Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review

School of Biological Sciences, Universiti Sains Malaysia, USM, Penang 11800, Malaysia
Agriculture 2021, 11(4), 297; https://doi.org/10.3390/agriculture11040297
Submission received: 5 March 2021
/
Revised: 24 March 2021
/
Accepted: 24 March 2021
/
Published: 30 March 2021
(This article belongs to the Section Crop Protection, Diseases, Pests and Weeds)
Abstract
:In tropical fruit crops, anthracnose is mainly caused by species belonging to the fungal genus, Colletotrichum. These phytopathogens can infect several parts of the fruit crops; however, infection during postharvest or ripening stages is responsible for major economic losses. Due to the formation of black to dark brown sunken lesions on the fruit surface, anthracnose reduces fruit quality and marketability. Among the most common tropical fruit crops susceptible to anthracnose are mango, papaya, banana, avocado, guava, and dragon fruit; these are economically relevant products in many developing countries. It is important to document that the newly recorded Colletotrichum spp. associated with fruit anthracnose can infect multiple hosts, but some species may be host-specific. By using multiple markers, many phylogenetic species of Colletotrichum have been reported as anthracnose-causing pathogens. Taking into account that disease management strategies strongly rely on adequate knowledge of the causative agents, updated information on Colletotrichum species and the hazard posed by the most recently identified species in tropical fruit plantations and harvested fruits becomes vital. Besides, the newly recorded species may be important for biosecurity and should be listed as quarantine pathogens, considering that tropical fruits are traded worldwide.
1. Introduction
Asia and countries of the Pacific region are major tropical fruit crop producers, followed by Latin America, the Caribbean, and Africa [1]. Minor producers include the United States and Oceania. The main tropical fruits are mango, pineapple, avocado, and papaya; with mango representing the most relevant production worldwide. The majority of tropical fruits are produced in India, the leading producer of mango and papaya, followed by Thailand, Mexico, China, Brazil, and Indonesia [2,3]. Minor tropical fruits include guava, longan, litchi, durian, passion fruit, rambutan, and mangosteen, with China and India being the largest producers [4].
Tropical fruit crops are susceptible to infection by Colletotrichum species, which typically cause anthracnose. Figure 1 summarizes the anthracnose disease cycle in tropical fruit crops. Anthracnose infection commonly occurs in the fields during the flowering and fruiting stages. Different factors may affect Colletotrichum infection, including humidity, temperature, fruit condition, and inoculum concentration [5,6].
Host infection generally begins with conidial germination and is followed by the formation of appressoria and penetration pegs, which are fungal structures that assist in the penetration into host tissues. In some cases, direct penetration occurs through wounds or natural openings [7,8].
After infection, many anthracnose pathogens adopt quiescence or latency, which is common in pathogens causing postharvest diseases, including Colletotrichum. During the latent period, anthracnose pathogens remain dormant within the host tissues until environmental conditions, and the host physiology are conducive for their reactivation and further development [9]. Reactivation occurs particularly when fruits ripen. Anthracnose symptoms often develop after harvest, during storage, transportation, and marketing [6,10].
Anthracnose pathogens infect not only fruits, but also other plant organs, including the leaves, flowers, twigs, and branches. The conidia and spores formed in these infected tissues are subsequently released and dispersed during rainy days through water splashes or during high humidity periods, thus becoming the primary inoculum for fruit infection at the preharvest stage [11]. The most visible anthracnose symptoms are black or dark brown sunken lesions containing conidial masses on the surface of infected fruits [7]. Small individual lesions may merge to produce larger lesions. These black or dark brown lesions on the surface appear unattractive to consumers and significantly reduce the market value of such fruits. Figure 2A–E show the anthracnose lesions form on the surface of several tropical fruit crops.
Before the application of molecular-based phylogenetic analysis using multiple markers for the taxonomic and systematic revision of Colletotrichum spp., only two species, Colletotrichum acutatum and Colletotrichum gloeosporioides, were reported to be associated with anthracnose symptoms in many tropical fruit crops. However, more recent phylogenetic analysis established that these two species belong to two complexes called “acutatum” and “gloeosporioides”, with several other species included within them [12,13]. Table 1 summarized the Colletotrichum species reported to be associated with anthracnose of tropical fruit crops from several countries. Many of the species were identified based on phylogenetic analysis of multiple markers.
Many Colletotrichum species that are part of these complexes are reported to cause anthracnose [14,15,16]. Some species are known to infect specific hosts, while others infect multiple hosts. Anthracnose pathogens that infect multiple hosts may indicate the development of cross-infection ability. Based on a cross-pathogenicity study by Phoulivong et al. (2012) [16], several species within the C. gloeosporioides complex were found to have the capacity to infect multiple hosts. Some examples include C. asianum, detected in infected chili, mango, and rose apple, and C. fructicola, a fungus initially reported to infect coffee berries, but now recognized as a phytopathogen in other plant species, such as chili, citrus fruits, rose apple, avocado, grapes, and papaya [17].
Previous studies (published before systematic revisinions of Colletotrichum genus based on phylogenetic analysis of multiple markers) indicate cross-infection among different anthracnose pathogens. For instance, Freeman et al. (2001) [18] showed that C. acutatum sensu lato isolated from strawberry is able to infect various fruit crops. Moreover, cross-infection studies by Sanders and Korsten (2003) [19] show that isolates of C. gloeosporioides sensu lato from mango could infect and produce symptoms in guava, chili, and papaya. Therefore, cross-infection of tropical fruits by various Colletotrichum species can occur in the field. Likewise, anthracnose pathogens may infect the same fruit crop in various countries. Therefore, it is important to identify these pathogens correctly and assign appropriate scientific names. Furthermore, information on the lifestyle and mode of infection of each species is key point to implement suitable control measures [14]. Moreover, a deeper knowledge of species distribution and population size can provide valuable insights into breeding strategies directed to achieve durable resistance to anthracnose, as well as to improve control methods.
Due to the economic importance of anthracnose in the context of tropical fruit production and commercialization, this review focuses on the current knowledge regarding Colletotrichum species associated with tropical fruit crops reported in several fruit-producing countries. Emphasis is laid on Colletotrichum species associated with mango, papaya, banana, avocado, guava, and dragon fruit because these fruit crops are cultivated in many tropical countries as an income source, contributing significantly to the economic well-being of their inhabitants.
2. Banana Anthracnose
Banana (Musa spp.) is one of the most important fruit crops and the most popular fruit consumed worldwide, with over 100 billion bananas eaten every year [98]. More than 150 countries have banana plantations; they are mainly distributed in Asia, Latin America, and Africa. The largest banana producer is India, followed by China, the Philippines, Ecuador, and Brazil [99].
International banana trade mainly involves the Cavendish type, which replaced the Gros Michel variety due to its resistance to Fusarium wilt. Currently, Cavendish is produced for export and local consumption worldwide in small farms, as well as in extensive plantations [100].
Anthracnose caused by Colletotrichum spp. is an important postharvest disease of bananas. Colletotrichum species infect banana in plantations and become latent pathogens. Bananas are often harvested before ripening; during storage, as the fruits ripen, anthracnose symptoms appear as brown or black lesions. Later, these lesions enlarge, become sunken, and produce spore masses. Wounds and scratches on banana peel caused by handling and transportation enhance the occurrence of anthracnose symptoms [101], which greatly impair the quality of bananas for export and local consumption.
Colletotrichum musae has long been associated with banana anthracnose worldwide [23,102,103,104,105,106,107]. In addition to anthracnose, C. musae can also cause stem-end, crown, and blossom-end rots in bananas [108]. Since C. musae is prevalent in bananas, Vieira et al. (2017) [23] suggested that this species might be host-specific to the plant. However, a detailed study on a global scale is required to confirm this hypothesis. Vieira et al. (2017) [23] have also developed C. musae species-specific primers for the rapid identification of this fungal species; this strategy results in cost savings compared to the sequencing of multiple genes. A study carried out by Li et al. (2019) [35] on mango anthracnose identified C. musae as one of the species involved, indicating that Colletotrichum species may not be host-specific to banana, as previously considered.
Among the main banana-producing countries, detailed studies on anthracnose pathogens were only conducted in Brazil, where five species were found to be associated with the disease [23]. Colletotrichum musae is still the most prevalent species reported in Brazil. Information regarding Colletotrichum spp. causing banana anthracnose in India, China, the Philippines, and Ecuador is rather scarce. The available data mainly consist of disease reports or newly recorded species; only one or two species are commonly reported to be involved.
Colletotrichum species causing banana anthracnose in Brazil were found to be C. siamense, C. tropicale, C. chrysophilum, and C. theobromicola [23]. Colletotrichum scovillei was reported in China [21], C. siamense in India [22] and Turkey [25], C. gloeosporioides in Ecuador [24] and C. chrysophilum in Mexico [26]. Other reported species include C. paxtonii [12], C. karstii [46], C. gloeosporioides sensu lato [20], and an undescribed species assigned to C. siamense sensu lato clade [106].
One of possible reasons lack of comprehensive studies on banana anthracnose might be many studies focusing on other diseases affecting bananas that are known to cause significant yield losses, such as wilt caused by Fusarium oxysporum Tropical Race 4, and Moko disease caused by Ralstonia solanacearum.
3. Mango Anthracnose
Mango (Mangifera indica L.) is planted mostly in Asia, particularly in India, which contributes to about 50% of the world’s mango production, followed by China, Thailand, Pakistan, and Indonesia. Brazil and Mexico are the largest mango producers in America, while Nigeria and Egypt are major producers in Africa [109]. For many of these countries, mango production is economically relevant.
Almost all mango cultivars grown in these countries are susceptible to anthracnose due to high temperature and humidity that characterize these tropical regions. The incidence of fruit anthracnose is almost 100% under wet conditions [110]. Not only fruits, but also the leaves, twigs, and flowers are affected by mango anthracnose. Leaf symptoms comprise black necrotic spots with irregular shapes on both sides of the leaves. Similar symptoms can appear on twigs and flowers. These black necrotic spots may coalesce to form larger infected areas. Infected tissues become dry; eventually, the infected parts of the plant die [5,110].
Colletotrichum gloeosporioides sensu lato is an important pathogen responsible for mango anthracnose worldwide [5,93,111]. In some cases, C. acutatum sensu lato has also been reported to be associated with mango anthracnose [5,112]. Phylogenetic analysis based only on internal transcribed spacer (ITS) sequences shows that C. gloeosporioides consists of diverse groups or species sub-populations, suggesting that other Colletotrichum spp. might be associated with mango anthracnose [111,113]. In contrast, several studies show that Colletotrichum isolates obtained from mango may consist of pathogenically and genetically distinct populations of C. gloeosporioides [114,115,116]. According to Ploetz (1999) [117], the C. gloeosporioides population on mango has restricted host range and is highly virulent only on mango.
After the reports published by Phoulivong et al. (2012) [16] and Weir et al. (2012) [13] describing phylogenetic analyses of the genus Colletotrichum using multiple markers, several species within C. gloeosporioides and C. acutatum complexes (including C. gloeosporioides) were reported to be associated with mango anthracnose. In addition, comprehensive studies on mango anthracnose pathogens using multilocus phylogenetic analysis were conducted in Brazil by Lima et al. (2013) [28] and Sharma et al. (2013) [29], in South China by Mo et al. (2018) [34] and in Mexico by Tovar-Pedraza et al. (2020) [37]. Other data on anthracnose-causing agents in mango have been communicated as first reports or disease notes.
In northeastern Brazil, five Colletotrichum species, namely C. asianum, C. fructicola, C. tropicale, C. karstii, and C. dianesei were found as anthracnose pathogens in mango; all species were reported for the first time in mango in Brazil [28]. Multiple genetic markers (glyceraldehyde-3-phosphatedehydrogenase (GAPDH), actin, β-tubulin, calmodulin, glutamine synthetase (GS), and the ITS region) were used to identify such species. Interestingly, only C. karstii could not infect two mango cultivars (Keith and Palmer) in a pathogenicity test carried out by Lima et al. (2015) [118], indicating that preference for certain mango cultivars may exist among Colletotrichum species. However, these authors found no host specificity in a cross-pathogenicity test, which included papaya, banana, guava, and bell pepper, indicating that these species have a broad host range [118].
Pardo-De la Hoz et al. (2016) [32] reported some level of host preference among Colletotrichum spp. associated with mango anthracnose in Colombia, which included C. asianum and C. gloeosporioides. Both species have also been reported as common mango anthracnose pathogens by Rojas et al. (2010) [27] in Panama and Krishnapillai and Wijeratnam (2014) [30] in Sri Lanka.
Many reports published in India, the largest mango producer in the world, point out C. gloeosporioides as the main causative agent of mango anthracnose, which, in some cases, might not be accurate. Based on restriction analysis and sequencing of the ITS region, Chowdappa and Kumar (2012) [111] reported that C. gloeosporioides associated with mango anthracnose in India comprise diverse subgroups. Pathogenicity tests demonstrate variation in the degree of virulence among C. gloeosporioides isolates, suggesting the existence of more than one species causing the disease. By using multigene phylogenetic analysis, Sharma et al. (2013) [29] later identified four phylogenetic species, namely, C. fragariae sensu stricto, C. fructicola, C. jasmine-sambac, and C. melanocaulon. Besides, five Colletotrichum lineages without species names were associated with mango anthracnose in India. Sharma et al. (2013) [29] also reported that none of the Colletotrichum isolates obtained from mango samples group with C. gloeosporioides sensu stricto, and are in line with the findings previously documented by Phoulivong et al. (2012) [16].
