
Corollospora mediterranea: A Novel Species Complex in the Mediterranean Sea



Mycotheca Universitatis Taurinensis, Department of Life Sciences and Systems Biology, University of Torino, Viale Mattioli 25, 10125 Turin, Italy
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(12), 5452; https://doi.org/10.3390/app11125452
Submission received: 12 May 2021
/
Revised: 10 June 2021
/
Accepted: 11 June 2021
/
Published: 11 June 2021
(This article belongs to the Special Issue Discovery and Research on Aquatic Microorganisms)
Abstract
:The genus Corollospora, typified by the arenicolous fungus Corollospora maritima, consists of twenty-five cosmopolitan species that live and reproduce exclusively in marine environments. Species of this genus are known to produce bioactive compounds and can be potentially exploited as bioremediators of oil spill contaminated beaches; hence their biotechnological importance. In this paper, nine fungal strains isolated in the Mediterranean Sea, from the seagrass Posidonia oceanica (L.) Delile, from driftwood and seawater contaminated by an oil spill, were investigated. The strains, previously identified as Corollospora sp., were examined by deep multi-loci phylogenetic and morphological analyses. Maximum-likelihood and Bayesian phylogeny based on seven genetic markers led to the introduction of a new species complex within the genus Corollospora: Corollospora mediterranea species complex (CMSC). The Mediterranean Sea, once again, proves an extraordinary reservoir of novel fungal species with a still undiscovered biotechnological potential.
1. Introduction
The last decades have seen an increasing interest in marine fungi due to the need to broaden our knowledge on aquatic biodiversity and to exploit these organisms as a source of novel bioactive molecules. Although more than 1800 species inhabiting the oceans have been described so far, most of the fungal diversity, estimated to exceed 10,000 taxa [1], is yet to be unveiled. Several marine habitats and substrates, both biotic and abiotic, are still being explored worldwide, leading to the discovery of new marine fungal lineages. Among Sordariomycetes, one of the ascomycetous classes mostly detected in the sea, the family Halosphaeriaceae (order Microascales) usually dominates this habitat, with 65 genera and 166 species occurring on driftwood, algae, and seagrasses worldwide [2]. While most of the genera of this family are represented by one or two species, the genus Corollospora, typified by Corollospora maritima Werderm, includes 25 arenicolous species typically found in beach sand, sea-foam, shell fragments, and algal thalli [3,4,5,6]. Besides their ability to rapidly degrade cellulose [7], species affiliated with this genus are known to produce bioactive metabolites [8,9] and can be potentially exploited as bioremediators of oil spill contaminated beaches [10]. For instance, the phthalide derivative corollosporine isolated from C. maritima demonstrated a strong antibacterial activity against Staphylococcus aureus, Bacillus subtilis, and Escherichia coli [11], whereas pulchellalactam produced by C. pulchella, along with the antimicrobial dioxopiperazines melinacidins II–IV and gancidin W [12], showed inhibitory activity against the tyrosine phosphatase CD45 in lymphocytes [13]. Weak antibacterial activity was also observed in fractions of C. lacera mycelial extract [14].
In general, this genus includes morphologically diverse species whose most distinctive features are the ascospore apical primary and equatorial secondary appendages, respectively formed from the epispore and by the fragmentation of the exospore layer [15,16,17,18]. Indeed, in reference to the ascospore appendage ontogeny, a revision of the genus was done by scanning and transmission electron microscope investigations [16]. In almost twenty years, other species of Corollospora were morphologically described [3,19,20,21,22,23,24]. With the upcoming molecular techniques, Campbell and collaborators [15] performed phylogenetic analysis of Corollospora spp. (and related taxa) based on 28S rDNA sequences and confirmed the monophyletic nature of the genus. However, the authors concluded that more genetic markers, including protein-coding genes, were necessary to resolve relationships among the species of Corollospora [15]. New sequence data, including 18S rDNA, ITS, and RPB1, relative to Corollospora spp., were then generated in the framework of the Fungal Barcoding Consortium and AFTOL project (Assembly Fungal Tree Of Life) [25,26].
Recently, nine unidentified strains belonging to the genus Corollospora were isolated in surveys aimed to investigate the underwater fungal diversity of the Mediterranean Sea: 4 isolates were retrieved from the seagrass Posidonia oceanica [27], 4 from seawater contaminated by oil spill [28], and 1 from submerged wood [29]. It is not uncommon, as it is in this case, to come across marine fungi that neither sporulate nor develop reproductive structures in axenic culture, leaving traditional morphology-based identification impossible. Consequently, the identification of these sterile mycelia must rely on molecular data [30,31,32,33].
With this study, we wish to provide an accurate phylogenetic placement of the Mediterranean strains by applying a combined multi-locus molecular phylogeny. Besides, the paper gives morphological insights into the strains that turned out to represent new species.
2. Materials and Methods
2.1. Fungal Isolates
The isolates analyzed in this study were previously retrieved from the Mediterranean Sea. In detail, 4 isolates derived from a site chronically contaminated by an oil spill in Sicily (Gela, Italy) [28] 1 from driftwood sampled in the seawater off Porto Badisco (Apulia, Italy) [29] and 4 from leaves of P. oceanica collected in good health condition in Tuscany (Elba island, Italy) [27] (Table 1). Originally, the strains were isolated on Corn Meal Agar medium supplemented with sea salts (CMASS; 3.5% w/v sea salt mix, Sigma-Aldrich, SL, USA, in ddH2O) and are preserved at the Mycotheca Universitatis Taurinensis (MUT), Italy.
2.2. Morphological Analysis
All isolates were pre-grown on Malt Extract Agar-sea water (MEASW; 20 g malt extract, 20 g glucose, 2 g peptone, 20 g agar in 1 L of seawater) for one month at 21 °C prior to inoculation in triplicate onto new MEASW Petri dishes (9 cm Ø). Petri dishes were incubated at 15 and/or 21 °C. The colony growth was monitored periodically for 28 days, while macroscopic and microscopic features were assessed at the end of the incubation period.
In an attempt to induce sporulation, sterile pieces of Quercus ruber cork and Pinus pinaster wood (species autochthonous to the Mediterranean area) were placed on 3-week old fungal colonies [34]. Petri dishes were further incubated for 4 weeks at 21 °C. Following, cork and wood specimens were transferred to 50 mL tubes containing 20 mL of sterile seawater. Samples were incubated at 21 °C for at least three months.
Morphological structures were observed, and images captured using an optical microscope (Leica DM4500B, Leica microsystems GmbH, Germany) equipped with a camera (Leica DFC320, Leica microsystems GmbH, Germany).
2.3. DNA Extraction, PCR Amplification, and Data Assembling
Approximately 100 mg of fresh mycelium were carefully scraped from MEASW plates, transferred to a 2 mL Eppendorf tube, and disrupted by the mean of an MM400 tissue lyzer (Retsch GmbH, Haan, Germany). Genomic DNA was extracted following the manufacturer’s instructions of a NucleoSpin kit (Macherey Nagel GmbH, Duren, DE, USA). The quality and quantity of DNA were measured spectrophotometrically (Infinite 200 PRO NanoQuant; TECAN, Switzerland); DNA samples were stored at −20 °C.
The partial sequences of seven genetic markers were amplified by PCR. Primer pairs ITS1/ITS4 [35], LR0R/LR7 [36], NS1/NS4 [35] were used to amplify the internal transcribed spacers, including the 5.8S rDNA gene (nrITS), 28S large ribosomal subunit (nrLSU) and 18S small ribosomal subunit (nrSSU). The translation elongation factor (TEF-1α), the β-tubulin (β-TUB) and the largest and second-largest subunits of RNA polymerase II (RPB1 and RPB2) were amplified by using the following primer pairs: EF-dF/EF-2218R [37], Bt2a/Bt2b [38], RPB1Af/RPB1Cr [39] and fRPB2-5F/fPB2-7R [40]. Amplifications were run in a T100 Thermal Cycler (Bio-Rad, Hercules, CA, USA) programmed as described in Table 2. Reaction mixtures consisted of 20–40 ng DNA template, 10× PCR Buffer (15 mM MgCl2, 500 mM KCl, 100 mM Tris-HCl, pH 8.3), 200 µM each dNTP, 1 μM each primer, 2.5 U Taq DNA Polymerase (Qiagen, Chatsworth, CA, USA), in 50 μL final volume. For problematic cases, additional MgCl2, BSA, and/or 2.5% DMSO facilitated the reaction.
