
How Do Prostomial Sensory Organs Affect Brain Anatomy? Phylogenetic Implications in Eunicida (Annelida)

Institute of Evolutionary Biology and Ecology, University of Bonn, An der Immenburg 1, 53121 Bonn, Germany
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2022, 10(11), 1707; https://doi.org/10.3390/jmse10111707
Submission received: 31 August 2022
/
Revised: 5 October 2022
/
Accepted: 7 November 2022
/
Published: 9 November 2022
(This article belongs to the Special Issue New Research Advances on Marine Invertebrates)
Abstract
:Eunicida is a taxon of marine annelids currently comprising the taxa Eunicidae, Onuphidae, Dorvilleidae, Oenonidae, Lumbrineridae, Histriobdellidae and Hartmaniella. Most representatives are highly mobile hunters sharing the presence of a sophisticated nervous system but differ in the number and shape of prostomial sensory organs (0–3 antennae; 0 or 2 palps; 0, 2 or 4 (+2) buccal lips; 0, 2 or 4 eyes; single-grooved or paired nuchal organs). This makes Eunicida an ideal model to study the following questions: Is the brain morphology affected by different specificities of prostomial sensory organs? Do similar numbers and shapes of prostomial sensory organs hint at close phylogenetic relationships among different eunicidan taxa? How can antennae, palps and buccal lips be differentiated? For the investigation of sensory organs and the nervous system, we performed immunohistochemistry, µCT, TEM, SEM, paraffin histology and semi-thin sectioning. Our results show that brain anatomy is mostly affected on a microanatomical level by sensory organs and that similar specificities of sensory organs support the latest phylogenetic relationships of Eunicida. Further, a reduction of antennae in Eunicida can be suggested and hypotheses about the presence of sensory organs in the stem species of Eunicida are made.
Keywords:
neuropil; nerve; reduction; nuchal organ; antenna; palp; eye; histology; electron microscopy1. Introduction
Eunicida currently comprises over 1000 species in the major taxa Eunicidae (Berthold, 1827), Onuphidae (Kinberg, 1865), Dorvilleidae (Chamberlin, 1919), Oenonidae (Kinberg, 1865), and Lumbrineridae (Schmarda, 1861) and the two minor subtaxa Histriobdellidae (Claus and Moquin-Tandon, 1884) and Hartmaniella (Imajima, 1977) [1,2,3,4,5,6]. Fossils date Eunicida back to the late Cambrian [6,7,8,9], underlining the great diversification this clade encountered over time [7]. All members of Eunicida are exclusively marine, conquering habitats ranging from shallow coastal lines to deep sea areas [7]. Most representatives of Eunicida have a predatory lifestyle: they hunt their prey actively down with their complex jaw apparatus [6,8,10]. Interaction with the environment and perception of chemical cues are performed by a set of diverse sensory organs such as head appendages (antennae and palps), buccal lips, pigment cup-shaped cerebral eyes, nuchal organs and lateral organs. The number of head appendages and eyes shows extreme variation between the different families of Eunicida and becomes visible even on the family level [3,5,11,12]: members of the family Eunicidae for example comprise species bearing five, three or one head appendage(s) and one pair of eyes, while members of the family Lumbrineridae lack head appendages as well as eyes completely. In between these extremes, diverse species-specific combinations of prostomial sensory organs are present.
All eunicidan prostomial sensory organs are innervated by nerves originating in the brain neuropil [11,13,14]. The brain is located in the prostomium and constitutes the central nervous system together with the circumoesophageal connectives and the ventral nervous system [10,13,15]. In the brain, neurites are frequently arranged in bundles with certain orientations, termed tracts or commissures [11,13,14,16]. Tracts and commissures serve as connectors, e.g., between nerves innervating sensory organs, so that different neuronal innervation patterns may become apparent among brains of different eunicidan taxa.
Since the 19th century, scientists have tried to describe the variety of head appendages in Eunicida using a set of different terms. Over time, the terminology changed according to new insights (homology hypotheses) and the application of new techniques (cLSM). A selection of these terms is given in Table S2. If compared to the latest nomenclature used, there is a historical consensus in terming the median and lateral antennae as “antennae”. Solely the studies of Monro [17] and Aiyar [18] are an exception to this in using the term “tentacle”.
Historical nomenclature of structures nowadays accepted as palps passed through greater changes. Many authors—former and present ones—use the term “antennae” to describe palps. Presumably led by the stunning same outer morphology of antennae and palps in most eunicidan taxa on the one-hand side (e.g., Figure 1) and prominent differences of eunicidan palps with, e.g., feeding and sensory palps of other annelid taxa on the other. With the advance of confocal laser scanning microscopy and generally improved imaging methods, the idea of differentiation by innervation patterns was put forward. Since palps in all annelid taxa are innervated by both the drcc and the vrcc and their commissures, this criterium served as defining feature for palps in Eunicida for the first time in the study of Orrhage and Müller [13]. Interestingly, former authors such as Hanström [12], Fauchald [19] and Fauchald and Rouse [20] used the term “palp” prior to this definition for head appendages in the “ventro-lateral” position in Dorvilleidae.
Buccal lips were described generally as the “anterior-most outgrowth” of the prostomium and similar to the classification of palps, many different terms were used to describe buccal lips over time. Most of the authors regarding all five head appendages as antennae termed buccal lips “palps”, e.g., Ehlers [5], Heider [21], Hanström [12], von Haffner [22,23], Paxton [24] and Orensanz [25]. The ventro-lateral antennae of Eunicidae and Onuphidae were homologized with the palps of Nereididae and Aphroditidae by Binard and Jeener [26] and Gustafson [27], who termed the eunicidan buccal lips consequently “faux-palpes”. Aiyar [18] who neither used the terms “antenna” nor “palp”, named the buccal lips neutrally “lobes”. This terminology for buccal lips was adopted by Åkesson [28] in labelling them “oral lobes”. Since Orrhage [14] introduced the term “buccal lip”, a consensus in the labelling of buccal lips exists. No homology of eunicidan buccal lips and palps of other annelid taxa was suggested by Binard and Jeener [24] and Orrhage [14]. The same conclusions were drawn by Zanol [29] due to differing innervation patterns. The dorsal buccal lips found in some taxa of Onuphidae passed through an intensive phase of renaming, too. From “palps” [12,22,23,24,30] over “antennae” [5,19,31] to the now-accepted term “ventral buccal lip” [6,13,14,20,29,32].
The differing numbers of head appendages make Eunicida an ideal model to study the following questions:
- Is the brain morphology affected by different numbers and shapes of prostomial sensory organs? It could be assumed, that species with many prostomial sensory organs possess complex innervation patterns for processing of stimuli, while species lacking eyes and head appendages may show a more simple brain morphology. However, it could also be hypothesized, that species with a less amount of prostomial sensory structures balance missing sensory input out by expanding another sense (e.g., chemosensation of nuchal organs) as von Haffner [11] already suggested.
- Do similar numbers and shapes of prostomial sensory organs hint at close phylogenetic relationships among different eunicidan taxa?
- How can head appendages (antennae and palps) as well as buccal lips be differentiated?
To provide answers to these research questions, we analyzed the innervation of sensory organs and reconstructed their tracts and commissures in the brain in a broad comparative morphological approach (paraffin histology, immunohistochemistry, ultrastructural analyses, scanning electron microscopy, micro-computed tomography and 3D reconstructions). Further, we compared the morphology of antennae, palps, buccal lips, eyes, and nuchal organs among the different taxa to evaluate if interspecific differences are present. These differences might explain possible variations in the innervation of sensory organs on the one hand, while on the other hand, they may serve as indicators for hypotheses of phylogenetic relationships in Annelida. Generally, morphological studies are still crucial to understand the evolution of divergent morphological traits, especially in taxa as diverse as Annelida. A mapping of morphological characters on recent molecular phylogenies of Eunicida provides a reliable base for the formulation of phylogenetic and evolutionary hypotheses, shedding new light on old evolutionary questions.
2. Materials and Methods
2.1. Animals
Specimens of Leodice (Eunice) torquata (Quatrefages, 1866), Lysidice (Nematonereis) unicornis (Grube, 1840) were collected in the intertidal zone of Le Cabellou in Concarneau (France, Brittany) in March 2017 (Table S1, Figure 1). Specimens of Scoletoma (Lumbrineris) tetraura (Schmarda, 1861), Lysidice ninetta (Audouin and Milne Edwards, 1833), Paucibranchia (Marphysa) bellii (Audouin and Milne Edwards, 1833) and Marphysa spec. were found in the intertidal zone in Concarneau (France, Brittany) in April 2017 (Table S1, Figure 1). Hyalinoecia tubicola (O.F. Müller, 1776) was collected in Bergen (Norway) in August 2017 (Table S1). Diopatra neapolitana (Delle Chiaje, 1841) was found in Zostera beds during low tide in the bay of Arcachon close to Le Petit Piquey (France, Nouvelle-Aquitaine) in 1995 (Table S1). Specimens of Dorvillea spec. were found in an aquarium in Leipzig (Germany) (Table S1, Figure 1). Specimens of Ophryotrocha siberti were collected in an aquarium in San Diego (USA) in 2015. Specimens of Arabella iricolor (Montagu, 1804) were collected in the intertidal zone in Le Cabellou in Concarneau (France, Brittany) in 2014 and 2015 (Table S1, Figure 1). Scoletoma (Lumbrineris) fragilis (O.F. Müller, 1776) was collected in Le Cabellou in Concarneau (France, Brittany). Histriobdella homari (Beneden, 1858) was collected from commercially bought Lobster in Bergen (Norway) in 2018 (Table S1).
2.2. Paraffin Histology
Specimens were relaxed in 7% MgCl2 mixed with seawater (1:1) followed by prefixation in 7% formaldehyde in seawater (1:1). Fixation was performed in the refrigerator overnight using Bouin’s fixative modified after Dubosque-Basil. The further steps applied follow the standardised methodology described by [33,34,35]. Serial sections (thickness 5 µm) of the animal’s prostomium and the peristomium were created using a Reichert-Jung Autocut 2050 microtome (Leica, Wetzlar). Specimens were either cut in cross, horizontal or sagittal sections and were stained in AZAN (for a detailed description see Beckers et al. [35]) or impregnated with silver modified after the protocol of Palmgren (1948, 1955, 1960) (Table S1).
2.3. Immunohistochemistry
One specimen of Scoletoma tetraura and two specimens of Lysidice ninetta were relaxed in a 7% MgCl2 solution mixed with seawater (1:1). Fixation was performed in a 4% paraformaldehyde solution (Electron Microscope Sciences, Hatfield, PA, USA) in seawater at 4 °C overnight. Further steps were conducted according to standardized immunohistological protocols. Samples were cut using a vibratome (Micron HM 650 V, Thermo Scientific, Dreieich, Germany) producing 60 µm thick sections. Sections were stained using FMRF-amide (rabbit; ImmunoStar, Hudson, WI, USA) at a dilution of 1:2000 and α-tubulin (mouse; Sigma-Aldrich, Germany) at a dilution of 1:500 as primary antibodies. The secondary antibodies Cy2 (goat anti-rabbit; ImmunoStar, Hudson, WI, USA) and Cy5 (goat anti-mouse; Sigma-Aldrich, Germany) were added in a dilution of 1:500 each. Sections were mounted on chromalaun/gelatine-covered glass slides and embedded with Elvanol (after Rodriguez and Deinhard (1960) modified after M. Bastmeier). Examination of the sections was performed using a Leica confocal laser scanning microscope (TCS SPE, Germany, excitation wavelength: 488 nm/635 nm) and images of the samples were taken via the software LAS AF 1.6.1 (Leica, Microsystems).
2.4. Micro-Computed Tomography (µCT)
Specimens of Lysidice ninetta, Leodice torquata and Hyalinoecia tubicola were investigated via micro-computed tomography (µCT) (Table S1). First, specimens were relaxed in a solution of 7% MgCl2 mixed with seawater (1:1). Prefixation was performed in a 7% formaldehyde solution mixed with seawater (1:1) and fixation was performed in Bouin‘s fixative overnight. Samples were washed with 70% ethanol and stained in a 0.3% phosphotungstic acid (PTA) solution in 70% ethanol for two weeks. Imaging was performed using a SkyScan 1272 µCT scanner (Burker, Kontich, Belgium), which was equipped with a Hamamatsu L11871 20 tungsten X-ray source (Hamamatsu, Japan) and a Ximea xiRAY 16 camera (XIMEA GmbH, Germany). The following scanning parameters were used: source voltage = 70 kV, source current = 142 µA, exposure time = 1430 ms, frames averaged = 10, flat field correction = activated and scanning time = 14–20 h.
2.5. Electron Microscopy
For scanning electron microscopy (SEM) trunk segments of Paucibranchia bellii, Marphysa spec., Lysidice unicornis, Lysidice ninetta, Leodice torquata, Hyalinoecia tubicola, Diopatra neapolitana, Ophryotrocha siberti, Arabella iricolor and Scoletoma tetraura were investigated focusing on the lateral organs (Table S1). Fixation and further processing of the samples were conducted according to the methodology described by Beckers et al. [33]. Dehydration was performed in an ascending acetone series if followed by critical-point drying (Bal-Tec CPD 030, Switzerland) or via an ascending ethanol series if treated with HMDS (hexamethyldisilazane). After mounting on aluminium stubs, samples were sputter coated with gold (SEM coating unit E5100, Polaron Equipment Ltd., Great Britain) and were examined via a scanning electron microscope (Leitz AMR 1000, FEI Verios 460 L).
The anterior part of one specimen of Paucibranchia bellii and Scoletoma fragilis was investigated via semi-thin sections using light microscopy (BX-51, Olympus). Additionally, Paucibranchia bellii was examined via ultra-thin sections in transmission electron microscopy (TEM). Fixation and further processing of the samples were performed according to the methodology described by Beckers et al. [33]. Then, 1 µm semi-thin and 70 nm thick ultra-thin sections were created using a Leica EM Ultracut 6 microtome and a diamond knife. Semi-thin sections were transferred to glass slides and stained with Toluidine blue. Ultra-thin sections were placed on formvar-coated single slot copper grids (1 × 2 mm) and stained with uranyl acetate and lead citrate automatically (QG-3100, Boeckler Instruments). Examination of the ultra-thin sections was conducted using a transmission electron microscope (Zeiss EM 10CR). Negative pictures documented on phosphor imaging plates were digitalized using an imaging plate scanner (Ditabis).
2.6. Data Analyses and 3D Reconstructions
Living specimens were photographed using a Canon 600D camera mounted on a Zeiss-Stemi 2000. Paraffin and semi-thin sections were analysed with an Olympus microscope (BX-51) that was equipped with an Olympus camera (Olympus cc12) and the dot slide system (2.2 Olympus, Hamburg). Automatic alignment of the images was performed using imod and imodalign (http://www.q-terra.de/biowelt/3drekon/guides/imod_first_aid.pdf, accessed on 5 October 2022). For the creation of an image stack the images were loaded in ImageJ 1.51 k (Wayne Rasband, National Institutes of Health, USA, http://imagej.nih.gov/ij, accessed on 5 October 2022) and segmentation was performed using TrackEM in ImageJ. The 3D reconstructions were created in Amira 5.3.1 (Thermo Fisher Scientific, Waltham, MA, USA).
Z-projections of the immunohistological image stacks were created using ImageJ 1.51 k and the function “maximum intensity”. If images needed to be stitched together, 3D stitching was performed in ImageJ 1.51 k.
Two-dimensional (2D) image slicing of the µCT data was performed using ImageJ 1.51 k and the plug-in Volume Viewer. The 3D rendering was conducted in Amira 5.3.1 and 3D reconstructions of the nervous systems of the samples were performed by manual segmentation and surface rendering.
Image processing and creation of schematic drawings were conducted in Adobe Photoshop (CS6) and Adobe Illustrator (CS6).
3. Results
The neuroanatomical nomenclature used in this study follows the terminology given by Richter et al. [36]. Without proposing a general homology with palps of other annelids, the term “palp” is used here to describe head appendages innervated by two or more nerve roots originating from the dorsal root of the circumoesophageal connective (drcc) and the ventral root of the circumoesophageal connective (vrcc) and their commissures [13]. According to Orrhage and Müller [13] and Orrhage [14], antennae are solely innervated by nerves emanating from the drcc. Based on the study of Zanol [29], we use the adjective “eunicid” in context with all members belonging to the family Eunicidae, while “eunicidan” refers to members of the order Eunicida.
3.1. The Central Nervous System
The central nervous system (cns) of Eunicida is basiepidermal (intraepidermal) and consists of a central neuropil (brain), the circumoesophageal connectives (cc) and the paired ventral nerve chord (vnc) (Figure 2A–F). The brain is located dorsally in the prostomium and is connected via the cc in the peristomium to the suboesophageal ganglion and the subsequent paired ventral nerve chord in the metameric trunk (Figure 2A–F). The cc splits in a dorsal (drcc) and a ventral root (vrcc) in the brain and the ganglionic ventral nervous system forms regular segmental commissures running through the trunk (Figure 2A–F).
Comparing brain positions among the different species studied, the brain of Arabella iricolor (Oenonidae), Scoletoma tetraura and Scoletoma fragilis (Lumbrineridae) is positioned a bit more posteriorly in the prostomium than in the remaining taxa (Figure 2G,H). This impression might be consolidated by the elongated prostomium of these taxa.
3.2. General Brain Anatomy
In all species investigated, the brain surrounds the intracerebral cavity (Figure 3A,C,F,G). Anteriorly to this point, the brain splits into smaller neuropils, from which many delicate nerves arise innervating the tip of the prostomium (Figure 2G,H). At the height of the intracerebral cavity, dorsal and ventral brain neuropil can be differentiated with regard to their relative topology (Figure 3A,C,F,G). The dorsal brain neuropil innervates all dorsally located sensory organs (such as antennae, eyes and nuchal organs) (Figure 3C,D,H,I) and inhabits the respective commissures and tracts, e.g., optical commissures and association tracts of antennae. Parts of the ventral brain neuropil innervate the buccal lips—if present. Palps are innervated by both, the dorsal and the ventral brain neuropil. The oesophageal ganglion is still part of the ventral brain neuropil and gives rise to the jaw nerves (Figure 3D,E,H,I). The dorsal root of the circumoesophageal connective (drcc) originates from the dorsal brain neuropil (Figure 3D,I). The ventral brain neuropil gives rise to the ventral root of the circumoesophageal connective (vrcc) (Figure 3I). In many species, e.g., in Lysidice ninetta, the dorsal and the ventral commissure of the ventral root of the circumoesophageal connective (dcvr/vcvr) can be differentiated (Figure 2D). With this, the ventral brain neuropil terminates. The dorsal brain neuropil proceeds posteriorly and innervates the nuchal organ before it terminates (Figure 2G).
Deviations from this brain morphology occur in Arabella iricolor and Scoletoma tetraura. In both species, the anteriorly located neuropil parts extend much more ventrally than in all other species examined (Figure 3H). In A. iricolor, the oesophageal ganglion originates from the ventral parts of these neuropils (Figure 3H,I). The oesophageal ganglion is connected to the circumoesophageal connective via a pair of distinct commissures (Figure 3G,I). Both the commissures as well as the oesophageal ganglion are surrounded by a thick layer of intermediate filaments (Figure 3G). The paired stomatogastric nerves originate from these commissures innervating the stomatogastric system including the oesophagus (Figure 3J). This situation was not observed in any other species examined. Further, the intracerebral cavity in A. iricolor is much smaller than in any other taxon investigated (Figure 3F,H). In S. tetraura the ventrally extended anterior neuropil parts fuse in the posterior course so that together with the dorsal brain neuropil it surrounds the intracerebral cavity.
Additionally, in A. iricolor, as well as S. tetraura, distinct neuropils occur at the lateral sides of the brain neuropil. In both species, these lateral neuropils extend dorso-ventrally and they are surrounded by a thick layer of somata of type 1 neurons (Figure 4A,D). The lateral neuropils are interconnected by a tract located in the dorsal brain neuropil (Figure 3F,G and Figure 4B,C,E). The tract is surrounded by intermediate filaments of glia cells (Figure 3B,F and Figure 4C). In A. iricolor, the dorsal nerves root in this tract and the dorsal brain neuropil (Figure 4C).
3.3. Ultrastructure of the Central Neuropil and the Somata
In all species investigated, the central neuropil consists of neurites and is surrounded by somata of different neuronal types, that are discriminated by size (Figure 5A–C,E). The neurites can have different diameters and orientations and parallel-oriented neurite bundles create commissures and tracts in the otherwise heterogeneous central neuropil (Figure 5A,D). Dense and lucent core vesicles comprising neuropeptides are present in the neurites (Figure 5F) and stabilizing intermediate filaments of glial cells surround the neurites (Figure 5D). Blood vessels may traverse the brain. A thin layer of glial cells and their processes isolate the central neuropil from the surrounding somata (Figure 5A,D). Each soma is isolated by several layers of glia cell processes (Figure 5D) while desmosomes connect adjacent glia cell processes (Figure 5D). A glial layer surrounding the neuronal somata is not visible (Figure 5A).
Somata of type 1 neurons (S1,) are densely packed and their nuclei appear as reddish–purple spherical structures with a size of 4.4–7.5 µm (Figure 5A,B). S1 neurons occur predominantly on the lateral sides of the central neuropil (Figure 5A). As somata of type 2 neurons (S2) possess more glia enwrapping than S1 neurons, S2 neurons appear less dense packed. Both types of neurons share a similar-sized nucleus (Figure 5A,C). S2 neurons occur constantly around the central neuropil surrounding it dorsally, laterally as well as ventrally (Figure 5A). Somata of type 3 neurons (S3) are enlarged and their nuclei are more prominent (13.2–26.3 µm) than the nuclei of S1 and S2 neurons (Figure 5A,E). The nucleolus can easily be identified in S3 neurons due to its brightly red stain while the nucleus itself appears light blue (Figure 5E). S3 neurons do not occur constantly around the central neuropil since they are refined to certain areas: they occur medio-dorsally and ventro-laterally to the central neuropil and only at the frontal part of the dorsal neuropil (Figure 5A).
