
A New Species of Diaphus Associated with Seamounts of the Emperor Chain, North-Western Pacific Ocean (Teleostei: Myctophiformes: Myctophidae) †

 and
and
Abstract
:1. Introduction
2. Materials and Methods
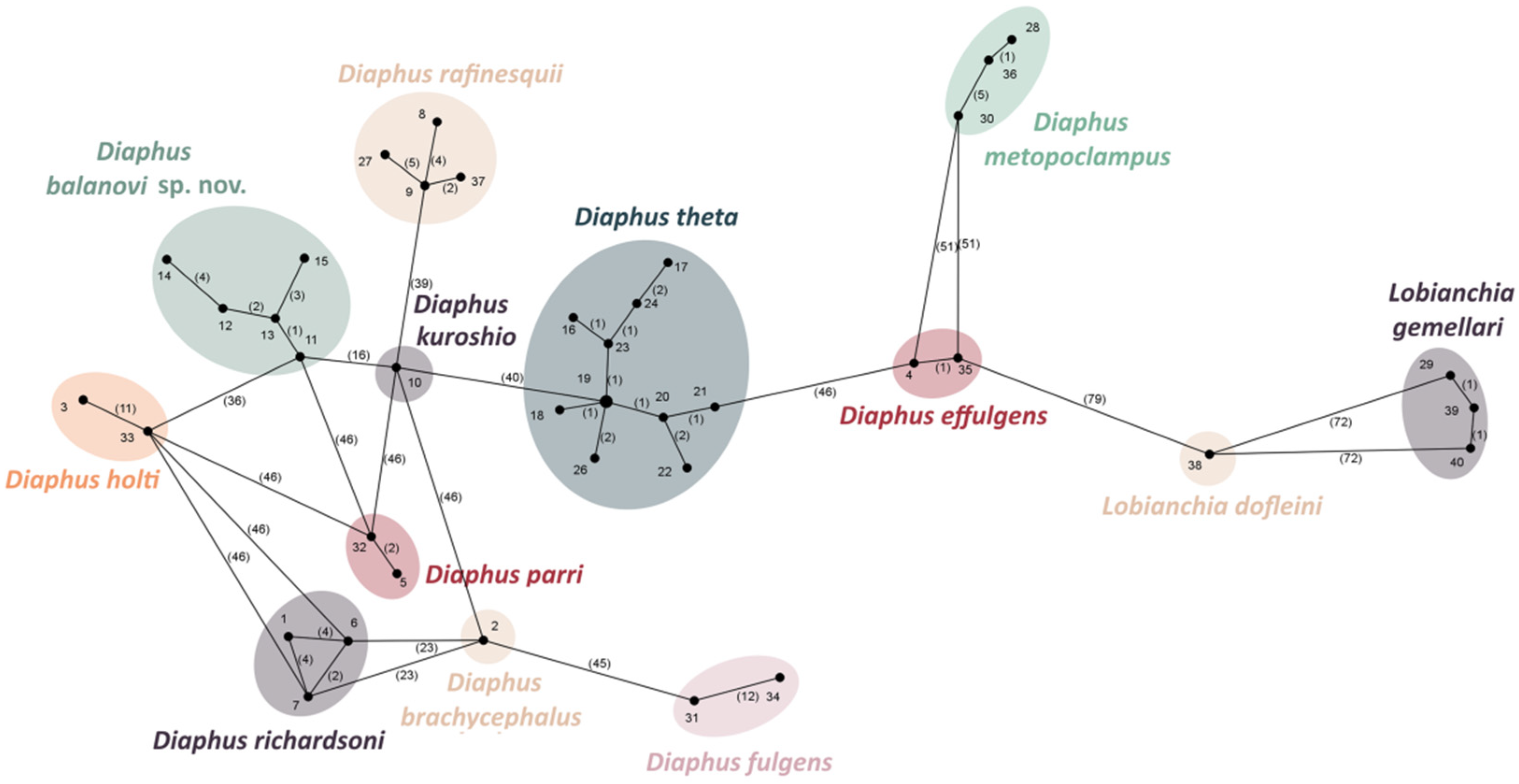
Molecular Data and Phylogenetic Analysis
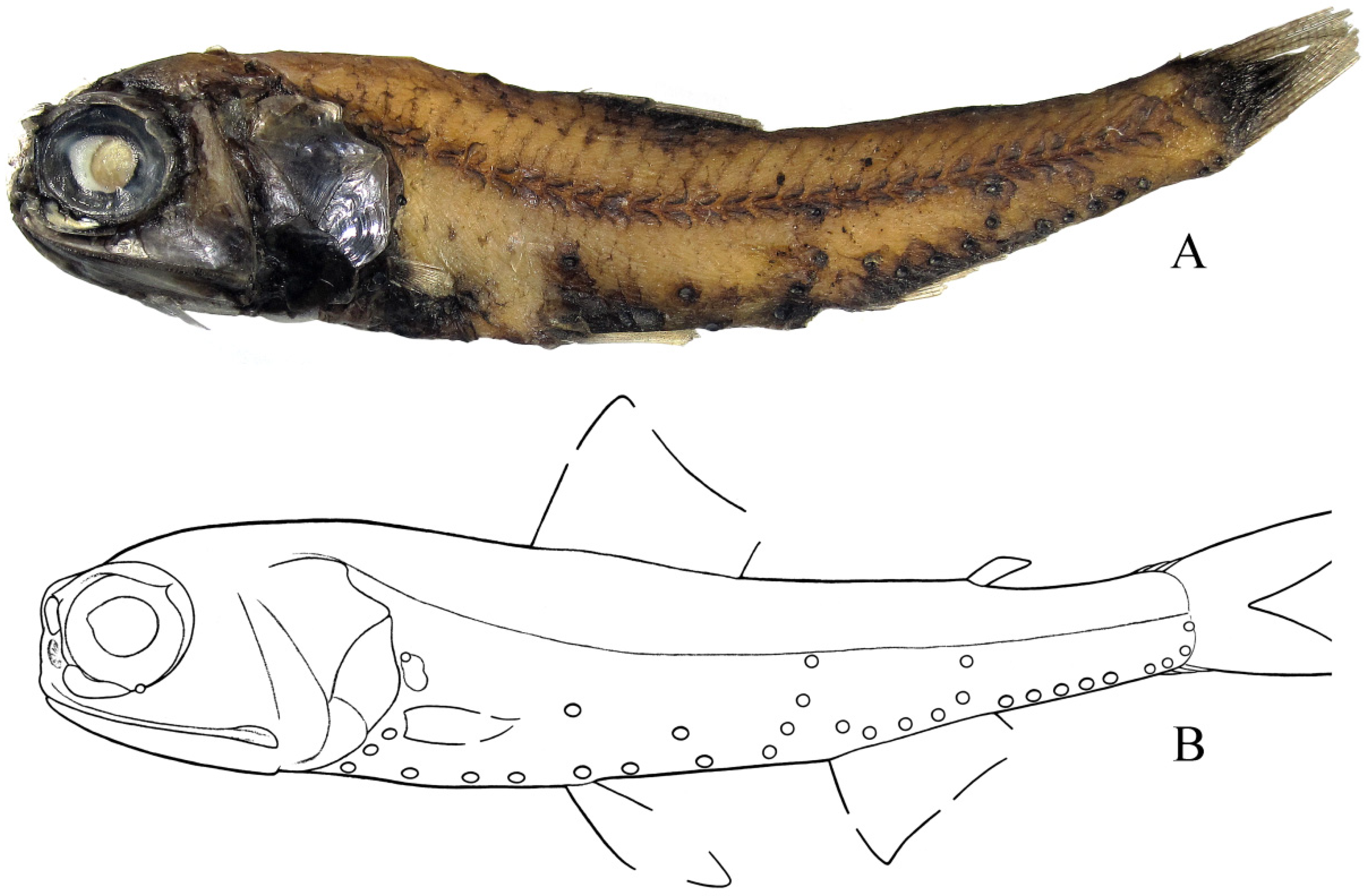