A multigene phylogenetic analysis carried out by Mo et al. (2018) [34] in different parts of Guangxi, south China, shows that three species of the C. gloeosporioides complex are pathogenic to mango fruits and its leaves. These species were identified as C. asianum, C. fructicola, and C. siamense. Later, Qin et al. (2017) [33] reported C. scovillei, a species within the C. acutatum complex, as another anthracnose-causal pathogen in mango leaves in Guangxi, China.
Among species in the C. gloeosporioides complex, C. asianum is the most common anthracnose pathogen in mango worldwide. This species has been reported in Brazil [28], Sri Lanka [30], Sanya City [33], and other areas of China [58], South Africa [119], Malaysia [31], Taiwan [36], Mexico [37], the Philippines [38] and Indonesia [120].
More Colletotrichum spp. were identified in association with mango anthracnose; this might be related to larger sampling areas, which may have allowed the access to more diseased-mango individuals and the detection of more isolates. In southern China, study on Colletotrichum spp. associated with mango anthracnose was reported by Li et al. (2019) [35], who analyzed infected mangoes from six provinces, Fujian, Guangdong, Guizhou, Hainan, Sichuan, and Yunan. In the study, 13 species are associated with mango anthracnose: C. asianum, C. cliviicola, C. cordylinicola, C. endophytica, C. fructicola, C. gigasporum, C. gloeosporioides, C. karstii, C. liaoningense, C. musae, C. scovillei, C. siamense, and C. tropicale. Two species, C. asianum and C. siamense, are the most common species identified, each accounting for 30% of the total species. Colletotrichum cordylinicola, C. endophytica, C. gigasporum, C. liaoningense, and C. musae were the first reported Colletotrichum spp. associated with mango anthracnose [35].
Wu et al. (2020) [36] reported on Colletotrichum spp. associated with mango anthracnose in Taiwan. These authors identified C. asianum, C. fructicola, C. siamense, C. tropicale, and C. scovillei, which were some of the species previously recognized by Li et al. (2019) [35] in China. Another comprehensive study of mango anthracnose was reported by Tovar-Pedraza et al. (2020) [37] in Mexico. Five species were identified using mating type Mat1-20 (ApMat) marker, namely C. alienum, C. asianum, C. fructicola, C. siamense, and C. tropicale. In terms of virulence, C. alienum and C. fructicola were the least virulent whereas C. siamense and C. asianum were the most virulent. Some of the species except C. aleinum have been reported as causal pathogens of mango anthracnose. Colletotrichum alienum was the first reported species associated with mango anthracnose worldwide at the time the report was published [37]. Later, Ahmad et al. (2021) [39] reported that C. alienum was associated with mango fruits cv. Jin-Hwang anthracnose in Beijing, China.
To date, 17 species of Colletotrichum are associated with mango anthracnose worldwide. By using, multiple markers phylogenetic analysis, it is likely more new species will be reported from other mango producing countries.
4. Papaya Anthracnose
It is widely accepted that papaya (Carica papaya L.) originated in Central America and southern Mexico. Currently, this fruit crop is planted commercially in many tropical countries, with India as the leading producer, followed by Brazil, Mexico, Indonesia, and the Dominican Republic [121]. Other papaya-producing countries are Thailand, the Philippines, China, Peru, and Nigeria.
Papaya is a climacteric fleshy fruit commonly harvested at the pre-climacteric stage. Therefore, the ripening stage starts after harvest, and the shelf life is relatively short. Several fruit rot pathogens may cause fruit damage during this period, including Colletotrichum spp. Though papaya anthracnose symptoms may occur both in fruits and leaves, Colletotrichum infection is usually more severe in fruits. Refrigerated papayas are particularly susceptible to disease; thus, fruits intended for export may develop anthracnose symptoms as they ripen [122].
In earlier studies, particularly in those conducted before application of multiple markers became usual tools for identification, C. gloeosporioides and C. capsici were indicated as common anthracnose pathogens in papaya in several regions and countries, including the Yucatan peninsula, Mexico [40]; Malaysia [41]; the Miyako Islands, Okinawa, Japan [42]; South Florida [43]; and Trinidad Island [47,48]. In addition to C. gloeosporioides and C. capsici, C. dematium has been found to be the causative agent of papaya anthracnose in Yucatan, Mexico [45].
To date, several species belonging to C. gloeosporioides, C. truncatum, C. magnum, and C. orchidearum complexes have been reported to be associated with papaya anthracnose, indicating that more than two Colletotrichum species are involved. Given that C. capsici is now a synonym of C. truncatum, the former name is no longer used in many publications.
In Australia, C. acutatum, C. simmondsii and C. queenslandicum have been described on papaya [12] while C. siamense in South Africa [13] and in China [61]. In India, using multigene phylogenetic analysis, several species have been identified as papaya anthracnose pathogens. The species reported include C. karstii [49], C. fructicola, C. gloeosporioides [51], and C. salsolae [53]. Most of the species reported are not host-specific and occur in many locations in papaya-producing countries.
Three species, namely, C. magna, C. gloeosporioides [44], and C. brevisporum [50], were reported as causative agents of papaya fruit rot and papaya anthracnose in Brazil. Although Nascimento et al. (2010) [44] referred to the disease as papaya fruit rot, Colletotrichum spp. were isolated from the lesions that constitute typical anthracnose symptoms, sometimes described as chocolate spots. Colletotrichum karstii has also been reported from papaya [46]. Papaya anthracnose caused by C. truncatum and C. okinawense was reported by Vieria et al. (2020) [123] and Dias et al. (2020) [59], respectively. Both species are the latest report on papaya anthracnose in Brazil.
Colletotrichum brevisporum and C. plurivorum have also been reported as a pathogens of papaya anthracnose in Taiwan. These pathogens were recovered from anthracnose lesions found on papaya fruits [55,57]. In addition to Taiwan and Brazil, C. brevisporum was also a causal pathogen of papaya anthracnose in China [58]. Another species, C. okinawense, which was first reported in Brazil, was found to be associated with papaya anthracnose in Taiwan [60].
Colletotrichum magnum was identified as causative agents of papaya anthracnose in Mexico [40] and Costa Rica [54]. Using the ITS region and specific primers, Molina-Chaves et al. (2017) [54] depicted C. truncatum, C. gloeosporioides sensu lato, and C. magnum as pathogens of papaya anthracnose in Guácimo, Costa Rica. Colletotichum trucatum has also been reported as papaya anthracnose in Korea [56].
Comprehensive studies focusing on the causative pathogens of papaya anthracnose are still lacking. Most available studies are either first disease reports or disease notes. A study on genetic variation in C. magnum was conducted by Pérez-Brito et al. (2018) [124] as an attempt to understand pathogenicity patterns and response to different fungicides. One of the reasons that may account for the lack of comprehensive studies focused on papaya anthracnose may be the occurrence of viral and bacterial diseases in papaya under field conditions, which frequently results in plant decay and decreased yields. Among viral diseases, papaya ringspot caused by the Papaya ringspot virus (PRSV) is the most serious and has been detected in many papaya producing countries in the tropics, as well as in subtropical areas. As for bacterial diseases, papaya dieback is a very destructive disease; and 100% yield losses have been recorded in Malaysia due to this pathogen [125].
In studies on control methods of papaya anthracnose, “C. gloeosporioides” is widely used as the causal pathogen. This should be treated with cautious as there are several other Colletotrichum species within C. gloeosporioides complex associated with papaya anthracnose (Table 1).
5. Dragon Fruit Anthracnose
Dragon fruit (Hylocereus sp.) is believed to have originated in Central and South America, and now this fruit crop is widely cultivated in many countries, including Vietnam, China, Mexico, Colombia, Nicaragua, Ecuador, Thailand, Malaysia, Indonesia, Australia, and United States [126]. China has also started a large-scale planting of dragon fruit, with 20,000 ha distributed in Guangdong and Guangxi provinces. Currently, the main dragon fruit producer is Vietnam, followed by Thailand, Taiwan, the Philippines, Malaysia, Sri Lanka, Australia, and Israel. In South America, dragon fruit is cultivated in Mexico, Ecuador, Colombia, Nicaragua, and Guatemala [127]. Two common species of cultivated dragon fruits are Hylocereus polyrhizus (red-fleshed) and Hylocereus undatus (white-fleshed).
The name “dragon fruit” probably derives from the fruit’s appearance, characterized by the presence of bracts or scales in the outer part [128]. This fruit is also known by other local names including strawberry pear or night-blooming cereus (English-speaking regions), pitahaya (Latin America), buah naga or buah mata naga (Malaysia), thanh long (Vietnam), kaeo mangkon or luk mangkon (Thailand), päniniokapunahou or päpipi pua (Hawaii), and paw wong fa kor (China).
Dragon fruit is often eaten fresh; its white, purple, or red flesh has a sweet taste (particularly the last one). Apart from being served as fruit salad, dragon fruit is used to flavor juices, sorbets, jams, yogurts, ice creams, jellies, candy, and dried fruit. Flower buds are used to make soups, or can be mixed in salads and tea preparations [129]. Besides, nutritional benefits have been assigned to this fruit as it contains vitamin C and other antioxidant metabolites, including betalains, flavonoids, and hydroxycinnamates, as well as fiber, iron, and magnesium.
Dragon fruit anthracnose caused by Colletotrichum affects the stems and fruits of Hylocereus spp. In earlier studies, C. gloeosporioides sensu lato is reported as the most common anthracnose pathogen in Hylocereus megalanthus in Brazil [64]; H. undatus is reported to be common in Okinawa Prefecture, Japan [62] and in Miami-Dade County, Florida, USA [63]. Colletotrichum gloeosporioides sensu lato has also been reported as an anthracnose-causing pathogen affecting the stems and fruits of H. polyrhizus, H. undatus, and Selenicereus megalanthus in Malaysia [65]. Using only ITS sequences, Lin et al. (2017) [70] identified three species, C. gloeosporioides, C. truncatum, and C. boninense, as anthracnose agents in H. polyrhizus, H. undatus, and H. costaricensis plants growing in several counties in Taiwan.
After the application of multiple markers for the identification of Colletotrichum species, several members within the C. gloeosporioides complex have been reported as anthracnose pathogens in Hylocereus spp. Ma et al. (2014) [66] reported C. gloeosporioides as an anthracnose pathogen in the young stems of H. undatus in China. In a later study, Zhao et al. (2018) [71] found C. siamense to be the causative agent of stem anthracnose in H. polyrhizus in China. Besides, C. aenigma and C. siamense are reported to be associated with stem and fruit anthracnose in H. undatus grown in Pathum Thani, Nakhon Pathom, and Samut Sakhon, Thailand [68]. Colletotrichum siamense is also responsible for fruit anthracnose in H. undatus growing in the Andaman Islands, India [72]. In Brazil, C. karstii was reported to be the causal pathogen of H. undatus stem anthracnose [73].
In addition to species within the C. gloeosporioides complex, C. truncatum is also reported as an anthracnose pathogen in Hylocereus spp. Colletotrichum truncatum is the causative agent of H. polyrhizus stem anthracnose in Malaysia [69], and H. undatus was also reported in fruits sold in a market in Yuanjiang County, Yunnan Province, China [67].
Seven Colletotrichum species were identified as causal anthracnose pathogens of different types of dragon fruits (Table 1). However, information on anthracnose pathogens associated with fruits and stems in several main producing countries including Vietnam, Indonesia and Si Lanka is still lacking.
6. Guava Anthracnose
Guava (Psidium guajava L.) is grown for its edible fruits that are rich in vitamin C and dietary fiber. Guava fruits are consumed fresh or as industrialized products, including purées, jams or marmalades, jellies, fruit pastes, juice, syrup, candy, and chutneys [130]. In addition, guava leaves are used in folk medicine owing to their medicinal properties that are useful in treating many ailments such as diarrhea, dysentery, gastroenteritis, hypertension, and diabetes, and to improve locomotor coordination [131].
This tropical fruit crop is native to Mexico, Central America, and South America, and receives different local names depending on the zones. For instance, it is known as jambu batu in Malay, amrood in Hindi, perakka in Malayalam, and farang in Thai. In French-speaking regions, guava is known as goyave or goyavier; Hawaiians call it kuawa, and in Portuguese-speaking areas, the fruit receives the name of goiaba or goiabeira [132].
India is the leading guava producer, with an estimated production of 17,650,000 metric tons annually, followed by Thailand and China. Other guava-producing countries are Pakistan, Mexico, Indonesia, Brazil, the Philippines, and Nigeria [133].
All guava-growing areas around the world are subjected to guava anthracnose. Fungi responsible for this disease infect guava fruits during pre- and post-harvest stages, particularly during high rainfall and high humidity periods. Young guava developing-flowers and fruits may also be infected. Anthracnose symptoms are obvious in mature fruits in the field, as well as in harvested fruits. Similar to that observed in other fruit crops, guava anthracnose symptoms consist mainly of sunken, dark necrotic lesions on the fruit surface. Spore masses are formed in these lesions under humid conditions [77].