3.4. Innervation Patterns of Antennae in the Brain
Antennal innervation patterns in the brain may differ among the species studied according to the number of antennae and palps present. Most complex innervation patterns can be found in Eunicidae bearing three antennae and two palps (Paucibranchia bellii, Marphysa spec., Leodice torquata) in possessing so-called association tracts (sensu Orrhage 1995), while association tracts seem to be missing in all other species examined (Figure 6).
3.4.1. Innervation Patterns in Eunicidae (Five Head Appendages)
In Pauchibranchia bellii, Marphysa spec. and Leodice torquata, antennae are innervated by one main nerve (man, lan) (Figure 7A). In the median antenna, this nerve splits into two main nerve roots (manr1) at the height of the ceratophore (Figure 7A and Figure 8B–D). The main nerve roots run along both sides of the antennae coelom towards the brain. Each manr1 splits into three minor nerve roots (manr2, manr3, atma) prior to entering the dorsal brain neuropil (Figure 7A). The manr2 and manr3 roots in the dorsal commissure of the dorsal root of the circumoesophageal connective (dcdr). The third minor nerve root is termed the association tract (atma) according to Orrhage (1995) and runs ventrally through the dorsal brain neuropil (Figure 7A and Figure 8A,D,E,G). Both association tracts of the median antenna (atma) fuse in the dorsal neuropil (Figure 7A and Figure 8E,G), and split again at the ventral side of the dorsal brain neuropil (Figure 7A and Figure 8B–D). There, each atma fuses with the respective association tract of the lateral antennae (atla) (Figure 7A and Figure 8B,D). The product of this fusion is the paired association tracts of the median and the lateral antennae (atmla) that proceed next to the intracerebral cavity in the anterior direction through the dorsal brain neuropil (Figure 7A and Figure 8D). In the anteriorly located part of the brain the paired atmla fuse in a larger area of intensively stained neurites. In this area, the dorsal roots of the palp nerves originate.
Similar to the innervation of the median antenna, the main nerve of the lateral antennae splits into two main nerve roots running alongside the antenna coelom (llanr, mlanr1) in P. bellii, Marphysa spec. and L. torquata (Figure 7A). Prior to entering the dorsal brain neuropil, both main nerve roots split into minor nerve roots. The lateral nerve root of the lateral antenna (llanr) splits into two minor nerve roots (latla1, matla1) (Figure 7A). The more medially located nerve root of the lateral antenna (mlanr1) splits into three minor nerve roots (mlanr2, latla2, matla2) (Figure 7A). Both lateral association tracts of the lateral antenna (latla1, latla2) fuse in the dorsal brain neuropil creating the first association tract of the lateral antenna (atla1) (Figure 7A). The atla1 proceeds ventrally and fuses with the dorsal fibril mass (dfm) (Figure 7A and Figure 8C). Both medially located association tracts of the lateral antenna (matla1, matla2) fuse in the dorsal brain neuropil, constituting the second association tract of the lateral antenna (atla2). The atla2 proceeds diagonally through the dorsal brain neuropil and fuses with the atma, creating the atmla (Figure 7A). Additionally, a further minor nerve root (mlanr3) splits from the matla2. This nerve root, the third median nerve root of the lateral antenna (mlanr3), roots in the horizontally proceeding commissure of the association system (cas) (sensu Orrhage 1995) (Figure 7A). The cas is a commissure that connects the matla2 of both lateral antennae via the mlanr3. The cas proceeds anteriorly originating in the dorsal brain neuropil.
3.4.2. Innervation Patterns in Hyalinoecia tubicola (Onuphidae)
The innervation pattern of the median antenna of Hyalinoecia tubicola (5 head appendages) concurs with that of eunicids bearing five head appendages (Figure 7). Due to the oblique cutting plane of the histological sections, tracking of association tracts was impeded in H. tubicola.
Lateral antennae in H. tubicola are innervated differently: although the lateral antenna is innervated by one main nerve (lan), too, it does not split into a pair of minor nerves. Instead, the lateral antenna nerve proceeds along the medially located side of the lateral antenna coelom towards the brain (Figure 7B). Prior to entering the dorsal brain neuropil, the lan roots at least into two minor nerve roots (lanr, atla). The more medially located nerve root (lanr) originates from the dcdr, while the association tract (atla) proceeds in a ventral direction through the dorsal brain neuropil (dnp) (Figure 7B). Before reaching the ventral part of the dorsal brain neuropil, the atla splits into two minor tracts, the median and the lateral association tract (latla, matla). The matla fuses with the respective association tract of the median antenna (atma) at the ventral side of the dorsal brain neuropil (Figure 7B). The product of this fusion is the paired association tracts of the median and the lateral antennae (atmla) (Figure 7B). The atmla run in the anterior direction through the dorsal brain neuropil and originate medially in the neuropil. The latla proceeds in the ventral direction through the dorsal brain neuropil rooting in the dorsal fibril mass (dfm) (Figure 7B).
3.4.3. Innervation Patterns in Eunicidae (Three and One Head Appendages)
In Lysidice ninetta (three antennae) and Lysidice unicornis (one median antenna), antennae are innervated by one main nerve (man/lan). As in P. belli, Marphysa spec. and L. torquata, the main nerves split at the height of the ceratophore into two nerve roots (2x manr1/llanr, mlanr1), running along the sides of the respective antenna coelom towards the dorsal brain neuropil. At least two minor nerve roots (manr2, manr3) are present in the median antenna of both species. The manr2 as well as the manr3 root in the dcdr. Distinct association tracts were not observed for the median antenna.
In L. ninetta, the lateral root of the lateral antennae nerve (llanr) splits into two minor nerve roots. Whether these nerve roots are association tracts is not clear, since tracking is impeded by the oblique cutting plane. At least one nerve root seems to proceed in ventral direction through the dorsal brain neuropil and could be interpreted as an association tract. As the association tracts (atla1) of P. belli, Marphysa spec. and L. torquata, it seems to originate at least in close proximity to the dorsal fibril mass (dfm). The other nerve root seems to originate more dorsally in the dorsal brain neuropil. The median root of the lateral antenna nerve (mlanr1) proceeds in one specimen of L. ninetta until the dorsal brain neuropil, but nerve roots could not be distinguished. In another specimen of the same species, the course of the mlanr1 could not be tracked or it simply shows no connection to the dorsal brain neuropil.
3.4.4. Innervation Patterns in Dorvillea spec. (Dorvilleidae)
In Dorvillea spec. (two lateral antennae, two palps) each antenna is innervated by one main nerve similar to the conditions found in the remaining species studied. Identification of antennal nerve roots is hampered by the oblique cutting plane of the histological sections. Nevertheless, two nerve roots (mlanr1, llanr) and several minor nerve roots could be identified as innervating the lateral antennae. The llanr is much more prominent than the relatively small and inconspicuous mlanr1, but both nerve roots proceed along the sides of the lateral antenna coelom towards the brain. The llanr roots at least with two minor nerve roots in the dorsal brain neuropil. The number of nerve roots of the mlanr1 as well as association tracts generally could not be identified.
3.4.5. Conditions in Oenonidae and Lumbrineridae (No Head Appendages)
Arabella iricolor (Oenonidae) lacks head appendages, but a pair of dorsal nerves (dn) is present resembling the median antenna nerves of the remaining taxa topologically (Figure 3F,H,I, Figure 4B,C and Figure 8F). The paired dorsal nerves surround the median coelomic cavity and root in a tract connecting the lateral neuropils (tlnp) as well as the dorsal brain neuropil (Figure 4B,C). At the insertion point of the dorsal nerves, the lateral neuropil tract forms a cushion-like neuropil area located medially in the dorsal brain neuropil (Figure 4C). This cushion-like neuropil area is surrounded by intermediate filaments of radial glia cells. Association tracts (sensu Orrhage 1995) are missing. Scoletoma tetraura and Scoletoma fragilis (Lumbrineridae) lack head appendages, association tracts and dorsal nerves (Figure 4E).
3.5. Prostomium and Prostomial Sensory Organs
In the examined representatives of Oenonidae and Lumbrineridae the prostomium is elongated and tapered (Figure 1G,H) while in Eunicidae and Onuphidae it is roundish with or without distinct buccal lips (Figure 1A–E). In Dorvilleidae the prostomium is very small and roundish (Figure 1F).
Prostomial sensory organs occur in species-specific combinations and morphology in Eunicida (Figure 1, Table 1). Antennae, palps, eyes and nuchal organ(s) are located on the posterior part of the prostomium dorsally to dorso-laterally while buccal lips are located ventrally (Figure 1 and Figure 2). The nuchal organ is the only prostomial sensory organ completely internalized: not more than the opening(s) of the nuchal cavity may be recognized from the outside (Figure 1).
3.6. Head Appendages (Antennae and Palps)
Paucibranchia bellii, Marphysa spec., Leodice torquata (Eunicidae) and Hyalinoecia tubicola (Onuphidae) possess one median antenna, one pair of lateral antennae and one pair of palps (Figure 1A,D,E, Table 1). One pair of lateral antennae and one pair of palps are present in Dorvillea spec. (Dorvilleidae) (Figure 1F, Table 1), while Lysidice ninetta (Eunicidae) bears one median antenna and one pair of lateral antennae (Figure 1C, Table 1). One median antenna is present in Lysidice unicornis (Eunicidae) (Figure 1B, Table 1). Arabella iricolor (Oenonidae), Scoletoma tetraura and Scoletoma fragilis (Lumbrineridae) lack head appendages (Figure 1F,G, Table 1 Outer Morphology of Antennae and Palps).
Antennae and palps are digitiform and possess a basal ceratophore (antennophore/palpophore) and a distal ceratostyle (antennal style/palpostyle) (Figure 1). Antennae and palps have a white-beige pigmented, smooth surface in P. bellii (Eunicidae), Marphysa spec. (Eunicidae), L. unicornis (Eunicidae), L. ninetta (Eunicidae) and Dorvillea spec. (Dorvilleidae) (Figure 1A–C,E,F). In L. torquata, the ceratophores and palpophores have a white pigmentated, smooth surface, while the ceratostyles and palpostyles are annulated in white and red (Figure 1D). In H. tubicola, the ceratophores and palpophores are annulated, while the ceratostyles and palpostyles have smooth surfaces (Figure 1E). Further, differences in the length of antennae and palps appear between the species: H. tubicola and Dorvillea spec. possess relatively long head appendages when compared to the size of their prostomia (Figure 1D,E). P. bellii, Marphysa spec. and L. torquata bear head appendages of moderate length (Figure 1A,D) while antennae of L. unicornis and L. ninetta are relatively short compared to the size of their prostomia (Figure 1B,C).
Inner Morphology of Antennae and Palps
Besides a similar outer morphology, antennae and palps share a similar inner anatomy in most of the species studied. Generally, one nerve consisting of longitudinal and circular directed nerve fibres can be found in the ceratostyles as well as the palpostyles (Figure 9A,B). Longitudinal nerve fibres are located medially and are surrounded by circular nerve fibres (Figure 9A). Nuclei of glial cells can be found in both kinds of nerve fibres (Figure 9A). Both nerve fibre-types run through the ceratostyle/palpostyle until the tip of the respective head appendage (Figure 9B). A coelomic cavity (here referred to as “antenna coelom” or “palpus coelom”) containing musculature is located in the ceratophores and the palpophores of all species investigated (e.g., Figure 9C–H). While the composition of antennae in Dorvillea spec. is the same as in the remaining taxa, its palps are differently organized: the coelomic cavity and the musculature are not limited to the palpophore, but extend until the tip of the respective head appendage (Figure 9F,G).
3.7. Innervation of Palps
In the studied Eunicidae and Onuphidae, palps are innervated by one main nerve (pn) running along the more medially located side of the palp coelom towards the brain (Figure 9D,E). Before entering the brain, the palp nerve splits into several minor nerve roots—in most species studied three minor palp nerve roots (pnr1, pnr2, pnr3) are present (Figure 7). The two more dorsally located palp nerve roots (pnr1, pnr2) originate from the dorsal brain neuropil (Figure 7 and Figure 9E). The more ventrally located palp nerve root originates from the ventral brain neuropil (Figure 7E and Figure 9E). The exact number of minor nerve roots was in some specimens hard to identify because oblique cutting planes hampered tracking.
In Pauchibranchia bellii, we found 3–4 minor nerve palp roots (3 rooting in the dnp (one of which rooting in the dfm) and probably 1 rooting in the vnp). For Marphysa spec. we could identify only one minor palp nerve root, rooting in the dfm (dorsal brain neuropil). Probably more nerve roots are present. In Leodice torquata, up to four or more minor palp nerve roots can be present depending on the specimen we are looking at. There were 1–2 nerve roots originating from the vnp, while 2–3 nerve roots were found rooting in the dnp. The exact number of palp nerve roots for Hyalinoecia tubicola could not be determined due to the oblique cutting plane.
In Dorvillea spec. two distinct palp nerves (dpn, vpn) innervate each palp (Figure 9F,G). Both palp nerves proceed from the basal to the distal end of each palp running along both sides of the coelomic cavity. The more ventrally located palp nerve (vpn) fuses via at least two minor nerve roots with the ventral brain neuropil, while the more dorsally located palp nerve (dpn) and its minor nerve roots originate from the dorsal brain neuropil. At least two minor nerve roots were observed for np.
3.8. Buccal Lips
Distinct buccal lips occur as paired structures in front of the mouth opening on the ventral side of the prostomium in the studied specimens of Eunicidae and Hyalinoecia tubicola (Onuphidae) (Figure 10A,B). In Eunicidae one pair of more or less rectangular-shaped buccal lips is present with a smooth surface (Figure 10A). In H. tubicola two pairs of buccal lips are present—one dorsal (dbl) and one ventral (vbl) pair –plus on additional pair of accessory buccal lips (abl) (Figure 10B). The more or less rectangular-shaped ventral buccal lips of H. tubicola are placed directly in front of the mouth opening and have a smooth surface. The dorsal buccal lips and the accessory buccal lips are spindle-shaped with a rough surface and are positioned in pairs anteriorly to the ventral buccal lips. Buccal lips are innervated by nerves (bln) originating from the ventral brain neuropil (Figure 10D,E). The buccal lip nerves proceed in the ventral direction alongside the buccal cavity (blc) and fuse eventually, building a loop. All remaining taxa studied belonging to the families of Dorvilleidae, Oenonidae and Lumbrineridae lack buccal lips (exemplified for Oenonidae, Figure 10C).
3.9. Eyes
Eyes are multicellular cerebral eyes of the pigment cup type and can be present as two pairs, one pair, or they are absent (Figure 1 and Figure 11, Table 1). In species possessing one pair of eyes (Paucibranchia bellii, Marphysa spec., Lysidice unicornis, Lysidice ninetta, Leodice torquata; Hyalinoecia tubicola), eyes are located between the lateral antennae and the palps (Figure 1A,D,E), or in a topologically equal position, if head appendages are lacking (Figure 1B,C,G). In species possessing two pairs of eyes (Dorvillea spec.; Arabella iricolor), the more anteriorly located pair of eyes resemble those of species with one pair of eyes in topology (Figure 11). The posteriorly located pair of eyes is smaller than the anterior pair of eyes (Figure 11B,E). In Dorvillea spec. the posterior pair of eyes is located dorsally to the lateral antennae (Figure 1F), while the posterior pair of eyes of A. iricolor can be found in an equivalent position (Figure 1G). In some of the studied specimens (A. iricolor, L. torquata) additional small eyes of the same type can be found next to the “regular” eyes (Figure 11B).
3.9.1. Inner Morphology of Eyes
The eyes of the examined Eunicida share the same inner structure. The microvilli of the photoreceptor cells project into the eye cup, which is made up of pigmented supportive cells (Figure 11A). The layer of pigmented supportive cells is surrounded by a dense layer of photoreceptor cells (Figure 11A). In Marphysa spec., L. ninetta, L. torquata, H. tubicola and Dorvillea spec. a cuticular invagination projects from the outer cuticle of the animal into the eye (Figure 11A,B,C). In all these species—except for Marphysa spec.—this invagination forms a lens-like structure inside the eye.
3.9.2. Innervation of Eyes
Each eye is innervated by one distinct optical nerve, which splits into fine nerves next to the eye (Figure 11A,B,E). The optical nerves fuse in the brain by forming an optical commissure (Figure 11C–E). In species possessing one pair of eyes, one optical commissure is present (Figure 11C,D), in species possessing two pairs of eyes two optical commissures are present (Figure 11E). The optical commissure connecting the more anteriorly located pair of eyes is positioned anteriorly to the second optical commissure as well as the dorsal commissure of the dorsal root of the circumoesophageal connectives (dcdr) (Figure 11E).
3.10. Nuchal Organs
All species studied possess a nuchal organ. The nuchal organ invaginates from the dorsal surface right behind the median antenna—if present—building either an unpaired or a paired ciliated cavity, the latter having separate openings. The shape of the nuchal organ can vary between families. The unpaired nuchal organ is either arc- or x-shaped (Figure 12A–C,F–I,L, Table 1), while the paired nuchal organ can be a simple invagination or an elaborate branching structure (Figure 12D,E,J,K, Table 1). All nuchal organs end blindly.
3.10.1. Morphology of Nuchal Organs
Nuchal organs share a similar composition among the species studied. A relatively thick layer of cuticle lines the nuchal cavity (Figure 12A–F) and patches of cilia extend into the nuchal cavity perforating the cuticle (Figure 12B,D). Two patches of cilia can be found in the paired as well as in the unpaired nuchal organ. In the arc-shaped nuchal organs, these ciliary patches are located on the ventral side of the nuchal organ (Figure 12A,B,H,I). In the branched and x-shaped nuchal organs, the ciliary patches are located in the dorsal branches of the nuchal cavity (Figure 12E,F,K,L). In all nuchal organs, the ciliary patches originate at the lateral sides of the nuchal cavity. More posteriorly, the ciliary patches enlarge until the basal side of the nuchal organ is covered nearly completely with cilia (Figure 12B). Only a small area in the middle of the basal side is not covered with cilia (Figure 12B). The ciliary patches can be everted into the nuchal cavity (Figure 12B).
3.10.2. Innervation of Nuchal Organs
Innervation of the ciliary patch cells is provided by fine nerves present in the tissue ventrally to the nuchal organ (Figure 12B,E–G). These fine nerves bundle in the paired nuchal organ nerves, which originate from the posterior-most part of the dorsal brain neuropil (Figure 2G and Figure 12E,F,M,O). No nuchal commissure was observed in the brain neuropil. Adjacent to the nuchal organ nerve, intermediate filaments of glial cells are present. Longitudinal and transversal musculature attaches at the lateral ends of the nuchal organ proceeding in the ventral direction until reaching the oesophagus. The nuchal cavity ends approximately at the height of the oesophagus and only musculature attaching posteriorly to the nuchal organ is still present. The ciliated patches of L. unicornis are covered by an orange-coloured layer in the AZAN-stained sections—possibly a secretion of gland cells.
3.10.3. Shape Differences of the Nuchal Organ
Regarding the shape of the nuchal organ, the following clusters can be found. The examined specimen of Eunicidae and Onuphidae possess an unpaired, arc-shaped nuchal organ (Figure 12A–C,H,I). The nuchal organ of Dorvillea spec. is a paired, simple invaginated structure (Figure 12D,J), A. iricolor possesses a paired, branched nuchal organ (Figure 12E,K,M,N) and the examined representatives of Lumbrineridae possess an unpaired, x-shaped nuchal organ (Figure 12F,L,O, Table 1).
Some peculiarities occur in the nuchal organs of Dorvilleidae, Oenonidae and Lumbrineridae. First of all, in Dorvillea spec. the nuchal cavities are interconnected anteriorly and they are connected to ciliary patches located on the lateral sides of the prostomium. Two of these ciliary patches are located on both sides of an elevation between the lateral antenna and the palp. More anteriorly, the elevation shrinks and the two ciliated patches fuse. In the posterior course, the more dorsally located ciliated patch is connected to the nuchal organ of the respective side (Figure 12J).
In A. iricolor (Oenonidae), the nuchal cavities start as simple invaginations resembling the arc-shaped condition of Eunicidae and Onuphidae but being separated. In the posterior course, the cavities branch due to lateral eversions. The shape of the nuchal organs resembles facing arrow heads (“>” “<”) at this point (Figure 12E,K). More posteriorly, the dorsal and ventral branches split eventually, so that four branches are present ending blindly.
Representatives of Lumbrineridae (S. fragilis, S. tetraura) possess a single-grooved, x-shaped nuchal organ (Figure 12F, Table 1). Similar to A. iricolor, the nuchal organ resembles the arc-shaped condition only shortly after its invagination. At this point, the lateral ends of the nuchal organ are enlarged building ciliated cavities already differing from the condition in Eunicidae and Onuphidae. These enlarged chambers are covered by cilia, while the remaining parts are covered by cuticles only. More posteriorly, the nuchal organ branches and resembles an “x” in its shape (Figure 12F,L). The branches are still connected to each other contrary to the condition in A. iricolor (Oenonidae). More posteriorly, the nuchal organ is separated in the middle and ends blindly.
4. Discussion
4.1. Phylogenetic Significance of Sensory Organs in Eunicida
Sensory organs in Eunicida are quite diverse and show variations in number, shape, and innervation. Therefore, it is possible to group certain characters (Table 2) and draw homology hypotheses about their evolutionary origin.
