
A Review on Genus Halichondria (Demospongiae, Porifera)


1
Marine Biological Research Centre, Department of Biology, University of Southern Denmark, 5230 Odense M, Denmark
2
Genetics, Ecology, and Evolution, Department of Biology, Aarhus University, 8000 Aarhus C, Denmark
*
Author to whom correspondence should be addressed.
J. Mar. Sci. Eng. 2022, 10(9), 1312; https://doi.org/10.3390/jmse10091312
Submission received: 14 August 2022
/
Revised: 5 September 2022
/
Accepted: 13 September 2022
/
Published: 16 September 2022
(This article belongs to the Special Issue Filter-Feeding in Marine Invertebrates)
Abstract
:Demosponges of the genus Halichondria Fleming (1828) are common in coastal marine ecosystems worldwide and have been well-studied over the last decades. As ecologically important filter feeders, Halichondria species represent potentially suitable model organisms to link and fill in existing knowledge gaps in sponge biology, providing important novel insights into the physiology and evolution of the sponge holobiont. Here we review studies on the morphology, taxonomy, geographic distribution, associated fauna, life history, hydrodynamic characteristics, and coordinated behavior of Halichondria species.
1. Introduction
The genus Halichondria Fleming (1828) [1] (Demospongiae, Porifera; subgenera Halichondria and Eumastia) contains the most common marine sponge species of the North Atlantic [2], including the common “bread-crumb” sponge Halichondria (Halichondria) panicea Pallas (1766) [3] and Bowerbank’s horny sponge H. bowerbanki Burton (1930) [4]. The most studied species, H. panicea, occurs in habitats covering a broad range of salinities, temperatures, turbidities, and flow conditions [5,6] and has been recorded in marine intertidal and sublittoral zones down to depths of more than 500 m [2]. Halichondria panicea provides substrate for many other marine organisms, including a large and varied associated fauna [7,8,9], symbiotic algae [10,11], and numerous bacteria [12,13]. The life histories of Halichondria spp. are characterized by different modes of asexual and sexual reproduction [14], with the latter revealing strong species- and habitat-specific adaptations [15,16,17,18]. Halichondria sponges are filter feeders capable of processing large volumes of seawater (up to six times their own body volume per minute [19]) and efficiently retaining small food particles [20], thus playing a key role in nutrient recycling of coastal marine ecosystems [8]. Modular arrangement of their leuconoid aquiferous systems [21,22] has made it possible to study the hydrodynamic properties of the sponge filter-pump, which may help to shed light on the evolution of complex filter-feeding systems in sponges (cf. [23]). Despite their apparently simple bauplan without a nervous or muscular system, Halichondria spp. show coordinated responses to changing environmental conditions, including phototactic responses of larvae [24], sponge body shape changes [25], and contractile behavior [22,26,27,28]. The detailed mechanisms underlying coordinated behavior in sponges are still unclear [29], but existing data for Halichondria points out the importance of cellular communication based on a neuronal-like ‘toolkit’ and could serve as a milestone towards an improved understanding of tissue organization in the first animals.
The vast majority of studies on Halichondria (a total of 11,100 research articles according to Google scholar) are based on H. panicea (36.4% of total research articles) with a focus on the biological and ecological aspects, whereas much fewer studies within these research fields have addressed other species, such as H. bowerbanki (4.0%), H. melanadocia Laubenfels (1936) [30] (1.5%), H. moorei Bergquist (1961) [31] (1.0%), or H. semitubulosa Lamarck (1814) [32] (0.2%, Table 1).
Other studies have explored the metabolite chemistry of Halichondria, mainly for the species H. okadai Kadota (1922) [34] (26.8%, Table 1), for undefined species (Halichondria sp./spp., 12.5%), or on a genus-level (6.5%), reflecting partially unresolved and still ongoing taxonomic revisions of Halichondria species [35]. Molecular biology, including studies on the sponge microbiome, has mainly been investigated on H. okadai, H. japonica Kadota (1922) [34] (2.3%), H. cylindrata Tanita & Hoshino (1989) [36] (1.6%), and H. oshoro Tanita (1961) [37] (0.7%). Few morphological studies exist for H. melanadocia and H. glabrata Keller (1891) [38] (0.1%), while research on the hydrodynamics of sponges has remained restricted to H. panicea and H. coerulea Berquist (1967) [39] (0.1%). Despite the relevance of comparative studies on sponge cell biology, most Halichondria species have remained understudied (2.1%, Table 1). The aim here is to provide a compilation of studies concerning sponges in the genus Halichondria and point out existing knowledge gaps that may aid in future studies of these ecologically important demosponges.
2. Morphology, Taxonomy, and Distribution
The genus Halichondria is placed in the animal phylum Porifera, class Demospongiae, subclass Heteroscleromorpha, order Suberitida, and family Halichondriidae. Growth forms of Halichondria species include encrusting, massive, occasionally irregularly branching, or digitate sponges with smooth or papillate surfaces. An important morphological character to separate the two subgenera, Halichondria and Eumastia, is the absence or presence of short conical papillae on the sponge surface, respectively [2]. Members of the genus Halichondria typically form chimneys of variable size (up to 5 cm high) with conspicuous, relatively large oscula (2–4 mm in diameter). They are characterized by their firm but compressible texture and variable color, from olive-green (due to symbiotic algae) over orange-yellow to creamy-yellow [2] (cf. Appendix A, Figure A1). The siliceous spicule skeleton of Halichondria consists exclusively of oxeas or oxea derivates in a wide size range, which are arranged in an ectosomal crust (200–300 µm thick) and appear scattered or in tight bundles in the choanosome along with spongin fibers [2,40]. While the functional cell morphology and number of cell types in Halichondria has remained largely unknown, 18 distinct cell types which comprise four major cell families, including contractile, digestive, and amoeboid-neuroid cells, have recently been described in the freshwater demosponge Spongilla lacustris [41].
Species identification is traditionally based on morphological characteristics, such as the shape and structure of the skeleton and the size and form of spicules [42], but several of these characters show strong intra-specific variation and are, therefore, of rather poor quality to distinguish species. For instance, a variety of growth forms are represented by H. panicea, ranging from thin encrusting (Figure A1a) to erect ramose (Figure A1b), which seems to depend on the intensity of ambient water currents [43] (cf. [44]). Moreover, an extensive overlap of spicule sizes in different species has been documented [2]. Molecular data used in phylogenetic studies includes complete mitochondrial genomes of several Halichondria species [45,46,47] and mitochondrial and ribosomal markers [48,49]. The classification of genus Halichondria, as defined in [2], is still in need of a major revision at an ordinal level [35,50], as classification based on morphology disagrees with phylogenetic analyses using molecular data. Overall, morphological, biochemical, and molecular characters applied in recent phylogenetic analyses seem to point out that Halichondria is nonmonophyletic [51,52,53,54].
To date, about 100 Halichondria species are accepted [33,55,56]. They occur in different types of marine habitats around the world, being widespread in European [4,11,57,58], American [2], and Brazilian coastal waters of the Atlantic [59], but also in parts of the Baltic Sea [60], the White Sea [61], and the Mediterranean Sea [62]. Halichondria species also occur in the North Pacific, including Alaska [63,64], Japan [65], Korea [42,66], and the South China Sea [67]. The closely related species H. panicea, H. bowerbanki, and other species in this complex may serve as a suitable model to illuminate possible speciation events due to their overlapping distribution in the North Atlantic, where H. panicea is mainly found in shallow, protected coastal regions of the eastern parts, and shows adaptation to frequent air exposure, while H. bowerbanki is most common in exposed habitats of the western parts, where it tolerates high levels of siltation [11]. A molecular study based on a part of the mitochondrial marker COI suggests that North East Pacific H. cf. panicea is genetically distant from and forms a sister group to a species complex consisting of European H. panicea and H. bowerbanki [53]. Halichondria panicea has also been reported from the Tropical Southwestern Atlantic, along with other species such as H. magniconulosa Hechtel (1965) [68], H. cebimarensis, H. tenebrica, H. migottea, H. sulfurea Carvalho & Hajdu (2001) [59] and H. marianae Santos et al. (2018) [69]. Common species in the Pacific Ocean are H. japonica [65], H. okadai, H. oshoro [70], H. gageoenesis and H. muanensis Kang & Sim (2008) [42], while H. panicea and H. bowerbanki have been reported from Alaska [63,64] and Korea [66], respectively. Revisions of the classification system should include more molecular data and more species and be used to reevaluate the morphological characters used in the traditional classification [50] (cf. [53,54]).