Amplicons, together with a GelPilot 1 kb plus DNA Ladder, were visualized on a 1.5% agarose gel stained with 5 mL 100 mL−1 ethidium bromide; PCR products were purified and sequenced at the Macrogen Europe Laboratory (Madrid, Spain). The resulting Applied Biosystem (ABI) chromatograms were inspected, trimmed, and assembled to obtain consensus sequences using Sequencer 5.0 (GeneCodes Corporation, Ann Arbor, MI, USA, http://www.genecodes.com acessed on 12 May 2021). Newly generated sequences were deposited in GenBank (Table 1).
2.4. Sequence Alignment and Phylogenetic Analysis
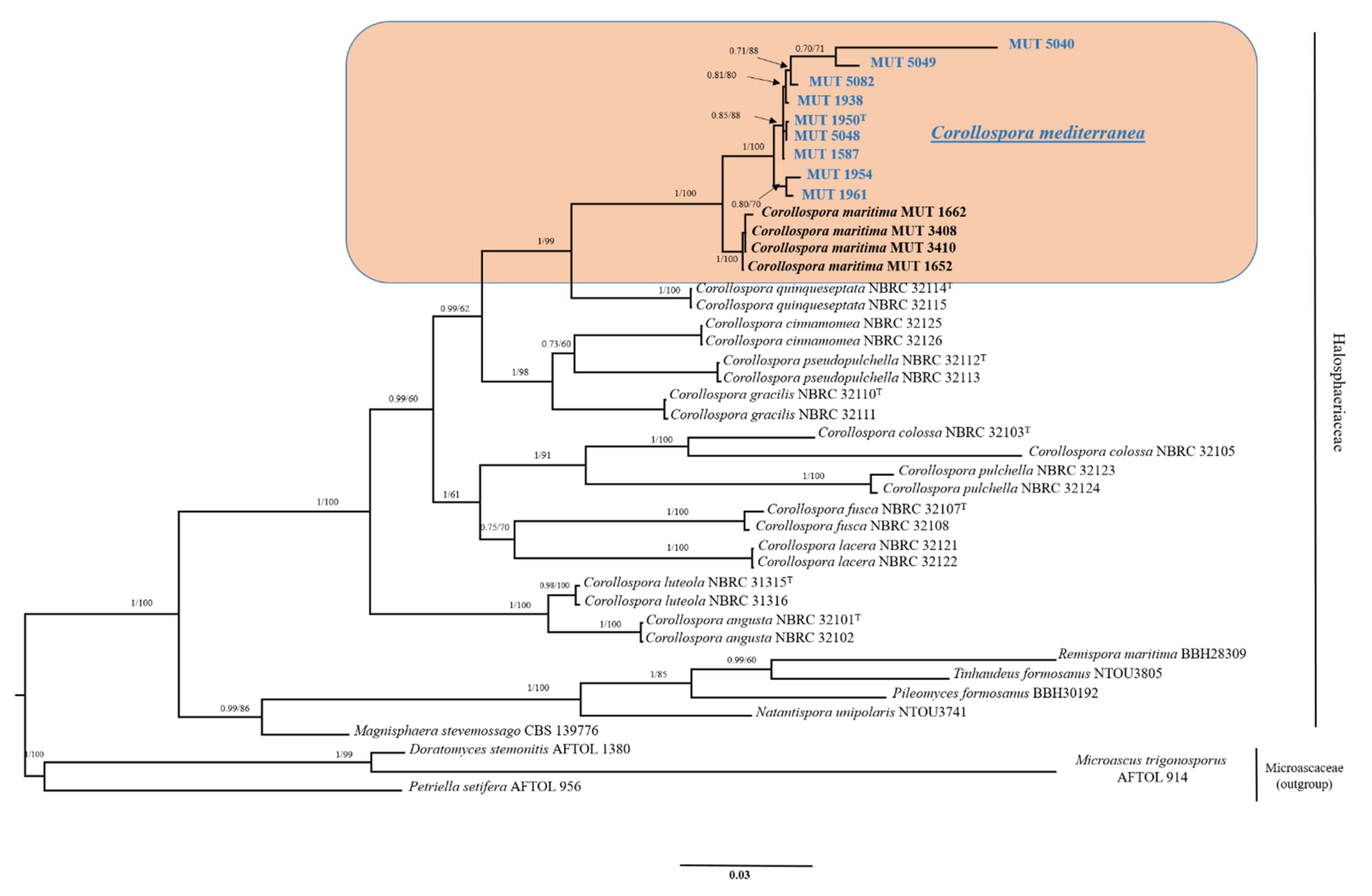
A dataset consisting of nrSSU, nrITS, nrLSU, and RPB1 was assembled based on BLASTn results and of the most recent phylogenetic studies focused on Halosphaeriaceae and Corollospora [30,41]. Reference sequences were retrieved from GenBank (Table 1). Sequences were aligned using MUSCLE (default conditions for gap openings and gap extension penalties), implemented in MEGA X (Molecular Evolutionary Genetics Analysis), visually inspected, and manually trimmed to delimit and discard ambiguously aligned regions. Since no incongruence was observed among single-loci phylogenetic trees, alignments were concatenated into a single data matrix with SequenceMatrix [42]. The best evolutionary model under the Akaike Information Criterion (AIC) was determined with jModelTest 2 [43]. Phylogenetic inference was estimated using Maximum Likelihood (ML) and Bayesian Inference (BI) criteria. The ML analysis was generated using RAxML v. 8.1.2 [44] under GTR + I + G evolutionary model and 1000 bootstrap replicates. Support values from bootstrapping runs (MLB) were mapped on the globally best tree using the “-f a” option of RAxML and “-x 12345” as a random seed to invoke the novel rapid bootstrapping algorithm. BI was performed with MrBayes 3.2.2 [45] with the same substitution model (GTR + I + G). The alignment was run for 10 million generations with two independent runs each containing four Markov Chains Monte Carlo (MCMC) and sampling every 100 iterations. The first 25% of generated trees were discarded as “burn-in”. A consensus tree was generated using the “sumt” function of MrBayes and Bayesian posterior probabilities (BPP) were calculated. Consensus trees were visualized in FigTree v. 1.4.2 (http://tree.bio.ed.ac.uk/software/figtree acessed on 12 May 2021). Three species of Microascaceae, namely Doratomyces stemonitis, Microascus trigonosporus, and Petriella setifera, were used to root the tree. Due to the topological similarity of the two resulting trees, only Bayesian analysis with MLB and BPP values was reported (Figure 1).