A mapping of characters on the latest phylogenetic tree of Eunicida (based on Tilic et al. [2]) delivers the following hypotheses (Figure 13):
- Antennae, palps, two pairs of multicellular adult eyes, nuchal organs and lateral organs seem to have already evolved in the common stem lineage of Eunicida. It is most parsimonious to assume, that the stem species of Eunicida possessed two pairs of multicellular adult eyes. In this scenario, a reduction of adult eyes took place independently in Lumbrineridae (complete loss of adult eyes), Eunicoidae (reduction to 1 pair of adult eyes) and Histriobdellidae (complete loss of adult eyes?). In the literature, there is no report about eyes in Histriobdellidae, but according to Rouse et al. [1], eyes may be present in Steineridrilus cirolanae (Führ, 1971). It can be assumed, that the stem species of Eunicida possessed a roundish prostomium and that in Oenonoidea a change of shape to a tapered, elongated prostomium occurred (Figure 13). This shift concurs with the hypothesized change of lifestyle from epi- to endobenthic.
- Antennae were reduced several times independently in Eunicida. A reduction of antennae took place once in Oenonoidea (sensu Tilic et al. [2]) with Lumbrineridae lacking antennae completely and Oenonidae having, at least in some species, remnants of either antennae, e.g., in Oenone (Lamarck, 1818), or antennae nerves (dorsal nerves), e.g., in Arabella iricolor. Dorsal nerves were observed by Zanol [20] in Schistomeringos pectinata (Perkins, 1979) (Dorvilleidae) and based on the similar topology and direction of nerves, she suggested a homology of dorsal nerves and median antenna nerves. Our results are in accordance with this hypothesis. Generally, it is assumed that nerves are attracted by the morphological structure (e.g., antennae) instead of initiating the formation of the structure itself [32] making a reduction of antennae even more plausible. Further, the median antenna was reduced completely in Dorvilleidae and in some specimens of Eunicidae the lateral antennae were reduced, e.g., in Nematonereis unicornis.
- Palps are homologous structures among Eunicida. A complete reduction of palps took place in Oenonoidea, supporting the sister group relationship of Lumbrineridae and Oenonidae. All other eunicidan families possess palps in the ground pattern. In some specimens of Eunicida palps have been reduced, e.g., in Lysidice ninetta and Nematonereis unicornis. Similar to the morphology of eunicidan antennae, the palp coelom as well as palp musculature are limited to the palpophore in most Eunicida. This is an unusual condition when compared to palps of other annelids. The palp coelom as well as the palp musculature are normally not limited to the palpophore but extend all the way up until the tip of the palp. The only eunicidan palps observed matching this pattern are those of Dorvillea spec.
- Buccal lips are not homologous to palps due to differing innervation patterns. The innervation of buccal lips was described by Orrhage [14] for Hyalinoecia tubicola: The dorsal buccal lips are innervated by nerves emanating from the lateral parts of the first ventral commissure of the ventral root of the circumoesophageal connective (vcvr1) while the ventral buccal lips are innervated by nerves rooting laterally in the third ventral commissure of the ventral root of the circumoesophageal connective (vcvr3).
- Buccal lips are an autapomorphy of Eunicoidea (sensu Tilic et al. [2]). In Eunicidae solely one pair of ventral buccal lips is present, while in Onuphidae additionally, one pair of dorsal buccal lips is present. In Hyalinoecia tubicola one pair of accessory buccal lips was observed besides the paired dorsal and ventral buccal lips.
- The shape of the nuchal organ can be used to infer phylogenetic relationships in the studied Eunicida. Oenonoidea have a branched nuchal organ while the nuchal organ of Eunicoidea is unpaired and arc-shaped. In Dorvilleidae nuchal organs were duplicated. In Histriobdellidae the nuchal organ is a paired, simple structure. The shape of the nuchal organ in the eunicidan ground pattern remains unresolved.
- Association tracts can be regarded as an autapomorphy of Eunicoidea as long not proven to be present in Histriobdellidae, too. Association tracts seem to be restricted to species possessing three antennae and one pair of palps (=species belonging to Eunicoidea), making it theoretically possible that association tracts might also be present in Histriobdellidae (=possessing three antennae + one pair of palps). However, in the histological serial section of Histriobdella homari we could not observe any association tracts.
4.2. Is the Brain shape Influenced by Sensory Structures?
The central nervous system of all examined Eunicida concurs with descriptions of the general annelid neuroanatomy [10,13,15,29,32,37]. Differences occur only in the condensed oesophageal ganglion of Arabella iricolor (Oenonidae): here, the oesophageal ganglion is much more prominent than in all other species investigated. It is additionally connected via commissures to the circumoesophageal connectives. Due to the condensed oesophageal ganglion, the brain of A. iricolor does not form a similar-shaped ring around the stomatogastric cavity as in all other examined species.
The brain shows a consistent morphology among the species studied: A dorsal and ventral brain neuropil can be distinguished topologically, separated by an intracerebral cavity [26,29]. As in other polychaetes, the brain can be defined as the frontal-most mass of neuropil surrounded by somata and can be interpreted as a “single entity” as already Malaquin [38] stated in his argumentation for an unsegmented brain [14]. The general brain anatomy is not affected to a great extent by sensory structures. Deviations occur only on a microanatomical level in the presence of sensory commissures, association tracts and lateral neuropils.
4.2.1. Optical Commissure
Species bearing one pair of eyes possess one optical commissure (Eunicidae, Onuphidae). Species with two pairs of eyes possess two optical commissures (Oenonidae, Dorvilleidae)—an anteriorly located one (aoc) and a posteriorly located one (poc). In Dorvillea spec. only the insertion points of the optical nerves could be observed and not the complete commissure due to an oblique cutting plane. The presence of an optical commissure was reported by Zanol [29] for Dorvilleidae only. The optical commissures are located dorsally in the brain and anteriorly to the dorsal commissure of the dorsal root of the circumoesophageal connective (dcdr) in all species investigated, which concurs with the findings of Orrhage and Müller [13].
4.2.2. Nuchal Commissure
A commissure of the nuchal organ nerves as Orrhage and Müller [13] and Zanol [29] reported, was not observed in any eunicidan specimen examined. The nuchal organ is innervated by nerves originating from the posterior-most part of the dorsal brain neuropil as typical for other annelids [13,29,39,40].
4.2.3. Association Tracts
Association tracts were found in species bearing five head appendages (Eunicidae, Onuphidae), while in species lacking head appendages (Oenonidae, Lumbrineridae) association tracts are not present. Interestingly, we did not observe any association tracts in Dorvilleidae (four head appendages). Together with the fact, that further studies on association tracts in Dorvilleidae are lacking, it could be assumed that association tracts might be an autapomorphy of Eunicoidea. Binard and Jeener [26] and Zanol [29] for example reported association systems (sensu Orrhage [14]) for Eunicidae and Onuphidae only—in Dorvilleidae, Zanol [29] did not find an association system. The association system is an anteriorly located structure composed of small cells [29] connected to association tracts. So, the association system seems to be involved in the processing of sensory stimuli transmitted by association tracts and in a further sense by head appendages. In our study, we observed anteriorly located areas of circularly arranged neurite bundles (atmla), in which the association tracts of the median and the lateral antennae anastomose. These areas might be association tracts on their own. If these areas concur with the association system sensu Orrhage [14] is a matter of debate—e.g., we did not observe a composition of small cells. At least in topology and the connection to association tracts both structures concur—presumably also in function as processors of sensory stimuli received by head appendages. We found the atmla only in species of Eunicoidea with five head appendages. Species with less than five head appendages belonging to Eunicidae (Lysidice unicornis, Lysidice ninetta) show neither a distinct atmla nor distinct association tracts in the histological sections. However, in L. ninetta one possible association tract was observed (one of the minor nerve roots of the lateral root of the lateral antennae nerve). Whether this observation consolidates a general difference in microanatomical processing of sensory stimuli received by head appendages between Eunicoidea and Dorvilleidae is a matter of debate. Clearly stated can be, that innervation patterns are more complex in species bearing five head appendages (Eunicoidea).
The number of association tracts found among the different species in this study is mostly in accordance with the pertinent literature [14]: In Eunicoidea one pair of association tracts is connected to the median antenna and in Onuphidae one pair of association tracts is linked to the lateral antennae, respectively. For the lateral antennae of Eunicidae, we found—contrary to the report of Orrhage [14] of one pair of association tracts—two pairs of association tracts. The additional lateral association tracts seem to anastomose with the ventrally located commissure (vc) in the dorsal brain neuropil of Eunicidae, resembling the dorsal fibril mass (dfm) of the schematic drawing of Orrhage [14] of H. tubicola. The other pair of lateral association tracts fuse with the pair of median association tracts, more or less similar to the drawing of Orrhage [14].
The presence of palps does not affect brain anatomy in Eunicida: no special brain compartments, commissures or tracts associated with palps were observable inside the brain neuropil. Sensory information gained by palps seems to be processed somewhere in the drcc and the vrcc—the commissures the palp nerves originate from. In contrast to conditions found in Polynoida and Nereididae [12,41,42,43], where each palp nerve is associated with mushroom body neuropil, palp nerves are not associated with mushroom bodies in Eunicida.
4.2.4. Lateral Neuropils
Another characteristic shared by the studied representatives of Oenonoidea is the paired lateral neuropils, which are lacking in Eunicoidea and Dorvilleidae. The lateral neuropils may further support the clade Oenonoidea. The lateral neuropils are interconnected via a tract in the brain neuropil, and in A. iricolor, the dorsal nerves originate from this tract. Further neuronal connections revealing the function of the lateral neuropils were not observed. A pair of lateral neuropils (called “dorso-ventral neuropil”) anteriorly in the brain of Eunicidae, Dorvilleidae, Oenonidae and Lumbrineridae (except for Onuphidae) was also observed by Zanol [29]. This structure seems not to match the neuropils we observed in Oenonidae and Lumbrineridae, since Zanol [29] described these neuropils as connecting the dorsal and ventral neuropils via several connections (dorsal fibril mass, dorsal commissure of the ventral root of the circumoesophageal connective, first ventral commissure of the ventral root of the circumoesophageal connective). Neither we observed lateral neuropils in other taxa than Oenonidae and Lumbrineridae nor we saw it connecting dorsal and ventral neuropil via the given connections. However, the lateral neuropils as well as Zanol’s dorso-ventral neuropils resemble polychaete mushroom bodies partially in shape and composition. Already Hanström [12] compared the small and chromatin-rich “Ganglienzellen” occurring in high amounts in the anterior region of the brains of Eunice norvegica (Linnaeus, 1767), Hyalinoecia tubicola and Diopatra cuprea (Bosc, 1802), with the cells of the corpora pedunculata. The presence of mushroom body-like structures in polychaete taxa was discussed already in the former literature [12,34,43] and it was pointed out, that mushroom body-like structures may be present in Eunicida [12]. However, the current literature (e.g., [1]) states that mushroom bodies are not present in Eunicida referring to the study of Bullock and Horridge [44]. The mushroom body-like structures in Eunicida might be remnants of mushroom bodies if a reduction is hypothesized. Since Phyllodocida, e.g., Nereis diversicolor, possess distinct mushroom bodies [42,43], a secondary reduction of mushroom bodies in Eunicida is not unlikely.
Regarding the presence of prominent neuropil areas such as the dorso-ventral neuropil and the lateral neuropils of Oenonidae and Lumbrineridae, the question arises, if these neuropils serve as compensators for lacking senses, e.g., of antennae, palps, eyes etc. Studies on this topic were conducted by von Haffner in 1962 [11]. His studies are based on a topographical tripartition of the brain using sensory organs as markers for the different areas (fore-brain = “palps” (=nowadays buccal lips), mid-brain = antennae and eyes, hind-brain = nuchal organ). Von Haffner [11] compared the size of the brain areas among Eunicida and proposed, that the extended hind-brain is associated with the extended nuchal organs in Oenonidae and Lumbrineridae. He deduced that the extension of both structures compensates for the missing sensory input of antennae and palps. Although we clearly saw shape differences and a generally bigger surface of the nuchal cavity in Oenonoidea when compared to the remaining Eunicida, we did not observe an extended posterior brain area (=hind-brain) in these taxa. Since the brain appeared as a continuous mass of neuropil in our study, we did not separate the brain according to Racovitza [45]. The only differentiation we applied was the distinction between a dorsal and a ventral neuropil. Both neuropils have equal sizes among the different species studied regardless of the number of prostomial sensory organs. Together with the fact, that we did not observe tracts connecting the nuchal organs with these neuropils, there is no reason to assume a compensational function of both neuropils in this aspect.
According to Zanol [29], shape differences in the brain occur among the families of Eunicida—especially in Onuphidae. She reports a different topology of the intracerebral cavity in Onuphidae (placed between the vcvr1 + vcvr2 and the vcvr3 + vcvr2) when compared to Eunicidae, Dorvilleidae, Oenonidae and Lumbrineridae (placed between the dcvr and the vcvr1). Furthermore, she observed an x-shaped connection in Onuphidae (between the vcvr1 + vcvr2 and the dcvr), which is lacking in the other families. Since our study focussed methodically on paraffin histology, the view we had on the brain differs from that of Zanol’s immunohistochemical studies, e.g., we did not clearly see all commissures of the roots of the circumoesophageal connectives and with this, we did not see different topologies of the intracerebral cavity or the x-shaped connection. Further, we did not recognize shape differences in the brains of Onuphidae and Eunicidae, but it must be considered, that we studied one adult specimen of Onuphidae (Hyalinoecia tubicola). Zanol [29] studied Onuphis (Nothria) iridescens (Johnson, 1901) for ventral dissection and sagittal cuts, and juveniles of Kinbergonuphis simoni (Santos, Day and Rice, 1981), so that species-specific differences or age effects can not be excluded. So, again brain morphology supports the clades Eunicoidea on the one-hand side and Oenonoidea on the other. The general brain anatomy of Dorvilleidae seems to be identical to Eunicidae and Onuphidae.
The general brain morphology seems not to be affected by the number of prostomial sensory organs because although differences in the shape of the brain might occur among the different families, they do not seem to be related to the number of prostomial sensory organs.
4.3. Phylogenetic Significance of Nerve Roots
Equal numbers and positions of nerve roots among the different families suggest a close relationship between Eunicidae and Onuphidae. The number of nerve roots is the same for the median antenna (4 main nerve roots) [13] as well as the palps (3 main nerve roots) in these taxa. In our study, the number of nerve roots of the lateral antennae differs among the families: while four nerve roots are present in Eunicidae, only two nerve roots were observed in Hyalinoecia tubicola. If this number is applicable to the complete clade of Onuphidae has to be evaluated by studying more specimens. It could be hypothesized that age effects may play a role, so that definitive innervation patterns may become apparent in the adult stage only. For example, Zanol [29] reported that juveniles of Eunicidae lack adult characters such as palps and lateral antennae and it could be suggested, that their innervation is not fully developed yet. Still, the specimen of H. tubicola we investigated seems to have already reached the adult stage as the remaining species studied. As the examined representatives of Onuphidae, Dorvilleidae lack minor nerves in the lateral antennae, so here also only two main nerve roots are present [29].
The palps of the examined Dorvilleidae differ in inner morphology as well as innervation from palps of other taxa as well as antennae of the same taxon. Dorvilleid palps are innervated by two distinct main nerves and possess a coelomic cavity and musculature extending from the palpophore to nearly the tip of the palpostyle. Whether this construction has a special function can only be speculated. Together with the exceptional innervation pattern, the differing inner palp morphology may support the phylogenetic placement of Dorvilleidae as a clade outside of Eunicoidea and Oenonoidea as proposed by Tilic et al. [2]. Whether Dorvilleidae forms a clade together with Histriobdellidae has still to be evaluated. According to Orrhage and Müller [13] and Purschke et al. [37], up to twelve different palp nerve roots can be present in Annelida, but until now no taxon studied possessed all twelve palp nerve roots [13,37]. The palp nerve roots we observed are the two thick main palp nerve roots originating from the drcc and the vrcc and some minor nerve roots (see Purschke et al. [37]).
4.4. Ecological Significance of Sensory Organs in Eunicidae
Antennae and palps are both presumably used to recognize prey in Eunicida. Eunice aphroditois (Pallas, 1788), commonly referred to as bobbit worm, for example, is an ambush predator and hides in the sediment—only its frontal part reaching out into the water column [46]. If potential prey gets accidentally in contact with the head appendages, the worm will capture it with its jaws. In endobenthic forms such as Oenonoidea head appendages and eyes are reduced in most species, which concurs with their lifestyle: in an endobenthic environment, the sensory function of head appendages, as well as optical senses, are of no great use. Generally, reductions are common in taxa-shifting lifestyles from epi- to endobenthic [47]. Small, interstitial eunicids as commonly found in Dorvilleidae (e.g., Neotenotrocha) may have reduced head appendages as well as innervation patterns due to progenesis [6,48]. Extreme differences in body sizes can frequently be observed in Eunicida, which are accompanied by different feeding modes and lifestyles: Small interstitial dorvilleids (1 mm) for example are microphagous “grazing” on algae and biofilms with their jaws (comparable to the radula of gastropods), while larger representatives, such as Eunice aphroditois (3 m), are macrophagous and predatory [49].
In the predatory Oenonoidea, the nuchal organ has a crucial function in detecting prey in this taxon. Von Haffner [11] hypothesised that the nuchal organ balances the missing sensory input of head appendages and eyes. In his studies, he theoretically separated the brain of Eunicida into different parts: a fore-brain, the mid-brain and a hind-brain. Based on this concept, he inferred, that the hind-brain in Lysidice ninetta and Nematonereis unicornis is bigger than in species bearing more head appendages. For Scoletoma (Lumbrineris) fragilis, he reported more prominent nuchal ganglia. We did not observe an extended posterior brain part or prominent nuchal commissures in any specimen.
While the surface of the nuchal cavity seems to be enlarged through foldings (x-shape) in the studied specimens of Oenonidae and Lumbrineridae, the receptive, ciliated surface is not enlarged when compared to the arc-shaped nuchal organs of Eunicoidea. Considering that in the examined specimens of Eunicoidea and Oenonoidea overall two ciliary patches of similar size are present, it could be assumed, that sensitivity of the nuchal organs is similar among these species. In contrast to this, the nuchal organs of Dorvillea spec. contain overall four ciliary patches—it shows a duplication of the common two ciliary patches. Here, the ventrally, laterally and dorsally located openings of the nuchal cavities may direct the water current directly onto the respective ciliary patch. This might enable Dorvillea spec. to detect the direction of, e.g., chemical cues ([39] for Saccocirrus species).
Lateral organs are present in every specimen of Eunicida and are regarded as characteristics belonging to the eunicidan ground pattern [4,37,50,51]. Generally, lateral organs are described as ciliated pits or papillae located between the noto- and the neuropodium in polychaetous annelids [4,52]. In our study, all lateral organs are based on the ventral side of the dorsal cirrus of the notopodium or an equivalent position in taxa lacking a notopodium, e.g., Lumbrineridae and Histriobdellidae. Additional differences in morphology of lateral organs occur in Hyalinoecia tubicola (Onuphidae) and Arabella iricolor (Oenonidae): here, the lateral organs are not only covered by cilia but also by microvilli. Moreover, the cilia of the lateral organ of Ophryotrocha siberti are longer than the ones in the remaining specimens studied. These characters support the hypothesis of species-specific morphologies of lateral organs in Eunicida. Hayashi and Yamane [53] reported differences in the histology of lateral organs of Marphysa sanguinea (Montagu, 1813) (Eunicidae) and Scoletoma (Lumbrineris) longifolia (Imajima and Higuchi, 1975) (Lumbrineridae) suggesting these to be species-specific.
4.5. High Metabolic Rate of the Brain
Regardless of the different lifestyles (endobenthic, epibenthic), the brain of the studied eunicidans exhibits the same indicators for a high metabolic rate. Blood vessels traverse the brain frequently indicating a high metabolic activity as typical for errant annelids. Moreover, three different kinds of neuronal somata surround the brain of the different species studied. Type one neurons are most abundant as typical for annelids [54]. Somata of type two and type three neurons generally contain densely arranged endoplasmatic reticula indicating a high metabolic rate [54]. Further, the high abundance of dense and lucent core vesicles in the brain neuropil indicates a high synaptic activity [34]. A high metabolic rate is a prerequisite for processing diverse sensory stimuli and for a predatory lifestyle. In all species examined these indicators (blood vessels, three types of neuronal somata, endoplasmatic reticula, dense and lucent core vesicles) can be found in the same abundancy and arrangement regardless of the number of prostomial sensory organs or lifestyle, which hint at a conserved status of these parameters.
4.6. Answers to Initial Questions
- Is the brain morphology affected by different numbers and shapes of prostomial sensory organs? The gross brain morphology is not affected by the number and shape of sensory organs, but the microanatomy may be affected in terms of commissures and tracts (e.g., association tracts, optical commissures).
- How can head appendages (antennae and palps) as well as buccal lips be differentiated? Differentiation is performed via innervation patterns: Antennae are solely innervated by nerves emanating from the drcc [13,14]. Palps are innervated by two or more nerve roots originating from the dorsal root of the circumoesophageal connective (drcc) and the ventral root of the circumoesophageal connective (vrcc) and their commissures [13]. Buccal lips are innervated solely by the ventral brain neuropil.