Colletotrichum gloeosporioides sensu lato has been reported as a common anthracnose pathogen in several guava-growing countries [74,75,77,83]. In Hawaii, C. gloeosporioides sensu lato has also been reported to infect guava leaves [76]. Intan Sakinah et al. (2014) [78] reported C. gloeosporioides sensu lato as the most common species causing anthracnose disease in guava fruit, but suggest that other species of the C. gloeosporioides complex may also be associated with guava anthracnose. Colletotrichum acutatum has also been reported to cause anthracnose disease in guava [134,135,136].
More recently, multiple gene phylogeny studies for the identification of Colletotrichum spp. indicate that several species belonging to C. gloeosporioides and C. acutatum complexes are associated with guava anthracnose. The species of the C. acutatum complex reported include C. simmondsii in Brazil [79], C. abscissum in Brazil and the USA [80,82], and C. guajavae in India [12]. Among the species of the C. gloeosporioides complex associated with guava, C. psidii was detected in Italy [13] and C. siamense in India [81]. In a pathogenicity study performed by Bragança et al. (2016) [82], C. nymphaeae isolated from apple fruits in Brazil could cause lesions on guava fruits, demonstrating the cross-pathogenicity of this species.
To summarize, several Colletotrichum species are found to be associated with guava anthracnose. The information here provided may be useful for the development of integrated disease management to control guava anthracnose, as some of the species involved have a wide host range.
7. Avocado Anthracnose
Avocado (Persea americana Mill.) is a common tropical fruit. This plant species originated in Central America, more specifically, in Mexico and Guatemala. Mexico is the main producer and exporter of avocado, followed by Netherlands, Peru, Spain, Chile, and Colombia [137]. Among other producing avocado countries are India, Indonesia, Israel, China, Kenya, Vietnam, the Philippine, Australia and New Zealand.
Avocado is considered a rich source of nutrients, particularly fatty acids such as oleic acid and palmitic acid, minerals, and vitamins. The plant also contains phytochemicals like tannins, alkaloids, phenols, saponins, and flavonoids, as well as lutein, which is the predominant carotenoid in avocado fruits [138]. Due to the presence of those compounds and many other phytochemicals, avocado has shown numerous medicinal properties, including antimicrobial, anti-inflammatory, analgesic, antihypoglycemic, antihypertensive, antihepatotoxic, anticonvulsant, and vasorelaxant effects [138].
Anthracnose may occur in avocado wherever this fruit crop is grown, particularly during the wet season and in high rainfall areas. Major infections occur on the fruit; however leaves and stems can also become infected. The dark lesions of variable size produced by anthracnose pathogens tend to expand rapidly on the fruit skin and also infect the pulp, causing rot [84].
Before the use of multiple gene phylogeny for the identification of Colletotrichum spp., C. gloeosporioides sensu lato was the most common species found in association with avocado anthracnose, followed by C. acutatum sensu lato [84]. However, based on molecular analysis, C. gloeosporioides was also reported as the causative agent of avocado fruit anthracnose in Mexico [85], Mersin Province, Turkey [88], and Ghana [90]. Hunupolagama et al. (2015) [86] identified C. gigasporum as a causative agent of avocado anthracnose in Sri Lanka based on four markers, ITS, actin (ACT), GAPDH, and β-tubulin. In Mexico, two species C. godetiae [87] and C. karstii [139] were identified as anthracnose pathogens on avocado. Both species were identified using ITS and GAPDH sequences.
A comprehensive study on Colletotrichum spp. associated with avocado anthracnose was conducted in Israel [91]. Using multiple genes/markers (ITS, ACT, ApMat, calmodulin [CAL], chitin synthase [CHS1], GAPDH, GS, HIS3, and β-tubulin), Sharma et al. (2017) [91] identified nine Colletotrichum species. Eight of these species, C. aenigma, C. alienum, C. fructicola, C. gloeosporioides sensu stricto, C. karstii, C. nupharicola, C. siamense, and C. theobromicola, had been reported before as avocado anthracnose pathogens in other avocado-producing countries. A new species, C. perseae, is reported in association with avocado anthracnose for the first time. While C. aenigma is the most virulent species in Israel, C. perseae sp. nov. is considered most dominant.
Some other studies conducted in different avocado-producing countries reported either the same species or different Colletotrichum species identified by Sharma et al. (2017) [91], to be responsible for avocado anthracnose. Fuentes-Aragón et al. (2018) [92] reported C. fructicola as the causal pathogen of avocado anthracnose in Hidalgo, Mexico and Giblin et al. (2018) [93] isolated and identified five species previously considered as C. gloeosporioides sensu lato from avocado fruit in eastern Australia: C. alienum, C. asianum, C. fructicola, C. karstii, and C. siamense. Shivas et al. (2016) [89] also reported the presence of C. alienum, C. fructicola, and C. siamense in avocado in Australia. Kwon et al. (2020) [94] identified C. kahawae subsp. cigarro as the isolate obtained from an imported avocado variety in a market in Jinju, South Korea. Uysal and Kurt (2020) [25] reported C. karstii as causal pathogen of avocado fruit and leaf anthracnose in Turkey. In south eastern Brazil, C. siamense and C. karstii were found to be associated avocado anthracnose [97].
Another comprehensive study on pathogen of avocado anthracnose was performed by Fuentes-Aragón et al. (2020) [96]. Using six markers (GAPDH, ITS, ACT, CHS-1, ApMat and β-tubulin), the study indicated 11 species were the causal pathogens of avocado anthracnose in Mexico, namely C. karstii, C. godetiae, C. siamense, C. fioriniae, C. cigarro, C. chrysophilum, C. jiangxiense, C. tropicale, C. nymphaeae), and two new lineages designated as Colletotrichum sp. 1 and Colletotrichum sp. 2. The most prevalent species was C. siamense and the most widespread was C. karstii.
According to existing reports, Colletotrichum species associated with avocado anthracnose are similar to those reported in other tropical fruit crops. They include C. asianum, C. fructicola, and C. siamense, species known to infect a wide range of hosts.
So far, there are a lack of reports on the Colletotrichum species associated with avocado anthracnose from other major avocado producing countries, including Indonesia, Dominican Republic, Peru, and Venezuela. Many studies on control methods in these countries referred to C. gloeosporioides sensu lato as the causal pathogen of avocado anthracnose, which might not be accurate.
8. Present and Future Management of Anthracnose
Fungicides, chemicals (e.g., benzimidazoles such as thiabendazole, benomyl, and carbendazim), and sterol inhibitors (e.g., imazalil, prochloraz, and propiconazole) have long been used to effectively control anthracnose disease in banana, mango, papaya, and avocado plants [140]. Benzimidazoles are often applied as dips or sprays to inhibit the anthracnose-causing fungus, Colletotrichum spp. [141], but large-scale and continuous fungicide use has led to fungal-resistance. For example, benzimidazole-resistant Colletotrichum has been detected in mangos and bananas [140]. The excessive use of fungicides also negatively affects human health and the environment, as chemical residues often contaminate the soil and water [142]. Biocontrol is an alternative, non-toxic method for controlling fruit crop anthracnose. Biocontrol agents (i.e., antagonistic microbes) such as yeast, bacteria, and filamentous fungi (particularly Trichoderma spp.) have shown promising results and are gaining popularity because of their direct post-harvest application to fruit surfaces [143].
Yeasts (unicellular fungi) have several characteristics that make them a desirable biocontrol agent. They grow rapidly on a wide range of substrates and have a high reproductive rate and simple nutritional requirements. Moreover, yeasts are not mycotoxigenic and can grow in high-sugar environments [144,145,146,147]. There is also an increasing demand for chemical-free or reduced chemical treatments to control anthracnose, which has led to the development of alternative methods that are safer for consumers (e.g., edible coatings from chitosan and essential oils). Generally recognized as safe (GRAS) salt treatments, nanomaterials, and cold plasma technology have also been explored. Often, these alternative approaches are used in combination for more effective anthracnose pathogen growth inhibition and disease severity reduction.
Chitosan emerged as a target for edible coating formulations because of its antifungal properties, and it is often combined with other compounds (i.e., essential oils). The antifungal efficacy of chitosan in solution (conventional chitosan) and chitosan in submicron dispersion were tested against C. gloeosporioides sensu lato, a dragon fruit anthracnose pathogen, by Asgar et al. (2013) [147]. When applied to the fruit, the chitosan treatments reduced the anthracnose symptoms and disease development. Combining chitosan and Cymbopogon citratus essential oil also has inhibitory effects against five anthracnose pathogens (C. asianum, C. siamense, C. fructicola, C. tropicale, and C. karstii) when inoculated on guava, mango, and papaya [148]. Braga et al. (2019) [149] combined chitosan and peppermint essential oils (Mentha piperita L and Mentha x villosa Huds) and documented anthracnose pathogen growth inhibition (C. gloeosporioides and C. brevisporum) on papaya in vitro and reduced anthracnose lesions after 10 days of storage.
GRAS inorganic and organic salts used to preserve food have been evaluated as an edible coating for anthracnose pathogen control and to reduce the amount of rotting fruit. Carbonates, sorbates, benzoates, and silicates have low toxicological effects and antifungal properties, and satisfactory results have been reported for anthracnose pathogen inhibition [150]. These results suggest that GRAS salts may be an alternative to post-harvest pathogen management.
De Costa and Gunawardhana (2012) [151] found that sodium bicarbonate reduced appressorium formation, spore production, germination, and pathogen mycelial growth of the banana anthracnose pathogen, C. musae, in vitro. Anthracnose lesions were also reduced by dipping the fruit into a 300 mM salt solution for 10 min. Jitareerat et al. (2018) [152] showed that sodium carbonate and potassium sorbate inhibited C. gloeosporioides and C. capsici spore germination. Moreover, when the fruit was placed in a potassium sorbate and hot water solution (55 °C for 5 min) and cooled in water, the disease severity was reduced without affecting the fruit quality. Kalupahana et al. (2020) [153] tested the effectiveness of sodium bicarbonate and sodium metabisulfite against the mango anthracnose pathogen, C. siamense, and found that both salts inhibited mycelial growth.
Nanomaterials, such as copper, silver, nickel, and magnesium, have antifungal properties and may be effective at managing anthracnose pathogens and post-harvest disease [154]. The efficacy of zinc oxide, magnesium oxide, and their composites (52–219 nm) were tested against papaya and avocado C. gloeosporioides. Conidial germination was inhibited and the fungal cells were damaged, indicating that the nanomaterials had an antifungal effect [154]. This was supported by Jagana et al. (2017) [155], who reported that copper, silver, nickel, and magnesium (68 nm) extracted from the leaves of the medicinal plants ajwain (Trachyspermum ammi) and neem (Azadirachta indica) inhibited the spore germination of C. musae isolated from banana. The severity of banana anthracnose was also reduced with 0.2% silver-neem.
Nanomaterials composite with other materials can also control mango anthracnose. Antifungal properties were reported when chitosan-silver composite (495–616 nm diameter) was used, which suppressed C. gloeosporioides conidial germination. An in-vivo study reported that 0.5% and 1% nanomaterial composite reduced anthracnose disease by 45.7% and 71.3%, respectively [156]. Neem extract was used to synthesize copper oxychloride-conjugated silver (21–25 nm) and treat C. gloeosporioides, resulting in pathogen growth suppression [157].
Cold plasma technology is another approach to inhibit anthracnose pathogens in tropical fruit. Cold plasma is a partially ionized gas, where a small subset of atoms and molecules are ionized by electrical discharges at atmospheric or sub-atmospheric pressure [158,159]. Studies using cold plasma technology have been performed on spoilage and mycotoxigenic fungi-contaminated food and feed with promising outcomes [159]. Siddique et al. (2018) [160] isolated C. alienum and C. fioriniae from avocados and treated them with cold plasma for 180 s or 360 s in open and sealed environments. In some treatments, the colony growth was reduced, and the conidial germination was inhibited, suggesting that cold plasma treatment may be an effective control for C. alienum and C. fioriniae in avocado. Cold plasma has also been used to decontaminate fruit containers and packaging. Misra et al. (2014) [161] used two gas mixtures (65% O2 + 16% N2 + 19% CO2 and 90% N2 + 10% O2) to decontaminate packaged and sealed strawberries, which reduced the microflora level from 5 to 3.0 log10 CFU/g in 300 s with no post-treatment changes to the packaging material.
Alternative methods to reduce post-harvest fruit crop losses are ongoing. Biocontrol agents, edible fruit coatings, and GRAS salt, cold plasma, and nanomaterial treatments have shown promising results, but they are not without challenges. The performance varies among fruit crops, and the formulations and costs need further investigation. Moreover, some methods are combined to improve efficacy, creating other issues, such as public and industry acceptance, product registration, and commercial viability.
9. Conclusions and Future Directions
Previously, anthracnose pathogens are often referred to as C. gloeosporioides or C. acutatum because, in many cases, the identification procedures did not include the use of multiple markers, and frequently, only the ITS region was analyzed. Thus, the data obtained may not reflect the true causal pathogens. Moreover, it is now accepted that C. gloeosporioides is not the most common anthracnose pathogen in tropical fruit crops, as previously thought.
