
Exploring the Potential Applications of Paecilomyceslilacinus 112
by
 , , , and
, , , and
Mariana Constantin
1,2, 
Iuliana Raut
1,
Ana-Maria Gurban
1,*,
Mihaela Doni
1,
Nicoleta Radu
1,3,
Elvira Alexandrescu
1 and
Luiza Jecu
1,* 1
National Institute for Research & Development in Chemistry and Petrochemistry—ICECHIM, Biotechnology Department, Spl. Independentei 202, 060021 Bucharest, Romania
2
Faculty of Pharmacy, Titu Maiorescu University, 16 Bd. Gh. Sincai, 040441 Bucharest, Romania
3
Faculty of Biotechnology, University of Agronomic Sciences and Veterinary Medicine of Bucharest, 59 Mărăşti Boulevard, 011464 Bucharest, Romania
*
Authors to whom correspondence should be addressed.
Appl. Sci. 2022, 12(15), 7572; https://doi.org/10.3390/app12157572
Submission received: 3 June 2022
/
Revised: 25 July 2022
/
Accepted: 25 July 2022
/
Published: 27 July 2022
(This article belongs to the Topic Advances in Environmental Biotechnology (AEB))
Abstract
:Microorganisms are widely used to obtain biostimulants that can facilitate the assimilation of nutrients, ensuring high crop yield and quality. A particular category of biostimulants are protein hydrolyzates (PH), obtained from microbial cultures grown on a nutrient medium. In the present study, Paecilomyces lilacinus 112, an endophytic fungus isolated from soil, was tested to determine its effect on the growth promotion of tomato seedlings in greenhouse conditions. Additionally, other beneficial features of the P. lilacinus isolate were evaluated via several tests: antagonism against plant pathogenic fungi, production of secondary useful metabolites, and solubilization of vital micronutrients. Out of the tested pathogens, P. lilacinus exhibited the highest antifungal activity against a Cladosporium isolate (inhibition of 66.3%), followed by Rhizoctonia. solani (52.53%), and Sclerotinia sclerotiorum (50.23%). Paecilomyces lilacinus 112 was able to secrete hydrolytic enzymes and siderophores, and solubilize zinc and phosphorus. In the tomato treatment, the application of PH obtained from fungal cultivation on a feather medium led to the following increases in plant growth parameters: 3.54-fold in plant biomass; 3.26-fold in plant height, 1.28-fold in plant diameter; 1.5-fold in the number of branches/plant; and 1.43-fold in the number of leaves/plant, as compared to water treatment. The application of this isolate can be of benefit to bioeconomy because keratin wastes are valorized and returned, in agriculture, contributing to renewable natural resources.
1. Introduction
Sustainable agriculture is achieved through reducing the consumption of fertilizers and agrochemicals, and replacing them using biological solutions as viable alternatives. The public opinion is exerting increasing pressure on governments to impose severe restrictions and regulations on the use of chemical compounds in agriculture. In addition to ecological and health considerations, it is necessary to capitalize on the natural, renewable, and widely accessible resources, and to reduce the consumption of conventional energy resources.
Biostimulants can facilitate the assimilation of nutrients, ensuring high crop yield and quality, and increasing the tolerance of the crop to abiotic stress. Biostimulants contain a large diversity of components, plant hormones, amino acids, betaines, peptides, proteins, carbohydrates, lipids, vitamins, nucleotides and nucleosides, humic substances, beneficial elements, phenolic compounds, and sterols. Microorganisms, such as fungi, bacteria, and yeasts, are widely used to obtain biostimulants, and living and/or non-living microorganisms, as well as various metabolites, could be included in these biostimulants. In general, the mode of action of biostimulants is still under analysis, due to the complexity and diversity of the source and extraction methods used. Biostimulants are obtained from raw materials of highly diverse origin through various technologies; their biological activity is complex and highly diverse, thus their application affecting the level of safety [1]. At present, the attention of scientists is focused on the elucidation of the biological basis of biostimulant function to ensure safe and widespread applications of these formulations in agriculture [2].
Protein hydrolyzates (PH) are a particular category of biostimulants, representing a mixture of polypeptides, oligopeptides, and free amino acids, resulting from chemical or enzymatic hydrolysis of plants or animal tissues. Additionally, protein hydrolyzates can be obtained from the cultivation of several fungal species on a suitable medium containing agroindustrial byproducts as carbon and energy sources. The microbiologically obtained keratin hydrolyzates are very useful, and several studies have reported their significant role in abiotic stress responses of plants [3,4,5].
Good starting materials to obtain protein hydrolyzates are keratin wastes, which are byproducts derived from commercial poultry processing plants, the leather industry, the wool and textile industry, and slaughterhouses. The main component is keratin, a rigid structural protein stabilized by a high degree of crosslinking by disulfide bonds, hydrophobic interactions, and hydrogen bonds.
Some applications of feathers have also been reported, ranging from composites, fibers, nano- and microparticles, electronic devices, but no major products on a large industrial scale [6,7,8,9]. For the same purpose, the exploitation of other types of keratin residues is worth investigating. Significant amounts of wool is generated in a number of different industries, including tanning and meat processing. Wool has limited use as a consequence of its lack of solubility and resistance to degradation, and it is considered a byproduct requiring disposal. Wool surface contains wax and grease, and also several impurities, such as inorganic mineral dirt and vegetable debris; thus, an expensive washing cycle is needed. There have been several proposed uses of wool, including filling materials for panels and boards, adsorption of heavy metals and soil amendment, but these are not yet applicable at the industrial level [10,11].
Our approach is based on the ability of fungal species to transform keratin wastes into a mixture of nitrogen and sulfur compounds with potential to be used as plant biostimulants. It is therefore a possible alternative to effectively reduce waste quantities and to obtain valuable products, meeting the demands of economic and ecological interest in the use of renewable resources, and reducing the use of conventional resources [12,13,14].
A potential opportunity to develop biobased commercial products for future agriculture is the use of endophytes and their secreted secondary metabolites. Since endophytes synthesize a large diversity of secondary metabolites (e.g., terpenoids, alkaloids, phenylpropanoids, aliphatic compounds, polyketides, and peptides), they are considered a “reservoir” of biological resources, with various applications in medicine, industry, and agriculture [15,16]. Among the endophytes is the genus Paecilomyces, a common saprobic filamentous fungi, which is a source of bioactive natural products with antimicrobial, antiviral, antitumoral, herbicidal, insecticidal, and cytotoxic activities [17].
The performed study aimed to highlight: (1) the properties of this microorganism from various points of view, as a stimulant of plant growth, a biocontrol agent against pathogens, and a producer of secondary useful metabolites (hydrolytic enzymes and siderophores), as well as the implication for solubilization of vital mineral micronutrients, such as phosphorus and zinc; (2) the effect of spent culture media, which contain protein hydrolyzates (PH), obtained from the submerged culturing of Paecilomyces lilacinus 112, using culture media formulated with different wastes (feathers or wool) on Lycopersicum esculentum.
2. Materials and Methods
2.1. Microorganisms
Paecilomyces lilacinus 112, a soil isolate from the Microbial Collection of Biotechnology Department of National Institute of Chemistry and Petrochemistry R&D of ICECHIM Bucharest, was used in the experimental studies. The microorganism was maintained on potato dextrose agar medium (PDA, Scharlau), and prior to use, their purity was controlled. The composition of the PDA medium was (g∙L−1): 4, peptone; 20, glucose, 15, agar; final pH = 5.6 at 25 °C.
2.2. Keratin Substrates (Feathers and Wool)
Chicken feathers (from local poultry) and wool (low-quality wool from sheep farming) were cleaned, sterilized with 3% ethanol, washed vigorously, and dried at 60 °C. The feathers and wool fibers were cut into small pieces for fungal cultivation. In order to be used as keratinase substrate in the enzymatic assay, the feathers were grounded until they became a fine powder [18].
2.3. Fungal Cultivation for Protein Hydrolyzate (PH) Preparation
Cultivation in liquid medium was performed in agitated Erlenmeyer flasks, on a rotary incubator shaker (Heidolph Unimax 1010) at 26 ± 2 °C for 21 days. The composition of the basal medium was as follows (g∙L−1): 0.1, KH2PO4; 0.1, CaCl2; 0.1, FeSO4 • 7H2O; 0.005, ZnSO4 • 7H2O. The medium contained a 1% (w/w) keratin source (wool or feathers). The control was considered the basal medium. The Erlenmeyer flasks were inoculated with 5 mL of fungal culture. Furthermore, after incubation, the culture broth was filtered through a 0.2 µm syringe, and supernatants were tested in experiments.
2.4. Antagonism versus Plant Pathogenic Fungi
The antagonism of Paecilomyces lilacinus 112 versus pathogens was tested using the dual culture technique on PDA medium in Petri plates [19]. The following pathogens, purchased from the German Collection of Microorganisms and Cell Cultures (DSMZ) (Braunschweig, Germany), were tested: Botrytis allii DSMZ 62081, Rhizoctonia solani DSM 2284, Fusarium graminearum DSM1095, and Sclerotinia sclerotiorum DSM 1946. In addition, two more isolates from the Microbial Collection of Biotechnology Department of National Institute of Chemistry and Petrochemistry R&D of ICECHIM Bucharest, Cladosporium sp. T2 and Alternaria alternata were also tested. Briefly, PDA medium poured into sterile Petri plates was incubated with a 5 mm culture disc of Paecilomyces lilacinus and each pathogen. Antagonist activity was observed after incubation at 25 ± 1 °C, for 3–5 days. Antagonist activity, expressed as inhibition range, was determined using the formula:
where I = inhibition; M = colony diameter for pathogen control; P = colony diameter of pathogen in dual cultures in presence of antagonist. For the accuracy of the assays, three independent experiments were carried out. Microscopy observations were obtained using an Olympus BX 51 microscope and a Quanta FEI 200 scanning electron microscope (SEM).
I (%) = (M − P)/M × 100
The photos were taken at the point of contact between the isolates of antagonist Paecilomyces and each phytopathogen.