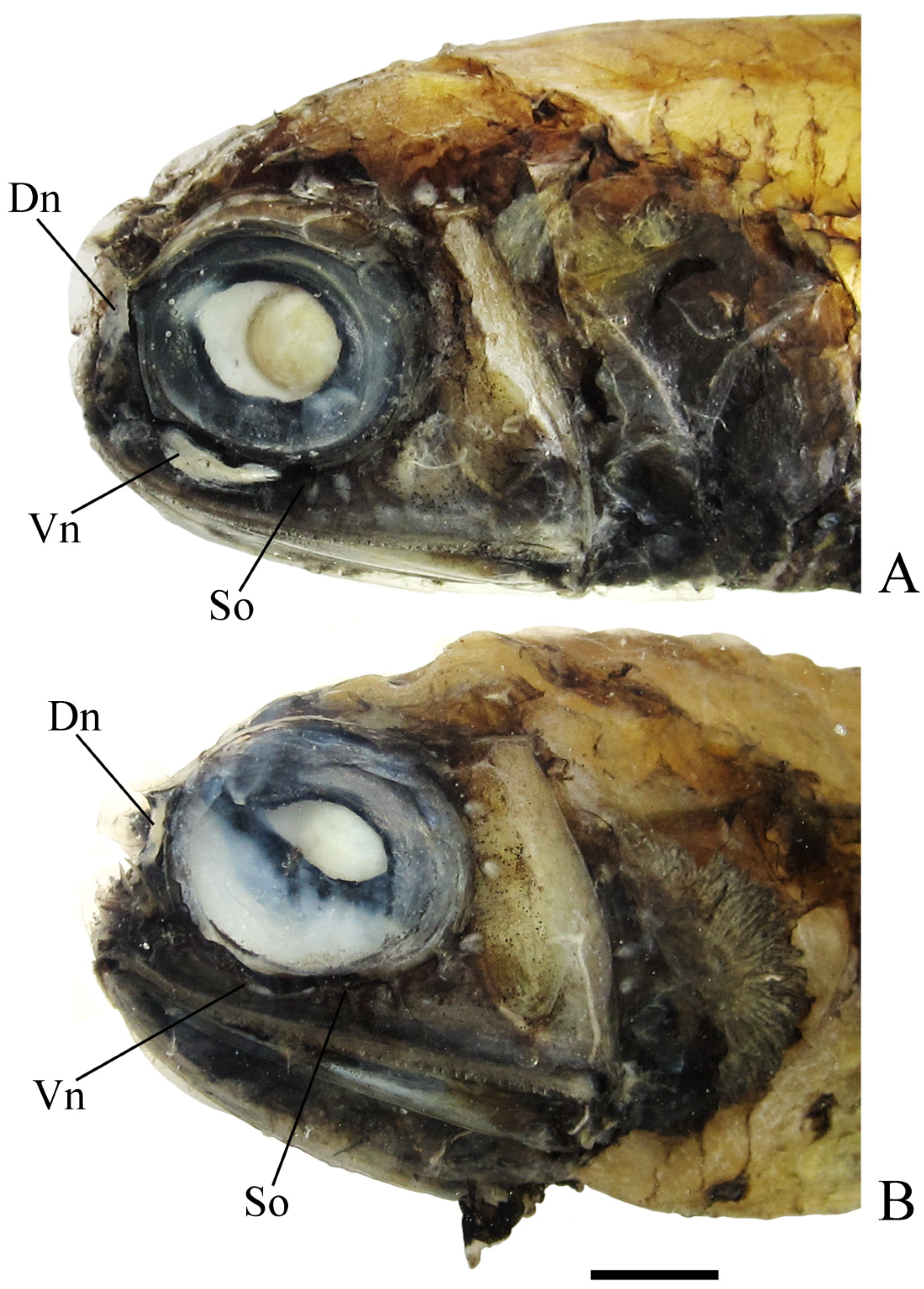
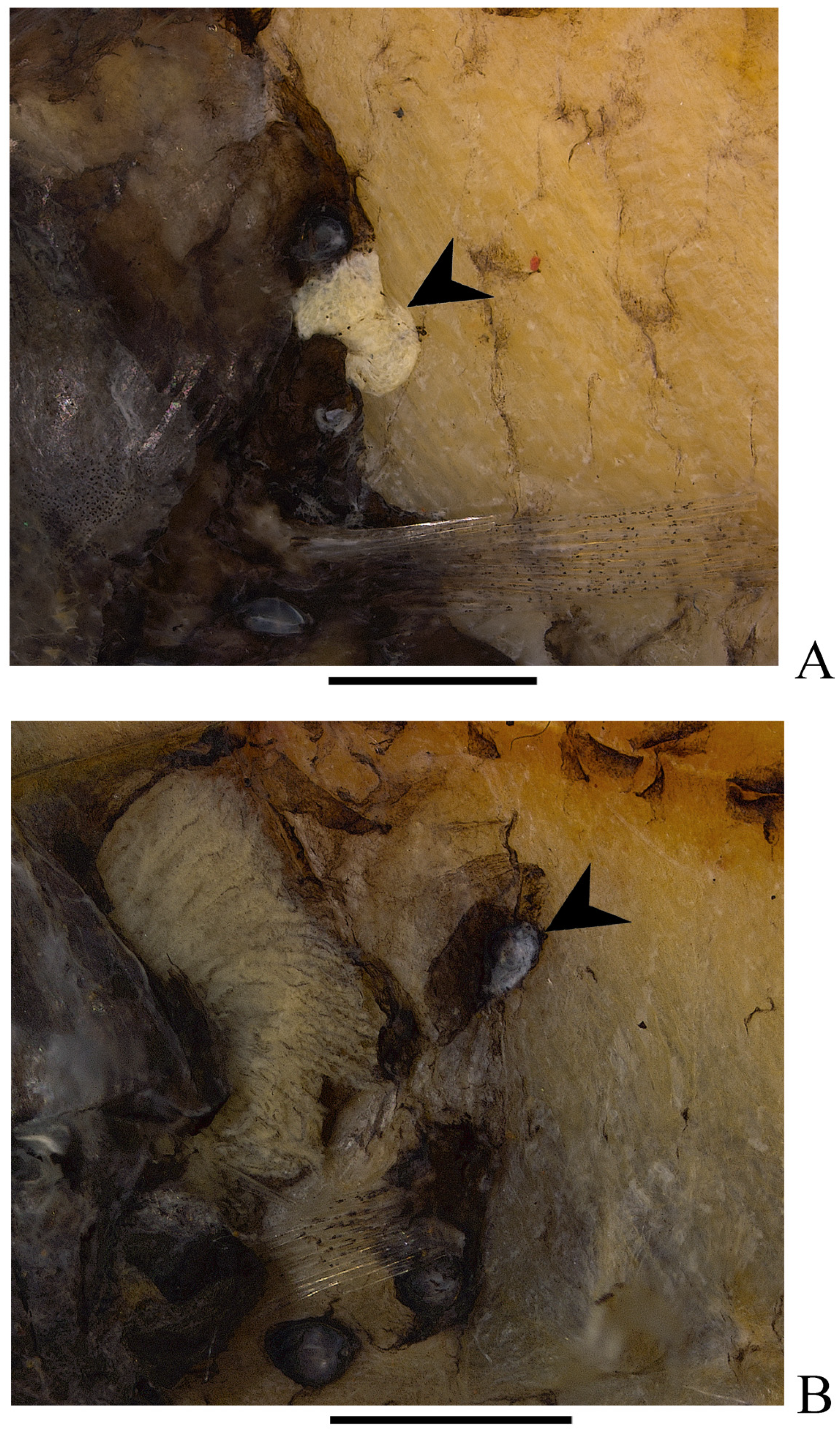
3. Results
Diaphus balanovi sp. nov.