Various Colletotrichum species can cause anthracnose in tropical fruit crops, thus becoming serious limiting factors in the production and marketing of these commodities. Table 2 shows diverse species of Colletotrichum associated with anthracnose of banana, papaya, mango, dragon fruits, guava, and avocado. Some of the Colletotrichum species not only infected the fruits but the stem and leaves as well indicated that other parts of the plants harbor inoculum sources for anthracnose infection on fruit crops. Several species including C. siamense, C. asianum, C. scovillei, C. gloeosporioides, C. karstii, C. fructicola, and C. tropicale can infect multiple hosts, demonstrated the possibility of cross infection to various types of fruit crops as well as other crops.
Since molecular phylogenetic analysis was applied for identification and characterization of Colletotrichum species, diverse species were reported to be associated with anthracnose of tropical fruits (Table 2). Many of the Colletotrichum species listed in Table 2 belong to different species complexes including C. gloeosporioides, C. acutatum and C. boninense complexes. Species in a species complex are closely related, and have similar behavior of host infection and colonization [12,13,46]. Thus, infection and colonization of various Colletotrichum spp. on different tropical fruit crops are also similar. In terms of virulence, anthracnose symptoms on different fruits may vary depending on the variety of the fruits, inoculum concentration, humidity and temperature [5]. Moreover, pathogenic variation of Colletotrichum spp. infected fruit crops has been demonstrated [162,163].
Diverse species of Colletotrichum causing anthracnose of fruit crops are also a quarantine concern. Banana, papaya, mango, dragon fruits, guava and avocado are exported and imported worldwide, and latent infection is part of the disease cycle of anthracnose pathogens. There are possibilities that the anthracnose pathogens can be distributed to other areas or regions. Therefore, it is important to document all the Colletotrichum spp. associated with anthracnose on different types of fruit crops.
Accurate identification and scientific name assignment of anthracnose pathogens are vital issues because precise taxonomic information enables us to classify a given species as a pathogen, saprophyte, or endophyte. The species involved in tropical fruits anthracnose may also have different presentations. It is well-known that effective disease management often depends on the proper identification of the causative pathogen.
The use of multiple markers allowed the recognition of an increasing number of Colletotrichum phylogenetic species, including species that cause anthracnose. However, for some of these phylogenetic species, information on the host range, pathogenicity, virulence variability, sensitivity to fungicides, and geographical distribution are still scarce. This situation may create a problem for plant pathologists, as many members of Colletotrichum are among the fungal species of quarantine concern in several countries. Keeping up to date with recently reported Colletotrichum species affecting tropical fruit crops is central to identify the risks posed by them.
Funding
Part of the research on anthracnose of Colletotrichum spp. in Malaysia was supported by a Research University Grant (1001/ PBIOLOGI / 811307) from the Universiti Sains Malaysia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
| ACT | actin |
| ApMat | mating type (Mat1-2) |
| CHS1 | chitin synthase |
| CAL | calmodulin |
| CFU | colony forming unit |
| GAPDH | glyceraldehyde-3-phosphatedehydrogenase |
| GRAS | generally recognized as safe |
| GS | glutamine synthetase |
| HIS3 | histone |
| ITS | internal transcribed spacer |
References
- OECD; FAO. OECD–FAO Agricultural Outlook 2019–2028, Part. II—Overview, Special Feature and Commodity Chapters; OECD Publishing: Paris, France, 2019. [Google Scholar]
- Altendorf, S. Minor. Tropical Fruits: Mainstreaming a Niche Market; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; pp. 69–76. [Google Scholar]
- OECD; FAO. OECD–FAO Agricultural Outlook 2020–2029; FAO: Rome, Italy; OECD Publishing: Paris, France, 2020. [Google Scholar]
- Altendorf, S. Major Tropical Fruits Statistical Compendium 2018; FAO: Rome, Italy, 2019. [Google Scholar]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Prusky, D. Pathogen quiescence in postharvest diseases. Annu. Rev. Phytopathol. 1996, 34, 413–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffries, P.; Dodd, J.C.; Jeger, M.J.; Plumbley, R.A. The biology and control of Colletotrichum species on tropical fruit crops. Plant Pathol. 1990, 39, 343–366. [Google Scholar] [CrossRef]
- Wharton, P.S.; Dieguez-Uribeondo, J. The biology of Colletotrichum acutatum. An. Jardín Botánico Madr. 2004, 61, 3–22. [Google Scholar]
- Talhinhas, P.; Mota-Capitao, C.; Martins, S.; Ramos, A.P.; Neves-Martins, J.; Guerra-Guimaraes, L.; Varzea, V.; Silva, M.C.; Sreenivasaprasad, S.; Oliveira, H. Epidemiology, histopathology and aetiology of olive anthracnose caused by Colletotrichum acutatum and C. gloeosporioides in Portugal. Plant Pathol. 2011, 60, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.W.; Emmanuel, C.J.; Emilda, D.; Terhem, R.B.; Shafia, A.; Tsamaidi, D.; Emblow, M.; Van, K.; Jan, A.L. Analysis of cryptic systemic Botrytis infections in symptomless hosts. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fitzell, R.D.; Peak, C.M. The epidemiology of anthracnose disease of mango: Inoculum sources, spore production and dispersal. Ann. Appl. Biol. 1984, 104, 53–59. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum—Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [Green Version]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Paul, P.W.J. Lifestyles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Phoulivong, S.; McKenzie, E.; Hyde, K. Cross infection of Colletotrichum species; a case study with tropical fruits. Curr. Res. Environ. Appl. Mycol. 2012, 2, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.J.; Sun, T.; Yang, Y.L.; Cai, L.; Hyde, K.D.; Bahkali, A.H.; Liu, Z.Y. Colletotrichum species on grape in Guizhou and Yunnan provinces, China. Mycoscience 2013, 54, 29–41. [Google Scholar] [CrossRef]
- Freeman, S.; Horowitz, S.; Sharon, A. Pathogenic and nonpathogenic lifestyles in Colletotrichum acutatum from strawberry and other plants. Phytopathology 2001, 91, 986–992. [Google Scholar] [CrossRef] [Green Version]
- Sanders, G.M.; Korsten, L. A comparative morphological study of South African avocado and mango isolates of Colletotrichum gloeosporioides. Can. J. Bot. 2003, 81, 877–885. [Google Scholar] [CrossRef]
- Intan Sakinah, M.A.; Suzianti, I.V.; Latiffah, Z. First report of Colletotrichum gloeosporioides causing anthracnose of banana (Musa spp.) in Malaysia. Plant Dis. 2013, 97, 991. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, J.S.; Yang, L.Y.; Wang, G.F.; Li, J.Q. First report of banana anthracnose caused by Colletotrichum scovillei in China. Plant Dis. 2016, 101, 381. [Google Scholar] [CrossRef]
- Kumar, V.S.; Nair, B.A.; Nair, P.V.R.; Annamalai, A.; Jaishanker, R.; Umamaheswaran, K.; Sooraj, N.P.; Peethambaran, C.K. First report of Colletotrichum siamense causing anthracnose of cliff Banana in India. Plant Dis. 2017, 101, 390. [Google Scholar] [CrossRef]
- Vieira, W.A.S.; Lima, W.G.; Nascimento, E.S.; Michereff, S.J.; Câmara, M.P.S.; Doyle, V.P. The impact of phenotypic and molecular data on the inference of Colletotrichum diversity associated with Musa. Mycologia 2017, 109, 912–934. [Google Scholar] [CrossRef] [PubMed]
- Riera, N.; Ramirez-Villacis, D.; Barriga-Medina, N.; Alvarez-Santana, J.; Herrera, K.; Ruales, C.; Leon-Reyes, A. First report of banana anthracnose caused by Colletotrichum gloeosporioides in Ecuador. Plant Dis. 2019, 103, 763. [Google Scholar] [CrossRef]
- Uysal, A.; Kurt, S. First report of fruit and leaf anthracnose caused by Colletotrichum karstii on avocado in Turkey. Crop. Prot. 2020, 133, 105145. [Google Scholar] [CrossRef]
- Fuentes-Aragón, D.; Rebollar-Alviter, A.; Osnaya-González, M.; Enciso-Maldonado, G.A.; González-Reyes, H.; Silva-Rojas, H.V. Multilocus phylogenetic analyses suggest the presence of Colletotrichum chrysophilum causing banana anthracnose in Mexico. J. Plant Dis. Prot. 2020, 1–7. [Google Scholar] [CrossRef]
- Rojas, E.I.; Rehner, S.A.; Samuels, G.J.; Van Bael, S.A.; Herre, E.A.; Cannon, A.; Chen, R.; Pang, J.; Wang, R.; Zhang, Y.; et al. Colletotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panama: Multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2010, 102, 1318–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, N.B.; Batista, M.V.A.; Morais Junior, M.A.; Barbosa, M.A.G.; Michereff, S.J.; Hyde, K.D.; Câmara, M.P.S. Five Colletotrichum species are responsible for mango anthracnose in northeastern Brazil. Fungal Divers. 2013, 61, 75–88. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar, N.; Weir, B.S.; Hyde, K.D.; Shenoy, B.D. Apmat gene marker can resolve Colletotrichum species: A case study with Mangifera indica. Fungal Divers. 2013, 61, 117–138. [Google Scholar] [CrossRef]
- Krishnapillai, N.; Wijeratnam, R.S.W. First report of Colletotrichum asianum causing anthracnose on Willard mangoes in Sri Lanka. New Dis. Rep. 2014, 29. [Google Scholar] [CrossRef] [Green Version]
- Latiffah, Z.; Nurul Zaadah, J.; Suzianti, I.V.; Intan Sakinah, M.A. Molecular characterization of Colletotrichum isolates associated with anthracnose of mango fruit. Sains Malays. 2015, 44, 651–656. [Google Scholar]
- Pardo-De la Hoz, C.J.; Calderon, C.; Rincon, A.M.; Cardenas, M.; Danies, G.; Lopez-Kleine Restrepo, S.; Jimenez, P. Species from the Colletotrichum acutatum, Colletotrichum boninense and Colletotrichum gloeosporioides species complexes associated with tree tomato and mango crops in Colombia. Plant Pathol. 2016, 65, 227–237. [Google Scholar] [CrossRef]
- Qin, L.P.; Huang, S.L.; Lin, S.H. First report of anthracnose of Mangifera indica caused by Colletotrichum asianum in Sanya city in China. Plant Dis. 2017, 101, 1038. [Google Scholar] [CrossRef]
- Mo, J.; Zhao, G.; Li, Q.; Solangi, G.S.; Tang, L.; Guo, T.; Huang, S.; Hsiang, T. Identification and characterization of Colletotrichum species associated with mango anthracnose in Guangxi, China. Plant Dis. 2018, 102, 1283–1289. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Bu, J.; Shu, J.; Yu, Z.; Tang, L.; Huang, S.; Guo, T.; Mo, J.; Luo, S.; Ghulam, S.S.; et al. Colletotrichum species associated with mango in southern China. Sci. Rep. 2019, 9, 18891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Chen, H.; Ni, H. Identification and characterization of Colletotrichum species associated with mango anthracnose in Taiwan. Eur. J. Plant Pathol. 2020, 157, 1–15. [Google Scholar] [CrossRef]