2.5. Enzymatic Activities
Chitinase activity was evaluated using a qualitative method, with colloidal chitin as a carbon source [20]. The Petri plates with suitable agar medium were inoculated with 5 mm culture disc of fungal isolate and incubated at 26 ± 2 °C for 7 days. The appearance of the red-violet color on medium containing bromocresol purple (Sigma-Aldrich, St. Louis, MI, USA) was considered a positive result. Three replications were achieved.
Cellulase activity was evaluated by qualitative method with carboxymethylcellulose (CMC) as cellulosic substrate [21]. The solid medium exhibited the following composition (g∙L−1): 1, yeast extract; 5, CMC; 15, agar. The Petri plates were inoculated with 5 mm culture disc of fungal isolate and incubated as in the chitinase assay. After the incubation, the plates on agar suitable medium were stained with Lugol’s solution (0.1%) for 15 min at 26 ± 2 °C. The formation of a transparent circular area around the fungal colony was considered a positive result. Three replicates were achieved.
Keratinase activity was evaluated via a qualitative method, with feather powder as the enzymatic substrate [22]. Chicken feathers were disinfected with 70% (v/v) ethylic alcohol, rinsed with sterile distilled water, dried overnight at 50 °C, and finally, milled in a ball mill (Retsch, model MM400, Haan, Germany). The solid medium possessed the following composition (g∙L−1): 10, feather powder; 0.1, KH2PO4; 0.01, CaCl2; 0.1, FeSO4 • 7H2O; 0.005, ZnSO4 • 7H20; pH = 7. The Petri plates were inoculated with 5 mm culture disc of fungal isolate and incubated as in the chitinase assay. The formation of a transparent circular area around the fungal colony was considered a positive result. Three replications were achieved.
2.6. Analysis of Plant Growth-Promoting Characteristics
The fungal isolate was tested in vitro for its zinc solubilizing ability using a plate assay [23]. The Petri plates were inoculated with 5 mm mycelial fragments from fresh cultures, and incubated in the dark at 26 ± 2 °C for 7 days to observe a clear halo zone around colonies. Three replications were achieved.
Phosphate solubilization was determined qualitatively on solid medium with Ca3(PO4)2 and bromocresol purple (Sigma-Aldrich) [24]. The strain was cultured in a medium with the following composition (g∙L−1): 0.3, MgSO4 • 7H2O; 0.004, MnSO4 • 7H2O; 0.002, FeSO4 • 7H2O; 20, NaCl; 0.5, yeast extract; 0.1, bromocresol purple as pH indicator; 5.0, Ca3(PO4)2; 16, agar; pH = 7.0. The phosphate solubilization was confirmed by the formation of a clearing zone around the colonies. The experimental protocol was similar to that of zinc solubilization. Three replications were achieved.
The qualitative test for siderophores production was carried out with the following culture medium (g∙L−1): 30, malt extract; 5, peptone; 0.05, 8- hydroxyquinoline; 15, agar [25]. The solid medium was inoculated with 5 mm mycelium fragment from fresh culture. The plates were incubated for 5 days, at 26–28 °C. The result was considered positive when the strain was able to grow on the culture medium containing 8-hydroxyquinoline.
Analysis of effects of protein hydrolyzates derived from P. lilacinus 112 under growth chamber conditions were carried out with Lycopersicum esculentum seeds (Balkonzauber-AS, purchased from Plant-shop.ro, Sacele, Romania). The seeds were planted in conic plastic cups (height = 9.0 cm; top diameter = 6.5 cm; bottom diameter = 4.0 cm) and incubated in a growth chamber (Micro Clima Series TM, Labs Economic Lux chamber, Snijders, The Netherlands) under controlled conditions, as follows: humidity approximately 69%; 26 °C as day temperature and 22 °C as night temperature; 10,000 Lux; day/night cycle as 16 h per day/8 h per night. The substrate for seedlings (100% natural, purchased commercially) presented the following physical–chemical properties: 66.67%, organic matter; 1.364%, nitrogen; 100 ppm, phosphorus; 140 ppm, potassium; 65%, humidity. Each plastic cup containing one seed was treated with 1 mL of filtrate from protein hydrolyzates, the liquid culture on keratin substrate after filtration with 0.2 µm filter. Fifty pots (plastic cups) were prepared for each variants tested. The treatment was applied by soil drenching over a period of 21 days, at weekly intervals. The control plants were treated with water. Additionally, a mineral medium (composition presented in Section 2.3) was used for the treatment of corresponding pots. After 21 days and four treatments, seedlings were collected and growth parameters (biomass, plant height and diameter, number of branches/plant, number of leaves/plant) were measured.
2.7. Soluble Protein Content of Protein Hydrolyzates
The Lowry method was used to determine the soluble protein content of the hydrolyzates [26]. Under alkaline conditions, copper complexes with proteins from solution were formed. When Folin phenol reagent (phospho-molybdic-phosphotungstic reagent) is added, it binds to proteins. The bound reagent is then slowly reduced and changes color from yellow to blue. The intensity of the blue color formed is directly proportional to the protein concentration of the solution. Using different aliquots of bovine serum albumin (Sigma-Aldrich) with known concentrations obtained from the standard curve, the unknown protein concentration of the sample was measured spectrophotometrically at 660 nm.
2.8. Scheme Used for Plant Treatments
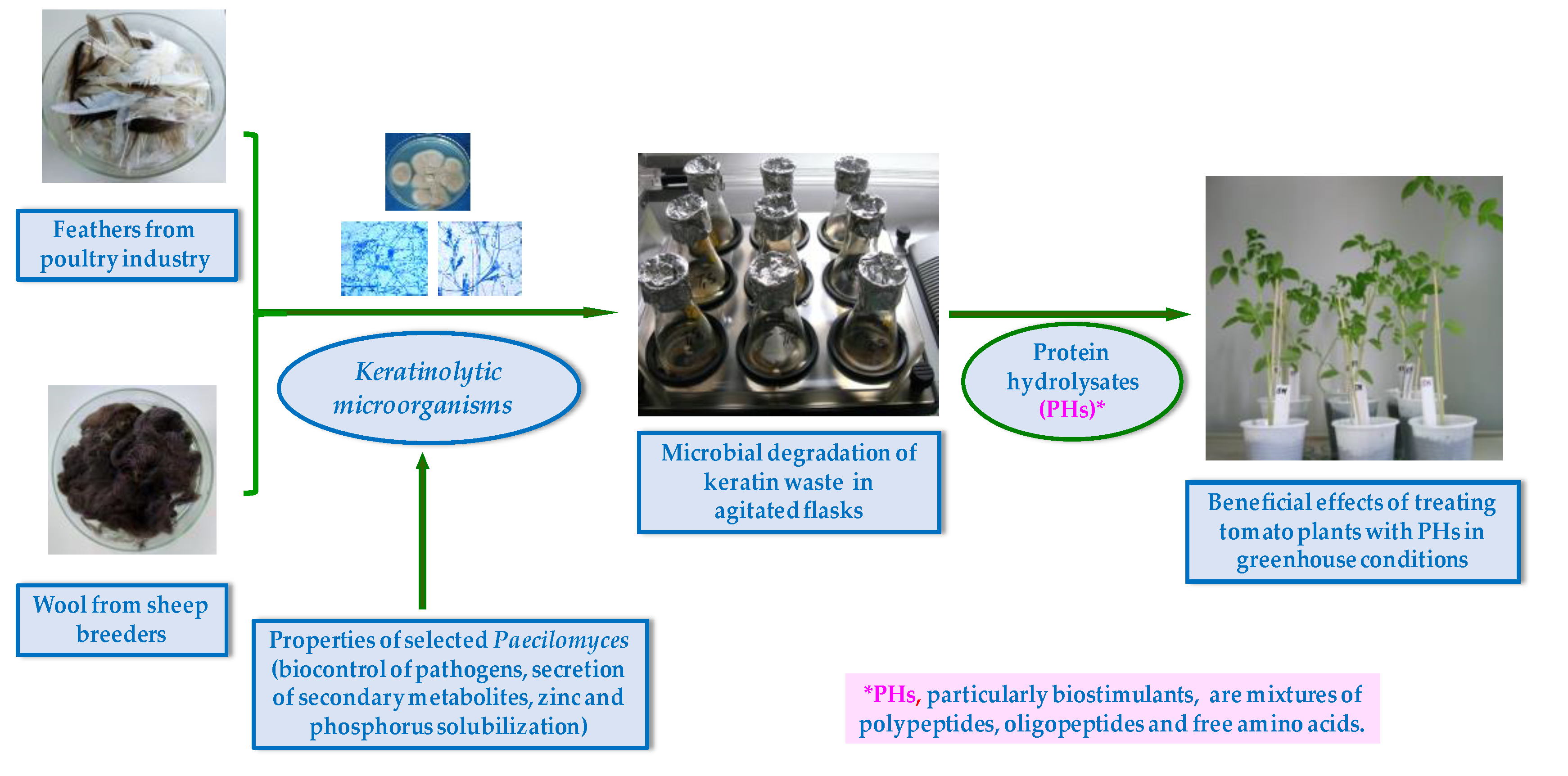
The experimental protocol using protein hydrolyzates (PHs) from Paecilomyces lilacinus 112 as a plant growth-promoting agent is presented in Figure 1. In summary, the process comprised the following steps: preparation of keratinous substrate (wool, feathers); preparation of Paecilomyces culture; fungal cultivation under optimum conditions (incubation, temperature, pH, agitation, mineral composition, keratin substrate concentration); obtainment of protein hydrolyzates; preparation of tomato seedlings and substrate for plant growth test; tomato seedling cultivation in growth chamber under controlled conditions; application of treatment with PHs; measurement of plant growth parameters, according to experimental protocol. In addition, it is worth mentioning the investigation of the fungal strain to highlight its characteristics as a plant biostimulant.