4. Discussion

Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becker, V.E. Miktofovye Ryby Mirovogo Okeana [Myctophid Fishes of the World Ocean]; Nauka: Moscow, Russia, 1983; pp. 1–246. [Google Scholar]
- Nafpaktitis, B.G.; Robertson, D.A.; Paxton, J.R. Four new species of the lanternfish genus Diaphus (Myctophidae) from the Indo-Pacific. N. Z. J. Mar. Freshw. Res. 1995, 29, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Schwarzhans, W. A comparative morphological study of the recent otoliths of the genera Diaphus, Idiolychnus and Lobianchia (Myctophidae). Palaeoichthyologica 2013, 13, 41–82. [Google Scholar]
- Poulsen, J.Y.; Byrkjedal, I.; Willassen, E.; Rees, D.; Takeshima, H.; Satoh, T.P.; Shinohara, G.; Nishida, M.; Miya, M. Mitogenomic sequences and evidence from unique gene rearrangements corroborate evolutionary relationships of Myctophiformes (Neoteleostei). BMC Evol. Biol. 2013, 13, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, V.E.; Prut’ko, V.G. A new species of the genus Diaphus (Myctophidae) from northwestern part of the Indian Ocean. Vopr. Ikhtiol. 1984, 24, 883–887. [Google Scholar]
- Kornilova, G.N. Lanternfishes. In Biologicheskije Resursy Indijskogo Okeana [Biological Resources of the Indian Ocean]; Parin, N.V., Novikov, N.P., Eds.; Nauka: Moscow, Russia, 1989; pp. 247–251. [Google Scholar]
- Shust, K.V.; Orlov, A.M. Prospects for developing lanternfishes stock. Rybn. Khoz. 2003, 2, 38–41. [Google Scholar]
- Novikov, N.P.; Kodolov, L.S.; Gavrilov, G.M. Preliminary list of fishes of the Emperor underwater ridge. In Ryby Otkrytogo Okeana [Fishes of the Open Ocean]; Parin, N.V., Ed.; Institut Okeanologii AN SSSR: Moscow, Russia, 1981; pp. 32–35. [Google Scholar]
- Belyaev, V.A.; Darniskiy, V.B. Features of oceanography and ichthyofauna composition on the Emperor Ridge. In Deep Sea 2003: Conference of the Governance and Management of Deep-Sea Fisheries. Part 1: Conference Reports; Shotton, R., Ed.; FAO: Rome, Italy, 2005; pp. 107–124. [Google Scholar]
- Mundy, B.C. Checklist of the fishes of the Hawaiian Archipelago. Bishop Mus. Bull. Zool. 2005, 6, 1–704. [Google Scholar]
- Kawaguchi, K.; Shimizu, H. Taxonomy and distribution of the lanternfishes, genus Diaphus (Pisces, Myctophidae) in the western Pacific, eastern Indian oceans and the southeast Asian seas. Bull. Ocean Res. Inst. Univ. Tokyo 1978, 10, 1–145. [Google Scholar]
- Shinohara, G.; Yabe, M.; Nakaya, K.; Anma, G.; Yamaguchi, S.; Amaoka, K. Deep-sea fishes collected from the North Pacific by the T/S Oshoro-Maru. Mem. Fac. Fish. Hokkaido Univ. 1994, 45, 48–80. [Google Scholar]
- Sassa, C.; Moser, H.G.; Kawaguchi, K. Horizontal and vertical distribution patterns of larval myctophid fishes in the Kuroshio Current region. Fish. Oceanogr. 2002, 11, 1–10. [Google Scholar] [CrossRef]
- Sassa, C.; Kawaguchi, K.; Kinoshita, T.; Watanabe, C. Assemblages of vertical migratory mesopelagic fish in the transitional region of the western North Pacific. Fish. Oceanogr. 2002, 11, 193–204. [Google Scholar] [CrossRef]
- Sassa, C.; Kawaguchi, K.; Oozeki, Y.; Kubota, H.; Sugisaki, H. Distribution patterns of larval myctophid fishes in the transition region of the western North Pacific. Mar. Biol. 2004, 144, 417–428. [Google Scholar] [CrossRef]