- Tovar-Pedraza, J.M.; Mora-Aguilera, J.A.; Nava-Díaz, C.; Lima, N.B.; Michereff, S.J.; Sandoval-Islas, J.S.; Câmara, M.P.S.; Téliz-Ortiz, D.; Leyva-Mir, S.G. Distribution and pathogenicity of Colletotrichum species associated with mango anthracnose in Mexico. Plant Dis. 2020, 104, 137–146. [Google Scholar] [CrossRef]
- Alvarez, L.V.; Hattori, Y.; Deocaris, C.C.; Mapanao, C.P.; Bautista, A.B.; Cano, M.J.B.; Naito, K.; Kitabata, S.; Motohashi, K.; Nakashim, C. Colletotrichum asianum causes anthracnose in Philippine mango cv. Carabao, Australas. Plant Dis. Notes 2020, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, T.; Wang, J.; Zheng, Y.; Mugizi, A.E.; Moosa, A.; Chengrong, N.; Liu, Y. First record of Colletotrichum alienum causing postharvest anthracnose disease of mango fruit in China. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Tapia-Tussell, R.; Quijano-Ramayo, A.; Cortes-Velazquez, A.; Lappe, P.; Larque-Saavedra, A.; Perez-Brito, D. PCR-based detection and characterization of the fungal pathogens Colletotrichum gloeosporioides and Colletotrichum capsici causing anthracnose in papaya (Carica papaya L.) in the Yucatan Peninsula. Mol. Biotech. 2008, 40, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Mahmud, T.M.M.; Kadir, J.; Abdul Rahman, R.; Begum, M.M. Major postharvest fungal diseases of papaya cv. ‘Sekaki’ in Selangor, Malaysia. Pertanika J. Trop. Agric. Sci. 2008, 31, 27–34. [Google Scholar]
- Yaguchi, Y.; Nakanishi, Y.; Saito, T.; Nakamura, S. Anthracnose of Carica papaya L. caused by Colletotrichum capsici. Ann. Phytopath. Soc. Jpn. 1995, 61, 222. [Google Scholar]
- Tarnowski, T.B.L.; Ploetz, R.C. First report of Colletotrichum capsici causing postharvest anthracnose on papaya in South Florida. Plant Dis. 2010, 94, 1065. [Google Scholar] [CrossRef]
- Nascimento, R.J.; Mizubuti, E.S.G.; Câmara, M.P.S.; Ferreira, M.F.; Maymon, M.; Freeman, S.; Michereff, S.J. First report of papaya fruit rot caused by Colletotrichum magna in Brazil. Plant Dis. 2010, 94, 1506. [Google Scholar] [CrossRef]
- Santamaría Basulto, F.; Díaz Plaza, R.; Gutiérrez Alonso, O.; Santamaría Fernández, J.; Larqué Saavedra, A. Control of two species of Colletotrichum causing anthracnose in Maradol papaya fruits. Rev. Mex. Cienc. Agríc. 2017, 2, 631–643. [Google Scholar]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Rampersad, S.N. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease of papaya in Trinidad. Plant Dis. 2011, 95, 1244–1254. [Google Scholar] [CrossRef] [Green Version]
- Maharaj, A.; Rampersad, S.N. Genetic Differentiation of Colletotrichum gloeosporioides and C. truncatum associated with anthracnose disease of papaya (Carica papaya L.) and bell pepper (Capsium annuum L.) based on ITS PCR-RFLP fingerprinting. Mol. Biotechnol. 2012, 50, 237–249. [Google Scholar] [CrossRef]
- Sharma, G.; Shenoy, B.D. Multigene sequence-based identification of Colletotrichum cymbidiicola, C. karstii and C. phyllanthi from India. Czech Mycol. 2013, 65, 79–88. [Google Scholar] [CrossRef]
- Vieira, W.A.S.; Nascimento, R.J.; Michereff, S.J.; Hyde, K.D.; Câmara, M.P.S. First report of papaya fruit anthracnose caused by Colletotrichum brevisporum in Brazil. Plant Dis. 2013, 97, 1659. [Google Scholar] [CrossRef] [PubMed]
- Saini, T.J.; Gupta, S.G.; Anandalakshmi, R. First report of papaya anthracnose caused by Colletotrichum fructicola in India. New Dis. Rep. 2016, 34, 7. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Tussell, R.; Cortes-Velazquez, A.; Valencia-Yah, T.; Navarro, C.; Espinoza, E.; Moreno, B.; Perez-Brito, D. First report of Colletotrichum magnum causing anthracnose in papaya in Mexico. Plant Dis. 2016, 100, 2323. [Google Scholar] [CrossRef]
- Saini, T.J.; Gupta, S.G.; Anandalakshmi, R. First report of papaya anthracnose caused by Colletotrichum salsolae in India. New Dis. Rep. 2017, 35, 27. [Google Scholar] [CrossRef] [Green Version]
- Molina-Chaves, A.; Gómez-Alpízar, L.; Umaña-Rojas, G. Identificación de especies del género Colletotrichum asociadas a la antracnosis en papaya (Carica papaya L.) EnCosta Rica. Agron. Costarric. 2017, 41, 69–80. [Google Scholar]
- Duan, C.H.; Pan, H.R.; Wang, C.C. First report of Colletotrichum brevisporum causing anthracnose on papaya in Taiwan. Plant Dis. 2018, 102, 2375–2376. [Google Scholar] [CrossRef]
- Aktaruzzaman, M.; Afroz, T.; Lee, Y.G.; Kim, B.S. Post-harvest anthracnose of papaya caused by Colletotrichum truncatum in Korea. Eur. J. Plant Pathol. 2018, 150, 259–265. [Google Scholar] [CrossRef]
- Sun, Y.C.; Damm, U.; Huang, C.J. Colletotrichum plurivorum, the causal agent of anthracnose fruit rot of papaya in Taiwan. Plant Dis. 2019, 103, 1040. [Google Scholar] [CrossRef]
- Liu, X.B.; Yanli, F.; Xiaolan, Z.; Huang, G.X. First report of papaya anthracnose caused by Colletotrichum brevisporum in China. Plant Dis. 2019, 103, 2473. [Google Scholar] [CrossRef]
- Dias, L.R.C.; Brito, R.A.S.; Melo, T.A.; Serra, I.M.R.S. First report of papaya fruit anthracnose caused by Colletotrichum okinawense in Brazil. Plant Dis. 2020, 4, 573. [Google Scholar] [CrossRef]
- Sun, Y.C.; Huang, C.J. Colletotrichum okinawense, the causal agent of postharvest anthracnose fruit rot of papaya in Taiwan. J. Plant Pathol. 2020, 102, 581–582. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, W.; Ning, P.; Guo, T.; Huang, S.P.; Tang, L.; Qili, L.; Mo, J. First report of anthracnose of papaya (Carica papaya L.) caused by Colletotrichum siamense in China. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Taba, S.; Mikami, D.; Takaesu, K.; Ooshiro, A.; Moromizato, Z.; Nakasone, S.; Kawano, S. Anthracnose of pitaya (Hylocereus undatus) by Colletotrichum gloeosporioides. Jpn. J. Phytopathol. 2006, 72, 25–27. [Google Scholar] [CrossRef]
- Palmateer, A.J.; Ploetz, R.C.; van Santen, E.; Correll, J.C. First occurrence of anthracnose caused by Colletotrichum gloeosporioides on Pitahaya. Plant Dis. 2007, 91, 631. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, L.M.; Rosa, D.D.; Basseto, M.A.; de Souza, H.G.; Furtado, E.L. First report of Colletotrichum gloeosporioides on Hylocereus megalanthus in Brazil. Australas. Plant Dis. 2008, 3, 96–97. [Google Scholar] [CrossRef] [Green Version]
- Masyahit, M.; Kamaruzaman, S.; Yahya, A.; Mohd Ghazali, M.S. The first report of the occurrence of anthracnose disease caused by (Penz.) Penz. & Sacc. on dragon fruit (Hylocereus spp.) in Peninsular Malaysia. Am. J. Appl. Sci. 2009, 6, 902–912. [Google Scholar]
- Ma, W.J.; Yang, X.; Wang, X.R.; Zeng, Y.S.; Liao, M.D.; Chen, C.J.; Sun, S.; Jia, D.M. First report of anthracnose disease on young stems of bawanghua (Hylocereus undatus) caused by Colletotrichum gloeosporioides in China. Plant Dis. 2014, 98, 991. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.W.; Wu, Y.X.; Ho, H.H.; Su, Y.Y.; Mao, Z.C.; He, P.F.; He, Y.Q. First report of dragon fruit (Hylocereus undatus) anthracnose caused by Colletotrichum truncatum in China. J. Phytopathol. 2014, 162, 272–275. [Google Scholar] [CrossRef]
- Meetum, P.; Leksomboon, C.; Kanjanamaneesathian, M. First report of Colletotrichum aenigma and C. siamense, the causal agents of anthracnose disease of dragon fruit in Thailand. J. Plant Pathol. 2015, 97, 391–403. [Google Scholar]
- Suzianti, I.V.; Intan Sakinah, M.A.; Latiffah, Z. Characterization and Pathogenicity of Colletotrichum truncatum causing stem anthracnose of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopathol. 2015, 163, 67–71. [Google Scholar]
- Lin, C.P.; Ann, P.J.; Huang, H.C.; Chang, T.T.; Tsai, J.N. Anthracnose of pitaya (Hylocereus spp.) caused by Colletotrichum spp., a new postharvest disease in Taiwan. J. Taiwan Agric. Res. 2017, 66, 171–183. [Google Scholar]
- Zhao, H.J.; Chen, S.C.; Chen, Y.F.; Zou, C.C.; Wang, X.L.; Wang, Z.H.; Liu, A.R.; Ahammed, G.J. First report of red dragon fruit (Hylocereus polyrhizus) anthracnose caused by Colletotrichum siamense in China. Plant Dis. 2018, 6, 862. [Google Scholar] [CrossRef]
- Abirami, K.; Sakthivel, K.; Sheoran, N.; Baskaran, V.; Gautam, R.K.; Jerard, B.A.; Kumar, A. Occurrence of anthracnose disease caused by Colletotrichum siamense on dragon fruit (Hylocereus undatus) in Andaman Islands, India. Plant Dis. 2019, 103, 768. [Google Scholar] [CrossRef]
- Nascimento, M.B.; Bellé, C.; Azambuja, R.M.; Maich, S.L.P.; Neves, C.G.; Souza-Junior, I.T.; Jacobsen, C.R.F.; Barros, D.R. First Report of Colletotrichum karstii causing anthracnose spot on pitaya (Hylocereus undatus) in Brazil. Plant Dis. 2019, 103, 2137. [Google Scholar] [CrossRef]
- Omar, A.A.W. Occurrence of Colletotrichum anthracnose disease of guava fruit in Egypt. Int. J. Pest. Manag. 2001, 47, 147–152. [Google Scholar]
- Amusa, N.A.; Ashaye, O.A.; Amadi, J.; Dapo, O.O. Guava fruit anthracnose and the effects on its nutritional and market values in Ibadan. Niger. J. Appl. Sci. 2005, 6, 539–542. [Google Scholar] [CrossRef]
- Keith, L.M.; Zee, F.T. Guava disease in Hawaii and the characterization of Pestalotiopsis spp. affecting guava. Acta Hortic. 2010, 849, 269–276. [Google Scholar] [CrossRef]
- Merida, M.; Palmateer, A.J. Florida Plant Disease Management Guide: Guava (Psidium guajava); PP-232. Plant Pathology Department, UF/IFAS Extension; Department of Agriculture, Cooperative Extension Service; Universidad de Florida: Gainesville, FL, USA, 2013. [Google Scholar]
- Intan Sakinah, M.A.; Suzianti, I.V.; Latiffah, Z. Molecular characterization and pathogenicity of Colletotrichum sp. from guava. Arch. Phytopathol. Plant Protect. 2014, 47, 1549–1556. [Google Scholar]
- Cruz, A.F.; Medeiros, N.L.; Benedet, G.L.; Araújo, M.B.; Uesugi, C.H.; da Ferreira, M.A.S.V.; Peixoto, J.R.; Blum, L.E.B. Control of post-harvest anthracnose infection in guava (Psidium guajava) fruits with phosphites, calcium chloride, acetyl salicylic acid, hot water, and 1-MCP. Hort. Environ. Biotechnol. 2015, 56, 330–340. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Hernández-Restrepo Sutton, D.A.; Acharya, K.; Barber, P.A.; Boekhout, T.; Dimitrov, R.A.; Dueñas Dutta, A.K.; Gené, J.; et al. Fungal planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar-Pinnaka, A.; Shenoy, B.D. Resolving the Colletotrichum Siamese species complex using ApMat marker. Fungal Divers. 2015, 71, 247–264. [Google Scholar] [CrossRef]
- Bragança, C.A.D.; Damm, U.; Baroncelli, R.; Massola Júnior, N.S.; Crous, P.W. Species of the Colletotrichum acutatum complex associated with anthracnose diseases of fruit in Brazil. Fungal Biol. 2016, 120, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Lan, C.; Huang, P.; Yu, D. PCR detection of Colletotrichum gloeosporioides in Psidium guajava. Australas. Plant Pathol. 2018, 47, 95–100. [Google Scholar] [CrossRef]
- Nelson, S. Anthracnose of avocado. Plant Disease. 2008. PD-58. Available online: https://www.ctahr.hawaii.edu/oc/freepubs/pdf/pd-58.pdf (accessed on 15 April 2020).