2.9. Statistical Analysis
The data obtained from experiments performed on Lycopersicum esculentum were analyzed with GraphPad Prism 5.0 software, with a confidence of 95%, by comparing experimental variants A02 and A03 with control (untreated) plants. Notations used: * p ≤ 0.05; ** 0.05% < p ≤ 0.01; *** 0.01 < p ≤ 0.001. Variant A02 represents the effects produced on plants by treatment with PH obtained from fungal culture on feather medium, and variant A03 describes the effects produced by PH obtained from fungal culture on wool medium.
3. Results
3.1. Antagonism against Plant Pathogenic Fungi
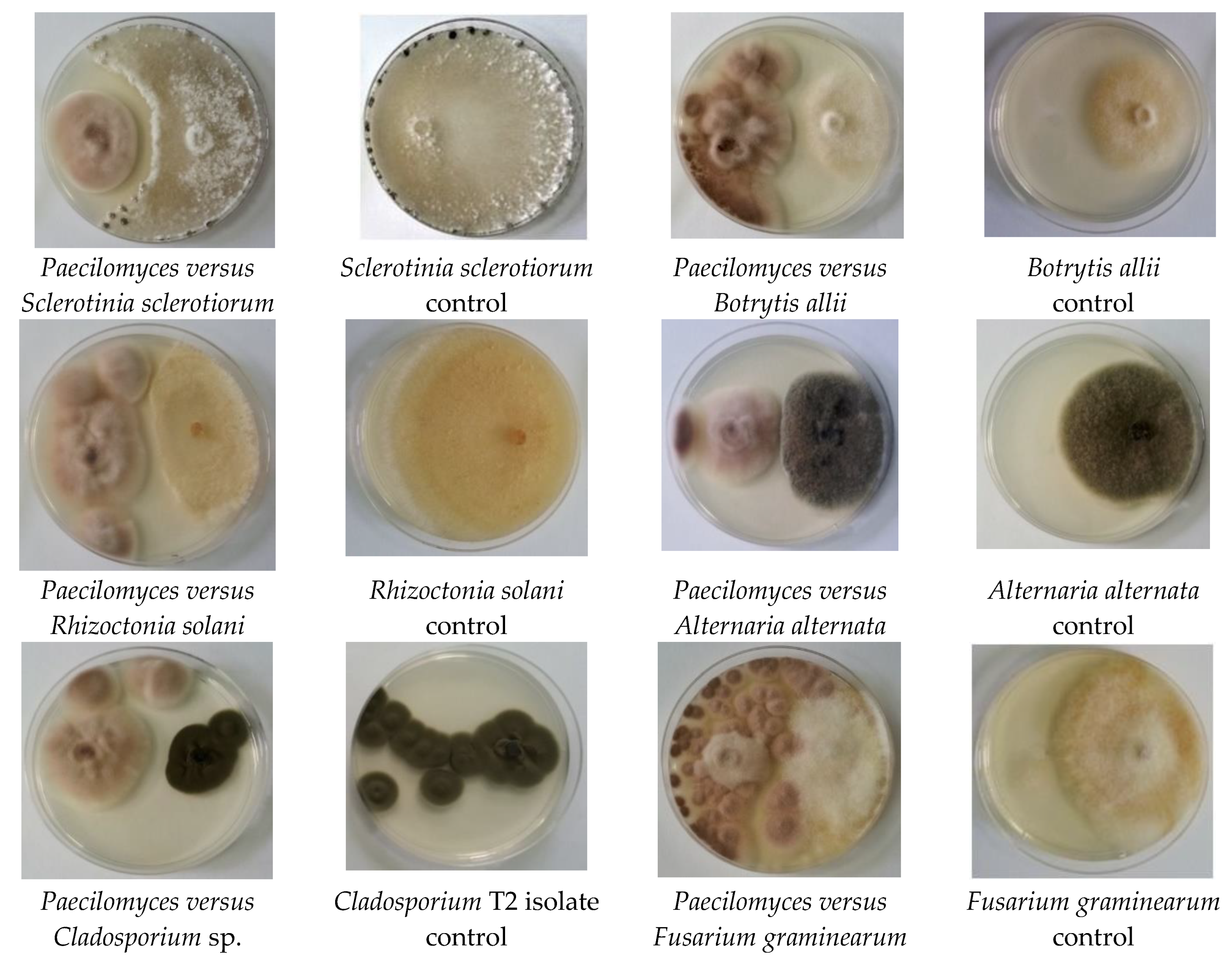
The antagonistic activity of Paecilomyces lilacinus 112 against plant pathogenic fungi was evaluated using the dual culture technique. The images of Petri plates with fungal cultures on a solid medium are presented in Figure 2. The values of the inhibition diameters are presented in Table 1. The findings reveal that Paecilomyces lilacinus 112 exhibited the highest antifungal activity against the Cladosporium T2 isolate (inhibition of 66.3%), followed by R. solani (52.53%), and Sclerotinia sclerotiorum (50.23%). The lowest activity was found against A. alternata and B. allii.
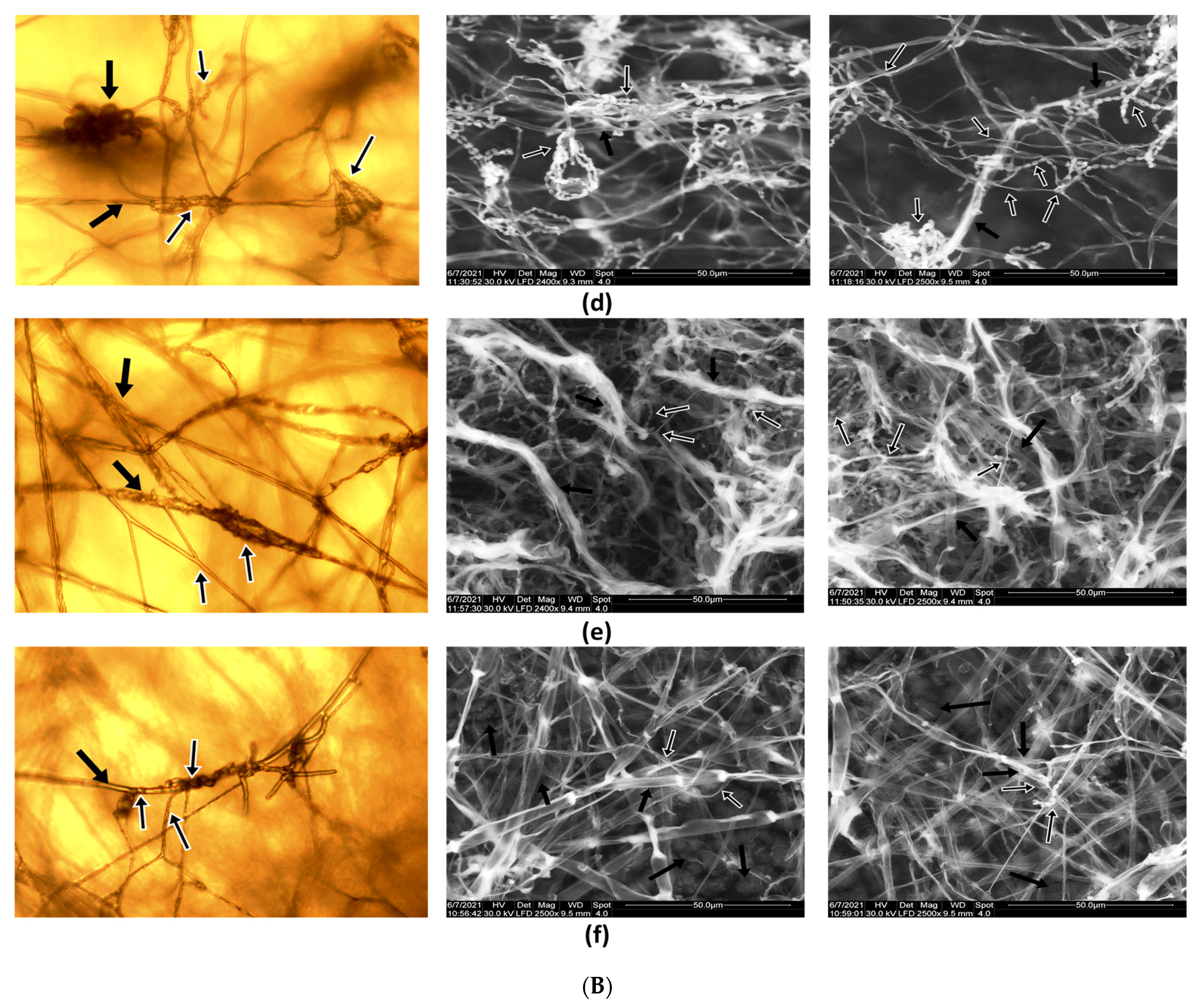
Microscopic observations of the interactions between Paecilomyces lilacinus 112 and plant pathogenic fungi are presented in Figure 3A,B. Alterations in the pathogen hyphae can be observed, for example, in R. solani, S. sclerotiorum, and B. allii..
The degree of in vitro invasion, covering, and pathogen mycelium destruction analyzed by the microscopic observations of the interaction between Paecilomyces lilacinus and the phytopathogens showed different stages of the mycoparasitism process as a mechanism of antagonism, such as dense coils of hyphae that tightly encircle (surround) the phytopathogens hyphae (Figure 3(Aa), and penetrating a hypha followed by mycolysis of the phytopathogen hyphae (Figure 3(Ac). These changes in the structure of the pathogen are also supported by the results obtained in the enzymatic tests, more specific by the secretion of hydrolytic enzymes by antagonistic Paecilomyces lilacinus.
In Figure 3B, it can be observed that to a certain degree, the mechanisms involved in biological control exercised by antagonistic Paecilomyces lilacinus to suppress the growth of pathogens; for example, mycoparasitism and competition for nutrients, represented by entangled hyphae of the Paecilomyces lilacinus on the surface of hyphae formed by phytopathogens.
