
The Impact of Sea Ice Loss on Benthic Communities of the Makarov Strait (Northeastern Barents Sea)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
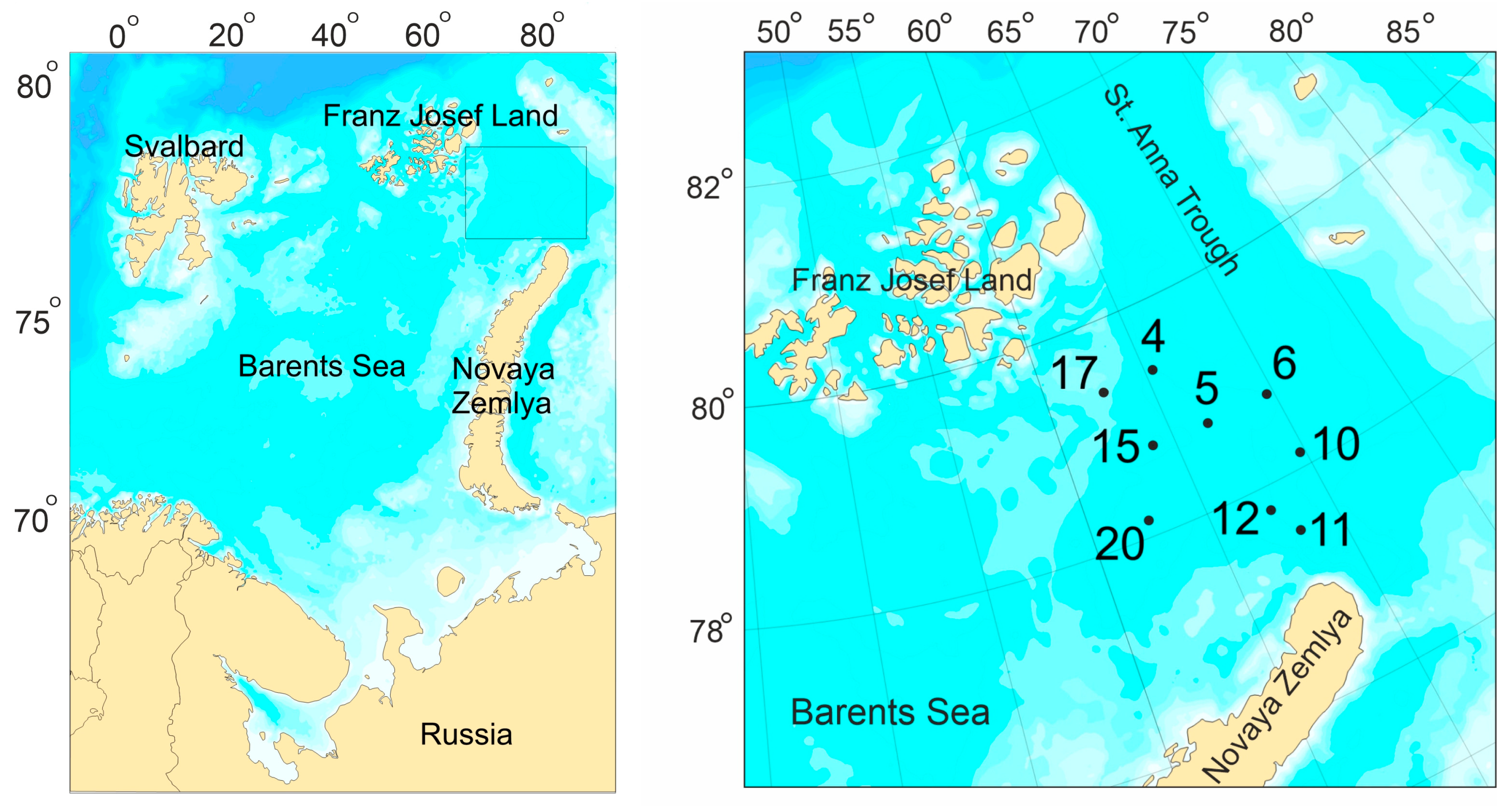
2. Materials and Methods
2.1. Sample Collection and Analysis
2.2. Diversity Indices
2.3. Statistical Analysis
3. Results
3.1. Environmental Conditions
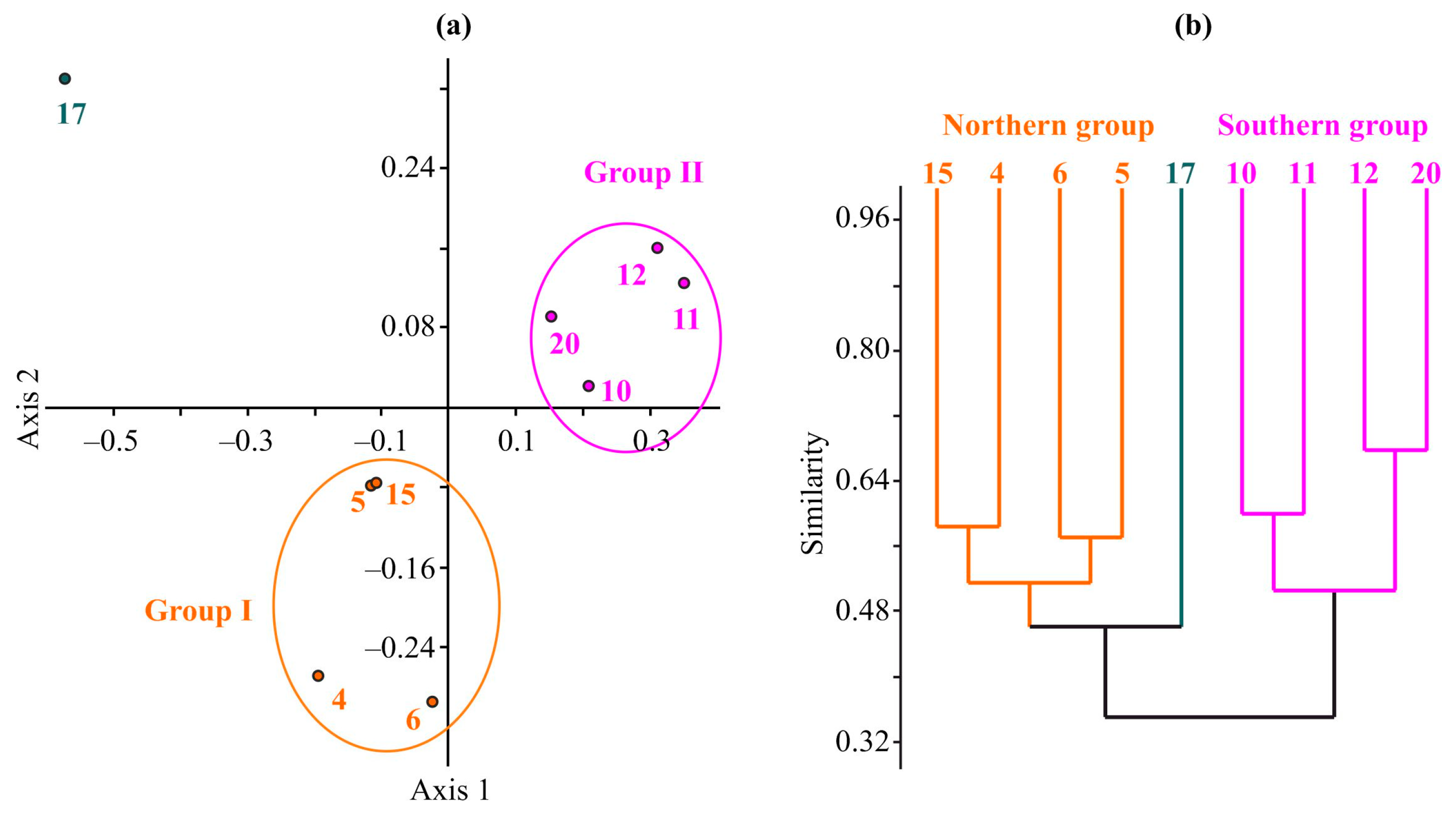
3.2. Benthic Diversity, Abundance, and Biomass
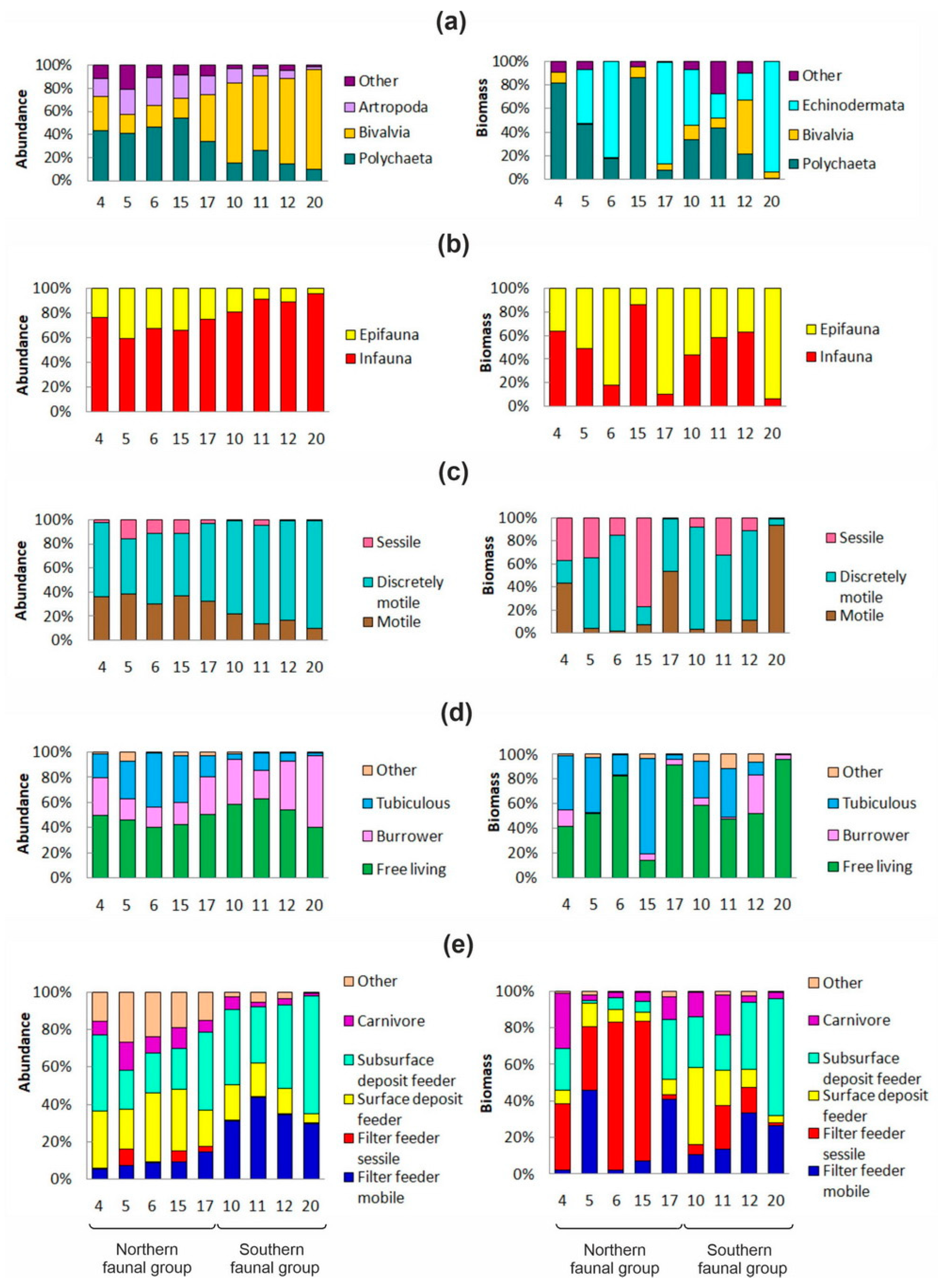
3.3. Faunal Groups
3.4. Benthic Communities within the Faunal Groups
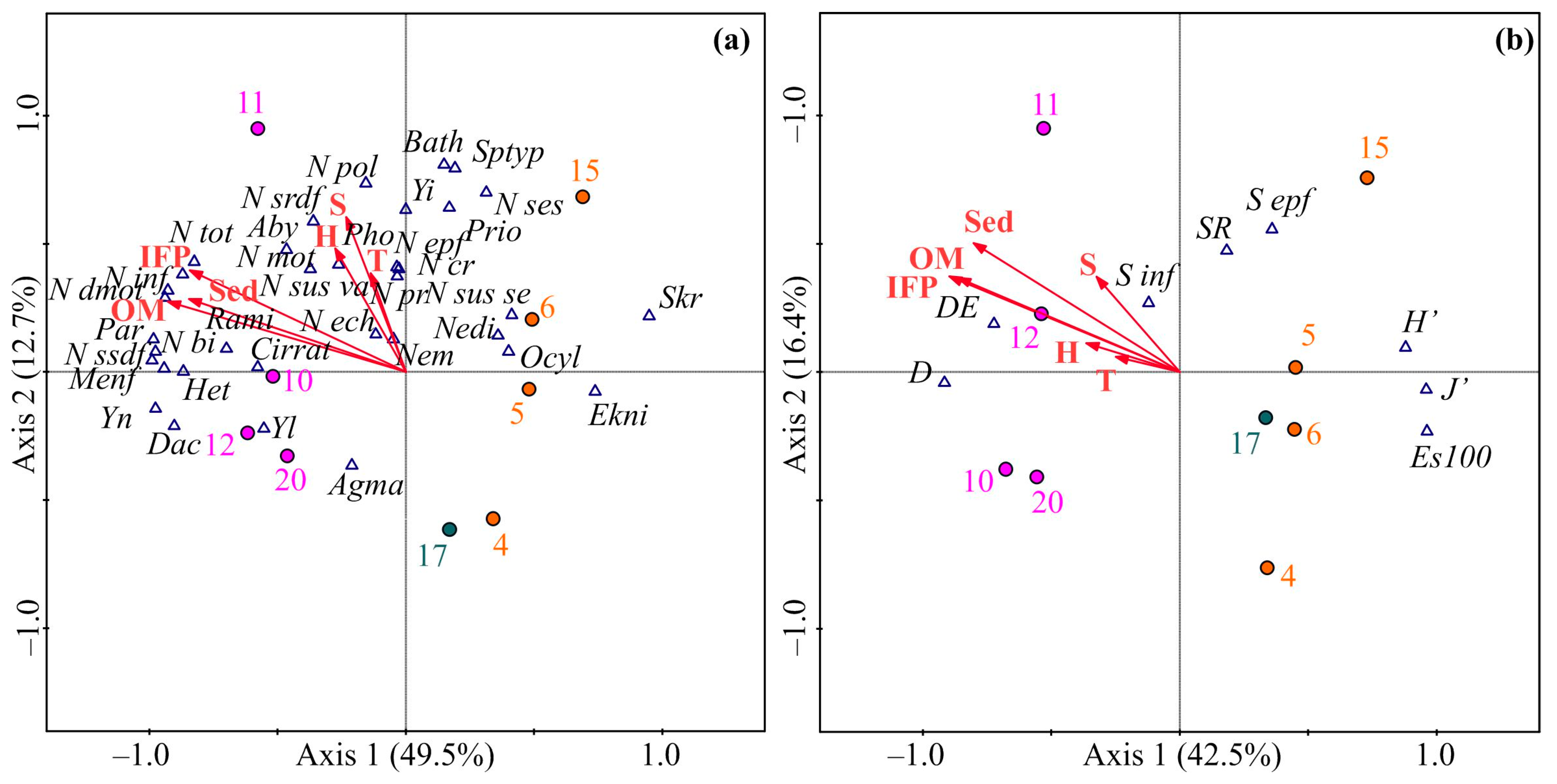
3.5. Environmental Factors driving Benthic Communities
4. Discussion
4.1. Environment in the Makarov Strait under Warming Conditions
4.2. Functioning of Macrobenthic Communities under Modern Climatic Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Station | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 4 | 5 | 6 | 10 | 11 | 12 | 15 | 17 | 20 | |
| Phylum Porifera | |||||||||
| Class Calcarea | |||||||||
| Clathrina blanca (Miklucho-Maclay, 1868) | + | + | |||||||
| Calcarea g. sp. | + | ||||||||
| Class Demospongiae | |||||||||
| Polymastia grimaldii (Topsent, 1913) | + | + | + | ||||||
| Polymastia sp. | + | + | + | ||||||
| Polymastiidae g. sp. | + | + | |||||||
| Phylum Cnidaria | |||||||||
| Class Hydrozoa | |||||||||
| Eudendrium vaginatum Allman, 1863 | + | ||||||||
| Eudendrium sp. 1 (capillare?) | + | ||||||||
| Eudendrium sp. 2 (album?) | + | ||||||||
| Halitholus yoldiaearcticae (Birula, 1897) | + | ||||||||
| Ptychogena crocea Kramp & Damas, 1925 | + | ||||||||
| Class Anthozoa | |||||||||
| Gersemia fruticosa (Sars, 1860) | + | + | + | ||||||
| Gersemia rubiformis (Ehrenberg, 1834) | + | + | |||||||
| Edwardsiidae g. sp. | + | + | + | ||||||
| Phylum Nemertea | |||||||||
| Nemertea g. sp. | + | + | + | + | + | + | + | + | + |
| Phylum Nematoda | |||||||||
| Nematoda g. sp. | + | + | + | + | + | + | + | + | |
| Phylum Annelida | |||||||||
| Class Polychaeta | |||||||||
| Abyssoninoe cf. abyssorum (McIntosh, 1885) | + | + | + | + | + | + | + | + | + |
| Aglaophamus malmgreni (Théel, 1879) | + | + | + | + | + | + | + | ||
| Ampharetidae g. sp. | + | + | + | ||||||
| Ampharete finmarchica (M. Sars, 1866) | + | + | |||||||
| Aphelochaeta sp. | + | + | |||||||
| Aricidea hartmanae (Strelzov, 1968) | + | + | + | + | |||||
| Aricidea (Strelzovia) quadrilobata Webster & Benedict, 1887 | + | + | + | + | |||||
| Aricidea nolani Webster & Benedict, 1887 | + | ||||||||
| Aricidea sp. | + | ||||||||
| Artacama proboscidea Malmgren, 1865 | + | ||||||||
| Bylgides sp. | + | ||||||||
| Chone infundibuliformis Krøyer, 1856 | + | + | |||||||
| Chone murmanica Lucash, 1910 | + | + | + | + | |||||
| Chone sp. | + | + | |||||||
| Cirratulidae g. sp. | + | + | + | + | + | + | + | + | |
| Cistenides hyperborea (Malmgren, 1865) | + | + | + | + | |||||
| Cossura longocirrata Webster & Benedict, 1887 | + | + | + | + | + | ||||
| Dipolydora caeca (Oersted, 1843) | + | ||||||||
| Enipo torelli (Malmgren, 1865) | + | ||||||||
| Eteone flava (Fabricius, 1780) | + | + | + | + | |||||
| Euchone analis (Krøyer, 1856) | + | + | |||||||
| Galathowenia oculata Zachs, 1923 | + | + | + | + | + | ||||
| Glyphanostomum pallescens (Théel, 1873) | + | + | + | + | |||||
| Harmothoe impar impar (Johnston, 1839) | + | ||||||||
| Hesionidae g. sp. | + | ||||||||
| Heteromastus filiformis (Claparède, 1864) | + | + | + | + | + | + | + | ||
| Laonice cirrata (M. Sars, 1851) | + | + | + | + | + | + | |||
| Laphania boecki Malmgren, 1865 | + | ||||||||
| Leitoscoloplos acutus (Verrill, 1873) | + | + | + | + | |||||
| Levinsenia gracilis (Tauber, 1879) | + | + | + | ||||||
| Lysippe labiata Malmgren, 1865 | + | + | |||||||
| Maldane sarsi Malmgren, 1867 | + | + | + | + | |||||
| Maldanidae g. sp. | + | ||||||||
| Melinna elisabethae McIntosh, 1922 | + | ||||||||
| Melinnopsis arctica (Annenkova, 1931) | + | ||||||||
| Myriochele heeri Malmgren, 1867 | + | + | + | + | + | + | |||
| Nephtys ciliata (Müller, 1779) | + | + | + | ||||||
| Nephtyidae g. sp. | + | + | |||||||
| Nereis zonata Malmgren, 1867 | + | + | |||||||
| Nothria hyperborea (Hansen, 1878) | + | + | + | ||||||
| Notomastus latericeus M. Sars, 1851 | + | + | + | + | |||||
| Notoproctus oculatus Arwidsson, 1906 | + | + | + | + | |||||
| Ophelina abranchiata Støp-Bowitz, 1948 | + | + | + | + | + | ||||
| Ophelina cylindricaudata (Hansen, 1878) | + | + | + | + | + | + | + | ||
| Owenidae g. sp. | + | + | + | + | |||||
| Parougia sp. | + | + | + | + | + | + | |||
| Pholoe assimilis (Muller, 1776) | + | + | + | + | + | + | + | ||
| Phyllodoce groenlandica Oersted, 1842 | + | + | + | ||||||
| Phyllodocidae g. sp. | + | ||||||||
| Polycirrus medusa Grube, 1850 | + | ||||||||
| Polynoidae g. sp. | + | + | |||||||
| Praxillella gracilis (M. Sars, 1861) | + | + | + | + | + | ||||