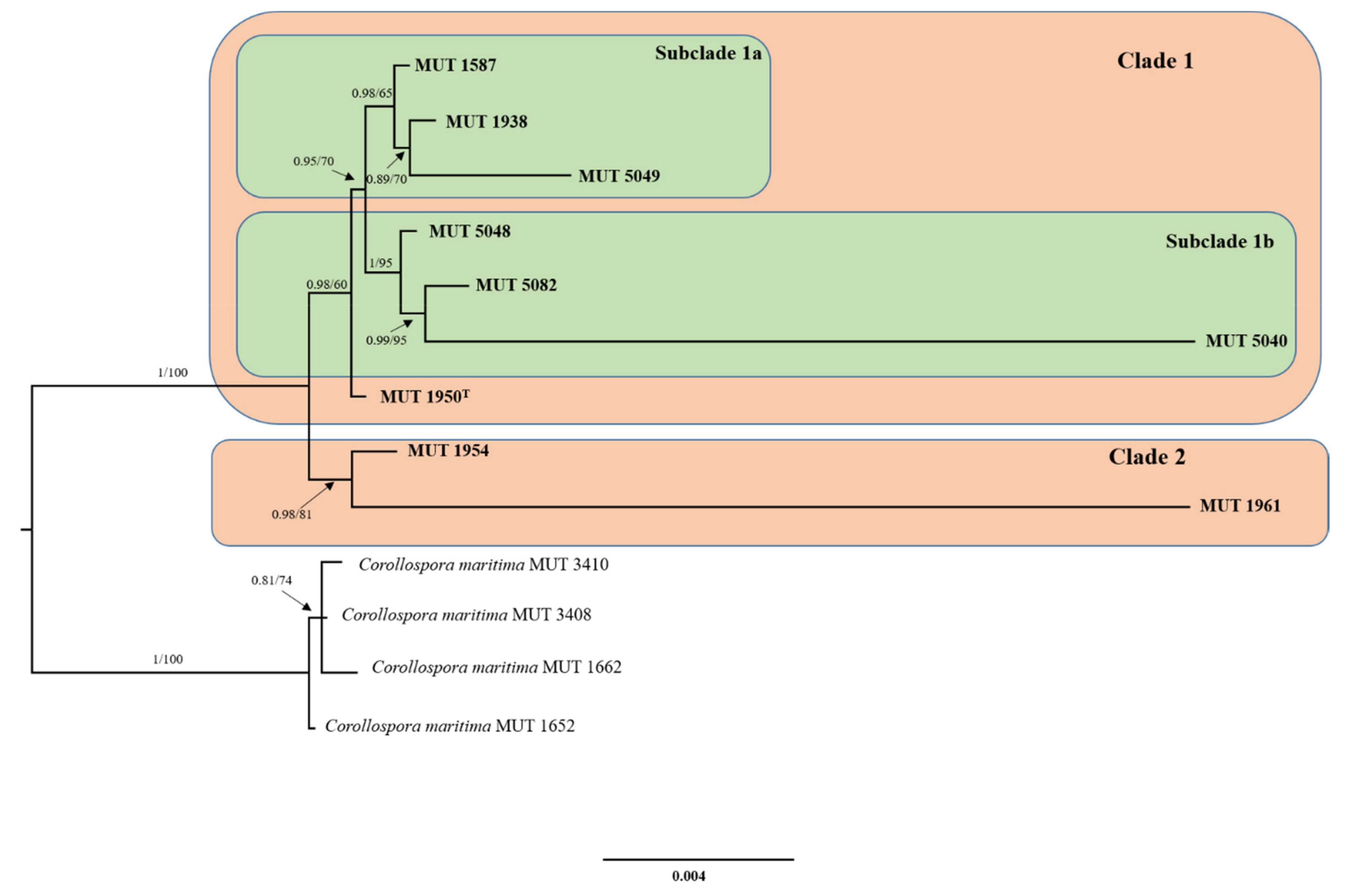
Following, a new phylogenetic analysis was conducted only on the strains investigated, whose relationship was unclear. To this aim, TEF-1α, β-TUB, and RPB2 sequences were added to the restricted dataset. Alignments and multi-loci phylogeny were conducted as described above. Sequence alignments and phylogenetic trees were deposited in TreeBASE (http://www.treebase.org, submission number S27921 and S27923, acessed on 12 May 2021).
3. Results
3.1. Phylogenetic Inference
Preliminary analyses carried out individually with nrITS, nrSSU, nrLSU, and RPB1 revealed no incongruence in the topology of the single-locus trees. The combined four-markers dataset–built based on BLASTn results and of recent phylogenetic studies [30,41]—Consisted of 42 taxa (including MUT isolates) that represented 9 genera and 20 species (Table 1). A total of 82 sequences (9 nrITS, 13 nrSSU, 13 nrLSU, 13 TEF-1α, 13 β-TUB, 11 RPB1, and 10 RPB2) were newly generated while 91 were retrieved from GenBank.
The dataset combining nrSSU, nrITS, nrLSU and RPB1 had an aligned length of 2431 characters, of which 1530 were conserved, 263 were parsimony-uninformative and 638 parsimony informative (TL = 2192, CI = 0.550901, RI = 0.758483, RC = 0.417849, HI = 0.449099). The strains under investigation, MUT 1587, MUT 1938, MUT 1950, MUT 1954, MUT 1961, MUT 5040, MUT 5048, MUT 5049 and MUT 5082 formed a strongly supported monophyletic lineage (BYPP = 1.00; MLB = 100%) close but well set apart from Corollospora marittima (Figure 1). Within this new group, MUT 1954 and MUT 1961 seemed to form a separated clade, as well as MUT 5040 and MUT 5049. However, the relationships between the taxa were unclear.
The additional dataset, implemented with the addition of TEF-1α, β-TUB, and RPB2 sequence data relative to the novel lineage and to Corollospora marittima only, had an aligned length of 4086 characters, of which 3693 were constant, 236 were parsimony-uninformative and 157 parsimony informative (TL = 452, CI = 0.843537, RI = 0.900217, RC = 0.759367, HI = 0.156463). Two clusters could be observed: the first grouped MUT 1954 and MUT 1961 while the second one was further split into two subclades that separated MUT 1587, MUT 1938, and MUT 5049 from MUT 5040, MUT 5048, and MUT 5082. Finally, MUT 1950, although included second cluster, was not part of any of the two subclades (Figure 2).
3.2. Taxonomy
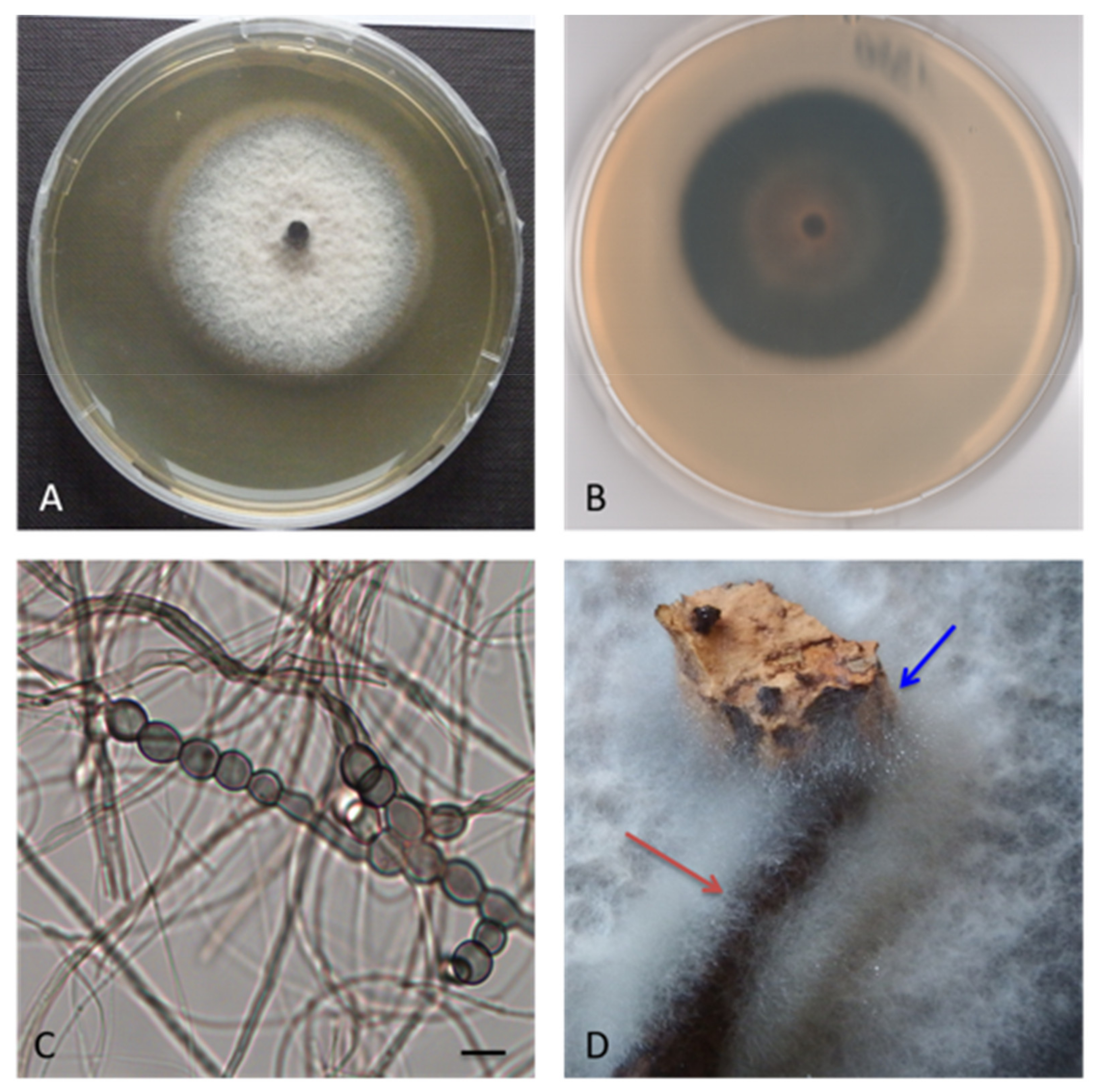
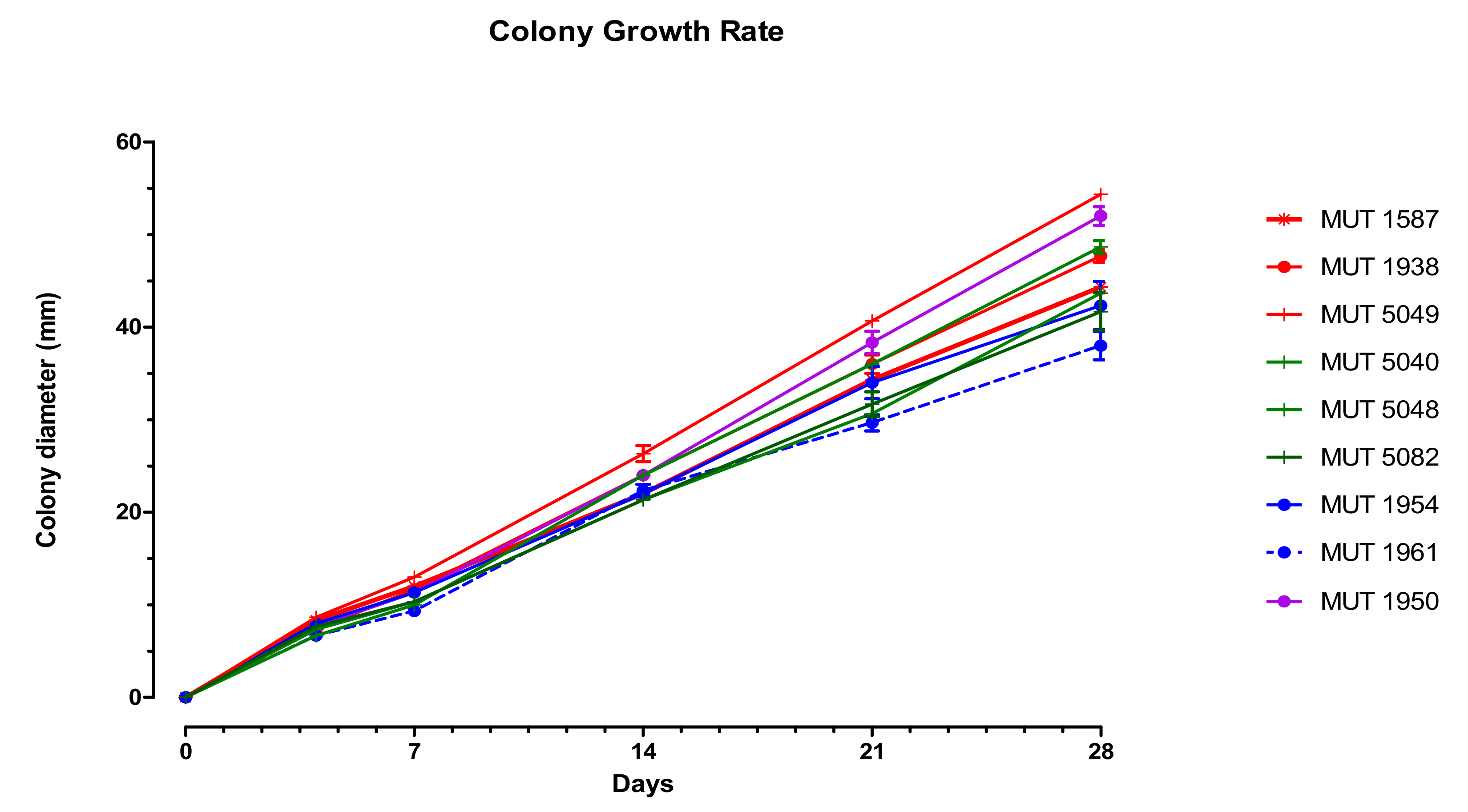