3.2. Secretion of Hydrolytic Enzymes
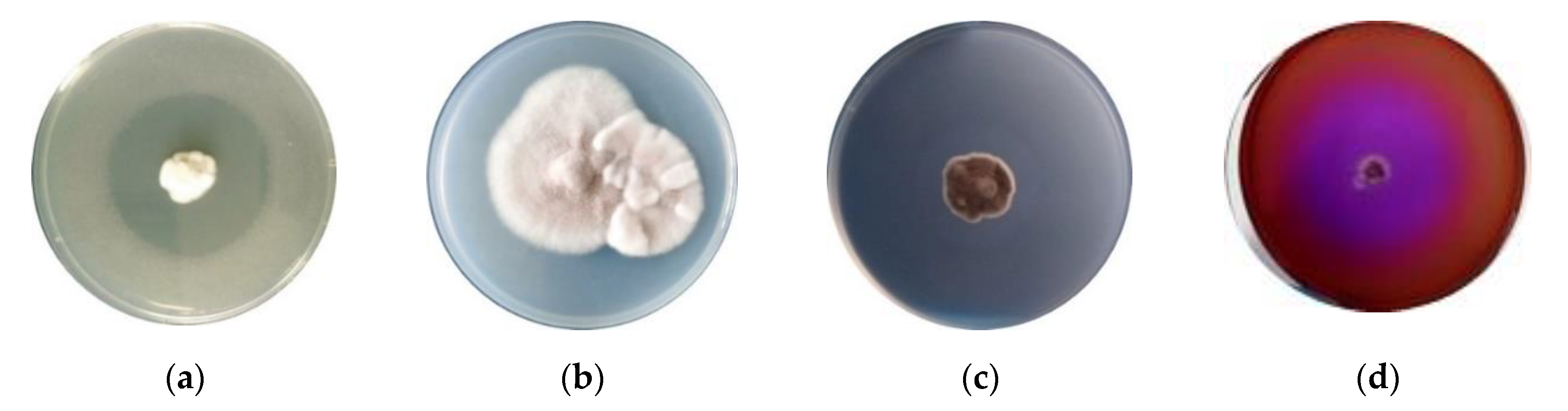
The production of hydrolytic enzymes by Paecilomyces lilacinus 112 was qualitatively determined using a qualitative method, whereas enzyme production was tested on solid agar media (halo on cup plate assay). Minimal appropriate medium was used. An enzymatic halo, referred to as a “degradation halo”, was used as an indicator of the presence of enzyme activity produced by fungal isolate through the culture medium. The appearance of a clear zone around the fungal colony indicated the degradation of carbon substrate due to enzyme activity (Figure 4a–c). Accordingly, the fungal isolate Paecilomyces lilacinus 112 showed a good level of hydrolytic enzymes (as chitinases, cellulases, and keratinases), which is a beneficial feature of microorganisms used as plant biostimulants.
3.3. Plant Growth-Promoting Characteristics
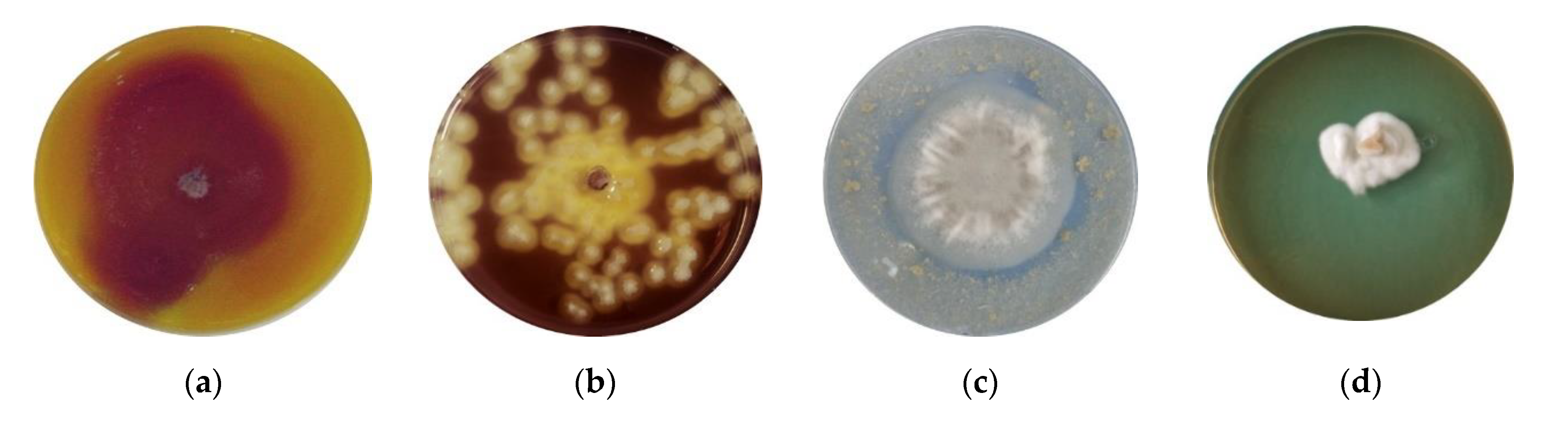
Siderophore production of Paecilomyces lilacinus 112 was determined on 8-hydroxychinoline medium, by incubating fungal mycelial disc of active culture. Figure 4d presents the positive results of a qualitative test, revealed as the growth of fungal isolate on a specific medium.
The results of the studies regarding the solubilization of zinc and phosphorous by Paecilomyces lilacinus are presented in Figure 5. Zinc solubilization was tested using diverse compounds, such as ZnO, ZnNO3 and ZnSO4 (Figure 5a–c). A clear zone of solubilization was obtained for ZnO, while the solubilizing capacity was lower for ZnNO3 and ZnSO4.
Phosphate solubilization by Paecilomyces lilacinus 112 from Ca3(PO4)2 was demonstrated by the formation of transparent zone surrounding fungal colonies (Figure 5d).
3.4. Soluble Protein of Fungal Hydrolyzates
The results of the Lowry assay applied to protein hydrolyzates are presented in Table 2. It can be observed that the amount of soluble protein in hydrolyzates from cultivation on medium with feathers is significantly higher compared to that obtained on medium with wool.
3.5. Effects of PH on Tomato Seedlings
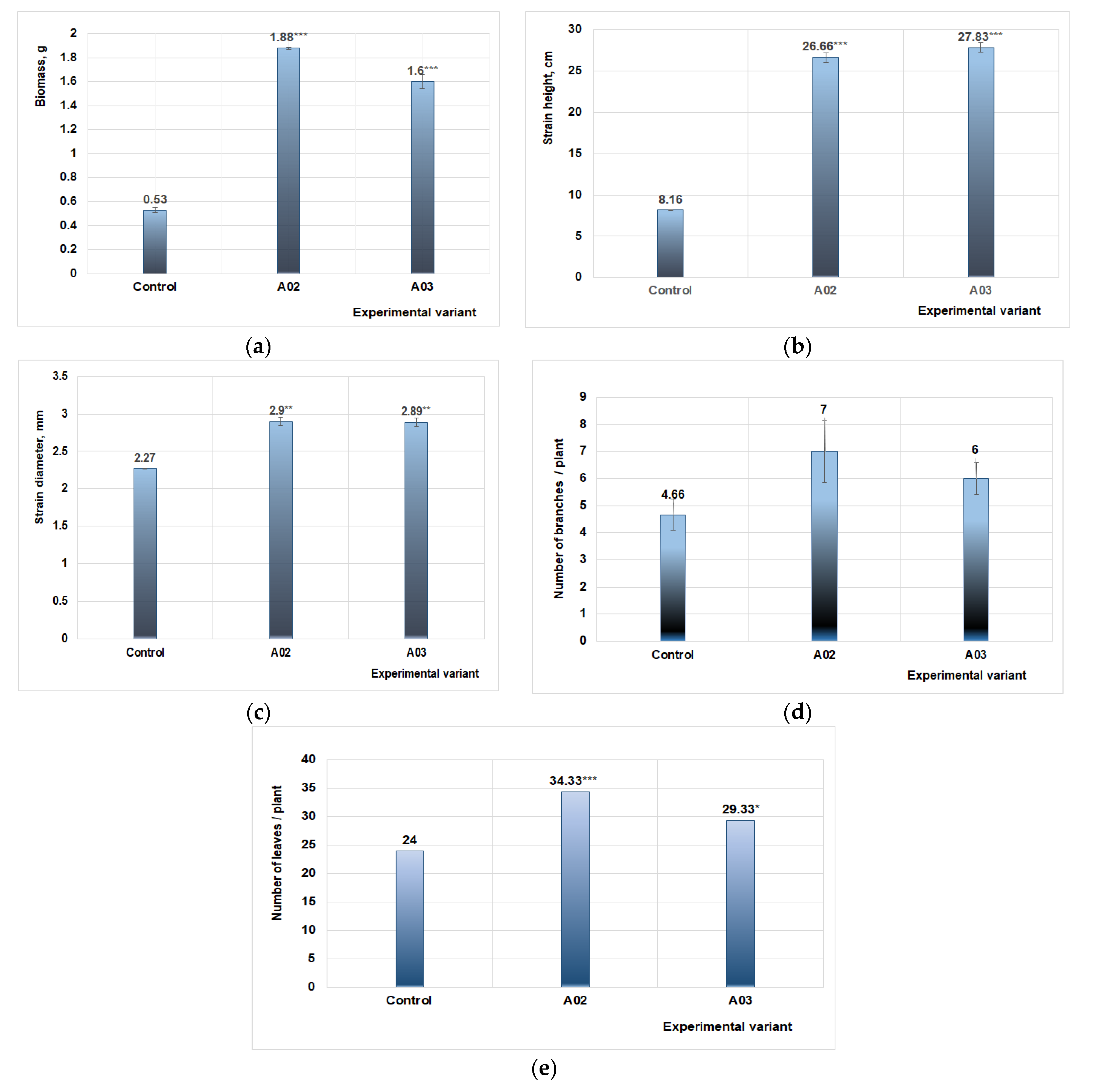
The studies performed on plant treatments with protein hydrolyzates from fungal culture on medium with keratin waste (wool and feathers) showed that they had a positive effect on plant growth and development. The images where this was most distinct were chosen to present the growth of tomato seedlings (Figure 6). The growth parameters, biomass, plant height and diameter, number of branches/plant, number of leaves/plant, were measured after four applications of PHs (Table 3).
