Morphometric and Meristic Characterization of Native Chame Fish (Dormitator latifrons) in Ecuador Using Multivariate Analysis

 , , , , , and
, , , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sample and Data Collection
2.2. Body Measurements
2.3. Fulton’s Condition Factor (K)
2.4. Statistical Analysis
3. Results and Discussion
3.1. Morphometric and Meristic Characteristics
3.2. Relationships Among Morphometric Measurements
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loor Risco, O. El Chame-Dormitator Latifrons- una Opción de Vida Para las Comunidades de Escasos Recursos Económicos en la Costa Ecuatoriana. 2000. Available online: https://es.scribd.com/document/229615586/EL-CHAME (accessed on 5 August 2020).
- Revelo, W. Aspectos biológicos y pesqueros de las principales especies de peces en el sistema hídrico de la provincia de los Ríos, durante 2009. Bol. Cient Tec. 2010, 20, 53–84. [Google Scholar]
- IUCN. Red List of Treatened Species.Version. Available online: www.iucnredlist.org.2011 (accessed on 31 July 2020).
- Barriga, R. Lista de peces de agua dulce e intermareales del Ecuador. Rev. Politécnica 2012, 30, 83–119. [Google Scholar]
- López Vera, M.R. Caracterización del Chame (Dormitator latrifrons) de la Subcuenca del Carrizal. Manabí, Ecuador. Master’s Thesis, University of Córdoba, Córdoba, Spain, 2017. [Google Scholar]
- Rodríguez, J.; Angón, E.; González, M.; Perea, J.; Barba, C.; García, A. Allometric relationship and growth models of juveniles of Cichlasoma festae (Perciforme: Cichlidae), a freshwater species native in Ecuador. Rev. Biol. Trop. 2017, 65, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- SOFIA. El estado mundial de la pesca y la acuicultura. Organización de las Naiones Unidas para la Agricultura y la Alimentación. FAO 2018, 250, 187–217. [Google Scholar]
- FAO. El estado Mundial de la Pesca y la Acuicultura 2012, Págs. 206–213. Roma. 231 págs. Available online: www.fao.org/docrep/016/i2727s/i2727s00.htm (accessed on 5 August 2020).
- Haz Alvarado, M. Producción y Exportación del Chame, Como Nueva Alternativa Comercial del Ecuador. Bachelor’s Thesis, ESCUELA SUPERIOR POLITECNICA DEL LITORAL (ESPOL), Guayaquil, Ecuador, 2002. [Google Scholar]
- Lopez, M.R.; Gonzalez, A.; Bellido, L.; Caez, J.; Gonzalez, M.; Angon, E.; Garcia, A. Modelizacion del crecimiento de Chame (dorminator latifrons) en edad juvenil en estanques de geomembrana con suelo. In Proceedings of the XII Congreso de la Federación Iberoamericana de Razas Criollas y Autóctonas, Veracruz, Mexico, 9–12 October 2019; pp. 267–271. [Google Scholar]
- Gonzalez, A.; Caez, J.; Rodriguez, J.; Machuca, C.; Gonzalez, M.; Lopez, M.; Garcia, A. Modelizacion del crecimiento de Chame (dorminator latifrons) en edad juvenil con tenología biofloc (BFT). In Proceedings of the XII Congreso de la Federación Iberoamericana de Razas Criollas y Autóctonas, Veracruz, Mexico, 9–12 October 2019; pp. 267–271. [Google Scholar]
- ECOCOSTAS. Estudio de Factibilidad Para la Implementación de un Centro de Capacitación Para el Cultivo de Chame en el Estuario del Río Cojimíes. 2006. Available online: http://www.tedein.com.ec/ecocostas/www.ecocostas.org/success/images/documentos/1236092848_FactibilidadCapacitacionCHAME.pdf (accessed on 15 August 2020).
- FAO. Phenotypic Characterization of Animal Genetic re-Sources; No.11; FAO Animal Production and Health Guidelines: Rome, Italy, 2012. [Google Scholar]
- González, M.A.; Rodriguez, J.M.; Angón, E.; Martínez, A.; Garcia, A.; Peña, F. Characterization of morphological and meristic traits and their variations between two different populations (wild and cultured) of Cichlasoma festae, a species native to tropical Ecuadorian rivers. Archiv. Anim. Breed. 2016, 59, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Caez, J.; González, A.; González, M.A.; Angón, E.; Rodríguez, J.M.; Peña, F.; Barca, C.; García, A. Application of multifactorial discriminant analysis in the morphostructural differentiation of wild and cultured populations of Vieja Azul (Andinoacara rivulatus). Turk. J. Zool. 2019, 4, 516–530. [Google Scholar] [CrossRef]
- Konings, A. Cichlids de América Central; TFH Publication, Inc.: Neptune, NJ, USA, 1989. [Google Scholar]
- Canadian Council on Animal Care. Guidelines on: The Care and Use of Fish Research, Teaching and Testing. 2005. Available online: https://www.ccac.ca/Documents/Standards/Guidelines/Fish.pdf (accessed on 31 July 2020).