Corollospora mediterranea sp. nov. A. Poli, E. Bovio, G.C. Varese and V. Prigione, MYCOBANK: MB 839640, Type. Italy, Sicily, Mediterranean Sea, Gela (CL), July 2013, from seawater contaminated by an oil spill, R. Denaro, MUT 1950 holotype, living culture permanently-preserved in metabolically inactively state by deep-freezing at MUT. Additional material examined. Italy, Sicily, Mediterranean Sea, Gela (CL), July 2013, from seawater contaminated by an oil spill, R. Denaro MUT 1938, MUT 1954 and MUT 1961. Italy, Apulia, Mediterranean Sea, Porto Badisco (LE), July 2011, from driftwood, L. Garzoli, MUT 1587. Italy, Tuscany, Mediterranean Sea, Elba Island (LI), from the seagrass Posidonia oceanica, March 2010, R. Mussat-Sartor and N. Nurra, MUT 5040, MUT 5048, MUT 5049 and MUT 5082. Etymology. In reference to the Mediterranean Sea. Description. Growing actively on Pinus pinaster and Quercus ruber cork. Hyphae two types observed, one thin (1 μm wide) and hyaline, one thicker (3 μm wide) and melanized. Chlamydospores numerous, in chain globose to subglobose, 4–13 × 4–11 μm diameter (Figure 3). Sexual morph not observed. Asexual morph with differentiated conidiogenesis not observed. Colony description. Colonies on MEASW attaining 38–54 mm diam after 28 days at 21 °C (Figure 4), mycelium white, grey, grey-green, sometimes with pinkish shades; dense and feltrose, occasionally umbonate in the middle, with radial grooves; reverse from light brown to dark green, occasionally with concentric rings, lighter to the edges. Neither soluble pigments nor exudates were observed (Figure 3).
4. Discussion
The description of the strains investigated in this study was particularly tricky and complicated since neither asexual nor sexual reproductive structures developed in axenic conditions. As a consequence, it was not possible to describe the range of diagnostic traits amongst these newly identified lineages.
To better characterize these fungi, we tried to mimic the saline environment by using a culture medium supplemented with seawater, since it is well known that only this method supports a measurable growth of vegetative mycelium [33]. The family Halospaheriaceae, to whom the genus Corollospora belongs, includes the largest number of marine lignicolous species. It was thus realistic to induce sporulation by placing wood and cork specimens on the colony surface prior to transfer them into seawater. Despite wood colonization, sporulation did not occur, whereas chlamydospores were abundantly produced. Likewise, strains of Corollospora sp. isolated from intertidal decayed wood of mangrove trees and driftwood in Saudi Arabia did not develop reproductive structures but only chlamydospores that were much bigger and not comparable to those observed in this study [30]. It must be considered that strictly vegetative growth with no sporulation is a common feature of marine fungi [46,47,48] that are likely to rely on their dispersion to hyphal fragments and/or chlamydospores. Most probably, a necessary requisite for fungi to develop reproductive structures is the occurrence of those environmental conditions these organisms are adapted to (e.g., high salinity, low temperature, high pressure, wet-dry cycles, etc.).