5. Conclusions
The gross morphology of the central nervous system is not influenced by the number of prostomial sensory organs and seems to be conserved among Eunicida when comparing only major components. When taking a closer look at more detailed morphologies, plasticity appears consolidating in apparent similarities between the studied specimens of Eunicidae + Onuphidae on the one-hand side and Oenonidae + Lumbrineridae on the other-hand side. Prostomial sensory organs shape the microanatomy of the brain in the studied representatives of Eunicidae and Onuphidae in a similar manner in terms of commissures and tracts. Further, the prostomial sensory organs such as antennae, palps, buccal lips, eyes and especially nuchal organs of both taxa occur in characteristic numbers and morphologies. In contrast to this, the studied representatives of Oenonidae and Lumbrineridae lack most of these prostomial sensory organs and the related commissures. Still, nuchal organs are present and based on a similar morphology, they could hint at a close relationship between Oenonidae and Lumbrineridae. Further, the presence of lateral neuropil areas and the dorso-ventrally extended brain shape could be used as further indicators for this relationship. When extrapolating the data gained in this study from species level to family rank, our data concur with the phylogeny of Eunicida proposing a sister group relationship of Eunicidae + Onuphidae (Eunicoidea) and Oenonidae + Lumbrineridae (Oenonoidea) [2]. In this phylogeny, Dorvilleidae builds a clade outside of Eunicoidae and Oenonoidea, which can be supported by the results gained in this study. If Dorvilleidae forms a clade together with Histriobdellidae has still to be evaluated. The dorsal nerves of Arabella iricolor could be homologized with nerves innervating antennae in Eunicidae and Onuphidae and—supported by equivalent observations in Dorvilleidae by Zanol [29]—it could be hypothesized that a reduction of antennae might have taken place.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jmse10111707/s1, Table S1. Specimens used in this study, sampling locality and methods applied. Overall 13 specimens of Eunicidae, 3 specimens of Onuphidae, 5 specimens of Dorvilleidae, 2 specimens of Oenonidae, 4 specimens of Lumbrineridae and 2 specimens of Histriobdellidae were used. cLSM: confocal laser scanning microscopy, LM: light microscopy, TEM: transmission electron microscopy, SEM: scanning electron microscopy; Table S2. Terminology of head appendages and buccal lips over time; Table S3. Links for downloading complete image stacks of the examined specimens.
Author Contributions
P.B. conceived the study. S.K. acquired and analyzed all data and wrote the manuscript. P.B. and T.B. reviewed and edited the draft. All authors have read and agreed to the published version of the manuscript.
Funding
The µCT-scanner was funded by the State of North Rhine-Westphalia and the Deutsche Forschungsgemeinschaft (INST 217/849-1 FUGG). The FEI Verios 460 L scanning electron microscope was funded by the State of North Rhine-Westphalia and the Deutsche Forschungsgemeinschaft (INST 217/784-1 FUGG).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
To facilitate future comparative analyses and to maintain data transparency, all image series of paraffin histology (AZAN + silver) are deposited in https://zenodo.org, accessed on 5 October 2022 (see Supplementary Material, Table S3). Complete image stacks can be downloaded using Zenodo_get (Völgyes and Lupton 2020). Physical sections are deposited in the histology collection of the Institute of Evolutionary Biology and Ecology of the University of Bonn as vouchers.
Acknowledgments
The authors thank the Station Marine de Concarneau for their support in collecting some of the examined animals. The authors thank Ekin Tilic for providing specimens of Arabella iricolor and Ophryotrocha siberti, and Conrad Helm for gently providing a specimen of Hyalinoecia tubicola. Further, we would like to thank Alexander Ziegler for his support with the µCT-scanner. We are very grateful to Gregor Kirfel from the Institute for Cell Biology in Bonn for his support with the FEI Verios 460 L scanning electron microscope.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rouse, G.W.; Pleijel, F.; Tilic, E. Annelida; Oxford University Press: Oxford, UK, 2022; pp. 103–125. [Google Scholar]
- Tilic, E.; Stiller, J.; Campos, E.; Pleijel, F.; Rouse, G.W. Phylogenomics resolves ambiguous relationships within Aciculata (Errantia, Annelida). Mol. Phylogenetics Evol. 2022, 166, 107339. [Google Scholar] [CrossRef]
- von Palubitzki, T.; Purschke, G. Ultrastructure of pigmented eyes in Onuphidae and Eunicidae (Annelida: Errantia: Eunicida) and its importance in understanding the evolution of eyes in Annelida. Zoomorphology 2020, 139, 1–19. [Google Scholar] [CrossRef]
- Rouse, G.; Pleijel, F. Polychaetes; Oxford University Press: Oxford, UK, 2001. [Google Scholar] [CrossRef]
- Ehlers, E. Die Borstenwürmer Nach Systematischen und Anatomischen Untersuchungen Dargestellt; Wilhelm Engelmann: Leipzig, Germany, 1868. [Google Scholar]
- Struck, T.H.; Purschke, G.; Halanych, K.M. Phylogeny of Eunicida (Annelida) and exploring data congruence using a partition addition bootstrap alteration (PABA) approach. Syst. Biol. 2006, 55, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; Carrera-Parra, L.F.; Steiner, T.M.; Amaral, A.C.Z.; Wiklund, H.; Ravara, A.; Budaeva, N. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity 2021, 13, 74. [Google Scholar] [CrossRef]
- Paxton, H. Phylogeny of Eunicida (Annelida) based on morphology of jaws. Zoosymposia 2009, 2, 241–264. [Google Scholar] [CrossRef] [Green Version]
- Kielan-Jaworowska, Z. Scolecodonts versus jaw apparatuses. Lethaia 1968, 1, 39–49. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Polychaeta, 2nd ed.; Fischer: Jena, Germany, 1996. [Google Scholar]
- von Haffner, K. Über die Abhängigkeit des Gehirnbaues von den Sinnesorganen: Vergleichende Untersuchungen an Euniciden (Polychaeta). Zool. Jb. Anat 1962, 80, 159–212. [Google Scholar]
- Hanström, B. Das zentrale und periphere Nervensystem des Kopflappens einiger Polychäten. Z. Morphol. Ökologie Tiere 1927, 7, 543–596. [Google Scholar] [CrossRef]
- Orrhage, L.; Müller, M.C.M. Morphology of the nervous system of Polychaeta (Annelida). Hydrobiologia 2005, 535–536, 79–111. [Google Scholar] [CrossRef]
- Orrhage, L. On the Innervation and Homologues of the Anterior End Appendages of the Eunicea (Polychaeta), with a Tentative Outline of the Fundamental Constitution of the Cephalic Nervous System of the Polychaetes. Acta Zool. 1995, 76, 229–248. [Google Scholar] [CrossRef]
- Hessling, R.; Purschke, G. Immunohistochemical (cLSM) and ultrastructural analysis of the central nervous system and sense organs in Aeolosoma hemprichi (Annelida, Aeolosomatidae). Zoomorphology 2000, 120, 65–78. [Google Scholar] [CrossRef]
- Beckers, P.; Helm, C.; Bartolomaeus, T. The anatomy and development of the nervous system in Magelonidae (Annelida)—Insights into the evolution of the annelid brain. BMC Evol. Biol. 2019, 19, 173. [Google Scholar] [CrossRef] [Green Version]
- Monro, C.C.A. Polychaete worms. In Discovery Reports; University Press: Cambridge, UK, 1931; Volume II, pp. 3–218. [Google Scholar]
- Aiyar, R.G. On the Anatomy of Marphysa Gravelyi Southern. Rec. Zool. Surv. India 1933, 35, 287–323. [Google Scholar] [CrossRef]
- Fauchald, K. The Polychaete Worms: Definitions and Keys to the Orders, Families and Genera; Natural History Museum of Los Angeles County, Science Series: Los Angeles, CA, USA, 1977. [Google Scholar]
- Fauchald, K.; Rouse, G.W. Polychaete systematics: Past and present. Zool. Scr. 1997, 26, 71–138. [Google Scholar] [CrossRef]
- Heider, K. Über Eunice. Systematisches, Kiefersack, Nervensystem. Z. Für Wiss. Zool. 1925, 125, 55–90. [Google Scholar]
- von Haffner, K. Über den Bau und den Zusammenhang der Wichtigsten Organe des Kopfendes von Hyalinoecia Tubicola Malmgren (Polychaeta, Eunicidae, Onuphiidae) mit Berücksichtigung der Gattung Eunice. Zool. Jahrbücher Abt. Anat. Ontog. Tiere 1959, 77, 113–192. [Google Scholar]
- von Haffner, K. Untersuchungen und Gedanken über das Gehirn und Kopfende von Polychaeten und ein Vergleich mit den Arthropoden. Mitt. Zool. Staatsinst. Zool. Mus. Hambg. 1959, 37, 1–29. [Google Scholar]
- Paxton, H. Generic revision and relationships of the family Onuphidae (Annelida: Polychaeta). Rec. Aust. Mus. 1986, 38, 1–74. [Google Scholar] [CrossRef] [Green Version]
- Orensanz, J.M. The eunicemorph polychaete annelids from Antarctic and Subantarctic seas: With addenda to the eunicemorpha of Argentina, Chile, New Zealand, Australia, and the southern Indian Ocean. Biol. Antarct. Seas XXI 1990, 52, 1–183. [Google Scholar] [CrossRef]
- Binard, A.; Jeener, R. Morphologie du lobe préoral des polychètes. Recl. De L‘Inst. Zool. Torley-Rousseau 1928, 2, 177–240. [Google Scholar]
- Gustafson, G. Anatomische Studien über die Polychaeten-Familien Amphinomidae und Euphrosynidae. Ph.D. Thesis, Almqvist & Wiksell, Stockholm, Sweden, 1930. [Google Scholar]
- Åkesson, B. The Embryology of the Polychaete Eunice kobiensis. Acta Zool. 1967, 48, 142–192. [Google Scholar] [CrossRef]
- Zanol, J. Homology of prostomial and pharyngeal structures in eunicida (Annelida) based on innervation and morphological similarities. J. Morphol. 2010, 271, 1023–1043. [Google Scholar] [CrossRef] [PubMed]
- Pruvot, G.; Racovitza, E.-G. Matériaux pour la faune des Annélides de Banyuls. Arch. Zool. Expérimentale Générale 1895, 3, 339–492. [Google Scholar]
- Fauvel, P. Faune de France: 5 Polychètes Errantes. Fédération Française des Sociétés de Sciences Naturelles: Paris, France, 1923; pp. 4–491. [Google Scholar]
- Müller, M.C.M. Polychaete nervous systems: Ground pattern and variations—cLS microscopy and the importance of novel characteristics in phylogenetic analysis. Integr. Comp. Biol. 2006, 46, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckers, P.; Helm, C.; Purschke, G.; Worsaae, K.; Hutchings, P.; Bartolomaeus, T. The central nervous system of Oweniidae (Annelida) and its implications for the structure of the ancestral annelid brain. Front. Zool. 2019, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Beckers, P.; Pein, C.; Bartolomaeus, T. Fine structure of mushroom bodies and the brain in Sthenelais boa (Phyllodocida, Annelida). Zoomorphology 2021, 3, 579. [Google Scholar] [CrossRef]
- Beckers, P.; Müller, C.; Wallnisch, C.; Bartolomaeus, T. Getting two birds with one stone: Combining immunohistochemistry and Azan staining in animal morphology. J. Biol. Methods 2021, 8, e153. [Google Scholar] [CrossRef]
- Richter, S.; Loesel, R.; Purschke, G.; Schmidt-Rhaesa, A.; Scholtz, G.; Stach, T.; Vogt, L.; Wanninger, A.; Brenneis, G.; Döring, C.; et al. Invertebrate neurophylogeny: Suggested terms and definitions for a neuroanatomical glossary. Front. Zool. 2010, 7, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Purschke, G.; Bleidorn, C.; Struck, T. Systematics, evolution and phylogeny of Annelida—A morphological perspective. Mem. Mus. Vic. 2014, 71, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Malaquin, A. Recherches Sur Les Syllidiens: Morphologie, Anatomie, Reproduction, Développement. Danel, L., Ed.; Lille, France, 1893; pp. 3–477. Available online: https://www.biodiversitylibrary.org/bibliography/99327 (accessed on 5 October 2022).
- Purschke, G. Ultrastructure of Nuchal Organs in Polychaetes (Annelida)—New Results and Review. Acta Zool. 1997, 78, 123–143. [Google Scholar] [CrossRef]
- Ax, P. Multicellular Animals: The Phylogenetic System of the Metazoa; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2000. [Google Scholar]
- Strausfeld, N.J.; Hansen, L.; Li, Y.; Gomez, R.S.; Ito, K. Evolution, Discovery, and Interpretations of Arthropod Mushroom Bodies. Learn. Mem. 1998, 5, 11–37. [Google Scholar] [CrossRef]
- Loesel, R.; Heuer, C.M. The mushroom bodies—Prominent brain centres of arthropods and annelids with enigmatic evolutionary origin. Acta Zool. 2010, 91, 29–34. [Google Scholar] [CrossRef]
- Heuer, C.M.; Müller, C.H.; Todt, C.; Loesel, R. Comparative neuroanatomy suggests repeated reduction of neuroarchitectural complexity in Annelida. Front. Zool. 2010, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullock & Horridge. Structure and Function of the Nervous System of Invertebrates; Freeman: San Francisco, CA, USA, 1965. [Google Scholar]
- Racovitza, E.G. Le lobe céphalique et l’encéphale des Annélides Polychètes (Anatomie, Morphologie, Histologie). Arch. Zool. Expérimentale Générale 1896, 3, 133–343. [Google Scholar]
- Huisman, J.; Dixon, R. Northern exposure: Marine life of the Kimberley. Landscope 2011, 26, 10–16. [Google Scholar]
- Purschke, G.; Nowak, K.H. Ultrastructure of pigmented eyes in Dorvilleidae (Annelida, Errantia, Eunicida) and their importance for understanding the evolution of eyes in polychaetes. Acta Zool. 2015, 96, 67–81. [Google Scholar] [CrossRef]
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, A.; Westheide, W.; Purschke, G.; Bleidorn, C.; Halanych, K.M. The evolution of annelids reveals two adaptive routes to the interstitial realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef]
- Purschke, G.; Tzetlin, A.B. Dorsolateral ciliary folds in the polychaete foregut: Structure, prevalence and phylogenetic significance. Acta Zool. 1996, 77, 33–49. [Google Scholar] [CrossRef]
- Beesley, P.L.; Ross, G.J.B.; Glasby, C.J.; Study, A.B.R. Polychaetes & Allies: The Southern Synthesis; CSIRO Publishing: Clayton, Australia, 2000. [Google Scholar]
- Budaeva, N.; Schepetov, D.; Zanol, J.; Neretina, T.; Willassen, E. When molecules support morphology: Phylogenetic reconstruction of the family Onuphidae (Eunicida, Annelida) based on 16S rDNA and 18S rDNA. Mol. Phylogenetics Evol. 2016, 94, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Purschke, G. Sense organs in polychaetes (Annelida). Hydrobiologia 2005, 535–536, 53–78. [Google Scholar] [CrossRef]
- Hayashi, I.; Yamane, S. Further Observation of a Recently Found Sense Organ in Some Euniciforms, with Special Reference to Lumbrineris Longifolia (Polychaeta, Lumbrineridae). Bull. Mar. Sci. 1997, 60, 564–574. [Google Scholar]
- Beckers, P.; Tilic, E. Fine structure of the brain in Amphinomida (Annelida). Acta Zool. 2021, 11, 2860. [Google Scholar] [CrossRef]
Figure 1.
Variation of head appendages (antennae, palps) and eyes in representatives of Eunicida. Living specimens (A–D,F–H), fixated specimens (E). (A) Paucibranchia bellii, dorsal view. Five head appendages—one median antenna (ma), one pair of lateral antennae (la) and one pair of palps (pa) are present. One pair of small red-pigmented eyes (ey) is located at the lateral basis of the lateral antennae. (B) Lysidice unicornis, lateral view. One median antenna (ma) and one pair of red-pigmented eyes (ey) are located on the prostomium. (C) Lysidice ninetta, dorsal view. Three head appendages are present (1 ma,1 pair of la) and one pair of relatively big red-pigmented eyes is located on the ventral basis of the lateral antennae. (D) Leodice torquata, dorsal view. Five head appendages are present (1 ma, 1 pair of la, 1 pair of pa). One pair of blackish-purple eyes (ey) is located on the ventral basis of the lateral antennae. (E) Hyalinoecia tubicola, lateral view. Five relatively long head appendages are present (1 ma, 1 pair of la, 1 pair of pa). One pair of black-pigmented eyes (ey) is located at the basis of the lateral antennae. The specimen was already fixed when the picture was taken—deviation in colouration of the specimen is possible. (F) Dorvillea spec., dorsal view. Four relatively long head appendages are present (1 pair of la, 1 pair of pa). Two pairs of red-pigmented eyes (ey) are located on the posterior edge of the prostomium. (G) Arabella iricolor, dorsal view. Head appendages are lacking, but two pairs of black-pigmented eyes (ey) are present on the prostomium. (H) Scoletoma tetraura, dorsal view. Lacks head appendages as well as eyes. ey = eye, la = lateral antenna, ma = median antenna, pa = palp, pe = peristomium, pr = prostomium.
Figure 1.
Variation of head appendages (antennae, palps) and eyes in representatives of Eunicida. Living specimens (A–D,F–H), fixated specimens (E). (A) Paucibranchia bellii, dorsal view. Five head appendages—one median antenna (ma), one pair of lateral antennae (la) and one pair of palps (pa) are present. One pair of small red-pigmented eyes (ey) is located at the lateral basis of the lateral antennae. (B) Lysidice unicornis, lateral view. One median antenna (ma) and one pair of red-pigmented eyes (ey) are located on the prostomium. (C) Lysidice ninetta, dorsal view. Three head appendages are present (1 ma,1 pair of la) and one pair of relatively big red-pigmented eyes is located on the ventral basis of the lateral antennae. (D) Leodice torquata, dorsal view. Five head appendages are present (1 ma, 1 pair of la, 1 pair of pa). One pair of blackish-purple eyes (ey) is located on the ventral basis of the lateral antennae. (E) Hyalinoecia tubicola, lateral view. Five relatively long head appendages are present (1 ma, 1 pair of la, 1 pair of pa). One pair of black-pigmented eyes (ey) is located at the basis of the lateral antennae. The specimen was already fixed when the picture was taken—deviation in colouration of the specimen is possible. (F) Dorvillea spec., dorsal view. Four relatively long head appendages are present (1 pair of la, 1 pair of pa). Two pairs of red-pigmented eyes (ey) are located on the posterior edge of the prostomium. (G) Arabella iricolor, dorsal view. Head appendages are lacking, but two pairs of black-pigmented eyes (ey) are present on the prostomium. (H) Scoletoma tetraura, dorsal view. Lacks head appendages as well as eyes. ey = eye, la = lateral antenna, ma = median antenna, pa = palp, pe = peristomium, pr = prostomium.

Figure 2.
Components and topology of the cns in Eunicida. The 3D reconstructions and volume renderings are based on µCT scans (A–F), cLSM scans of sagittal sections (G,H). (A) Leodice torquata, dorsal view. (B) L. torquata, lateral view. (C) Lysidice ninetta, dorsal view. (D) L. ninetta, lateral view. (E) Hyalinoecia tubicola, dorsal view. (F) H. tubicola, lateral view. (G) Scoletoma tetraura, lateral view. A fine net of frontal nerves (fn) originating from the brain neuropil innervates the anterior part of the prostomium. The dorsal and the ventral brain neuropil (dnp, vnp) can be distinguished. Nuchal organ nerves (non) originate from the posterior-most part of the dorsal brain neuropil (dnp) and innervate the nuchal organ (indicated by green arrowhead). (H) S. tetraura, lateral view. From the ventral brain neuropil, the oesophageal nerves (on) originate at height of the mouth opening (purple arrowhead). br: brain; ca: caudal; cc: circumoesophageal connective; cns: central nervous system; dnp: dorsal brain neuropil; ey: eye; fn: frontal nerves; fr: frontal; jn: jaw nerves; ma: median antenna; mo: mouth opening; no: nuchal organ; non: nuchal organ nerve; on: oesophageal nerves; pe: peristomium; pr: prostomium; vnc: ventral nerve chord; vnp: ventral brain neuropil; green arrow: nuchal cavity; pink arrow: mouth opening.
Figure 2.
Components and topology of the cns in Eunicida. The 3D reconstructions and volume renderings are based on µCT scans (A–F), cLSM scans of sagittal sections (G,H). (A) Leodice torquata, dorsal view. (B) L. torquata, lateral view. (C) Lysidice ninetta, dorsal view. (D) L. ninetta, lateral view. (E) Hyalinoecia tubicola, dorsal view. (F) H. tubicola, lateral view. (G) Scoletoma tetraura, lateral view. A fine net of frontal nerves (fn) originating from the brain neuropil innervates the anterior part of the prostomium. The dorsal and the ventral brain neuropil (dnp, vnp) can be distinguished. Nuchal organ nerves (non) originate from the posterior-most part of the dorsal brain neuropil (dnp) and innervate the nuchal organ (indicated by green arrowhead). (H) S. tetraura, lateral view. From the ventral brain neuropil, the oesophageal nerves (on) originate at height of the mouth opening (purple arrowhead). br: brain; ca: caudal; cc: circumoesophageal connective; cns: central nervous system; dnp: dorsal brain neuropil; ey: eye; fn: frontal nerves; fr: frontal; jn: jaw nerves; ma: median antenna; mo: mouth opening; no: nuchal organ; non: nuchal organ nerve; on: oesophageal nerves; pe: peristomium; pr: prostomium; vnc: ventral nerve chord; vnp: ventral brain neuropil; green arrow: nuchal cavity; pink arrow: mouth opening.

Figure 3.