A significant increase in plant biomass was obtained, with values three times higher compared to the control. Additionally, positive results were obtained from the number of branches/plant, indicating an increase of 33% using PHs from feather medium, and of 29% for PHs from wool medium, respectively (Figure 6).
Regarding the plant diameter, the obtained values for both cultures on keratin media were similar, with an increase of 1.28-fold and 1.27-fold, for PH from feather medium and wool medium, respectively. To some extent, a more significant difference between values for experimental keratin medium variants was obtained for the number of leaves per plant, an increase of 1.43-fold for feather medium, and only a 1.22-fold increase for wool medium. All the above-described increases are reported to control the value for each growth parameter.
3.6. Statistical Analysis
The data from the plant analysis showed that the best results were obtained by applying the treatment with PH from fungal culture grown on the feather medium. The values of growth parameters were highly significant compared to the control, while the effect on plant diameter presented only moderate statistically values (Figure 7).
The measurements show that the number of branches generated by the tomato plants under study varies in relation to the control variant (Figure 7d). The best results were obtained for variant A02, representing the effects on plants produced by treatment with PH from fungal culture on feather medium, followed by variant A03, which indicated effects produced by PH from fungal strain cultured on wool medium.
4. Discussion
There is a growing interest in the more efficient operation of fungal endophytes Paecilomyces for beneficial activities in the agricultural domain. Comprehensive reports underline the importance of studies dedicated to Paecilomyces sp., fungi with huge potential for development in various applications based on the chemical diversity and biological activity of secondary metabolites secreted [17,27].
The first aspect studied was the antagonism expressed versus plant pathogens. In general, the biocontrol activity of Paecilomyces is well documented, with the members of the Paecilomyces genus acting through direct mechanisms, parasitism, competition or antibiosis, and indirect, involving plant protection through induced systemic resistance mechanism. There are studies that show the effectiveness of Paecilomyces against different species of phytopathogenic bacteria, such as Xanthomonas campestris [28] and Pectobacterium carotovorum [29], but mainly against phytopathogenic fungi Alternaria solani [30], Fusarium oxysporum [31], Sclerotium rolfsii [32], Macrophomina phaseolina, Pythium aphanidermatum, R. solani, etc. Moreover, many studies are dedicated to biological control against nematodes, especially Meloidogyne spp. [33,34]. In a recently published comprehensive review, it is argued that the taxonomic study on Paecilomyces sp. as biological control agent against bacteria, phytopathogenic fungi, nematodes, and numerous pests will provide a significant contribution to the obtainment of microbial formulations useful in agriculture [35,36]. For example, in support of this assertion, an ethanol extract of the endophytic fungus Paecilomyces variotii (trade name ZhiNengCong (ZNC)) was presented as an efficient biocontrol agent [37].
In the present study, the criterion for the selection of the pathogenic was their aggressiveness and notoriety. Hence, F. graminearum [38], B. allii [39], and R. solani [40] are known to be pathogenic fungi causing diseases in a large number of economically important plants. In addition, Cladosporium sp. [41] and Alternaria alternata [42] from The Microbial Collection of Biotechnology Department from National Institute of Chemistry and Petrichemistry R&D ICECHIM of Bucharest present characteristics and behavior of pathogens, infecting the aerial parts of plants. In our study, the highest antagonistic activity of Paecilomyces lilacinus 112 was expressed against the Cladosporium T2 isolate. It is known that Cladosporioides are widely distributed, infecting fruits (mandarin, cantaloupe, pomegranate) and vegetables (especially tomato) [43], causing economic losses in agricultural crops. Taking into account all these possible damages produced by the Cladosporium genus, it can be underlined that Paecilomyces lilacinus 112 can be useful as a biocontrol agent of the Cladosporium pathogen. In addition, we have obtained a 52.53% inhibition of R. solani, which can be considered a better result compared with other Paecilomyces strains, such as P. marquandii, Paecilomyces sp. AE-3, and Paecilomyces sulphurellus, which produced inhibition levels of 39%, 50%, and 21%, respectively [44]. Moreno-Gaviria et al. [37] demonstrated the high capacity of P. variotii CDG33 to reduce the severity of diseases caused by different aerial and soil phytopathogens Pythium aphanidermatum (81.5%), Fusarium solani (72%), Botrytis cinerea (51%), Sclerotinia sclerotiorum (40%), and Rhizoctonia solani (28%). By comparison, our Paecilomyces lilacinus 112 was much more active against Rhizoctonia solani (52.53% inhibition) and Sclerotinia sclerotiorum (50.23%), but less active against pathogens belonging to genres Botrytis and Fusarium.
Filamentous fungi produce a wide array of secondary metabolites; among them, siderophores play a significant role in promoting the growth of several plants by increasing Fe uptake in plants [45,46,47]. Siderophores are able to capture insoluble ferric ions (Fe+3) from different habitats and transport them into cell membranes through specific receptors. Given the importance of siderophore production, in our experimental study we evaluated the ability of Paecilomyces lilacinus 112 to secrete these secondary metabolites. The qualitative test performed by culturing Paecilomyces lilacinus 112 on 8-hydroxyquinoline-based medium presented a positive result, which is a good starting point for further application in the agriculture field. Similar studies showed the production of the siderophore by a P. variotii isolate [35]. Furthermore, a Paecilomyces variotii produces siderophores, mostly ferrirubin, a member of the class of ferrichrome iron (III) chelates [48,49].
Phosphorus (P) is one of the most important minerals for plant growth. In most cases, phosphorus deficiency in the soil is alleviated by the addition of phosphate fertilizers; however, sometimes there are problems with the fixation of added P from chemical fertilizers. The soil-fixed P can be solubilized through the activity of microorganisms, namely fungi or bacteria, in connection with certain factors, such as the mechanism of solubilization, molecular genetics, and capacity to release P into the soil [50,51]. Our results reveal that Paecilomyces lilacinus 112 has the ability to solubilize P, the appearance of the transparent zone surrounding fungal colonies in a solid medium with Ca3(PO4)2 presenting a positive result. Other isolates belonging to the Paecilomyces genus have been reported to function similarly. Paecilomyces marquandii was described as an efficient phosphate solubilizer, especially grown in media with a wide range of carbon (fructose, galactose, glucose, glycerol, lactose, maltose, mannose, starch, and sucrose) and nitrogen sources (ammonium chloride, sulphate, nitrates, and urea) [52].
The complete growth and development of plants also depends on zinc, another essential micronutrient active in enzymatic reactions, carbohydrate metabolism, and the synthesis of proteins and auxins. Zinc existing in the soil is partially available since it is in insoluble form, and only cations Zn+2 can be taken up by plants [53,54]. In our tests, the Paecilomyces lilacinus 112 isolate exhibited zones of solubilization in the in vitro qualitative tests carried out with three insoluble zinc sources (zinc oxide, zinc nitrate, and zinc carbonate). Another study also reported the use of a Paecilomyces isolate as a potent bioinoculant for agricultural crops, an alternative to synthetic Zn sources [23].
The capability of Paecilomyces lilacinus 112 to solubilize phosphorus and zinc is significant feature of plant growth-promoting agents. These properties could ameliorate the uptake of phosphate and zinc by the plants, resulting in increase in the vegetative and biological parameters.
Endophytic fungi produce metabolic compounds important for their own biology and interactions with host plant. They secrete a wide range of hydrolytic extracellular enzymes that support their ecological role in combating pathogens through an important mechanism for fungal growth inhibition [55,56,57,58]. It is considered that endophytic fungal strains could represent a valuable resource for the production of cellulolytic and hemicellulolytic enzymes [59,60]. The hydrolytic enzymes can inhibit cell wall synthesis, perforating the cell membrane or degrading cell walls of pathogens. Due to the relevance of hydrolytic enzymes for an efficient biological control, chitinase and cellulase activities were analyzed through qualitative tests on agar specific medium. Chitinases can degrade the fungal cell wall, while cellulases cleave β-1,4-D glucan bonds of cellulose, the major polysaccharide component of the cell wall. Additionally, keratinolytic activity was analyzed since the carbon source in culture medium is keratin, a complex and structurally stable protein resistant to the degradation. Paecilomyces lilacinus 112 exhibited good enzymatic activities, as chitinases, cellulases, and keratinases, which ensure an antifungal activity against phytopathogenic fungi. This result is consistent with other studies that have reported the production of hydrolytic enzymes at the members of Paecilomyces genus. The fungus P. marquandii, through its keratinolytic enzymes, was able to hydrolyze keratin waste to form soluble products with a high level of nitrogen, mainly common amino acids. The obtained mixture can be used as foliar fertilizers [61]. Metabolites secreted by endophytic Paecilomyces variotii SJ1 have great potential for improving the crops quality. The biological activity of processed mycelium through ultrasonic extraction was tested on field potatoes and a significant increase in culture yield by 4.4–10.8% was obtained [62].
The effect of culture supernatants obtained from Paecilomyces lilacinus 112 on tomato plant growth was investigated. Tomato plants were chosen as they are one of the most important vegetables worldwide, and are also considered a strategic crop. Annually, there are huge losses of tomato crops due to various pathogens and pests, and it is often difficult to control the destructive action of pathogens since they have various mode of attack. In addition, there is already a resistance against common fungicides, and the development of new chemical formulations is expensive and overrides environmental regulations. In this context, it is advantageous to use beneficial microorganisms.
