 25 January 2023).
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, C.; Huang, J.; Kuai, X.; Shao, X. The potential therapeutic role of Lactobacillus reuteri for treatment of inflammatory bowel disease. Am. J. Transl. Res. 2020, 12, 1569–1583. [Google Scholar] [PubMed]
- Baral, K.C.; Bajracharya, R.; Lee, S.H.; Han, H.K. Advancements in the Pharmaceutical Applications of Probiotics: Dosage Forms and Formulation Technology. Int. J. Nanomed. 2021, 16, 7535–7556. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [PubMed]
- Kaillasapathy, K.; Chin, J. Survavival and therapeutic potential of probiotic organism with reference to Lactobacillis acidophilus and Bifidobacterium spp. Inmunol. Cell Biol. 2000, 78, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Tabasco, R.; Paarup, T.; Janer, C.; Peláez, C.; Requena, T. Selective enumeration and identification of mixed cultures of Streptococcus thermophilus, Lactobacillus delbrueckii subsp. bulgaricus, L. acidophilus, L. paracasei subsp. paracasei and Bifidobacterium lactis in fermented milk. Int. Dairy J. 2007, 17, 1107–1114. [Google Scholar]
- Peng, X.; Ed-Dra, A.; Song, Y.; Elbediwi, M.; Nambiar, R.B.; Zhou, X.; Yue, M. Lacticaseibacillus rhamnosus alleviates intestinal inflammation and promotes microbiota-mediated protection against Salmonella fatal infections. Front. Immunol. 2022, 13, 973224. [Google Scholar] [CrossRef]
- Jung, J.Y.; Han, S.S.; Kim, Z.H.; Kim, M.H.; Kang, H.K.; Jin, H.M.; Lee, M.H. In-Vitro Characterization of Growth Inhibition against the Gut Pathogen of Potentially Probiotic Lactic Acid Bacteria Strains Isolated from Fermented Products. Microorganisms 2021, 9, 2141. [Google Scholar] [CrossRef]
- Liu, W.; Pang, H.; Zhang, H.; Cai, Y. Biodiversity of Lactic Acid Bacteria. In Lactic Acid Bacteria; Zhang, H., Cai, Y., Eds.; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. Liver Physiol. 1999, 276, G941–G950. [Google Scholar]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Moreno Guerrero, S.S.; Ramírez Pacheco, A.; García Garibay, M.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Choi, J.W.; Jhun, J.; Kwon, J.Y.; Lee, B.I.; Yang, C.W.; Park, S.H.; Cho, M.L. Lactobacillus acidophilus Improves Intestinal Inflammation in an Acute Colitis Mouse Model by Regulation of Th17 and Treg Cell Balance and Fibrosis Development. J. Med. Food 2018, 21, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanism of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Nagpal, R.; Verma, V.; Kumar, A.; Kaur, N.; Hemalatha, R.; Gautam, S.K.; Singh, B. Probiotic metabolites as epigenetic targets in the prevention of colon cancer. Nutr. Rev. 2013, 71, 23–34. [Google Scholar] [CrossRef]
- Sionov, R.V.; Steinberg, D. Targeting the Holy Triangle of Quorum Sensing, Biofilm Formation, and Antibiotic Resistance in Pathogenic Bacteria. Microorganisms 2022, 10, 1239. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Cruz-Guerrero, A.; Hernández-Sánchez, H.; Rodríguez-Serrano, G.; Gómez-Ruiz, L.; García-Garibay, M.; Figueroa-González, I. Commercial probiotic bacteria and prebiotic carbohydrates: A fundamental study on prebiotics uptake, antimicrobials production and inhibition of pathogens. J. Sci. Food Agric. 2014, 94, 2246–2252. [Google Scholar] [CrossRef]
- Stivala, A.; Carota, G.; Fuochi, V.; Furneri, P.M. Lactobacillus rhamnosus AD3 as a Promising Alternative for Probiotic Product. Biomolecules 2021, 11, 94. [Google Scholar] [CrossRef]
- Handley, P.S.; Harty, D.W.; Wyatt, J.E.; Brown, C.R.; Doran, J.P.; Gibbs, A.C. A comparison of the adhesion, coaggregation and cell-surface hydrophobicity properties of fibrillar and fimbriate strains of Streptococcus salivarius. J. Gen. Microbiol. 1987, 133, 3207–3217. [Google Scholar] [CrossRef]