- Silva-Rojas, H.V.; Ávila-Quezada, G.D. Phylogenetic and morphological identification of Colletotrichum boninense: A novel causal agent of anthracnose in avocado. Plant Pathol. 2011, 60, 899–908. [Google Scholar] [CrossRef]
- Hunupolagama, D.M.; Wijesundera, R.L.C.; Chandrasekharan, N.V.; Wijesundera, W.S.S.; Kathriarachchi, H.S.; Fernando, T.H.P.S. Characterization of Colletotrichum isolates causing avocado anthracnose and first report of C. gigasporum infecting avocado in Sri Lanka. Plant Pathol. Quar. 2015, 5, 132–143. [Google Scholar] [CrossRef]
- Hernández-Lauzardo, A.N.; Campos-Martínez, A.; Velázquez-del Valle, M.G.; Flores-Moctezuma, H.E.; Suárez-Rodríguez, R.; Ramírez-Trujillo, J.A. First report of Colletotrichum godetiae causing anthracnose on avocado in Mexico. Plant Dis. 2015, 99, 555. [Google Scholar] [CrossRef]
- Akgul, D.S.; Awan, Q.N.; Guler, P.G.; Önelge, N. First report of anthracnose and stem end rot diseases caused by Colletotrichum gloeosporioides and Neofusicoccum australe on avocado fruits in Turkey. Plant Dis. 2016, 100, 1792. [Google Scholar] [CrossRef]
- Shivas, R.G.; Tan, Y.P.; Edwards, J.; Dinh, Q.; Maxwell, A.; Andjic, V.; Liberato, J.R.; Anderson, C.; Beasley, D.R.; Bransgrove, K.; et al. Colletotrichum species in Australia. Australas. Plant Pathol. 2016, 45, 447–464. [Google Scholar] [CrossRef]
- Honger, J.O.; Offei, S.K.; Oduro, K.A.; Odamtten, G.T.; Nyaku, S.T. Identification and molecular characterisation of Colletotrichum species from avocado, citrus and pawpaw in Ghana. S. Afr. J. Plant Soil 2016, 33, 1–9. [Google Scholar] [CrossRef]
- Sharma, G.; Maymon, M.; Freeman, S. Epidemiology, pathology and identification of Colletotrichum including a novel species associated with avocado (Persea americana) anthracnose in Israel. Sci. Rep. 2017, 7, 15839. [Google Scholar] [CrossRef]
- Fuentes-Aragón, D.; Juárez-Vázquez, S.B.; Vargas-Hernández, M.; Silva-Rojas, H.V. Colletotrichum fructicola, a member of Colletotrichum gloeosporioides sensu lato, is the causal agent of anthracnose and soft rot in avocado ruits cv. “Hass”. Mycobiology 2018, 46, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Giblin, F.R.; Tan, Y.P.; Mitchell, R.; Coates, L.M.; Irwin, J.A.G.; Shivas, R.G. Colletotrichum species associated with pre-and post-harvest diseases of avocado and mango in eastern Australia. Australas. Plant Pathol. 2018, 47, 269–276. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Choi, O.; Lee, Y.; Kim, S.; Kang, B.; Kim, J. Anthracnose on postharvest avocado caused by Colletotrichum kahawae subsp. ciggaro in South Korea. Can. J. Plant Pathol. 2020, 42, 508–513. [Google Scholar] [CrossRef]
- Ayvar-Serna, S.; Díaz-Nájera, J.F.; Mateo, V.H.; Camacho-Tapia, M.; Valencia-Rojas, G.A.; Lima, N.B.; Tovar-Pedraza, J.M. First report of Colletotrichum jiangxiense causing avocado anthracnose in Mexico. Plant Dis. 2021, 105, 502. [Google Scholar] [CrossRef]
- Fuentes-Aragón, D.; Silva-Rojas, H.V.; Guarnaccia, V.; Mora-Aguilera, J.A.; Aranda-Ocampo, S.; Bautista-Martínez, N.; Téliz-Ortíz, D. Colletotrichum species causing anthracnose on avocado fruit in Mexico: Current status. Plant Pathol. 2020, 69, 1513–1528. [Google Scholar] [CrossRef]
- Soares, M.G.O.; Alves, E.; Silveira, A.L.; Pereira, F.D.; Guimarães, S.S.C. Colletotrichum siamense is the main aetiological agent of anthracnose of avocado in south-eastern Brazil. Plant Pathol. 2021, 70, 154–166. [Google Scholar] [CrossRef]
- Bananalink. All about Banana. 2020. Available online: http://www.bananalink.org.uk/all-about-bananas (accessed on 21 May 2020).
- Agriexchange. Banana Report. 2020. Available online: https://agriexchange.apeda.gov.in/Weekly_eReport/Banana_Report.pdf (accessed on 21 May 2020).
- Food and Agriculture Organization of the United Nations (FAO). Banana Market Review February 2020 Snapshot; FAO: Rome, Italy, 2020. [Google Scholar]
- Jeger, M.J.; Eden-Green, S.; Thresh, J.M.; Johannson, A.; Waller, J.M.; Brown, A.E. Banana diseases. In Banana and Plantains; Gowen, S.R., Ed.; Chapman and Hall: London, UK, 1995; pp. 3117–3381. [Google Scholar]
- Meredith, D.S. Studies on Gloeosporium musarum (Gke and massee) causing storage rots of Jamaican bananas. Anthracnose and its chemical control. Ann. Appl. Biol. 1960, 48, 279–290. [Google Scholar] [CrossRef]
- Chillet, M.O.; Hubert, L.; de Bellaire, L.D.L. Relationship between physiological age, ripening and susceptibility of banana to wound anthracnose. Crop. Prot. 2007, 26, 1078–1082. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Roshdy, S.; Amin, O.E.; Rabani, M. First morphogenetic identication of the fungal pathogen Colletotrichum musae (Phyllachoraceae) from imported bananas in Saudi Arabia. Genet. Mol. Res. 2010, 9, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.Y.; Noireung, P.; Liu, F.; Hyde, K.D.; Moslen, M.A.; Bahkali, A.H.; Abd-Elsalam, K.A.; Cai, L. Epitypification of Colletotrichum musae, the causative agent of banana anthracnose. Mycoscience 2011, 52, 376–382. [Google Scholar] [CrossRef]
- Udayanga, D.; Manamgoda, D.S.; Liu, X.Z.; Chukeatirote, E.; Hyde, K.D. What are the common anthracnose pathogens of tropical fruits? Fungal Divers. 2013, 61, 165–179. [Google Scholar] [CrossRef]
- Balendres, M.A.; Mendoza, J.; Dela Cueva, F. Characteristics of Colletotrichum musae PHBN0002 and the susceptibility of popular banana cultivars to postharvest anthracnose. Indian Phytopathol. 2020, 73, 57–64. [Google Scholar] [CrossRef]
- Sangeetha, G.; Usharani, S.; Muthukumar, A. Significance of Lasiodiplodia theobromae and Colletotrichum musae in causing crown rot in banana and their reaction on some commercial banana cultivars. Indian J. Hort. 2010, 67, 21–25. [Google Scholar]
- Patil, R.S.; Deshmukh, R.G.; Bhaskar, K.R.; Jahagirdar, S.W. Growth and export performance of mango in India. Int. J. Curr. Microbiol. Appl. Sci. 2018, 6, 2667–2673. [Google Scholar]
- Arauz, L.F. Mango anthracnose: Economic impact and current options for integrated management. Plant Dis. 2000, 84, 600–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdappa, P.; Mohan Kumar, S.P. Existence of two genetically distinct populations of Colletotrichum gloeosporioides Penz in mango from India. Pest. Manag. Hort. Ecosyst. 2012, 18, 161–170. [Google Scholar]
- Jayasinghe, C.K.; Fernando, T.H.P.S. First report of Colletotrichum acutatum on Mangifera indica in Sri Lanka. J. Sci. Bio. Sci. 2009, 38, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Kamle, M.; Kumar, P.; Gupta, V.K.; Tiwari, A.K.; Misra, A.K.; Pandey, B.K. Identification and phylogenetic correlation among Colletotrichum gloeosporioides pathogen of anthracnose for mango. Biocatal. Agric. Biotechnol. 2013, 2, 285–287. [Google Scholar] [CrossRef]
- Alahakoon, P.W.; Brown, A.E.; Sreenivasaprasad, S. Cross-infection potential of genetic groups of Colletotrichum gloeospoirides on tropical fruits. Physiol. Mol. Plant. Pathol. 1994, 44, 93–103. [Google Scholar] [CrossRef]
- Hayden, H.L.; Pegg, K.G.; Aitken, E.A.B.; Irwin, J.A.G. Genetic relationships as assessed bimolecular markers and cross- infection among strains of Colletotrichum gloeosporioides. Aust. J. Bot. 1994, 42, 9–18. [Google Scholar] [CrossRef]
- Waller, J.M.; Bridge, P.D. Recent advances in understanding Colletotrichum diseases of some tropical perennial crops. In Colletotrichum: Host Specificity, Pathology, and Host-Pathogen Interaction; Prusky, D., Freeman, S., Dickman, M.B., Eds.; APS Press: St Paul, MN, USA, 2000; pp. 337–345. [Google Scholar]
- Ploetz, R. Anthracnose: The most important disease in much of the mango-producing world. In PLP News, The Newsletter of the Plant Pathology Department; The University of Florida: Gainseville, FL, USA, 1999; Volume 3, pp. 1–2. [Google Scholar]
- Lima, W.G.; Sposito, M.B.; Amorim, L.; Golcalves, F.P.; de Filho, P.A.M. Comparative epidemiology of Colletotrichum species from mango in northeastern Brazil. Eur. J. Plant Pathol. 2015, 141, 679–688. [Google Scholar] [CrossRef]
- Sharma, G.; Gryzenhout, M.; Hyde, K.D.; Pinnaka, A.K.; Shenoy, B.D. First report of Colletotrichum asianum causing mango anthracnose in South Africa. Plant Dis. 2015, 99, 725. [Google Scholar] [CrossRef]
- Benatar, G.V.; Wibowo, A.; Suryant. First report of Colletotrichum asianum associated with mango fruit anthracnose in Indonesia. Crop. Prot. 2021, 141, 105432. [Google Scholar] [CrossRef]
- FAOSTAT. 2017. Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Tropical_Fruits/Documents/CA2895EN (accessed on 15 April 2020).
- Dickman, K.B.; Part, V. Papaya: Anthracnose. In Compendium of Tropical Fruit Diseases; Ploetz, R.C., Zentmeyer, G.A., Nishijima, W.T., Rohrbach, K.G., Ohr, H.D., Eds.; APS Press: St Paul, MN, USA, 1994; pp. 58–59. [Google Scholar]
- Vieria dos Santos, W.A.; dos Santos Nunes, A.; Veloso, J.S.; Machado, A.R.; Queiroz Balbino, V.; da Silva, A.C.; Medeiros Gomes, A.Â.; Doyle, V.P.; Câmara, M.P.S. Colletotrichum truncatum causing anthracnose on papaya fruit (Carica papaya) in Brazil. Australas. Plant Dis. Notes 2020, 15. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Brito, D.; Cortes-Velázquez, A.; Valencia-Yah, T.; Magaña-Álvarez, A.; Navarro, C. Genetic variation of Colletotrichum magnum isolated from Carica papaya as revealed by DNA fingerprinting. J. Microbiol. 2018, 6, 813–821. [Google Scholar] [CrossRef]
- Sekeli, R.; Hamid, M.H.; Razak, R.A.; Wee, C.Y.; Ong-Abdullah, J. Malaysian Carica papaya L. var. Eksotika: Current research strategies fronting challenges. Front. Plant Sci. 2018, 9, 1380. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.J.; Paull, R.E. Overall Dragon Fruit Production and Global Marketing. FFTC Agricultural Policy Platform (FFTC-AP). 2018. Available online: https://ap.fftc.org.tw/article/1596 (accessed on 15 April 2020).
- Yacob, A. Dragonfruit: The Next Potential Major Tropical Fruit? 2019. Available online: https://www.itfnet.org/v1/2019/10/dragonfruit-the-next-potential-major-tropical-fruit/ (accessed on 15 April 2020).
- Hoa, T.T.; Clark, C.J.; Waddell, B.C.; Woolf, A.B. Postharvest quality of dragon fruit (Hylocereus undatus) following disinfesting hot air treatments. Postharvest Biol. Technol. 2006, 41, 62–69. [Google Scholar] [CrossRef]
- Gunasena, H.P.M.; Pushpakumara, D.K.N.G.; Kariyawasam, M. Dragon fruit (Hylocerus undatus [Haw.] Britton and Rose). In Underutilized Fruit Trees in Sri Lanka; Pushpakumara, D.K.N.G., Gunasena, H.P.M., Singh, V.P., Eds.; South Asia Regional Office; World Agroforestry Centre (International Council for Research in Agroforestry–ICRAF): New Delhi, India, 2007; Volume 1, pp. 110–141. [Google Scholar]
- Leite, K.M.S.C.; Tadiotti, A.C.; Baldochi, D.; Oliveira, O.M.M.F. Partial purification, heat stability and kinetic characterization of the pectinmethylesterase from Brazilian guava, Paluma cultivars. Food Chem. 2006, 94, 565–572. [Google Scholar] [CrossRef]
- Naseer, S.; Naureen, N.; Hussain, S.; Pervaiz, M.; Rahman, M. The phytochemistry and medicinal value of Psidium guajava (guava). Clin. Phytosci. 2018, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Morton, J. Guava (Psidium guajava L.). In Fruits of Warm Climates; Morton: Miami, FL, USA, 1987; pp. 356–363. Available online: http://www.hort.purdue.edu/newcrop/morton/guava.html# (accessed on 20 April 2020).
- Pariona, A. Top Guava Producing Countries in the World. 2017. Available online: https://www.worldatlas.com/articles/top-guava-producing-countries-in-the-world.html (accessed on 20 May 2020).
- Das, M.; Bora, K.N. Ultrastructural studies on infection processes by Colletotrichum acutatum on guava fruit. Indian Phytopathol. 1998, 51, 353–356. [Google Scholar]
- Peres, N.A.; Timmer, L.W.; Adaskaveg, J.; Correll, J.C. Life cycles of Colletotrichum acutatum. Plant Dis. 2005, 89, 784–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, A.R.; Lourenco, S.; Amorim, L. Infection of guava by C. gleosporioides and C. acutatum under different temperature and wetting periods. Trop. Plant Pathol. 2008, 33, 265–272. [Google Scholar]
- Workman, D. Avocados Export by Country. 2020. Available online: http://www.worldstopexports.com/avocados-exports-by-country/ (accessed on 1 July 2020).