- Kurnosova, A.S.; Somov, A.A.; Kanzeparova, A.N.; Zuev, M.A.; Orlova, S.Y.; Kurnosov, D.S.; Orlov, A.M. Mesopelagic micronekton and macroplankton and the conditions of its habitat at three stations in the Northeastern Pacific Ocean. Oceanology 2022, 62. in press. [Google Scholar]
- Somov, A.A.; Kanzeparova, A.N.; Vazhova, A.S.; Khleborodov, A.S.; Zuev, M.A.; Slabinsky, A.M.; Orlova, S.Y.; Kurnosov, D.S.; Belyaev, V.A.; Orlov, A.M. Some preliminary results of research on Emperor Seamounts in April, 2019. Tr. VNIRO 2019, 175, 208–219. [Google Scholar] [CrossRef]
- Hubbs, C.L.; Lagler, K.F. Fishes of the Great Lakes region. Bull. Cranbrook Inst. Sci. 1958, 26, 1–213. [Google Scholar]
- Nafpaktitis, B.G. Taxonomy and distribution of lanternfishes, genera Lobianchia and Diaphus, in the North Atlantic. Dana Rep. 1968, 73, 1–131. [Google Scholar]
- Nafpaktitis, B.G. Systematics and distribution of lanternfishes of the genera Lobianchia and Diaphus (Myctophidae) in the Indian Ocean. Sci. Bull. Nat. Hist. Mus. Los Angel. Co. 1978, 30, 1–92. [Google Scholar]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Phil. Trans. Roy. Soc. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Silva, J.W.; Costa, M.; Valente, V.; Sousa, J.D.F.; Paçó-Larson, M.; Espreafico, E.; Camargo, S.; Monteiro, E.; Holanda, A.D.J.; Zago, M.; et al. PCR template preparation for capillary DNA sequencing. Biotechniques 2001, 30, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; Heled, J.; Kearse, M.; Markowitz, S.; et al. Geneious v5.4. 2011. Available online: http://www.geneious.com (accessed on 24 September 2021).
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Villesen, P. FaBox: An online toolbox for fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denton, J.S. Seven-locus molecular phylogeny of Myctophiformes (Teleostei; Scopelomorpha) highlights the utility of the order for studies of deep-sea evolution. Mol. Phylogen. Evol. 2014, 76, 270–292. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Parin, N.V.; Sokolovskiy, A.S. Vidovoy sostav semeistva Melanostomiatidae (Osteichthyes) v zone techenija Kurosio [Species composition of the family Melanostomiatidae (Osteichthyes) in the Kuroshio Current Zone]. Tr. Inst. Okeanol. AN SSSR 1976, 104, 237–249. [Google Scholar]
- Nafpaktitis, B.G.; Backus, R.H.; Craddock, J.E.; Haedrich, R.L.; Robinson, B.H.; Karnella, C. Family Myctophidae. In Fishes of the Western North Atlantic; Gibbs, R.H., Ed.; Sears Foundation for Marine Research: New Haven, MD, USA, 1977; pp. 13–265. [Google Scholar]
- Kawaguchi, K.; Nafpaktitis, B.G. A new lanternfish, Diaphus kuroshio (family Myctophidae), from the Kuroshio waters off Japan. Jpn. J. Ichth. 1978, 25, 89–91. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Sample | No. Haplotype | Genbank Accession Number | Sampling Date | Locality | Country | Source |
|---|---|---|---|---|---|---|
| Lobianchia dofleini (outgroup 1) | ||||||
| 1 | 38 | SCAFB1054-09 | 8 September 2007 | Nova Scotia | Canada | BOLD Systems |
| 2 | 38 | MAECO457-09 | 28 June 2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| Lobianchia gemellari (outgroup 2) | ||||||