The phylogenetic analysis based on ribosomal genes (nrITS, nrLSU, nrSSU) and RPB1, shows a clear distance between the strains under investigation and the other species of Corollospora, Corollospora maritima being the closest one. This new and strongly supported clade may include one or more novel species. Hypothetically, the tree highlights the presence of three clusters that include: (i) MUT 1954 and MUT 1961; (ii) MUT 1938, MUT 5040, MUT 5049 and MUT 5082; (iii) MUT 1587, MUT 1950, and MUT 5048 (Figure 1). Bearing in mind the conclusion drawn by Campbell et al. [15] that indicated the need of sequencing more genetic markers to clarify the relations among the species of the genus Corollospora, a dataset focused on this new group, together with C. maritima, was built with the addition of three more protein-coding genes, namely RPB2, TEF-1α and β-TUB (Figure 2). From one side, this strongly supported tree points out once more the distance from C. maritima, strengthening the idea that we are dealing with a new species. It is undeniably complicated to define a fungal species, and Corollospora mediterranea was here established following the recommendations outlined by Jeewon and Hyde [49], who, dealing with the issue of the species boundaries and identification of new taxa, pointed out a number of key elements to follow. Notably, all the ITS sequences (including 5.8S) analyzed are longer than the minimum 450 base pairs required and display a percentage of identity with the closest relative C. maritima < 95%. In addition, as recommended, the strongly supported phylogenetic tree includes the minimum number (4–5) of closely related taxa of the same genus (Figure 1). A thorough inspection of the tree shown in Figure 2 reveals the presence of two clusters: the former (Clade 1) consists of MUT 1950 and two subclusters (MUT 5040, MUT 5048 and MUT 5082; MUT 1587, MUT 1938, MUT 5049), the latter (Clade 2) includes MUT 1954 and MUT 1961. On the other hand, the lack of distinct micro-morphological traits (neither sexual nor asexual morph with differentiated conidiogenesis detected) and the huge variability observed among the colonies (in terms of growth rate, texture, surface and reverse color) lead us to introduce the C. mediterranea species complex (CMSC), since we could not discern the species boundaries with certainty (Figure 5 and Figure 6). Indeed, the term “species complex” comes to help taxonomists when: (i) a group of organisms may represent more than one species; (ii) morphological features are overlapping due to extreme variability; (iii) the species may be somehow related although no certain assumption can be assessed. Furthermore, no clear correlation between colony features, growth rates, source of isolation, and/or phylogenetic position was noticed. In general, researchers introduce a species complex when facing a problematic topic. This is the case for example of Fusarium oxysporum species complex (FOSC) [50,51], Fusarium solani species complex (FSSC) [52], Wallemia sebi species complex (WSSC) [53], Colletotrichum acutatum species complex (CASC) [54] and many others. Most of these cases were resolved with a revision of the species complex, where individual species were identified based on a multi-loci phylogeny. However, in our study, this approach did not lead to a sharp resolution of the complex. The genome sequencing of all the strains investigated would be an option to sort out this intriguing issue. This approach would possibly reveal those genetic regions that may allow an easy distinction of the species within the complex [55]. On the other hand, the same goal could be achieved by the analysis of secondary metabolites [53]. An investigation of this sort is important also from another point of view: it is now recognized that marine fungi are a reservoir of novel active metabolites that can be harnessed for pharmaceutical, nutraceutical, cosmetic, and environmental purposes.
Kirk and Gordon [10] demonstrated that a number of strains of C. maritima, C. lacera, and C. intermedia were capable of using hexadecane as the sole carbon source, and assumed that the genus Corollospora could find utility in petroleum degradation. Supporting this idea is the isolation of 4 strains of C. mediterranea (MUT 1938, MUT 1950, MUT 1954, and MUT 1961) from a site chronically contaminated by an oil spill in the Mediterranean Sea [28]. The aforementioned findings indicate the great versatility and adaptability of these fungi to hydrocarbons-contaminated environments, with the consequent possibility of being used as bioremediators. In addition, knowing that the species of Corollospora belong to a family of lignicolous fungi and that are rich in cellulase and lignin-degrading enzymes [7], their retrieval from leaves of the seagrass P. oceanica (MUT 5040, MUT 5048, MUT 5049, and MUT 5082) and from driftwood (MUT 1587) does not come as a surprise. It is, therefore, reasonable to assume a role of C. mediterranea in degrading and recycling organic matter, making them available for other organisms.
5. Conclusions
In conclusion, the retrieval of C. mediterranea from different substrates, localities and in different sampling campaigns indicates its constant presence in (at least) the Mediterranean Sea and points out how the marine environments is still largely uninvestigated from a mycological point of view. Therefore, it is more and more glaring that the Oceans are a huge reservoir of unidentified microorganisms with a valuable biotechnological potential not yet disclosed. In the next future, we would aim at resolving the species complex, by applying more approaches to induce sexual and/or asexual sporulation. In parallel, all the strains studied in this work, will be investigated for the production of new and powerful bioactive molecules.
Author Contributions
Conceptualization, A.P., E.B., and V.P.; methodology, A.P., E.B., and V.P.; software, A.P.: validation, A.P., I.P., and V.P.; formal analysis, A.P., E.B., and V.P.; investigation, A.P., E.B., and V.P.; resources, V.P. and G.C.V.; data curation, A.P., I.P., and V.P.; writing—original draft preparation, A.P. and V.P.; writing—review and editing, A.P., V.P., and G.C.V.; visualization, A.P. and V.P.; supervision, V.P. and G.C.V.; project administration, A.P., V.P., and G.C.V.; funding acquisition, G.C.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the University of Torino (ex 60%).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Sequence data are available at Genbank (NCBI) under the accession numbers reported in the manuscript.
Acknowledgments
The authors would like to thank JRU MIRRI-IT for technical and scientific support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jones, E.B.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J.; et al. An online resource for marine fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Jones, E.G.; Ju, W.-T.; Lu, C.-L.; Guo, S.-Y.; Pang, K.-L. The halosphaeriaceae revisited. Bot. Mar. 2017, 60, 453–468. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Nagahama, T.; Abdel-Aziz, F.A. Two new Corollospora species and one new anamorph based on morphological and molecular data. Mycoscience 2009, 50, 147–155. [Google Scholar] [CrossRef]
- Hsieh, S.Y.; Moss, S.T.; Jones, E.B.G. Ascoma development in the marine ascomycete Corollospora gracilis (Halosphaeriales, Hypocreomycetidae, Sordariomycetes). Bot. Mar. 2007, 50, 302–313. [Google Scholar] [CrossRef]
- Walker, A.K.; Robicheau, B.M. Fungal diversity and community structure from coastal and barrier island beaches in the United States Gulf of Mexico. Sci. Rep. 2021, 11, 3889. [Google Scholar] [CrossRef] [PubMed]
- Velez, P.; Gasca-Pineda, J.; Nakagiri, A.; Hanlin, R.T.; Gonzalez, M.C. Genetic diversity and population structure of Corollospora maritima sensu lato: New insights from population genetics. Bot. Mar. 2016, 59, 307–320. [Google Scholar] [CrossRef]
- Raghukumar, S. Fungi in Coastal and Oceanic Marine Ecosystems: Marine Fungi; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Overy, D.P.; Bayman, P.; Kerr, R.G.; Bills, G.F. An assessment of natural product discovery from marine (sensu strictu) and marine-derived fungi. Mycology 2014, 5, 145–167. [Google Scholar] [CrossRef]
- Overy, D.P.; Rama, T.; Oosterhuis, R.; Walker, A.K.; Pang, K.L. The Neglected Marine Fungi, Sensu stricto, and Their Isolation for Natural Products’ Discovery. Mar. Drugs 2019, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Kirk, P.W.; Gordon, A.S. Hydrocarbon degradation by fiilamentous marine higher fungi. Mycologia 1988, 80, 776–782. [Google Scholar] [CrossRef]