3. The Holobiont Halichondria
Halichondria spp. occur on a variety of inorganic and organic hard substrates, including mussel banks, small stones and rocks, and macroalgae [8,9,43,71]. The sponges themselves provide habitat for a diverse associated fauna and various symbiotic microorganisms. The associated epi- and endofauna of H. panicea include various Arthropoda such as skeleton shrimps (Caprella spp.) and copepods, but also molluscs, e.g., the scallop Chlamys varia, annelids, platyhelminths, and demersal fish that prey almost exclusively upon sponge epifauna [7,8,9,10]. Symbiosis with the dinoflagellate Prorocentrum lima has been documented in H. okadai [72,73], and H. panicea seems to harbor (intracellular) green algae [10,11]. However, many Halichondria species have not been investigated, indicating numerous other yet undiscovered symbiotic interactions, e.g., with dinoflagellates, cryptophytes, microalgae, and diatoms [73]. While the growth of pathogenic bacteria on H. panicea can cause sponge mortality under stagnant flow conditions [74], sponges harbor diverse microbial assemblages that contribute positively to host metabolism and defense [12,75,76]. Halichondria spp. are characterized as low microbial abundance (LMA) sponges with high variability in their bacterial diversity across species and environments [12,13,76]. While only 7 operational taxonomic units (OTUs) of microorganisms have been identified in H. okadai from Korea [77], about 500 OTUs were detected in H. panicea and H. (Eumastia) sitiens Schmidt (1870) [78] from the White Sea [76], respectively, and 1779 OTUs seem to be unique to H. bowerbanki from the mid-Atlantic region of the eastern United States [13]. The microbiome of H. panicea is dominated by a core taxon of Alphaproteobacteria within the class Amylibacter which has recently been named ‘Candidatus Halichondribacter symbioticus’ [12,76,79,80,81,82]. Transmission of bacterial symbionts occurs in a mixed vertical (i.e., direct through reproduction) and horizontal mode (i.e., indirect through the environment) in H. bowerbanki; it is likely to vary across Halichondria species [13]. Metagenomics have revealed that distinct viromes with low similarity to known viral sequences are associated with H. panicea and H. sitiens, suggesting the existence of bacterial antiphage systems in sponges [76].
Halichondria sponges and their microbial symbionts produce a broad spectrum of mainly symbiont-derived bioactive metabolites [83] with cytotoxic or cell growth-inhibiting properties. Substances isolated from Halichondria sponges include halichondrin B and okadaic acid in H. okadai [72,84,85] or gymnostatins and dankastatins from an H. japonica-derived fungal strain [86] which may additionally serve Halichondria sponges as a defense mechanism against pathogens, predators, and biofouling [73,87]. Okadaic acid is a biotoxin known for its cyto-, neuro-, immune-, embryo-, and genotoxicity in marine animals [87,88,89] and has been suggested to protect the demosponge Suberites domuncula from bacterial and parasitic infections [87]. Epibiotic H. panicea can negatively affect the heart performance of blue mussels (Mytilus edulis), which may be due to the sponges’ release of excretory/secretory products. Such substances with cytotoxic properties and antimicrobial activity seem to benefit H. panicea in the competition for space and food across benthic fouling communities [90]. Neuroactive bacteria-derived compounds in H. panicea [73] suggest the relevance of symbiotic interactions for essential physiological processes such as coordinated behavior. The natural variability of sponge-microbe associations in Halichondria seems to provide a meaningful framework for modeling symbiotic interactions in metazoans (cf. [91]). In H. bowerbanki, for instance, changes in microbial communities after exposure to thermal stress have been documented [92], pointing out the relevance of future studies on sponges for assessing possible shifts in symbiont community composition and structure in response to global warming.
4. Life History
The life histories of Halichondria species typically include a reproductive period of 2–3 months in temperate regions [15,71,93]. Halichondria spp. are ovoviviparous and characterized by asynchronous gameto- and embryogenesis, while habitat-specific differences include successive hermaphroditism in White Sea populations of H. panicea and H. sitiens [18], simultaneous hermaphroditism in H. panicea and H. bowerbanki from the southwest coast of the Netherlands [16], incomplete gonochorism in Halichondria sp. from Mystic Estuary, US [15], or gonochorism in H. panicea from Kiel Bight, Germany [17]. In temperate regions, environmental parameters such as temperature and salinity drive the onset of sexual reproduction in H. panicea [17]. Differentiation of gametes from somatic cells has been observed in both H. panicea and H. semitubulosa, indicating the development of spermatocytes from choanocytes or archaeocytes, a process that may be species-dependent [62,94]. The larvae of Halichondria species are typically of parenchymella type and sometimes contain choanocyte chambers before settlement [24,95]. The release of Halichondria larvae seems to follow a light cue, being triggered by the onset of darkness in the temperate species H. panicea [96], while tropical H. melanadocia release larvae on exposure to light following a period of dark adaptation [24]. Phototactic responses of larvae range from positive to neutral to negative before settlement upon various hard substrates [24] (Figure A2a,b).
The growth of Halichondria sponges is dependent on temperature [70] and the concentration of available food, which mainly consists of bacteria and phytoplankton [97]. Pumping rates of H. panicea increase linearly with temperature and require relatively low energy demands for filtering large volumes of seawater [20,98], as expressed by F/R-ratios ≥15.6 L H2O (mL O2)−1, which are comparable to other filter-feeding marine invertebrates [19]. In contrast, the energetic cost of growth is high in sponges [20,99], with exponential growth at a maximum rate of 4% d−1 in H. panicea under natural conditions [100]. The weight-specific growth of H. panicea is constant over sponge size, which has been pointed out as a unique feature among most other filter-feeding invertebrates, reflecting the modular organization of sponges [100]. A study of H. panicea from the Western Baltic Sea suggested that stored glycogen reserves fueled sexual reproduction and that the sponges degenerated in the end of the following year after reproduction [71]. Tissue regression and high mortality during the colder months of the year have also been reported for temperate Halichondria sp. from the Mystic and Thames estuaries, US [57,101] and for Halichondria bowerbanki from New England, US [102], respectively, while the longevity of H. okadai in Japanese waters may exceed 3 years when considering asexual reproduction, i.e., fission and fusion of sponge fragments [14]. Halichondria panicea is capable of rapid regeneration of damaged parts, as expressed in ≥3-fold increased growth rates in response to predation [103] or during the reorganization of the aquiferous system in explant cuttings within approximately 6–10 days [22] (Figs. A2c-f), while other species, such as H. magniconulosa, seem to regenerate at slower rates [104]. Several Halichondria species, including H. lutea Alcolado (1984) [105], H. magniconulosa, and H. melanadocia have been recognized as important members of the Caribbean mangrove and coral reef communities, where they are preyed upon by fish [106,107]. H. panicea can also serve as a food source for hermit crabs, shrimp, large isopods (e.g., Idothea sp.), or the nudibranch Archidoris montereyensis, which may appear in such high density that it can eliminate large and long-lived sponge populations [63,64]. Halichondria sponges play an important role in nutrient recycling of coastal marine ecosystems due to their unique ability to retain small particles (≤0.1 µm) [20,108]. Regular tissue sloughing has been observed in H. panicea in response to sedimentation of organic material and settlement of small organisms on the sponge surface [109], along with seasonal remineralization of released H. panicea biomass following reproduction [8]. As the water pumping activity of H. panicea leads to an accumulation of pollutants, such as heavy metals, in direct proportion to ambient concentrations, their potential use as biomonitoring organisms has been proposed [40,110].
5. Hydrodynamics
As for other demosponges, the aquiferous system of Halichondria is leuconoid [21,40,111] and characterized by choanocytes organized in small spherical chambers which create a unidirectional flow of ambient water through a complex canal system [112,113]. The aquiferous elements of Halichondria act like a sieve for particles of variable size due to their aperture diameters (Figure A3a). As documented for H. panicea, they include numerous inhalant openings (ostia; 7–32 µm) through which seawater is drawn into incurrent canals (50–200 µm), finer incurrent canal branches (prosodi; 5 µm), and the prosopyles (1–4 µm) of choanocyte chambers (18–35 µm; Figure A3b) [113]. Here, choanocytes retain small food particles ≤0.1 µm [20] on their microvilli collars (Figure A4a). Each choanocyte chamber of H. panicea contains about 40–120 choanocytes at an estimated choanocyte chamber density of 18,000 mm−3 [113]. Water leaves choanocyte chambers through an apopyle (7–17 µm; Figure A4b) via excurrent canals (140–450 µm), which drain into an atrium (2.1 mm) from where the water exits the sponge in an excurrent jet through the osculum (1.2 mm) [113] (but see also [21]).
