
Spermatological Characterization of the Cestode Meggittina gerbilli (Cyclophyllidea: Catenotaeniidae), a Parasite of Gerbils, Gerbillus gerbillus and Gerbillus campestris (Rodentia: Muridae) in Tunisia
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
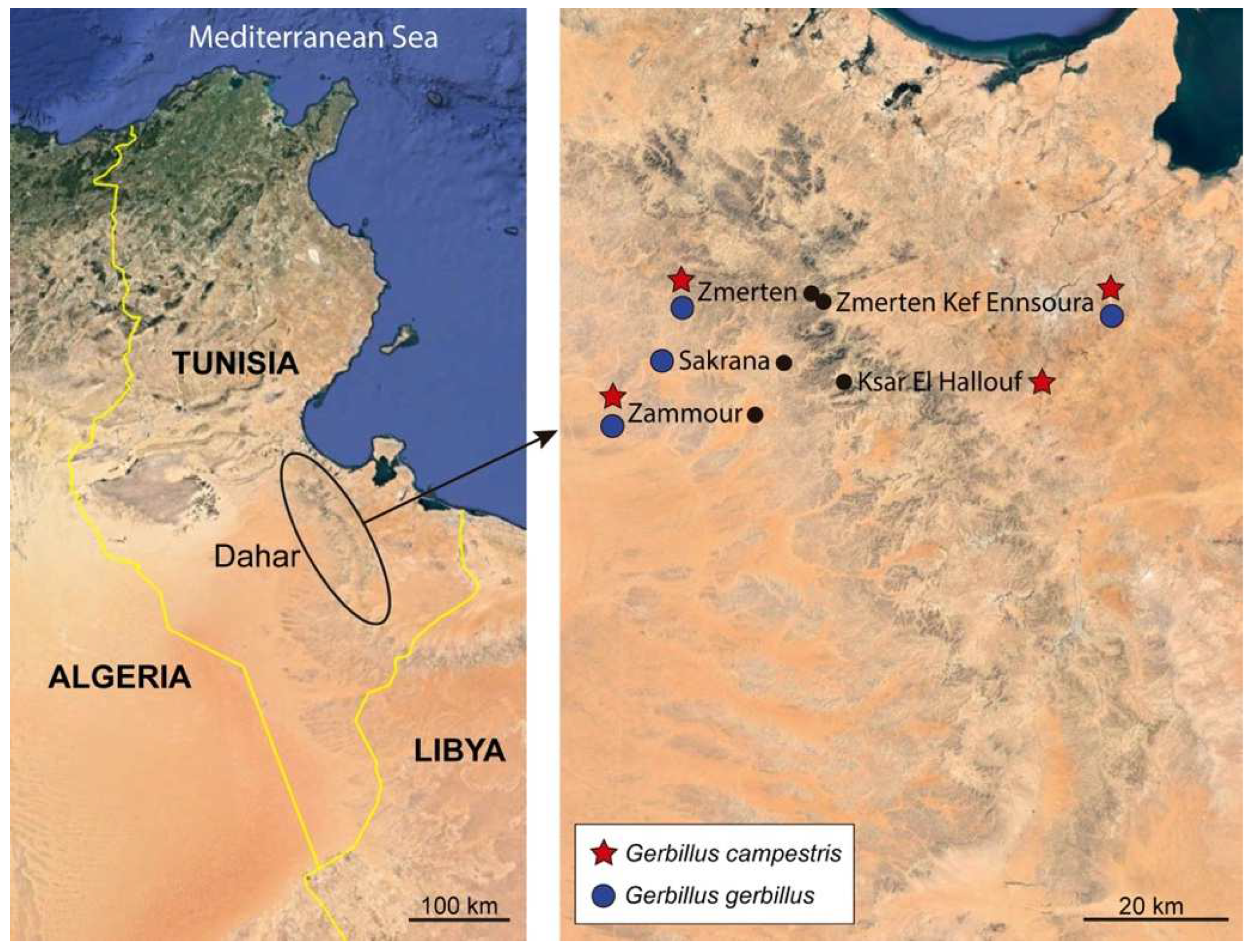
2.1. Specimens
2.2. Species Identification
2.3. Transmission Electron Microscopy Study
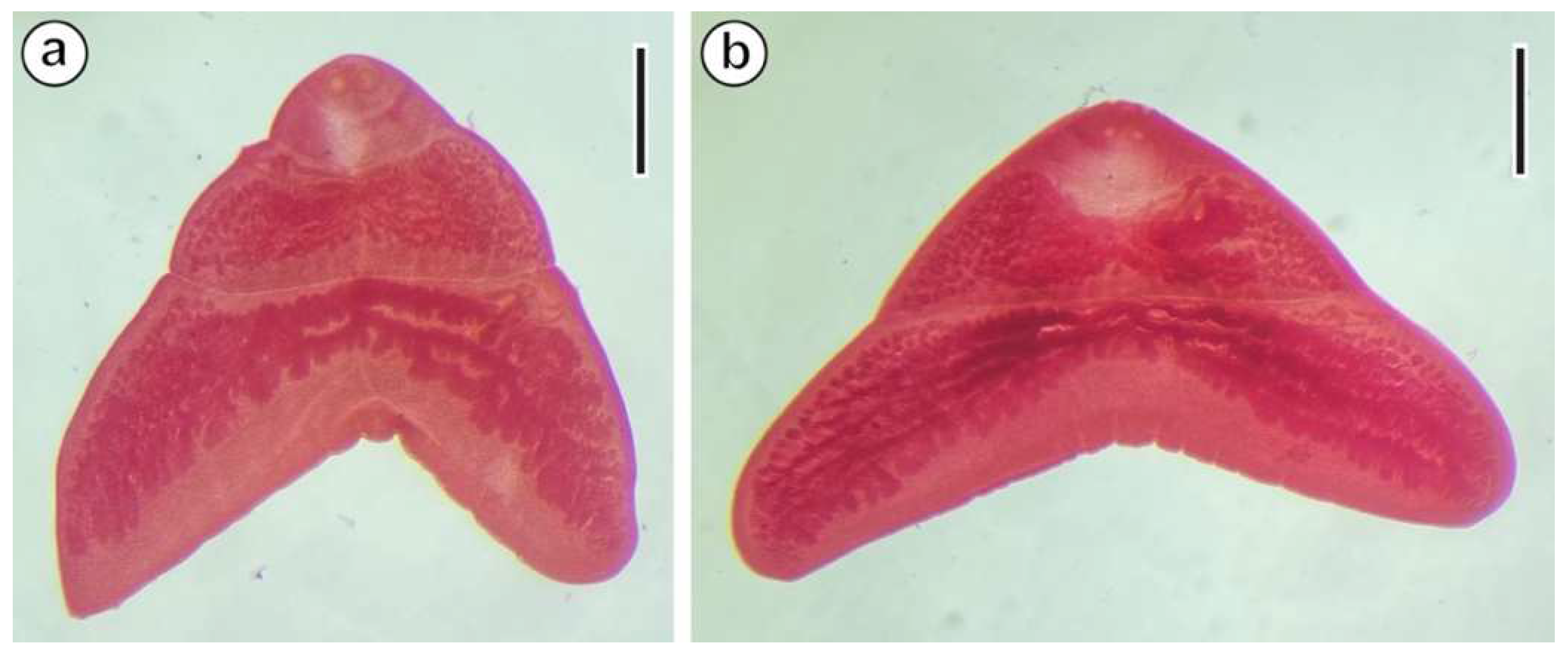
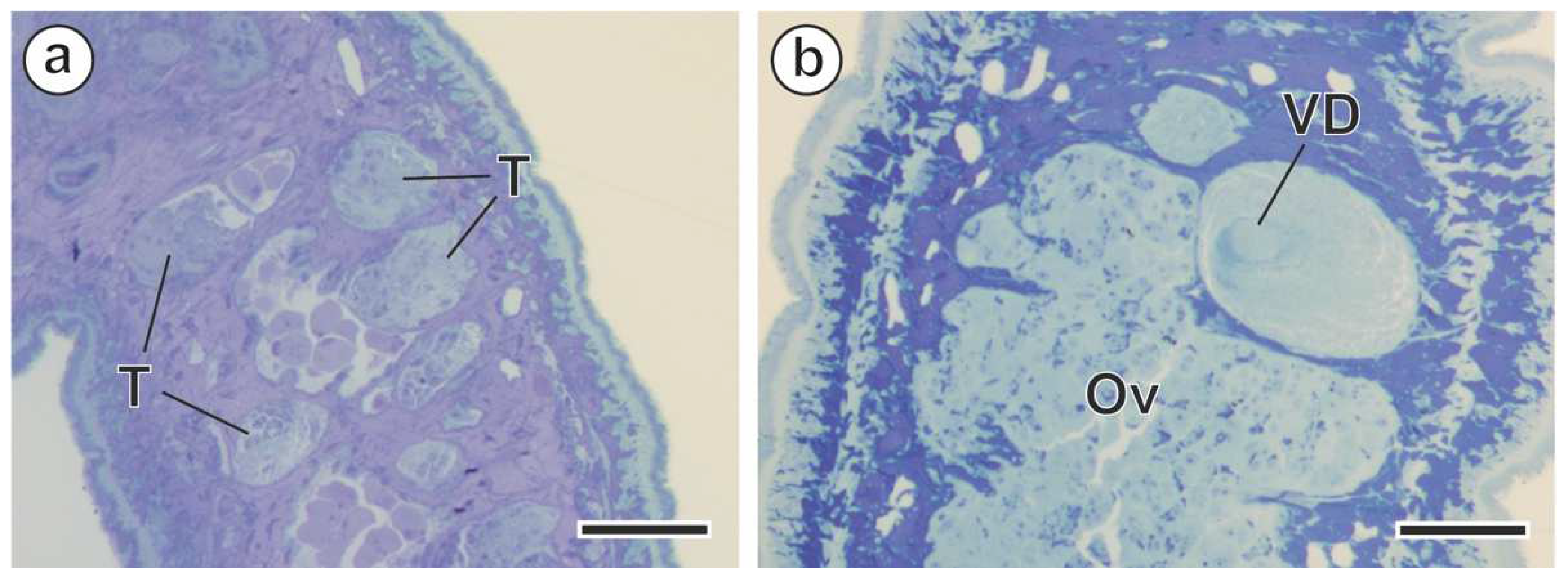
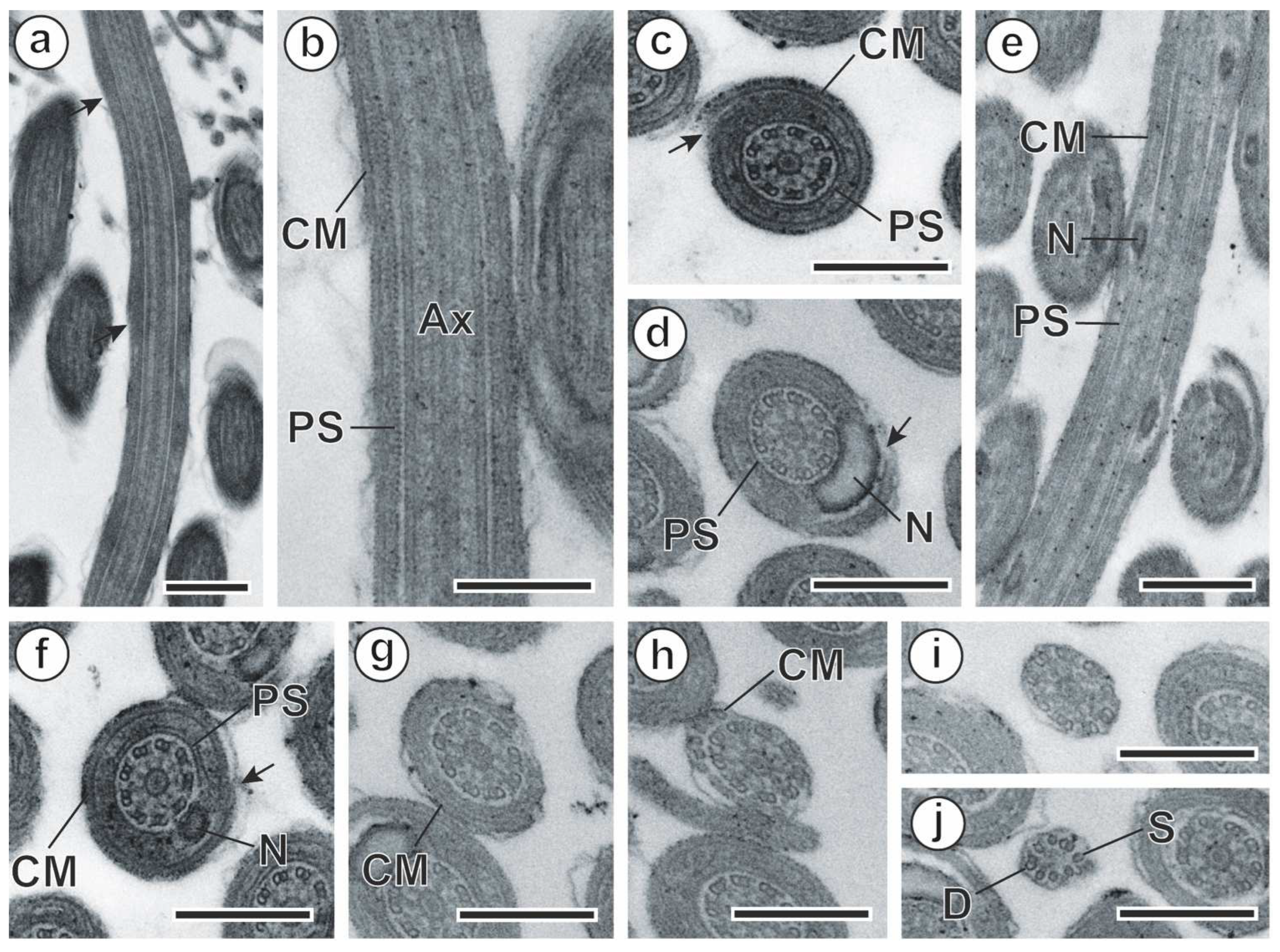
3. Results
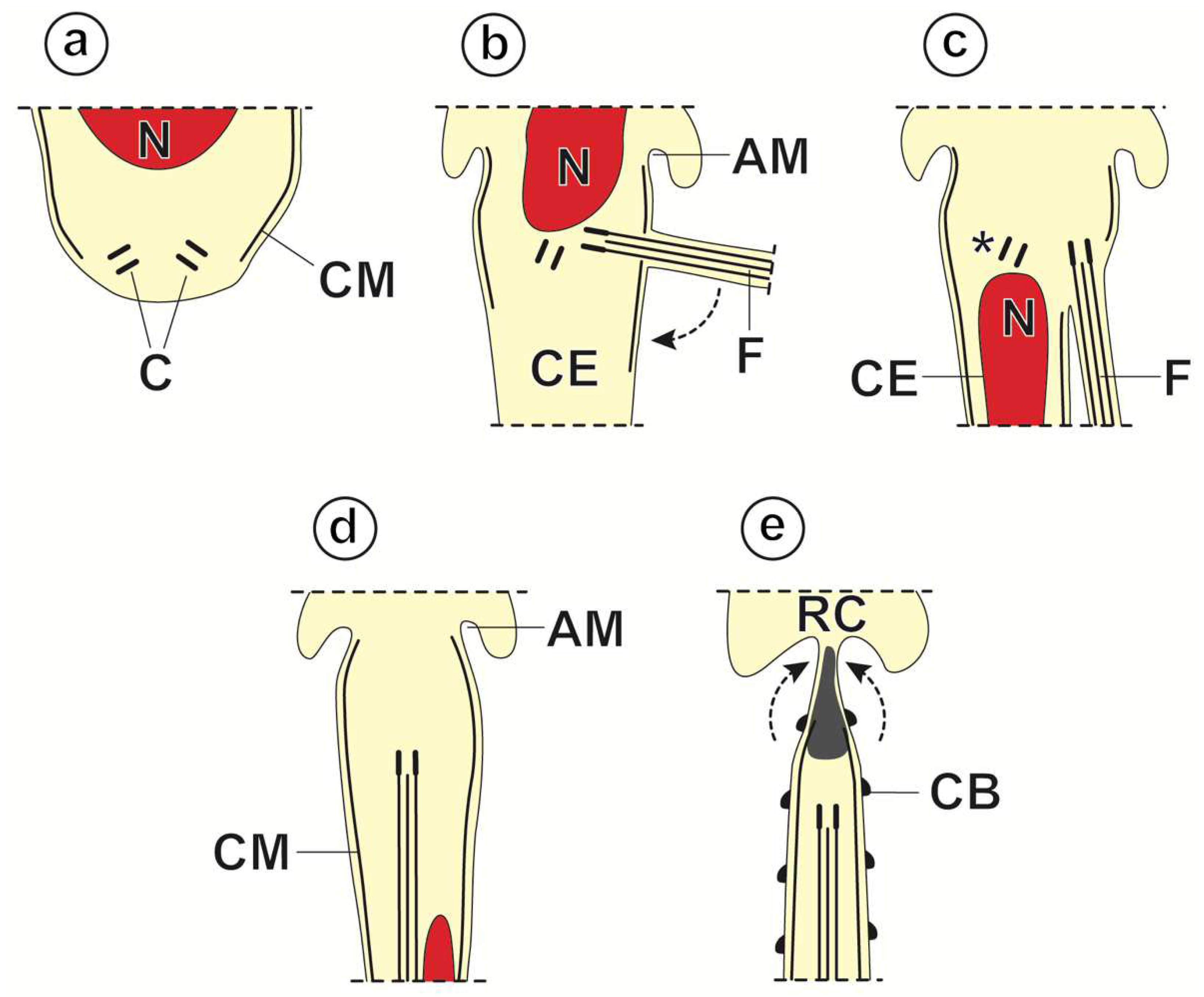
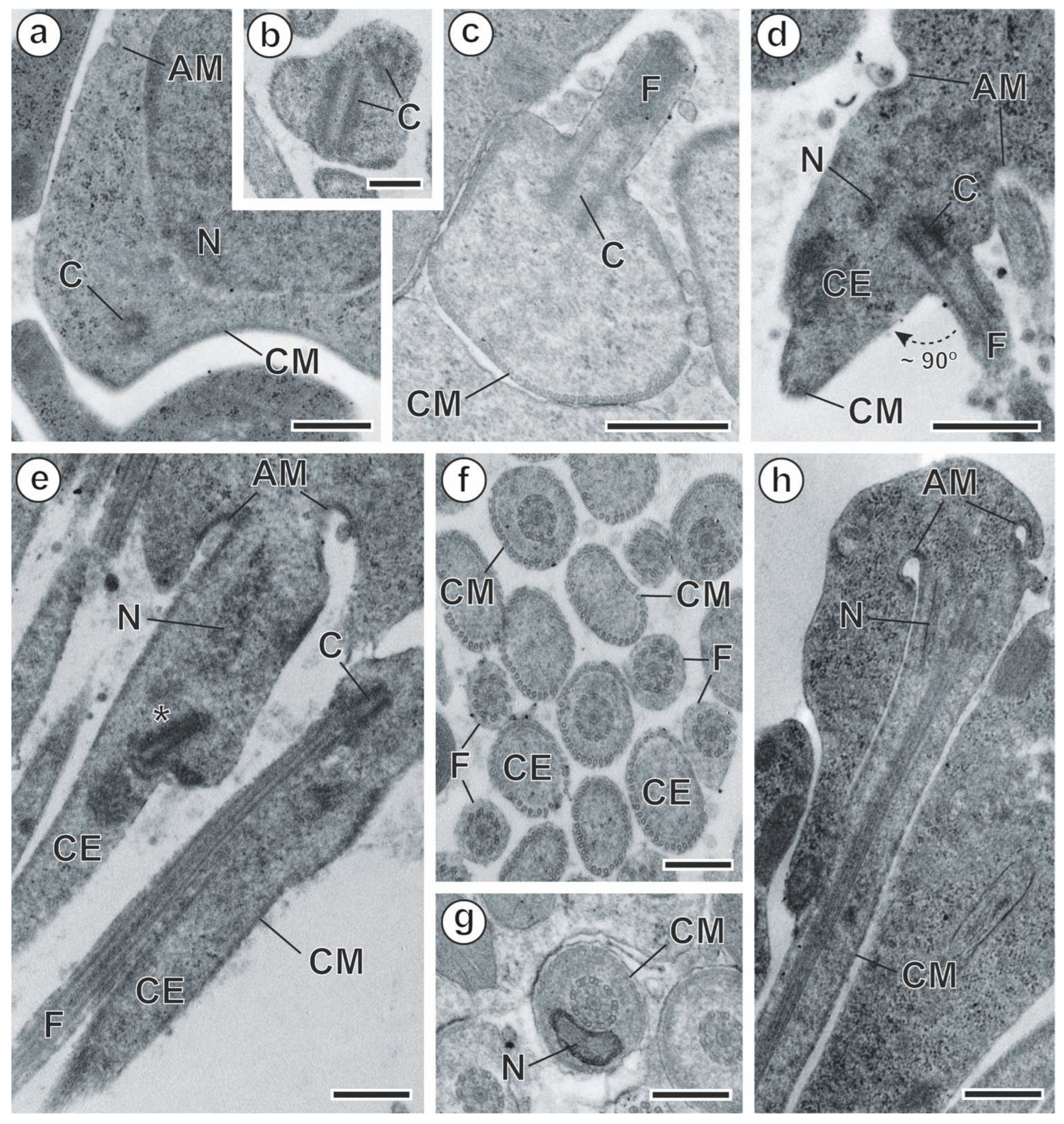
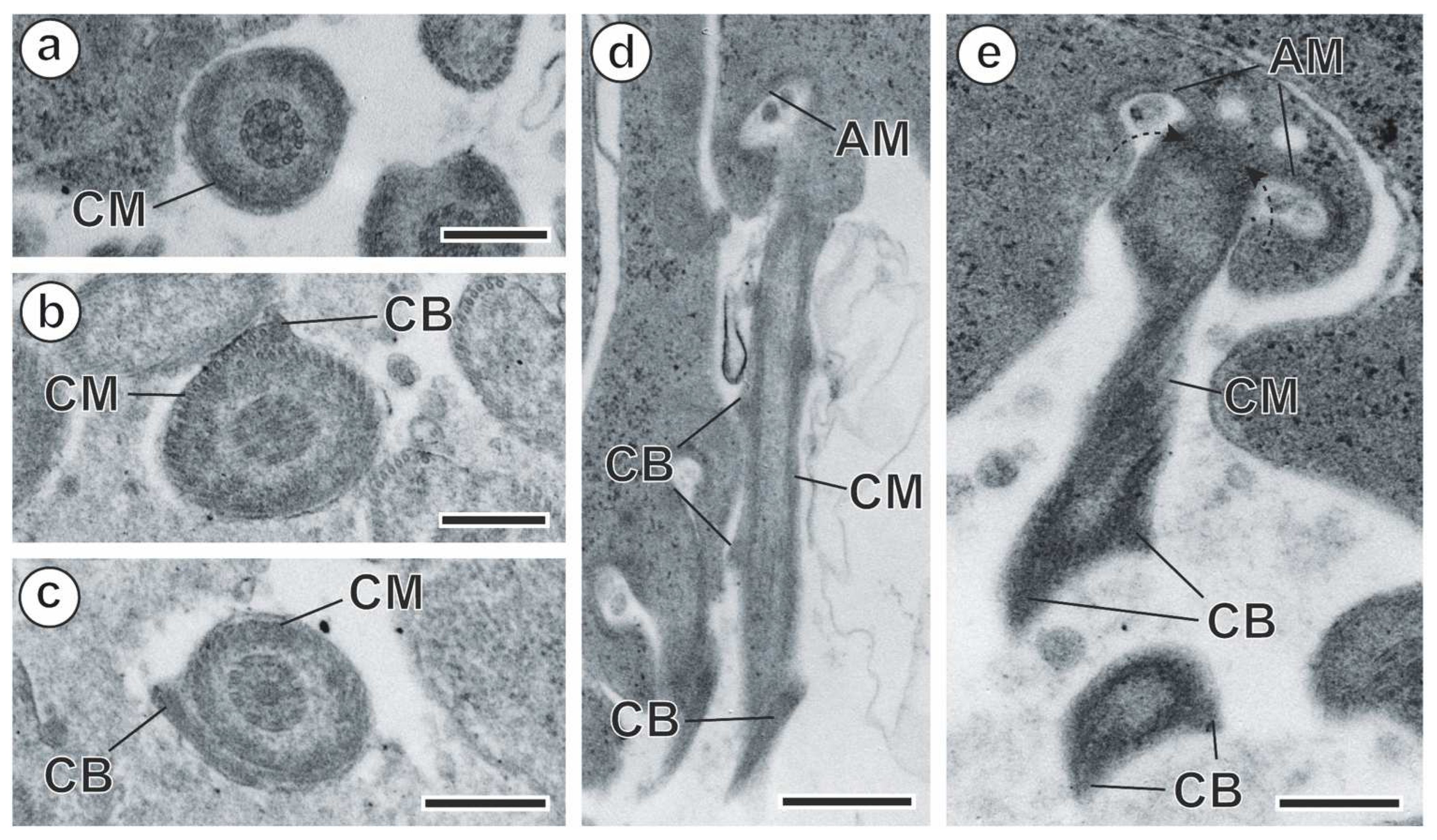
3.1. Spermiogenesis
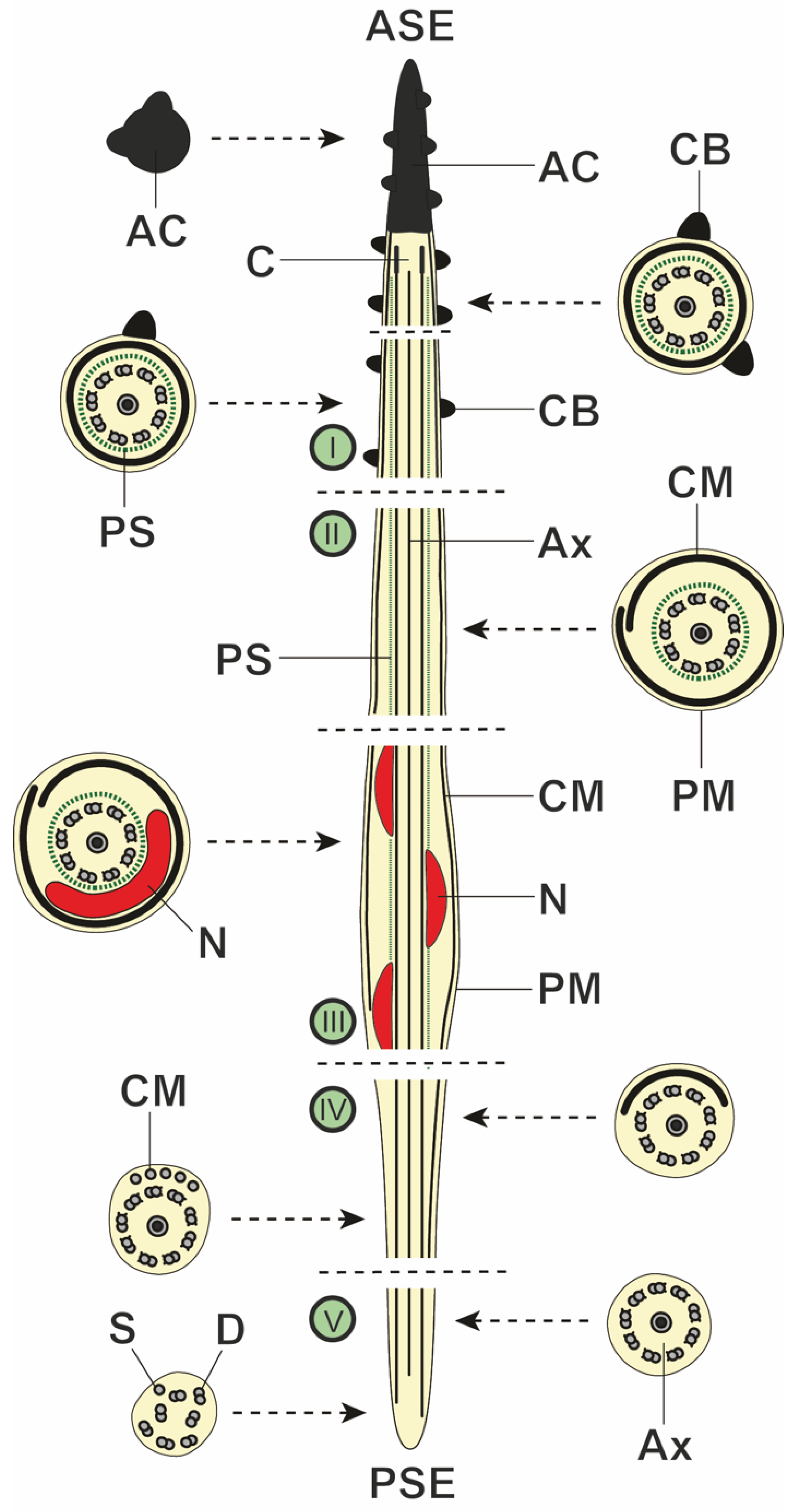
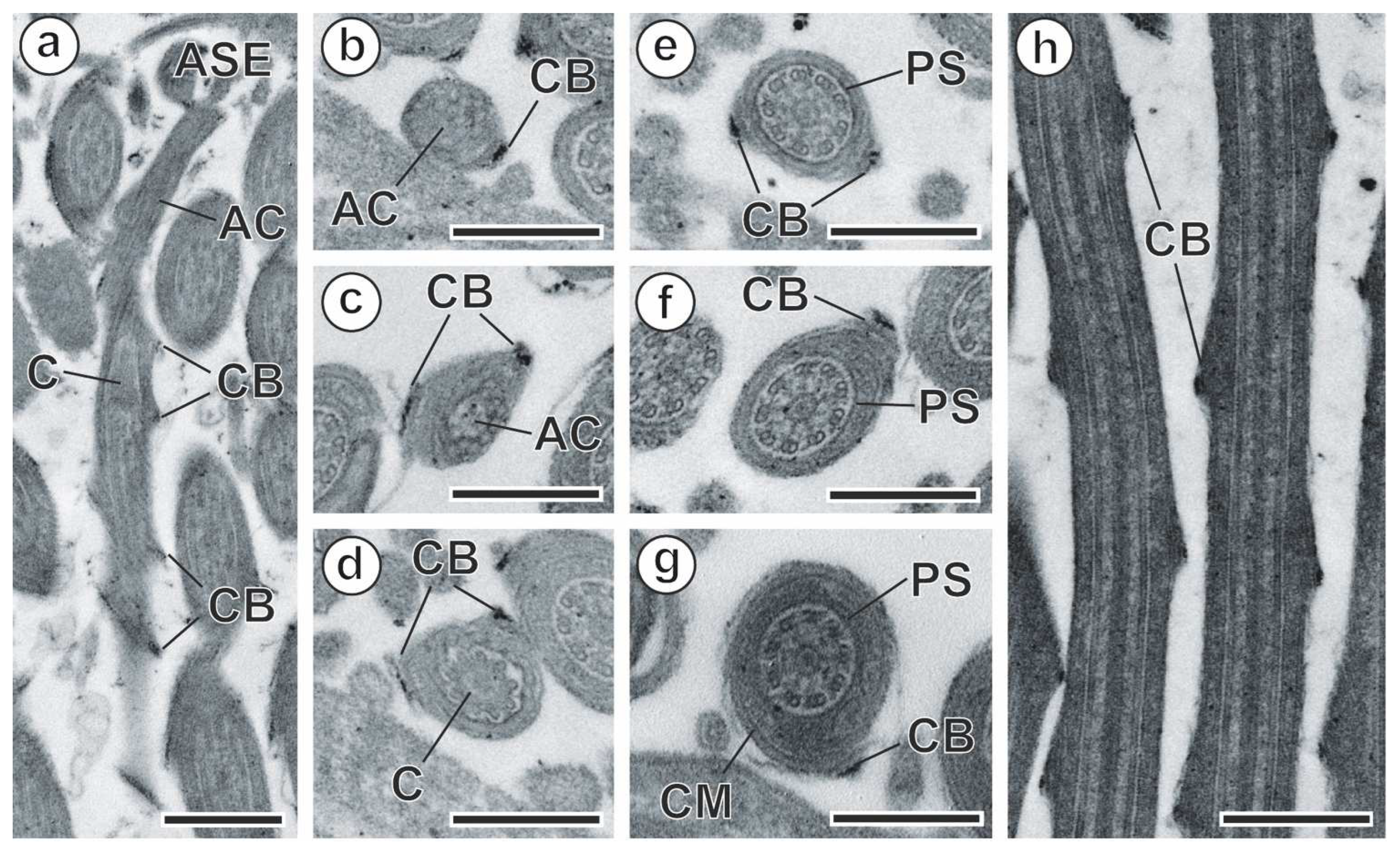
3.2. Spermatozoon
4. Discussion
4.1. Spermiogenesis
4.2. Spermatozoon
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haukisalmi, V.; Hardman, L.M.; Henttonen, H. Taxonomic review of cestodes of the genus Catenotaenia Janicki, 1904 in Eurasia and molecular phylogeny of the Catenotaeniidae (Cyclophyllidea). Zootaxa 2010, 2489, 1–33. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Ribas, A.; Junker, K.; Spickett, A.; Matthee, S.; Henttonen, H.; Jrijer, J.; Halajian, A.; Anders, J.L.; Nakao, M. Molecular systematics and evolutionary history of catenotaeniid cestodes (Cyclophyllidea). Zool. Scr. 2018, 47, 221–230. [Google Scholar] [CrossRef]
- Wertheim, G. A new anoplocephalid cestode from the gerbil. Parasitology 1954, 44, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, G.; Schmidt, G.D.; Greenberg, Z. Witenbergitaenia sinaica gen. n., sp. n. (Anoplocephalidae) and other Cestodes from small Mammals in Israel and in the Sinai Peninsula. Bull. Mus. Natn. Hist Nat. Paris 4e Sér. 1986, 8, 543–550. [Google Scholar] [CrossRef]
- Global Cestode Database. Available online: https://tapewormdb.uconn.edu (accessed on 15 September 2023).
- Lynsdale, J.A. On a remarkable new cestode, Meggittina baeri gen. et sp. nov. (Anoplocephalinae) from Rodents in Southern Rhodesia. J. Helminthol. 1953, 27, 129–142. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Ogunsuri, R.A.; Nwude, N.; Aliu, Y. Helminths of the African giant rat (Cricetomys gambianus Waterhouse) in Zaria, Nigeria. Rev. Elev. Méd. vét. Pays trop. 1984, 37, 304–307. [Google Scholar]
- Jrijer, J.; Neifar, L. Meggittina numida n. sp. (Cyclophyllidea: Catenotaeniidae), a parasite of the Shaw’s jird Meriones shawi (Duvernoy) (Rodentia: Gerbillinae) in Tunisia. Syst. Parasitol. 2014, 88, 167–174. [Google Scholar] [CrossRef]
- Wolfgang, R.W. Helminth parasites of reptiles, birds, and mammals in Egypt. II. Catenotaenia aegyptica sp. nov. from myomorph rodents, with additional notes on the genus. Can. J. Zool. 1956, 34, 6–20. [Google Scholar]
- Jrijer, J.; Bordes, F.; Morand, S.; Neifar, L. Gastro-intestinal helminth parasites of the common North African gerbil Meriones shawi (Duvernoy) in Tunisia: Parasites diversity and habitat anthropization effect. Trop. Biomed. 2016, 33, 702–710. [Google Scholar]