- Liberra, K.; Jansen, R.; Lindequist, U. Corollosporine, a new phthalide derivative from the marine fungus Corollospora maritima Werderm. 1069. Pharmazie 1998, 53, 578–581. [Google Scholar] [CrossRef]
- Furuya, K.; Okudaira, M.; Shindo, T.; Sato, A. Corollospora pulchella, a marine fungus producing antibiotics, melinacidins III, IV and gancidin W. Annu. Rep. Sankyo Res. Lab. 1985, 37, 140–142. [Google Scholar]
- Alvi, K.A.; Casey, A.; Nair, B.G. Pulchellalactam: A CD45 protein tyrosine phosphatase inhibitor from the marine fungus Corollospora pulchella. J. Antibiot. 1998, 51, 515–517. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, S.E.; Gurusamy, G.S.; Piorko, A.; Strongman, D.B.; Hu, T.M.; Wright, J.L.C. Isolation of sterols from the marine fungus Corollospora lacera. Can. J. Microbiol. 2004, 50, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.; Shearer, C.A.; Mitchell, J.I.; Eaton, R.A. Corollospora revisited: A molecular approach. In Fungi in Marine Environment; Hyde, K.D., Ed.; (Fungal Diversity Research Series); Fungal Diversity Press: Hong Kong, China, 2002; Volume 7, pp. 15–33. [Google Scholar]
- Jones, E.G.; Johnson, R.; Moss, S. Taxonomic studies of the Halosphaeriaceae: Corollospora Werdermann. Bot. J. Linn. Soc. 1983, 87, 193–212. [Google Scholar] [CrossRef]
- Lutley, M.; Wilson, I.M. Development and fine structure of ascospores in the marine fungus Ceriosporopsis halima. Trans. Br. Mycol. Soc. 1972, 58, 393–402. [Google Scholar] [CrossRef]
- Lutley, M.; Wilson, I.M. Observations on the fine structure of ascospores of marine fungi: Halosphaeria appendiculata, Torpedospora radiata and Corollospora maritima. Trans. Br. Mycol. Soc. 1972, 59, 219–227. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Sakayaroj, J.; Suetrong, S.; Somrithipol, S.; Pang, K.L. Classification of marine Ascomycota, anamorphic taxa and Basidiomycota. Fungal Divers. 2009, 35, 187. [Google Scholar]
- Koch, J. Some Lignicolous Marine Fungi From Thailand, including 2 New Species. Nord. J. Bot. 1986, 6, 497–499. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Volkmannkohlmeyer, B. Corollospora armoricana sp. nov An Arenicolous Ascomycete from Brittany (France). Can. J. Bot. Rev. Can. Bot. 1989, 67, 1281–1284. [Google Scholar] [CrossRef]
- McKeown, T.A.; Moss, S.T.; Jones, E.B.G. Ultrastructure of the melanized ascospores of Corollospora fusca. Can. J. Bot. Rev. Can. Bot. 1996, 74, 60–66. [Google Scholar] [CrossRef]
- Jones, E.B.G. Fifty years of marine mycology. Fungal Divers. 2011, 50, 73–112. [Google Scholar] [CrossRef]
- Sundari, R.; Vikineswary, S.; Yusoff, M.; Jones, E.B.G. Corollospora besarispora, a new arenicolous marine fungus from Malaysia. Mycol. Res. 1996, 100, 1259–1262. [Google Scholar] [CrossRef]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panno, L.; Bruno, M.; Voyron, S.; Anastasi, A.; Gnavi, G.; Miserere, L.; Varese, G.C. Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. New Biotechnol. 2013, 30, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Bovio, E.; Gnavi, G.; Prigione, V.; Spina, F.; Denaro, R.; Yakimov, M.; Calogero, R.; Crisafi, F.; Varese, G.C. The culturable mycobiota of a Mediterranean marine site after an oil spill: Isolation, identification and potential application in bioremediation. Sci. Total Environ. 2017, 576, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Gnavi, G.; Tamma, F.; Tosi, S.; Varese, G.C.; Picco, A.M. Sink or swim: Updated knowledge on marine fungi associated with wood substrates in the Mediterranean Sea and hints about their potential to remediate hydrocarbons. Prog. Oceanogr. 2015, 137, 140–148. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Hodhod, M.S.; Bahkali, A.H.A.; Jones, E.B.G. Marine fungi of Saudi Arabia. Bot. Mar. 2014, 57, 323–335. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Wanasinghe, D.N.; Devadatha, B.; Abeywickrama, P.; Jones, E.B.G.; Chomnunti, P.; Sarma, V.V.; Hyde, K.D.; Lumyong, S.; McKenzie, E.H.C. Modern taxonomic approaches to identifying diatrypaceous fungi from marine habitats, with a novel genus Halocryptovalsa Dayarathne & KDHyde, gen. nov. Cryptogam. Mycol. 2020, 41, 21–67. [Google Scholar] [CrossRef]
- Jones, E.G.; Pang, K.-L. (Eds.) Marine Fungi and Fungal-Like Organisms; Walter de Gruyter: Berlin, Germany, 2012. [Google Scholar]
- Poli, A.; Bovio, E.; Ranieri, L.; Varese, G.C.; Prigione, V. News from the Sea: A New Genus and Seven New Species in the Pleosporalean Families Roussoellaceae and Thyridariaceae. Diversity 2020, 12, 144. [Google Scholar] [CrossRef] [Green Version]
- Panebianco, C.; Tam, W.Y.; Jones, E.B.G. The effect of pre-inoculation of balsa wood by selected marine fungi and their effect on subsequent colonisation in the sea. Fungal Divers. 2002, 10, 77–88. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenetics Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Raja, H.A.; Baker, T.R.; Little, J.G.; Oberlies, N.H. DNA barcoding for identification of consumer-relevant mushrooms: A partial solution for product certification? Food Chem. 2017, 214, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sakayaroj, J.; Pang, K.L.; Jones, E.B.G. Multi-gene phylogeny of the Halosphaeriaceae: Its ordinal status, relationships between genera and morphological character evolution. Fungal Divers. 2011, 46, 87–109. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Gnavi, G.; Garzoli, L.; Polil, A.; Prigione, V.; Burgaud, G.; Varese, G.C. The culturable mycobiota of Flabellia petiolata: First survey of marine fungi associated to a Mediterranean green alga. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Poli, A.; Prigione, V.; Gnavi, G.; Varese, G.C. Peacock’s tail with a fungal cocktail: First assessment of the mycobiota associated with the brown alga Padina pavonica. Fungal Ecol. 2018, 35, 87–97. [Google Scholar] [CrossRef]
- Poli, A.; Vizzini, A.; Prigione, V.; Varese, G.C. Basidiomycota isolated from the Mediterranean Sea—Phylogeny and putative ecological roles. Fungal Ecol. 2018, 36, 51–62. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Kistler, H.C. Genetic diversity in the plant-pathogenic fungus Fusarium oxysporum. Phytopathology 1997, 87, 474–479. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenetics Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- O’Donnell, K. Molecular phylogeny of the Nectria haematococca-Fusarium solani species complex. Mycologia 2000, 92, 919–938. [Google Scholar] [CrossRef]
- Jancic, S.; Nguyen, H.D.T.; Frisvad, J.C.; Zalar, P.; Schroers, H.J.; Seifert, K.A.; Gunde-Cimerman, N. Taxonomic Revision of the Wallemia sebi Species Complex. PLoS ONE 2015, 10, e0125933. [Google Scholar] [CrossRef] [Green Version]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matute, D.R.; Sepulveda, V.E. Fungal species boundaries in the genomics era. Fungal Genet. Biol. 2019, 131, 9. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic inference based on a combined nrITS, nrSSU, nrLSU, RPB1 dataset. The tree is rooted in three species of Miroascaceae. Branch numbers indicate BYPP and BS values; T Type Strain; Bar = expected changes per site (0.03).
Figure 1.
Phylogenetic inference based on a combined nrITS, nrSSU, nrLSU, RPB1 dataset. The tree is rooted in three species of Miroascaceae. Branch numbers indicate BYPP and BS values; T Type Strain; Bar = expected changes per site (0.03).

Figure 2.
Phylogenetic inference based on a combined nrITS, nrSSU, nrLSU, RPB1, RPB2, TEF-1α, and βTUB dataset. The tree is midrooted. Branch numbers indicate BYPP and BS values; T Type Strain; Bar = expected changes per site (0.004).
Figure 2.
Phylogenetic inference based on a combined nrITS, nrSSU, nrLSU, RPB1, RPB2, TEF-1α, and βTUB dataset. The tree is midrooted. Branch numbers indicate BYPP and BS values; T Type Strain; Bar = expected changes per site (0.004).

Figure 3.
Corollospora mediterranea sp. nov. MUT 1950 (holotype), the 28-days-old colony at 21 °C on MEASW (A) and reverse (B); chlamydospores in the chain (C); mycelial growth on Pinus pinaster wood (red arrow) and Quercus ruber cork (blue arrow) (D). Scale bars: 10 μm.
Figure 3.
Corollospora mediterranea sp. nov. MUT 1950 (holotype), the 28-days-old colony at 21 °C on MEASW (A) and reverse (B); chlamydospores in the chain (C); mycelial growth on Pinus pinaster wood (red arrow) and Quercus ruber cork (blue arrow) (D). Scale bars: 10 μm.