Each osculum represents a functional unit of aquiferous elements in a certain sponge volume (cf. Figure A2b–d), thus characterizing Halichondria sponges with multiple oscula as an array of several autonomous aquiferous modules [22,114,115]. The pumping rate of each aquiferous module is directly proportional to the density of choanocyte chambers in H. panicea [22], implying constant choanocyte densities for different Halichondria species. However, module size seems to determine the volume-specific pumping rates of H. panicea, which can reach a maximum of 15 mL min−1 (cm3 sponge)−1 in growing modules, as observed in single-osculum explants [26,27] (Figure A2c,d), while the modules in multi-oscula explants seem to pump at a lower maximum rate of 3 mL min−1 (cm3 sponge)−1 [22], probably due to a decrease in choanocyte chamber density with increasing module volume [116]. Based on the volume-specific pumping rate and choanocyte chamber density of H. panicea, the pumping rate per choanocyte chamber in a multi-oscula sponge can be estimated to (3/18,000)/60 = 2.78 × 10−6 mm3 s−1 = 2778 µm−3 s−1, and thus the pumping rate per choanocyte at an average of 80 choanocytes per chamber [113], to (2778/80 = 35 µm3 s−1). This value is in range with a previous estimate of (4.46 × 10−6 mm3 s−1/95 = 47 µm3 s−1) for the demosponge Haliclona permollis [113,117] (their Table 1, respectively). A recent hydrodynamic model on the pump characteristics of leuconoid sponges assumed the presence of flagellar vanes along with a glycocalyx mesh which distally connects the microvilli collars of choanocytes, as has been shown for the freshwater sponge Spongilla lacustris [118,119], in order to deliver observed pump pressures [23]. These ultra-structural features of choanocytes have so far not been documented in Halichondria (cf. Figure A4a), pointing out the need for further studies on ultrastructure and hydrodynamic properties, which may provide valuable insight into the evolution of demosponge filter-pump systems (cf. [120]).
6. Coordinated Behavior
At least three different basic cell types are found in Halichondria species, including choanocytes, pinacocytes, and amoeboid (mesohyl) cells [24,121]. The coordinated behavior of sponge cells mediates the hydrodynamic and physical properties of the aquiferous system required for efficient filter feeding under different environmental conditions. Communication between motile cells is the basic principle underlying continuous tissue reorganization, regeneration, and microscale movements in sponges [122,123,124,125]—a topic which has, unfortunately, so far only been addressed by a few studies on Halichondria spp. Continuous tissue remodeling in H. panicea, as expressed by fusion, shape changes, and movement of sponges, has been observed in aquaria and intertidal rocky pools [25]. Halichondria japonica explants have been shown to fuse with explants of the same sponge, while they reject cells from other H. japonica sponges or from H. okadai [126]. Several types of mesohyl cells seem to be involved in this process of “self and nonself” recognition in H. japonica, including amoeboid archaeocytes, motile (granule-rich) gray cells, and collencytes [126]. Recent work on H. panicea points out the importance of cellular transport for the removal of inedible particles from the aquiferous system [27]. At the same time, sponge sandwich cultures may provide a suitable method (Figure A2e,f) for studying the cell types and mechanisms mediating capture, transport, and digestion/removal of edible and inedible particles (Figure A5).
Coordinated behavior further includes contraction of various parts of the aquiferous system, including the osculum [26], in- and excurrent canals, ostia and apopylar openings of the choanocyte chambers, resulting in reduction and temporal shut down of the water flow through single-osculum explants of H. panicea [27,28]. Contractile behavior is common among sponges and seems to follow species-specific cycles of distinct frequency and intensity [127,128,129,130,131] which can be expressed in asynchronous patterns across conspecifics in H. panicea [28,132]. Contractions can occur spontaneously in H. panicea explants under undisturbed conditions in the laboratory and can be induced by chemical messengers (γ-aminobutyric acid and L-glutamate) or by mechanical stimulation with inedible particles [28]. Coordinated contractions of different aquiferous modules in H. panicea explants with multiple oscula have been observed in response to external stimuli [22]. Peristaltic-like waves of contraction travel through the sponge, resulting in osculum closure at speeds of up to 233 nm s−1 in H. panicea (15 °C) [28]. Comparatively, observed contraction speeds of up to 12 µm s−1 in the marine demosponge Tethya wilhema (26 °C) [129] and 122 µm s−1 in the freshwater demosponge Ephydatia muelleri (21 °C) [131] seem considerably higher, emphasizing the need for future studies on the contraction kinetics of Halichondria species. During contractions, H. panicea shows reduced pumping activity [19,26,27], an associated decrease in respiration rates [132], and local internal oxygen depletion [133]. These physiological changes have been suggested as adaptations to variable environmental conditions, including food limitation [134], resuspension of sediment during storm events [135] (cf. [136]), seasonal changes in water temperature, changes in illumination period, spawning events of other sponge species [128], and facilitation of suitable habitat for specific symbiotic microorganisms [132,133]. Contractions may serve Halichondria sponges as an important mechanism to protect the sponge filter-pump in distinct aquiferous modules from clogging and damage and seem to be mediated by exo- and endopinacocytes [22,27,28,134,137], while the underlying cellular pathways have remained unclear. Previous studies have described contractile epithelial cells in sponges that function based on a ‘toolkit’ of neuronal-like elements, including sensory cilia, conduction pathways, and signaling molecules [41,134,138,139,140]. The pinacocytes of other demosponges exhibit actomyosin-based contractility [41,130,137,139,141,142], and myosin type II has been isolated from cells of H. okadai [143].
It is likely that communication between sponge cells in Halichondria is based on the extracellular spreading of chemical messengers [41,123,144], neuronal-like receptors [145], and cell contacts via cellular processes/membrane junctions [146,147,148]. As the abovementioned examples emphasize, cellular communication pathways require further attention in future studies. More detailed information on the functional cell morphology of Halichondria, as can now be accessed using whole-body single-cell RNA sequencing (cf. [41]), is needed to shed light on the principles underlying coordinated behavior in sponges. We encourage future work on the LMA demosponge H. panicea as a model organism to revisit functional coordination pathways with an integral perspective on the underlying morphological structures combining molecular, cytological, and physiological techniques.
7. Conclusions
Halichondria sponges are well-studied and the literature represents a strong base for our present understanding of the ecology and physiology of demosponges. Previous work has mainly focused on H. panicea, paving the foundations for modeling sponge-microbe interactions, hydrodynamics of the sponge filter pump, and cell communication in demosponges. We encourage future research to fill in present knowledge gaps regarding the functional cell morphology and filter-pump characteristics of H. panicea, along with comparative studies including other Halichondria species, to improve and verify existing models based on this ubiquitous demosponge genus.
Author Contributions
Conceptualization, J.G. and P.F.; writing—original draft preparation, J.G.; writing—review & editing, P.F.; visualization, J.G.; project administration, P.F.; funding acquisition, P.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Independent Research Fund, grant number 8021-00392B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to three anonymous reviewers for providing valuable feedback on the manuscript. We further thank Héloïse Hamel and Janni Magelund Degn Larsen for supplementary photographs. Stereo-, light and scanning electron microscopy (SEM) images were acquired at the Marine Biological Research Centre, Kerteminde, University of Southern Denmark, and at the Interdisciplinary Nanoscience Center (iNANO), Aarhus University, Denmark.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
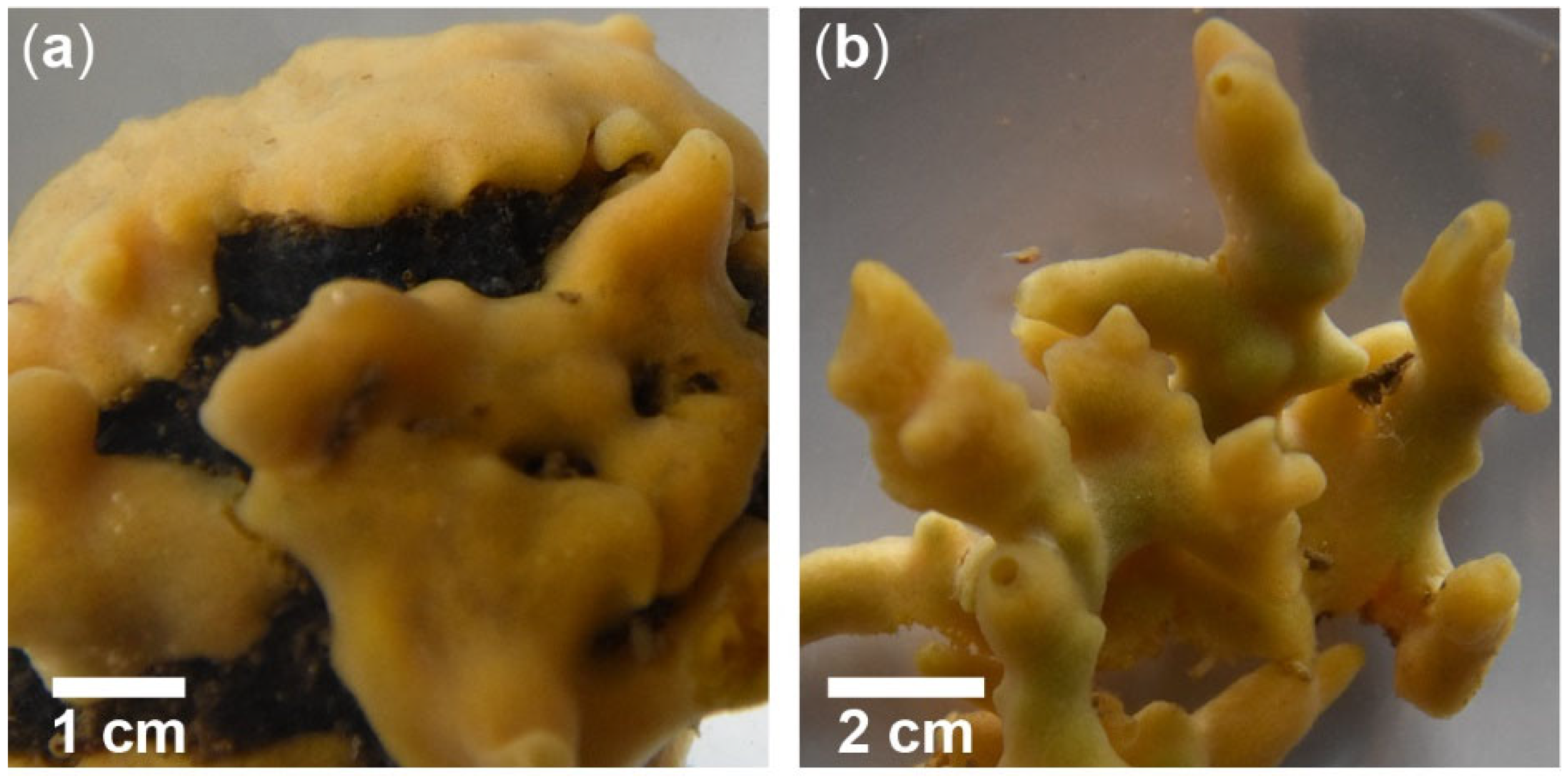
Figure A1.
