Preliminary Evidence for the Role Played by South Westerly Wind Strength on the Marine Diatom Content of an Antarctic Peninsula Ice Core (1980–2010)

 , , , ,
, , , ,
Abstract
:1. Introduction
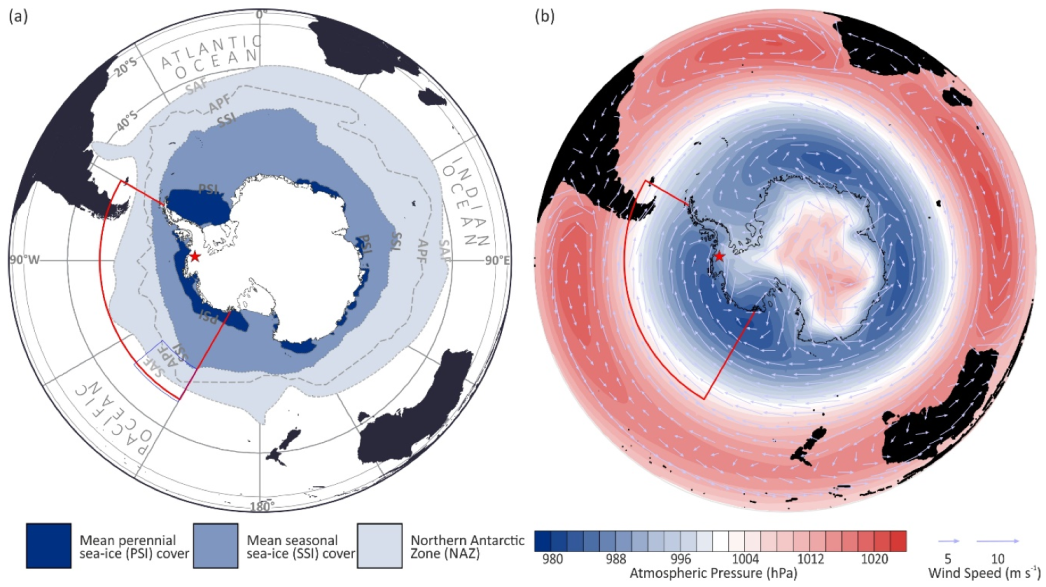
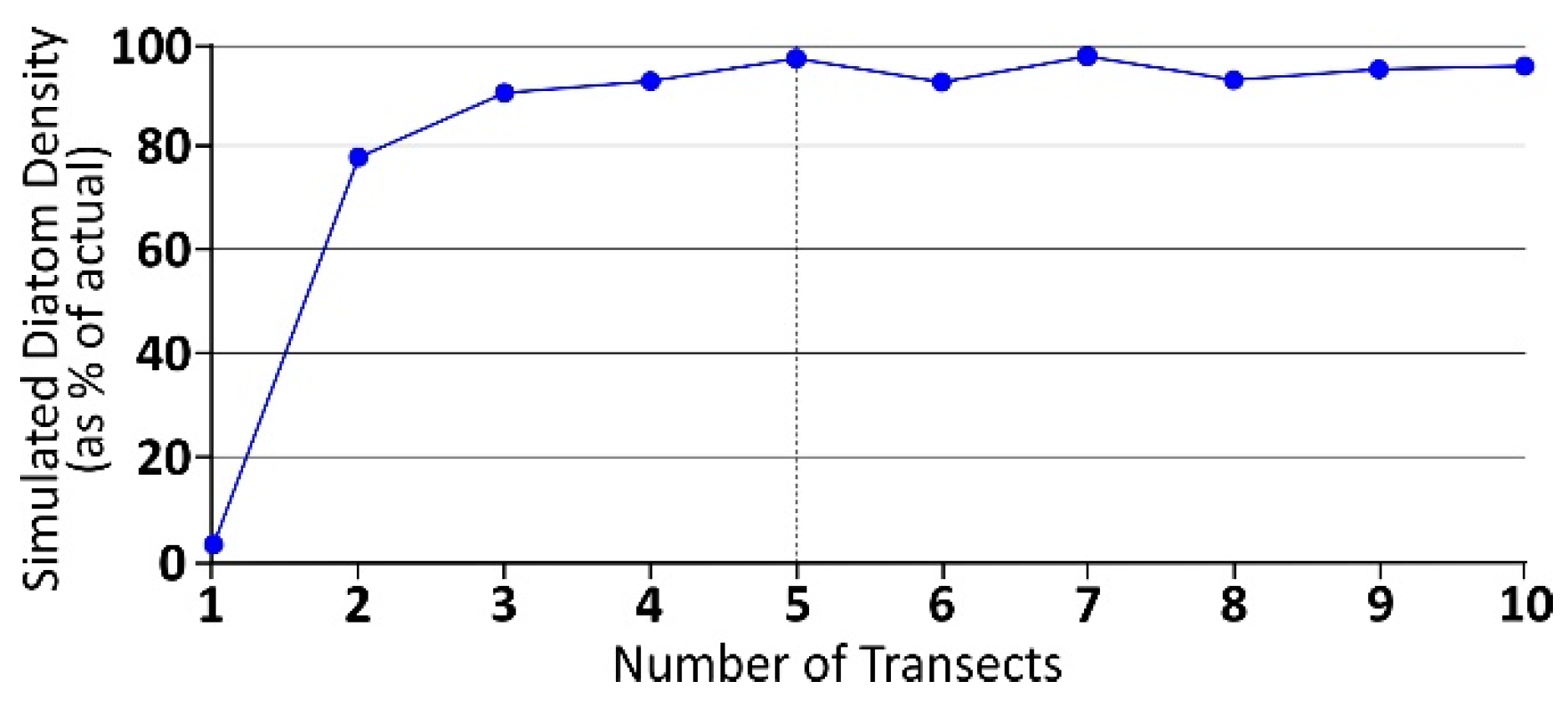
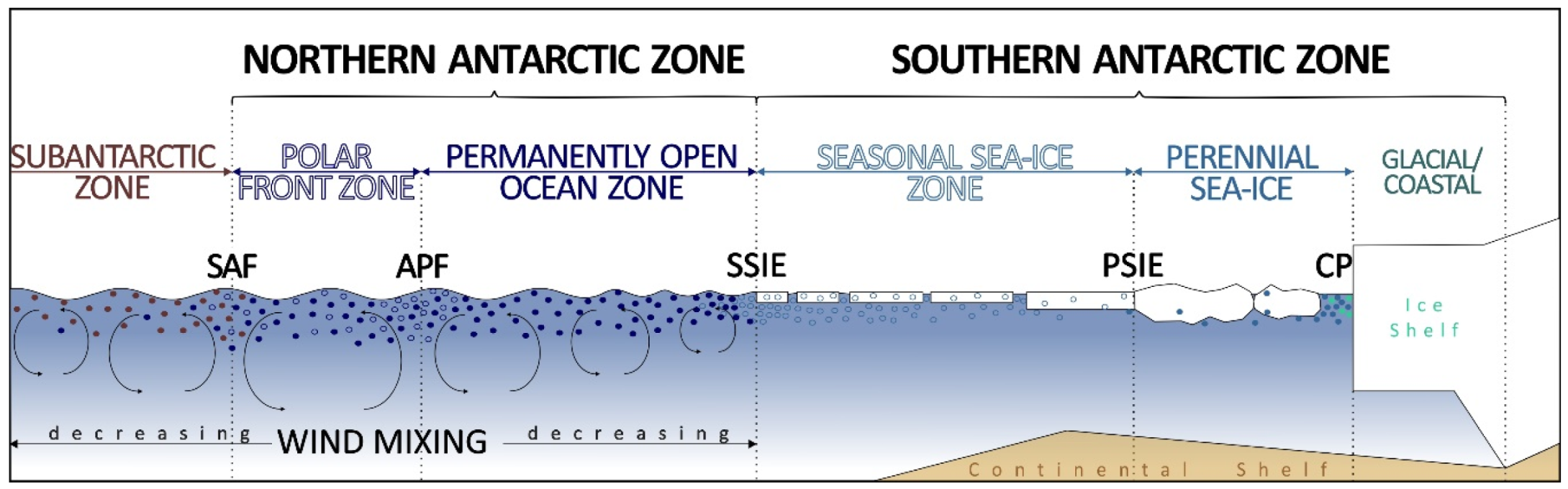
2. Materials and Methods
3. Results
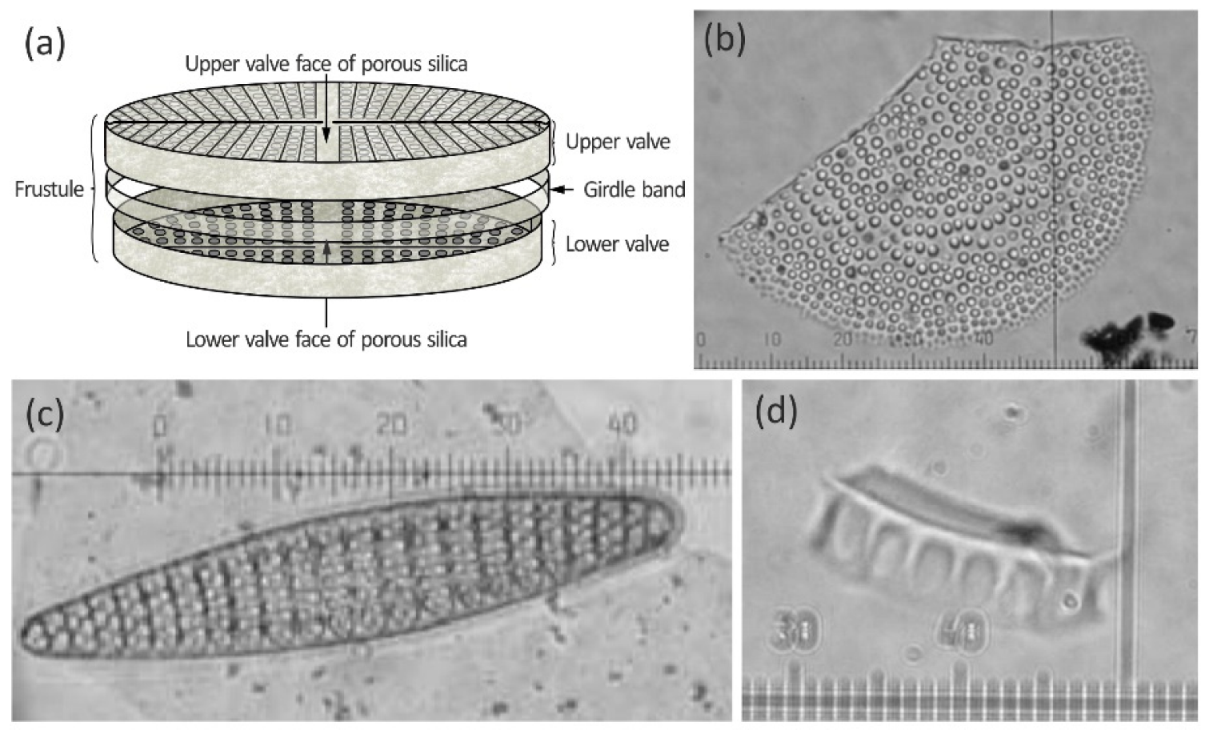
3.1. Diatom Assemblage Composition and Ecological Associations
3.1.1. Fragilariopsis kerguelensis
3.1.2. Thalassiosira lentiginosa
3.1.3. Thalassiothrix Group
3.1.4. Diatom Preservation
3.1.5. Summary of Ecological Affinities
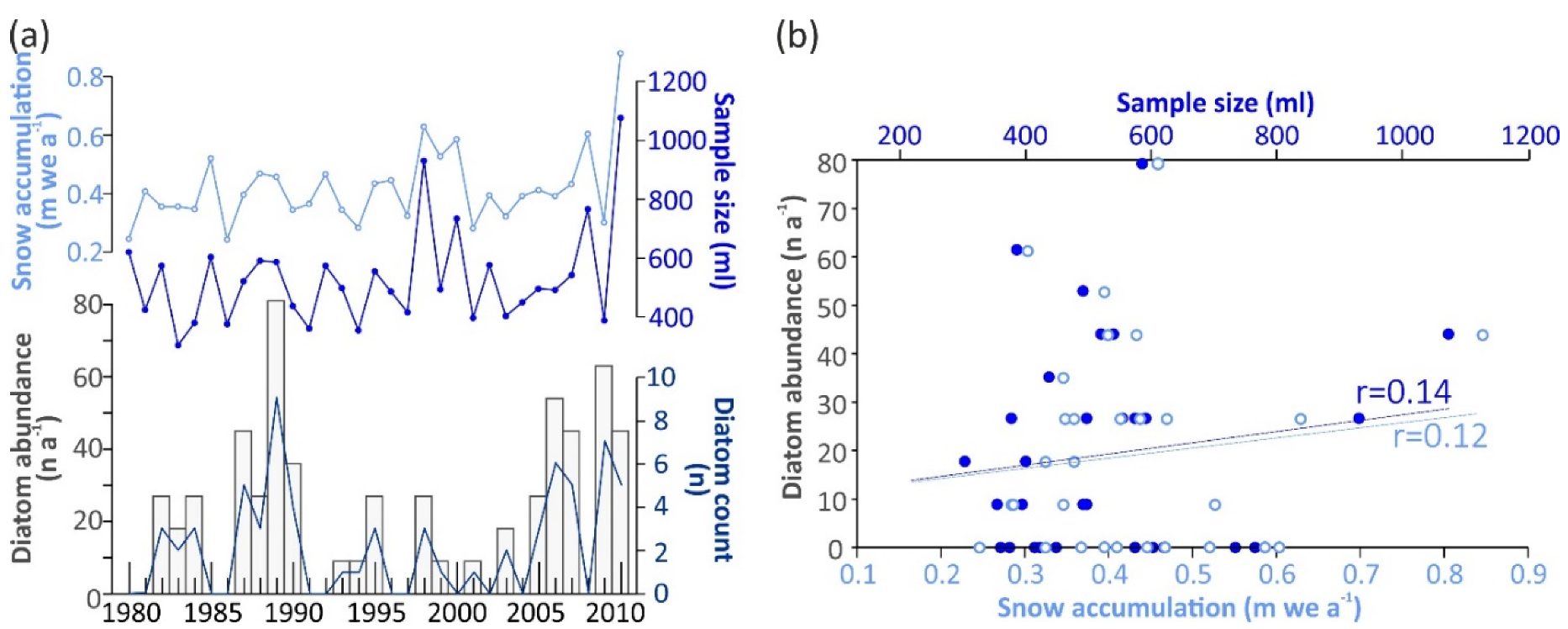
3.2. Diatom Abundance and Variability
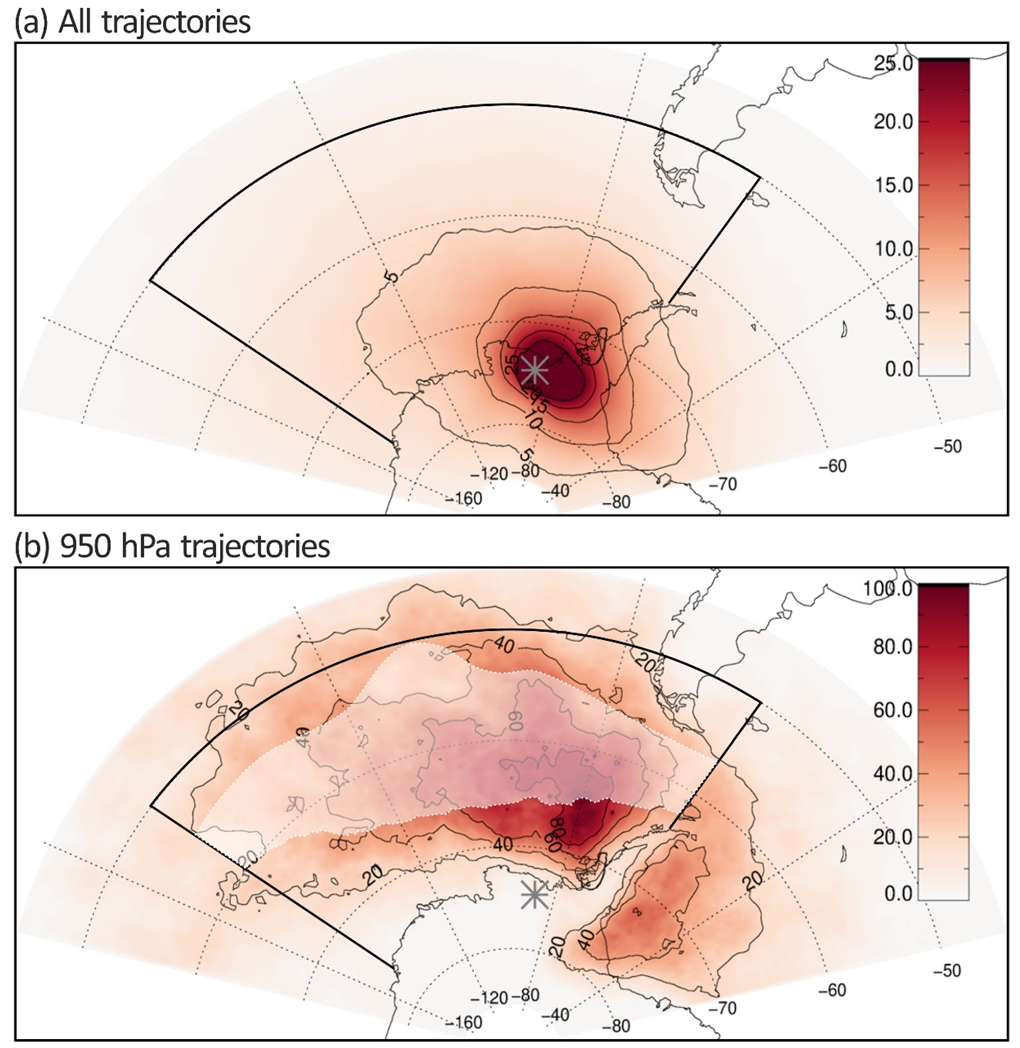
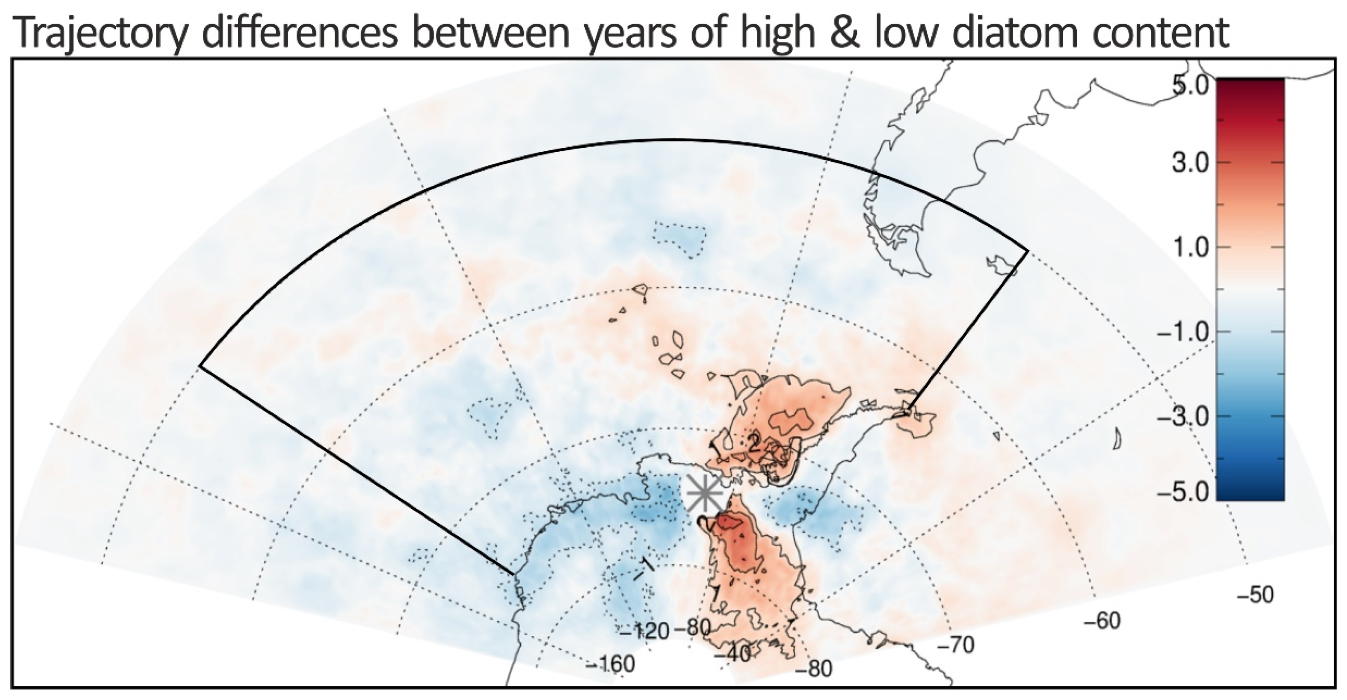
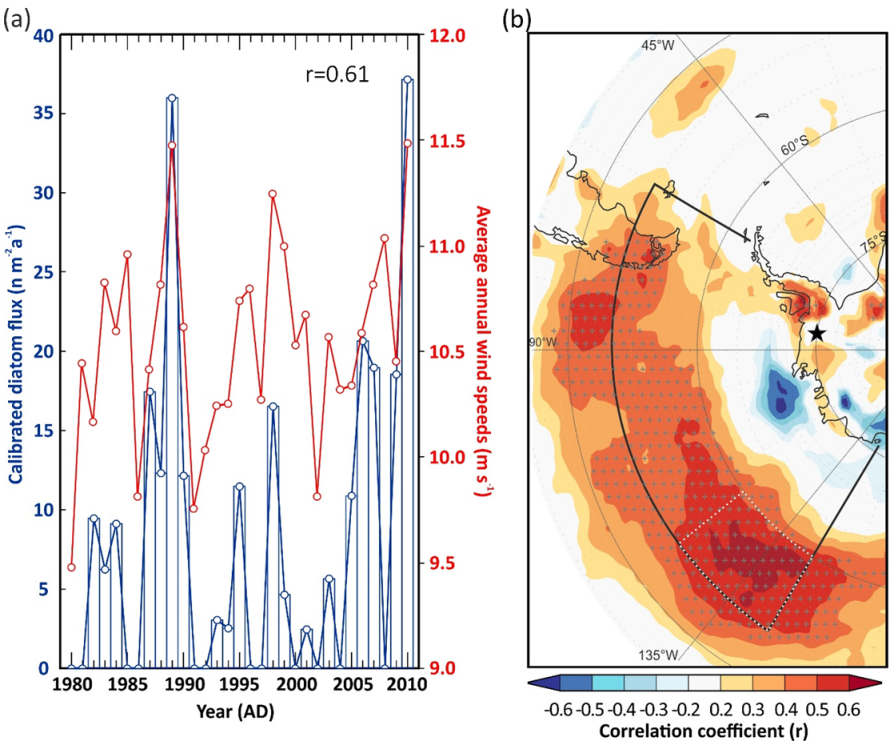
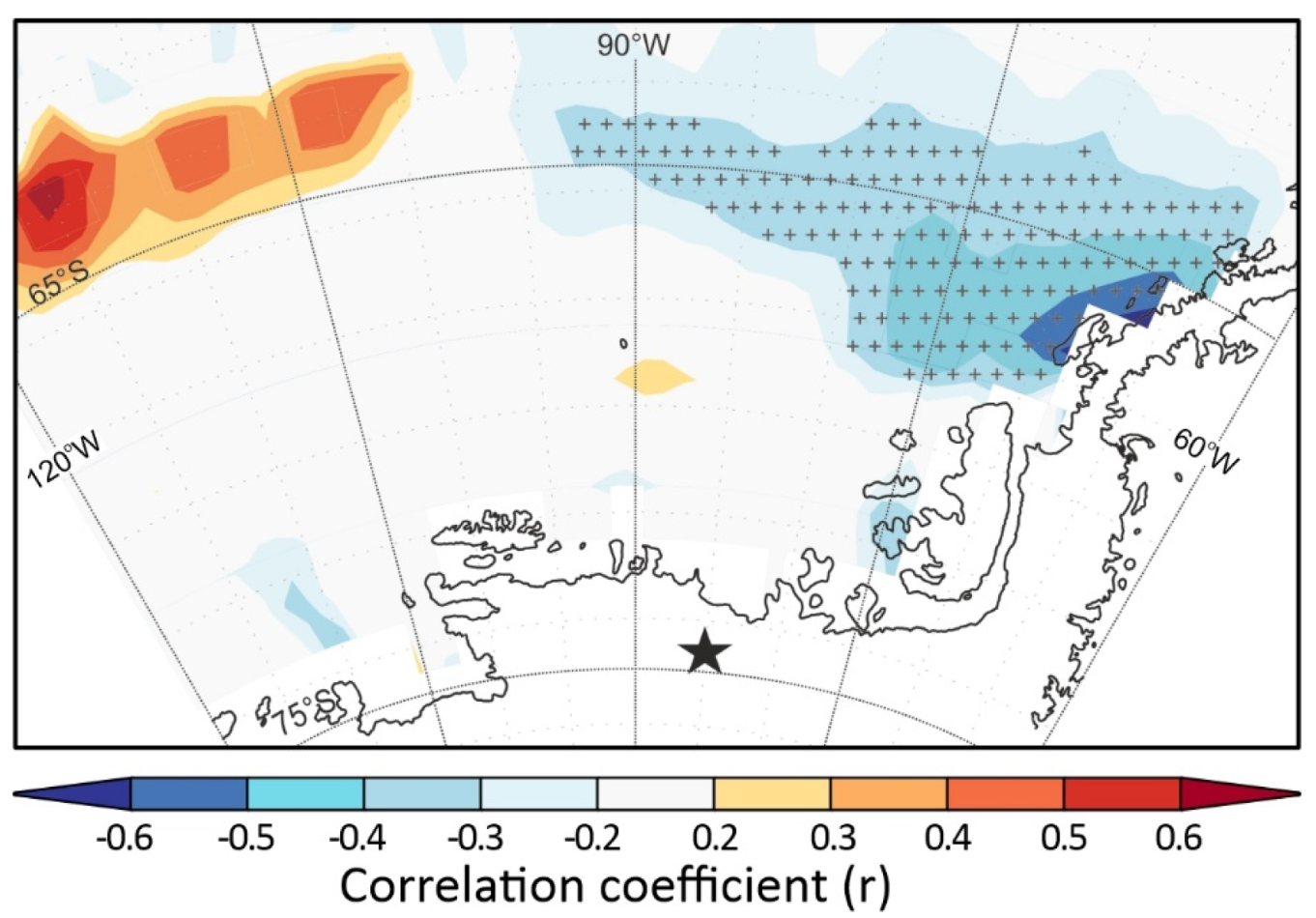
3.3. Air-Mass Circulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Marshall, G.J.; Orr, A.; Van Lipzig, N.P.M.; King, J.C. The Impact of a Changing Southern Hemisphere Annular Mode on Antarctic Peninsula Summer Temperatures. J. Clim. 2006, 19, 5388–5404. [Google Scholar] [CrossRef]
- Thomas, E.R.; Hosking, J.S.; Tuckwell, R.; Warren, R.A.; Ludlow, E.C. Twentieth century increase in snowfall in coastal West Antarctica. Geophys. Res. Lett. 2015, 42, 9387–9393. [Google Scholar] [CrossRef] [Green Version]
- Cook, A.J.; Holland, P.R.; Meredith, M.P.; Murray, T.; Luckman, A.; Vaughan, D. Ocean forcing of glacier retreat in the western Antarctic Peninsula. Science 2016, 353, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Spence, P.; Griffies, S.M.; England, M.; Hogg, A.M.; Saenko, O.A.; Jourdain, N.C. Rapid subsurface warming and circulation changes of Antarctic coastal waters by poleward shifting winds. Geophys. Res. Lett. 2014, 41, 4601–4610. [Google Scholar] [CrossRef] [Green Version]
- Rignot, E. Changes in West Antarctic ice stream dynamics observed with ALOS PALSAR data. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef] [Green Version]