The Lowry test showed that the quality of hydrolyzates from cultivation on chicken feathers, in terms of soluble protein content, is superior to those obtained on wool medium. This aspect is supported by the results of the tests performed in greenhouse conditions. Paecilomyces lilacinus 112 was able to produce beneficial effects on tomato plant growth and development, the higher values of parameters growth were obtained in plant treatment with PH obtained from fungal cultivation on medium with 1% feathers. We obtained the following increases in the analyzed parameters: 3.54-fold in plant biomass; 3.26-fold in plant height; 1.28-fold in plant diameter; 1.5-fold in the number of branches/plant; and 1.43-fold in the number of leaves/plant, compared to treatment of plants with water.
Similar benefits have been observed as a result of the application of other species of the genus Paecilomyces. Thus, Baron et al. [63] demonstrated that Paecilomyces lilacinus strains can significantly increase growth promotion parameters of maize, bean, and soybean plants, the greatest performance being for the strains characterized by their P solubilization ability. Some Paecilomyces strains were selected for their potential as biofertilizers after test trials. Hence, P. lilacinus strain LSM 65 provided a significant increase in maize plant height compared to the control. The interactions between each plant species and microbial strains are very complex and depend on the location of the fungus, mode of resistance induction, and production of phytohormones. As novel biostimulant, extracts of mycelium from Paecilomyces variotii integrated with diammonium phosphate coated with a biobased polyurethane polymer were analyzed in a three-year field trial. The treatments significantly increased maize total root length by 178.2% and average yield by 9.65% [64].
5. Conclusions
Paecilomyces lilacinus 112 presents the ability to produce hydrolytic enzymes involved in pathogen inhibition, secreting useful secondary metabolites and solubilizing essential micronutrients, such as phosphorus and zinc. Our data corroborate with encouraging results from greenhouse trials regarding the beneficial effects of protein hydrolyzates upon plants, indicating its potential as a plant biostimulant. The use of keratinolytic fungi in producing protein hydrolyzates, via culturing on industrial byproducts and waste rich in nitrogen, as biostimulants for plants, could represent a promising and sustainable biomass valorization processes, following circular economy principles.
Author Contributions
L.J., I.R. and M.C. conceived and designed the experiments; M.C., I.R., E.A. and A.-M.G. performed the experiments; L.J., M.D., A.-M.G. and N.R. analyzed the data; L.J., A.-M.G. and M.D. wrote the paper; L.J., A.-M.G. and N.R. reviewed and edited. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by MCID within the Core-Program, project number PN 19.23.01.01/2019 and by a national grant of UEFISCDI, through project number PN-III-P2-2.1-PED-2019-0991, within PNCDI III.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank to the Ministry of Research, Innovation and Digitization of Romania, through Program 1—Development of the national research and development system, Subprogram 1.2—Institutional performance—Projects to finance excellence in RDI, Contract no. 15PFE/2021.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci 2019, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for Plant Growth and Mitigation of Abiotic Stresses: A Metabolomics Perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Du Jardin, P. Plant biostimulants: Definition; concept; main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Drobek, M.; Fra, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Reddy, N. Non-food industrial applications of poultry feathers. Waste Manag. 2015, 45, 91–107. [Google Scholar] [CrossRef]
- Tesfaye, T.; Sithole, B.; Ramjugernath, D. Valorisation of chicken feathers: A review on recycling and recovery route—Current status and future prospects. Clean Technol. Environ. Policy 2017, 19, 2363–2378. [Google Scholar] [CrossRef]
- Khumalo, M.; Tesfaye, T.; Sithole, B.; Ramjugernath, D. Possible Beneficiation of Waste Chicken Feathers via Conversion into Biomedical Applications. Int. J. Chem. Sci. 2019, 17, 298. [Google Scholar] [CrossRef]
- Šafari, R.; Zemljič, L.F.; Novak, M.; Dugonik, B.; Bratina, B.; Gubeljak, N.; Bolka, S.; Strnad, S. Preparation and Characterisation of Waste Poultry Feathers Composite Fibreboards. Materials 2020, 13, 4964. [Google Scholar] [CrossRef] [PubMed]
- Marchelli, F.; Rovero, G.; Curti, M.; Arato, E.; Bosio, B.; Moliner, C. An Integrated Approach to Convert Lignocellulosic and Wool Residues into Balanced Fertilisers. Energies 2021, 14, 497. [Google Scholar] [CrossRef]
- Rajabinejad, H.; Bucişcanu, I.I.; Maier, S.S. Current approaches for raw wool waste management and unconventional valorization: A review. Environ. Eng. Manag. J. 2019, 18, 1439–1456. [Google Scholar]
- He, Z.; Sun, R.; Tang, Z.; Bu, T.; Wu, Q.; Li, C.; Chen, H. Biodegradation of Feather Waste Keratin by the Keratin-Degrading Strain Bacillus subtilis 8. J. Microbiol. Biotechnol. 2018, 28, 314–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joardar, J.C.; Rahman, M.M. Poultry feather waste management and effects on plant growth. Int. J. Recycl. Org. Waste Agric. 2017, 7, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Ghaffar, I.; Imtiaz, A.; Hussain, A.; Arshad Javid, A.; Jabeen, F.; Akmal, M.; Qaz, J.I. Microbial production and industrial applications of keratinases: An overview. Int. Microbiol. 2018, 21, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Mousa, W.K.; Raizada, M.N. The diversity of anti-microbial secondary metabolites produced by fungal endophytes: An interdisciplinary perspective. Front. Microbiol. 2013, 4, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadiji, A.E.; Babalola, O.O. Exploring the potentialities of beneficial endophytes for improved plant growth. Saudi J. Biol. Sci. 2020, 27, 3622–3633. [Google Scholar] [CrossRef]
- Dai, Z.B.; Wang, X.; Hong, G. Secondary Metabolites and Their Bioactivities Produced by Paecilomyces. Molecules 2020, 25, 5077. [Google Scholar] [CrossRef]
- Akpor, O.B.; Odesola, D.E.; Thomas, R.E.; Oluba, O.M. Chicken feather hydrolysate as alternative peptone source for microbial cultivation. F1000Research 2019, 7, 1918. [Google Scholar] [CrossRef]
- Islam, M.A.; Nain, Z.; Alam, M.K.; Banu, N.A.; Islam, M.R. In vitro study of biocontrol potential of rhizospheric Pseudomonas aeruginosa against Fusarium oxysporum f. sp. Cucumerinum. Egypt. J. Biol. Pest Control 2018, 28, 90. [Google Scholar] [CrossRef]
- Sakpetch, P.; Aran, H.; Kuwahara, Y.; Komeda, H.; Asano, Y. Isolation of indigenous antagonistic microorganism to inhibit Rigidoporus microporus and other plant pathogens and analysis of the bioactive compounds. Biol. Control 2018, 124, 53–60. [Google Scholar] [CrossRef]
- Yoon, J.H.; Park, J.E.; Dong, Y.S.; Hong, S.B.; Ko, S.J.; Kim, S.H. Comparison of dyes for easy detection of extracellular cellulases in fungi. Mycobiology 2007, 35, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Lateefa, A.; Adelerea, I.A.; Gueguim-Kana, B.E.B. Bacillus safensis LAU 13: A new source of keratinase and its multi-functional biocatalytic applications. Biotechnol. Biotechnol. Equip. 2015, 29, 5463. [Google Scholar] [CrossRef]
- Anitha, S.; Padma, D.S.N.; Sunith, K.K. Isolation and identification of zinc solubilizing fungal isolates from agricultural fields. Indian J. Agric. Sci. 2015, 85, 1638–1642. [Google Scholar]
- Kaur, G.; Reddy, M.S. Role of Phosphate-Solubilizing Fungi in Sustainable Agriculture. In Developments in Fungal Biology and Applied Mycology; Satyanarayana, T., Deshmukh, S., Johri, B., Eds.; Springer: Singapore, 2017; pp. 391–412. [Google Scholar] [CrossRef]
- Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control. 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 256–275. [Google Scholar] [CrossRef]
- Li, X.Q.; Xu, K.; Liu, X.M.; Zhang, P. Systematic Review on Secondary Metabolites of Paecilomyces Species: Chemical Diversity and Biological Activity. Planta Med. 2020, 86, 805–821. [Google Scholar] [CrossRef]
- Suárez-Estrella, F.; Arcos-Nievas, M.A.; López, M.J.; Vargas-García, M.C.; Moreno, J. Biological control of plant pathogens by microorganisms isolated from agro-industrial composts. Biol. Control 2013, 67, 509–515. [Google Scholar] [CrossRef]
- Nesha, R.; Siddiqui, Z.A. Effects of Paecilomyces lilacinus and Aspergillus niger alone and in combination on the growth, chlorophyll contents and soft rot disease complex of carrot. Sci. Hortic. 2017, 218, 258–264. [Google Scholar] [CrossRef]
- Varma, P.K.; Gandhi, S.K.; Surender, S. Biological control of Alternaria solani, the causal agent of early blight of tomato. J. Biol. Control 2008, 22, 67–72. [Google Scholar]
- Sivakumar, T.; Eswaran, A.; Balabaskar, P. Bioefficacy of antagonists against for the management of Fusarium oxysporum f. sp. lycopersici and Meloidogyne incognita disease complex of tomato under field condition. Plant Arch. 2008, 8, 373–377. [Google Scholar]
- Yang, F.; Abdelnabby, H.; Xiao, Y. A mutant of the nematophagous fungus Paecilomyces lilacinus (Thom) is a novel biocontrol agent for Sclerotinia sclerotiorum. Microb. Pathog. 2015, 89, 169–176. [Google Scholar] [CrossRef]
- Al-Hazmi, A.S.; Dawabah, A.A.M.; Al-Nadhari, S.N.; Al-Yahya, F.A. Comparative efficacy of different approaches to managing Meloidogyne incognita on green bean. Saudi J. Bio. Sci. 2017, 24, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Dahlin, P.; Eder, R.; Consoli, E.; Krauss, J.; Kiewnick, S. Integrated control of Meloidogyne incognita in tomatoes using fluopyram and Purpureocillium lilacinum strain 251. Crop Prot. 2019, 124, 104874. [Google Scholar] [CrossRef]