- Davoodabadi, A.; Soltan Dallal, M.M.; Lashani, E.; Tajabadi Ebrahimi, M. Antimicrobial Activity of Lactobacillus spp. Isolated from Fecal Flora of Healthy Breast-Fed Infants Against Diarrheagenic Escherichia coli. Jundishapur J. Microbiol. 2015, 8, e27852. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.J.; Fishbein, S.R.S.; Dantas, G. Antimicrobial resistance in enteric bacteria: Current state and next-generation solutions. Gut Microbes 2020, 12, 1799654. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Lai, C.C.; Huang, H.L.; Huang, W.Y.; Toh, H.S.; Weng, T.C.; Chuang, Y.C.; Lu, Y.C.; Tang, H.J. Antimicrobial Activity of Lactobacillus Species Against Carbapenem-Resistant Enterobacteriaceae. Front. Microbiol. 2019, 10, 789. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Lai, C.C.; Huang, H.L.; Su, Y.T.; Chiu, Y.H.; Toh, H.S.; Chiang, S.R.; Chuang, Y.C.; Lu, Y.C.; Tang, H.J. Antimicrobial ability and mechanism analysis of Lactobacillus species against carbapenemase-producing Enterobacteriaceae. J. Microbiol. Immunol. Infect. 2021, 54, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Factories 2020, 19, 168. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 6, 1007–1015. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Lee, S.W. Antimicrobial Activity of Ligilactobacillus animalis SWLA-1 and Its Cell-Free Supernatant against Multidrug-Resistant Bacteria and Its Potential Use as an Alternative to Antimicrobial Agents. Microorganisms 2023, 11, 182. [Google Scholar] [CrossRef]
- Lima, A.A.M.; Medeiros, P.H.Q.S.; Havt, A. Enteroaggregative Escherichia coli subclinical and clinical infections. Curr. Opin. Infect. Dis. 2018, 31, 433–439. [Google Scholar] [CrossRef]
- Navarro-Garcia, F.; Elias, W.P. Autotransporters and virulence of enteroaggregative E. coli. Gut Microbes 2011, 2, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Reniero, R.; Cocconcelli, P.; Bottazzi, V.; Morelli, L. High frequency of conjugation in Lactobacillus mediated by an aggregation-promoting factor. J. Gen. Microbiol. 1992, 138, 763–768. [Google Scholar] [CrossRef]
- Schachtsiek, M.; Hammes, W.P.; Hertel, C. Characterization of Lactobacillus coryniformis DSM 20001T surface protein Cpf mediating coaggregation with and aggregation among pathogens. Appl. Environ. Microbiol. 2004, 70, 7078–7085. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque, T.M.R.; Garcia, E.F.; de Oliveira Araújo, A.; Magnani, M.; Saarela, M.; de Souza, E.L. In Vitro Characterization of Lactobacillus Strains Isolated from Fruit Processing By-Products as Potential Probiotics. Probiotics Antimicrob. Protein 2018, 10, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Ghane, M.; Babaeekhou, L.; Ketabi, S.S. Antibiofilm Activity of Kefir Probiotic Lactobacilli Against Uropathogenic Escherichia coli (UPEC). Avicenna J. Med. Biotechnol. 2020, 12, 221–229. [Google Scholar]
- Almeida, M.E.; Pessoa, W.F.B.; Melgaço, A.C.C.; Ramos, L.P.; Rezende, R.P.; Romano, C.C. In vitro selection and characterization of probiotic properties in eight lactobacillus strains isolated from cocoa fermentation. An. Acad. Bras. Cienc. 2022, 94, e20220013. [Google Scholar] [CrossRef]
- Mohanty, D.; Panda, S.; Kumar, S.; Ray, P. In vitro evaluation of adherence and anti-infective property of probiotic Lactobacillus plantarum DM 69 against Salmonella enterica. Microb. Pathog. 2019, 126, 212–217. [Google Scholar] [CrossRef]
- Colautti, A.; Orecchia, E.; Comi, G.; Iacumin, L. Lactobacilli, a Weapon to Counteract Pathogens through the Inhibition of Their Virulence Factors. J. Bacteriol. 2022, 204, e0027222. [Google Scholar] [CrossRef]
- Liu, J.; Hu, D.; Chen, Y.; Huang, H.; Zhang, H.; Zhao, J.; Gu, Z.; Chen, W. Strain-specific properties of Lactobacillus plantarum for prevention of Salmonella infection. Food Funct. 2018, 9, 3673–3682. [Google Scholar] [CrossRef]
- Shim, Y.H.; Lee, S.J.; Lee, J.W. Antimicrobial activity of lactobacillus strains against uropathogens. Pediatr. Int. 2016, 58, 1009–1013. [Google Scholar] [CrossRef]
- Yi, L.; Li, J.; Liu, B.; Wang, Y. Advances in research on signal molecules regulating biofilms. World J. Microbiol. Biotechnol. 2019, 35, 130. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, J.; Qu, J.; Liu, J.; Yin, P.; Zhang, G.; Shang, D. Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 2019, 41, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Menge, C. Molecular Biology of Escherichia coli Shiga Toxins’ Effects on Mammalian Cells. Toxins 2020, 12, 345. [Google Scholar] [CrossRef] [PubMed]
- Lagrafeuille, R.; Miquel, S.; Balestrino, D.; Vareille-Delarbre, M.; Chain, F.; Langella, P.; Forestier, C. Opposing effect of Lactobacillus on in vitro Klebsiella pneumoniae in biofilm and in an in vivo intestinal colonisation model. Benef. Microbes 2018, 9, 87–100. [Google Scholar] [CrossRef]
- Ríos-Muñiz, D.; Cerna-Cortés, J.F.; Morán-García, N.; Meza-Segura, M.; Estrada-García, T. Escherichia coli enterotoxigénica y enteroagregativa: Prevalencia, patogénesis y modelos múridos. Gac. Médica Mex. 2019, 155, 410–416. [Google Scholar] [CrossRef]
- Lee, J.E.; Lee, N.K.; Paik, H.D. Antimicrobial and anti-biofilm effects of probiotic Lactobacillus plantarum KU200656 isolated from kimchi. Food Sci. Biotechnol. 2020, 30, 97–106. [Google Scholar] [CrossRef]
- Fernández, M.S.M.; Rios, J.C.; Vasconcelos, B.M.; Lourenço, M.L.M.C.; Matos, M.N.C.; Cavalcante, R.M.B.; de Almeida, M.V.A.; Costa, R.A.; Carneiro, V.A. Effect of Lactobacillus spp. cell-free supernatant against planktonic growth and biofilm formation of foodborne Escherichia coli isolates. Lett. Appl. Microbiol. 2023, 76, ovac006. [Google Scholar]
- Shi, S.; Gong, L.; Yu, H.; He, G.; Zhang, J.; Han, Y.; Liu, Y.; Hu, J.; Dong, J.; Liu, J.; et al. Antagonistic activity and mechanism of Lactobacillus rhamnosus SQ511 against Salmonella enteritidis. 3 Biotech 2022, 12, 126. [Google Scholar] [CrossRef]
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga Toxin Producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef]
- Navarro-Garcia, F. Enteroaggregative Escherichia coli plasmid-encoded toxin. Future Microbiol. 2010, 5, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Bruballa, A.C.; Shiromizu, C.M.; Bernal, A.M.; Pineda, G.E.; Sabbione, F.; Trevani, A.S.; Bentancor, L.V.; Ramos, M.V.; Fernández-Brando, R.J.; Muñoz, M.J.; et al. Role of Shiga Toxins in Cytotoxicity and Immunomodulatory Effects of Escherichia coli O157:H7 during Host-Bacterial Interactions in vitro. Toxins 2020, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Aditya, A.; Peng, M.; Young, A.; Biswas, D. Antagonistic Mechanism of Metabolites Produced by Lactobacillus casei on Lysis of Enterohemorrhagic Escherichia coli. Front. Microbiol. 2020, 11, 574422. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha-Ramírez, L.M.; Hernández-Chiñas, U.; Moreno-Guerrero, S.S.; Ramírez-Pacheco, A.; Eslava, C.A. In Vitro Effect of the Cell-Free Supernatant of the Lactobacillus casei Strain IMAU60214 against the Different Pathogenic Properties of Diarrheagenic Escherichia coli. Microorganisms 2023, 11, 1324. https://doi.org/10.3390/microorganisms11051324
Rocha-Ramírez LM, Hernández-Chiñas U, Moreno-Guerrero SS, Ramírez-Pacheco A, Eslava CA. In Vitro Effect of the Cell-Free Supernatant of the Lactobacillus casei Strain IMAU60214 against the Different Pathogenic Properties of Diarrheagenic Escherichia coli. Microorganisms. 2023; 11(5):1324. https://doi.org/10.3390/microorganisms11051324
Chicago/Turabian StyleRocha-Ramírez, Luz María, Ulises Hernández-Chiñas, Silvia Selene Moreno-Guerrero, Arturo Ramírez-Pacheco, and Carlos A. Eslava. 2023. "In Vitro Effect of the Cell-Free Supernatant of the Lactobacillus casei Strain IMAU60214 against the Different Pathogenic Properties of Diarrheagenic Escherichia coli" Microorganisms 11, no. 5: 1324. https://doi.org/10.3390/microorganisms11051324