Figure 4.
Growth rate on MEASW of the strains investigated over 28 days. The symbol indicates the isolation substrate (asterisk: driftwood; full circle: oil spill; vertical line: leaves of Posidonia oceanica); the color indicates the position in the phylogenetic tree (red: Clade 1, Subclade 1a; green: Clade 1, Subclade 1b; purple: Clade 1; blue: Clade 2).
Figure 4.
Growth rate on MEASW of the strains investigated over 28 days. The symbol indicates the isolation substrate (asterisk: driftwood; full circle: oil spill; vertical line: leaves of Posidonia oceanica); the color indicates the position in the phylogenetic tree (red: Clade 1, Subclade 1a; green: Clade 1, Subclade 1b; purple: Clade 1; blue: Clade 2).

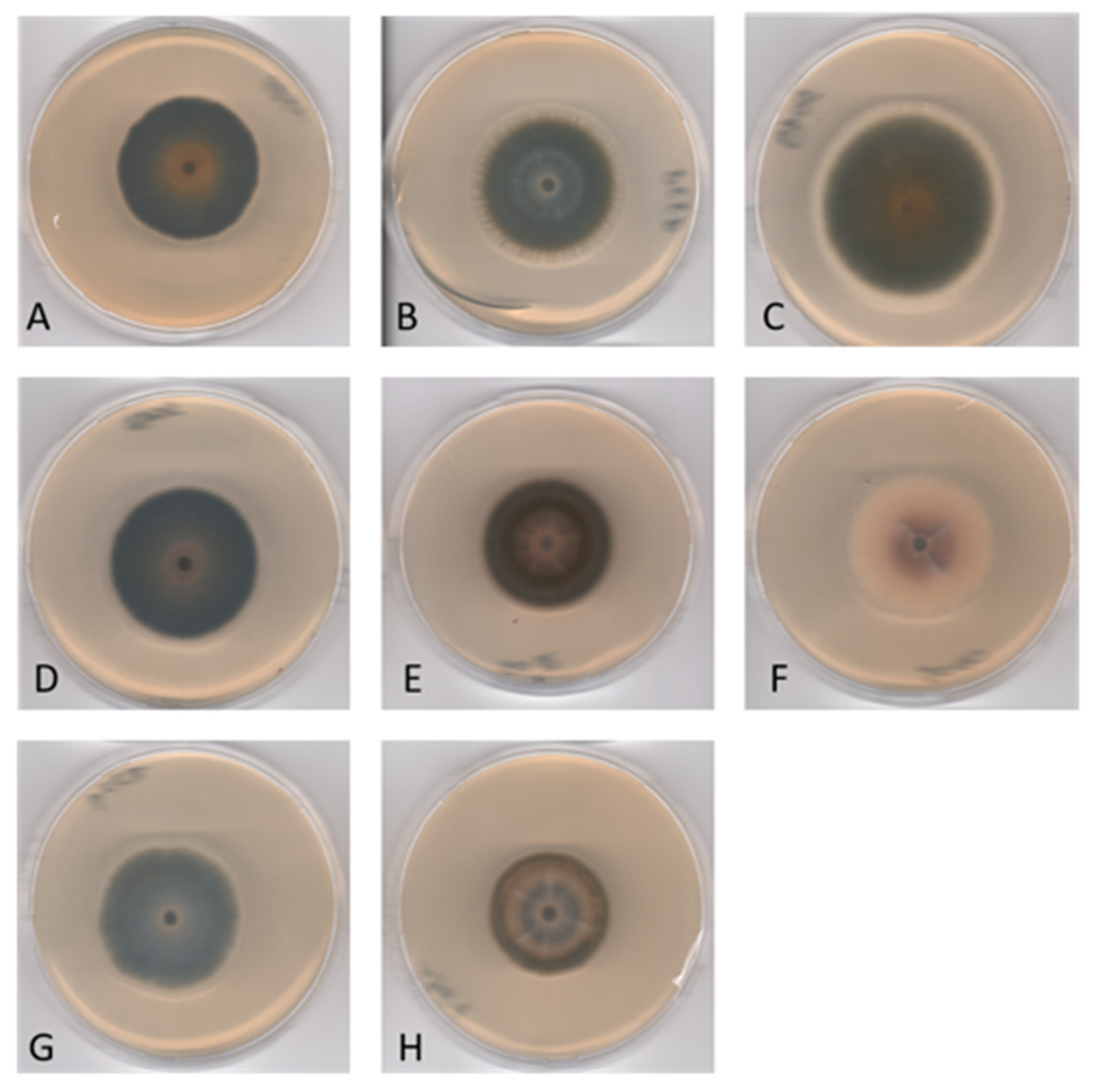
Figure 5.
Corollospora mediterranea sp. nov. 28-days-old colonies at 21 °C on MEASW: (A) MUT 1587, (B) MUT 1938, (C) MUT 5049 (strains belonging to the Clade 1, Subclade 1a); (D) MUT 5048, (E) MUT 5082, (F) MUT 5040 (strains belonging to the Clade 1, Subclade 1b); (G) MUT 1954, (H) MUT 1961 (strains belonging to the Clade 2).
Figure 5.
Corollospora mediterranea sp. nov. 28-days-old colonies at 21 °C on MEASW: (A) MUT 1587, (B) MUT 1938, (C) MUT 5049 (strains belonging to the Clade 1, Subclade 1a); (D) MUT 5048, (E) MUT 5082, (F) MUT 5040 (strains belonging to the Clade 1, Subclade 1b); (G) MUT 1954, (H) MUT 1961 (strains belonging to the Clade 2).

Figure 6.
Corollospora mediterranea sp. nov. reverse of 28-days-old colonies at 21 °C on MEASW: (A) MUT 1587, (B) MUT 1938, (C) MUT 5049 (strains belonging to the Clade 1, Subclade 1a); (D) MUT 5048, (E) MUT 5082, (F) MUT 5040 (strains belonging to the Clade 1, Subclade 1b); (G) MUT 1954, (H) MUT 1961 (strains belonging to the Clade 2).
Figure 6.
Corollospora mediterranea sp. nov. reverse of 28-days-old colonies at 21 °C on MEASW: (A) MUT 1587, (B) MUT 1938, (C) MUT 5049 (strains belonging to the Clade 1, Subclade 1a); (D) MUT 5048, (E) MUT 5082, (F) MUT 5040 (strains belonging to the Clade 1, Subclade 1b); (G) MUT 1954, (H) MUT 1961 (strains belonging to the Clade 2).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Dataset used for phylogenetic analysis. Genbank sequences include newly generated nrITS, nrLSU, nrSSU, RPB1, RPB2, TEF-1α, and βTUB amplicons (in bold) relative to the novel species Corollospora mediterranea.
Table 1.
Dataset used for phylogenetic analysis. Genbank sequences include newly generated nrITS, nrLSU, nrSSU, RPB1, RPB2, TEF-1α, and βTUB amplicons (in bold) relative to the novel species Corollospora mediterranea.