Growth forms of Halichondria panicea Pallas (1766) in the inlet to Kerteminde Fjord, Denmark (55°26′59″ N, 10°39′41″ E). (a) Growing on a piece of rope, collected in November 2020 and (b) with finger-shaped chimneys, found on wood in November 2020. Pictures: Héloïse Hamel.
Figure A1.
Growth forms of Halichondria panicea Pallas (1766) in the inlet to Kerteminde Fjord, Denmark (55°26′59″ N, 10°39′41″ E). (a) Growing on a piece of rope, collected in November 2020 and (b) with finger-shaped chimneys, found on wood in November 2020. Pictures: Héloïse Hamel.

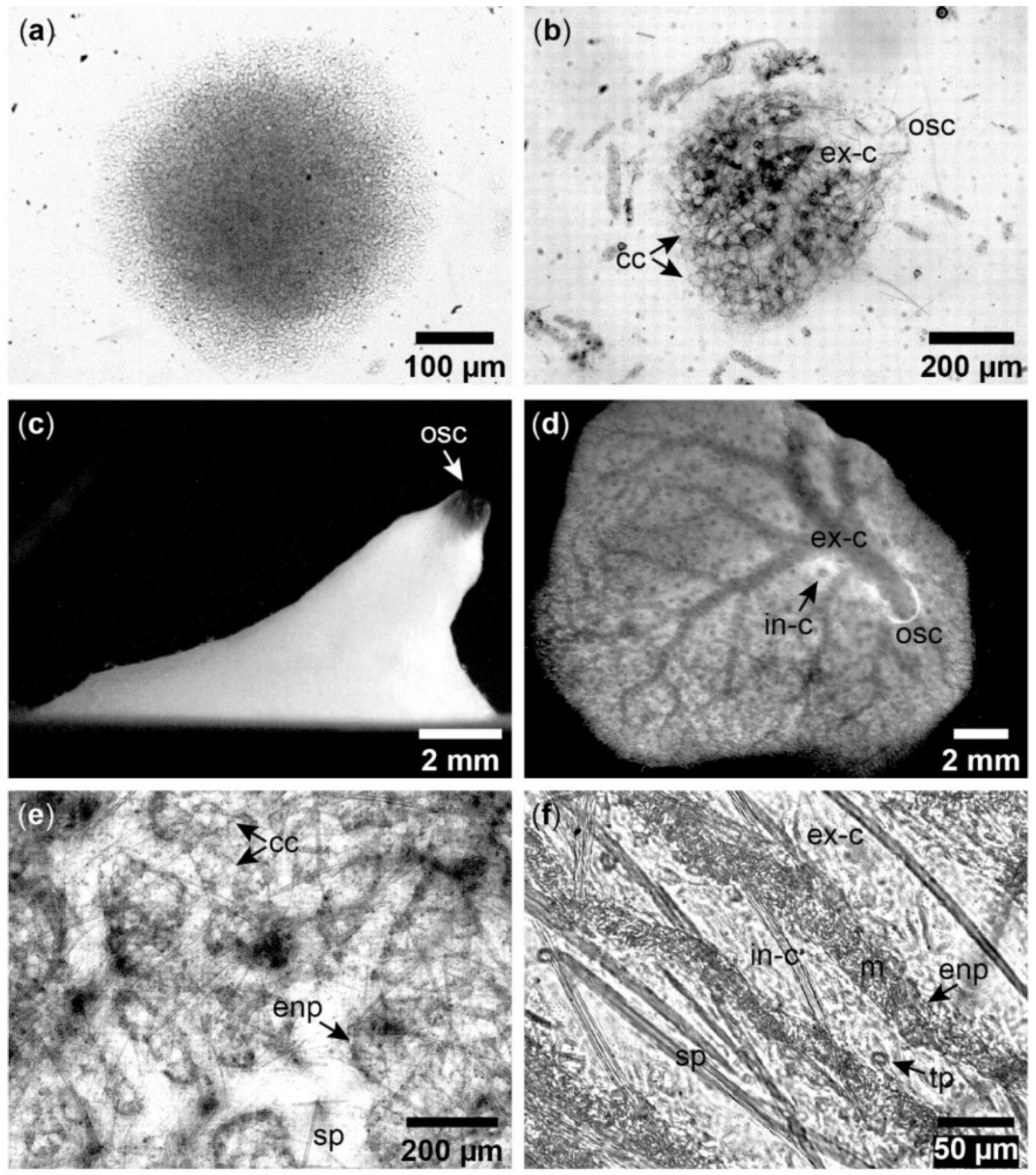
Figure A2.
Aquiferous module formation in Halichondria panicea. (a) Sponge cells after larval settlement, (b) development of choanocyte chambers (cc), excurrent canals (ex-c) and an osculum (osc) in a juvenile sponge, (c) single-osculum explant (side-view), (d) explant (top-view) with visible incurrent (in-c) and excurrent canals (ex-c), (e) sandwich culture with choanocyte chambers (cc), spicules (sp), and endopinacoderm (enp) lining aquiferous canals, (f) sandwich culture after addition of edible particles (tp) for tracing water flow in the incurrent canal (in-c) which is separated from the flow in the excurrent canal (ex-c) by endopinacocytes (enp) and mesohyl (m).
Figure A2.
Aquiferous module formation in Halichondria panicea. (a) Sponge cells after larval settlement, (b) development of choanocyte chambers (cc), excurrent canals (ex-c) and an osculum (osc) in a juvenile sponge, (c) single-osculum explant (side-view), (d) explant (top-view) with visible incurrent (in-c) and excurrent canals (ex-c), (e) sandwich culture with choanocyte chambers (cc), spicules (sp), and endopinacoderm (enp) lining aquiferous canals, (f) sandwich culture after addition of edible particles (tp) for tracing water flow in the incurrent canal (in-c) which is separated from the flow in the excurrent canal (ex-c) by endopinacocytes (enp) and mesohyl (m).