- Rignot, E.; Mouginot, J.; Morlighem, M.; Seroussi, H.; Scheuchl, B. Widespread, rapid grounding line retreat of Pine Island, Thwaites, Smith, and Kohler glaciers, West Antarctica, from 1992 to 2011. Geophys. Res. Lett. 2014, 41, 3502–3509. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, H.D.; Arthern, R.J.; Vaughan, D.; Edwards, L.A. Extensive dynamic thinning on the margins of the Greenland and Antarctic ice sheets. Nature 2009, 461, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C.; Edwards, T.; Durand, G.; Payne, A.J.; Peyaud, V.; Hindmarsh, R. Potential sea-level rise from Antarctic ice-sheet instability constrained by observations. Nature 2015, 528, 115–118. [Google Scholar] [CrossRef]
- Dutrieux, P.; De Rydt, J.; Jenkins, A.; Holland, P.R.; Ha, H.K.; Lee, S.H.; Steig, E.J.; Ding, Q.; Abrahamsen, P.; Schröder, M. Strong Sensitivity of Pine Island Ice-Shelf Melting to Climatic Variability. Science 2014, 343, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, H.D.; Ligtenberg, S.R.M.; Fricker, H.A.; Vaughan, D.; Broeke, M.R.V.D.; Padman, L. Antarctic ice-sheet loss driven by basal melting of ice shelves. Nature 2012, 484, 502–505. [Google Scholar] [CrossRef]
- Bromwich, D.H.; Fogt, R. Strong Trends in the Skill of the ERA-40 and NCEP–NCAR Reanalyses in the High and Midlatitudes of the Southern Hemisphere, 1958–2001. J. Clim. 2004, 17, 4603–4619. [Google Scholar] [CrossRef]
- Raphael, M.N.; Marshall, G.J.; Turner, J.; Fogt, R.; Schneider, D.; Dixon, D.A.; Hosking, J.S.; Jones, J.; Hobbs, W.R. The Amundsen Sea Low: Variability, Change, and Impact on Antarctic Climate. Bull. Am. Meteorol. Soc. 2016, 97, 111–121. [Google Scholar] [CrossRef]
- Alley, R.B. Reliability of ice-core science: Historical insights. J. Glaciol. 2010, 56, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Legrand, M.; Mayewski, P. Glaciochemistry of polar ice cores: A review. Rev. Geophys. 1997, 35, 219–243. [Google Scholar] [CrossRef] [Green Version]
- Delmonte, B.; Paleari, C.I.; Ando’, S.; Garzanti, E.; Andersson, P.S.; Petit, J.R.; Crosta, X.; Narcisi, B.; Baroni, C.; Salvatore, M.C.; et al. Causes of dust size variability in central East Antarctica (Dome B): Atmospheric transport from expanded South American sources during Marine Isotope Stage 2. Quat. Sci. Rev. 2017, 168, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Dixon, D.; Mayewski, P.A.; Goodwin, I.; Marshall, G.J.; Freeman, R.; Maasch, K.A.; Sneed, S.B. An ice-core proxy for northerly air mass incursions into West Antarctica. Int. J. Clim. 2011, 32, 1455–1465. [Google Scholar] [CrossRef] [Green Version]
- Bullard, J.; Baddock, M.; Bradwell, T.; Crusius, J.; Darlington, E.; Gaiero, D.; Gassó, S.; Gisladottir, G.; Hodgkins, R.; McCulloch, R.; et al. High-latitude dust in the Earth system. Rev. Geophys. 2016, 54, 447–485. [Google Scholar] [CrossRef] [Green Version]
- Lambert, F.; Delmonte, B.; Petit, J.-R.; Bigler, M.; Kaufmann, P.R.; Hutterli, M.A.; Stocker, T.F.; Rüth, U.; Steffensen, J.P.; Maggi, V. Dust-climate couplings over the past 800,000 years from the EPICA Dome C ice core. Nature 2008, 452, 616–619. [Google Scholar] [CrossRef] [Green Version]
- Wolff, E.; Barbante, C.; Becagli, S.; Bigler, M.; Boutron, C.; Castellano, E.; De Angelis, M.; Federer, U.; Fischer, H.; Fundel, F.; et al. Changes in environment over the last 800,000 years from chemical analysis of the EPICA Dome C ice core. Quat. Sci. Rev. 2010, 29, 285–295. [Google Scholar] [CrossRef]
- Armand, L.; Crosta, X.; Romero, O.; Pichon, J.-J. The Biogeography of Major Diatom Taxa in Southern Ocean Sediments: 1. Sea Ice Related Species. Palaeogeogr. Palaeoclim. Palaeoecol. 2005, 223, 93–126. [Google Scholar] [CrossRef]
- Crosta, X.; Romero, O.; Armand, L.; Pichon, J.-J. The biogeography of major diatom taxa in Southern Ocean sediments: 2. Open ocean related species. Palaeogeogr. Palaeoclim. Palaeoecol. 2005, 223, 66–92. [Google Scholar] [CrossRef]
- Armand, L.; Crosta, X.; Romero, O.; Pichon, J.-J. The Biogeography of Major Diatom Taxa in Southern Ocean Surface Sediments: 3. Tropical/Subtropical Species. Palaeogeogr. Palaeoclim. Palaeoecol. 2005, 223, 49–65. [Google Scholar] [CrossRef]
- Zielinski, U.; Gersonde, R. Diatom distribution in Southern Ocean surface sediments (Atlantic sector): Implications for paleoenvironmental reconstructions. Palaeogeogr. Palaeoclim. Palaeoecol. 1997, 129, 213–250. [Google Scholar] [CrossRef]
- Spaulding, S.A.; McKnight, D. Diatoms as indicators of environmental change in antarctic freshwaters. In The Diatoms: Applications for the Environmental and Earth Sciences; Smol, J.P., Stoermer, E.F., Eds.; Cambridge University Press: New York, NY, USA, 2010; pp. 267–283. [Google Scholar] [CrossRef]
- Crosta, X.; Koç, N. Diatoms: From Micropaleontology to Isotope Geochemistry. In Developments in Marine Geology; Hilaire-Marcel, C., de Vernal, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Lowe, R.; Stoermer, E.F.; Smol, J.P. (Eds.) The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Romero, O.E.; Dupont, L.; Wyputta, U.; Jahns, S.; Wefer, G. Temporal variability of fluxes of eolian-transported freshwater diatoms, phytoliths, and pollen grains off Cape Blanc as reflection of land-atmosphere-ocean interactions in northwest Africa. J. Geophys. Res. Space Phys. 2003, 108. [Google Scholar] [CrossRef]
- Welch, H.E.; Muir, D.C.G.; Billeck, B.N.; Lockhart, W.L.; Brunskill, G.J.; Kling, H.J.; Olson, M.P.; Lemoine, R.M. Brown snow: A long-range transport event in the Canadian Arctic. Environ. Sci. Technol. 1991, 25, 280–286. [Google Scholar] [CrossRef]
- Budgeon, A.; Roberts, D.; Gasparon, M.; Adams, N. Direct evidence of aeolian deposition of marine diatoms to an ice sheet. Antarct. Sci. 2012, 24, 527–535. [Google Scholar] [CrossRef]
- Fritz, S.C.; Brinson, B.E.; Billups, W.E.; Thompson, L.G. Diatoms at >5000 Meters in the Quelccaya Summit Dome Glacier, Peru. Arctic. Antarct. Alp. Res. 2015, 47, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Papina, T.S.; Blyakharchuk, T.; Eichler, A.; Malygina, N.; Mitrofanova, E.; Schwikowski, M. Biological proxies recorded in a Belukha ice core, Russian Altai. Clim. Past 2013, 9, 2399–2411. [Google Scholar] [CrossRef] [Green Version]
- Tormo, R.; Recio, D.; Silva, I.; Muñoz, A. A quantitative investigation of airborne algae and lichen soredia obtained from pollen traps in south-west Spain. Eur. J. Phycol. 2001, 36, 385–390. [Google Scholar] [CrossRef] [Green Version]
- McKay, R.; Barrett, P.; Harper, M.; Hannah, M. Atmospheric transport and concentration of diatoms in surficial and glacial sediments of the Allan Hills, Transantarctic Mountains. Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 260, 168–183. [Google Scholar] [CrossRef]
- Ram, M.; Gayley, R.I. Insoluble particles in polar ice: Identification and measurement of the insoluble background aerosol. Geophys. Res. Lett. 1994, 21, 437–440. [Google Scholar] [CrossRef]
- Delmonte, B.; Baroni, C.; Andersson, P.S.; Narcisi, B.; Salvatore, M.; Petit, J.-R.; Scarchilli, C.; Frezzotti, M.; Albani, S.; Maggi, V. Modern and Holocene aeolian dust variability from Talos Dome (Northern Victoria Land) to the interior of the Antarctic ice sheet. Quat. Sci. Rev. 2013, 64, 76–89. [Google Scholar] [CrossRef]
- Burckle, L.H.; Kellogg, D.E.; Kellogg, T.B.; Fastook, J.L. A mechanism for emplacement and concentration of diatoms in glacigenic deposits. Boreas 2008, 26, 55–60. [Google Scholar] [CrossRef]
- Scherer, R.P. Pleistocene Collapse of the West Antarctic Ice Sheet. Science 1998, 281, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Stroeven, A.P.; Prentice, M.J.; Kleman, J. Recycled Marine Microfossils in Glacial Tills of the Sirius Group at Mount Fleming: Transport Mechanisms and Pathways. Antarct. J. U. S. 1997, 32, 1. [Google Scholar]
- Scherer, R.P.; DeConto, R.M.; Pollard, D.; Alley, R.B. Windblown Pliocene diatoms and East Antarctic Ice Sheet retreat. Nat. Commun. 2016, 7, 12957. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, S.; Larocque-Tobler, I.; Richard, P.J.; Pienitz, R.; St-Onge, G.; Fye, F. Diatom-inferred wind activity at Lac du Sommet, southern Québec, Canada: A multiproxy paleoclimate reconstruction based on diatoms, chironomids and pollen for the past 9500 years. Holocene 2011, 21, 925–938. [Google Scholar] [CrossRef]
- Wang, L.; Lu, H.; Liu, J.; Gu, Z.; Mingram, J.; Chu, G.; Li, J.; Rioual, P.; Negendank, J.F.W.; Han, J.; et al. Diatom-based inference of variations in the strength of Asian winter monsoon winds between 17,500 and 6000 calendar years B.P. J. Geophys. Res. Space Phys. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.R.; Bracegirdle, T.; Turner, J.; Wolff, E. A 308 year record of climate variability in West Antarctica. Geophys. Res. Lett. 2013, 40, 5492–5496. [Google Scholar] [CrossRef] [Green Version]
- Nye, J.F. Correction Factor for Accumulation Measured by the Thickness of the Annual Layers in an Ice Sheet. J. Glaciol. 1963, 4, 785–788. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.R.; Bracegirdle, T. Precipitation pathways for five new ice core sites in Ellsworth Land, West Antarctica. Clim. Dyn. 2014, 44, 2067–2078. [Google Scholar] [CrossRef]
- Thomas, E.R.; Van Wessem, M.; Roberts, J.; Isaksson, E.; Schlosser, E.; Fudge, T.J.; Vallelonga, P.; Medley, B.; Lenaerts, J.T.M.; Bertler, N.A.N.; et al. Regional Antarctic snow accumulation over the past 1000 years. Clim. Past 2017, 13, 1491–1513. [Google Scholar] [CrossRef] [Green Version]
- Fetterer, F.; Knowles, K.; Meier, W.; Savoie, M.; Windnagel, A.K. Antarctic February and September 30-Year Means (1981–2010). In Sea Ice Index, Version 3; National Snow and Ice Data Centre: Boulder, CO, USA, 2017. [Google Scholar]
- Reanalysis Map of Mean Annual Wind Speed (M/S) for the Interval 1979–2015. In Climate Reanalyzer; Climate Change Institute, University of Maine: Orono, ME, USA, 2018; Available online: https://climatereanalyzer.org/ (accessed on 25 February 2020).
- Dee, D.P.; Uppala, S.M.; Simmons, A.J.; Berrisford, P.; Poli, P.; Kobayashi, S.; Andrae, U.; Balmaseda, M.A.; Balsamo, G.; Bauer, P.; et al. The Era-Interim Reanalysis: Configuration and Performance of the Data Assimilation System. Q. J. Roy. Meteorol. Soc. 2011, 137, 553–597. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E.E. Chapter 2: Marine Diatoms. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 5–385. [Google Scholar] [CrossRef]
- Johansen, J.; Fryxell, G.A. The genus Thalassiosira (Bacillariophyceae): Studies on species occurring south of the Antarctic Convergence Zone. Phycologia 1985, 24, 155–179. [Google Scholar] [CrossRef]
- Cefarelli, A.O.; Ferrario, M.E.; Almandoz, G.O.; Atencio, A.G.; Akselman, R.; Vernet, M. Diversity of the diatom genus Fragilariopsis in the Argentine Sea and Antarctic waters: Morphology, distribution and abundance. Polar Boil. 2010, 33, 1463–1484. [Google Scholar] [CrossRef] [Green Version]
- Armand, L.; Cornet-Barthaux, V.; Mosseri, J.; Quéguiner, B. Late summer diatom biomass and community structure on and around the naturally iron-fertilised Kerguelen Plateau in the Southern Ocean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 653–676. [Google Scholar] [CrossRef]
- Esper, O.; Gersonde, R. Quaternary surface water temperature estimations: New diatom transfer functions for the Southern Ocean. Palaeogeogr. Palaeoclim. Palaeoecol. 2014, 414, 1–19. [Google Scholar] [CrossRef]
- Esper, O.; Gersonde, R. New tools for the reconstruction of Pleistocene Antarctic sea ice. Palaeogeogr. Palaeoclim. Palaeoecol. 2014, 399, 260–283. [Google Scholar] [CrossRef]
- Abram, N.J.; Thomas, E.R.; McConnell, J.R.; Mulvaney, R.; Bracegirdle, T.; Sime, L.C.; Aristarain, A.J. Ice core evidence for a 20th century decline of sea ice in the Bellingshausen Sea, Antarctica. J. Geophys. Res. Space Phys. 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.R.; Dennis, P.; Bracegirdle, T.; Franzke, C. Ice core evidence for significant 100-year regional warming on the Antarctic Peninsula. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef] [Green Version]
- Hamsher, S.; Kopalová, K.; Kociolek, J.P.; Zidarova, R.; Van De Vijver, B. Kopalov The genus Nitzschia on the South Shetland Islands and James Ross Island. Fottea 2016, 16, 79–102. [Google Scholar] [CrossRef] [Green Version]
- Scott, F.J.; Marchant, H.J. (Eds.) Antarctic Marine Protists; Australian Biological Resources Study; CSIRO Publishing: Clayton, Australia, 2005. [Google Scholar]
- Allen, C.S.; Pike, J.; Pudsey, C.J. Last glacial–interglacial sea-ice cover in the SW Atlantic and its potential role in global deglaciation. Quat. Sci. Rev. 2011, 30, 2446–2458. [Google Scholar] [CrossRef]
- Hasle, G.R. The biogeography of some marine planktonic diatoms. Deep. Sea Res. Oceanogr. Abstr. 1976, 23. [Google Scholar] [CrossRef]
- Moore, J.K.; Abbott, M.R. Phytoplankton chlorophyll distributions and primary production in the Southern Ocean. J. Geophys. Res. Space Phys. 2000, 105, 28709–28722. [Google Scholar] [CrossRef]
- Mohan, R.; Quarshi, A.A.; Meloth, T.; Sudhakar, M. Diatoms from the Surface Waters of the Southern Ocean during the Austral Summer of 2004. Curr. Sci. 2011, 100, 1323–1327. [Google Scholar]
- Bathmann, U.; Scharek, R.; Klaas, C.; Dubischar, C.; Smetacek, V. Spring development of phytoplankton biomass and composition in major water masses of the Atlantic sector of the Southern Ocean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 1997, 44, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Assmy, P.; Smetacek, V.; Montresor, M.; Klaas, C.; Henjes, J.; Strass, V.; Arrieta, J.; Bathmann, U.; Berg, G.M.; Breitbarth, E.; et al. Thick-shelled, grazer-protected diatoms decouple ocean carbon and silicon cycles in the iron-limited Antarctic Circumpolar Current. Proc. Natl. Acad. Sci. USA 2013, 110, 20633–20638. [Google Scholar] [CrossRef] [Green Version]
- Burckle, L.H.; Cirilli, J. Origin of Diatom Ooze Belt in the Southern Ocean: Implications for Late Quaternary Paleoceanography. Micropaleontology 1987, 33, 82–86. [Google Scholar] [CrossRef]
- Fischer, G.; Gersonde, R.; Wefer, G. Organic carbon, biogenic silica and diatom fluxes in the marginal winter sea-ice zone and in the Polar Front Region: Interannual variations and differences in composition. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 1721–1745. [Google Scholar] [CrossRef]
- Hasle, G.R. Family Bacillariaceae: The Genera Thalassiothrix and Trichotoxon. In Polar Marine Diatoms; Medlin, L., Priddle, J., Eds.; British Antarctic Survey: Cambridge, UK, 1990; pp. 133–136. [Google Scholar]
- Smetacek, V.; Klaas, C.; Menden-Deuer, S.; Rynearson, T. Mesoscale distribution of dominant diatom species relative to the hydrographical field along the Antarctic Polar Front. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 3835–3848. [Google Scholar] [CrossRef]
- Rigual-Hernández, A.S.; Trull, T.W.; Bray, S.G.; Cortina, A.; Armand, L. Latitudinal and temporal distributions of diatom populations in the pelagic waters of the Subantarctic and Polar Frontal zones of the Southern Ocean and their role in the biological pump. Biogeosciences 2015, 12, 5309–5337. [Google Scholar] [CrossRef] [Green Version]
- Warnock, J.P.; Scherer, R.P. Diatom species abundance and morphologically-based dissolution proxies in coastal Southern Ocean assemblages. Cont. Shelf Res. 2015, 102, 1–8. [Google Scholar] [CrossRef]
- Kellogg, D.E.; Kellogg, T.B. Glacial/Interglacial Variations in the Flux of Atmospherically Transported Diatoms in Taylor Dome Ice Core. Antarct. J. U. S. 1996, 31, 68–70. [Google Scholar]
- Burckle, L.H.; Gayley, R.I.; Ram, M.; Petit, J.-R. Diatoms in Antarctic ice cores: Some implications for the glacial history of Antarctica. Geology 1988, 16, 326. [Google Scholar] [CrossRef]
- Hosking, J.S.; Orr, A.; Marshall, G.J.; Turner, J.; Phillips, T.; Hosking, J.S. The Influence of the Amundsen–Bellingshausen Seas Low on the Climate of West Antarctica and Its Representation in Coupled Climate Model Simulations. J. Clim. 2013, 26, 6633–6648. [Google Scholar] [CrossRef] [Green Version]
- De Leeuw, G.; Andreas, E.L.; Anguelova, M.D.; Fairall, C.W.; Lewis, E.; O’Dowd, C.; Schulz, M.; Schwartz, S.E. Production flux of sea spray aerosol. Rev. Geophys. 2011, 49. [Google Scholar] [CrossRef] [Green Version]
- Grythe, H.; Ström, J.; Krejci, R.; Quinn, P.O.; Stohl, A. A review of sea-spray aerosol source functions using a large global set of sea salt aerosol concentration measurements. Atmospheric Chem. Phys. Discuss. 2014, 14, 1277–1297. [Google Scholar] [CrossRef] [Green Version]
- Troitskaya, Y.; Kandaurov, A.; Ermakova, O.; Kozlov, D.; Sergeev, D.; Zilitinkevich, S. Bag-breakup fragmentation as the dominant mechanism of sea-spray production in high winds. Sci. Rep. 2017, 7, 1614. [Google Scholar] [CrossRef]
- O’Dowd, C.; De Leeuw, G. Marine aerosol production: A review of the current knowledge. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2007, 365, 1753–1774. [Google Scholar] [CrossRef] [Green Version]
- Montiel, F.; Squire, V.; Bennetts, L. Reflection and transmission of ocean wave spectra by a band of randomly distributed ice floes. Ann. Glaciol. 2015, 56, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Salter, M.; Nilsson, E.D.; Butcher, A.; Bilde, M. On the seawater temperature dependence of the sea spray aerosol generated by a continuous plunging jet. J. Geophys. Res. Atmos. 2014, 119, 9052–9072. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, D.E.; Kellogg, T.B. Recent Diatom Distributions in the Amundsen Sea. Antarct. J. U. S. 1986, 161–162. [Google Scholar]
- Pike, J.; Allen, C.S.; Leventer, A.; Stickley, C.E.; Pudsey, C.J. Comparison of contemporary and fossil diatom assemblages from the western Antarctic Peninsula shelf. Mar. Micropaleontol. 2008, 67, 274–287. [Google Scholar] [CrossRef]
- Turner, J.; Phillips, T.; Hosking, J.S.; Marshall, G.J.; Orr, A.; Hosking, J.S. The Amundsen Sea low. Int. J. Clim. 2012, 33, 1818–1829. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.R.; Kwok, R.; Kwok, R. Wind-driven trends in Antarctic sea-ice drift. Nat. Geosci. 2012, 5, 872–875. [Google Scholar] [CrossRef]
- Thomas, E.R.; Abram, N.J. Ice core reconstruction of sea ice change in the Amundsen-Ross Seas since 1702 A.D. Geophys. Res. Lett. 2016, 43, 5309–5317. [Google Scholar] [CrossRef] [Green Version]
- Rousseaux, C.S.; Gregg, W.W. Interannual Variation in Phytoplankton Primary Production at A Global Scale. Remote. Sens. 2013, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Arrigo, K.R.; Dijken, G.L.; Bushinsky, S.M. Primary production in the Southern Ocean, 1997–2006. J. Geophys. Res. Space Phys. 2008, 113. [Google Scholar] [CrossRef]
- Soppa, M.; Völker, C.; Bracher, A. Diatom Phenology in the Southern Ocean: Mean Patterns, Trends and the Role of Climate Oscillations. Remote. Sens. 2016, 8, 420. [Google Scholar] [CrossRef] [Green Version]
- De Jong, J.; Schoemann, V.; Maricq, N.; Mattielli, N.; Langhorne, P.; Haskell, T.; Tison, J.-L. Iron in land-fast sea ice of McMurdo Sound derived from sediment resuspension and wind-blown dust attributes to primary productivity in the Ross Sea, Antarctica. Mar. Chem. 2013, 157, 24–40. [Google Scholar] [CrossRef]
- Planquette, H.; Statham, P.J.; Fones, G.R.; Charette, M.A.; Moore, M.; Salter, I.; Menet-Nedelec, F.; Taylor, S.L.; French, M.; Baker, A.; et al. Dissolved iron in the vicinity of the Crozet Islands, Southern Ocean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 1999–2019. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diatom Taxa | Sample Years | Number of Specimens | Total |
|---|---|---|---|
| Fragilariopsis kerguelensis | 2006 (2) 1994 1990 (2) 1989 (2) 1988 (2) | 2 1 2 2 2 | 9 |
| Thalassiosira lentiginosa | 2006 1995 1993 | 1 1 1 | 3 |
| Thalassiothrixgp | 2006 1989 | 1 1 | 2 |
| Eucampia antarctica | 2005 1989 | 1 1 | 2 |
| Fragilariopsisspp.* | 1998 1988 | 1 1 | 2 |
| Chaetoceros rs | 1982 | 1 | 1 |
| Dactyliosolengb | 1989 | 1 | 1 |
| Odontella sp. | 2005 | 1 | 1 |
| Nitzschia sp. | 1998 | 1 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allen, C.S.; Thomas, E.R.; Blagbrough, H.; Tetzner, D.R.; Warren, R.A.; Ludlow, E.C.; Bracegirdle, T.J. Preliminary Evidence for the Role Played by South Westerly Wind Strength on the Marine Diatom Content of an Antarctic Peninsula Ice Core (1980–2010). Geosciences 2020, 10, 87. https://doi.org/10.3390/geosciences10030087
Allen CS, Thomas ER, Blagbrough H, Tetzner DR, Warren RA, Ludlow EC, Bracegirdle TJ. Preliminary Evidence for the Role Played by South Westerly Wind Strength on the Marine Diatom Content of an Antarctic Peninsula Ice Core (1980–2010). Geosciences. 2020; 10(3):87. https://doi.org/10.3390/geosciences10030087
Chicago/Turabian StyleAllen, Claire S., Elizabeth R. Thomas, Hilary Blagbrough, Dieter R. Tetzner, Richard A. Warren, Emily C. Ludlow, and Thomas J. Bracegirdle. 2020. "Preliminary Evidence for the Role Played by South Westerly Wind Strength on the Marine Diatom Content of an Antarctic Peninsula Ice Core (1980–2010)" Geosciences 10, no. 3: 87. https://doi.org/10.3390/geosciences10030087