General brain anatomy. Histological cross sections (5 µm, AZAN) of Leodice torquata (A,B) and Arabella iricolor (F,G). Three-dimensional reconstructions based on histological serial sections of Lysidice ninetta (C–E) and A. iricolor (H–J). at: association tract; atmla: association tracts of the median and the lateral antennae; bc: buccal cavity; ca: caudal; cc: circumoesophageal connective; d: dorsal; dcvr: dorsal commissure of the ventral root of the circumoesophageal connective; dn: dorsal nerves; dnp: dorsal brain neuropil; drcc: dorsal root of the circumoesophageal connective; ey: eye; fr: frontal; ic: intracerebral commissure; if: intermediate filaments; jn: jaw nerves; lan: lateral antenna nerve; man: median antenna nerve; no: nuchal organ; oc: optical commissure; oegc: connective of the oesophageal ganglion; oen: oesophageal nerves; on: optical nerve; stn: stomatogastric nerves v: ventral; vcvr: ventral commissure of the ventral root of the circumoesophageal connective; vnp: ventral brain neuropil; vrcc: ventral root of the circumoesophageal connective.
Figure 3.
General brain anatomy. Histological cross sections (5 µm, AZAN) of Leodice torquata (A,B) and Arabella iricolor (F,G). Three-dimensional reconstructions based on histological serial sections of Lysidice ninetta (C–E) and A. iricolor (H–J). at: association tract; atmla: association tracts of the median and the lateral antennae; bc: buccal cavity; ca: caudal; cc: circumoesophageal connective; d: dorsal; dcvr: dorsal commissure of the ventral root of the circumoesophageal connective; dn: dorsal nerves; dnp: dorsal brain neuropil; drcc: dorsal root of the circumoesophageal connective; ey: eye; fr: frontal; ic: intracerebral commissure; if: intermediate filaments; jn: jaw nerves; lan: lateral antenna nerve; man: median antenna nerve; no: nuchal organ; oc: optical commissure; oegc: connective of the oesophageal ganglion; oen: oesophageal nerves; on: optical nerve; stn: stomatogastric nerves v: ventral; vcvr: ventral commissure of the ventral root of the circumoesophageal connective; vnp: ventral brain neuropil; vrcc: ventral root of the circumoesophageal connective.

Figure 4.
Lateral brain neuropils in Arabella iricolor (Oenonidae) and Scoletoma tetraura (Lumbrineridae). Histological cross sections (5 µm, AZAN) (A,C,D). Three-dimensional reconstructions based on histological serial sections (B,E). (A) The lateral brain neuropil (lnp) is surrounded by a dense layer of somata of type 1 neurons (S1) and intermediate filaments (if) of glia cells in A. iricolor. (B) 3D reconstruction of the lnp and the tract (tlnp) connecting both neuropils in the brain (np) in A. iricolor. The dorsal nerves (dn) root partially in the tlnp. Frontal view. (C) The dn root in the tlnp and the dorsal brain neuropil (dnp) in A. iricolor. The tract is surrounded by intermediate filaments (if) of glia cells. (D) A dense layer of somata of type 1 neurons (S1) surround the lnp in S. tetraura. (E) A 3D reconstruction of the lnp and the tlnp connecting both neuropils in the brain (np) of S. tetraura. Frontal view. d: dorsal; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dn: dorsal nerves; dnp: dorsal brain neuropil; if: intermediate filaments; lnp: lateral brain neuropil; np: neuropil; S1: somata of type 1 neurons; tlnp: tract of the lateral brain neuropils; v: ventral; vnp: ventral brain neuropil.
Figure 4.
Lateral brain neuropils in Arabella iricolor (Oenonidae) and Scoletoma tetraura (Lumbrineridae). Histological cross sections (5 µm, AZAN) (A,C,D). Three-dimensional reconstructions based on histological serial sections (B,E). (A) The lateral brain neuropil (lnp) is surrounded by a dense layer of somata of type 1 neurons (S1) and intermediate filaments (if) of glia cells in A. iricolor. (B) 3D reconstruction of the lnp and the tract (tlnp) connecting both neuropils in the brain (np) in A. iricolor. The dorsal nerves (dn) root partially in the tlnp. Frontal view. (C) The dn root in the tlnp and the dorsal brain neuropil (dnp) in A. iricolor. The tract is surrounded by intermediate filaments (if) of glia cells. (D) A dense layer of somata of type 1 neurons (S1) surround the lnp in S. tetraura. (E) A 3D reconstruction of the lnp and the tlnp connecting both neuropils in the brain (np) of S. tetraura. Frontal view. d: dorsal; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dn: dorsal nerves; dnp: dorsal brain neuropil; if: intermediate filaments; lnp: lateral brain neuropil; np: neuropil; S1: somata of type 1 neurons; tlnp: tract of the lateral brain neuropils; v: ventral; vnp: ventral brain neuropil.

Figure 5.
Brain neuropil and different types of neuronal somata. Ultra-thin cross sections (70 nm) of Paucibranchia bellii (A,B) and histological cross sections (5 µm, AZAN) of Scoletoma tetraura (C–F). (A) Distribution of different types of somata (S1, S2, S3) around the brain neuropil (np). (B) Somata of type 1 neurons (S1). (C) Somata of type 2 neurons (S2). (D) Somata of type 3 neurons (S3) with prominent nucleoli (nuc) in the nucleus. © The brain neuropil (np) consists of neurites (nt) of different sizes and orientations. Processes of glial cells (pgc) surround the central neuropil (np) creating a layer between the central neuropil (np) and the neuronal somata (so). Nuclei of the glial cells (ngc) are brick-shaped and processes of the glial cells (pgc) can be distinguished. Neuronal somata (so) are interconnected by desmosomes (ds). (F) Dense (dcv) and lucent core vesicles (lcv) are present in the brain containing neurotransmitters. dcv: dense core vesicles; ds: desmosomes; gc: glia cells; if: intermediate filaments; lcv: lucent core vesicles; ngc: nucleus of glia cell; np: neuropil; nt: neurite; nu: nucleus; nuc: nucleolus; pgc: processes of glia cell; S1: somata of type 1 neurons; S2: somata of type 2 neurons; S3: somata of type 3 neurons; so: somata.
Figure 5.
Brain neuropil and different types of neuronal somata. Ultra-thin cross sections (70 nm) of Paucibranchia bellii (A,B) and histological cross sections (5 µm, AZAN) of Scoletoma tetraura (C–F). (A) Distribution of different types of somata (S1, S2, S3) around the brain neuropil (np). (B) Somata of type 1 neurons (S1). (C) Somata of type 2 neurons (S2). (D) Somata of type 3 neurons (S3) with prominent nucleoli (nuc) in the nucleus. © The brain neuropil (np) consists of neurites (nt) of different sizes and orientations. Processes of glial cells (pgc) surround the central neuropil (np) creating a layer between the central neuropil (np) and the neuronal somata (so). Nuclei of the glial cells (ngc) are brick-shaped and processes of the glial cells (pgc) can be distinguished. Neuronal somata (so) are interconnected by desmosomes (ds). (F) Dense (dcv) and lucent core vesicles (lcv) are present in the brain containing neurotransmitters. dcv: dense core vesicles; ds: desmosomes; gc: glia cells; if: intermediate filaments; lcv: lucent core vesicles; ngc: nucleus of glia cell; np: neuropil; nt: neurite; nu: nucleus; nuc: nucleolus; pgc: processes of glia cell; S1: somata of type 1 neurons; S2: somata of type 2 neurons; S3: somata of type 3 neurons; so: somata.

Figure 6.
Generalized schemes of the innervation patterns of species bearing different numbers of antennae, palps, and eyes and conditions in species lacking these sensory organs. The asterisk (*) in picture D indicates the inclusion of data from Zanol [20]. (A) Innervation patterns found in Eunicidae and Onuphidae bearing 5 head appendages (Pauchibranchia bellii, Marphysa spec., Leodice torquata, Hyalinoecia tubicola). (B) Innervation patterns found in Lysidine ninetta (3 head appendages). (C) Innervation patterns of Nematonereis unicornis (1 head appendage). (D) Innervation patterns of Dorvillea spec. (4 head appendages). (E) Innervation patterns of Arabella iricolor (lacking head appendages, but possessing two pairs of eyes). (F) Conditions found in Scoletoma tetraura and Scoletoma fragilis (lacking head appnendages and eyes). aoc: anterior optical commissure; atla: association tract of the lateral antennae; atma: association tract of the median antenna; bln: buccal lip nerve; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dn: dorsal nerve; e: eye; ic: intracerebral cavity; lan: lateral antenna nerve; lnp: lateral neuropil; lnpc: lateral neuropil commissure; lnpc1: first commissure of the lateral neuropils; lnpc2: second commissure of the lateral neuropils; man: median antenna nerve; np: brain neuropil; oc: optical commissure; pn: palp nerve; pnr: palp nerve root; poc: posterior optical commissure.
Figure 6.
Generalized schemes of the innervation patterns of species bearing different numbers of antennae, palps, and eyes and conditions in species lacking these sensory organs. The asterisk (*) in picture D indicates the inclusion of data from Zanol [20]. (A) Innervation patterns found in Eunicidae and Onuphidae bearing 5 head appendages (Pauchibranchia bellii, Marphysa spec., Leodice torquata, Hyalinoecia tubicola). (B) Innervation patterns found in Lysidine ninetta (3 head appendages). (C) Innervation patterns of Nematonereis unicornis (1 head appendage). (D) Innervation patterns of Dorvillea spec. (4 head appendages). (E) Innervation patterns of Arabella iricolor (lacking head appendages, but possessing two pairs of eyes). (F) Conditions found in Scoletoma tetraura and Scoletoma fragilis (lacking head appnendages and eyes). aoc: anterior optical commissure; atla: association tract of the lateral antennae; atma: association tract of the median antenna; bln: buccal lip nerve; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dn: dorsal nerve; e: eye; ic: intracerebral cavity; lan: lateral antenna nerve; lnp: lateral neuropil; lnpc: lateral neuropil commissure; lnpc1: first commissure of the lateral neuropils; lnpc2: second commissure of the lateral neuropils; man: median antenna nerve; np: brain neuropil; oc: optical commissure; pn: palp nerve; pnr: palp nerve root; poc: posterior optical commissure.

Figure 7.
Schematic drawings of association tracts and nerve roots (light grey, reddish) of species bearing five head appendages based on a serial histological cross section of the dorsal brain neuropil (dark grey). (A) Eunicidae (Marphysa spec., Paucibranchia bellii and Leodice torquata). (B) Onuphidae (Hyalinoecia tubicola). atla: association tract of the lateral antenna; atla1: first association tract of the lateral antenna; atla2: second association tract of the lateral antenna; atma: association tract of the median antenna; atmla: association tract of the median and the lateral antennae; cas: commissure of the association system; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dfm: dorsal fibril mass; dnp: dorsal brain neuropil; drcc: dorsal root of the circumoesophageal connective; lan: lateral antenna nerve; lanr: nerve root of the lateral antenna; latla: lateral association tract of the lateral antenna; latla1: lateral root of the first association tract of the lateral antenna; latla2: lateral root of the second association tract of the lateral antenna; llanr: lateral root of the lateral antenna nerve; man: median antenna nerve; matla: median association tract of the lateral antenna; matla1: median root of the first association tract of the lateral antenna; matla2: median root of the second association tract of the lateral antenna; manr1: first root of the median antenna nerve; manr2: second root of the median antenna nerve; manr3: third root of the median antenna nerve; mlanr: median root of the lateral antenna nerve; mlanr2: second median root of the lateral antenna nerve; mlanr3: third median root of the lateral antenna nerve; pn: palp nerve; pnr1: first root of the palp nerve; pnr2: second root of the palp nerve; pnr3: third root of the palp nerve.
Figure 7.
Schematic drawings of association tracts and nerve roots (light grey, reddish) of species bearing five head appendages based on a serial histological cross section of the dorsal brain neuropil (dark grey). (A) Eunicidae (Marphysa spec., Paucibranchia bellii and Leodice torquata). (B) Onuphidae (Hyalinoecia tubicola). atla: association tract of the lateral antenna; atla1: first association tract of the lateral antenna; atla2: second association tract of the lateral antenna; atma: association tract of the median antenna; atmla: association tract of the median and the lateral antennae; cas: commissure of the association system; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dfm: dorsal fibril mass; dnp: dorsal brain neuropil; drcc: dorsal root of the circumoesophageal connective; lan: lateral antenna nerve; lanr: nerve root of the lateral antenna; latla: lateral association tract of the lateral antenna; latla1: lateral root of the first association tract of the lateral antenna; latla2: lateral root of the second association tract of the lateral antenna; llanr: lateral root of the lateral antenna nerve; man: median antenna nerve; matla: median association tract of the lateral antenna; matla1: median root of the first association tract of the lateral antenna; matla2: median root of the second association tract of the lateral antenna; manr1: first root of the median antenna nerve; manr2: second root of the median antenna nerve; manr3: third root of the median antenna nerve; mlanr: median root of the lateral antenna nerve; mlanr2: second median root of the lateral antenna nerve; mlanr3: third median root of the lateral antenna nerve; pn: palp nerve; pnr1: first root of the palp nerve; pnr2: second root of the palp nerve; pnr3: third root of the palp nerve.

Figure 8.
Association tracts of Pauchibranchia bellii, Marphysa spec., Leodice torquata and dorsal nerves of Arabella iricolor. Histological cross sections (5 µm), silver staining (A–C,E,G), AZAN staining (F). Three-dimensional reconstructions based on histological serial sections (D). Pictures (B,C,E,G) show association tracts in a consecutive anterior-posterior course. (A) Marphysa spec. The nerves of the median and the lateral antenna (man, lan) root in the dorsal commissure of the dorsal root of the circumoesophageal connective (dcdr) and the respective association tracts (atma, atla). The association tract of the median antenna (atma) proceeds in a ventral direction through the dorsal brain neuropil (dnp). The association tract of the lateral antenna (atla) runs diagonally through the dorsal brain neuropil. (B) Leodice torquata. The association tracts of the lateral antennae (atla) fuse with the association tracts of the median antenna (atma) in the ventral part of the dorsal brain neuropil (dnp). (C) L. torquata. The association tracts of both lateral antenna (atla) fuse in the ventral brain neuropil creating a ventral commissure (vc). (D) Paucibranchia bellii, frontal view. The association tracts of the median antenna (atma) and the lateral antennae (atla) fuse in two association tracts (atmla). The association tracts of the median and the lateral antennae (atmla) proceed in the anterior direction through the dorsal brain neuropil. (E) L. torquata. Both nerves of the median antenna (man) fuse in one association tract (atma). (F) Arabella iricolor. A pair of dorsal nerves (dn) originates from the dorsal brain neuropil (dnp) in a topologically similar position as the median antenna nerves of antennae-bearing species. (G) L. torquata. The association tract of the median antenna (atma) splits into two tracts, which will fuse with the respective association tracts of the lateral antenna (atla) creating the association tracts of the median and the lateral antennae (atmla). atla: association tract of the lateral antenna; atma: association tract of the median antenna; atmla: association tract of the median and the lateral antennae; d: dorsal; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dn: dorsal nerves; dnp: dorsal brain neuropil; ey: eye; la: lateral antenna; lan: lateral antenna nerve; ma: median antenna; man: median antenna nerve; np: neuropil; on: optical nerve; pn: palp nerve; vc: ventral commissure; v: ventral.
Figure 8.
Association tracts of Pauchibranchia bellii, Marphysa spec., Leodice torquata and dorsal nerves of Arabella iricolor. Histological cross sections (5 µm), silver staining (A–C,E,G), AZAN staining (F). Three-dimensional reconstructions based on histological serial sections (D). Pictures (B,C,E,G) show association tracts in a consecutive anterior-posterior course. (A) Marphysa spec. The nerves of the median and the lateral antenna (man, lan) root in the dorsal commissure of the dorsal root of the circumoesophageal connective (dcdr) and the respective association tracts (atma, atla). The association tract of the median antenna (atma) proceeds in a ventral direction through the dorsal brain neuropil (dnp). The association tract of the lateral antenna (atla) runs diagonally through the dorsal brain neuropil. (B) Leodice torquata. The association tracts of the lateral antennae (atla) fuse with the association tracts of the median antenna (atma) in the ventral part of the dorsal brain neuropil (dnp). (C) L. torquata. The association tracts of both lateral antenna (atla) fuse in the ventral brain neuropil creating a ventral commissure (vc). (D) Paucibranchia bellii, frontal view. The association tracts of the median antenna (atma) and the lateral antennae (atla) fuse in two association tracts (atmla). The association tracts of the median and the lateral antennae (atmla) proceed in the anterior direction through the dorsal brain neuropil. (E) L. torquata. Both nerves of the median antenna (man) fuse in one association tract (atma). (F) Arabella iricolor. A pair of dorsal nerves (dn) originates from the dorsal brain neuropil (dnp) in a topologically similar position as the median antenna nerves of antennae-bearing species. (G) L. torquata. The association tract of the median antenna (atma) splits into two tracts, which will fuse with the respective association tracts of the lateral antenna (atla) creating the association tracts of the median and the lateral antennae (atmla). atla: association tract of the lateral antenna; atma: association tract of the median antenna; atmla: association tract of the median and the lateral antennae; d: dorsal; dcdr: dorsal commissure of the dorsal root of the circumoesophageal connective; dn: dorsal nerves; dnp: dorsal brain neuropil; ey: eye; la: lateral antenna; lan: lateral antenna nerve; ma: median antenna; man: median antenna nerve; np: neuropil; on: optical nerve; pn: palp nerve; vc: ventral commissure; v: ventral.

Figure 9.
Inner morphology and innervation of antennae and palps. Histological cross sections (5 µm) stained in AZAN (A,E–H) or silver (C,D). Horizontal section (5 µm, AZAN) (B). (A) Leodice torquata (lateral antenna). Antennae as well as palps possess one main nerve, which consists of medially located longitudinal nerve fibres (ln) and surrounding circular nerve fibres (cn). Both types of nerve fibres possess nuclei of glia cells (gcn). (B) Leodice torquata (palp). Longitudinal nerve fibres (ln) and circular nerve fibres (cn) proceed nearly until the tip of the respective head appendage. (C) Marphysa spec.: The main nerve of the median antenna and the lateral antenna splits at height of the coelomic cavity (co) into two main roots (median antenna: 2x manr1; lateral antenna: mlanr1, llanr), respectively. Each main nerve root runs alongside the antenna coelom (palp coelom) (co) towards the brain (np). Prior to entering the brain, the main nerve roots split into several minor nerve roots (e.g., manr2, manr3, atma), some of which originate from the dcdr or proceed as association tracts (e.g., atma) through the dorsal brain neuropil. Each antenna or palp coelom (co) contains musculature (mu). (D) Marphysa spec.: The palp nerve (pn) proceeds along the dorsally located side of the palp coelom (co) and splits into minor nerve roots prior to entering the brain (np). (E) Palp nerve roots exemplified for Leodice torquata. The palp nerve (pn) passes the palp coelom (co) and splits at least into three minor palp nerve roots (pnr) before entering the brain. The two more dorsally located palp nerve roots originate from the dorsal brain neuropil (dnp), and the more ventrally located palp nerve root originates from the ventral brain neuropil (vnp). (F,G) In Dorvillea spec. each palp is innervated by two palp nerves (dpn, vnp). The more dorsally located palp nerve (dpn) originates from the dorsal brain neuropil (dnp), and the more ventrally located palp nerve (vpn) originates from the ventral brain neuropil (vnp). Both nerves proceed together with the coelomic cavity (co) and musculature (mu) through the palp. (H) Separation of palps into palpophore (pp) and palpostyle (ps) exemplified for Hyalinoecia tubicola. The palp is composed of an annulated basal palpophore (pp) and a distal palpostyle (ps). One palp nerve (pn) innervates the palp. atma = association tract of the median antenna, ca = caudal, cn = circular nerve fibres, co = coelom, cu = cuticle, d = dorsal, dcdr = dorsal commissure of the dorsal root of the circumoesophageal connective, dfm = dorsal fibril mass, dnp = dorsal brain neuropil, dpn = dorsal palp nerve, ep = epidermis, ey = eye, fr = frontal, gcn = nuclei of glia cells, la = lateral antenna, lan = lateral antenna nerve, llanr = lateral root of the lateral antenna nerve, ln = longitudinal nerve fibres, ma = median antenna, manr1 = first root of the median antenna nerve, manr2 = second root of the median antenna nerve, manr3 = third root of the median antenna nerve, mlanr1 = median root of the lateral antenna nerve, mu = musculature, np = neuropil, p = palp, pn = palp nerve, pnr = roots of palp nerve, pp = palpophore, ps = palpostyle, S3 = somata of class 3 neurons, sc = stomatogastric cavity, so = somata, v = ventral, vnp = ventral brain neuropil, vpn = ventral palp nerve.
Figure 9.