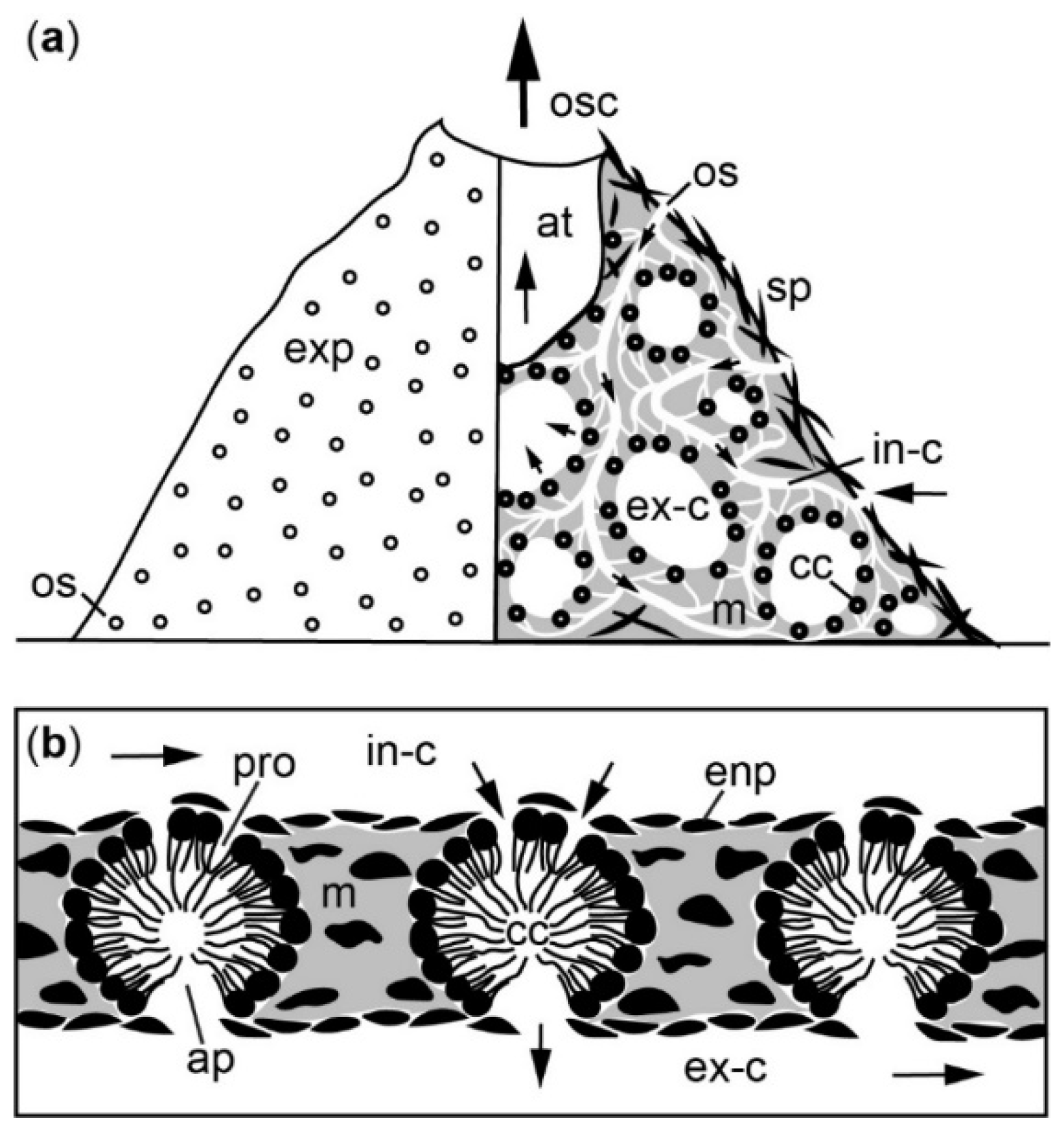
Figure A3.
Schematic illustration of the aquiferous system in a functional module of Halichondria panicea. (a) Left: external surface with ostia (open circles), right: canal system with choanocyte chambers (black circles) and flow direction towards osculum indicated by arrows (b) water flow (arrows) through choanocyte chambers (cf. [111,117], their Figures 9d and 2b, respectively). Abbreviations: exp = exopinacoderm, os = ostium, in-c = incurrent canal, enp = endopinacoderm, pro = prosopyle, cc = choanocyte chamber, ap = apopyle, m = mesohyl, sp = spicule, ex-c = excurrent canal, at = atrium, osc = osculum.
Figure A3.
Schematic illustration of the aquiferous system in a functional module of Halichondria panicea. (a) Left: external surface with ostia (open circles), right: canal system with choanocyte chambers (black circles) and flow direction towards osculum indicated by arrows (b) water flow (arrows) through choanocyte chambers (cf. [111,117], their Figures 9d and 2b, respectively). Abbreviations: exp = exopinacoderm, os = ostium, in-c = incurrent canal, enp = endopinacoderm, pro = prosopyle, cc = choanocyte chamber, ap = apopyle, m = mesohyl, sp = spicule, ex-c = excurrent canal, at = atrium, osc = osculum.

Figure A4.
Halichondria panicea. SEM of cryo-fractured explants. (a) Choanocyte chamber with choanocytes (c) and their microvilli collars (mv) surrounding the flagellum (fl), (b) the fracture shows components of the aquiferous system with prosopyles (pro) and apopyles (ap) connected to incurrent (in-c) and excurrent canals (ex-c), respectively, embedded in mesohyl (m) with choanocyte chambers (cc) and spicules (sp).
Figure A4.
Halichondria panicea. SEM of cryo-fractured explants. (a) Choanocyte chamber with choanocytes (c) and their microvilli collars (mv) surrounding the flagellum (fl), (b) the fracture shows components of the aquiferous system with prosopyles (pro) and apopyles (ap) connected to incurrent (in-c) and excurrent canals (ex-c), respectively, embedded in mesohyl (m) with choanocyte chambers (cc) and spicules (sp).

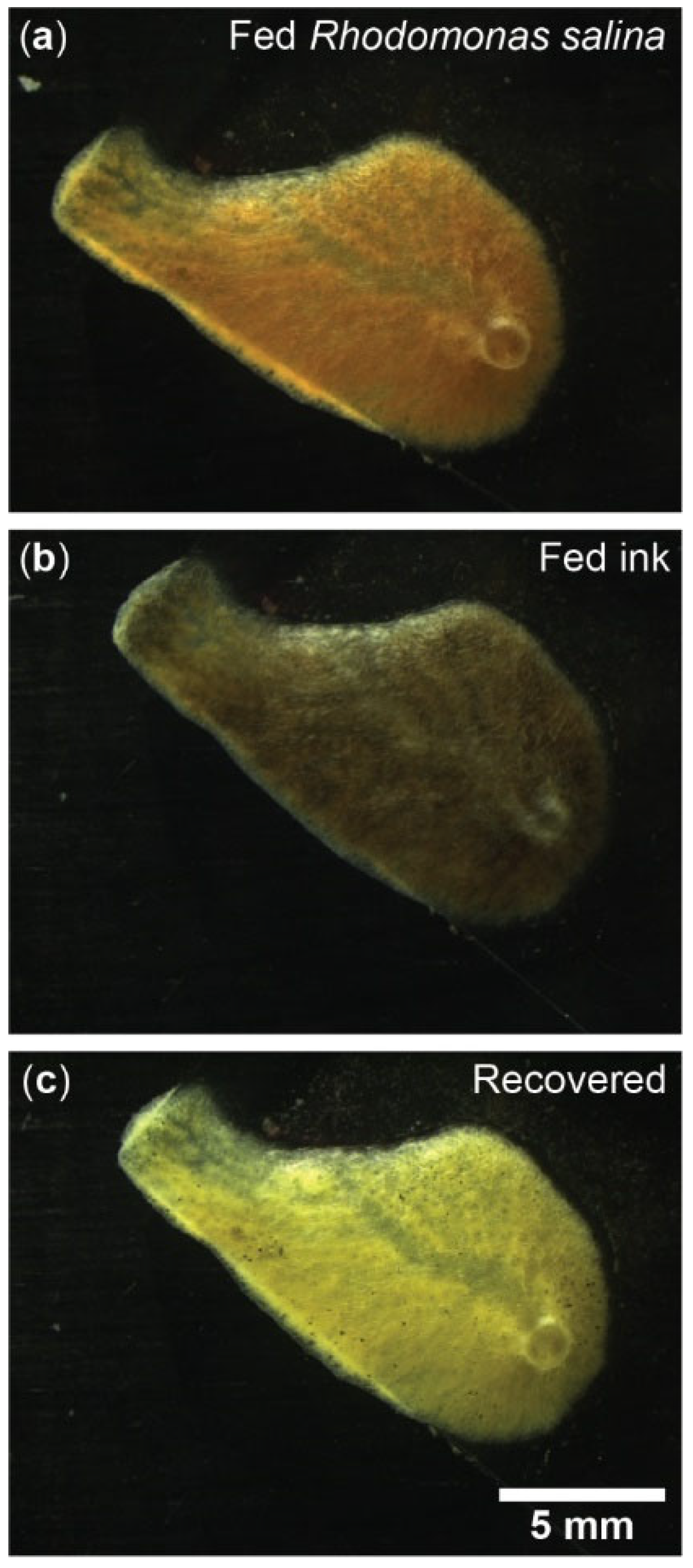
Figure A5.
Exposure of Halichondria panicea to different particle types. Single-osculum explant (top-view) after (a) feeding on Rhodomonas salina (Cryptophyceae); note the red color originating from added algae, (b) exposure to inedible ink (Pelikan Scribtol, 2 × 104-fold diluted) for 1 h; note black color, and (c) recovery in particle-free seawater for 24 h. Pictures: Janni Magelund Degn Larsen.
Figure A5.
Exposure of Halichondria panicea to different particle types. Single-osculum explant (top-view) after (a) feeding on Rhodomonas salina (Cryptophyceae); note the red color originating from added algae, (b) exposure to inedible ink (Pelikan Scribtol, 2 × 104-fold diluted) for 1 h; note black color, and (c) recovery in particle-free seawater for 24 h. Pictures: Janni Magelund Degn Larsen.