| Species | Strain | Source | nrITS | nrSSU | nrLSU | TEF-1α | RPB1 | RPB2 | βTUB |
|---|---|---|---|---|---|---|---|---|---|
| Microascales | |||||||||
| Halosphaeriaceae | |||||||||
| Corollospora angusta | NBRC 32102 | Sea foam | JN943383 | JN941667 | JN941478 | -- | JN992397 | -- | -- |
| NBRC 32101 T | Seafoam | JN943381 | JN941668 | JN941477 | -- | -- | -- | -- | |
| C. cinnamomea | NBRC 32125 | Beach sand | AB361023 | JN941666 | JN941479 | -- | JN992396 | -- | -- |
| NBRC 32126 | Beach sand | -- | JN941665 | JN941480 | -- | JN992395 | -- | -- | |
| C. colossa | NBRC 32103 T | Seafoam | -- | JN941664 | JN941481 | -- | JN992394 | -- | -- |
| NBRC 32105 | Seafoam | JN943445 | JN941663 | JN941482 | -- | -- | -- | -- | |
| C. fusca | NBRC 32107 T | Seafoam | JN943382 | JN941662 | JN941483 | -- | JN992393 | -- | -- |
| NBRC 32108 | Seafoam | JN943385 | JN941661 | JN941484 | -- | JN992392 | -- | -- | |
| C. gracilis | NBRC 32110 T | Seafoam | -- | JN941660 | JN941486 | -- | JN992390 | -- | -- |
| NBRC 32111 | Seafoam | JN943386 | JN941659 | JN941487 | -- | JN992389 | -- | -- | |
| C. lacera | NBRC 32121 | Sea foam | -- | JN941658 | JN941488 | -- | JN992388 | -- | -- |
| NBRC 32122 | Seafoam | -- | JN941657 | JN941489 | -- | JN992387 | -- | -- | |
| C. luteola | NBRC 31315 T | Sea foam | -- | JN941656 | JN941490 | -- | -- | -- | -- |
| NBRC 31316 | Sargassum giganteifolium | -- | JN941655 | JN941491 | -- | -- | -- | -- | |
| C. maritima | MUT 1652 | Driftwood | MW543050 * | MW543046 * | MW543054 * | MW577242 * | MW577246 * | MW577250 * | MW556316 * |
| MUT 1662 | Submerged wood | MW543051 * | MW543047 * | MW543055 * | MW577243 * | MW577247 * | MW577251 * | MW556317 * | |
| MUT 3408 | Submerged wood | MW543052 * | MW543048 * | MW543056 * | MW577244 * | MW577248 * | MW577252 * | MW556318 * | |
| MUT 3410 | Submerged wood | MW543053 * | MW543049 * | MW543057 * | MW577245 * | MW577249 * | MW577253 * | MW556319 * | |
| Corollospora mediterranea sp. nov. | MUT 1587 | Driftwood | KF915998 | MW584971 * | MW584962 * | MW703375 * | MW645216 * | MW666025 * | MW727528 * |
| MUT 1938 | Oil-contaminated seawater | MW582548 * | MW584966 * | MW584957 * | MW703370 * | MW645212 * | n.d. | MW727523 * | |
| MUT 1950 T | Oil-contaminated seawater | KU935664 | MW584972 * | MW584963 * | MW703376 * | MW645217 * | MW666026 * | MW727529 * | |
| MUT 1954 | Oil-contaminated seawater | KU935659 | MW584973 * | MW584964 * | MW703377 * | MW645218 * | MW666027 * | MW727530 * | |
| MUT 1961 | Oil-contaminated seawater | KU935662 | MW584974 * | MW584965 * | MW703378 * | MW645219 * | MW666028 * | MW727531 * | |
| MUT 5040 | P. oceanica | MW582549 * | MW584967 * | MW584958 * | MW703371 * | MW645213 * | MW666022 * | MW727524 * | |
| MUT 5048 | P. oceanica | MW582550 * | MW584968 * | MW584959 * | MW703372 * | MW645214 * | MW666023 * | MW727525 * | |
| MUT 5049 | P. oceanica | MW582551 * | MW584969 * | MW584960 * | MW703373 * | n.d. | n.d. | MW727526 * | |
| MUT 5082 | P. oceanica | MW582552 * | MW584970 * | MW584961 * | MW703374 * | MW645215 * | MW666024 * | MW727527 * | |
| C. pseudopulchella | NBRC 32113 | Seafoam | -- | JN941651 | JN941495 | -- | JN992383 | -- | -- |
| NBRC 32112 T | Seafoam | -- | JN941652 | JN941494 | -- | JN992384 | -- | -- | |
| C. pulchella | NBRC 32123 | Beach sand | JN943446 | JN941650 | JN941496 | -- | JN992382 | -- | -- |
| NBRC 32124 | Beach sand | -- | JN941649 | JN941497 | -- | JN992381 | -- | -- | |
| C. quinqueseptata | NBRC 32114 T | Sea foam | -- | JN941648 | JN941498 | -- | JN992380 | -- | -- |
| NBRC 32115 | Sargassum sagamianum | -- | JN941647 | JN941499 | -- | JN992379 | -- | -- | |
| Magnisphaera stevemossago | CBS 139776 | -- | KT278691 | KT278704 | -- | -- | KT278740 | -- | |
| Natantispora unipolaris | NTOU3741 | KM624523 | KM624521 | KM624522 | -- | -- | -- | -- | |
| Pileomyces formosanus | BBH30192 | JX003862 | KX686803 | KX686804 | -- | -- | -- | -- | |
| Remispora maritima | BBH28309 | -- | HQ111002 | HQ111012 | -- | -- | -- | -- | |
| Tinhaudeus formosanus | NTOU3805 | KT159895 | KT159897 | KT159899 | -- | -- | -- | -- | |
| Microascaceae | -- | -- | -- | -- | -- | -- | -- | ||
| Cephalotrichum stemonitis | AFTOL-ID 1380 | n.d | -- | DQ836901 | DQ836907 | DQ836916 | -- | -- | -- |
| Microascus trigonosporus | AFTOL-ID 914 | n.d | DQ491513 | DQ471006 | DQ470958 | DQ471077 | DQ471150 | DQ470908 | -- |
| Petriella setifera | CBS 437.75 | Driftwood | -- | DQ471020 | DQ470969 | -- | DQ842034 | DQ836883 | -- |
* = newly generated sequences; n.d. = not determined; T Type Strain. AFTOL = Assembly Fungal Tree Of Life; BBH = BIOTEC Bangkok Herbarium; CBS = Centraalbureau voor Schimmelculture; MUT = Mycotheca Universitatis Taurinensis; NBRC = Nite Biological Resource Centre; NTOU = National Taiwan Ocean University.
Table 2.
Genetic markers, primers, and thermocycler conditions were used in this study.
| Forward and Reverse Primers | Thermocycler Conditions | References | |
|---|---|---|---|
| ITS | ITS1-ITS4 | 95 °C: 5 min, (95 °C: 40 s, 55 °C: 50 s, 72 °C: 50 s) × 35 cycles; 72 °C: 8 min; 4 °C: ∞ | [35] |
| LSU | LR0R-LR7 | 95 °C: 5 min, (95 °C: 1 min, 50 °C: 1 min, 72 °C: 2 min) × 35 cycles; 72 °C: 10 min; 4 °C: ∞ | [36] |
| SSU | NS1-NS4 | 95 °C: 5 min, (95 °C: 1 min, 50 °C: 1 min, 72 °C: 2 min) × 35 cycles; 72 °C: 10 min; 4 °C: ∞ | [35] |
| TEF-1α | EF-dF/EF-2218R | 95 °C: 5 min, (95 °C: 1 min, 50 °C: 1 min; 72 °C: 2 min) × 40 cycles, 72 °C: 10 min; 4 °C: ∞ | [37] |
| βTUB | Bt2a-Bt2b | 94 °C: 4 min, (94 °C: 35 sec, 58 °C: 35 s, 72 °C: 50 s) × 35 cycles; 72 °C: 5 min; 4 °C: ∞ | [38] |
| RPB1 | RPB1Af-RPB1Cr | 96 °C: 5 min, (94 °C: 30 s, 52 °C: 30 s, 72 °C: 1 min) × 40 cycles; 72 °C: 8 min; 4 °C: ∞ | [39] |
| RPB2 | fRPB2-5F/fPB2-7cR | 94 °C: 3 min, (94 °C: 30 s; 55 °C: 30 s; 72 °C: 1 min) × 40 cycles, 72 °C: 10 min; 4 °C: ∞ | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Poli, A.; Bovio, E.; Perugini, I.; Varese, G.C.; Prigione, V. Corollospora mediterranea: A Novel Species Complex in the Mediterranean Sea. Appl. Sci. 2021, 11, 5452. https://doi.org/10.3390/app11125452
AMA Style
Poli A, Bovio E, Perugini I, Varese GC, Prigione V. Corollospora mediterranea: A Novel Species Complex in the Mediterranean Sea. Applied Sciences. 2021; 11(12):5452. https://doi.org/10.3390/app11125452
Chicago/Turabian StylePoli, Anna, Elena Bovio, Iolanda Perugini, Giovanna Cristina Varese, and Valeria Prigione. 2021. "Corollospora mediterranea: A Novel Species Complex in the Mediterranean Sea" Applied Sciences 11, no. 12: 5452. https://doi.org/10.3390/app11125452
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.