- Diodatti, F.C.; Fonseca de Freitas, R.T.; Freato, T.A.; Pérez Ribeiro, P.A.; Solis Murgas, L.D. Parámetros morfométricos en el rendimiento de los componentes corporales de tilapia del Nilo (Oreochromis Niloticus). An. Vet. 2008, 24, 45–55. [Google Scholar]
- Turan, C.; Oral, M.; Öztürk, B.; Düzgüne, E. Morphometric and meristic variation between stocks of Bluefish (Pomatomus saltatrix) in the Black, Marmara, Aegean and northeastern Mediterranean Seas. Fish. Res. 2006, 79, 139–147. [Google Scholar] [CrossRef]
- Elliott, N.G.; Haskard, K.; Koslow, J.A. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. J. Fish Biol. 1995, 46, 202–220. [Google Scholar] [CrossRef]
- Tudela, S. Morphological variability in a Mediterranean, genetically homogeneous population of the European anchovy, Engraulis encrasicolus. Fish. Res. 1999, 42, 229–243. [Google Scholar] [CrossRef]
- Reist, J.D. An empirical evaluation of several univariate methods that adjust for size variation in morphometric variation. Can. J. Zool. 1985, 63, 1429–1439. [Google Scholar] [CrossRef]
- Florencio, A.; Serrano, M. Algunos aspectos sobre la biologia del chame, Dormitator latifrons. Rev. Cienc. Mar. Limnol. 1981, 1, 73–81. [Google Scholar]
- Sasi, H.; Berber, S. Age, growth and some biological characteristics of white bream (Blicca bjoerk na L., 1758) in Uluabat lake, in Northwestern of Anatolia. Asian J. Anim. Vet. Adv. 2012, 7, 262–267. [Google Scholar] [CrossRef]
- Solomon, S.O.; Okomoda, V.T.; Ogbenyikwu, A.I. Intraspecific morphological variation between cultured and wild Clarias gariepinus (Burchell) (Clariidae.; Siluriformes). Arch. Pol. Fisheries 2015, 23, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Wimberger, P.H. Plasticity of fish body shape, the effects of diet, development, family and age in two species of Geophagus (Pisces: Cichlidae). Biol. J. Linn. Soc. 1992, 45, 197–218. [Google Scholar] [CrossRef]
- Lowe-McConnell, R.H. Fish Communities in Tropical Freshwaters; Longman: London, UK, 1975; p. 331. [Google Scholar]
- Welcomme, R.L. Fisheries Ecology qf Floodplain Rivers; Longman: London, UK, 1979; p. 317. [Google Scholar]
- Chang, B.D. Seasonal variations in growth, condition and gonads of Dormitator Zatifvons (Richardson) in the Chone River Basin, Ecuador. J. Fish Biol. 1984, 24, 637–648. [Google Scholar] [CrossRef]
- Ujjania, N.C.; Kohli, M.P.S. Landmark-based morphometric analysis for selected species of indian major carp (catla catla.; ham. 1822). Int. J. Food Agric. Vet. Sci. 2011, 1, 64–74. [Google Scholar]
- Ochoa Ubilla, B.Y.; Mendoza Nieto, K.X.; Vivas Moreira, R.; Zambrano, J.U.; Ferrer-Sánchez, Y. Ecuador Structure of catch sizes and length-weight ratio of native fish in the Abras de Mantequilla wetland, Ecuador (in Spanish). Cien. Tecnol. UTEQ 2016, 9, 19–27. [Google Scholar]
- Turan, C.; Yalcin, S.; Turan, F.; Okur, E.; Akyurt, I. Morphometric comparisons of African catfish, Clarias gariepinus populations in Turkey. Folia Zool. Praha 2005, 54, 165–172. [Google Scholar]