References
- Fleming, J. A History of British Animals: Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca, and Radiata of the United Kingdom, Including the Indigenous, Extirpated, and Extinct Kinds, Together with Periodical and Occasional Visitors; Bell & Bradfute: London, UK, 1828; pp. 506–524. [Google Scholar]
- Erpenbeck, D.; Van Soest, R.W. Family Halichondriidae Gray, 1867. In Systema Porifera, 1st ed.; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; pp. 787–815. [Google Scholar]
- Pallas, P.S. Elenchus Zoophytorum Sistens Generum Adumbrationes Generaliores et Specierum Cognitarum Succintas Descriptiones, Cum Selectis Auctorum Synonymis; Fransiscum Varrentrapp: The Hague, The Netherlands, 1766; p. 451. [Google Scholar]
- Burton, M. Additions to the sponge fauna at Plymouth. J. Mar. Biolog. Assoc. U. K. 1930, 16, 489–508. [Google Scholar] [CrossRef]
- Hiscock, K.; Jones, H. Halichondria (Halichondria) bowerbanki Bowerbank’s halichondria. In Marine Life Information Network: Biology and Sensitivity Key Information Reviews; Tyler-Walters, H., Hiscock, K., Eds.; Marine Biological Association of the United Kingdom: Plymouth, UK, 2007; pp. 1–14. [Google Scholar] [CrossRef]
- Hiscock, K. Halichondria (Halichondria) panicea breadcrumb sponge. In Marine Life Information Network: Biology and Sensitivity Key Information Reviews; Tyler-Walters, H., Hiscock, K., Eds.; Marine Biological Association of the United Kingdom: Plymouth, UK, 2008; pp. 1–16. [Google Scholar] [CrossRef]
- Forester, A.J. The association between the spongeHalichondriapanicea (Pallas) and scallop Chlamys varia (L.): A commensal-protective mutualism. J. Exp. Mar. Biol. Ecol. 1979, 36, 1–10. [Google Scholar] [CrossRef]
- Barthel, D. On the ecophysiology of the spongeHalichondriapanicea in Kiel Bight. 2. Biomass, production, energy budget and integration in environmental processes. Mar. Ecol. Prog. Ser. 1988, 43, 87–93. [Google Scholar] [CrossRef]
- Peattie, M.E.; Hoare, R. The sublittoral ecology of the Menai Strait: II. The spongeHalichondriapanicea (Pallas) and its associated fauna. Estuar. Coast. Shelf Sci. 1981, 13, 621–635. [Google Scholar] [CrossRef]
- Long, E.R. The associates of four species of marine sponges of Oregon and Washington. Pac. Sci. 1968, 22, 347–351. [Google Scholar]
- Vethaak, A.D.; Cronie, R.J.A.; Van Soest, R.W.M. Ecology and distribution of two sympatric, closely related sponge species,Halichondriapanicea (Pallas, 1766) and H. bowerbanki Burton, 1930 (Porifera, Demospongiae), with remarks on their speciation. Bijdr. Dierkd. 1982, 52, 82–102. [Google Scholar] [CrossRef]
- Knobloch, S.; Jóhannsson, R.; Marteinsson, V. Bacterial diversity in the marine spongeHalichondriapanicea from Icelandic waters and host-specificity of its dominant symbiont “Candidatus Halichondribacter symbioticus”. FEMS Microbiol. Ecol. 2019, 95, fiy220. [Google Scholar] [CrossRef]
- Sacristán-Soriano, O.; Winkler, M.; Erwin, P.; Weisz, J.; Harriott, O.; Heussler, G.; Hill, M. Ontogeny of symbiont community structure in two carotenoid-rich, viviparous marine sponges: Comparison of microbiomes and analysis of culturable pigmented heterotrophic bacteria. Environ. Microbiol. Rep. 2019, 11, 249–261. [Google Scholar] [CrossRef]
- Tanaka, K. Growth dynamics and mortality of the intertidal encrusting spongeHalichondriaokadai (Demospongiae, Halichondrida). Mar. Biol. 2002, 140, 383–389. [Google Scholar]
- Fell, P.E.; Jacob, W.F. Reproduction and development ofHalichondriasp. in the Mystic Estuary, Connecticut. Biol. Bull. 1979, 156, 62–75. [Google Scholar] [CrossRef]
- Wapstra, M.; Van Soest, R.W.M. Sexual reproduction, larval morphology and behaviour in demosponges from the southwest of the Netherlands. In Taxonomy of Porifera, 1st ed.; Vacelet, J., Boury-Esnault, N., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 281–307. [Google Scholar]
- Witte, U.; Barthel, D.; Tendal, O. The reproductive cycle of the spongeHalichondriapanicea Pallas (1766) and its relationship to temperature and salinity. J. Exp. Mar. Biol. Ecol. 1994, 183, 41–52. [Google Scholar] [CrossRef]
- Gerasimova, E.I.; Ereskovsky, A.V. Reproduction of two species ofHalichondria(Demospongiae: Halichondriidae) in the White Sea. In Porifera Research—Biodiversity, Innovation and Sustainability. Série Livros, 1st ed.; Custódio, M.R., Ed.; Museu Nacional: Rio de Janeiro, Brazil, 2007; Volume 28, pp. 327–333. [Google Scholar]
- Riisgård, H.U.; Kumala, L.; Charitonidou, K. Using the F/R-ratio for an evaluation of the ability of the demospongeHalichondriapanicea to nourish solely on phytoplankton versus free-living bacteria in the sea. Mar. Biol. Res. 2016, 12, 907–916. [Google Scholar] [CrossRef]
- Thomassen, S.; Riisgård, H.U. Growth and energetics of the spongeHalichondriapanicea. Mar. Ecol. Prog. Ser. 1995, 128, 239–246. [Google Scholar] [CrossRef]
- Vogel, S. Current-induced flow through the sponge, Halichondria. Biol. Bull. 1974, 147, 443–456. [Google Scholar] [CrossRef]
- Kealy, R.A.; Busk, T.; Goldstein, J.; Larsen, P.S.; Riisgård, H.U. Hydrodynamic characteristics of aquiferous modules in the demospongeHalichondriapanicea. Mar. Biol. Res. 2019, 15, 531–540. [Google Scholar] [CrossRef]
- Asadzadeh, S.S.; Larsen, P.S.; Riisgård, H.U.; Walther, J.H. Hydrodynamics of the leucon sponge pump. J. R. Soc. Interface 2019, 16, 20180630. [Google Scholar] [CrossRef]
- Woollacott, R.M. Structure and swimming behavior of the larva ofHalichondriamelanadocia (Porifera: Demospongiae). J. Morphol. 1990, 205, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Burton, M. Observations on littoral sponges, including the supposed swarming of larvae, movement and coalescence in mature individuals, longevity and death. Proc. Zool. Soc. Lond. 1949, 118, 893–915. [Google Scholar] [CrossRef]
- Kumala, L.; Riisgård, H.U.; Canfield, D.E. Osculum dynamics and filtration activity studied in small single-osculum explants of the demospongeHalichondriapanicea. Mar. Ecol. Prog. Ser. 2017, 572, 117–128. [Google Scholar] [CrossRef]
- Goldstein, J.; Riisgård, H.U.; Larsen, P.S. Exhalant jet speed of single-osculum explants of the demospongeHalichondriapanicea and basic properties of the sponge-pump. J. Exp. Mar. Biol. Ecol. 2019, 511, 82–90. [Google Scholar] [CrossRef]
- Goldstein, J.; Bisbo, N.; Funch, P.; Riisgård, H.U. Contraction-expansion and the effects on the aquiferous system in the demospongeHalichondriapanicea. Front. Mar. Sci. 2020, 7, 113. [Google Scholar] [CrossRef]
- Abe, T.; Sahin, F.P.; Akiyama, K.; Naito, T.; Kishigami, M.; Miyamoto, K.; Sakakibara, Y.; Uemura, D. Construction of a metagenomic library for the marine spongeHalichondriaokadai. Biosc. Biotechnol. Biochem. 2012, 76, 633–639. [Google Scholar] [CrossRef]
- Laubenfels, M.W. A discussion of the sponge fauna of the Dry Tortugas in particular and the West Indies in general, with material for a revision of the families and orders of the Porifera. Publ. Carnegie Instit. Wash. 1936, 467, 1–225. [Google Scholar]
- Bergquist, P.R. A collection of Porifera from Northern New Zealand, with descriptions of seventeen new species. Pac. Sci. 1961, 15, 33–48. [Google Scholar]
- Lamarck, J.B. Sur les polypiers empâtés. Ann. Mus. Natl. Hist. Nat. 1814, 20, 294–312. [Google Scholar]
- Van Soest, R.W.; Boury-Esnault, N.; Hooper, J.N.; Rützler, K.; de Voogd, N.J.; de Glasby, B.A.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schoenberg, C.; et al. Halichondria Fleming, 1828. World Porifera Database. World Register of Marine Species 2020. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=131807 (accessed on 6 July 2022).
- Kadota, J. Observations of two new species of the genus Reniera of monaxonid sponges. Zool. Mag. 1922, 34, 700–711. [Google Scholar]
- Erpenbeck, D.; Hall, K.; Alvarez, B.; Büttner, G.; Sacher, K.; Schätzle, S.; Schuster, A.; Vargas, S.; Hooper, J.N.A.; Wörheide, G. The phylogeny of halichondrid demosponges: Past and present re-visited with DNA-barcoding data. Org. Divers. Evol. 2012, 12, 57–70. [Google Scholar] [CrossRef]
- Tanita, S.; Hoshino, T. The Demospongiae of Sagami Bay; Biological Laboratory, Imperial Household: Tokyo, Japan, 1989; p. 197.