- Moreno-Gavíra, A.; Huertas, V.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Paecilomyces and Its Importance in the Biological Control of Agricultural Pests and Diseases. Plants 2020, 9, 1746. [Google Scholar] [CrossRef]
- Moreno-Gavíra, A.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Biocontrol Effects of Paecilomyces variotii against Fungal Plant Diseases. J. Fungi 2021, 7, 415. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liu, H.; Jiang, D.; Wang, L.; Jiang, Y.; Tang, S.; Hou, X.; Han, X.; Chu, Z.; Ding, X. Paecilomyces variotii extracts (ZNC) enhance plant immunity and promote plant growth. Plant Soil 2019, 441, 383–397. [Google Scholar] [CrossRef]
- Rauwane, M.E.; Ogugua, U.V.; Kalu, C.M.; Ledwaba, L.K.; Woldesemayat, A.A.; Ntushelo, K. Pathogenicity and Virulence Factors of Fusarium graminearum Including Factors Discovered Using Next Generation Sequencing Technologies and Proteomics. Microorganisms 2020, 8, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, M.; Hassan, M.; Abo-Elyousr, K.A.M. Biological control of Botrytis allii by Trichoderma viride on onion (Allium cepa). World Appl. Sci. J. 2014, 32, 522–526. [Google Scholar] [CrossRef]
- Erlacher, A.; Cardinale, M.; Grosch, R.; Grube, M.; Berg, G. The impact of the pathogen Rhizoctonia solani and its beneficial counterpart Bacillus amyloliquefaciens on the indigenous lettuce microbiome. Front. Microbiol. 2014, 5, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swett, C.L.; Hamby, K.A.; Hellman, E.M.; Carignan, C.; Bourret, T.B.; Koivunen, E.E. Characterizing members of the Cladosporium cladosporioides species complex as fruit rot pathogens of red raspberries in the mid-Atlantic and co-occurrence with Drosophila suzukii (spotted wing drosophila). Phytoparasitica 2019, 47, 415–428. [Google Scholar] [CrossRef]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria Toxins: Potential Virulence Factors and Genes Related to Pathogenesis. Front. Microbiol. 2017, 8, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, S.; Thomas, C.M. Molecular Interactions Between Tomato and the Leaf Mold Pathogen Cladosporium fulvum. Ann. Rev. Phytopathol. 2005, 43, 395–436. [Google Scholar] [CrossRef] [PubMed]
- Demirci, E.; Dane, E.; Eken, C. In vitro antagonistic activity of fungi isolated from sclerotia on potato tubers against Rhizoctonia solan. Turk. J. Biol. 2011, 35, 457–462. [Google Scholar] [CrossRef]
- Abegaz, B.M.; Kinfe, H.H. Secondary metabolites, their structural diversity, bioactivity, and ecological functions: An overview. Phys. Sci. Rev. 2019, 4, 20180100. [Google Scholar] [CrossRef]
- Shuikan, A.M.; Hozzein, W.N.; Alzharani, M.M.; Sandouka, M.N.; Al Yousef, S.A.; Alharbi, S.A.; Damra, E. Enhancement and Identification of Microbial Secondary Metabolites. In Extremophilic Microbes and Metabolites—Diversity, Bioprospecting and Biotechnological Applications; Afef, N., Ameur, C., Haïtham, S., Hadda, I.O., Eds.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Holinsworth, B.; Martin, J.D. Siderophore production by marine derived fungi. BioMetals 2009, 22, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Renshaw, J.C.; Robson, G.D.; Trinci, A.P.; Wiebe, M.G.; Livens, F.R.; Collison, D.; Taylor, R.J. Fungal siderophores: Structures, functions and applications. Mycol. Res. 2002, 106, 1123–1142. [Google Scholar] [CrossRef]
- Billah, M.; Khan, M.; Bano, A.; Ul Hassan, T.; Asia Munir, A.; Gurmani, A.R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Mitra, D.; Snđelković, S.; Panneerselvam, P.; Senapati, A.; Vasić, T.; Ganeshamurthy, A.N.; Chauhan, M.; Uniyal, N.; Mahakur, B.; Radha, T.K. Phosphate-Solubilizing Microbes and Biocontrol Agent for Plant Nutrition and Protection: Current Perspective. Commun. Soil Sci. Plant. Anal. 2020, 51, 645–657. [Google Scholar] [CrossRef]
- Ahuja, A.; D’Souza, F.D. Bioprocess for Solubilization of Rock Phosphate on Starch Based Medium by Paecilomyces marquandii Immobilized on Polyurethane Foam. Appl. Biochem. Biotechnol. 2009, 152, 1. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, P.; Kashyap, P.L.; Pandiyan, K.; Bhardwaj, A.K. Zinc-Solubilizing Microbes for Sustainable Crop Production: Current Understanding, Opportunities, and Challenges. In Phytobiomes. Current Insights and Future Vistas; Solanki, M., Kashyap, P., Kumari, B., Eds.; Springer: Singapore, 2020; pp. 281–298. [Google Scholar] [CrossRef]
- Bhatt, K.; Maheshwari, D.K. Zinc solubilizing bacteria (Bacillus megaterium) with multifarious plant growth promoting activities alleviates growth in Capsicum annuum L. 3 Biotech 2020, 36, 10. [Google Scholar] [CrossRef] [PubMed]
- Farouk, H.M.; Attia, E.Z.; El-Katatny, M.H. Hydrolytic enzyme production of endophytic fungi isolated from soybean (Glycine max). J. Mod. Res. 2020, 2, 1–7. [Google Scholar] [CrossRef]
- Lopez-Llorca, L.V.; Carbonell, T.; Gomez-Vidal, S. Degradation of insect cuticle by Paecilomyces farinosus proteases. Mycol. Prog. 2002, 1, 249–256. [Google Scholar] [CrossRef]
- Cavello, I.A.; Crespo, J.M.; García, S.S.; Zapiola, J.M.; Luna, M.F.; Cavalitto, S.F. Plant growth promotion activity of keratinolytic fungi growing on a recalcitrant waste known as Hair Waste. Biotechnol. Res. Intern. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.; Mishra, J.; Dwivedi, S.K.; Arora, N.K. Chapter 10. Microbial Enzymes in Biocontrol of Phytopathogens. In Microbial Enzymes: Roles and Applications in Industries Microorganisms for Sustainability 11; Arora, N.K., Mishra, J., Mishra, V., Eds.; Springer Nature Singapore Pte Ltd.: Berlin/Heidelberg, Germany, 2020; pp. 259–285. [Google Scholar] [CrossRef]
- Roblet, D.; Delabona, P.S.; Mergel, C.M.; Rojas, J.D.; Costa, P.S.; Pimente, I.C.; Vicente, V.A.; Pradella, J.G.; Padilla, G. The capability of endophytic fungi for production of hemicellulases and related enzymes. BMC Biotechnol. 2013, 13, 94. [Google Scholar] [CrossRef] [Green Version]
- Correa, R.C.G.; Rhoden, S.A.; Mota, T.R.; Azevedo, J.L.; Pamphile, J.A.; Souza, C.G.M.; Polizeli, M.L.T.M.; Bracht, A.; Peralta, R.M. Endophytic fungi: Expanding the arsenal of industrial enzyme producers. J. Ind. Microbiol. Biotechnol. 2014, 41, 1467–1478. [Google Scholar] [CrossRef]
- Vesela, M.; Friederich, J. Amino Acid and Soluble Protein Cocktail from Waste Keratin Hydrolysed by a Fungal Keratinase of Paecilomyces marquandii. Biotechnol. Bioprocess. Eng. 2009, 14, 84–90. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, C.; Shi, L.; Liu, Z.; Zhou, D.; Meng, H.; Zhao, H.; Li, F.; Zhang, M. A Technical System for the Large-Scale Application of Metabolites from Paecilomyces variotii SJ1 in Agriculture. Front. Bioeng. Biotechnol. 2021, 9, 671879. [Google Scholar] [CrossRef] [PubMed]
- Baron, N.C.; de Souza Pollo, A.; Rigobelo, E.C. Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 2020, 8, e9005. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, Z.; Qu, Z.; Zhou, H.; Qi, Y.; Liu, Z.; Zhang, M. Maize yield and root morphological characteristics affected by controlled-release diammonium phosphate and Paecilomyces variotii extracts. Field Crops Res. 2020, 255, 107862. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of plant treatments with protein hydrolyzates obtained from Paecilomyces lilacinus cultured on keratin wastes.
Figure 1.
Schematic representation of plant treatments with protein hydrolyzates obtained from Paecilomyces lilacinus cultured on keratin wastes.

Figure 2.
Illustration of the antagonism of Paecilomyces lilacinus 112 versus plant pathogenic fungi.
Figure 2.
Illustration of the antagonism of Paecilomyces lilacinus 112 versus plant pathogenic fungi.

Figure 3.
(A) Observations of the antagonism mechanism between Paecilomyces lilacinus 112 and plant pathogenic fungi under microscope light (left) and SEM (right). (a) Paecilomyces lilacinus versus Alternaria alternata: Paecilomyces presents ovoid conidia forming chains that can be intertangled (white arrow); septate and thin hyphae of Paecilomyces are coiled or attach upon Alternaria hyphae; septate and large hyphae of Alternaria (black arrow); acropetal chains of conidia (white arrow). (b) Paecilomyces lilacinus versus Cladosporium T2 isolate: Paecilomyces develops thin hyphae that are coiled around Cladosporium filaments (white arrow); Cladosporium large hyphae, conidiophores, and conidia in branched acropetal chains (black arrow). (c) Paecilomyces lilacinus versus Rhizoctonia solani: Paecilomyces thin hyphae coiled or attached onto R. solani hyphae (white arrow); Paecilomyces conidiophores, phialides, and long chains of ovoid conidia. Deformation of R. solani hyphae caused by Paecilomyces activity; R. solani large hyphae (black arrow). (B) Observations of the antagonism mechanism between Paecilomyces lilacinus 112 and pathogens under microscope light (left) and SEM (right). (d) Paecilomyces lilacinus versus Botrytis allii: B. allii presents large hyphae and conidia (black arrow). Paecilomyces thin hyphae are coiled or attached on the B. allii hyphae; conidiophores, phialides, and long chains of ovoid conidia (white arrow). Deformation of pathogen hyphae caused by Paecilomyces activity. (e) Paecilomyces lilacinus sp. versus Fusarium graminearum: Thin hyphae and long chains of ovoid conidia from Paecilomyces (white arrow). Large hyphae of F. graminearum (black arrow). (f) Paecilomyces lilacinus versus Sclerotinia sclerotiorum: Large hyphae and mature melanized sclerotia of S. sclerotiorum (black arrow). Paecilomyces hyphae coiled or attached on S. sclerotiorum hyphae (white arrow); chains of ovoid conidia. Deformation of S. sclerotiorum hyphae caused by Paecilomyces activity.