- Ngbolua, K.; Ngiala, G.B.; Liyongo, C.I.; Ashande, C.M.; Lufuluabo, G.L.; Mukiza, J.; Mpiana, P.T. A mini review on the phytochemistry and pharmacology of the medicinal plant species Persea americana Mill. (Lauraceae). Discov. Phytomed. 2019, 6, 102–111. [Google Scholar] [CrossRef]
- Velázquez-del Valle, M.G.; Campos-Martínez, A.; Flores-Moctezuma, H.E.; Suárez-Rodríguez, R.; Ramírez-Trujillo, J.A.; Hernández-Lauzardo, A.N. First report of avocado anthracnose caused by Colletotrichum karstii in Mexico. Plant Dis. 2016, 100, 534. [Google Scholar] [CrossRef]
- Eckert, J.W. Recent developments in the chemical control of postharvest diseases. Acta Hortic. 1990, 269, 477–494. [Google Scholar] [CrossRef]
- Coates, L.M.; Johnson, G.I.; Dale, M. Postharvest pathology of fruit and vegetables. In Plant Pathogens and Plant Diseases; Brown, J., Ogle, H., Eds.; Rockvale Publications: Burlington, MA, USA, 1997; pp. 533–547. [Google Scholar]
- Northover, J.; Zhou, T. Control of rhizopus rot of peaches with postharvest treatments of tebuconazole, fludioxonil, and Pseudomonas syringae. Can. J. Plant Pathol. 2002, 24, 144–153. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control. 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Wisniewski, M.; Wilson, C.; Droby, S.; Chalutz, E.; El Ghaouth, A.; Stevens, C. Postharvest biocontrol: New concepts and applications. In Biological Control: A Global Perspective; Vincent., C., Goettal, M.S., Lazarovits, G., Eds.; CABI: Cambridge, MA, USA, 2007; pp. 262–273. [Google Scholar]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef]
- Asgar, A.; Noosheen, Z.; Sivakumar, M.; Yasmeen, S.; Alderson, P.G.; Mehdi, M. Effectiveness of submicron chitosan dispersions in controlling anthracnose and maintaining quality of dragon fruit. Postharvest Biol. Technol. 2013, 86, 147–153. [Google Scholar]
- De Oliveira, K.A.R.; Berger, L.R.R.; de Araújo, S.A.; Câmara, M.P.S.; de Souza, E.L. Synergistic mixtures of chitosan and Mentha piperita L. essential oil to inhibit Colletotrichum species and anthracnose development in mango cultivar Tommy Atkins. Food Microbiol. 2017, 66, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Braga, S.; Lundgren, G.; Samara, M.; Tavares Josean, T.; Willie Anderson, V.; Câmara, M.; Evandro, S. Application of coatings formed by chitosan and Mentha essential oils to control anthracnose caused by Colletotrichum gloesporioides and C. brevisporum in papaya (Carica papaya L.) fruit. Int. J. Biol. Macromol. 2019, 139, 631–639. [Google Scholar] [CrossRef]
- Palou, L. Postharvest treatments with GRAS salts to control fresh fruit decay. Horticulturae 2018, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- De Costa, D.M.; Gunawardhana, H.M.D.M. Effects of sodium bicarbonate on pathogenicity of Colletotrichum musae and potential for controlling postharvest diseases of banana. Postharvest Biol. Technol. 2012, 68, 54–63. [Google Scholar] [CrossRef]
- Jitareerat, P.; Sripong, K.; Masaya, K.; Aiamlaor, S.; Uthairatanakij, A. Combined effects of food additives and heat treatment on fruit rot disease and quality of harvested dragon fruit. Agric. Nat. Resour. 2018, 5, 543–549. [Google Scholar] [CrossRef]
- Kalupahana, K.I.M.; Kuruppu, M.; Dissanayake, P.K. Effect of essential oils and GRAS compounds on postharvest disease control in mango (Mangifera indica L.cv Tom EJC). J. Agric. Sci. Sri Lanka 2020, 15, 207–221. [Google Scholar] [CrossRef]
- De la Rosa-García, S.; Martínez-Torres, P.; Gomez-Cornelio, S.; Corral-Aguado, M.; Quintana, P.; Gómez-Ortíz, N. Antifungal activity of ZnO and MgO nanomaterials and their mixtures against Colletotrichum gloeosporioides strains from tropical fruit. J. Nanomater. 2018, 3498527. [Google Scholar] [CrossRef] [Green Version]
- Jagana, D.; Hegde, Y.R.; Lella, R. Green nanoparticles: A novel approach for the management of banana anthracnose caused by Colletotrichum musae. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1749–1756. [Google Scholar] [CrossRef]
- Chowdappa, P.; Gowda, S.; Chethana, C.S.; Madhura, S. Antifungal activity of chitosan-silver nanoparticle composite against Colletotrichum gloeosporoides associated with mango anthracnose. Afr. J. Microbiol. Res. 2014, 8, 1803–1812. [Google Scholar]
- Raghavendra, S.N.; Raghu, H.S.; Divyashree, K.; Rajeshwara, A.N. Antifungal efficiency of copper oxychloride-conjugated silver nanoparticles against Colletotrichum gloeosporioides which causes anthracnose disease. Asian J. Pharm. Clin. Res. 2019, 12, 230–233. [Google Scholar] [CrossRef]
- Turner, M. Physics of cold plasma. In Cold Plasma in Food and Agriculture; Misra, N.N., Schluter, O., Cullen, P.J., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 17–51. [Google Scholar]
- Misra, N.N.; Barun, Y.; Roopesh, M.S.; Cheorun, J. Cold plasma for effective fungal and mycotoxin control in Foods: Mechanisms, inactivation effects and applications. Compr. Rev. Food Sci. Saf. 2019, 18, 106–120. [Google Scholar] [CrossRef] [Green Version]
- Siddique, S.S.; Hardy, G.J.; Bayliss, K.L. Advanced technologies for controlling postharvest diseases of fruit. Acta Hortic. 2018, 1194, 193–200. [Google Scholar] [CrossRef]
- Misra, N.N.; Moiseev, T.; Patil, S.; Pankaj, S.K.; Bourke, P.; Mosnier, J.P.; Keener, K.M.; Cullen, P.J. Cold plasma in modified atmospheres for post-harvest treatment of strawberries. Food Bioprocess Tech. 2014, 7, 3045–3054. [Google Scholar] [CrossRef]
- Talhinhas, P.; Gonçalves, E.; Sreenivasaprasad, S.; Oliveira, H. Virulence diversity of anthracnose pathogens (Colletotrichum acutatum and C. gloeosporioides Species Complexes) on eight olive cultivars commonly grown in Portugal. Eur. J. Plant Pathol. 2014, 142, 73–83. [Google Scholar] [CrossRef]
- Baroncelli, R.; Zapparata, A.; Sarrocco, S.; Sukno, S.A.; Lane, C.R.; Thon, M.R.; Vannacci, G.; Holub, E.; Sreenivasaprasad, S. Molecular diversity of anthracnose pathogen populations associated with UK strawberry production suggests multiple introductions of three different Colletotrichum species. PLoS ONE 2015, 10, e0129140. [Google Scholar] [CrossRef]
Figure 1.
Anthracnose disease cycle of tropical fruit crops. (Drawn by Latiffah Zakaria).

Figure 2.
Anthracnose symptoms on several tropical fruit crops. (A) Guava, (B) dragon fruits, (C) banana, (D) mango, (E) papaya. (Photographs taken by Latiffah Zakaria and postgraduate students).
Figure 2.
Anthracnose symptoms on several tropical fruit crops. (A) Guava, (B) dragon fruits, (C) banana, (D) mango, (E) papaya. (Photographs taken by Latiffah Zakaria and postgraduate students).


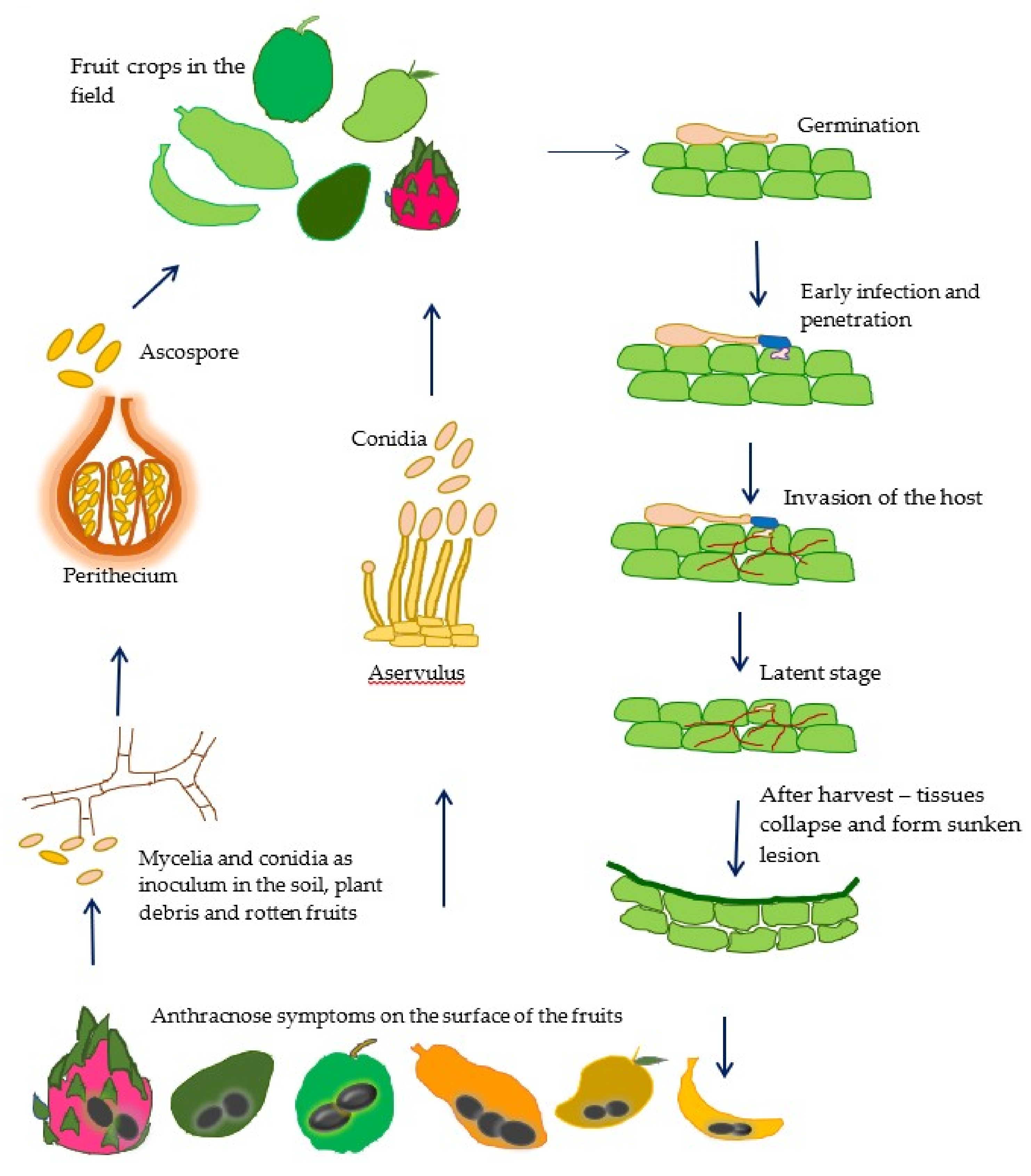{kind=link}
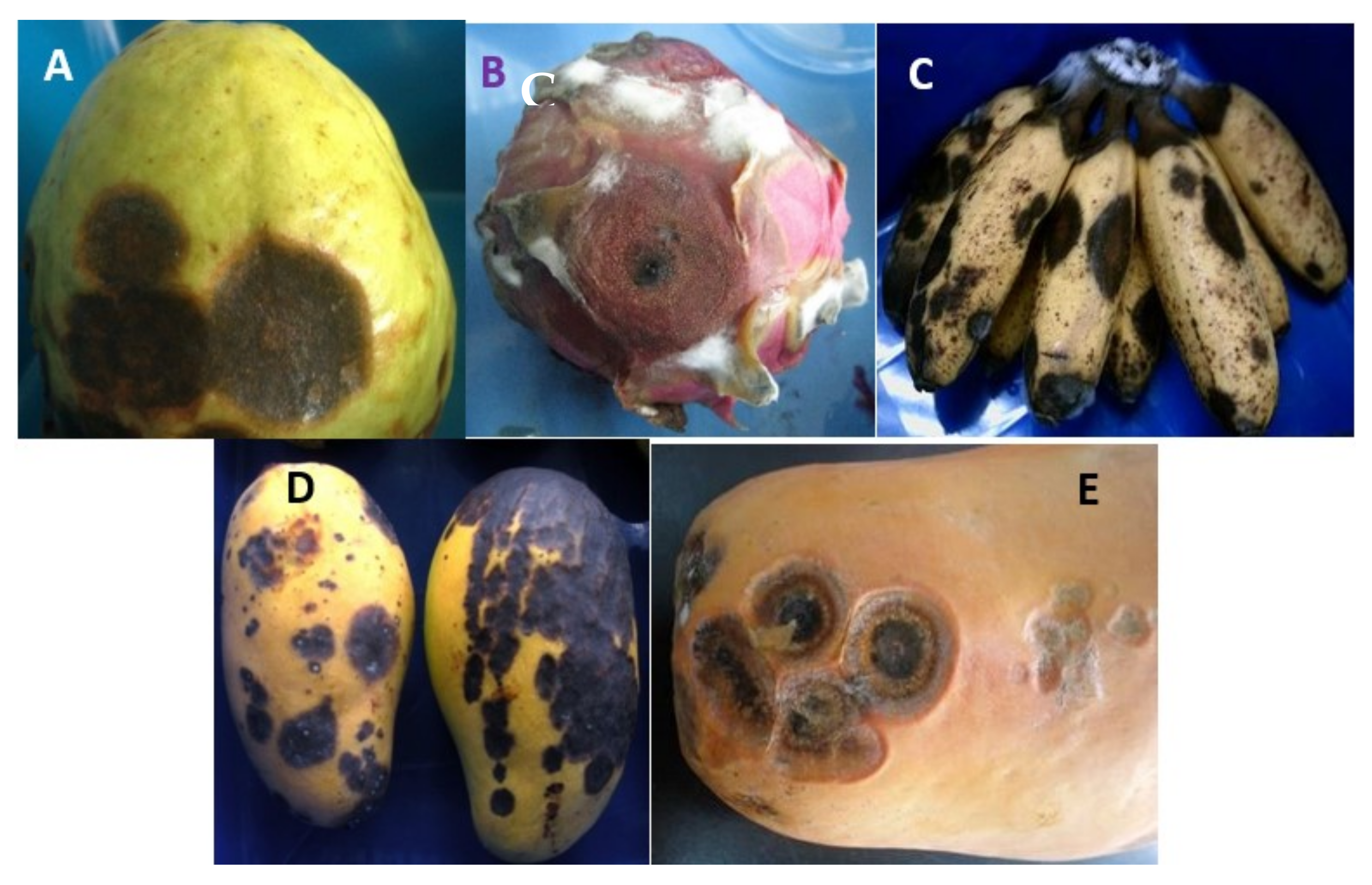{kind=link}
Table 1.