- Tanita, S. Two sponges obtained by the Training Ship ‘Oshoro Maru’ in the Eastern Behring Sea. Bull. Fish. Sci. Hokkaido Univ. 1961, 11, 183–187. [Google Scholar]
- Keller, C. Die Spongienfauna des Rothen Meeres (II. Hälfte). Z. Wiss. Zool. 1891, 52, 294–368. [Google Scholar]
- Bergquist, P.R. Additions to the sponge fauna of the Hawaiian Islands. Micronesica 1967, 3, 159–174. [Google Scholar]
- Olesen, T.M.E.; Weeks, J.M. Accumulation of Cd by the marine spongeHalichondriapanicea Pallas: Effects upon filtration rate and its relevance for biomonitoring. Bull. Environ. Contam. Toxicol. 1994, 52, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Musser, J.M.; Schippers, K.J.; Nickel, M.; Mizzon, G.; Kohn, A.B.; Pape, C.; Ronchi, P.; Papadopoulos, N.; Tarashansky, A.J.; Hammel, J.U.; et al. Profiling cellular diversity in sponges informs animal cell type and nervous system evolution. Science 2021, 374, 717–723. [Google Scholar] [CrossRef]
- Kang, D.W.; Sim, C.J. Two new sponges of the genusHalichondria(Halichondrida: Halichondriidae) from Korea. Anim. Cells Syst. 2008, 12, 65–68. [Google Scholar] [CrossRef]
- Barthel, D. Influence of different current regimes on the growth form ofHalichondriapanicea Pallas. In Fossil and Recent Sponges, 1st ed.; Reitner, J., Keupp, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 387–394. [Google Scholar]
- Bell, J.J.; Barnes, D.K. The influences of bathymetry and flow regime upon the morphology of sublittoral sponge communities. J. Mar. Biolog. Assoc. U. K. 2000, 80, 707–718. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Huang, D. The complete mitochondrial genome of spongeHalichondria(Halichondria) sp. (Demospongiae, Suberitida, Halichondriidae). Mitochondrial DNA B Resour. 2016, 1, 512–514. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.J.; Jung, Y.H.; Yu, C.; An, Y.R.; Han, D.; Kang, D.W. The complete mitochondrial genome of spongeHalichondriaokadai (Demospongiae, Suberitida, Halichondriidae) from Korea water. Mitochondrial DNA B Resour. 2017, 2, 873–874. [Google Scholar] [CrossRef]
- Kim, H.; Kang, D.W.; Yu, C.; Jung, Y.H.; Yoon, M.; Kim, H.J. The complete mitochondrial genome of spongeHalichondriasp. (Demospongiae, Suberitida, Halichondriidae) from Dok-do. Mitochondrial DNA B Resour. 2019, 4, 1729–1730. [Google Scholar] [CrossRef] [Green Version]
- Erpenbeck, D.; Breeuwer, J.; van der Velde, H.; van Soest, R. Unravelling host and symbiont phylogenies of halichondrid sponges (Demospongiae, Porifera) using a mitochondrial marker. Mar. Biol. 2002, 141, 377–386. [Google Scholar]
- Erpenbeck, D.; Duran, S.; Rützler, K.; Paul, V.J.; Hooper, J.N.; Wörheide, G. Towards a DNA taxonomy of Caribbean demosponges: A gene tree reconstructed from partial mitochondrial CO1 gene sequences supports previous rDNA phylogenies and provides a new perspective on the systematics of Demospongiae. J. Mar. Biolog. Assoc. U. K. 2007, 87, 1563–1570. [Google Scholar] [CrossRef]
- Morrow, C.C.; Picton, B.E.; Erpenbeck, D.; Boury-Esnault, N.; Maggs, C.A.; Allcock, A.L. Congruence between nuclear and mitochondrial genes in Demospongiae: A new hypothesis for relationships within the G4 clade (Porifera: Demospongiae). Mol. Phylogenetics Evol. 2012, 62, 174–190. [Google Scholar] [CrossRef]
- Alvarez, B.; Crisp, M.D.; Driver, F.; Hooper, J.N.; Van Soest, R.W. Phylogenetic relationships of the family Axinellidae (Porifera: Demospongiae) using morphological and molecular data. Zool. Scr. 2000, 29, 169–198. [Google Scholar] [CrossRef]
- Castellanos, L.; Zea, S.; Osorno, O.; Duque, C. Phylogenetic analysis of the order Halichondrida (Porifera, Demospongiae), using 3β-hydroxysterols as chemical characters. Biochem. Syst. Ecol. 2003, 31, 1163–1183. [Google Scholar] [CrossRef]
- Erpenbeck, D. A molecular comparison of Alaskan and North East AtlanticHalichondriapanicea (Pallas 1766) (Porifera: Demospongiae) populations. Boll. Mus. Ist. Biol. Univ. Genova 2004, 68, 319–325. [Google Scholar]
- Erpenbeck, D.; Breeuwer, J.A.; Van Soest, R.W. Identification, characterization and phylogenetic signal of an elongation factor-1 alpha fragment in demosponges (Metazoa, Porifera, Demospongiae). Zool. Scr. 2005, 34, 437–445. [Google Scholar] [CrossRef]
- Hooper, J.N.A.; van Soest, R.W.M. Systema Porifera. A Guide to the Classification of Sponges; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; Volume 1, pp. 1–1101. [Google Scholar]
- Alvarez, B.; Hooper, J.N. Taxonomic revision of the order Halichondrida (Porifera: Demospongiae) of northern Australia. Family Halichondriidae. Beagle Rec. Mus. Art Galleries North. Territ. 2011, 27, 55–84. [Google Scholar] [CrossRef]
- Fell, P.E.; Lewandrowski, K.B. Population dynamics of the estuarine sponge,Halichondriasp., within a New England eelgrass community. J. Exp. Mar. Biol. Ecol. 1981, 55, 49–63. [Google Scholar] [CrossRef]
- Picton, B.E.; Goodwin, C.E. Sponge biodiversity of Rathlin Island, Northern Ireland. J. Mar. Biolog. Assoc. U. K. 2007, 87, 1441–1458. [Google Scholar] [CrossRef]
- Carvalho, M.D.S.; Hajdu, E. Comments on brazilianHalichondriaFleming (Halichondriidae, Halichondrida, Demospongiae), with the description of four new species from the São Sebastião Channel and its environs (Tropical Southwestern Atlantic). Rev. Bras. Zool. 2001, 18, 161–180. [Google Scholar] [CrossRef]
- Barthel, D. Population dynamics of the spongeHalichondriapanicea (Pallas) in Kiel Bight. In Marine Eutrophication and Population Dynamics: 25th European Marine Biology Symposium; Colombo, G., Ed.; Olsen & Olsen: Fredensborg, Denmark, 1992; pp. 203–209. [Google Scholar]
- Khalaman, V.V.; Komendantov, A.Y. Experimental study of the ability of the spongeHalichondriapanicea (Porifera: Demospongiae) to compete for a substrate in shallow-water fouling communities of the White Sea. Biol. Bull. Russ. Acad. Sci. 2016, 43, 69–74. [Google Scholar] [CrossRef]
- Gaino, E.; Lepore, E.; Rebora, M.; Mercurio, M.; Sciscioli, M. Some steps of spermatogenesis inHalichondriasemitubulosa (Demospongiae, Halichondriidae). Ital. J. Zool. 2007, 74, 117–122. [Google Scholar] [CrossRef]
- Knowlton, A.L.; Highsmith, R.C. Convergence in the time-space continuum: A predator-prey interaction. Mar. Ecol. Prog. Ser. 2000, 197, 285–291. [Google Scholar] [CrossRef]
- Wulff, J. Ecological interactions and the distribution, abundance, and diversity of sponges. Adv. Mar. Biol. 2012, 61, 273–344. [Google Scholar]
- Hoshino, S. Systematic status ofHalichondriajaponica (Kadota) (Demospongiae, Halichondrida) from Japan. Boll. Mus. Ist. Biol. Univ. Genova 2004, 68, 373–379. [Google Scholar]
- Jeon, Y.J.; Sim, C.J. A new record of genusHalichondria(Demospongiae: Halichondrida: Halichondriidae) from Korea. Anim. Syst. Evol. Diversity 2009, 25, 137–139. [Google Scholar] [CrossRef]
- Li, Z.; Hu, Y.; Liu, Y.; Huang, Y.; He, L.; Miao, X. 16S rDNA clone library-based bacterial phylogenetic diversity associated with three South China Sea sponges. World J. Microbiol. Biotechnol. 2007, 23, 1265–1272. [Google Scholar] [CrossRef]
- Hechtel, G.J. A systematic study of the Demospongiae of Port Royal, Jamaica. Bull. Peabody Mus. Nat. Hist. 1965, 20, 1–103. [Google Scholar]