| 3 | 29 | FCFMT219-09 | 15 July 2007 | Mediterranean Sea | Malta | BOLD Systems |
| 4 | 40 | SCAFB1032-09 | 7 September 2007 | Nova Scotia | Canada | BOLD Systems |
| 5 | 39 | MAECO460-09 | 28 June 2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| Diaphus metopoclampus (sister group 1) | ||||||
| 6 | 28 | FCFMT123-09 | 16 June 2007 | Mediterranean Sea | Malta | BOLD Systems |
| 7 | 30 | FOAK120-10 | 15 June 2008 | Tasman Sea | Australia | BOLD Systems |
| 8 | 36 | MAECO111-06 | 30 June2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| Diaphus effulgens (sister group 2) | ||||||
| 9 | 4 | GBGCA11052-15 | 24 May 2000 | New England | USA | BOLD Systems |
| 10 | 4 | UKFBJ499-08 | 24 May 2000 | New England | USA | BOLD Systems |
| 11 | 35 | MAECO105-06 | 28 June 2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| Diaphus holti (sister group 3) | ||||||
| 12 | 33 | GBMIN125128-17 | July 2010 | Balearic Islands | Spain | BOLD Systems |
| 13 | 3 | GBGCA5261-13 | July 2010 | Balearic Islands | Spain | BOLD Systems |
| Diaphus brachycephalus (sister group 4) | ||||||
| 14 | 2 | MFLE267-13 | 26 January 2007 | Caribbean Sea | Belize | BOLD Systems |
| 15 | 2 | FOAO932-15 | 16 September 2014 | Western Indian Ocean | na | BOLD Systems |
| Diaphus fulgens (sister group 5) | ||||||
| 16 | 31 | GBGCA11055-15 | 2 December 2006 | Eastern tropical Pacific | na | BOLD Systems |
| 17 | 34 | LIDMA1136-12 | 10 March 1985 | Eastern tropical Pacific | na | BOLD Systems |
| Diaphus parri (sister group 6) | ||||||
| 18 | 5 | GBGCA11063-15 | na | na | na | BOLD Systems |
| 19 | 32 | GBGCA11065-15 | na | na | na | BOLD Systems |
| 20 | 5 | GBGCA11064-15 | na | na | na | BOLD Systems |
| Diaphus rafinesquii (sister group 7) | ||||||
| 21 | 37 | MAECO115-06 | 30 June 2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| 22 | 8 | MAECO116-06 | 28 June 2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| 23 | 9 | MAECO117-06 | 28 June 2004 | Mid-Atlantic Ridge | na | BOLD Systems |
| Diaphus richardsoni (sister group 8) | ||||||
| 24 | 6 | GBGCA11068-15 | na | na | na | BOLD Systems |
| 25 | 7 | LFSCS086-18 | 25 April 2014 | South China Sea | China | BOLD Systems |
| 26 | 1 | ANGBF22462-19 | 18 December 2010 | Southwestern Pacific | Australia | BOLD Systems |
| Diaphus kuroshio (sister group 9) | ||||||
| 27 | 10 | OL894503 | 12 April 2019 | Emperor Seamounts | na | NCBI |
| Diaphus theta (sister group 10) | ||||||
| 28 | 24 | DSFIB184-12 | 15 September 2011 | Strait of Georgia | Canada | BOLD Systems |
| 29 | 24 | FMV566-11 | 23 May 2006 | Sailish Sea | USA | BOLD Systems |
| 30 | 19 | TZFPB409-05 | 22 October 2005 | SW Vancouver Isl. | Canada | BOLD Systems |
| 31 | 16 | OL894509 | 28 March 2019 | NE Pacific | na | NCBI |
| 32 | 17 | OL894510 | 28 March 2019 | NE Pacific | na | NCBI |
| 33 | 18 | OL894511 | 28 March 2019 | NE Pacific | na | NCBI |
| 34 | 19 | OL894512 | 28 March 2019 | NE Pacific | na | NCBI |
| 35 | 19 | OL894513 | 28 March 2019 | NE Pacific | na | NCBI |
| 36 | 20 | OL894514 | 28 March 2019 | NE Pacific | na | NCBI |
| 37 | 21 | OL894515 | 31 March 2019 | NE Pacific | na | NCBI |
| 38 | 22 | OL894516 | 31 March 2019 | NE Pacific | na | NCBI |
| 39 | 23 | OL894517 | 31 March 2019 | NE Pacific | na | NCBI |
| 40 | 20 | OL894518 | 31 March 2019 | NE Pacific | na | NCBI |
| 41 | 24 | OL894519 | 31 March 2019 | NE Pacific | na | NCBI |
| 42 | 19 | OL894520 | 31 March 2019 | NE Pacific | na | NCBI |
| 43 | 25 | OL894521 | 31 March 2019 | NE Pacific | na | NCBI |
| 44 | 23 | OL894522 | 31 March 2019 | NE Pacific | na | NCBI |
| 45 | 26 | OL894523 | 31 March 2019 | NE Pacific | na | NCBI |
| Diaphus balanovi sp. nov. (main group) | ||||||
| 46 | 11 | OL894504 | 12 April 2019 | Emperor Seamounts | na | NCBI |
| 47 | 12 | OL894505 | 11 April 2019 | Emperor Seamounts | na | NCBI |
| 48 | 13 | OL894506 | 11 April 2019 | Emperor Seamounts | na | NCBI |
| 49 | 14 | OL894507 | 11 April 2019 | Emperor Seamounts | na | NCBI |
| 50 | 15 | OL894508 | 11 April 2019 | Emperor Seamounts | na | NCBI |
| Character | Holotype | Paratypes (n = 34) | Holotype + Paratypes | ||
|---|---|---|---|---|---|
| n * | Mean ± Error of Mean | Standard Deviation | |||
| SL, mm | 96 | 83–98 | - | - | - |
| In % of SL | |||||
| Head length | 30.2 | 27.6–31.8 | 33 | 29.6 ± 0.19 | 1.07 |
| Snout length | 2.6 | 2.6–3.6 | 32 | 3.2 ± 0.05 | 0.28 |
| Horizontal diameter of orbit | 10.4 | 9.6–11.9 | 33 | 10.7 ± 0.10 | 0.58 |
| Interorbital width | 10.4 | 8.5–12.1 | 29 | 10.3 ± 0.15 | 0.81 |
| Length of upper jaw | 19.8 | 17.7–20.5 | 31 | 19.3 ± 0.14 | 0.80 |
| Eye—maxilla distance | 7.3 | 5.5–8.5 | 31 | 6.6 ± 0.13 | 0.72 |
| Greatest body depth | 21.9 | 19.7–24.4 | 28 | 21.8 ± 0.23 | 1.24 |
| Least depth of caudal peduncle | 9.4 | 8.9–10.8 | 30 | 9.7 ± 0.11 | 0.60 |
| Caudal peduncle length | 20.8 | 17.1–21.8 (25.7) | 31 | 20.0 ± 0.29 | 1.60 |
| Distance between origins of pectoral and pelvic fins | 16.7 | 12.8–19.8 | 27 | 16.3 ± 0.34 | 1.76 |
| Distance between origins of pelvic and anal fins | 24.0 | 19.8–26.5 | 27 | 22.2 ± 0.30 | 1.58 |
| Predorsal distance | 43.8 | 42.5–46.9 | 27 | 44.7 ± 0.22 | 1.16 |
| Prepelvic distance | 44.8 | 41.5–48.2 | 29 | 45.7 ± 0.29 | 1.58 |
| Preadipose distance | 80.2 | 77.9–83.5 | 31 | 81.0 ± 0.28 | 1.59 |
| Dorsal-adipose distance | 20.8 | 16.9–20.8 (25.0) | 27 | 19.6 ± 0.32 | 1.64 |
| Length of dorsal-fin base | 14.9 | 13.8–19.6 | 28 | 16.3 ± 0.25 | 1.33 |
| Length of anal-fin base | 14.1 | 12.2–15.4 | 28 | 13.5 ± 0.16 | 0.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prokofiev, A.M.; Emelyanova, O.R.; Orlov, A.M.; Orlova, S.Y. A New Species of Diaphus Associated with Seamounts of the Emperor Chain, North-Western Pacific Ocean (Teleostei: Myctophiformes: Myctophidae) . J. Mar. Sci. Eng. 2022, 10, 65. https://doi.org/10.3390/jmse10010065
Prokofiev AM, Emelyanova OR, Orlov AM, Orlova SY. A New Species of Diaphus Associated with Seamounts of the Emperor Chain, North-Western Pacific Ocean (Teleostei: Myctophiformes: Myctophidae) . Journal of Marine Science and Engineering. 2022; 10(1):65. https://doi.org/10.3390/jmse10010065
Chicago/Turabian StyleProkofiev, Artem M., Olga R. Emelyanova, Alexei M. Orlov, and Svetlana Y. Orlova. 2022. "A New Species of Diaphus Associated with Seamounts of the Emperor Chain, North-Western Pacific Ocean (Teleostei: Myctophiformes: Myctophidae) " Journal of Marine Science and Engineering 10, no. 1: 65. https://doi.org/10.3390/jmse10010065