Inner morphology and innervation of antennae and palps. Histological cross sections (5 µm) stained in AZAN (A,E–H) or silver (C,D). Horizontal section (5 µm, AZAN) (B). (A) Leodice torquata (lateral antenna). Antennae as well as palps possess one main nerve, which consists of medially located longitudinal nerve fibres (ln) and surrounding circular nerve fibres (cn). Both types of nerve fibres possess nuclei of glia cells (gcn). (B) Leodice torquata (palp). Longitudinal nerve fibres (ln) and circular nerve fibres (cn) proceed nearly until the tip of the respective head appendage. (C) Marphysa spec.: The main nerve of the median antenna and the lateral antenna splits at height of the coelomic cavity (co) into two main roots (median antenna: 2x manr1; lateral antenna: mlanr1, llanr), respectively. Each main nerve root runs alongside the antenna coelom (palp coelom) (co) towards the brain (np). Prior to entering the brain, the main nerve roots split into several minor nerve roots (e.g., manr2, manr3, atma), some of which originate from the dcdr or proceed as association tracts (e.g., atma) through the dorsal brain neuropil. Each antenna or palp coelom (co) contains musculature (mu). (D) Marphysa spec.: The palp nerve (pn) proceeds along the dorsally located side of the palp coelom (co) and splits into minor nerve roots prior to entering the brain (np). (E) Palp nerve roots exemplified for Leodice torquata. The palp nerve (pn) passes the palp coelom (co) and splits at least into three minor palp nerve roots (pnr) before entering the brain. The two more dorsally located palp nerve roots originate from the dorsal brain neuropil (dnp), and the more ventrally located palp nerve root originates from the ventral brain neuropil (vnp). (F,G) In Dorvillea spec. each palp is innervated by two palp nerves (dpn, vnp). The more dorsally located palp nerve (dpn) originates from the dorsal brain neuropil (dnp), and the more ventrally located palp nerve (vpn) originates from the ventral brain neuropil (vnp). Both nerves proceed together with the coelomic cavity (co) and musculature (mu) through the palp. (H) Separation of palps into palpophore (pp) and palpostyle (ps) exemplified for Hyalinoecia tubicola. The palp is composed of an annulated basal palpophore (pp) and a distal palpostyle (ps). One palp nerve (pn) innervates the palp. atma = association tract of the median antenna, ca = caudal, cn = circular nerve fibres, co = coelom, cu = cuticle, d = dorsal, dcdr = dorsal commissure of the dorsal root of the circumoesophageal connective, dfm = dorsal fibril mass, dnp = dorsal brain neuropil, dpn = dorsal palp nerve, ep = epidermis, ey = eye, fr = frontal, gcn = nuclei of glia cells, la = lateral antenna, lan = lateral antenna nerve, llanr = lateral root of the lateral antenna nerve, ln = longitudinal nerve fibres, ma = median antenna, manr1 = first root of the median antenna nerve, manr2 = second root of the median antenna nerve, manr3 = third root of the median antenna nerve, mlanr1 = median root of the lateral antenna nerve, mu = musculature, np = neuropil, p = palp, pn = palp nerve, pnr = roots of palp nerve, pp = palpophore, ps = palpostyle, S3 = somata of class 3 neurons, sc = stomatogastric cavity, so = somata, v = ventral, vnp = ventral brain neuropil, vpn = ventral palp nerve.

Figure 10.
Anatomy and innervation of buccal lips. Ventral view of the prostomium (A,B). Histological cross sections (5 µm, AZAN) (D,E). (A) Paired buccal lips (bl) of Leodice torquata. (B) In Hyalinoecia tubicola two pairs of buccal lips are present—one pair of dorsal buccal lips (dbl) and one pair of ventral buccal lips. Additionally, one pair of accessory buccal lips (abl) are present. (C) In Arabella iricolor buccal lips are absent. (D,E) Innervation of buccal lips (bl) in L. torquata. Buccal lips (bl) are innervated by nerves (bln) originating from the ventral brain neuropil (vnp). The nerves run along the cavity of the buccal lip (blc). abl = accessory buccal lips, bl = buccal lip, blc = coelomic cavity of the buccal lip, bln = buccal lip nerve, dbl = dorsal buccal lip, dnp = dorsal brain neuropil, mo = mouth opening, vbl = ventral buccal lip, vnp = ventral brain neuropil.
Figure 10.
Anatomy and innervation of buccal lips. Ventral view of the prostomium (A,B). Histological cross sections (5 µm, AZAN) (D,E). (A) Paired buccal lips (bl) of Leodice torquata. (B) In Hyalinoecia tubicola two pairs of buccal lips are present—one pair of dorsal buccal lips (dbl) and one pair of ventral buccal lips. Additionally, one pair of accessory buccal lips (abl) are present. (C) In Arabella iricolor buccal lips are absent. (D,E) Innervation of buccal lips (bl) in L. torquata. Buccal lips (bl) are innervated by nerves (bln) originating from the ventral brain neuropil (vnp). The nerves run along the cavity of the buccal lip (blc). abl = accessory buccal lips, bl = buccal lip, blc = coelomic cavity of the buccal lip, bln = buccal lip nerve, dbl = dorsal buccal lip, dnp = dorsal brain neuropil, mo = mouth opening, vbl = ventral buccal lip, vnp = ventral brain neuropil.

Figure 11.