Species of Colletotrichum associated with tropical fruit crops reported in several countries.
Table 1.
Species of Colletotrichum associated with tropical fruit crops reported in several countries.
| Fruit Crop | Reported Colletotrichum spp. | Country | References |
|---|---|---|---|
| Banana (Musa spp.) | C. paxtonii | St Lucia | Damm et al. (2012a) [12] |
| C. gloeosporioides sensu lato | Malaysia | Intan Sakinah et al. (2013) [20] | |
| C. scovillei | Hainan Province, China | Zhou et al. (2016) [21] | |
| C. siamense | India | Kumar et al. (2017) [22] | |
| C. siamense, C. tropicale, C. chrysophilum, C. theobromicola | Brazil | Vieira et al. (2017) [23] | |
| C. gloeosporioides | Ecuador | Riera et al. (2019) [24] | |
| C. siamense | Turkey | Uysal and Kurt (2020) [25] | |
| C. chrysophilum | Mexico | Fuentes-Aragón et al. (2020) [26] | |
| Mango (Mangifera indica L.) | C. asianum and C. gloeosporioides | Panama | Rojas et al. (2010) [27] |
| C. asianum, C. fructicola, C. tropicale, C. karstii, C. dianesei | northeastern Brazil | Lima et al. (2013) [28] | |
| C. fragariae sensu stricto, C. fructicola, C. jasmine-sambac, C. melanocaulon | India | Sharma et al. (2013) [29] | |
| C. asianum, C. gloeosporioides | Sri Lanka. | Krishnapillai and Wijeratnam (2014) [30] | |
| C. asianum | Malaysia | Latiffah et al. (2015) [31] | |
| C. asianum, C. gloeosporioides | Colombia | Pardo-De la Hoz et al. (2016) [32] | |
| C. asianum | Sanya City, China | Qin et al. (2017) [33] | |
| C. asianum, C. fructicola, C. siamense | Guangxi, South China | Mo et al. (2018) [34] | |
| C. scovillei (mango leaves) | Guangxi, China | Qin et al. (2017) [33] | |
| C. asianum, C. cliviicola, C. cordylinicola, C. endophytica, C. fructicola, C. gigasporum, C. gloeosporioides, C. karstii, C. liaoningense, C. musae, C. scovillei, C. siamense, C. tropicale | southern China | Li et al. (2019) [35] | |
| C. asianum, C. fructicola, C. siamense, C. tropicale, C. scovillei | Taiwan | Wu et al. (2020) [36] | |
| C. alienum, C. asianum, C. fructicola, C. siamense, C. tropicale | Mexico | Tovar-Pedraza et al. (2020) [37] | |
| C. asianum | Philippine | Alvarez et al. (2020) [38] | |
| C. alienum | Beijing, China | Ahmad et al. (2021) [39] | |
| Papaya (Carica papaya L.) | C. gloeosporioides, C. capsici | Yucatan peninsula, Mexico | Tapia-Tussell et al. (2008) [40] |
| C. gloeosporioides, C. capsici | Malaysia | Rahman et al. (2008) [41] | |
| C. gloeosporioides, C. capsici | Miyako Islands, Okinawa, Japan | Yaguchi et al. (2008) [42] | |
| C. gloeosporioides, C. capsici | South Florida | Tarnowski and Ploetz (2010) [43] | |
| C. magna, C. gloeosporioides | Brazil | Nascimento et al. (2010) [44] | |
| C. gloeosporioides, C. capsici, C. dematium | Yucatan, Mexico | Santamaría Basulto et al. (2017) [45] | |
| C. acutatum, C. simmondsii | Australia | Damm et al. (2012a) [12] | |
| C. karstii | Brazil | Damm et al. (2012b) [46] | |
| C. queenslandicum | Australia | Weir et al. (2012) [13] | |
| C. siamense | South Africa | Weir et al. (2012) [13] | |
| C. gloeosporioides, C. capsici | Trinidad Island | Rampersad (2011) [47], Maharaj and Rampersad (2013) [48] | |
| C. karstii | India | Sharma and Shenoy (2013) [49] | |
| C. brevisporum | Brazil | Vieira et al. (2013) [50] | |
| C. fructicola, C. gloeosporioides | India | Saini et al. (2016) [51] | |
| C. magnum | Mexico | Tapia-Tussell et al. (2016) [52] | |
| C. salsolae | India | Saini et al. (2017) [53] | |
| C. truncatum, C. gloeosporioides sensu lato, C. magnum | Costa Rica | Molina-Chaves et al. (2017) [54] | |
| C. brevisporum | Taiwan | Duan et al. (2018) [55] | |
| C. truncatum | Korea | Aktaruzzaman et al. (2018) [56] | |
| C. plurivorum | Taiwan | Sun et al. (2019) [57] | |
| C. brevisporum | China | Liu et al. (2019) [58] | |
| C. truncatum | Brazil | Vieira et al. (2017) [23] | |
| C. okinawense | Brazil | Dias et al. (2020) [59] | |
| C. okinawense | Taiwan | Sun and Huang (2020) [60] | |
| C. siamense | China | Zhang et al. (2021) [61] | |
| Dragon fruits (Hylocereus spp.) | C. gloeosporioides sensu lato (H. undatus) | Okinawa Prefecture, Japan | Taba et al. (2006) [62] |
| C. gloeosporioides sensu lato (H. undatus) | Miami-Dade County, Florida, USA | Palmateer et al. (2007) [63] | |
| C. gloeosporioides sensu lato (H. megalanthus) | Brazil | Takahashi et al. (2008) [64] | |
| C. gloeosporioides sensu lato (H. polyrhizus, H. undatus, Selenicereus megalanthus) | Malaysia | Masyahit et al. (2009) [65] | |
| C. gloeosporioides (H. undatus young stems) | China | Ma et al. (2014) [66] | |
| C. truncatum (H. undatus fruits) | Yuanjiang County, Yunnan Province, China | Guo et al. (2014) [67] | |
| C. aenigma and C. siamense (H. undatus stem and fruit) | Thailand | Meetum et al. (2015) [68] | |
| C. truncatum (H. polyrhizus stem) | Malaysia | Suzianti et al. (2015) [69] | |
| C. gloeosporioides, C. truncatum, C. boninense (H. polyrhizus, H. undatus, H. costaricensis) | Taiwan | Lin et al. (2017) [70] | |
| C. siamense (H. polyrhizus stem) | China | Zhao et al. (2018) [71] | |
| C. siamense (H. undatus) | Andaman Islands, India | Abirami et al. (2019) [72] | |
| C. karstii (H. undatus stem) | Brazil | Nascimento et al. (2019) [73] | |
| Guava (Psidium guajava L.) | C. gloeosporioides sensu lato | Egypt | Omar (2001) [74] |
| C. gloeosporioides sensu lato | Ibadan, Nigeria | Amusa et al. (2005) [75] | |
| C. gloeosporioides | Hawaii | Keith and Zee (2010) [76] | |
| C. psidii | Italy | Weir et al. (2012) [13] | |
| C. guajavae | India | Damm et al. (2012a) [12] | |
| C. gloeosporioides sensu lato | Florida, USA | Merida and Palmateer (2013) [77] | |
| C. gloeosporioides sensu lato | Malaysia | Intan Sakinah et al. (2014) [78] | |
| C. simmondsii | Brazil | Cruz et al. (2015) [79] | |
| C. abscissum | USA | Crous et al. (2015) [80] | |
| C. siamense | India | Sharma et al. (2015b) [81] | |
| C. abscissum | Brazil | Bragança et al. (2016) [82] | |
| C. gloeosporioides sensu lato | China | Yao et al. (2018) [83] | |
| Avocado (Persea americana Mill.) | C. gloeosporioides sensu lato | USA | Nelson (2008) [84] |
| C. gloeosporioides, C. acutatum, C. boninense | Mexico | Silva-Rojas and Avila-Quezada (2011) [85] | |
| C. gigasporum | Sri Lanka | Hunupolagama et al. (2015) [86] | |
| C. godetiae | Mexico | Hernandez-Lauzardo et al. (2015) [87] | |
| C. gloeosporioides | Mersin Province, Turkey | Akgul et al. (2016) [88] | |
| C. alienum, C. fructicola, C. siamense | Australia | Shivas et al. (2016) [89] | |
| C. gloeosporioides, C. siamense | Ghana | Honger et al. (2016) [90] | |
| C. aenigma, C. alienum, C. fructicola, C. gloeosporioides sensu stricto, C. karstii, C. nupharicola, C. siamense, C. theobromicola, C. perseae | Israel | Sharma et al. (2017) [91] | |
| C. fructicola | Hidalgo, Mexico | Fuentes-Aragón et al. (2018) [92] | |
| C. alienum, C. asianum, C. fructicola, C. karstii, C. siamense | eastern Australia | Giblin et al. (2018) [93] | |
| C. kahawae subsp. cigarro | Jinju, South Korea | Kwon et al. (2020) [94] | |
| C. karstii | Turkey | Uysal and, Kurt (2020) [25] | |
| C. jiangxiense | Mexico | Ayvar-Serna et al. (2020) [95] | |
| C. karstii, C. godetiae, C. siamense, C. fioriniae, C. cigarro, C. chrysophilum, C. jiangxiense, C. tropicale, C. nymphaeae,Colletotrichum sp. 1, Colletotrichum sp. 2 | Mexico | Fuentes-Aragón et al. (2020) [96] | |
| C. siamense, C. kartsii | south eastern Brazil | Soares et al. (2021) [97] |
Table 2.
Diversity of Colletotrichum spp. and the infected plant parts.
| Fruit Crop | Colletotrichum spp. | Infected Parts |
|---|---|---|
| Banana (Musa spp.) | C. scovillei,C. gloeosporioides,C. siamense, C. tropicale, C. chrysophilum, C. paxtonii C. theobromicola | Fruit |
| C. musae | Ripe fruit, but has been reported from leaves and root of Musa spp. | |
| Mango (Mangifera indica L.) | C. fructicola, C. tropicale, C. fragariae sensu stricto, C. jasmine-sambac, C. melanocaulon, C. alienum | Fruit |
| C. asianum, C. karstii, C. scovillei, C. scovillei, C. fructicola, C. siamense, C. cliviicola, C. musae C. cordylinicola, C. endophytica, C. gigasporum, C. liaoningense, C. tropicale | Fruit and leaves | |
| C. gloeosporioides | Fruit, leaves, and inflorescence | |
| Papaya (Carica papaya L.) | C. magna, C. gloeosporioides, C. dematium,
C. acutatum, C. simmondsii, C. karstii, C. queenslandicum, C. siamense, C. salsolae, C. magnum, C. brevisporum, C. fructicola, C. plurivorum, C. okinawense, C. siamense | Fruit |
| Dragon fruits (Hylocereus spp.) | C. gloeosporioides sensu lato, C. truncatum, C. aenigma, C. siamense | Fruit and stem |
| C. gloeosporioides | Young stem | |
| C. boninense | Fruit | |
| C. karstii | Stem | |
| Guava (Psidium guajava L.) | C. gloeosporioides sensu lato, C. psidii, C. guajavae, C. simmondsii, C. abscissum, C. siamense | Fruit |
| C. gloeosporioides | Fruit and leaves | |
| Avocado (Persea americana Mill.) | C. gloeosporioides sensu lato, C. nymphaeae C. gloeosporioides, C. gigasporum, C. karstii, C. godetiae, C. alienum, C. fructicola, C. siamense, C. aenigma, C. alienum, C. perseae C. nupharicola, C. theobromicola, C. tropicale. C. kahawae subsp. cigarro, C. jiangxiense, C. cigarro, C. chrysophilum | Fruit |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zakaria, L. Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review. Agriculture 2021, 11, 297. https://doi.org/10.3390/agriculture11040297
AMA Style
Zakaria L. Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review. Agriculture. 2021; 11(4):297. https://doi.org/10.3390/agriculture11040297
Chicago/Turabian StyleZakaria, Latiffah. 2021. "Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review" Agriculture 11, no. 4: 297. https://doi.org/10.3390/agriculture11040297
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.