| Praxillella praetermissa (Malmgren, 1865) | + | + | + | ||||||
| Praxillura longissima Arwidsson, 1906 | + | + | + | + | |||||
| Prionospio cirrifera (Wirén, 1883) | + | + | + | + | + | + | + | + | + |
| Pseudoscalibregma parvum (Hansen, 1878) | + | + | |||||||
| Scalibregma inflatum Rathke, 1843 | + | + | + | + | + | + | |||
| Scolelepis aggr. korsuni Sikorski, 1994 | + | ||||||||
| Sosane wireni (Hessle, 1917) | + | + | + | ||||||
| Sphaerodoropsis philippi (Fauvel, 1911) | + | + | + | + | + | ||||
| Sphaerodoridium kolchaki Gagaev, 2015 | + | + | |||||||
| Spio limicola Verrill, 1879 | + | + | |||||||
| Spiochaetopterus typicus M. Sars, 1856 | + | + | + | + | + | + | + | + | |
| Spiophanes kroeyeri Grube, 1860 | + | + | + | + | + | + | |||
| Syllidae g. sp. | + | + | |||||||
| Syllis cornuta Rathke, 1843 | + | ||||||||
| Thelepus cincinnatus (Fabricius, 1780) | + | ||||||||
| Terebellidae g. sp. | + | + | |||||||
| Terebellides stroemi Sars, 1835 | + | + | + | + | + | + | |||
| Terebellides gracilis Malm, 1874 | + | ||||||||
| Trochochaeta carica Birula, 1879 | + | ||||||||
| Subclass Echiura | |||||||||
| Hamingia arctica Danielsen & Koren, 1881 | + | ||||||||
| Phylum Sipuncula | |||||||||
| Class Sipunculidea | |||||||||
| Golfingia (Golfingia) vulgaris vulgaris (de Blainville, 1827) | + | + | + | + | |||||
| Golfingia (Golfingia) sp. | + | ||||||||
| Nephasoma (Nephasoma) capilleforme (Murina, 1973) | + | ||||||||
| Nephasoma (Nephasoma) diaphanes diaphanes (Gerould, 1913) | + | + | + | + | + | + | + | + | |
| Nephasoma (Nephasoma) lilljeborgi (Danielssen & Koren, 1880) | + | + | + | + | |||||
| Nephasoma (Nephasoma) sp. | + | + | |||||||
| Phascolion (Phascolion) strombus strombus (Montagu, 1804) | + | + | + | ||||||
| Phylum Arthropoda | |||||||||
| Class Malacostraca | |||||||||
| Order Amphipoda | |||||||||
| Aceroides (Aceroides) latipes (G.O. Sars, 1882) | + | ||||||||
| Arrhis phyllonyx (M. Sars, 1858) | + | + | + | ||||||
| Centromedon calcaratus G. O. Sars, 1879 | + | ||||||||
| Centromedon typhlops (G. O. Sars, 1879) | + | + | |||||||
| Centromedon sp. | + | ||||||||
| Halice abyssi Stappers, 1911 | + | ||||||||
| Haploops tenuis Kanneworff, 1966 | + | + | + | ||||||
| Haploops sp. | + | ||||||||
| Harpinia mucronata G.O. Sars, 1879 | + | + | + | + | + | ||||
| Harpinia pectinata G.O. Sars, 1891 | + | + | |||||||
| Harpiniopsis similis Stephensen, 1925 | + | ||||||||
| Hippomedon sp. | + | ||||||||
| Lysianassidae g. sp. | + | ||||||||
| Paraphoxus oculatus (G.O. Sars, 1879) | + | ||||||||
| Pardaliscella boecki (Malm, 1870) | + | ||||||||
| Paroediceros lynceus (M. Sars, 1858) | + | ||||||||
| Pleusymtes pulchella (G.O. Sars, 1876) | + | ||||||||
| Pontoporeia femorata Krøyer, 1842 | + | ||||||||
| Order Cumacea | |||||||||
| Ektonodiastylis nimia (Hansen, 1920) | + | + | + | + | + | + | + | + | + |
| Eudorella emarginata (Krøyer, 1846) | + | + | + | + | |||||
| Eudorella gracilis Sars, 1871 | + | + | + | + | |||||
| Eudorella sp. | + | ||||||||
| Leptostylis gorbunovi Zimmer, 1946 | + | ||||||||
| Leucon (Leucon) acutirostris G.O. Sars, 1864 | + | + | + | + | + | ||||
| Leucon (Leucon) nasica (Krøyer, 1841) | + | ||||||||
| Leucon (Alytoleucon) pallidus G.O. Sars, 1864 | + | + | + | ||||||
| Order Decapoda | |||||||||
| Chionoecetes opilio juv. (O. Fabricius, 1788) | + | ||||||||
| Order Isopoda | |||||||||
| Caecognathia elongata (Krøyer, 1847) | + | + | |||||||
| Calathura brachiata (Stimpson, 1854) | + | + | |||||||
| Desmosomatidae g. sp. | + | + | |||||||
| Ilyarachna bicornis Hansen, 1916 | + | ||||||||
| Ilyarachninae g. sp. | + | + | |||||||
| Order Tanaidacea | |||||||||
| Akanthophoreus sp. | + | ||||||||
| Chauliopleona armata (Hansen, 1913) | + | + | |||||||
| Pseudotanais affinis Hansen, 1887 | + | ||||||||
| Pseudotanais lilljeborgii Sars, 1882 | + | + | |||||||
| Pseudosphyrapus anomalus (Sars, 1869) | + | + | + | + | + | + | |||
| Pseudotanais sp. | + | ||||||||
| Typhlotanais sp. | + | ||||||||
| Class Ostracoda | |||||||||
| Heterocyprideis sorbyana (Jones, 1857) | + | + | + | ||||||
| Philomedes globosus (Lilljeborg, 1853) | + | + | + | ||||||
| Rabilimis mirabilis (Brady,1968) Hazel, 1967 | + | + | + | + | + | + | + | + | + |
| Class Pycnogonida | |||||||||
| Nymphon hirtipes Bell, 1855 | + | ||||||||
| Phylum Mollusca | |||||||||
| Class Bivalvia | |||||||||
| Astarte crenata (Gray, 1842) | + | + | + | + | + | + | + | ||
| Axinopsida orbuculata (G.O. Sars, 1878) | + | + | |||||||
| Bathyarca glacialis (Gray, 1842) | + | + | + | + | + | + | + | + | |
| Ciliatocardium ciliatum (Fabricius, 1780) | + | + | + | + | |||||
| Cuspidaria glacialis (G.O. Sars, 1878) | + | + | + | + | + | + | |||
| Dacrydium vitreum (Møller, 1842) | + | + | + | + | + | + | + | + | |
| Ennucula tenuis (Montagu, 1808) | + | + | + | ||||||
| Macoma calcarea (Gmelin,1791) | + | + | + | + | |||||
| Mendicula ferruginosa (Forbes, 1844) | + | + | + | + | + | + | + | + | |
| Nuculana pernula (O.F. Müller, 1779) | + | + | + | + | |||||
| Parathyasira equalis (Verrill & Bush, 1898) | + | + | + | + | + | + | + | + | |
| Yoldiella annenkovae (Gorbunov, 1946) | + | + | + | + | + | ||||
| Yoldiella frigida (Torell, 1859) | + | + | + | + | |||||
| Yoldiella intermedia (M. Sars, 1865) | + | + | + | + | + | + | |||
| Yoldiella lenticula (Møller, 1842) | + | + | + | + | + | + | |||
| Yoldiella nana (M. Sars, 1865) | + | + | + | + | + | + | + | + | + |
| Yoldiella solidula Warén, 1989 | + | ||||||||
| Class Caudofoveata | |||||||||
| Caudofoveata g. sp. | + | + | + | ||||||
| Class Gastropoda | |||||||||
| Cylichna alba (T. Brown, 1827) | + | + | + | ||||||
| Diaphana hiemalis (Couthouy, 1839) | + | + | |||||||
| Frigidoalvania janmayeni (Friele, 1878) | + | ||||||||
| Lepeta coeca (O.F. Müller, 1776) | |||||||||
| Margarites groenlandicus (Gmelin, 1791) | |||||||||
| Onoba aculeus (A. Gould, 1841) | + | ||||||||
| Onoba leptalea (Verrill, 1884) | + | ||||||||
| Propebela harpularia (Couthuoy, 1838) | + | + | + | + | + | + | + | + | |
| Punctulum wyvillethomsoni (Friele, 1877) | + | + | + | + | + | ||||
| Retusa obtusa (Montagu, 1803) | + | + | |||||||
| Testudinalia testudinalis (O.F. Müller, 1776) | + | ||||||||
| Class Scaphopoda | |||||||||
| Siphonodentalium lobatum (G.B. Sowerby II, 1860) | + | ||||||||
| Phylum Bryozoa | |||||||||
| Class Gymnolaemata | |||||||||
| Alcyonidium gelatinosum (Linneus, 1761) | + | ||||||||
| Alcyonidium radicellatum Kluge, 1946 | + | + | |||||||
| Bugulopsis peachii (Busk, 1851) | + | ||||||||
| Copidozoum smitti (Kluge, 1946) | + | + | |||||||
| Escharoides jacksonii (Waters, 1900) | + | ||||||||
| Eucratea loricata (Linneus, 1758) | + | + | + | ||||||
| Leieschara subgracilis (d’Orbigny, 1853) | + | ||||||||
| Parasmittina jeffreysii (Norman, 1876) | + | ||||||||
| Pseudoflustra solida (Stimpson, 1854) | + | ||||||||
| Sarsiflustra abyssicola (G.O. Sars, 1872) | + | ||||||||
| Schizoporella costata Kluge, 1962 | + | ||||||||
| Scrupocellaria intermedia Norman, 1893 | + | ||||||||
| Scrupocellaria minor Kluge, 1915 | + | ||||||||
| Stomacrustula cruenta (Busk, 1854) | |||||||||
| Uschakovia gorbunovi Kluge, 1946 | + | ||||||||
| Class Stenolaemata | |||||||||
| Crisia eburneodenticulata Smitt ms in Busk, 1875 | + | + | + | ||||||
| Crisiella sp. | + | ||||||||
| Exidmonea atlantica (Forbs in Jonston, 1847) | + | ||||||||
| Phylum Echinodermata | |||||||||
| Class Asteroidea | |||||||||
| Ctenodiscus crispatus (Bruzelius, 1805) | + | + | + | + | |||||
| Pontaster tenuispinus (Düben & Koren, 1846) | + | ||||||||
| Class Echinoidea | |||||||||
| Strongylocentrotus pallidus (G.O. Sars, 1872) | + | ||||||||
| Class Holothuroidea | |||||||||
| Chiridota laevis (Fabricius, 1780) | + | ||||||||
| Molpadia arctica (Marenzeller von, 1877) | + | ||||||||
| Myriotrochus eurycyclus Heding, 1935 | + | ||||||||
| Myriotrochus rinkii Steenstrup, 1851 | + | ||||||||
| Class Ophiuroidea | |||||||||
| Gorgonocephalus arcticus Leach, 1819 | + | + | |||||||
| Ophiacantha bidentata (Bruzelius, 1805) | + | + | + | ||||||
| Ophiopus arcticus Ljungman, 1867 | + | ||||||||
| Ophiocten sericeum (Forbes, 1852) | + | ||||||||
| Ophiopleura borealis Danielssen & Koren, 1877 | + | + | + | + | + | ||||
| Ophioscolex glacialis Müller & Troschel, 1842 | + | + | + | ||||||
| Ophiuroidea g. sp. | + | + | |||||||
| Phylum Chordata | |||||||||
| Class Ascidiacea | |||||||||
| Molgula sp. | + | ||||||||
| Polycarpa comata (Alder, 1863) | + | ||||||||
References
- Kędra, M.; Grebmeier, J.M. Ecology of Arctic Shelf and Deep Ocean Benthos. In Arctic Ecology; Thomas, D.N., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021; pp. 325–355. [Google Scholar]
- Wassmann, P.; Reigstad, M.; Haug, T.; Rudels, B.; Carroll, M.L.; Hop, H.; Gabrielsen, G.W.; Falk-Petersen, S.; Denisenko, S.G.; Arashkevich, E.; et al. Food webs and carbon flux in the Barents Sea. Prog. Oceanogr. 2006, 71, 232–287. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar]
- Jakobsen, T.; Ozhigin, V.K. (Eds.) The Barents Sea: Ecosystem, Resources, Management: Half a Century of Russian-Norwegian Co-Operation; Tapir Academic Press: Trondheim, Norway, 2011. [Google Scholar]
- Cochrane, S.K.; Denisenko, S.G.; Renaud, P.E.; Emblow, C.S.; Ambrose, W.G., Jr.; Ellingsen, I.H.; Skarðhamar, J. Benthic macrofauna and productivity regimes in the Barents Sea—Ecological implications in a changing Arctic. J. Sea Res. 2009, 61, 222–233. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar]
- Reigstad, M.; Carroll, J.; Slagstad, D.; Ellingsen, I.; Wassmann, P. Intra-regional comparison of productivity, carbon flux and ecosystem composition within the northern Barents Sea. Prog. Oceanogr. 2011, 90, 33–46. [Google Scholar]
- Cochrane, S.K.J.; Pearson, T.H.; Greenacre, M.; Costelloe, J.; Ellingsen, I.H.; Dahle, S.; Gulliksen, B. Benthic fauna and functional traits along a Polar Front transect in the Barents Sea—Advancing tools for ecosystem-scale assessments. J. Mar. Syst. 2012, 94, 204–217. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Shellfish as biosensors in online monitoring of aquatic ecosystems: A review of Russian studies. Fishes 2023, 8, 102. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibionts of an introduced king crab in the Barents Sea: A second five-year study. Diversity 2023, 15, 29. [Google Scholar]
- Evseeva, O.Y.; Dvoretsky, A.G. Shallow-water bryozoan communities in a glacier fjord of West Svalbard, Norway: Species composition and effects of environmental factors. Biology 2023, 12, 185. [Google Scholar]
- Eriksen, E.; Benzik, A.N.; Dolgov, A.V.; Skjoldal, H.R.; Vihtakari, M.; Johannesen, E.; Prokhorova, T.A.; Keulder-Stenevik, F.; Prokopchuk, I.; Strand, E. Diet and trophic structure of fishes in the Barents Sea: The Norwegian-Russian program “year of stomachs” 2015—establishing a baseline. Prog. Oceanogr. 2020, 183, 102262. [Google Scholar]
- Pavlova, L.V.; Dvoretsky, A.G. Prey selectivity in juvenile red king crabs from the coastal Barents Sea. Diversity 2022, 14, 568. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 1280–1290. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Cucumaria in Russian waters of the Barents Sea: Biological aspects and aquaculture potential. Front. Mar. Sci. 2021, 8, 613453. [Google Scholar]
- Morata, N.; Michaud, E.; Poullaouec, M.A.; Devesa, J.; Le Goff, M.; Corvaisier, R.; Renaud, P.E. Climate change and diminishing seasonality in Arctic benthic processes. Phil. Trans. R. Soc. A 2020, 378, 20190369. [Google Scholar]
- Onarheim, I.H.; Eldevik, T.; Smedsrud, L.H.; Stroeve, J.C. Seasonal and regional manifestation of Arctic sea ice loss. J. Clim. 2018, 31, 4917–4932. [Google Scholar]
- Dvoretsky, V.G.; Vodopianova, V.V.; Bulavina, A.S. Effects of climate change on chlorophyll a in the Barents Sea: A long-term assessment. Biology 2023, 12, 119. [Google Scholar]
- Renaud, P.E.; Morata, N.; Carroll, M.L.; Denisenko, S.G.; Reigstad, M. Pelagic–benthic coupling in the western Barents Sea: Processes and time scales. Deep Sea Res. Part II 2008, 55, 2372–2380. [Google Scholar] [CrossRef]
- Barton, B.I.; Lenn, Y.D.; Lique, C. Observed Atlantification of the Barents Sea causes the polar front to limit the expansion of winter sea ice. J. Phys. Oceanogr. 2018, 48, 1849–1866. [Google Scholar] [CrossRef]
- Carroll, M.L.; Denisenko, S.G.; Renaud, P.E.; Ambrose, W.G., Jr. Benthic infauna of the seasonally ice-covered western Barents Sea: Patterns and relationships to environmental forcing. Deep Sea Res. Part II 2008, 55, 2340–2351. [Google Scholar] [CrossRef]
- Denisenko, S.G. Biodiversity and Bioresources of Macrozoobenthos of the Barents Sea: Structure and Long-Term Changes; Nauka: St. Petersburg, Russia, 2013. (In Russian) [Google Scholar]
- Lyubina, O.S.; Strelkova, N.A.; Lubin, P.A.; Frolova, E.A.; Dikaeva, D.R.; Zimina, O.L.; Akhmetchina, O.Y.; Manushin, I.E.; Nekhaev, I.O.; Frolov, A.A.; et al. Modern quantitative distribution of zoobenthos along on the transect “Kola Section”. Trans. Kola Sci. Cent. RAS 2016, 3, 64–91. (In Russian) [Google Scholar]
- Evseeva, O.Y.; Ishkulova, T.G.; Dvoretsky, A.G. Environmental drivers of an intertidal bryozoan community in the Barents Sea: A case study. Animals 2022, 12, 552. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, L.V.; Zuyev, Y.A.; Dvoretsky, A.G. Shallow-water benthic communities on soft bottoms of a sub-Arctic fjord (Southern Barents Sea, Russia) along a gradient of ecological factors. Diversity 2023, 15, 84. [Google Scholar] [CrossRef]
- Brotskaya, V.A.; Zenkevich, L.A. Quantitative accounting of the bottom fauna of the Barents Sea. Tr. VNIRO 1939, 4, 5–126. (In Russian) [Google Scholar]
- Kiyko, O.A.; Pogrebov, V.B. Long-term benthic population change (1920–1930s-present) in the Barents and Kara seas. Mar. Poll. Bull. 1997, 35, 322–332. [Google Scholar] [CrossRef]
- Panteleeva, N.N.; Rodimov, A.A. Analysis of bottom fauna communities in the glacial waters of Novaya Zemlya and in the St. Anna Trough. In Scientific Report of MMBI Expedition “Yasnogorsk-95”. Marine Biological and Geological Studies off Novaya Zemlya Glaciers and in St. Anna Trough; Korsun, S.A., Ed.; KSC RAS: Apatity, Russia, 1996; pp. 35–40. (In Russian) [Google Scholar]
- Gurevich, V.I. Modern Sedimentogenesis and Geoecology of the Western Arctic Shelf of Eurasia; Scientific World: Moscow, Russia, 2002. (In Russian) [Google Scholar]
- Loeng, H.; Ozhigin, V.; Adlandsvik, B. Water fluxes through the Barents Sea. ICES J. Mar. Sci. 1997, 54, 310–317. [Google Scholar] [CrossRef]
- Schauer, U.; Loeng, H.; Rudels, B.; Ozhigin, V.K.; Dieck, W. Atlantic Water flow through the Barents and Kara Seas. Deep Sea Res. Part I 2002, 49, 2281–2298. [Google Scholar] [CrossRef]
- Ozhigin, V.K.; Ivshin, V.A.; Trofimov, A.G.; Karsakov, A.L.; Anciferov, M.Y. The Barents Sea Water: Structure, Circulation, Variability; PINRO: Murmansk, Russia, 2016. (In Russian) [Google Scholar]
- Tarasov, G.A.; Ivanov, G.; Kukina, N.A.; Matishov, G.G. The granulometrical structure of surface sediments in the St. Anna Trough area. Ber. Polarforsch. 1999, 342, 124–133. [Google Scholar]
- Korsun, S.; Hald, M.; Panteleeva, N.; Tarasov, G. Biomass of Foraminifera in the St. Anna Trough, Russian arctic continental margin. Sarsia 1998, 83, 419–431. [Google Scholar] [CrossRef]
- Glyaznetsova, Y.S.; Nemirovskaya, I.A. Features of bitumoid distribution in bottom sediments of the Barents Sea. Oceanology 2020, 60, 831–839. [Google Scholar] [CrossRef]
- Shevchenko, V.P.; Ivanov, G.; Zernova, V.V. Vertical particle fluxes in the St. Anna Trough and in the eastern Barents Sea in august–september 1994. Ber. Polarforsch. 1999, 342, 46–54. [Google Scholar]
- Dvoretsky, V.G.; Venger, M.P.; Vashchenko, A.V.; Maksimovskaya, T.M.; Ishkulova, T.G.; Vodopianova, V.V. Pelagic bacteria and viruses in a high Arctic region: Environmental control in the autumn period. Biology 2022, 11, 845. [Google Scholar] [CrossRef]
- Matishov, G.G.; Berdnikov, S.V.; Zhichkin, A.P. Atlas of Climatic Changes in Large Marine Ecosystems of the Northen Hemisphere (1873–2013); Region 1. The Eastern Arctic seas. Region. Region 2. The Black Sea, the Sea of Azov, and the Caspian Sea; SSC RAS: Rostov, Russia, 2014. (In Russian) [Google Scholar]
- Matishov, G.G.; Dzhenyuk, S.L.; Denisov, V.V.; Zhichkin, A.P.; Moiseev, D.V. Taking into account of the Barents Sea climate secular dynamics in course of marine activity planning. Trans. Kola Sci. Cent. 2013, 1, 56–71. (In Russian) [Google Scholar]
- Parkinson, C.L. Spatially mapped reductions in the length of the Arctic sea ice season. Geophys. Res. Lett. 2014, 41, 4316–4322. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Copepod communities off Franz Josef Land (northern Barents Sea) in late summer of 2006 and 2007. Polar Biol. 2011, 34, 1231–1238. [Google Scholar] [CrossRef]
- Dvoretsky, V.G. Distribution of Calanus species off Franz-Josef Land (Arctic Barents Sea). Polar Sci. 2011, 5, 361–373. [Google Scholar] [CrossRef]
- Istoshin, Y.V. Marine Hydrometry; Hydrometeorological Publishing House: Leningrad, USSR, 1967. (In Russian) [Google Scholar]
- Noorany, I. Classification of marine sediments. J. Geotech. Eng. 1989, 115, 23–37. [Google Scholar] [CrossRef]
- Wollenburg, J.E.; Kuhnt, W. The response of benthic foraminifers to carbon flux and primary production in the Arctic Ocean. Mar. Micropaleontol. 2000, 40, 189–231. [Google Scholar] [CrossRef]
- Hulbert, S.H. The non-concept of species diversity. A critique and alternative parameters. Ecology 1971, 52, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Golikov, A.N.; Scarlato, O.A.; Averintsev, V.G.; Menshutkina, T.V.; Novikov, O.K.; Sheremetevsky, A.M. Ecosystems of the Novosibirsk shallow water and some patterns of their distribution and functioning. In Ecosystems of the Novosibirsk Shallow Water and Fauna of the Laptev Sea and Adjacent Waters of the Arctic Ocean: Collection of Scientific Works. Research of the Fauna of Seas; Golikov, A.N., Ed.; Nauka: Saint Petersburg, Russia, 1990; Volume 37, pp. 4–79. (In Russian) [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Macdonald, T.A.; Burd, B.J.; Macdonald, V.I.; van Roodselaar, A. Taxonomic and feeding guild classification for the marine benthic macroinvertebrates of the Strait of Georgia, British Columbia. Can. Tech. Rep. Fish. Aquat. Sci. 2010, 2874, 1–63. [Google Scholar]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Ann. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef]
- Gagaev, S.Y. Polychaetes (Annelida: Polychaeta) of the Kara Sea. Proc. Zool. Inst. RAS 2021, 325, 183–196. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and Canodraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Aksenov, P.V.; Ivanov, V.V. “Atlantification” as a Possible Cause for Reducing of the Sea-Ice Cover in the Nansen Basin in winter. Arct. Antarct. Res. 2018, 64, 42–54. (In Russian) [Google Scholar] [CrossRef]
- Alekseev, G.; Glok, N. Influence of the equatorial North Atlantic on the sea ice shrinking in the Arctic. Preprint. 2020. Available online: http://old.aari.ru/resources/meteoarct/docs/02_Alekseev_Genrikh_SSTAtlSIEArct.pdf (accessed on 20 May 2023).
- Pearson, T.H.; Rosenberg, R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev. 1978, 16, 229–311. [Google Scholar]
- Vetrov, A.A.; Romankevich, E.A. Primary production and fluxes of organic carbon to the seabed in the Eurasian arctic seas, 2003–2012. Dokl. Earth Sci. 2014, 454, 44–46. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L. Continued increases in Arctic Ocean primary production. Prog. Oceanogr. 2015, 136, 60–70. [Google Scholar] [CrossRef]
- Lewis, K.M.; van Dijken, G.L.; Arrigo, K.R. Changes in phytoplankton concentration now drive increased Arctic Ocean primary. Science 2020, 369, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Kodryan, K.V.; Torgunova, N.I.; Agatova, A.I. Variability of dissolved and particulate organic matter of the Barents Sea during the period of variability of its ice cover. Tr. VNIRO 2017, 169, 91–103. (In Russian) [Google Scholar]
- Makarevich, P.; Druzhkova, E.; Larionov, V. Primary producers of the Barents Sea. In Diversity of Ecosystems; Mahamane, A., Ed.; In Tech: Rijeka, Croatia, 2012; pp. 367–392. [Google Scholar]
- Zatsepin, A.G.; Poyarkov, S.G.; Kremenetskiy, V.V.; Nedospasov, A.A.; Shchuka, S.A.; Baranov, V.I.; Kondrashov, A.A.; Korzh, A.O. Hydrophysical features of deep water troughs in the western Kara Sea. Oceanology 2015, 55, 472–484. [Google Scholar] [CrossRef]
- Pokrovsky, O.S.; Viers, J.; Dupre, B.; Chabaux, F.; Gaillardet, J.; Audry, S.; Prokushkin, A.S.; Shirokova, L.S.; Kirpotin, S.N.; Lapitsky, S.A.; et al. Biogeochemistry of carbon, major and trace elements in watersheds of Northern Eurasia drained to the Arctic Ocean: The change of fluxes, sources and mechanisms under the climate warming prospective. Comptes Rendus Geosci. 2012, 344, 663–677. [Google Scholar] [CrossRef]
- Matishov, G.G.; Matishov, D.G.; Moiseev, D.V. Inflow of Atlantic-origin waters to the Barents Sea along glacial troughs. Oceanologia 2009, 51, 321–340. [Google Scholar] [CrossRef]
- Makhotin, M.S.; Ivanov, V.V. Distribution of Atlantic water masses in the Barents Sea according to observations and numerical modeling. Hydromet. Res. Cent. Russ. Fed. 2016, 361, 169–191. (In Russian) [Google Scholar]
- Murdmaa, I.O. Oceanic facies. In Ocean Geology. Sedimentation and Magmatism of the Ocean; Monin, A.S., Lisitsyn, A.P., Eds.; Nauka: Moscow, Russia, 1979; pp. 269–306. (In Russian) [Google Scholar]
- Denisenko, N.V.; Denisenko, S.G. Zoobenthos of the Barents Sea. In The Barents Sea System; Lisitzin, A.P., Ed.; GEOS: Moscow, Russia, 2021; pp. 352–369. (In Russian) [Google Scholar]
- Lyubina, O.S.; Dikaeva, D.R.; Frolova, E.A.; Frolov, A.A.; Zimina, O.L.; Akhmetchina, O.Y.; Garbul, E.A. Dynamics of benthic communities in the central depression of the Barents Sea. Biol. Bull. 2010, 37, 523–531. [Google Scholar] [CrossRef]
- Zimina, O.L.; Frolova, E.A.; Dikaeva, D.R.; Akhmetchina, O.Y.; Garbul, E.A.; Frolov, A.A.; Nekhaev, I.O. Fauna and distribution of abundance and biomass of zoobenthos in the northern Barents Sea in April and May 2016. Trans. Kola Sci. Cent. RAS 2017, 2, 66–81. (In Russian) [Google Scholar]
- Zakharov, D.V.; Manushin, I.E.; Strelkova, N.A.; Pavlov, V.A.; Nosova, T.B. Diet of the snow crab in the Barents Sea and macrozoobenthic communities in the area of its distribution. Tr. VNIRO 2018, 172, 70–90. (In Russian) [Google Scholar] [CrossRef]
- Renaud, P.E.; Ambrose, W.G.; Węsławski, J.M. Benthic communities in the polar night. In Polar Night Marine Ecology. Advances in Polar Ecology; Berge, J., Johnsen, G., Cohen, J., Eds.; Springer: Cham, Switzerland, 2020; Volume 4, pp. 161–179. [Google Scholar]
- Yao, K.S.; He, Y.J.; Van den Brink, P.J.; Dai, Y.N.; Yang, Y.; Cui, Y.D. Multivariate approaches to assess the drivers of benthic macroinvertebrate communities and biotic indices in a highly urbanized fluvial-estuarine system. Ecol. Indic. 2022, 139, 108956. [Google Scholar] [CrossRef]
- Piepenburg, D.; Ambrose, W.G.; Brandt, A.; Renaud, P.E.; Ahrens, M.J.; Jensen, P. Benthic community patterns reflect water column processes in the Northeast Water Polynya (Greenland). J. Mar. Syst. 1997, 10, 467–482. [Google Scholar] [CrossRef]
- Ambrose, W.G., Jr.; Renaud, P.E.; Cochrane, S.K.; Denisenko, S.G.; Skarðhamar, J. Polychaete diversity patterns on two Arctic shelves: Impacts of ice and primary production? Zoosymposia 2009, 2, 457–485. [Google Scholar] [CrossRef]
- Novikov, M.A. Atlas of Bottom Sediments Pollution in the Barents Sea; PINRO: Murmansk, Russia, 2021. (In Russian) [Google Scholar]
- Galkin, S.V.; Vedenin, A.A.; Minin, K.V.; Rogacheva, A.V.; Molodtsova, T.N.; Rajskiy, A.K.; Kucheruk, N.V. Macrobenthos of the Southern Part of St. Anna Trough and the Adjacent Kara Sea Shelf. Oceanology 2015, 55, 614–622. [Google Scholar] [CrossRef]
- Dauwe, B.; Herman, P.M.J.; Heip, C.H.R. Community structure and bioturbation potential of macrofauna at four North Sea stations with contrasting food supply. Mar. Ecol. Prog. Ser. 1998, 173, 67–83. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Gallardo, V.A.; Mayor, S.; Neira, C.; Vásquez, C.; Sellanes, J.; Rivas, M.; Soto, A.; Carrasco, F.; Baltazar, M. Effects of dissolved oxygen and fresh organic matter on the bioturbation potential of macrofauna in sublittoral sediments off Central Chile during the 1997/1998 El Niño. Mar. Ecol. Prog. Ser. 2000, 202, 81–99. [Google Scholar] [CrossRef]
- Filatova, Z.A.; Zenkevich, L.A. Quantitative distribution of the bottom fauna of the Kara Sea. Tr. Vsesoyuznogo Gidrobiol. Obs. 1957, 8, 3–62. (In Russian) [Google Scholar]
- Kudryavtseva, O.Y. Review of Ichthyofauna in Franz-Josef Land region. Trans. Kola Sci. Cent. 2014, 4, 222–251. (In Russian) [Google Scholar]
- Astafyev, D.A.; Tolstikov, A.V.; Naumova, L.A.; Kabalin, M.Y. Promising XXI-century trends of oil and gas prospecting offshore Russia. Vesti Gazov. Nauk. 2018, 4, 4–18. (In Russian) [Google Scholar]




| No. | H | T, °C | S | Sediment | IFP | OM Influx | |
|---|---|---|---|---|---|---|---|
| Characteristics | Score | ||||||
| 4 | 427 | −0.25 | 34.79 | Liquid brown silt, foraminiferal sand, soft brown clay | 1 | 3.0 ± 0.3 | 1 |
| 5 | 343 | −0.2 | 34.9 | Brown sandy silt, foraminiferal sand, soft brown clay | 2 | 4.4 ± 0.5 | 1 |
| 6 | 507 | −0.2 | 34.9 | Brown silt, foraminiferal sand, soft brown clay | 1 | 4.6 ± 0.6 | 1 |
| 10 | 445 | −0.1 | 34.9 | Brown sandy silt, gray clay | 3 | 6.8 ± 1.1 | 2 |
| 11 | 502 | −0.3 | 34.9 | Thin layer of brown silt, gray clay, stones (occasional) | 4 | 7.8 ± 1.2 | 3 |
| 12 | 430 | −0.2 | 34.9 | Thin layer of brown silt, gray clay, stones (occasional) | 4 | 7.6 ± 1.2 | 3 |
| 15 | 361 | −0.3 | 34.9 | Brown sandy silt, foraminiferal sand, soft brown clay | 2 | 4.2 ± 0.4 | 1 |
| 17 | 186 | −1.2 | 34.7 | Brown sandy silt, foraminiferal sand, brown clay | 2 | 3.2 ± 0.6 | 1 |
| 20 | 392 | −0.3 | 34.9 | Brown sandy silt, gray clay, low content of Foraminifera | 3 | 7.0 ± 0.9 | 2 |
| No. | SR/α | ES100 | H’ | J’ | D | DE | N | B | Main Contributors (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | B | R | |||||||||
| 4 | 26/48 | 6.2 | 4.6 | 0.84 | 0.06 | −0.23 | 770 | 5 | Yoldiella nana (15%) | Aglaophamus malmgreni (25%) S. typicus (35%) | A. malmgreni (23%) |
| 5 | 49/85 | 6.2 | 5.4 | 0.84 | 0.04 | −0.46 | 1370 | 41 | Prionospio cirrifera (8%) Y. nana (8%) | S. typicus (32%) | S. typicus (44%) |
| 6 | 40/67 | 5.4 | 5.0 | 0.82 | 0.05 | −0.66 | 1230 | 82 | P. cirrifera (11%) Spiochaetopterus typicus (11%) | Ophiopleura borealis (81%) | S. typicus (48%) |
| 10 | 43/68 | 1.5 | 3.6 | 0.58 | 0.17 | −0.10 | 4640 | 39 | Y. nana (32%) Mendicula ferruginosa (23%) | O. borealis (47%) | Laonice cirrata (15%), Y.nana (15%) |
| 11 | 46/76 | 1.4 | 3.8 | 0.61 | 0.15 | 0.01 | 5580 | 72 | M. ferruginosa (31%) Y. nana (18%) | S. typicus (20%) | S. typicus (25%) |
| 12 | 41/69 | 1.6 | 3.5 | 0.58 | 0.19 | 0.06 | 4190 | 15 | Y. nana (33%) M. ferruginosa (25%) | O. borealis (23%) | Y. nana (23%) |
| 15 | 37/61 | 5.4 | 4.9 | 0.83 | 0.05 | −0.51 | 1120 | 18 | P. cirrifera (12%) Ektonodiastylis nimia (10%) | S. typicus (73%) | S. typicus (49%) |
| 17 | 37/68 | 5.4 | 4.5 | 0.74 | 0.09 | −0.38 | 1250 | 48 | E. nimia (13%) | Ctenodiscus crispatus 49%) O. borealis (30%) | C. crispatus (16%) A. malmgreni (15%) |
| 20 | 19/33 | 1.8 | 2.7 | 0.54 | 0.25 | −0.43 | 1870 | 64 | Y. nana (42%) M. ferruginosa (22%) | Strongylocentrotus pallidus (93%) | Y. nana (22%) |
| Taxa | FO | Stations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 5 | 6 | 10 | 11 | 12 | 15 | 17 | 20 | ||
| Yoldiella nana (Bi) | 96 | 113/0.12 | 103/0.04 | 63/0.05 | 1487/1.71 | 997/0.72 | 1393/2.01 | 27/0.02 | 313/0.66 | 780/1.48 |
| Prionospio cirrifera (Pol) | 96 | 77/0.09 | 103/0.09 | 133/0.20 | 50/0.06 | 187/0.36 | 153/0.35 | 130/0.23 | 43/0.06 | 23/0.04 |
| Abyssoninoe cf. abyssorum (Pol) | 93 | 57/0.49 | 23/0.10 | 13/0.04 | 67/0.41 | 177/1.09 | 67/0.45 | 80/0.35 | 37/0.39 | 30/0.18 |
| Ektonodiastylis nimia (Cum) | 89 | 67/0.02 | 167/0.04 | 127/0.04 | 53/0.01 | 23/0.00 | 47/0.01 | 107/0.04 | 157/0.03 | 7/0.00 |
| Mendicula ferruginosa (Bi) | 85 | 10/0.00 | 30/0.01 | 43/0.0.01 | 1057/0.69 | 1720/1.24 | 1043/0.67 | 0 | 93/0.06 | 403/0.34 |
| Cirratulidae g. sp. (Pol) | 81 | 47/0.07 | 0 | 13/0.05 | 60/0.23 | 100/0.47 | 30/0.24 | 10/0.04 | 50/0.05 | 17/0.10 |
| Nephasoma (Nephasoma) diaphanes diaphanes (Sip) | 74 | 27/0.03 | 67/0.91 | 23/0.07 | 20/0.02 | 23/0.06 | 20/0.02 | 13/0.04 | 17/0.05 | 0 |
| Spiochaetopterus typicus (Pol) | 70 | 7/1.83 | 93/13.08 | 133/12.46 | 37/1.03 | 230/14.67 | 17/0.65 | 60/12.89 | 3/0.21 | 0 |
| Rabilimis mirabilis (Ost) | 70 | 10/0.01 | 13/0.01 | 7/0.00 | 20/0.01 | 30/0.02 | 183/0.10 | 17/0.01 | 7/0.00 | 30/0.02 |
| Aglaophamus malmgreni (Pol) | 67 | 10/0.97 | 0 | 3/0.01 | 33/0.05 | 0 | 33/0.13 | 23/0.49 | 13/0.96 | 13/0.07 |
| Parathyasira equalis (Bi) | 67 | 13/0.03 | 7/0.01 | 10/0.05 | 297/1.08 | 663/2.91 | 347/1.33 | 0 | 13/0.06 | 127/0.77 |
| Nemertini g. sp. (Nem) | 67 | 10/0.22 | 13/0.11 | 3/0.02 | 17/0.05 | 10/0.05 | 17/0.01 | 17/0.08 | 10/0.01 | 3/0.00 |
| Dacrydium vitreum (Bi) | 63 | 3/0.02 | 7/0.01 | 3/0.02 | 53/0.07 | 17/0.01 | 27/ | 0 | 10/0.00 | 20/0.02 |
| Bathyarca glacialis (Bi) | 56 | 3/0.05 | 10/0.06 | 10/0.08 | 17/0.21 | 13/0.23 | 3/0.00 | 27/0.64 | 0 | 3/0.00 |
| Yoldiella lenticula (Bi) | 56 | 37/0.19 | 20/0.03 | 0 | 0 | 60/0.16 | 143/0.64 | 0 | 3/0.01 | 270/0.78 |
| Heteromastus filiformis (Pol) | 56 | 7/0.01 | 13/0.13 | 0 | 83/0.30 | 103/0.21 | 53/0.12 | 0 | 13/0.05 | 13/0.02 |
| Spiophanes kroeyeri (Pol) | 56 | 23/0.12 | 17/0.02 | 50/0.11 | 0 | 3/0.00 | 0 | 67/0.30 | 10/0.01 | 0 |
| Pholoe assimilis (Po) | 56 | 0 | 47/0/04 | 17/0.01 | 0 | 30/0.02 | 33/0.02 | 10/0.01 | 3/0.00 | 0 |
| Yoldiella intermedia (Bi) | 52 | 0 | 0 | 47/0.14 | 33/0.13 | 20/0.18 | 10/0.03 | 43/0.22 | 7/1.01 | 0 |
| Ophelina cylindricaudata (Pol) | 52 | 3/0.00 | 7/0.01 | 13/0.01 | 0 | 7/0.01 | 0 | 30/0.02 | 23/0.02 | 53/0.04 |
| Characteristics | Community | ||||
|---|---|---|---|---|---|
| Southern Complex | Northern Complex | ||||
| I | II | III | IV | V | |
| SR | 96 | 73 | 117 | 82 | 68 |
| N | 5110 ±340 | 3030 ± 590 | 1300 ± 110 | 945 ± 110 | 1250 |
| B | 55 ± 17 | 39 ± 18 | 61 ± 16 | 11 ± 3 | 48 |
| H’ | 3.7 ± 0.2 | 3.2 ± 0.4 | 5.2 ± 0.2 | 4.8 ± 0.1 | 4.5 |
| J’ | 0.59 ± 0.1 | 0.56 ± 0.02 | 0.83 ± 0.01 | 0.84 ± 0.01 | 0.74 |
| D | 0.17 ± 0.02 | 0.21 ± 0.06 | 0.05 ± 0.00 | 0.06 ± 0.00 | 0.09 |
| DE | 0.04 ± 0.03 | −0.26 ± 0.16 | −0.56 ± 0.10 | −0.37 ± 0.14 | −0.38 |
| Binf:Bepf | 1:1 | 1:5 | 1:2.4 | 4:1 | 1.5:1 |
| Dominants by respiration rate | Spiochaetopterus typicus (18%) Laonice cirrata (10%) | Y. nana (23%) Strongylocentrotus pallidus (18%) Parathyasira equalis (10%) | S. typicus (46%) O. borealis (21%) | S. typicus (31%) Aglaophamus malmgreni (15%) Abyssoninoe cf. abyssorum (15%) | Ctenodiscus crispatus (17%) Aglaophamus malmgreni (13%) |
| Dominants by biomass | Ophiopleura borealis (25%) S. typicus (15%) | S. pallidus (47%) O. borealis (12%) Y. nana (8%) | O. borealis (40%) S. typicus (24%) Ophiacantha bidentata (22%) | S. typicus (55%) Aglaophamus malmgreni (11%) | Ctenodiscus crispatus (49%) O. borealis (30%) |
| Dominants by abundance | Mendicula ferruginosa (27%) Yoldiella nana (24%) | Y. nana (36%) M. ferruginosa (24%) | Ektonodiastylis nimia (11%) Prionospio cirrifera (9%) | Prionospio cirrifera (11%) | Y. nana (25%) |
| Dominating trophic groups by biomass | Surface deposit feeder (31%) Sub-surface deposit feeder (24%) Carnivores (18%) | Sub-surface deposit feeders (50%) | Sessile suspension feeders (58%) | Sessile suspension feeders (57%) | Mobile suspension feeders (41%) Sub-surface deposit feeders (33%) |
| Characteristic species | Maldane sarsi, Praxillella gracilis, Golfingia (Golfingia) vulgaris vulgaris, Y. nana, Parathyasira equalis, M. ferruginosa | Parathyasira equalis, M. ferruginosa, Yoldiella lenticula, O. borealis | Nephasoma (Nephasoma) diaphanes diaphanes, Prionospio cirrifera, Spiophanes kroeyeri, Harpinia mucronata, Ektonodiastylis nimia, Y. nana, M. ferruginosa | Prionospio cirrifera, Spiophanes kroeyeri, Notomastus latericeus, Astarte crenata, Bathyarca glacialis, Ektonodiastylis nimia | Ophiopleura borealis, Y. nana, Astarte crenata, |
| Depth, m | 445–502 | 392–430 | 343–507 | 361–427 | 186 |
| Stations | 10, 11 | 12, 20 | 5, 6 | 4, 15 | 17 |
| Variable | Abundance | Diversity | |||||
|---|---|---|---|---|---|---|---|
| EV | F | P | Variable | EV | F | P | |
| OM | 44 | 0.001 | 5.44 | OM | 36 | 0.038 | 4.01 |
| S | 7 | 0.474 | 0.86 | T | 11 | 0.458 | 0.77 |
| IFP | 7 | 0.499 | 0.9 | Sed | 3 | 0.734 | 0.25 |
| T | 7 | 0.494 | 0.82 | S | 7 | 0.483 | 0.62 |
| H | 6 | 0.613 | 0.57 | IFP | 3 | 0.736 | 0.22 |
| Sed | 5 | 0.738 | 0.41 | H | 2 | 0.806 | 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlova, L.V.; Dvoretsky, A.G.; Frolov, A.A.; Zimina, O.L.; Evseeva, O.Y.; Dikaeva, D.R.; Rumyantseva, Z.Y.; Panteleeva, N.N. The Impact of Sea Ice Loss on Benthic Communities of the Makarov Strait (Northeastern Barents Sea). Animals 2023, 13, 2320. https://doi.org/10.3390/ani13142320
Pavlova LV, Dvoretsky AG, Frolov AA, Zimina OL, Evseeva OY, Dikaeva DR, Rumyantseva ZY, Panteleeva NN. The Impact of Sea Ice Loss on Benthic Communities of the Makarov Strait (Northeastern Barents Sea). Animals. 2023; 13(14):2320. https://doi.org/10.3390/ani13142320
Chicago/Turabian StylePavlova, Lyudmila V., Alexander G. Dvoretsky, Alexander A. Frolov, Olga L. Zimina, Olga Yu. Evseeva, Dinara R. Dikaeva, Zinaida Yu. Rumyantseva, and Ninel N. Panteleeva. 2023. "The Impact of Sea Ice Loss on Benthic Communities of the Makarov Strait (Northeastern Barents Sea)" Animals 13, no. 14: 2320. https://doi.org/10.3390/ani13142320