- Hockley, A.R. On Skrjabinotaenia cricetomydis n. sp. (Cestoda, Anoplocephalata) from the gambian pouched rat, Nigeria. J. Helminthol. 1961, 35, 233–254. [Google Scholar] [CrossRef]
- Jones, A. Some catenotaeniids from African rodents. Parasitology 1984, 89, XXI. [Google Scholar]
- Khemiri, H.; Jrijer, J.; Neifar, L.; Nouira, S. A survey study on the helminth parasites of two wild jirds, Meriones shawi and M. libycus (Rodentia: Gerbillinae), in Tunisian desert areas. Eur. Zool. J. 2017, 84, 303–310. [Google Scholar] [CrossRef]
- Tenora, F.; Mas-Coma, S.; Murai, E.; Feliu, C. The System of Cestodes of the Suborder Catenotaeniata Spassky, 1963. Parasit. Hung. 1980, 13, 39–57. [Google Scholar]
- Euzet, L.; Świderski, Z.; Mokhtar-Maamouri, F. Ultrastructure comparée du spermatozoïde des Cestodes. Relations avec la phylogénèse. Ann. Parasitol. 1981, 56, 247–259. [Google Scholar] [CrossRef]
- Świderski, Z. Three types of spermiogenesis in Cestodes. In Proceedings of the 11th International Congress on Electron Microscopy, Kyoto, Japan, 31 August–7 September 1986; pp. 2959–2960. [Google Scholar]
- Justine, J.-L. Phylogeny of parasitic Platyhelminthes: A critical study of synapomorphies proposed on the basis of the ultrastructure of spermiogenesis and spermatozoa. Can. J. Zool. 1991, 69, 1421–1440. [Google Scholar] [CrossRef]
- Justine, J.-L. Cladistic study in the Monogenea (Platyhelminthes), based upon a parsimony analysis of spermiogenetic and spermatozoal ultrastructural characters. Int. J. Parasitol. 1991, 21, 821–838. [Google Scholar] [CrossRef]
- Justine, J.-L. Spermatozoal ultrastructure and phylogeny of the parasitic Platyhelminthes. Mém. Mus. Natn. Hist. Nat. 1995, 166, 55–86. [Google Scholar]
- Bâ, C.T.; Marchand, B. Spermiogenesis, spermatozoa and phyletic affinities in the Cestoda. Mém. Mus. Natn. Hist. Nat. 1995, 166, 87–95. [Google Scholar]
- Justine, J.-L. Spermatozoa as phylogenetic characters for the Eucestoda. J. Parasitol. 1998, 84, 385–408. [Google Scholar] [CrossRef]
- Justine, J.-L. Spermatozoa as phylogenetic characters for the Platyhelminthes. In Interrelationships of Platyhelminthes; Littlewood, D.T.J., Bray, R.A., Eds.; Taylor & Francis: London, UK, 2001; pp. 231–238. [Google Scholar]
- Levron, C.; Miquel, J.; Oros, M.; Scholz, T. Spermatozoa of tapeworms (Platyhelminthes, Eucestoda): Advances in ultrastructural and phylogenetic studies. Biol. Rev. 2010, 85, 523–543. [Google Scholar] [CrossRef]
- Bakhoum, A.J.S.; Miquel, J.; Ndiaye, P.I.; Justine, J.-L.; Falchi, A.; Bâ, C.T.; Marchand, B.; Quilichini, Y. Advances in spermatological characters in the Digenea: Review and proposal of spermatozoa models and their phylogenetic importance. Adv. Parasitol. 2017, 98, 111–165. [Google Scholar] [PubMed]
- Justine, J.-L.; Poddubnaya, L.G. Spermiogenesis and spermatozoon ultrastructure in basal polyopisthocotylean monogeneans, Hexabothriidae and Chimaericolidae, and their significance for the phylogeny of the Monogenea. Parasite 2018, 25, 7. [Google Scholar] [CrossRef] [PubMed]
- Bâ, A.; Bâ, C.T.; Quilichini, Y.; Dieng, T.; Marchand, B. Ultrastructure of the spermatozoon of Taeniarhynchus saginatus (syn. Taenia saginata) (Goeze, 1782) Weinland, 1858 (Cestoda, Taeniidae) an intestinal parasite of human. Parasitol. Res. 2011, 108, 831–836. [Google Scholar] [PubMed]
- Taeleb, A.A.; Gamil, I.S. Spermatozoon ultrastructure of the davaineid cestode Raillietina (R.) vinagoi Ortlepp, 1938 from Egypt. Acta Parasitol. Glob. 2011, 2, 11–19. [Google Scholar]
- Yoneva, A.; Levron, C.; Nikolov, P.N.; Mizinska, Y.; Mariaux, J.; Georgiev, B.B. Spermiogenesis and spermatozoon ultrastructure of the paruterinid cestode Notopentorchis sp. (Cyclophyllidea). Parasitol. Res. 2012, 111, 135–142. [Google Scholar] [CrossRef]
- Bâ, A.; Ndiaye, P.I.; Bâ, C.T.; Miquel, J. Ultrastructure of the spermatozoon of Anomotaenia quelea (Mettrick, 1961) (Cestoda, Cyclophyllidea, Dilepididae), an intestinal parasite of Quelea quelea (Aves, Ploceidae) in Senegal. Zool. Anz. 2013, 253, 119–125. [Google Scholar] [CrossRef]
- Miquel, J.; Khallaayoune, K.; Azzouz-Maache, S.; Pétavy, A.-F. Spermatological characteristics of the genus Taenia inferred from the ultrastructural study on Taenia hydatigena. Parasitol. Res. 2015, 114, 201–208. [Google Scholar] [CrossRef]
- Miquel, J.; Świderski, Z.; Azzouz-Maache, S.; Pétavy, A.-F. Echinococcus multilocularis Leuckart, 1863 (Taeniidae): New data on sperm ultrastructure. Parasitol. Res. 2016, 115, 2269–2275. [Google Scholar] [CrossRef]
- Miquel, J.; Świderski, Z.; Feliu, C. Spermatozoon ultrastructure of Thysanotaenia congolensis (Cyclophyllidea, Anoplocephalidae, Inermicapsiferinae): Phylogenetic implications. Parasitol. Res. 2016, 115, 3083–3091. [Google Scholar] [CrossRef]
- Hidalgo, C.; Miquel, J.; Torres, J.; Marchand, B. Ultrastructural study of spermiogenesis and the spermatozoon in Catenotaenia pusilla, an intestinal parasite of Mus musculus. J. Helminthol. 2000, 74, 73–81. [Google Scholar] [CrossRef]
- Miquel, J.; Bâ, C.T.; Marchand, B. Ultrastructure of the spermatozoon of Skrjabinotaenia lobata (Cyclophyllidea, Catenotaeniidae), intestinal parasite of Apodemus sylvaticus (Rodentia, Muridae). J. Submicr. Cytol. Pathol. 1997, 29, 521–526. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high pH as an electronopaque stain in electronmicroscopy. J. Cell. Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Yoneva, A.; Georgieva, K.; Mizinska, Y.; Nikolov, P.N.; Georgiev, B.B.; Stoitsova, S.R. Ultrastructure of spermiogenesis and mature spermatozoon of Anonchotaenia globata (von Linstow, 1879) (Cestoda, Cyclophyllidea, Paruterinidae). Acta Zool. 2010, 91, 184–192. [Google Scholar] [CrossRef]
- Yoneva, A.; Georgieva, K.; Nikolov, P.N.; Mizinska, Y.; Georgiev, B.B.; Stoitsova, S.R. Ultrastructure of spermiogenesis and mature spermatozoon of Triaenorhina rectangula (Cestoda: Cyclophyllidea: Paruterinidae). Folia Parasitol. 2009, 56, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, P.I.; Miquel, J.; Marchand, B. Ultrastructure of spermiogenesis and spermatozoa of Taenia parva Baer, 1926 (Cestoda, Cyclophyllidea, Taeniidae), a parasite of the common genet (Genetta genetta). Parasitol. Res. 2003, 89, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Miquel, J.; Świderski, Z.; Foronda, P.; Torres, J.; Feliu, C. Ultrastructure of spermatogenesis of Taenia taeniaeformis (Batsch, 1786) (Cestoda, Cyclophyllidea, Taeniidae) and comparison of spermatological characters in the family Taeniidae Ludwig, 1886. Acta Parasitol. 2009, 54, 230–243. [Google Scholar] [CrossRef]
- Yoneva, A.; Georgieva, K.; Mizinska, Y.; Georgiev, B.B.; Stoitsova, S.R. Ultrastructure of spermiogenesis and mature spermatozoon of Skrjabinoporus merops (Cyclophyllidea, Metadilepididae). Acta Parasitol. 2006, 51, 200–208. [Google Scholar] [CrossRef]
- Ndiaye, P.I.; Agostini, S.; Miquel, J.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon in the genus Joyeuxiella Fuhrmann, 1935 (Cestoda, Cyclophyllidea, Dipylidiidae): Comparative analysis of J. echinorhynchoides (Sonsino, 1889) and J. pasqualei (Diamare, 1893). Parasitol. Res. 2003, 91, 175–186. [Google Scholar] [CrossRef]
- Miquel, J.; Bâ, C.T.; Marchand, B. Ultrastructure of spermiogenesis of Dipylidium caninum (Cestoda, Cyclophyllidea, Dipylidiidae), an intestinal parasite of Canis familiaris. Int. J. Parasitol. 1998, 28, 1453–1458. [Google Scholar] [CrossRef]
- Miquel, J.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Anoplocephaloides dentata (Cestoda, Cyclophyllidea, Anoplocephalidae), an intestinal parasite of Arvicolidae rodents. J. Parasitol. 1998, 84, 1128–1136. [Google Scholar] [CrossRef]
- Miquel, J.; Świderski, Z.; Młocicki, D.; Eira, C.; Marchand, B. Spermatogenesis in the anoplocephalid cestode Gallegoides arfaai (Mobedi et Ghadirian, 1977) Tenora et Mas-Coma, 1978. Acta Parasitol. 2005, 50, 132–144. [Google Scholar]
- Eira, C.; Miquel, J.; Vingada, J.; Torres, J. Spermiogenesis and spermatozoon ultrastructure of the cestode Mosgovoyia ctenoides (Cyclophyllidea: Anoplocephalidae), an intestinal parasite of Oryctolagus cuniculus (Lagomorpha: Leporidae). J. Parasitol. 2006, 92, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Bâ, A.; Quilichini, Y.; Ndiaye, P.I.; Bâ, C.T.; Marchand, B. Ultrastructure of the spermatozoon of Bothriocotyle sp. (Cestoda: Bothriocephalidea), a parasite of Schedophilus velaini (Sauvage, 1879) (Perciformes: Centrolophidae) in Senegal. J. Parasitol. 2012, 98, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Bruňanská, M.; Poddubnaya, L.G. Spermatological characters of the spathebothriidean tapeworm Didymobothrium rudolphii (Monticelli, 1890). Parasitol. Res. 2010, 106, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Bruňanská, M.; Fagerholm, H.-P.; Nebesářová, J.; Kostič, B. Ultrastructure of the mature spermatozoon of Eubothrium rugosum (Batsch, 1786) with a re-assessment of the spermatozoon ultrastructure of Eubothrium crassum (Bloch, 1779) (Cestoda: Bothriocephalidea). Helminthologia 2010, 47, 257–263. [Google Scholar] [CrossRef]
- Bruňanská, M.; Matey, V.; Nebesářová, J. Ultrastructure of the spermatozoon of the diphyllobothriidean cestode Cephalochlamys namaquensis (Cohn, 1906). Parasitol. Res. 2012, 111, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Bruňanská, M.; Matoušková, M.; Jasinská, R.; Nebesářová, J.; Poddubnaya, L.G. Heteromorphism of sperm axonemes in a parasitic flatworm, progenetic Diplocotyle olrikii Krabbe, 1874 (Cestoda, Spathebothriidea). Parasitol. Res. 2020, 119, 177–187. [Google Scholar] [CrossRef]
- Levron, C.; Yoneva, A.; Kalbe, M. Spermatological characters in the diphyllobothriidean Schistocephalus solidus (Cestoda). Acta Zool. 2013, 94, 240–247. [Google Scholar] [CrossRef]
- Marigo, A.M.; Eira, C.; Bâ, C.T.; Miquel, J. Spermatological characters of Acanthobothrium crassicolle Wedl, 1855 (Tetraphyllidea, Onchobothriidae), a parasite of the common stingray Dasyatis pastinaca. Acta Parasitol. 2011, 56, 191–201. [Google Scholar] [CrossRef]
- Marigo, A.M.; Świderski, Z.; Bâ, C.T.; Miquel, J. Spermiogenesis and ultrastructure of the spermatozoon of the trypanorhynch cestode Aporhynchus menezesi (Aporhynchidae), a parasite of the velvet belly lanternshark Etmopterus spinax (Elasmobranchii: Etmopteridae). Folia Parasitol. 2011, 58, 69–78. [Google Scholar] [CrossRef]
- Marigo, A.M.; Levron, C.; Bâ, C.T.; Miquel, J. Ultrastructural study of spermiogenesis and the spermatozoon of the proteocephalidean cestode Barsonella lafoni de Chambrier et al., 2009, a parasite of the catfish Clarias gariepinus (Burchell, 1822) (Siluriformes, Clariidae). Zool. Anz. 2012, 251, 147–159. [Google Scholar] [CrossRef]
- Marigo, A.M.; Delgado, E.; Torres, J.; Bâ, C.T.; Miquel, J. Spermiogenesis and spermatozoon ultrastructure of the bothriocephalidean cestode Clestobothrium crassiceps (Rudolphi, 1819), a parasite of the teleost fish Merluccius merluccius (Gadiformes: Merlucciidae). Parasitol. Res. 2012, 110, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Miquel, J.; Świderski, Z. Spermatological characteristics of the Trypanorhyncha inferred from new ultrastructural data on species of Tentaculariidae, Eutetrarhynchidae, and Progrillotiidae. C. R. Biol. 2013, 336, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, P.I.; Quilichini, Y.; Bâ, A.; Bâ, C.T.; Marchand, B. Ultrastructural study of the male gamete of Glossobothrium sp. (Cestoda: Bothriocephalidea: Triaenophoridae) a parasite of Schedophilus velaini (Perciformes: Centrolophidae) in Senegal. Tissue Cell 2012, 44, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Šípková, L.; Levron, C.; Freeman, M.; Scholz, T. Spermiogenesis and spermatozoon of the tapeworm Parabothriocephalus gracilis (Bothriocephalidea): Ultrastructural and cytochemical studies. Acta Parasitol. 2010, 55, 58–65. [Google Scholar] [CrossRef]
- Šípková, L.; Levron, C.; Oros, M.; Justine, J.-L. Spermatological characters of bothriocephalideans (Cestoda) inferred from an ultrastructural study on Oncodiscus sauridae and Senga sp. Parasitol. Res. 2011, 108, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Yoneva, A.; Kuchta, R.; Scholz, T. Spermiogenesis and sperm ultrastructure of two species of Duthiersia, parasites of monitors, with a review of spermatological characters in the Diphyllobothriidea (Cestoda). Zool. Anz. 2013, 252, 486–494. [Google Scholar] [CrossRef]
- Cielocha, J.J.; Yoneva, A.; Cantino, M.E.; Daniels, S.; Jensen, K. Ultrastructure of the sperm of Adelobothrium sp. (Cestoda: Lecanicephalidea). Invertebr. Biol. 2013, 132, 315–325. [Google Scholar] [CrossRef]
- Cielocha, J.J.; Martinez, E.; Jackson, A.; Yoneva, A. Characterization of spermatozoon ultrastructure in Tetragonocephalum sp. (Cestoda: Lecanicephalidea: Tetragonocephalidae) from the whipray, Urogymnus asperrimus (Dasyatidae: Urogymninae). J. Parasitol. 2022, 108, 500–510. [Google Scholar] [CrossRef]
- Bruňanská, M.; Kostič, B. Revisiting caryophyllidean type of spermiogenesis in the Eucestoda based on spermatozoon differentiation and ultrastructure of Caryophyllaeus laticeps (Pallas, 1781). Parasitol. Res. 2012, 110, 141–149. [Google Scholar] [CrossRef]
- Bruňanská, M.; Bílý, T.; Nebesářová, J. Nippotaenia mogurndae Yamaguti et Myiata, 1940 (Cestoda, Nippotaeniidea): First data on spermiogenesis and sperm ultrastructure. Parasitol. Res. 2015, 114, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Bruňanská, M.; Matoušková, M.; Nebesářová, J.; Mackiewicz, J.S.; Poddubnaya, L.G. First ultrastructural and cytochemical data on the spermatozoon and its differentiation in progenetic and adult Archigetes sieboldi Leuckart, 1878 (Cestoda, Caryophyllidea, Caryophyllaeidae). Parasitol. Res. 2019, 118, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Matoušková, M.; Bílý, T.; Bruňanská, M.; Mackiewicz, J.S.; Nebesářová, J. Ultrastructure, cytochemistry and electron tomography analysis of Caryophyllaeides fennica (Schneider, 1902) (Cestoda: Lytocestidae) reveals novel spermatology characteristics in the Eucestoda. Parasitol. Res. 2018, 117, 3091–3102. [Google Scholar] [CrossRef] [PubMed]
- Matoušková, M.; Bílý, T.; Bruňanská, M.; Oros, M.; Kostič, B.; Nebesářová, J. New data on spermiogenesis and trepaxonematan axoneme in basal tapeworms (Cestoda, Caryophyllidea, Lytocestidae) parasitizing cyprinid fishes. Sci. Rep. 2019, 9, 12881. [Google Scholar] [CrossRef] [PubMed]
- Matoušková, M.; Bruňanská, M.; Nebesářová, J.; Poddubnaya, L.G. Ultrastructure and cytochemistry of the mature spermatozoon of Khawia armeniaca (Cholodkovsky, 1915) (Caryophyllidea: Lytocestidae) a parasite of Capoeta capoeta sevangi (De Filippi, 1865) (Teleostei, Cyprinidae). Helminthologia 2020, 57, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Yoneva, A.; Levron, C.; Oros, M.; Orosová, M.; Scholz, T. Ultrastructure of spermiogenesis and mature spermatozoon of Breviscolex orientalis (Cestoda: Caryophyllidea). Parasitol. Res. 2011, 108, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Yoneva, A.; Levron, C.; Ash, A.; Scholz, T. Spermatological characters of monozoic tapeworms (Cestoda: Caryophyllidea), including first data on a species from the indomalayan catfish. J. Parasitol. 2012, 98, 423–430. [Google Scholar] [CrossRef]
- Yoneva, A.; Levron, C.; Oros, M.; Orosová, M.; Scholz, T. Spermiogenesis and spermatozoon ultrastructure of Hunterella nodulosa (Cestoda: Caryophyllidea), a monozoic parasite of suckers (Catostomidae) in North America. Folia Parasitol. 2012, 59, 179–186. [Google Scholar] [CrossRef]
- Marigo, A.M.; Bâ, C.T.; Miquel, J. Spermiogenesis and spermatozoon ultrastructure of the dilepidid cestode Molluscotaenia crassiscolex (von Linstow, 1890), an intestinal parasite of the common shrew Sorex araneus. Acta Zool. 2011, 92, 116–125. [Google Scholar] [CrossRef]
- Azzouz-Draoui, N.; Mokhtar-Maamouri, F. Ultrastructure comparée de la spermiogenèse et du spermatozoïde de Echinobothrium affine Diesing, 1863 et E. harfordi Mac Vicar, 1976 (Cestoda, Diphyllidea). Bull. Soc. Sci. Nat. Tunisie 1986, 18, 9–20. [Google Scholar]
- Marigo, A.M.; Eira, C.; Bâ, C.T.; Miquel, J. Spermiogenesis and spermatozoon ultrastructure of the diphyllidean cestode Echinobothrium euterpes (Neifar, Tyler and Euzet 2001) Tyler 2006, a parasite of the common guitarfish Rhinobatos rhinobatos. Parasitol. Res. 2011, 109, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Bâ, C.T.; Marchand, B. Ultrastructure of the spermatozoon of Sandonella sandoni (Cestoda, Proteocephalidea, Sandonellinae). Invertebr. Reprod. Dev. 1994, 25, 9–17. [Google Scholar] [CrossRef]
- Xylander, W.E.R. Ultrastructural studies on the reproductive system of Gyrocotylidea and Amphilinidea (Cestoda): Spermatogenesis, spermatozoa, testes and vas deferens of Gyrocotyle. Int. J. Parasitol. 1989, 19, 897–905. [Google Scholar] [CrossRef]
- Bruňanská, M.; Poddubnaya, L.G.; Xylander, W.E.R. Spermatozoon cytoarchitecture of Amphilina foliacea (Platyhelminthes, Amphilinidea). Parasitol. Res. 2012, 111, 2063–2069. [Google Scholar] [CrossRef] [PubMed]
- Bâ, C.T.; Marchand, B. Ultrastructural particularities of the spermatozoon of Stilesia globipunctata (Cestoda) parasite of the small intestine of sheep and goats in Senegal. J. Submicr. Cytol. Pathol. 1992, 24, 29–34. [Google Scholar]
- Bâ, C.T.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Mathevotaenia herpestis (Cestoda), intestinal parasite of Atelerix albiventrix in Sénégal. Acta Zool. 1994, 75, 167–175. [Google Scholar] [CrossRef]
- Miquel, J.; Marchand, B. Ultrastructure of the spermatozoon of Dipylidium caninum (Cestoda, Cyclophyllidea, Dilepididae), an intestinal parasite of Canis familiaris. Parasitol. Res. 1997, 83, 349–355. [Google Scholar] [CrossRef]
- Miquel, J.; Świderski, Z.; Marchand, B. Spermatological characters in the Dipylidiidae Stiles, 1896 (Cestoda, Cyclophyllidea). Acta Parasitol. 2005, 50, 65–73. [Google Scholar]
- Yoneva, A.; Miquel, J.; Świderski, Z.; Georgieva, K.; Mizinska, Y.; Georgiev, B.B. Ultrastructure of spermiogenesis and mature spermatozoon of Angularella beema (Clerc, 1906) (Cestoda, Cyclophyllidea, Dilepididae). Acta Parasitol. 2006, 51, 264–272. [Google Scholar] [CrossRef]
- Yoneva, A.; Świderski, Z.; Georgieva, K.; Nikolov, P.N.; Mizinska, Y.; Georgiev, B.B. Spermiogenesis and sperm ultrastructure of Valipora mutabilis Linton, 1927 (Cestoda, Cyclophyllidea, Gryporhynchidae). Parasitol. Res. 2008, 103, 1397–1405. [Google Scholar] [CrossRef]
- Bâ, C.T.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Vampirolepis microstoma (Cestoda, Hymenolepididae), intestinal parasite of Rattus rattus. Microsc. Res. Tech. 1998, 42, 218–225. [Google Scholar] [CrossRef]
- Sène, A.; Bâ, C.T.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Nomimoscolex sp. (Cestoda, Proteocephalidea) intestinal parasite of Clarotes laticeps (Fish, Teleost) in Senegal. J. Submicrosc. Cytol. Pathol. 1997, 29, 115–124. [Google Scholar] [PubMed]
- Bâ, C.T.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Aporina delafondi (Cyclophyllidea, Anoplocephalidae) intestinal parasite of turtle doves in Senegal. Int. J. Parasitol. 1994, 24, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Bâ, A.; Bâ, C.T.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Sudarikovina taterae (Cestoda, Cyclophyllidea, Anoplocephalidae) intestinal parasite of Tatera gambiana (Rodentia, Gerbillidae). J. Submicrosc. Cytol. Pathol. 2000, 32, 137–144. [Google Scholar]
- Miquel, J.; Ndiaye, P.I.; Feliu, C. Crest-like bodies in the spermatozoon of Hymenolepididae cestodes from Iberian rodents. Rev. Ibér. Parasitol. 2007, 67, 27–33. [Google Scholar]
- Miquel, J.; Feliu, C.; Marchand, B. Ultrastructure of spermiogenesis and the spermatozoon of Mesocestoides litteratus (Cestoda, Mesocestoididae). Int. J. Parasitol. 1999, 29, 499–510. [Google Scholar] [CrossRef]
- Miquel, J.; Eira, C.; Świderski, Z.; Conn, D.B. Mesocestoides lineatus (Goeze, 1782) (Mesocestoididae): New data on sperm ultrastructure. J. Parasitol. 2007, 93, 545–552. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily, | Spermiogenesis | Spermatozoon Characteristics | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species, and | Characteristics | CB | |||||||||||||
| Reference | Type 1 | PF | FR | SR | IB | Type 2 | n | T | L | AC | CM | N | DG | PS | IW |
| Catenotaeniinae | |||||||||||||||
| Catenotaenia pusilla [33] | III | + | + (~45°) | – | – | VI | 2 | 75 | ≠ | 1750 × 225 | 40° | Sp | – | + | – |
| Skrjabinotaeniinae | |||||||||||||||
| Meggittina gerbilli Present study | III | + | + (~90°) | – | – | VI | 2 | 70 | ≠ | >1150 | 30° | Sp | – | + | – |
| Skrjabinotaenia lobata [34] | ? | ? | ? | ? | ? | VI | 2 | 60–80 | ≠ | 2500 × 200 | 40° | Sp | – | + | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aouina, F.; Kacem, H.; Martín-Carrillo, N.; Foronda, P.; Miquel, J. Spermatological Characterization of the Cestode Meggittina gerbilli (Cyclophyllidea: Catenotaeniidae), a Parasite of Gerbils, Gerbillus gerbillus and Gerbillus campestris (Rodentia: Muridae) in Tunisia. Animals 2024, 14, 12. https://doi.org/10.3390/ani14010012
Aouina F, Kacem H, Martín-Carrillo N, Foronda P, Miquel J. Spermatological Characterization of the Cestode Meggittina gerbilli (Cyclophyllidea: Catenotaeniidae), a Parasite of Gerbils, Gerbillus gerbillus and Gerbillus campestris (Rodentia: Muridae) in Tunisia. Animals. 2024; 14(1):12. https://doi.org/10.3390/ani14010012
Chicago/Turabian StyleAouina, Faouzi, Hichem Kacem, Natalia Martín-Carrillo, Pilar Foronda, and Jordi Miquel. 2024. "Spermatological Characterization of the Cestode Meggittina gerbilli (Cyclophyllidea: Catenotaeniidae), a Parasite of Gerbils, Gerbillus gerbillus and Gerbillus campestris (Rodentia: Muridae) in Tunisia" Animals 14, no. 1: 12. https://doi.org/10.3390/ani14010012