- Vreven, E.J.; Adèpo-Gourène, B.; Agnèse, J.F.; Teugels, G.G. Morphometric and allozyme in populations and cultured strains of the Nile tilapia k (Teleostei, Cichlidae). Belg. J. Zool. 1998, 128, 23–34. [Google Scholar]
- King, M. Review of “Fisheries Biologya, Assessment and Managenement, Second edition. Rev. Fish. Biol. Fish. 2008, 8, 451–452. [Google Scholar]
- Rodriguez Montes de Oca, G.; Medina-Hernández, E.; Velázquez-Sandoval, J.; López-López, V.; Reyes, J.C.; Dabrowski, K.; Haws, M. Production of “Chame” (Dormitator latifrons, Pisces: Eleotridae) larvae using GnRHa and LHRHa. Rev. Colom. Cienc. Pecua. 2012, 25, 422–429. [Google Scholar]
- Stearns, S.C. The genetic basis of differences in life history traits among 6 populations of mosquitofish (Gambusia affinis) that shared ancestors in 1905. Evolution 1983, 37, 618–627. [Google Scholar] [PubMed]
- Martínez-Lendech, N.; Martínez-Falcón, A.P.; Schmitter-Soto, J.J.; Mejía-Mojica, H.; Sorani-Dalbón, V.; Cruz-Ruíz, G.I.; Mercado-Silva, N. Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico. Diversity 2020, 12, 187. [Google Scholar] [CrossRef]



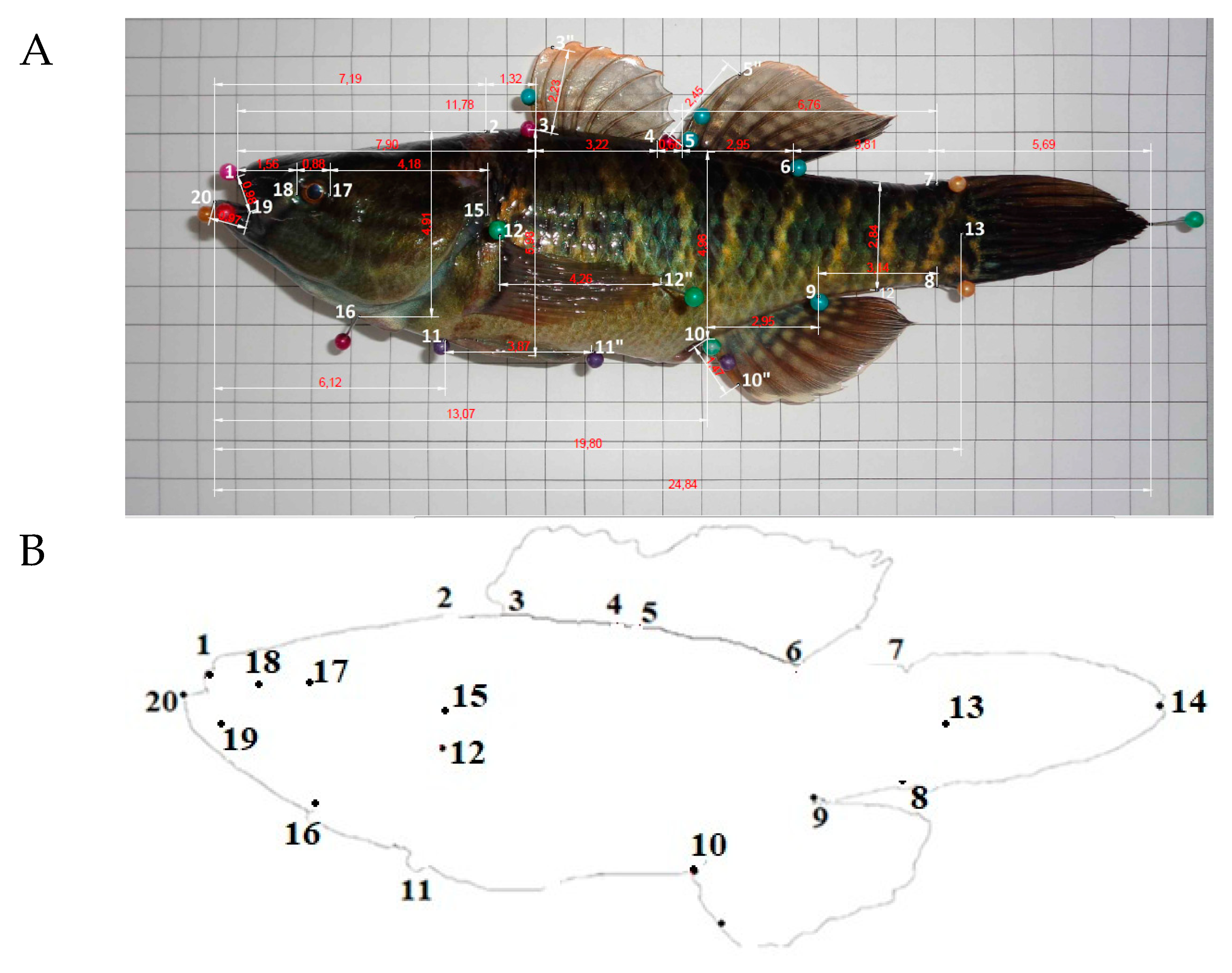{kind=link}
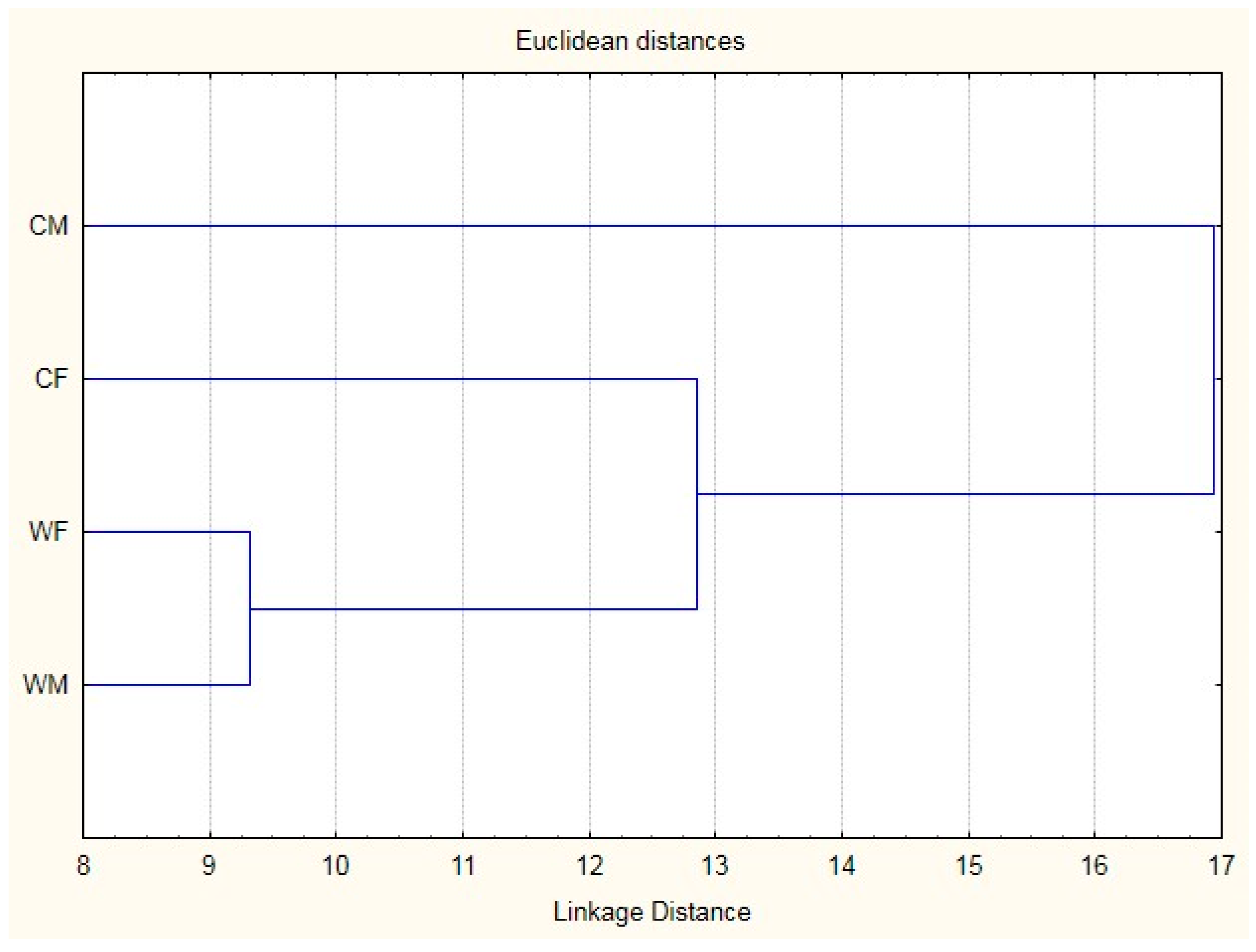{kind=link}
| Character | Description | Acronyms |
|---|---|---|
| Body weight | Total weight including gut and gonads | BW |
| Total length | Tip of the upper jaw to the caudal end of the caudal fin | TL |
| Standard length | Tip of the upper jaw to the tail base | SL |
| Head length | From the front of the upper lip to the posterior end of the opercula membrane | HL |
| Eye diameter | The greatest bony diameter of the orbit | ED |
| Preorbital length | Front of the upper lip to cranial eye edge | Pre-OL |
| Predorsal length | Front of the upper lip to the origin of the dorsal fin | Pre-DL |
| Prepectoral length | Front of the upper lip to the origin of the pectoral fin | Pre-PcL |
| Preanal length | Front of the upper lip to the origin of the anal fin | Pre-AL |
| Dorsal fin length | From base of first dorsal spine to base of last dorsal ray | DFL |
| Dorsal fin ray length | From base to tip of the fifth dorsal ray | DFRL |
| Pectoral fin length | From base to tip of the pectoral fin | PcFL |
| Pelvic fin length | From base to tip of the pelvic fin | PvFL |
| Anal fin length | From base of first anal spine to base of last anal ray | AFL |
| Anal fin ray length | From base to tip of the last anal ray | AFRL |
| Upper jaw length | Straight line measurement between the snout tip and posterior edge of maxilla | UJL |
| Body depth 1 | Body depth at the level of the first ray of the dorsal fin | AC1 |
| Body depth 2 | Body depth at the level of the first ray of the anal fin | AC2 |
| Body depth 3 | Body depth at the level of the first radius of the caudal fin | AC3 |
| Body perimeter 1 | Body perimeter at the level of the first ray of the dorsal fin | P1 |
| Body perimeter 2 | Body perimeter at the level of the first radius of the anal fin | P2 |
| Body perimeter 3 | Body perimeter at the level of the last ray of the dorsal fin | P3 |
| Body width 1 | Straight line measurement from side to side at the level of the base of first dorsal spine | LC1 |
| Body width 2 | Straight line measurement from side to side at the level of the base of first anal spine | LC2 |
| Body width 3 | Straight line measurement from side to side at the level of the base of last dorsal ray | LC3 |
| Body width 4 | Straight line measurement from side to side at the level of the base of last anal ray | LC4 |
| Dorsal fin rays | Number of thorns in the dorsal fin | DFR |
| Radius dorsal fin | Number of cartilages found in the space between thorns from the dorsal fin | RDF |
| Pectoral fin rays | Number of thorns in the pectoral fin | PcFR |
| Radius pectoral fin | Number of cartilages found in the space between thorns in the pectoral fin | RPcF |
| Pelvic fin rays | Number of thorns in the pelvic fin | PvFR |
| Radius pelvic fin | Number of cartilages found in the space between thorns in the pelvic fin | RPvF |
| Anal fin rays | Number of thorns in the anal fin | AFR |
| Radius anal fin | Number of cartilages found in the space between thorns in the anal fin | RAF |
| Caudal fin rays | Number of thorns in the caudal fin | CFR |
| Radius caudal fin | Number of cartilages found in the space between thorns in the caudal fin | RCF |
| Character 1 | All (N = 300) | Cultured (N = 150) | Wild (N = 150) | Males (N = 153) | Females (N = 147) | p Value 2 | ||
|---|---|---|---|---|---|---|---|---|
| System Production (A) | Sex (B) | A × B | ||||||
| BW (g) | 173.13 (31.00) | 209.89 (16.8) | 136.38 (31.27) | 176.89 (33.26) | 169.22 (28.37) | 88.57 *** | 1.37 ns | 0.98 ns |
| K | 2.41 (23.23) | 2.48 (30.21) | 2.35 (10.76) | 2.48 (27.30) | 2.34 (17.10) | 1.48 ns | 1.57 ns | 1.99 ns |
| TL (cm) | 24.65 (12.93) | 26.46 (9.54) | 22.85 (11.99) | 24.78 (13.35) | 24.53 (12.59) | 46.38 *** | 0.37 ns | 0.01 ns |
| SL (cm) | 19.23 (13.24) | 20.65 (9.97) | 17.81 (12.22) | 19.22 (13.72) | 19.24 (12.87) | 44.11 *** | 0.00 ns | 0.05 ns |
| HL (cm) | 6.47 (15.88) | 6.93 (12.61) | 6.00 (16.02) | 6.66 (16.61) | 6.26(14.42) | 27.05 *** | 5.39 * | 0.01 ns |
| ED (cm) | 0.88 (18.86) | 0.98 (17.14) | 0.78 (11.33) | 0.87 (22.42) | 0.88 (14.56) | 52.64 *** | 0.01 ns | 0.80 ns |
| Pre-OL (cm) | 1.21 (20.23) | 1.31 (18.28) | 1.11 (18.69) | 1.23 (20.78) | 1.18 (19.54) | 20.68 *** | 1.67 ns | 0.60 ns |
| Pre-DL_1 (cm) | 7.68 (13.88) | 8.25 (10.95) | 7.11 (12.75) | 7.64 (14.65) | 7.72 (13.18) | 38.99 *** | 0.13 ns | 0.67 ns |
| Pre-DL_2 (cm) | 11.53 (12.99) | 12.33 (10.49) | 10.73 (11.61) | 11.42 (13.71) | 11.63 (12.3) | 39.24 *** | 0.48 ns | 0.41 ns |
| Pre-PcL (cm) | 6.45 (13.77) | 6.88 (11.09) | 6.02 (13.19) | 6.55 (14.77) | 6.34 (12.47) | 31.20 *** | 2.25 ns | 0.07 ns |
| Pre-AL (cm) | 12.95 (14.67) | 15.91 (77.75) | 11.74 (11.71) | 12.89 (15.88) | 14.81 (85.69) | 5.61 *** | 1.09 ns | 0.97 ns |
| DFL_1 (cm) | 3.08 (15.26) | 3.30 (13.85) | 2.85 (12.82) | 3.10 (15.73) | 3.06 (14.87) | 29.03 *** | 0.33 ns | 0.03 ns |
| DFL_2 (cm) | 0.77 (30.16) | 0.78 (33.66) | 0.76 (26.34) | 0.69 (26.54) | 0.85 (29.52) | 0.08 ns | 12.68 ** | 0.00 ns |
| DFRL_1 (cm) | 2.22 (18.50) | 2.40 (16.97) | 2.04 (16.43) | 2.27 (20.95) | 2.17 (15.17) | 22.23 *** | 1.79 ns | 0.21 ns |
| DFRL _2 (cm) | 2.48 (19.69) | 2.72 (19.02) | 2.24 (13.94) | 2.64 (20.73) | 2.31 (15.08) | 39.10 *** | 19.74 *** | 5.39 * |
| PcFL (cm) | 4.51 (15.45) | 4.92 (12.07) | 4.09 (12.85) | 4.55 (16.70 | 4.46 (14.06) | 54.24 *** | 1.00 ns | 0.01 ns |
| PvFL (cm) | 4.01 (17.86) | 4.49 (12.64) | 3.53 (13.73) | 4.15 (19.04) | 3.86 (15.61) | 92.34 *** | 9.27 ** | 1.30 ns |
| AFL (cm) | 2.75 (19.09) | 2.75 (18.91) | 2.76 (19.46) | 2.85 (18.26) | 2.65 (19.45) | 0.00 ns | 3.78 ns | 0.30 ns |
| AFRL (cm) | 2.03 (35.80) | 2.54 (25.35) | 1.53 (23.74) | 2.11 (34.48) | 1.95 (37.20) | 96.13 *** | 3.09 ns | 0.60 ns |
| UJL (cm) | 1.12 (20.18) | 1.21 (19.77) | 1.03 (16.78) | 1.14 (21.01) | 1.09 (19.07) | 18.41 *** | 1.93 ns | 0.16 ns |
| AC1 (cm) | 6.45 (17.92) | 7.34 (9.74) | 5.57 (13.85) | 6.45 (18.15) | 6.46 (17.85) | 139.49 *** | 0.03 ns | 0.48 ns |
| AC2 (cm) | 5.35 (14.81) | 5.81 (10.62) | 4.88 (13.8) | 5.38 (15.2) | 5.31 (14.51) | 51.23 *** | 0.49 ns | 1.33 ns |
| AC3 (cm) | 2.93 (14.14) | 3.17 (10.8) | 2.69 (12.27) | 2.92 (14.51) | 2.94 (13.9) | 52.72 *** | 0.08 ns | 1.93 ns |
| P1 (cm) | 14.85 (13.63) | 16.22 (7.45) | 13.47 (12.76) | 14.90 (13.67) | 14.79(13.72) | 84.33 *** | 0.33 ns | 0.13 ns |
| P2 (cm) | 12.23 (14.58) | 13.49 (8.18) | 10.98 (12.87) | 12.41 (15.23) | 12.05 (13.81) | 100.02 *** | 2.68 ns | 1.05 ns |
| P3 (cm) | 6.79 (11.97) | 7.06 (7.29) | 6.52 (14.74) | 6.83 (9.47) | 6.75 (14.24) | 11.91 ** | 0.34 ns | 0.08 ns |
| LC1 (cm) | 3.29 (17.75) | 3.77 (5.56) | 2.81 (15.12) | 3.32 (17.61) | 3.26 (18.03) | 202.69 *** | 1.62 ns | 0.07 ns |
| LC2 (cm) | 2.89 (13.95) | 3.16 (6.30) | 2.61 (14.15) | 2.83 (13.68) | 2.94 (14.05) | 84.37 *** | 3.14 ns | 0.11 ns |
| LC3 (cm) | 2.10 (16.16) | 2.32 (10.23) | 1.88 (14.83) | 2.09 (17.20) | 2.11 (15.17) | 70.30 *** | 0.05 ns | 0.00 ns |
| LC4 (cm) | 1.14 (21.33) | 1.19 (25.89) | 1.11 (14.03) | 1.11 (10.78) | 1.19 (27.49) | 2.40 ns | 2.16 ns | 3.19 ns |
| Character 1 | All (N = 300) | Cultured (N = 150) | Wild (N = 150) | Males (N = 153) | Females (N = 147) | p Value 2 | ||
|---|---|---|---|---|---|---|---|---|
| System Production (A) | Sex (B) | A × B | ||||||
| DFR-1 | 6.98 (2.02) | 6.98 (2.03) | 6.98 (2.03) | 6.98 (2.01) | 6.98 (2.05) | n.a | 0.81 ns | 1.94 ns |
| RDF-1 | 5.98 (2.35) | 5.98 (2.36) | 5.98 (2.36) | 5.98 (2.34) | 5.98 (2.39) | n.a | 0.81 ns | 1.94 ns |
| DFR-2 | 9.98 (2.00) | 10.02 (1.41) | 9.94 (2.41) | 9.98 (1.40) | 9.98 (2.50) | 4.00 * | 0.93 ns | 5.12 ns |
| RDF-2 | 8.98 (2.23) | 9.02 (1.57) | 8.94 (2.68) | 8.98 (1.56) | 8.98 (2.77) | 4.00 * | 0.93 ns | 5.12 ns |
| PcRF | 15.04 (3.26) | 15.16 (3.62) | 14.92 (2.65) | 14.94 (3.11) | 15.14 (3.30) | 6.12 * | 4.24 * | 10.40 * |
| RPcF | 14.04 (3.50) | 14.16 (3.87) | 13.92 (2.84) | 13.94 (3.34) | 14.14 (3.54) | 6.12 * | 4.24 * | 10.40 * |
| PvRF | 5.00 (0) | 5 (0) | 5 (0) | 5 (0) | 5 (0) | n.a | n.a | n.a |
| RPvF | 4.00 (0) | 4 (0) | 4 (0) | 4 (0) | 4 (0) | n.a | n.a | n.a |
| AFR | 10.84 (4.09) | 10.70 (4.72) | 10.98 (2.90) | 10.78 (3.85) | 10.90 (4.29) | 10.31 ** | 1.44 ns | 15.56 ** |
| RAF | 9.86 (4.56) | 9.74 (5.41) | 9.98 (3.19) | 9.82 (4.42) | 9.90 (4.72) | 7.59 ** | 0.61 ns | 10.41 * |
| CFR | 15.04 (3.12) | 15.06 (3.65) | 15.02 (2.51) | 15.14 (3.24) | 14.94 (2.87) | 0.49 ns | 4.36 * | 5.57 ns |
| RCF | 14.04 (3.35) | 14.06 (3.91) | 14.02 (2.69) | 14.14 (3.47) | 13.94 (3.07) | 0.49 ns | 4.36 * | 5.47 ns |
| Character 1 | All (N = 300) | Cultured (N = 150) | Wild (N = 150) | Males (N = 153) | Females (N = 147) | p Value 2 | ||
|---|---|---|---|---|---|---|---|---|
| System Production (A) | Sex (B) | A × B | ||||||
| BW/SL | 8.84 (23.81) | 10.19 (15.38) | 7.48 (22.14) | 9.03 (26.05) | 8.63 (20.97) | 69.70 *** | 2.10 ns | 1.76 ns |
| TL/SL | 1.28 (1.64) | 1.28 (1.80) | 1.28 (1.47) | 1.29 (1.69) | 1.28 (1.35) | 0.11 ns | 14.54 *** | 0.68 ns |
| HL/SL | 0.34 (7.69) | 0.34 (7.62) | 0.34 (7.83) | 0.35 (7.28) | 0.33 (6.83) | 0.00 ns | 17.48 *** | 0.05 ns |
| ED/SL | 0.05 (15.62) | 0.05 (14.87) | 0.04 (15.89) | 0.05 (17.39) | 0.05 (13.75) | 4.21 * | 0.15 ns | 0.90 ns |
| Pre-OL/SL | 0.06 (17.58) | 0.06 (17.08) | 0.06 (18.21) | 0.06 (16.15) | 0.06 (18.98) | 0.16 ns | 1.60 ns | 0.42 ns |
| Pre-DL_1/SL | 0.40 (4.25) | 0.40 (4.69) | 0.40 (3.79) | 0.40 (4.36) | 0.40 (4.09) | 0.01 ns | 1.33 ns | 2.74 ns |
| Pre-DL_2/SL | 0.60 (2.91) | 0.60 (3.48) | 0.60 (2.12) | 0.59 (2.84) | 0.61 (2.74) | 4.28 * | 9.33 ** | 2.22 ns |
| Pre-PcL/SL | 0.34 (6.65) | 0.33 (7.09) | 0.34 (6.19) | 0.34 (5.96) | 0.33 (7.03) | 0.87 ns | 5.59 * | 0.12 ns |
| Pre-AL/SL | 0.67 (4.76) | 0.69 (4.87) | 0.66 (3.78) | 0.67 (5.35) | 0.68 (4.06) | 18.81 *** | 1.30 ns | 0.64 ns |
| DFL_1/SL | 0.16 (8.48) | 0.16 (9.56) | 0.16 (7.34) | 0.16 (8.06) | 0.16 (8.94) | 0.13 ns | 0.73 ns | 0.05 ns |
| DFL_2/SL | 0.04 (29.66) | 0.04 (32.63) | 0.04 (25.81) | 0.04 (26.63) | 0.04 (28.82) | 5.86 * | 13.74 *** | 0.00 ns |
| DFRL_1/SL | 0.12 (12.51) | 0.12 (14.56) | 0.11 (10.1) | 0.12 (14.24) | 0.11 (9.97) | 0.24 ns | 2.57 ns | 0.70 ns |
| DFRL_2/SL | 0.13(13.43) | 0.13 (16.54) | 0.13 (8.43) | 0.14 (13.00) | 0.12 (9.85) | 4.75 * | 33.16 *** | 9.03 ** |
| PcFL/SL | 0.23(7.64) | 0.24 (7.78) | 0.23 (7.12) | 0.24 (7.84) | 0.23 (7.35) | 4.80 * | 2.57 ns | 0.35 ns |
| PvFL/SL | 0.21 (10.62) | 0.22 (10.39) | 0.20(8.32) | 0.22 (10.69) | 0.20 (9.27) | 28.65 *** | 17.75 *** | 3.32 ns |
| AFL/SL | 0.14 (15.35) | 0.13 (13.96) | 0.15 (12.89) | 0.15 (15.00) | 0.14 (14.77) | 34.28 *** | 9.05 ** | 0.59 ns |