- Santos, G.G.; Nascimento, E.; Pinheiro, U. Halichondriidae Gray, 1867 from the Northeastern Brazil with description of a new species. Zootaxa 2018, 4379, 556–566. [Google Scholar] [CrossRef]
- Kim, H.S.; Parkc, B.J.; Sim, C.J. Marine sponges in South Korea (I). Korean J. Syst. Zool. 1986, 11, 37–47. [Google Scholar]
- Barthel, D. On the ecophysiology of the spongeHalichondriapanicea in Kiel Bight. I. Substrate specificity, growth and reproduction. Mar. Ecol. Prog. Ser. 1986, 32, 291–298. [Google Scholar] [CrossRef]
- Kobayashi, J.; Ishibashi, M. Bioactive metabolites of symbiotic marine microorganisms. Chem. Rev. 1993, 93, 1753–1769. [Google Scholar] [CrossRef]
- Lee, Y.K.; Lee, J.H.; Lee, H.K. Microbial symbiosis in marine sponges. J. Microbiol. 2001, 39, 254–264. [Google Scholar]
- Hummel, H.; Sepers, A.B.; De Wolf, L.; Melissen, F.W. Bacterial growth on the marine spongeHalichondriapanicea induced by reduced waterflow rate. Mar. Ecol. Prog. Ser. 1988, 42, 195–198. [Google Scholar] [CrossRef]
- Hentschel, U.; Hopke, J.; Horn, M.; Friedrich, A.B.; Wagner, M.; Hacker, J.; Moore, B.S. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 2002, 68, 4431–4440. [Google Scholar] [CrossRef]
- Rusanova, A.; Fedorchuk, V.; Toshchakov, S.; Dubiley, S.; Sutormin, D. An interplay between viruses and bacteria associated with the White Sea sponges revealed by metagenomics. Life 2021, 12, 25. [Google Scholar] [CrossRef]
- Jeong, J.B.; Kim, K.H.; Park, J.S. Sponge-specific unknown bacterial groups detected in marine sponges collected from Korea through barcoded pyrosequencing. J. Microbiol. Biotechnol. 2015, 25, 1–10. [Google Scholar] [CrossRef]
- Schmidt, O. Grundzüge einer Spongien-Fauna des atlantischen Gebietes; Wilhelm Engelmann: Leipzig, Germany, 1870; pp. 1–88. [Google Scholar]
- Wichels, A.; Würtz, S.; Döpke, H.; Schütt, C.; Gerdts, G. Bacterial diversity in the breadcrumb spongeHalichondriapanicea (Pallas). FEMS Microbiol. Ecol. 2006, 56, 102–118. [Google Scholar] [CrossRef]
- Steinert, G.; Rohde, S.; Janussen, D.; Blaurock, C.; Schupp, P.J. Host-specific assembly of sponge-associated prokaryotes at high taxonomic ranks. Sci. Rep. 2017, 7, 2542. [Google Scholar] [CrossRef]
- Naim, M.A.; Morillo, J.A.; Sørensen, S.J.; Waleed, A.A.S.; Smidt, H.; Sipkema, D. Host-specific microbial communities in three sympatric North Sea sponges. FEMS Microbiol. Ecol. 2014, 90, 390–403. [Google Scholar] [CrossRef]
- Strehlow, B.W.; Schuster, A.; Francis, W.R.; Canfield, D.E. Metagenomic data forHalichondriapanicea from Illumina and nanopore sequencing and preliminary genome assemblies for the sponge and two microbial symbionts. BMC Res. Notes 2022, 15, 135. [Google Scholar] [CrossRef]
- Sun, J.F.; Wu, Y.; Yang, B.; Liuc, Y. Chemical constituents of marine sponge Halichondria sp. from south China sea. Chem. Nat. Compd. 2015, 51, 975–977. [Google Scholar] [CrossRef]
- Hirata, Y.; Uemura, D. Halichondrins-antitumor polyether macrolides from a marine sponge. Pure Appl. Chem. 1986, 58, 701–710. [Google Scholar] [CrossRef]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; Van Engen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus Halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Amagata, T.; Tanaka, M.; Yamada, T.; Minoura, K.; Numata, A. Gymnastatins and dankastatins, growth inhibitory metabolites of a Gymnascella species from aHalichondriasponge. J. Nat. Prod. 2008, 71, 340–345. [Google Scholar] [CrossRef]
- Prego-Faraldo, M.V.; Valdiglesias, V.; Méndez, J.; Eirín-López, J.M. Okadaic acid meet and greet: An insight into detection methods, response strategies and genotoxic effects in marine invertebrates. Mar. Drugs 2013, 11, 2829–2845. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.L.; Zhao, X.Y.; Ji, L.D.; Xu, J. Okadaic acid (OA): Toxicity, detection and detoxification. Toxicon 2019, 160, 1–7. [Google Scholar] [CrossRef]
- Corriere, M.; Soliño, L.; Costa, P.R. Effects of the marine biotoxins okadaic acid and dinophysistoxins on fish. J. Mar. Sci. Eng. 2021, 9, 293. [Google Scholar] [CrossRef]
- Khalaman, V.V.; Sharov, A.N.; Kholodkevich, S.V.; Komendantov, A.Y.; Kuznetsova, T.V. Influence of the White Sea spongeHalichondriapanicea (Pallas, 1766) on physiological state of the blue mussel Mytilus edulis (Linnaeus, 1758), as evaluated by heart rate characteristics. J. Evol. Biochem. Physiol. 2017, 53, 225–232. [Google Scholar] [CrossRef]
- Pita, L.; Fraune, S.; Hentschel, U. Emerging sponge models of animal-microbe symbioses. Front. Microbiol. 2016, 7, 2102. [Google Scholar] [CrossRef]
- Lemoine, N.; Buell, N.; Hill, A.; Hill, M. Assessing the utility of sponge microbial symbiont communities as models to study global climate change: A case study withHalichondriabowerbanki. In Porifera Research: Biodiversity, Innovation and Sustainability; Série Livros, 1st ed.; Custódio, M.R., Ed.; Museu Nacional: Rio de Janeiro, Brazil, 2007; Volume 28, pp. 419–425. [Google Scholar]
- Frøhlich, H.; Barthel, D. Silica uptake of the marine spongeHalichondriapanicea in Kiel Bight. Mar. Biol. 1997, 128, 115–125. [Google Scholar] [CrossRef]
- Barthel, D.; Detmer, A. The spermatogenesis ofHalichondriapanicea (Porifera, Demospongiae). Zoomorphology 1991, 110, 9–15. [Google Scholar] [CrossRef]
- Sokolova, A.M.; Pozdnyakov, I.R.; Ereskovsky, A.V.; Karpov, S.A. Kinetid structure in larval and adult stages of the demosponges Haliclona aquaeductus (Haplosclerida) andHalichondriapanicea (Suberitida). Zoomorphology 2019, 138, 171–184. [Google Scholar] [CrossRef]
- Amano, S. Larval release in response to a light signal by the intertidal spongeHalichondriapanicea. Biol. Bull. 1986, 171, 371–378. [Google Scholar] [CrossRef]
- Lüskow, F.; Riisgård, H.U.; Solovyeva, V.; Brewer, J.R. Seasonal changes in bacteria and phytoplankton biomass control the condition index of the demospongeHalichondriapanicea in temperate Danish waters. Mar. Ecol. Prog. Ser. 2019, 608, 119–132. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Thomassen, S.; Jakobsen, H.; Weeks, J.M.; Larsen, P.S. Suspension feeding in marine spongesHalichondriapanicea and Haliclona urceolus: Effects of temperature on filtration rate and energy cost of pumping. Mar. Ecol. Prog. Ser. 1993, 96, 177–188. [Google Scholar] [CrossRef]
- Koopmans, M.; Martens, D.; Wijffels, R.H. Growth efficiency and carbon balance for the sponge Haliclona oculata. Mar. Biotechnol. 2010, 12, 340–349. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Larsen, P.S. Actual and model-predicted growth of sponges—with a bioenergetic comparison to other filter-feeders. J. Mar. Sci. Eng. 2022, 10, 607. [Google Scholar] [CrossRef]
- Fell, P.E.; Parry, E.H.; Balsamo, A.M. The life histories of sponges in the Mystic and Thames estuaries (Connecticut), with emphasis on larval settlement and postlarval reproduction. J. Exp. Mar. Biol. Ecol. 1984, 78, 127–141. [Google Scholar] [CrossRef]
- Hartman, W.D. Natural history of the marine sponges of southern New England. Bull. Peabody Mus. Yale 1958, 12, 1–155. [Google Scholar]
- Knowlton, A.L.; Highsmith, R.C. Nudibranch-sponge feeding dynamics: Benefits of symbiont-containing sponge to Archidoris montereyensis (Cooper, 1862) and recovery of nudibranch feeding scars byHalichondriapanicea (Pallas, 1766). J. Exp. Mar. Biol. