
Development of a New Marine Fish Continuous Cell Line Derived from Brain of Red Sea Bream (Pagrosomus major) and Its Application to Fish Virology and Heavy Metal Toxicology
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Establishment and Subculture of the RSBB Cell Line
2.2. Characterization of the RSBB Cell Line
2.2.1. Cryopreservation Efficiency
2.2.2. Immunophenotyping Characterization
2.2.3. Chromosome Analysis
2.2.4. Cell Growth Characteristics
2.2.5. Cell Transfection
2.3. Virology
2.3.1. Virus Sensitivity
2.3.2. PCR Detection for the Viruses
2.4. Metal Cytotoxicity to RSBB Cells
2.4.1. Cadmium Chloride (CdCl2) Exposure and Viability Test of RSBB Cells
2.4.2. Cell Apoptosis or Necrosis Induced by CdCl2
3. Results
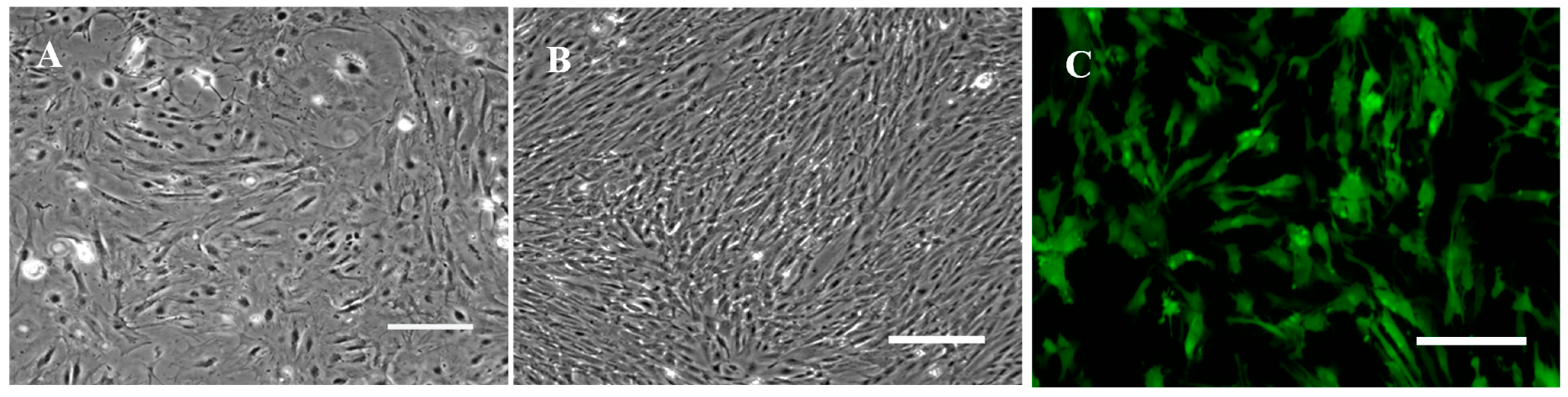
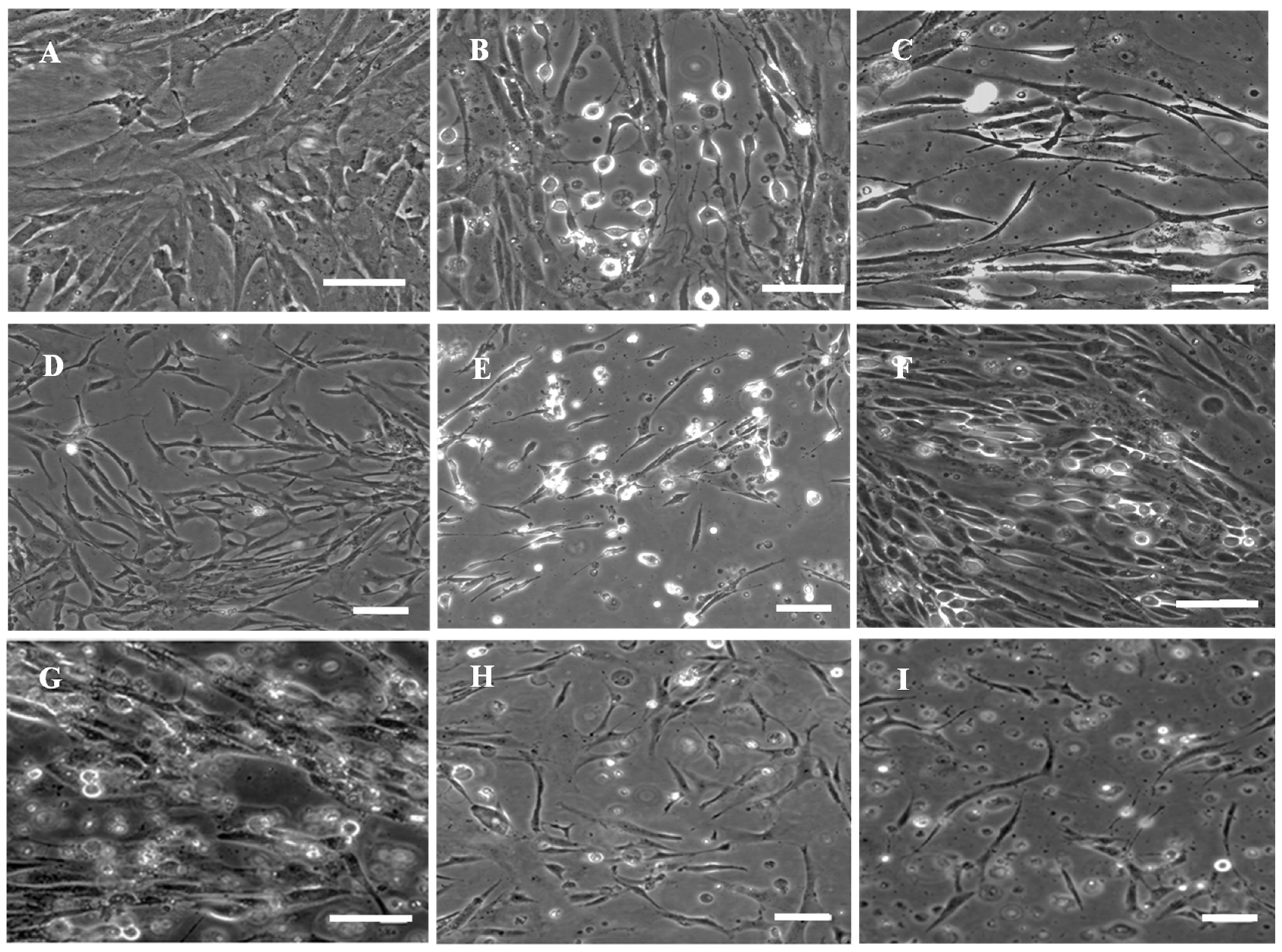
3.1. Primary Culture and Subculture of RSBB Cells
3.2. Cell Thawing Efficiency of Cryopreserved RSBB Cells
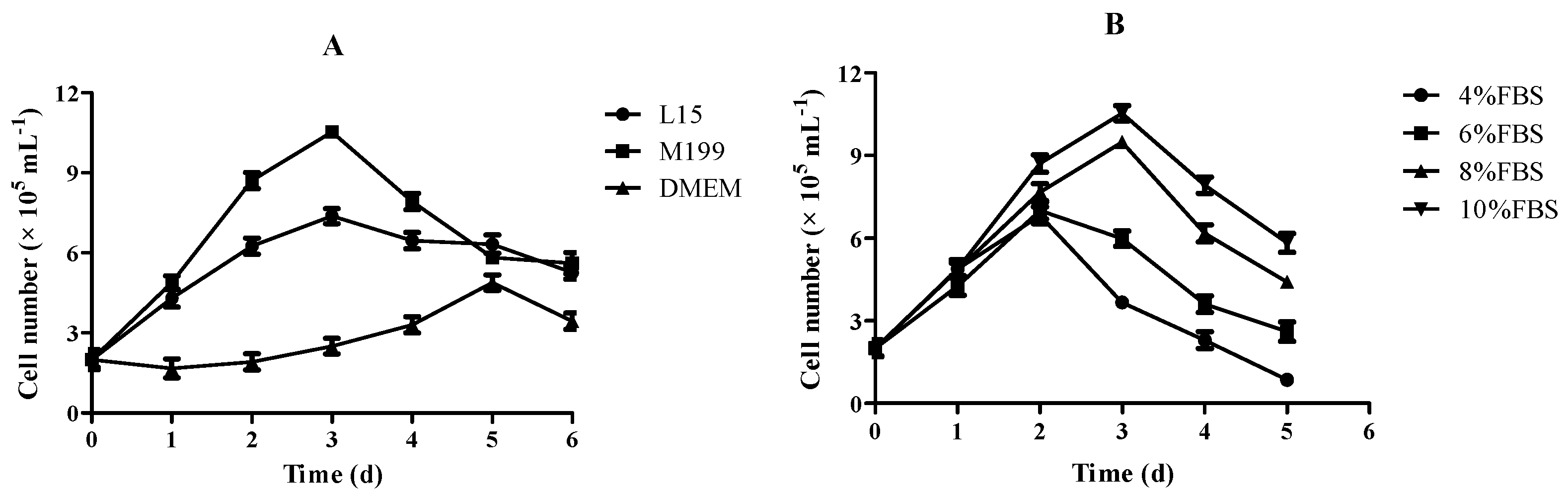
3.3. Effect of Culture Medium and FBS on Growth of RSBB Cells
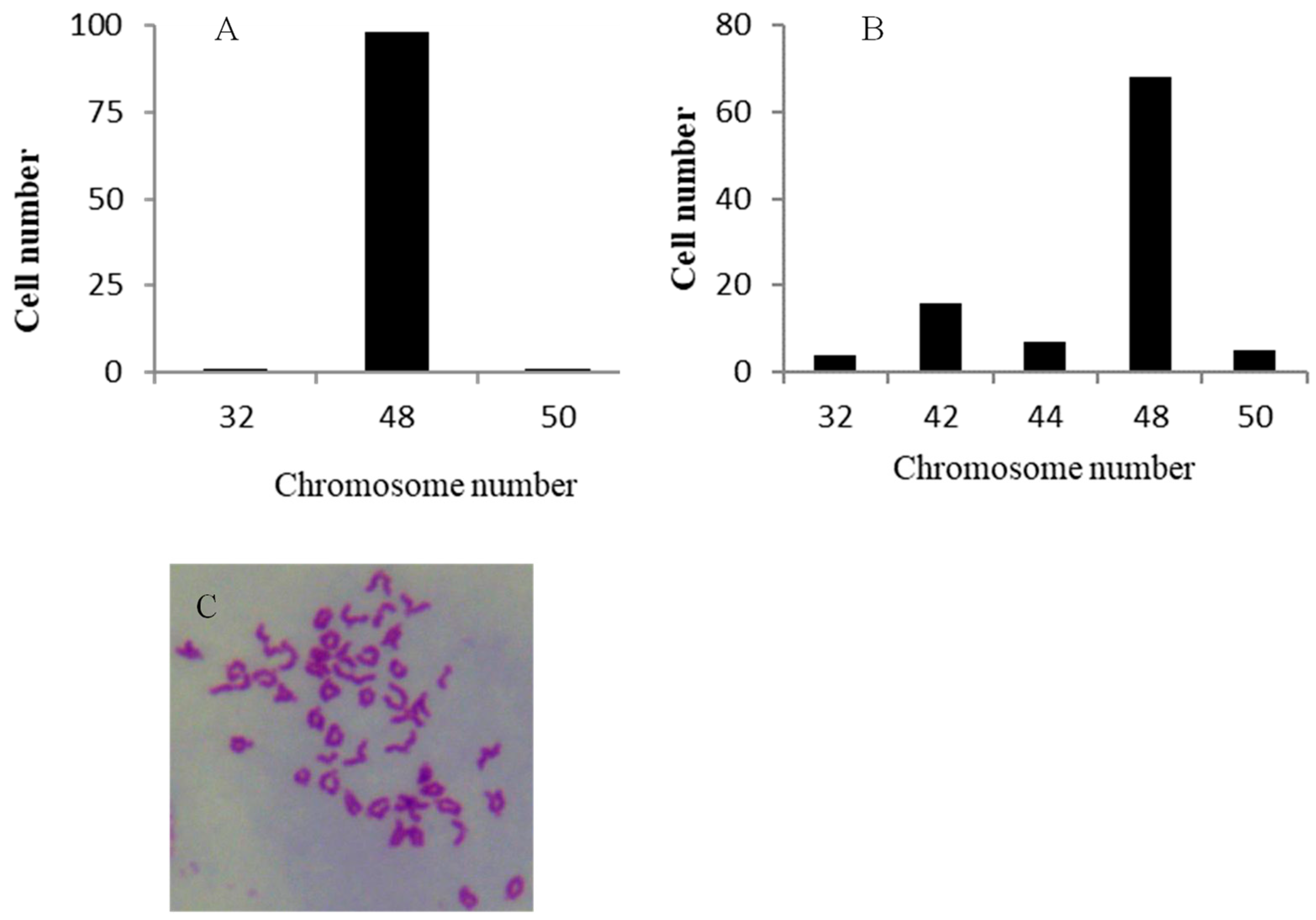
3.4. Chromosome Analysis
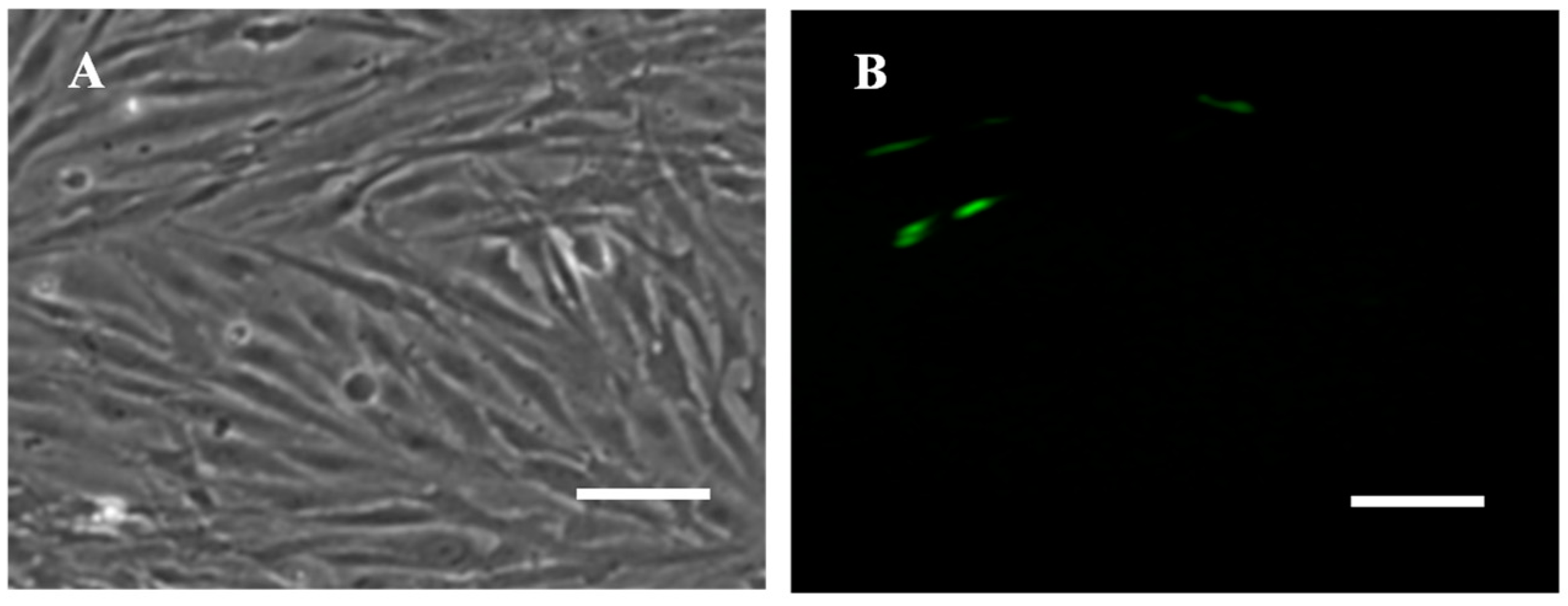
3.5. Transfection Efficiency
3.6. Virus Susceptibility
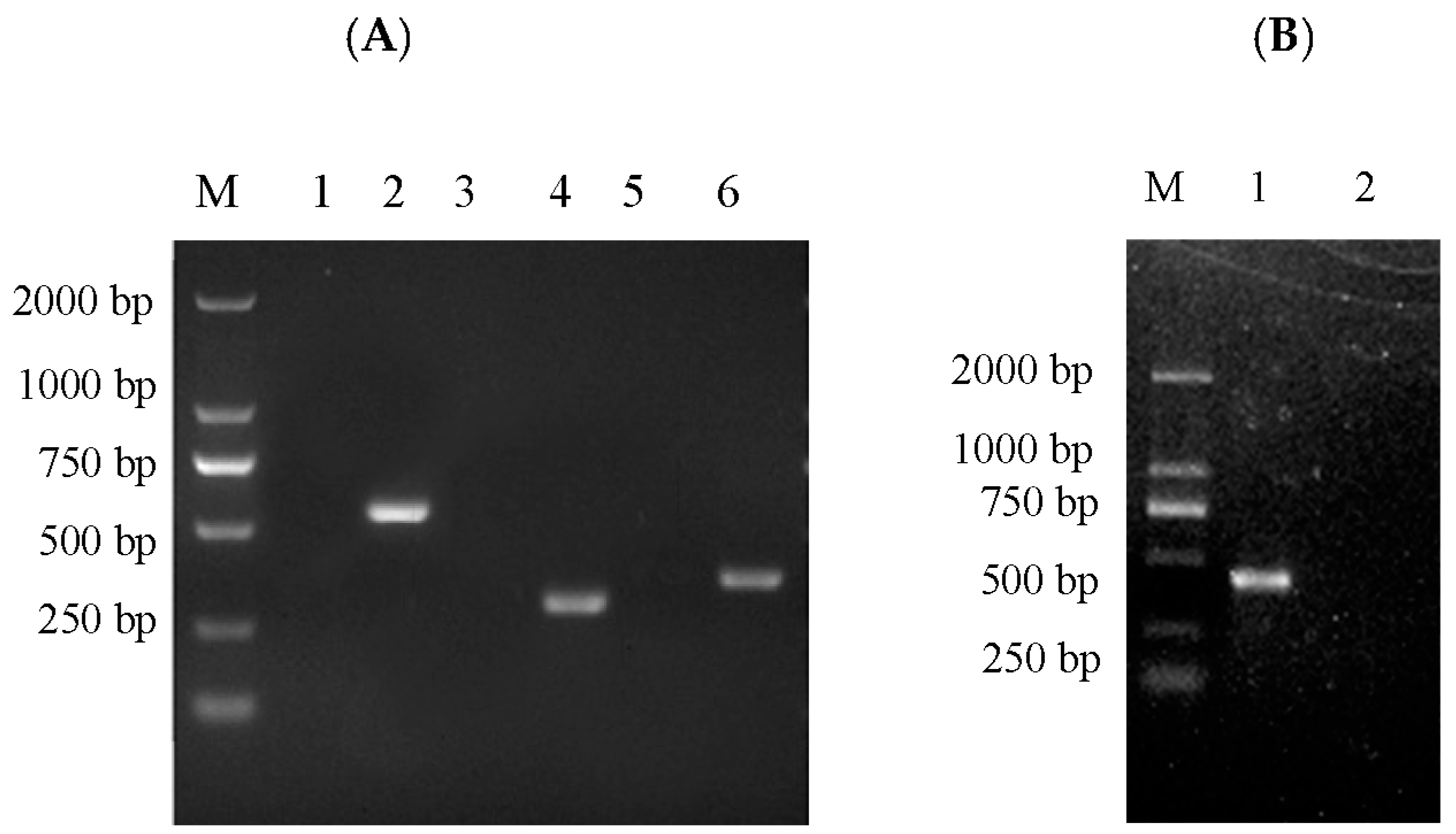
3.7. PCR Detection for Virus Specificity
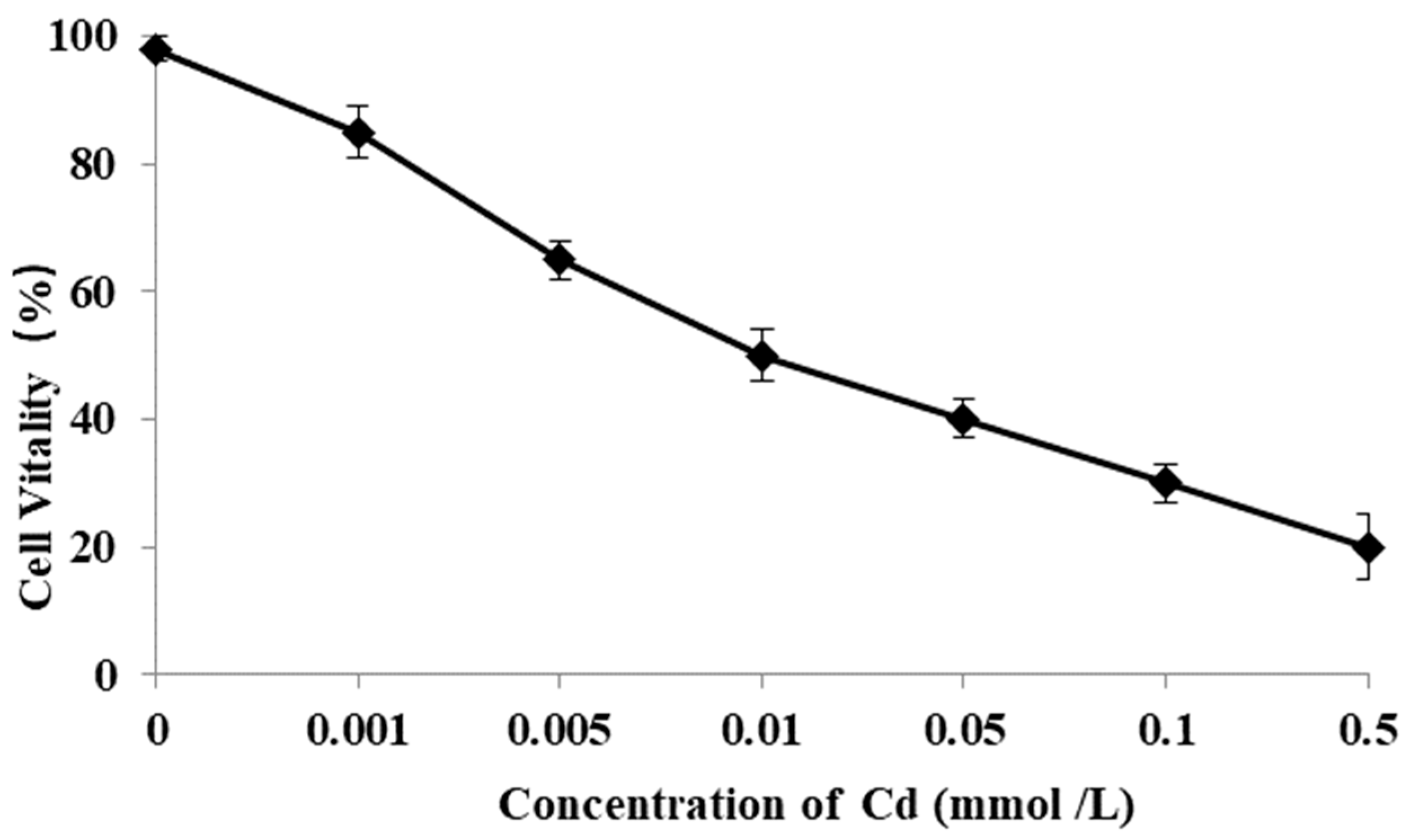
3.8. Cell Vitality Affected by Cd
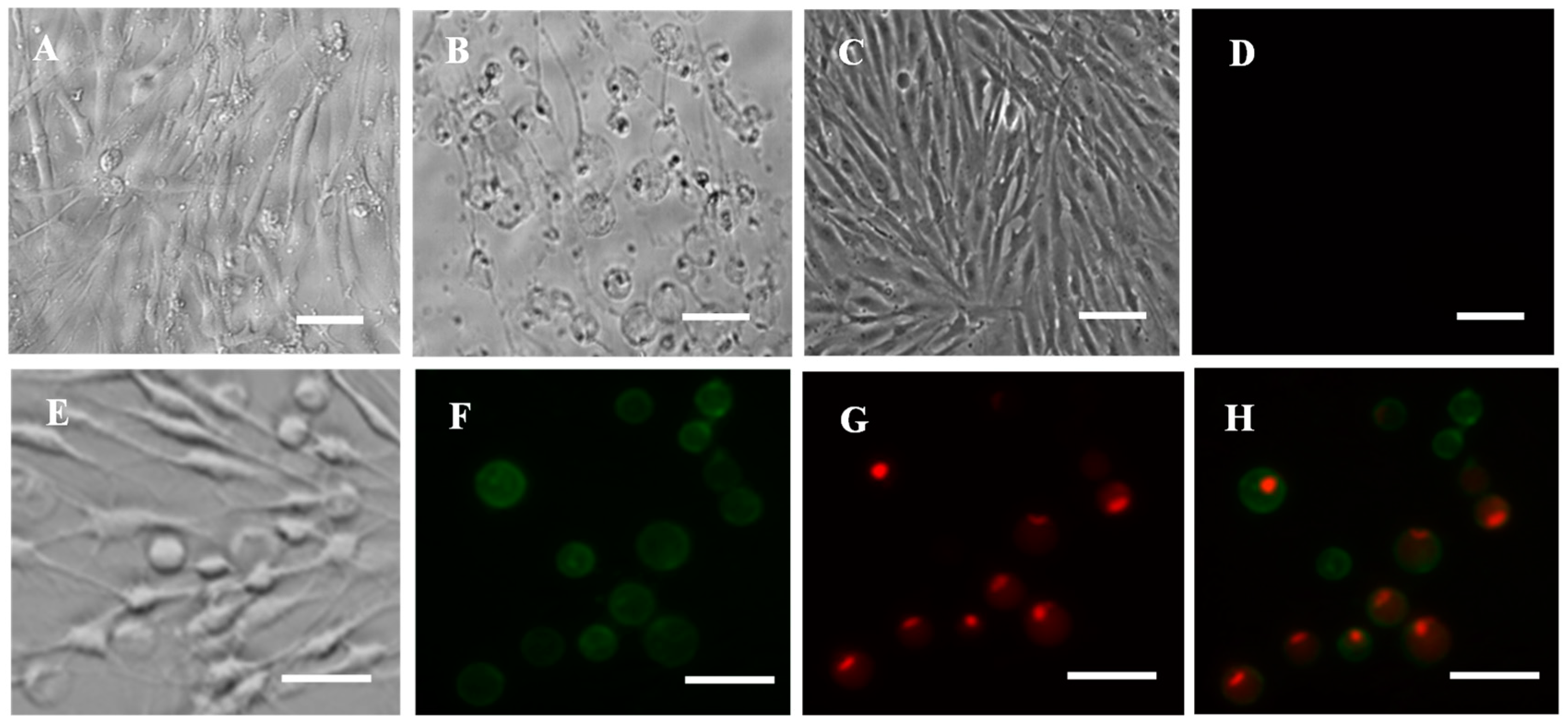
3.9. Cell Apoptosis and Necrosis Induced by Cd
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Uysal, K.; Köse, E.; Bülbül, M.; Dönmez, M.; Erdoğan, Y.; Koyun, M.; Ömeroğlu, Ç.; Özmal, F. The comparison of heavy metal accumulation ratios of some fish species in Enne Dame Lake (Kütahya/Turkey). Environ. Monit. Assess. 2009, 157, 355–362. [Google Scholar] [CrossRef]
- Fent, K. Fish cell lines as versatile tools in ecotoxicology: Assessment of cytotoxicity, cytochrome P4501A induction potential and estrogenic activity of chemicals and environmental samples. Toxicol. In Vitro 2001, 15, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Segner, H. Cytotoxicity Assays with Fish Cells as an Alternative to the Acute Lethality Test with Fish. Altern. Lab. Anim. 2004, 32, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, P.; Chaves-Pozo, E.; Meseguer, J.; Esteban, M.; Cuesta, A. Establishment of a new teleost brain cell line (DLB-1) from the European sea bass and its use to study metal toxicology. Toxicol. In Vitro 2017, 38, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Bloch, S.R.; Vo, N.T.K.; Walsh, S.K.; Chen, C.; Lee, L.E.J.; Hodson, P.V.; Bols, N.C. Development of a cell line from the American eel brain expressing endothelial cell properties. Vitr. Cell. Dev. Biol.-Anim. 2016, 52, 395–409. [Google Scholar] [CrossRef]
- Imajoh, M.; Ikawa, T.; Oshima, S.-I. Characterization of a new fibroblast cell line from a tail fin of red sea bream, Pagrus major, and phylogenetic relationships of a recent RSIV isolate in Japan. Virus Res. 2007, 126, 45–52. [Google Scholar] [CrossRef]
- Qin, Q.; Wu, T.; Jia, T.; Hegde, A.; Zhang, R. Development and characterization of a new tropical marine fish cell line from grouper, Epinephelus coioides susceptible to iridovirus and nodavirus. J. Virol. Methods 2006, 131, 58–64. [Google Scholar] [CrossRef]
- Frerichs, G.N.; Rodger, H.D.; Peric, Z. Cell Culture Isolation of Piscine Neuropathy Nodavirus from Juvenile Sea Bass, Dicentrarchus labrax. J. Gen. Virol. 1996, 77 Pt 9, 2067–2071. [Google Scholar] [CrossRef]
- Tong, S.-L.; Miao, H.-Z.; Li, H. Three new continuous fish cell lines of SPH, SPS and RSBF derived from sea perch (Lateolabrax japonicus) and red sea bream (Pagrosomus major). Aquaculture 1998, 169, 143–151. [Google Scholar] [CrossRef]
- Chen, S.; Ye, H.; Sha, Z.; Hong, Y. Derivation of a pluripotent embryonic cell line from red sea bream blastulas. J. Fish Biol. 2003, 63, 795–805. [Google Scholar] [CrossRef]
- Lopez-Jimena, B.; del Carmen Alonso, M.; Thompson, K.D.; Adams, A.; Infante, C.; Castro, D.; Borrego, J.J.; Garcia-Rosado, E. Tissue distribution of Red Spotted Grouper Nervous Necrosis Virus (RGNNV) genome in experimentally infected juvenile European seabass (Dicentrarchus labrax). Vet. Microbiol. 2011, 154, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jimena, B.; García-Rosado, E.; Thompson, K.D.; Adams, A.; Infante, C.; Borrego, J.J.; del Carmen Alonso, M. Distribution of red-spotted grouper nervous necrosis virus (RGNNV) antigens in nervous and non-nervous organs of European seabass (Dicentrarchus labrax) during the course of an experimental challenge. J. Veter.-Sci. 2012, 13, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Starkey, W.G.; Ireland, J.H.; Muir, K.F.; Shinn, A.P.; Richards, R.H.; Ferguson, H.W. Isolation of nodavirus from Scottish farmed halibut, Hippoglossus hippoglossus (L). J. Fish Dis. 2000, 23, 418–422. [Google Scholar] [CrossRef]
- Bloch, B.; Gravningen, K.; Larsen, J. Encephalomyelitis among turbot associated with a picornavirus-like agent. Dis. Aquat. Org. 1991, 10, 65–70. [Google Scholar] [CrossRef]
- Lewis, L.M.; Lall, S.P.; Witten, P.E. Morphological descriptions of the early stages of spine and vertebral development in hatchery-reared larval and juvenile Atlantic halibut (Hippoglossus hippoglossus). Aquaculture 2004, 241, 47–59. [Google Scholar] [CrossRef]
- Bovo, G.; Nishizawa, T.; Maltese, C.; Borghesan, F.; Mutinelli, F.; Montesi, F.; De Mas, S. Viral encephalopathy and retinopathy of farmed marine fish species in Italy. Virus Res. 1999, 63, 143–146. [Google Scholar] [CrossRef]
- Fu, X.; Li, N.; Lai, Y.; Luo, X.; Wang, Y.; Shi, C.; Huang, Z.; Wu, S.; Su, J. A novel fish cell line derived from the brain of Chinese perch Siniperca chuatsi: Development and characterization. J. Fish Biol. 2015, 86, 32–45. [Google Scholar] [CrossRef]
- Thangaraj, R.S.; Ravi, C.; Kumar, R.; Dharmaratnam, A.; Saidmuhammed, B.V.; Pradhan, P.K.; Sood, N. Derivation of two tilapia (Oreochromis niloticus) cell lines for efficient propagation of Tilapia Lake Virus (TiLV). Aquaculture 2018, 492, 206–214. [Google Scholar] [CrossRef]
- Freshney, R.I. Culture of animal cells. In A Manual of Basic Technique; Freshney, R.I., Ed.; Wiley/Liss Press: New York, NY, USA, 1994; pp. 387–389. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Hu, X.; Fu, X.; Li, N.; Dong, X.; Zhao, L.; Lan, J.; Ji, W.; Zhou, W.; Ai, T.; Wu, S.; et al. Transcriptomic analysis of Mandarin fish brain cells infected with infectious spleen and kidney necrosis virus with an emphasis on retinoic acid-inducible gene 1-like receptors and apoptosis pathways. Fish Shellfish. Immunol. 2015, 45, 619–629. [Google Scholar] [CrossRef]
- Servili, A.; Bufalino, M.R.; Nishikawa, R.; de Melo, I.S.; Muñoz-Cueto, J.A.; Lee, L.E. Establishment of long term cultures of neural stem cells from adult sea bass, Dicentrarchus labrax. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 152, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.-C.; Teng, Y.-C.; Wang, C.-S.; Lu, C.-H. Establishment and characterization of three cell lines derived from the rockfish grouper Epinephelus quoyanus: Use for transgenic studies and cytotoxicity testing. Aquaculture 2009, 294, 147–151. [Google Scholar] [CrossRef]
- Liu, X.F.; Wu, Y.H.; Wei, S.N.; Wang, N.; Li, Y.Z.; Zhang, N.W.; Li, P.F.; Qin, Q.W.; Chen, S.L. Establishment and characterization of a brain-cell line from kelp grouper Epinephelus moara. J. Fish Biol. 2018, 92, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Z.; Wang, Q.; Zeng, W.; Li, Y.; Yin, J.; Bergmann, S.M.; Zhu, X. Establishment of a brain cell line obtained from hybrids of Channa argus × Channa maculata for the detection of tilapia lake virus. Microb. Pathog. 2020, 138, 103810. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Palacios, M.; Esteban, M.; Cuesta, A. Establishment of a brain cell line (SaB-1) from gilthead seabream and its application to fish virology. Fish Shellfish. Immunol. 2020, 106, 161–166. [Google Scholar] [CrossRef]
- Ahmed, V.I.; Babu, V.S.; Chandra, V.; Nambi, K.; Thomas, J.; Bhonde, R.; Hameed, A.S. A new fibroblastic-like cell line from heart muscle of the Indian major carp (Catla catla): Development and characterization. Aquaculture 2009, 293, 180–186. [Google Scholar] [CrossRef]
- Ziniu, Y.; Xiaoyu, K.; Zongyong, X. Studies on karyotypes of fishes of economic importance in coastal waters of Shandong peninsula. J. Fish. Sci. China 1995, 2, 1–6. [Google Scholar]
- Krumschnabel, G.; Ebner, H.L.; Hess, M.W.; Villunger, A. Apoptosis and necroptosis are induced in rainbow trout cell lines exposed to cadmium. Aquat. Toxicol. 2010, 99, 73–85. [Google Scholar] [CrossRef]
- Morcillo, P.; Cordero, H.; Meseguer, J.; Esteban, M.Á.; Cuesta, A. In vitro immunotoxicological effects of heavy metals on European sea bass (Dicentrarchus labrax L.) head-kidney leucocytes. Fish Shellfish. Immunol. 2015, 47, 245–254. [Google Scholar] [CrossRef]
- Maracine, M.; Segner, H. Cytotoxicity of metals in isolated fish cells: Importance of the cellular glutathione status. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 120, 83–88. [Google Scholar] [CrossRef]
- Doan, Q.K.; Vandeputte, M.; Chatain, B.; Morin, T.; Allal, F. Viral encephalopathy and retinopathy in aquaculture: A review. J. Fish Dis. 2017, 40, 717–742. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.K.; Baeck, G.W.; Kim, J.H.; Choresca, C.H.; Park, S.C. Molecular Detection of Betanodavirus in Wild Marine Fish Populations in Korea. J. Veter.-Diagn. Investig. 2008, 20, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Lin, Q.; Liang, H.; Liu, L.; Huang, Z.; Li, N.; Su, J. The biological features and genetic diversity of novel fish rhabdovirus isolates in China. Arch. Virol. 2017, 162, 2829–2834. [Google Scholar] [CrossRef]
- He, J.G.; Wang, S.P.; Zeng, K.; Huang, Z.J.; Chan, S.M. Systemic disease caused by an iridovirus-like agent in cultured mandarinfish, Siniperca chuatsi (Basilewsky), in China. J. Fish Dis. 2000, 23, 219–222. [Google Scholar] [CrossRef]
- Iwamoto, T.; Nakai, T.; Mori, K.I.; Arimoto, M.; Furusawa, I. Cloning of the fish cell line SSN-1 for piscine nodaviruses. Dis. Aquat. Org. 2000, 43, 81–89. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Primer Sequences | Accession Number | Fragment Length/bp |
|---|---|---|---|
| NNV | R3:5′-CGAGTCAACACGGGTGGAGA-3′ | MN496376.1 | 420 |
| F2:5′-CGTGTCAGTCATGTGTCGCT-3′ | |||
| SGIV | F:5′-CGGGCAAGAGTTTTCGGTC-3′ | MK107821.1 | 372 |
| R:5′-AACGGCAACGGGAGCACTA-3′ | |||
| ISKNV | F:5′-CGTGAGACCGTGCGTAGT-3′ | AF371960 | 562 |
| R:5′-AGGGTGACGGTCGATATG-3′ | |||
| SCRV | F:5′-CTGAATCTCCAAGAATGGAAAACC-3′ | DQ399789.1 | 317 |
| R:5′-AATTTTGGCCAACTCGATTC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Fu, X.; Zhang, M.; Liang, H.; Niu, Y.; Lin, Q.; Ma, B.; Liu, L.; Li, N. Development of a New Marine Fish Continuous Cell Line Derived from Brain of Red Sea Bream (Pagrosomus major) and Its Application to Fish Virology and Heavy Metal Toxicology. Animals 2023, 13, 3524. https://doi.org/10.3390/ani13223524
Luo X, Fu X, Zhang M, Liang H, Niu Y, Lin Q, Ma B, Liu L, Li N. Development of a New Marine Fish Continuous Cell Line Derived from Brain of Red Sea Bream (Pagrosomus major) and Its Application to Fish Virology and Heavy Metal Toxicology. Animals. 2023; 13(22):3524. https://doi.org/10.3390/ani13223524
Chicago/Turabian StyleLuo, Xia, Xiaozhe Fu, Min Zhang, Hongru Liang, Yinjie Niu, Qiang Lin, Baofu Ma, Lihui Liu, and Ningqiu Li. 2023. "Development of a New Marine Fish Continuous Cell Line Derived from Brain of Red Sea Bream (Pagrosomus major) and Its Application to Fish Virology and Heavy Metal Toxicology" Animals 13, no. 22: 3524. https://doi.org/10.3390/ani13223524