Anatomy and innervation of the eyes. Histological cross sections (5 µm), AZAN staining (A,B), silver staining (C), 3D reconstructions (D,E). (A) Paucibranchia bellii. The eye is composed of a lens-like structure (lls) in the inner, covered by a layer of sensory microvilli (smv), pigmented supportive cells (psc), and photoreceptor cells (prc). Unpigmented sensory cells (usc) and microvilli (mv) cover the lens-like structure as the uppermost layer. The eye is situated basally to the epidermis (ep). A cuticular channel (cuc) can be identified projecting from the cuticle (cu) into the eye. The eye is innervated by an optical nerve (on) originating from the brain (np). The optical nerve (oc) splits next to the eye into minor nerves. (B) Arabella iricolor. Two pairs of eyes are present, the ventral one being bigger than the dorsal one. Both eye pairs are innervated by one optical nerve (on), respectively. (C,D) Lysidice ninetta. In the brain (np), the optical nerves are connected by an optical commissure (oc). (E) Arabella iricolor. Two optical commissures can be differentiated in the brain (np)—an anterior optical commissure (aoc) connects the optical nerves of the more ventrally located pair of eyes and a posterior optical commissure (poc) connects the optical nerves of the more dorsally located pair of eyes. ae = additional eye, aoc = anterior optical commissure, cu = cuticle, cuc = cuticular channel, dn = dorsal nerve, ep = epidermis, ey = eye, lan = lateral antenna nerve, lls = lens-like structure, man = median antenna nerve, mv = microvilli, no = nuchal organ, np = neuropil, on = optical nerve, poc = posterior optical commissure, prc = photoreceptor cell, psc = pigmented sensory cell, smv = sensory microvilli, usc = unpigmented sensory cells.
Figure 11.
Anatomy and innervation of the eyes. Histological cross sections (5 µm), AZAN staining (A,B), silver staining (C), 3D reconstructions (D,E). (A) Paucibranchia bellii. The eye is composed of a lens-like structure (lls) in the inner, covered by a layer of sensory microvilli (smv), pigmented supportive cells (psc), and photoreceptor cells (prc). Unpigmented sensory cells (usc) and microvilli (mv) cover the lens-like structure as the uppermost layer. The eye is situated basally to the epidermis (ep). A cuticular channel (cuc) can be identified projecting from the cuticle (cu) into the eye. The eye is innervated by an optical nerve (on) originating from the brain (np). The optical nerve (oc) splits next to the eye into minor nerves. (B) Arabella iricolor. Two pairs of eyes are present, the ventral one being bigger than the dorsal one. Both eye pairs are innervated by one optical nerve (on), respectively. (C,D) Lysidice ninetta. In the brain (np), the optical nerves are connected by an optical commissure (oc). (E) Arabella iricolor. Two optical commissures can be differentiated in the brain (np)—an anterior optical commissure (aoc) connects the optical nerves of the more ventrally located pair of eyes and a posterior optical commissure (poc) connects the optical nerves of the more dorsally located pair of eyes. ae = additional eye, aoc = anterior optical commissure, cu = cuticle, cuc = cuticular channel, dn = dorsal nerve, ep = epidermis, ey = eye, lan = lateral antenna nerve, lls = lens-like structure, man = median antenna nerve, mv = microvilli, no = nuchal organ, np = neuropil, on = optical nerve, poc = posterior optical commissure, prc = photoreceptor cell, psc = pigmented sensory cell, smv = sensory microvilli, usc = unpigmented sensory cells.

Figure 12.
Nuchal organs. Histological cross sections (5 µm) stained in AZAN (A–G) and immunohistochemically stained section (60 µm) (H). Schematic drawings (I–M), 3D reconstructions based on histological sections (N–P). (A) Paucibranchia bellii, unpaired, arc-shaped nuchal organ. (B) Lysidice ninetta, unpaired, arc-shaped nuchal organ. (C) Hyalinoecia tubicola, unpaired, arc-shaped nuchal organ. (D) Dorvillea spec., paired nuchal organs. (E) Arabella iricolor, paired, branched nuchal organ. (F) Scoletoma tetraura, unpaired, x-shaped nuchal organ. (G) L. ninetta, nuchal organ nerves (non) emanating from the brain (np) and innervating the nuchal organ. (H,I) Unpaired, arc-shaped nuchal organ in Eunicidae and Onuphidae. (J) Paired nuchal organ of Dorvillea spec. (K) Paired, branched nuchal organ of Oenonidae. (L) Unpaired, branched nuchal organ of Lumbrineridae. (M) Topology of the paired, branched nuchal organ in A. iricolor, lateral view. The nuchal organ (no) is located dorsally to dorsal brain neuropil (dnp) and is innervated by nerves (non) emanating from the posterior part of the brain. (N) Topology of the paired, branched nuchal organ in A. iricolor, dorsal view. (O) Topology of the unpaired, branched nuchal organ in S. tetraura, lateral view. The nuchal organ (no) is located dorsally to the posterior part of the dorsal brain neuropil (dnp). White arrow = cilia. ca = caudal, cu = cuticle, cp = ciliary patch, d = dorsal, dnp = dorsal brain neuropil, ey = eye, fr = frontal, m = musculature, nc = nuchal cavity, no = nuchal organ, non = nuchal organ nerve, np = neuropil, v = ventral.
Figure 12.
Nuchal organs. Histological cross sections (5 µm) stained in AZAN (A–G) and immunohistochemically stained section (60 µm) (H). Schematic drawings (I–M), 3D reconstructions based on histological sections (N–P). (A) Paucibranchia bellii, unpaired, arc-shaped nuchal organ. (B) Lysidice ninetta, unpaired, arc-shaped nuchal organ. (C) Hyalinoecia tubicola, unpaired, arc-shaped nuchal organ. (D) Dorvillea spec., paired nuchal organs. (E) Arabella iricolor, paired, branched nuchal organ. (F) Scoletoma tetraura, unpaired, x-shaped nuchal organ. (G) L. ninetta, nuchal organ nerves (non) emanating from the brain (np) and innervating the nuchal organ. (H,I) Unpaired, arc-shaped nuchal organ in Eunicidae and Onuphidae. (J) Paired nuchal organ of Dorvillea spec. (K) Paired, branched nuchal organ of Oenonidae. (L) Unpaired, branched nuchal organ of Lumbrineridae. (M) Topology of the paired, branched nuchal organ in A. iricolor, lateral view. The nuchal organ (no) is located dorsally to dorsal brain neuropil (dnp) and is innervated by nerves (non) emanating from the posterior part of the brain. (N) Topology of the paired, branched nuchal organ in A. iricolor, dorsal view. (O) Topology of the unpaired, branched nuchal organ in S. tetraura, lateral view. The nuchal organ (no) is located dorsally to the posterior part of the dorsal brain neuropil (dnp). White arrow = cilia. ca = caudal, cu = cuticle, cp = ciliary patch, d = dorsal, dnp = dorsal brain neuropil, ey = eye, fr = frontal, m = musculature, nc = nuchal cavity, no = nuchal organ, non = nuchal organ nerve, np = neuropil, v = ventral.