| AFRL/SL | 0.11 (29.76) | 0.12 (21.59) | 0.09 (28.78) | 0.11 (29.45) | 0.10 (29.69) | 47.93 *** | 3.46 ns | 0.52 ns |
| UJL/SL | 0.06 (17.46) | 0.06 (17.02) | 0.06 (18.05) | 0.06 (15.69) | 0.06 (19.17) | 0.00 ns | 1.60 ns | 0.34 ns |
| AC1/SL | 0.33 (9.51) | 0.36 (7.55) | 0.31 (5.87) | 0.33 (9.17) | 0.33 (9.96) | 87.57 *** | 0.13 ns | 0.38 ns |
| AC2/SL | 0.28 (7.30) | 0.28 (7.83) | 0.27 (6.49) | 0.28 (7.17) | 0.28 (7.43) | 3.25 ns | 1.63 ns | 2.60 ns |
| AC3/SL | 0.15(5.64) | 0.15 (6.21) | 0.15 (4.88) | 0.15 (5.39) | 0.15 (5.93) | 2.29 ns | 0.23 ns | 5.28 * |
| P1/SL | 0.77 (9.72) | 0.79 (11.82) | 0.76 (5.98) | 0.78 (10.3) | 0.77 (9.12) | 5.09 * | 0.59 ns | 0.57 ns |
| P2/SL | 0.64 (10.23) | 0.66 (11.95) | 0.62 (6.34) | 0.65 (11.23) | 0.63 (8.79) | 10.78 ** | 3.29 ns | 2.08 ns |
| P3/SL | 0.36 (11.50) | 0.34 (10.37) | 0.37 (11.63) | 0.36 (9.56) | 0.35 (13.33) | 8.92 ** | 0.42 ns | 0.29 ns |
| LC1/SL | 0.17 (12.18) | 0.18 (9.84) | 0.16 (8.99) | 0.17 (13.06) | 0.17 (11.11) | 66.14 *** | 1.84 ns | 0.00 ns |
| LC2/SL | 0.15 (8.62) | 0.15 (9.47) | 0.15 (6.79) | 0.15 (8.38) | 0.15 (8.54) | 8.13 ** | 4.52 * | 0.04 ns |
| LC3/SL | 0.11 (10.39) | 0.11 (11.08) | 0.11 (8.24) | 0.11 (11.47) | 0.11 (9.25) | 10.81 *** | 0.05 ns | 0.21 ns |
| LC4/SL | 0.06 (23.59) | 0.06 (32.8) | 0.06 (9.58) | 0.06 (12.32) | 0.06 (30.38) | 2.33 ns | 1.75 ns | 2.34 ns |
| Character 1 | All (N = 300) | Cultured (N = 150) | Wild (N = 150) | Males (N = 153) | Females (N = 147) | p Value 2 | ||
|---|---|---|---|---|---|---|---|---|
| System Production (A) | Sex (B) | A × B | ||||||
| BW (g) | 8.84 (23.81) | 10.19 (15.38) | 7.48 (22.14) | 9.03 (26.05) | 8.63 (20.97) | 69.70 *** | 2.10 ns | 1.76 ns |
| K | 0.13 (32.16) | 0.12 (41.73) | 0.13 (20.41) | 0.13 (36.33) | 0.13 (26.34) | 1.55 ns | 1.05 ns | 1.66 ns |
| TL (cm) | 1.28 (1.64) | 1.28 (1.80) | 1.28 (1.47) | 1.29 (1.69) | 1.28 (1.35) | 0.11 ns | 14.54 *** | 0.68 ns |
| HL (cm) | 0.34 (7.69) | 0.34 (7.62) | 0.34 (7.83) | 0.35 (7.28) | 0.33 (6.83) | 0.00 ns | 17.48 *** | 0.05 ns |
| ED (cm) | 0.05 (15.62) | 0.05 (14.87) | 0.04 (15.89) | 0.05 (17.39) | 0.05 (13.75) | 4.21 * | 0.15 ns | 0.90 ns |
| Pre-OL (cm) | 0.06 (17.58) | 0.06 (17.08) | 0.06 (18.21) | 0.06 (16.15) | 0.06 (18.98) | 0.16 ns | 1.60 ns | 0.42 ns |
| Pre-DL_1 (cm) | 0.40 (4.25) | 0.40 (4.69) | 0.40 (3.79) | 0.40 (4.36) | 0.40 (4.09) | 0.01 ns | 1.33 ns | 2.74 ns |
| Pre-DL_2 (cm) | 0.60 (2.91) | 0.60 (3.48) | 0.60 (2.12) | 0.59 (2.84) | 0.61 (2.74) | 4.28 * | 9.33 ** | 2.22 ns |
| Pre-PcL (cm) | 0.34 (6.65) | 0.33 (7.09) | 0.34 (6.19) | 0.34 (5.96) | 0.33 (7.03) | 0.87 ns | 5.59 * | 0.12 ns |
| Pre-AL (cm) | 0.67 (4.76) | 0.69 (4.87) | 0.66 (3.78) | 0.67 (5.35) | 0.68 (4.06) | 18.81 *** | 1.30 ns | 0.64 ns |
| DFL_1 (cm) | 0.16 (8.48) | 0.16 (9.56) | 0.16 (7.34) | 0.16 (8.06) | 0.16 (8.94) | 0.13 ns | 0.73 ns | 0.05 ns |
| DFL_2 (cm) | 0.04 (29.66) | 0.04 (32.63) | 0.04 (25.81) | 0.04 (26.63) | 0.04 (28.82) | 5.86 * | 13.74 *** | 0.00 ns |
| DFRL_1 (cm) | 0.12 (12.51) | 0.12 (14.56) | 0.11 (10.1) | 0.12 (14.24) | 0.11 (9.97) | 0.24 ns | 2.57 ns | 0.70 ns |
| DFRL _2 (cm) | 0.13 (13.43) | 0.13 (16.54) | 0.13 (8.43) | 0.14 (13.00) | 0.12 (9.85) | 4.75 * | 33.16 *** | 9.03** |
| PcFL (cm) | 0.23 (7.64) | 0.24 (7.78) | 0.23 (7.12) | 0.24 (7.84) | 0.23 (7.35) | 4.80 * | 2.57 ns | 0.35 ns |
| PvFL (cm) | 0.21 (10.62) | 0.22 (10.39) | 0.20 (8.32) | 0.22 (10.69) | 0.20 (9.27) | 28.65 *** | 17.75 *** | 3.32 ns |
| AFL (cm) | 0.14 (15.35) | 0.13 (13.96) | 0.15 (12.89) | 0.15 (15.00) | 0.14 (14.77) | 34.28 *** | 9.05 ** | 0.59 ns |
| AFRL (cm) | 0.11 (29.76) | 0.12 (21.59) | 0.09 (28.78) | 0.11 (29.45) | 0.10 (29.69) | 47.93 *** | 3.46 ns | 0.52 ns |
| UJL (cm) | 0.06 (17.46) | 0.06 (17.02) | 0.06 (18.05) | 0.06 (15.69) | 0.06 (19.17) | 0.00 ns | 1.60 ns | 0.34 ns |
| AC1 (cm) | 0.33 (9.51) | 0.36 (7.55) | 0.31 (5.87) | 0.33 (9.17) | 0.33 (9.96) | 87.57 *** | 0.13 ns | 0.38 ns |
| AC2 (cm) | 0.28 (7.30) | 0.28 (7.83) | 0.27 (6.49) | 0.28 (7.17) | 0.28 (7.43) | 3.25 ns | 1.63 ns | 2.60 ns |
| AC3 (cm) | 0.15 (5.64) | 0.15 (6.21) | 0.15 (4.88) | 0.15 (5.39) | 0.15 (5.93) | 2.29 ns | 0.23 ns | 5.28* |
| P1 (cm) | 0.77 (9.72) | 0.79 (11.82) | 0.76 (5.98) | 0.78 (10.3) | 0.77 (9.12) | 5.09 * | 0.59 ns | 0.57 ns |
| P2 (cm) | 0.64 (10.23) | 0.66 (11.95) | 0.62 (6.34) | 0.65 (11.23) | 0.63 (8.79) | 10.78 ** | 3.29 ns | 2.08 ns |
| P3 (cm) | 0.36 (11.50) | 0.34 (10.37) | 0.37 (11.63) | 0.36 (9.56) | 0.35 (13.33) | 8.92 ** | 0.42 ns | 0.29 ns |
| LC1 (cm) | 0.17 (12.18) | 0.18 (9.84) | 0.16 (8.99) | 0.17 (13.06) | 0.17 (11.11) | 66.14 *** | 1.84 ns | 0.00 ns |
| LC2 (cm) | 0.15 (8.62) | 0.15 (9.47) | 0.15 (6.79) | 0.15 (8.38) | 0.15 (8.54) | 8.13 ** | 4.52 * | 0.04 ns |
| LC3 (cm) | 0.11 (10.39) | 0.11 (11.08) | 0.11 (8.24) | 0.11 (11.47) | 0.11 (9.25) | 10.81 ** | 0.05 ns | 0.21 ns |
| LC4 (cm) | 0.06 (23.59) | 0.06 (32.8) | 0.06 (9.58) | 0.06 (12.32) | 0.06 (30.38) | 2.33 ns | 1.75 ns | 2.34 ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Martinez, A.; Lopez, M.; Molero, H.M.; Rodriguez, J.; González, M.; Barba, C.; García, A. Morphometric and Meristic Characterization of Native Chame Fish (Dormitator latifrons) in Ecuador Using Multivariate Analysis. Animals 2020, 10, 1805. https://doi.org/10.3390/ani10101805
Gonzalez-Martinez A, Lopez M, Molero HM, Rodriguez J, González M, Barba C, García A. Morphometric and Meristic Characterization of Native Chame Fish (Dormitator latifrons) in Ecuador Using Multivariate Analysis. Animals. 2020; 10(10):1805. https://doi.org/10.3390/ani10101805
Chicago/Turabian StyleGonzalez-Martinez, Ana, Mario Lopez, Hebert Mario Molero, Jorge Rodriguez, Martin González, Cecilio Barba, and Anton García. 2020. "Morphometric and Meristic Characterization of Native Chame Fish (Dormitator latifrons) in Ecuador Using Multivariate Analysis" Animals 10, no. 10: 1805. https://doi.org/10.3390/ani10101805