Figure 3.
(A) Observations of the antagonism mechanism between Paecilomyces lilacinus 112 and plant pathogenic fungi under microscope light (left) and SEM (right). (a) Paecilomyces lilacinus versus Alternaria alternata: Paecilomyces presents ovoid conidia forming chains that can be intertangled (white arrow); septate and thin hyphae of Paecilomyces are coiled or attach upon Alternaria hyphae; septate and large hyphae of Alternaria (black arrow); acropetal chains of conidia (white arrow). (b) Paecilomyces lilacinus versus Cladosporium T2 isolate: Paecilomyces develops thin hyphae that are coiled around Cladosporium filaments (white arrow); Cladosporium large hyphae, conidiophores, and conidia in branched acropetal chains (black arrow). (c) Paecilomyces lilacinus versus Rhizoctonia solani: Paecilomyces thin hyphae coiled or attached onto R. solani hyphae (white arrow); Paecilomyces conidiophores, phialides, and long chains of ovoid conidia. Deformation of R. solani hyphae caused by Paecilomyces activity; R. solani large hyphae (black arrow). (B) Observations of the antagonism mechanism between Paecilomyces lilacinus 112 and pathogens under microscope light (left) and SEM (right). (d) Paecilomyces lilacinus versus Botrytis allii: B. allii presents large hyphae and conidia (black arrow). Paecilomyces thin hyphae are coiled or attached on the B. allii hyphae; conidiophores, phialides, and long chains of ovoid conidia (white arrow). Deformation of pathogen hyphae caused by Paecilomyces activity. (e) Paecilomyces lilacinus sp. versus Fusarium graminearum: Thin hyphae and long chains of ovoid conidia from Paecilomyces (white arrow). Large hyphae of F. graminearum (black arrow). (f) Paecilomyces lilacinus versus Sclerotinia sclerotiorum: Large hyphae and mature melanized sclerotia of S. sclerotiorum (black arrow). Paecilomyces hyphae coiled or attached on S. sclerotiorum hyphae (white arrow); chains of ovoid conidia. Deformation of S. sclerotiorum hyphae caused by Paecilomyces activity.


Figure 4.
Formation of halos owing to production of hydrolytic enzymes and siderophores by Paecilomyces lilacinus on agar plates. (a) Chitinolytic activity (arrow—zone of enzymatic activity); (b) cellulolytic activity (brown color of medium, iodine staining of cellulose medium); (c) keratinolytic activity (arrow—clear zone of enzymatic activity); (d) production of siderophores on medium with 8-hydroxychinoline.
Figure 4.
Formation of halos owing to production of hydrolytic enzymes and siderophores by Paecilomyces lilacinus on agar plates. (a) Chitinolytic activity (arrow—zone of enzymatic activity); (b) cellulolytic activity (brown color of medium, iodine staining of cellulose medium); (c) keratinolytic activity (arrow—clear zone of enzymatic activity); (d) production of siderophores on medium with 8-hydroxychinoline.

Figure 5.
Formation of solubilization zone owing the activity of Paecilomyces lilacinus. (a) ZnO (arrow—clear zone of solubilization); (b) ZnNO3 (arrow—discrete zone of solubilization); (c) ZnSO4 (arrow—pale zone of solubilization); (d) phosphorous solubilization (arrow—violet-purple color of medium as solubilization area).
Figure 5.
Formation of solubilization zone owing the activity of Paecilomyces lilacinus. (a) ZnO (arrow—clear zone of solubilization); (b) ZnNO3 (arrow—discrete zone of solubilization); (c) ZnSO4 (arrow—pale zone of solubilization); (d) phosphorous solubilization (arrow—violet-purple color of medium as solubilization area).

Figure 6.
Aspects of plants treated with protein hydrolyzates from Paecilomyces lilacinus 112 in growth chamber conditions. (a) Tomato plants after four treatments with PH; (b) tomato plants one week after the end of treatment with PH; (c) tomato plants treated with PH from fungal culture on wool medium; (d) tomato plants treated with PH from fungal culture on feather medium.
Figure 6.
Aspects of plants treated with protein hydrolyzates from Paecilomyces lilacinus 112 in growth chamber conditions. (a) Tomato plants after four treatments with PH; (b) tomato plants one week after the end of treatment with PH; (c) tomato plants treated with PH from fungal culture on wool medium; (d) tomato plants treated with PH from fungal culture on feather medium.

Figure 7.
The influence on tomato plants of treatments with PH obtained from Paecilomyces lilacinus cultured on keratin medium. (a) Influence on production of biomass (average values). (b) Influence on plant height (average values). (c) Influence on plant diameter (average values). (d) Influence on number of branches per plant (average values). (e) Influence on number of plants leaves (average values). Experimental variants: control (A01)—data of plant treated with water; A02—data of plant treated with PH from P. lilacinus cultured on feather medium; A03—data of plant treated with PH from P. lilacinus cultured on wool medium.
Figure 7.
The influence on tomato plants of treatments with PH obtained from Paecilomyces lilacinus cultured on keratin medium. (a) Influence on production of biomass (average values). (b) Influence on plant height (average values). (c) Influence on plant diameter (average values). (d) Influence on number of branches per plant (average values). (e) Influence on number of plants leaves (average values). Experimental variants: control (A01)—data of plant treated with water; A02—data of plant treated with PH from P. lilacinus cultured on feather medium; A03—data of plant treated with PH from P. lilacinus cultured on wool medium.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Interaction of Paecilomyces lilacinus 112 with tested pathogens.
| Antagonistic Fungal Isolate | Pathogenic Fungi | Diameter of Pathogen Colony in Presence of Antagonist (cm) * | Diameter of Pathogen Colony (Control) (cm) * | Inhibition of Tested Pathogens (%) |
|---|---|---|---|---|
| Paecilomyces lilacinus 112 | Rhizoctonia solani | 3.8 ± 0.1 | 8.0 ± 0.1 | 52.5 ± 2.1 |
| Sclerotinia sclerotiorum. | 4.2 ± 0.1 | 8.5 ± 0.1 | 50.2 ± 1.7 | |
| Fusarium graminearum | 3.0 ± 0.1 | 5.8 ± 0.1 | 47.1 ± 1.9 | |
| Botrytis allii | 3.6 ± 0.1 | 5.1 ± 0.2 | 29.4 ± 1.9 | |
| Cladosporium T2 | 2.7 ± 0.1 | 8.1 ± 0.1 | 66.3 ± 1.3 | |
| Alternaria alternata | 4.0 ± 0.1 | 5.9 ± 0.1 | 32.2 ± 1.7 |
* Values are the average of three independent experiments ± standard deviations.
Table 2.
Content of soluble protein in fungal hydrolyzates.
| Protein Hydrolyzate | Soluble Protein (mg/mL) |
|---|---|
| Paecilomyces lilacinus 112 cultivated on feather medium | 0.230 |
| Paecilomyces lilacinus 112 cultivated on wool medium | 0.110 |
| Paecilomyces lilacinus 112 cultivated on medium without keratin (control) | 0.002 |
Table 3.
Evaluation of treatment effects applied to tomato seedlings.
| Plant Treatments | Growth Parameters of Plants | ||||
|---|---|---|---|---|---|
| Biomass * (g) | Plant Height * (cm) | Plant Diameter * (mm) | Number of Branches/Plants * | Number of Leaves/Plants * | |
| Treatment with PH obtained from Paecilomyces lilacinus cultured on feather medium | 1.8 ± 0.2 | 26.6 ± 0.4 | 2.9 ± 0.3 | 7.0 ± 0.3 | 34.3 ± 0.3 |
| Treatment with PH obtained from Paecilomyces lilacinus cultured on wool medium | 1.6 ± 0.3 | 27.8 ± 0.4 | 2.8 ± 0.2 | 6.0 ± 0.3 | 29.3 ± 0.4 |
| Treatment with water | 0.5 ± 0.3 | 8.1 ± 0.3 | 2.2 ± 0.2 | 4.6 ± 0.2 | 24.0 ± 0.3 |
* Values are the average of ten independent experiments ± standard deviations.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Constantin, M.; Raut, I.; Gurban, A.-M.; Doni, M.; Radu, N.; Alexandrescu, E.; Jecu, L. Exploring the Potential Applications of Paecilomyceslilacinus 112. Appl. Sci. 2022, 12, 7572. https://doi.org/10.3390/app12157572
AMA Style
Constantin M, Raut I, Gurban A-M, Doni M, Radu N, Alexandrescu E, Jecu L. Exploring the Potential Applications of Paecilomyceslilacinus 112. Applied Sciences. 2022; 12(15):7572. https://doi.org/10.3390/app12157572
Chicago/Turabian StyleConstantin, Mariana, Iuliana Raut, Ana-Maria Gurban, Mihaela Doni, Nicoleta Radu, Elvira Alexandrescu, and Luiza Jecu. 2022. "Exploring the Potential Applications of Paecilomyceslilacinus 112" Applied Sciences 12, no. 15: 7572. https://doi.org/10.3390/app12157572
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.