Figure 13.
Characters mapped on the latest phylogenetic tree of Eunicidae (Tilic et al. [2]). Dotted lines indicate unresolved phylogenetic positions: either Dorvilleidae and Histriobdellidae are a clade and sister group to the remaining Eunicidae, or Histriobdellidae is a sister taxon to all remaining Eunicidae. Numbers code for characters listed in the character matrix (see Table 2).
Figure 13.
Characters mapped on the latest phylogenetic tree of Eunicidae (Tilic et al. [2]). Dotted lines indicate unresolved phylogenetic positions: either Dorvilleidae and Histriobdellidae are a clade and sister group to the remaining Eunicidae, or Histriobdellidae is a sister taxon to all remaining Eunicidae. Numbers code for characters listed in the character matrix (see Table 2).


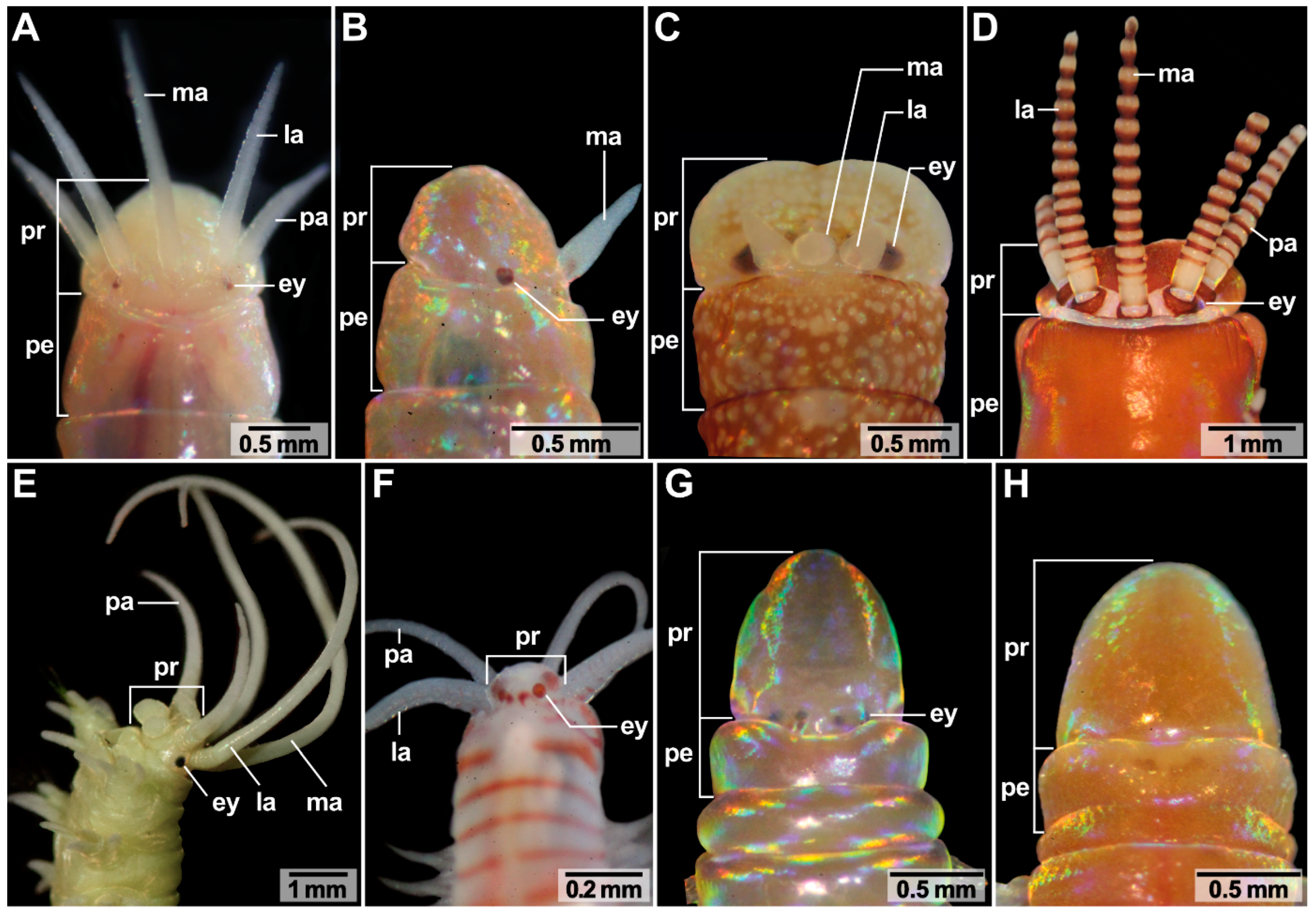{kind=link}
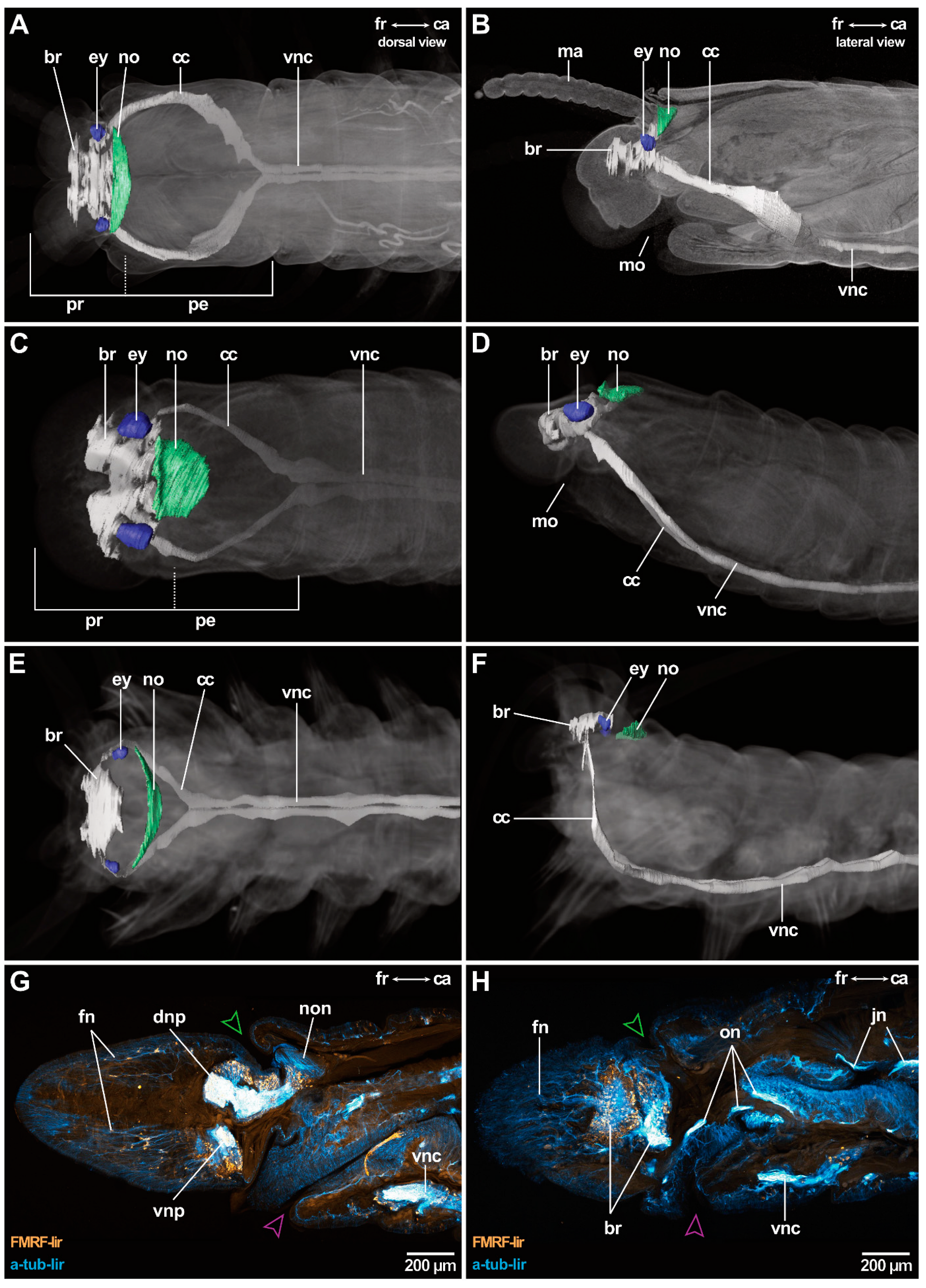{kind=link}
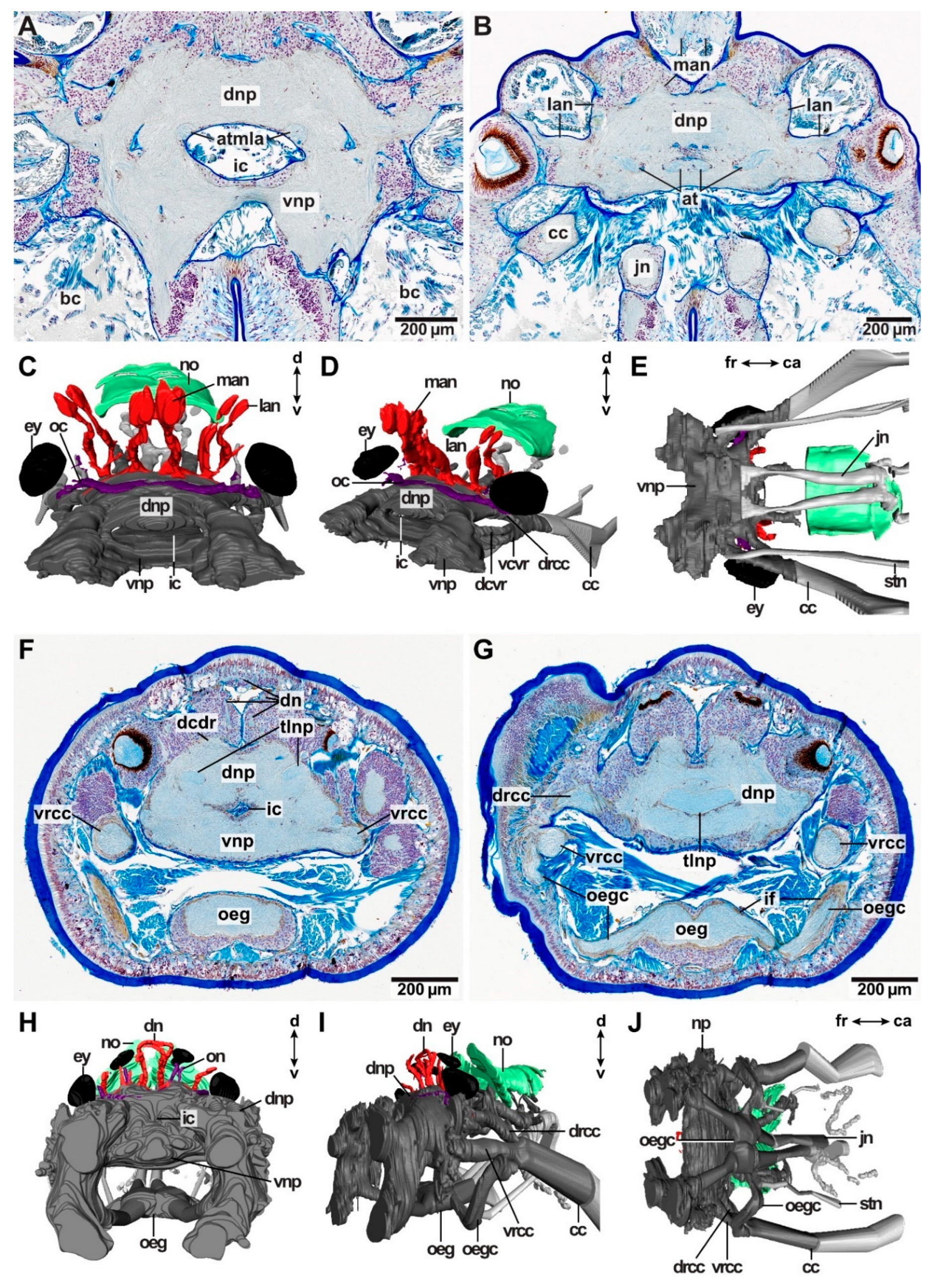{kind=link}
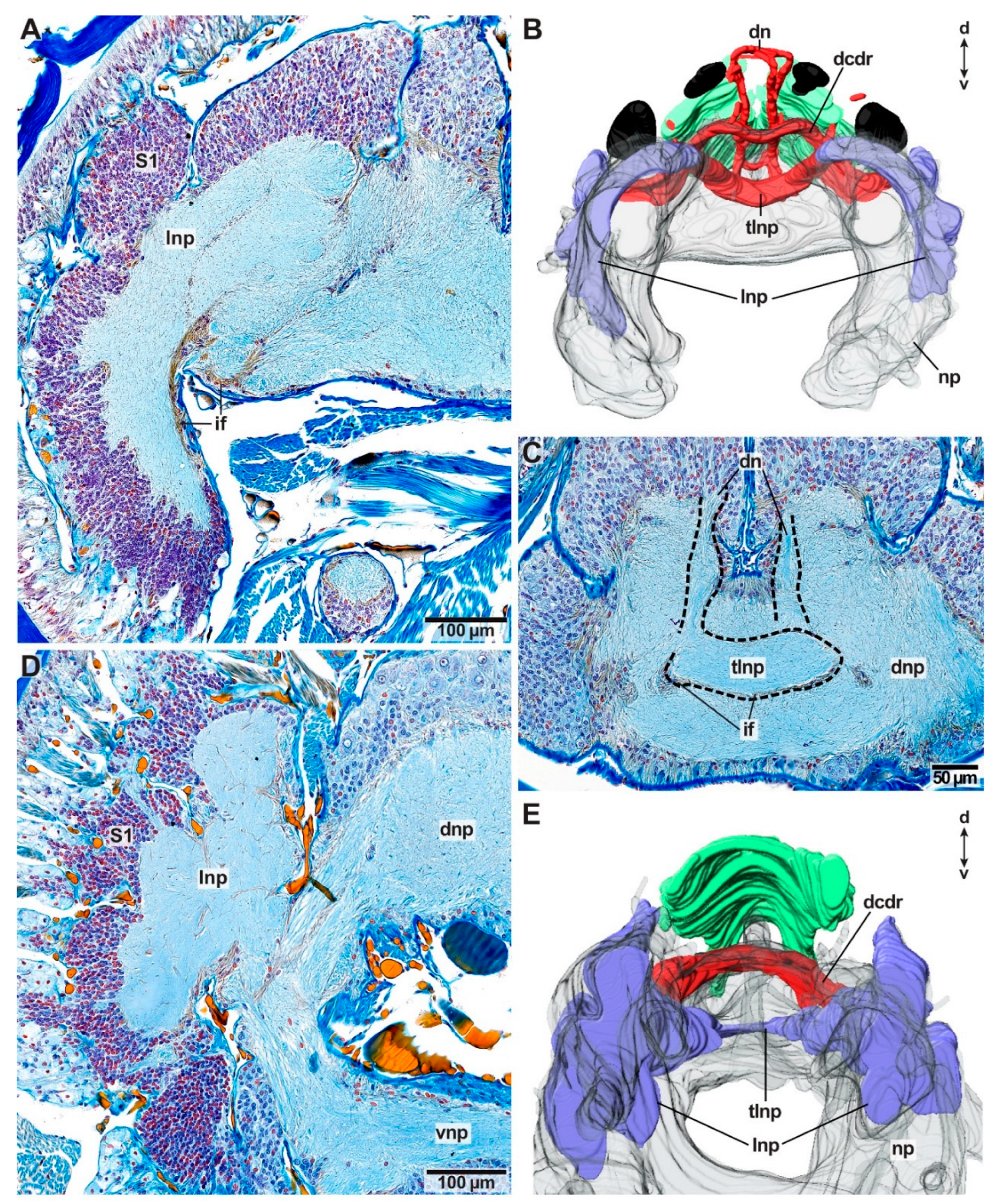{kind=link}
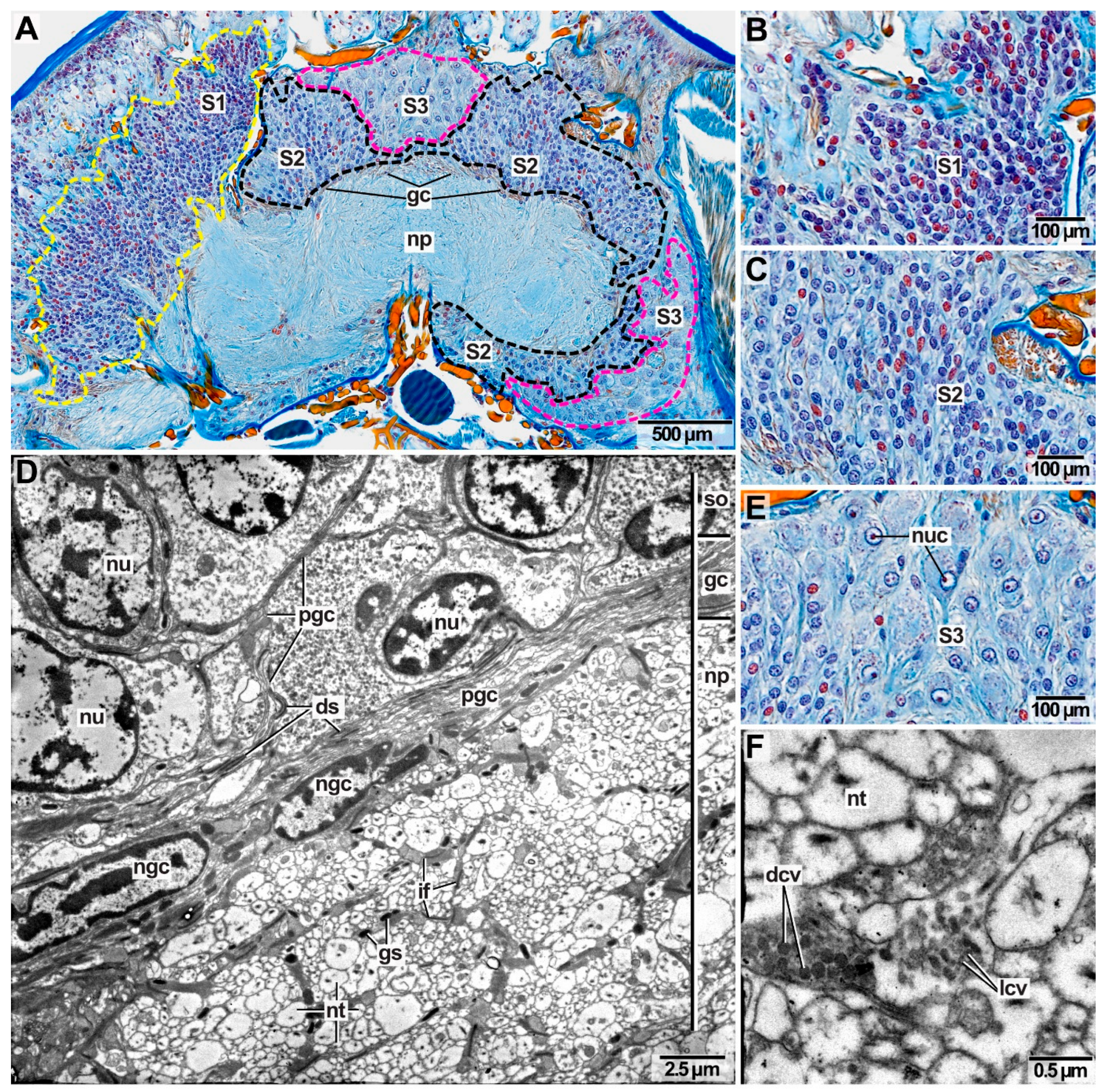{kind=link}
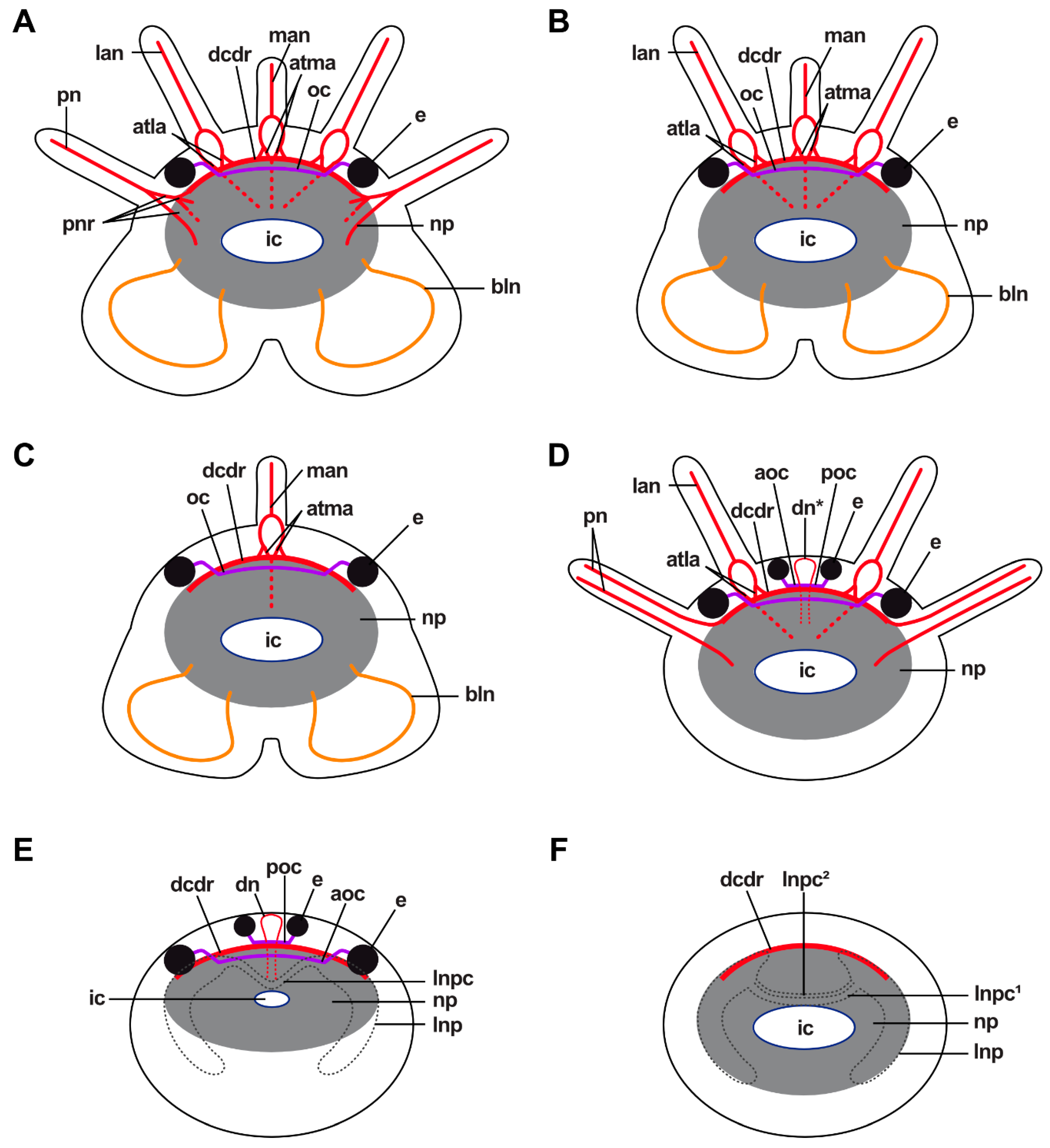{kind=link}
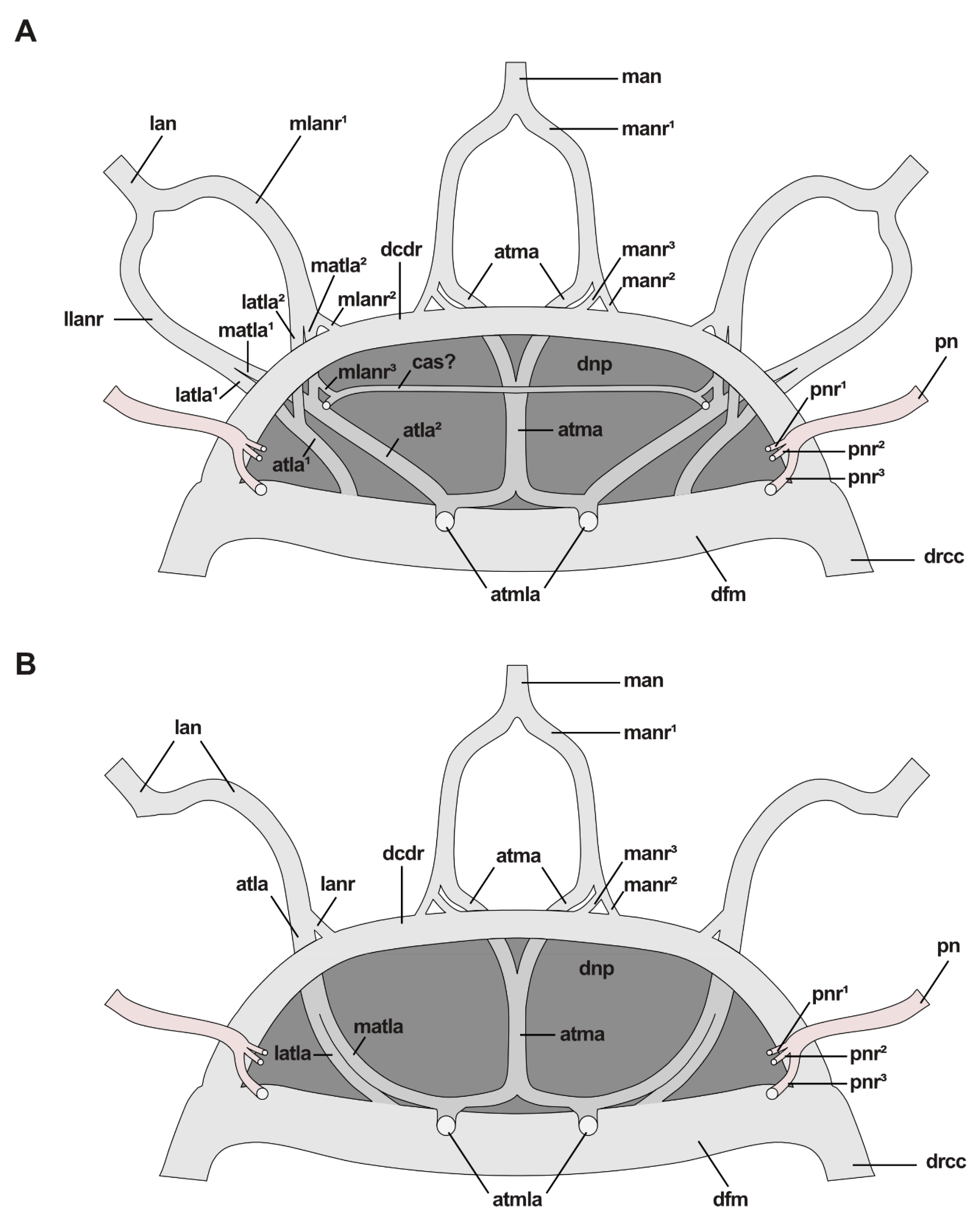{kind=link}
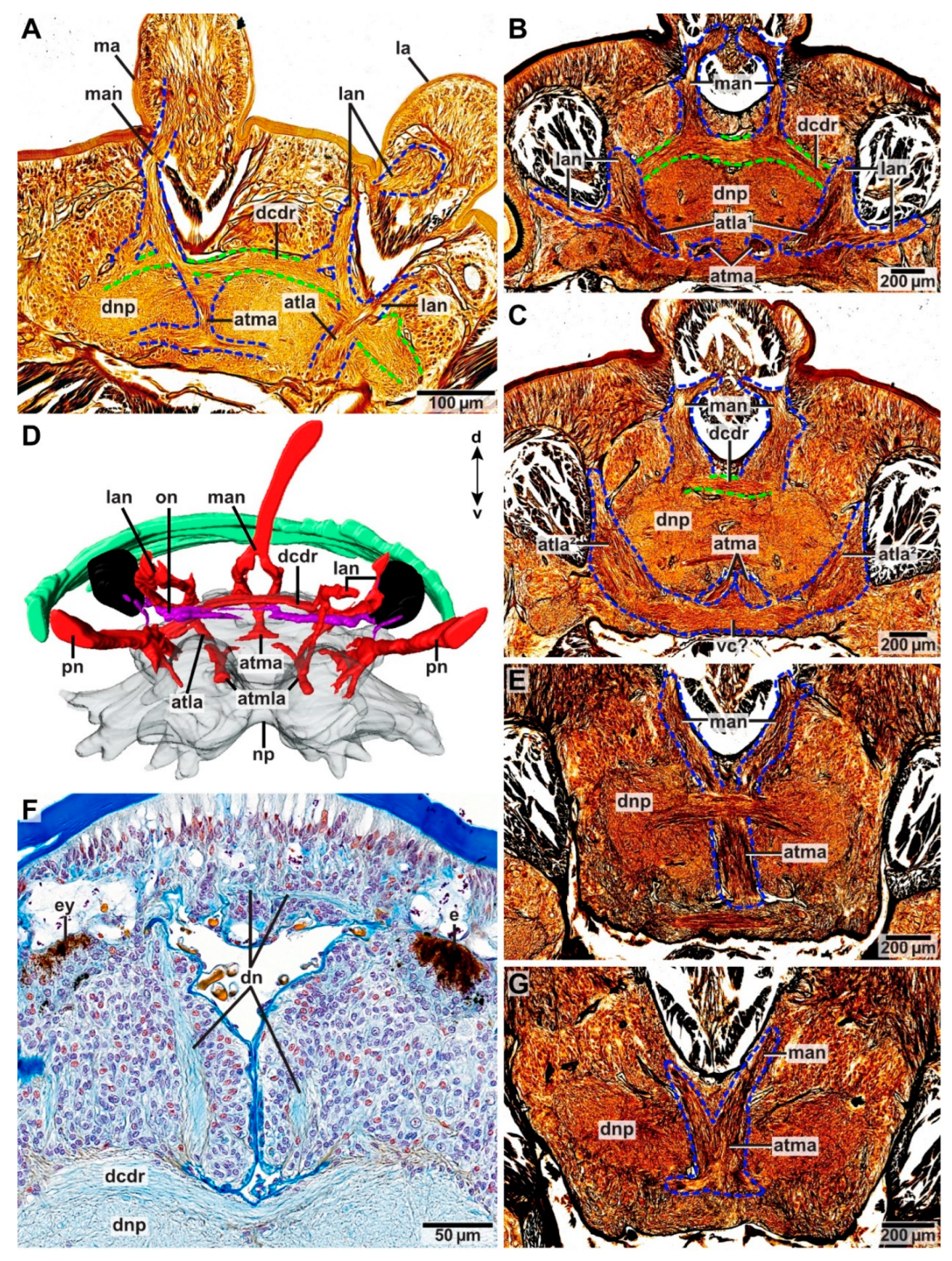{kind=link}
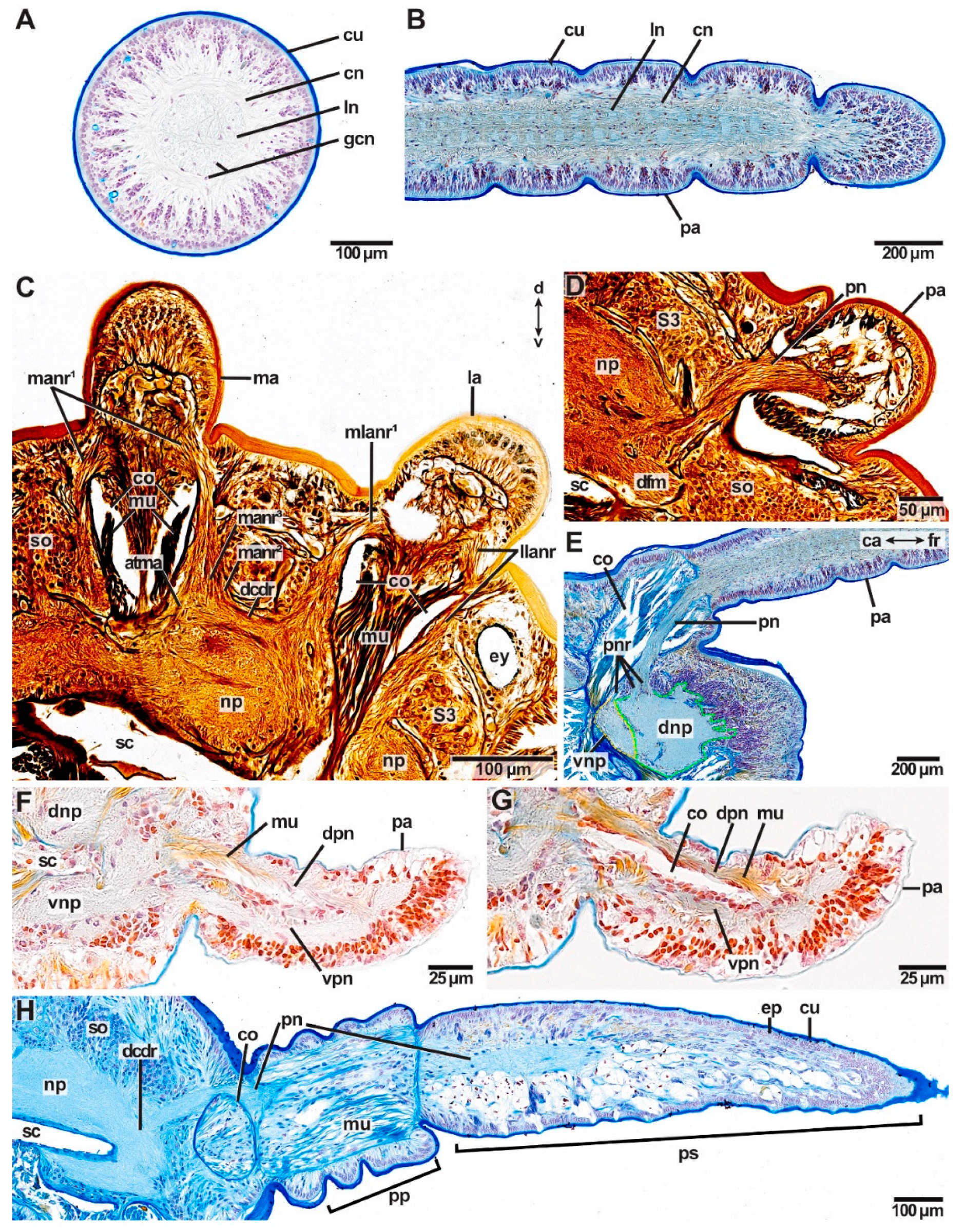{kind=link}
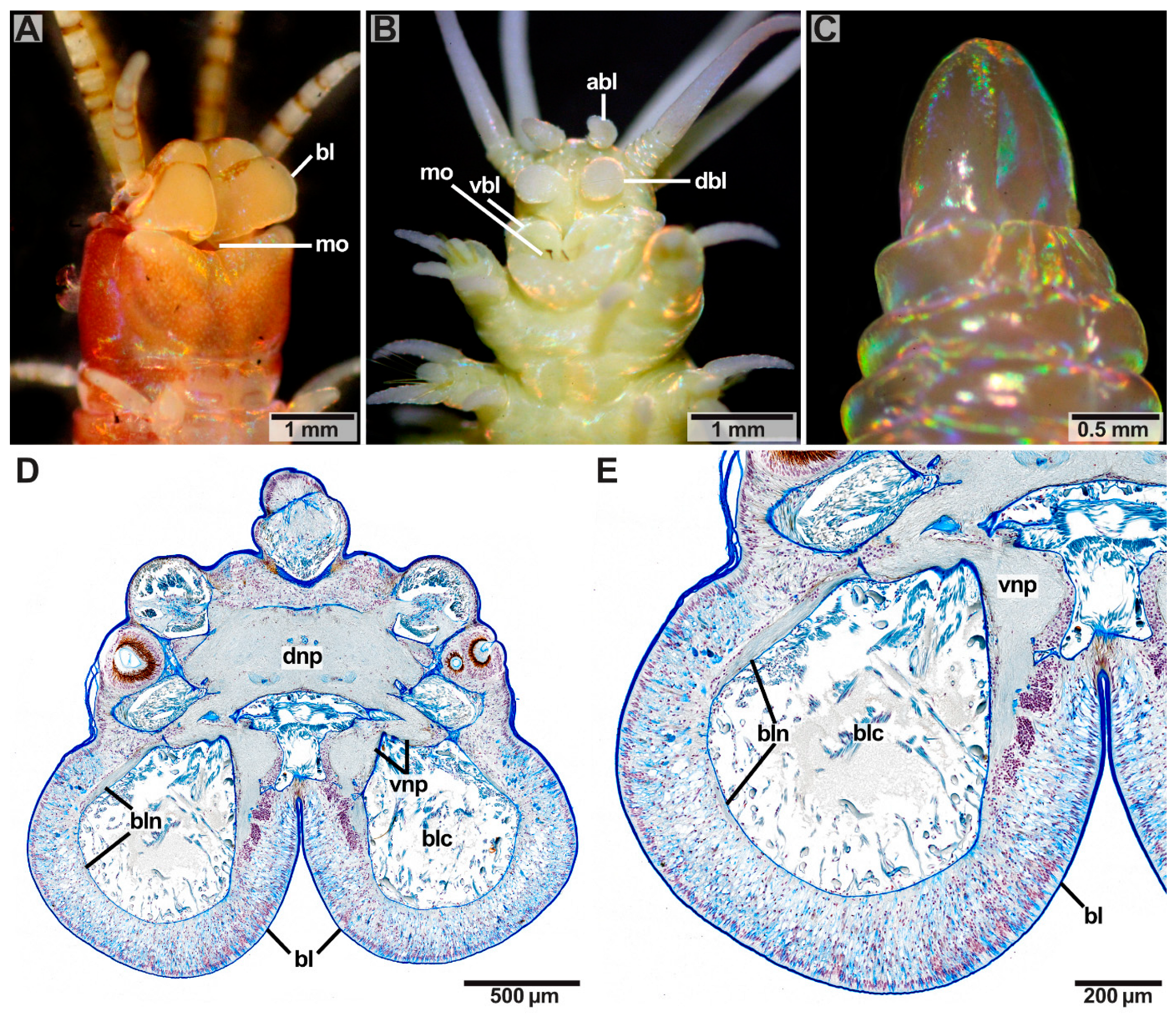{kind=link}
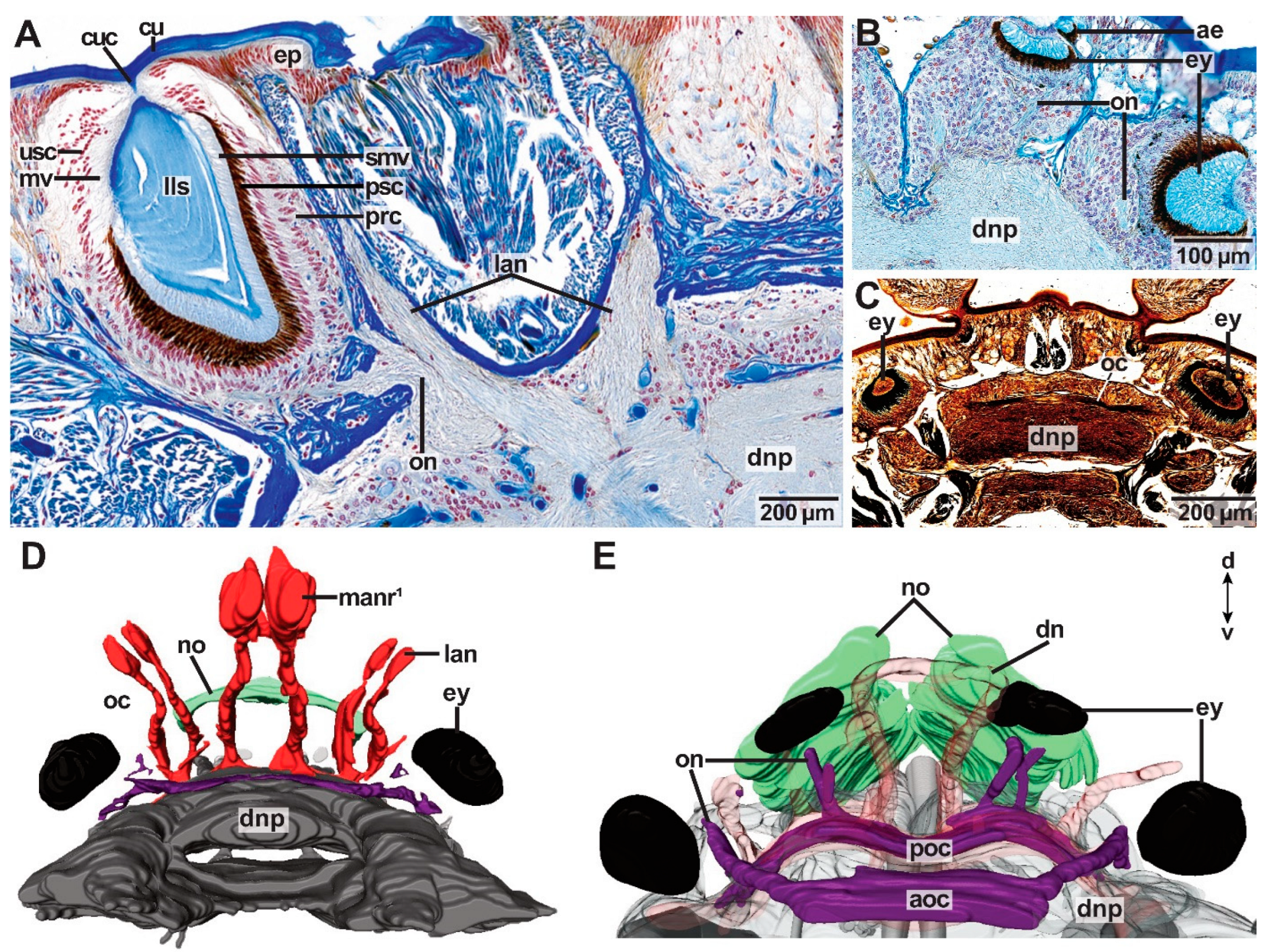{kind=link}
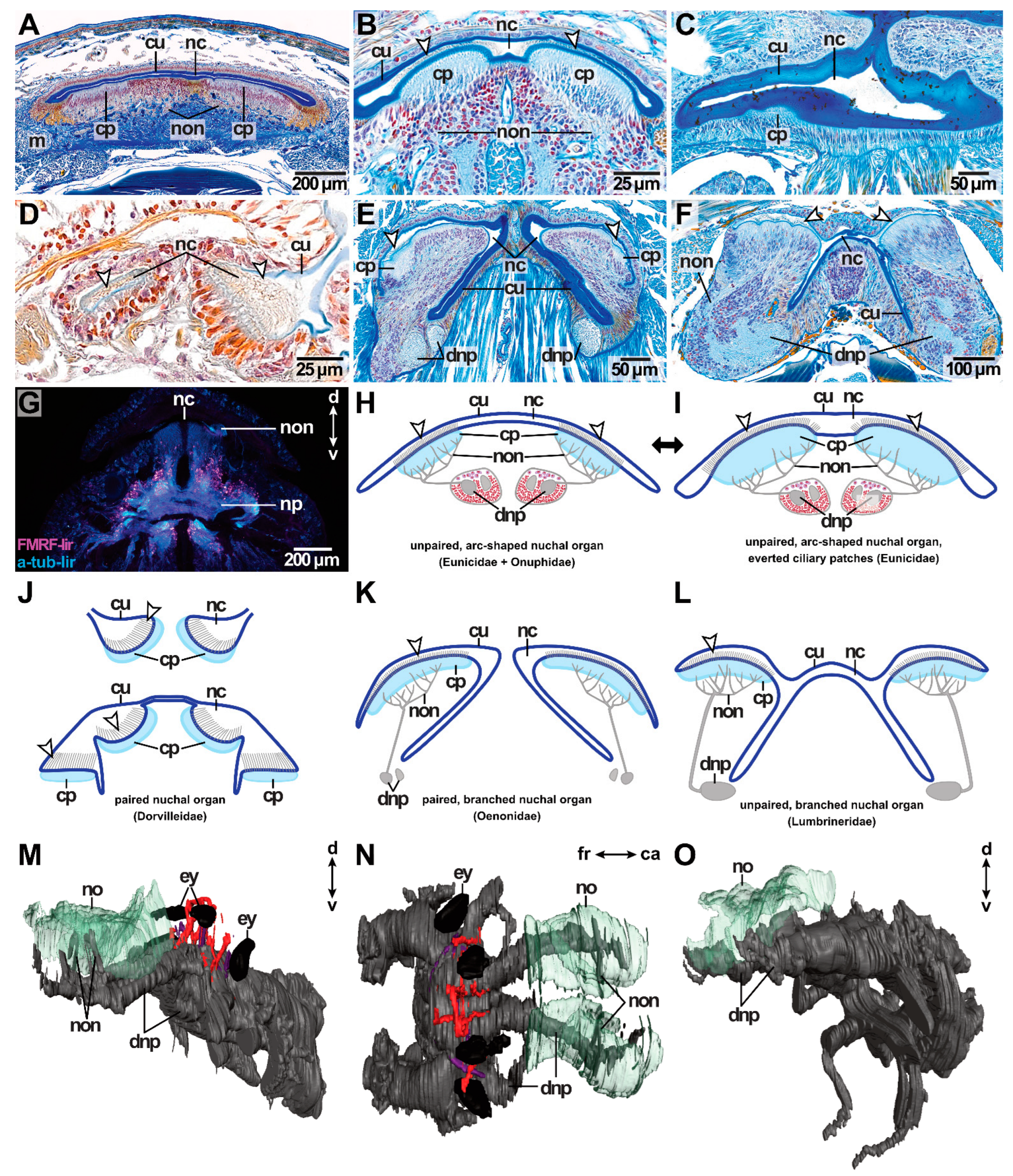{kind=link}
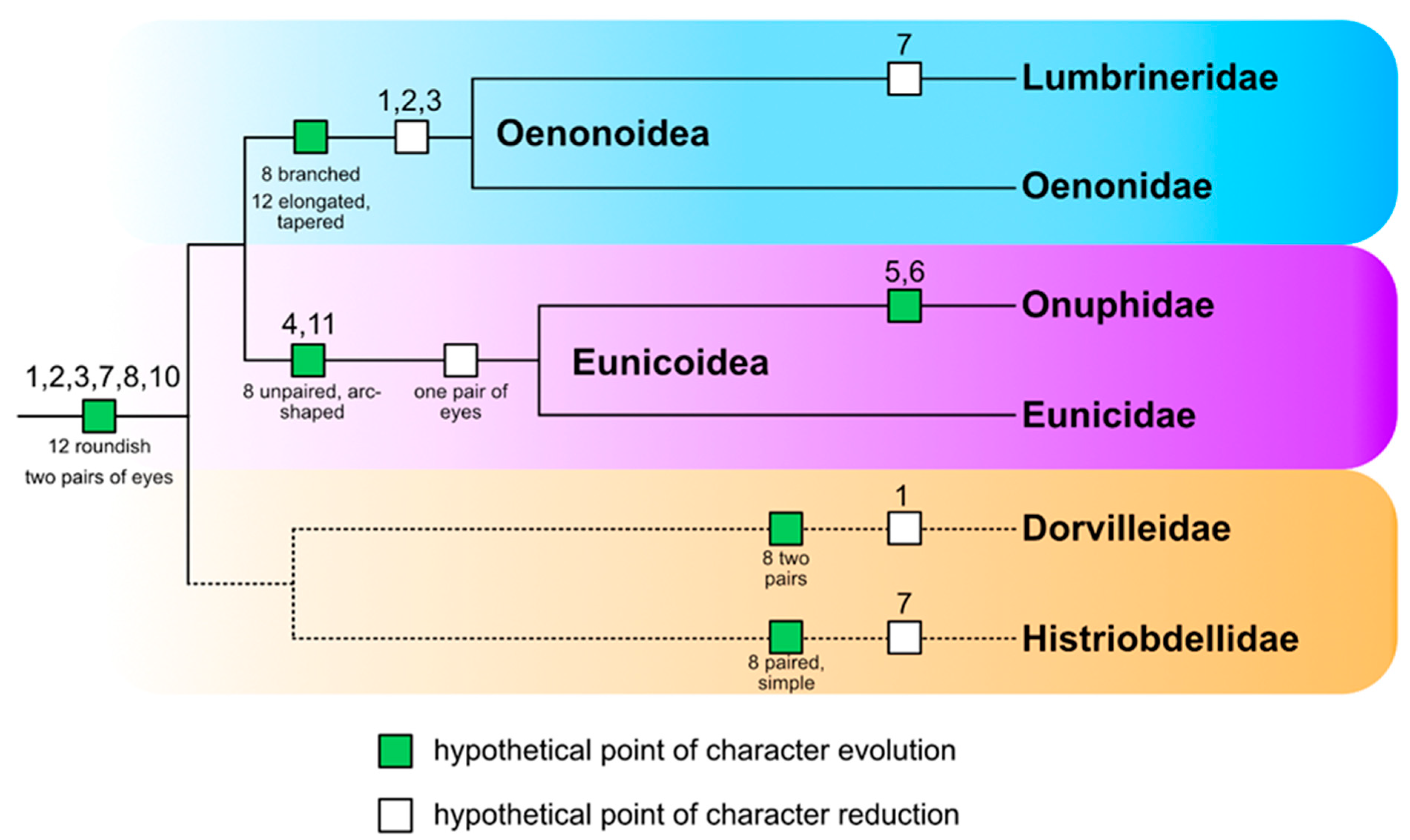{kind=link}
Table 1.
Number of sensory prostomial structures (antennae, palps, buccal lips, eyes and nuchal organ) in the examined Eunicida.
Table 1.
Number of sensory prostomial structures (antennae, palps, buccal lips, eyes and nuchal organ) in the examined Eunicida.
| Species | Median Antenna | Lateral Antennae | Palps | Buccal Lips | Eyes | Nuchal Organ |
|---|---|---|---|---|---|---|
| Pauchibranchia bellii | 1 | 2 | 2 | 2 | 2 | unpaired, arc-shaped |
| Marphysa spec. | 1 | 2 | 2 | 2 | 2 | unpaired, arc-shaped |
| Lysidice unicornis | 1 | 0 | 0 | 2 | 2 | unpaired, arc-shaped |
| Lysidine ninetta | 1 | 2 | 0 | 2 | 2 | unpaired, arc-shaped |
| Leodice torquata | 1 | 2 | 2 | 2 | 2 | unpaired, arc-shaped |
| Hyalinoecia tubicola | 1 | 2 | 2 | 2 | 2 | unpaired, arc-shaped |
| Dorvillea spec. | 0 | 2 | 2 | 0 | 4 | paired, simple |
| Arabella iricolor | 0 | 0 | 0 | 0 | 4 (5) | paired, branched |
| Scoletoma tetraura | 0 | 0 | 0 | 0 | 0 | unpaired, x-shaped |
| Scoletoma fragilis | 0 | 0 | 0 | 0 | 0 | unpaired, x-shaped |
| Histriobdella homari | 1 | 2 | 2 | 0? | 0? | “oval, very small pits” |
Table 2.
Character matrix. Depicted is the distribution of characters among the six studied families of Eunicida. The presence of a character is indicated by “+”, while its absence (or reduction) is marked with “-”. abl: accessory buccal lips, at: association tracts, dbl: dorsal buccal lips, ey: eye, la: lateral antenna, lo: lateral organ, ma: median antenna, no: nuchal organ, pa: palp, pr: prostomium, vbl: ventral buccal lips.
Table 2.
Character matrix. Depicted is the distribution of characters among the six studied families of Eunicida. The presence of a character is indicated by “+”, while its absence (or reduction) is marked with “-”. abl: accessory buccal lips, at: association tracts, dbl: dorsal buccal lips, ey: eye, la: lateral antenna, lo: lateral organ, ma: median antenna, no: nuchal organ, pa: palp, pr: prostomium, vbl: ventral buccal lips.
| Tree Code | Character | Lumbrineridae | Oenonidae | Onuphidae | Eunicidae | Dorvilleidae | Histriobdellidae |
|---|---|---|---|---|---|---|---|
| 1 | ma | - | - | + | + | - | + |
| 2 | la | - | - | + | + | + | + |
| 3 | pa | - | - | + | + | + | + |
| 4 | vbl | - | - | + | + | - | - |
| 5 | dbl | - | - | + | - | - | - |
| 6 | abl | - | - | + | - | - | - |
| 7 | ey | - | + 2 pairs | + 1 pair | + 1 pair | + 2 pairs | - |
| 8 | no | + | + | + | + | + 2 pairs | + |
| 9 | shape no | unpaired, x-shaped | paired, branched | unpaired, arc-shaped | unpaired, arc-shaped | paired, simple | paired, simple |
| 10 | lo | + | + | + | + | + | + |
| 11 | at | - | - | + | + | - | ? |
| 12 | shape pr | elongated, tapered | elongated, tapered | roundish | roundish | roundish | roundish |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kuhl, S.; Bartolomaeus, T.; Beckers, P. How Do Prostomial Sensory Organs Affect Brain Anatomy? Phylogenetic Implications in Eunicida (Annelida). J. Mar. Sci. Eng. 2022, 10, 1707. https://doi.org/10.3390/jmse10111707
AMA Style
Kuhl S, Bartolomaeus T, Beckers P. How Do Prostomial Sensory Organs Affect Brain Anatomy? Phylogenetic Implications in Eunicida (Annelida). Journal of Marine Science and Engineering. 2022; 10(11):1707. https://doi.org/10.3390/jmse10111707
Chicago/Turabian StyleKuhl, Sabrina, Thomas Bartolomaeus, and Patrick Beckers. 2022. "How Do Prostomial Sensory Organs Affect Brain Anatomy? Phylogenetic Implications in Eunicida (Annelida)" Journal of Marine Science and Engineering 10, no. 11: 1707. https://doi.org/10.3390/jmse10111707
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.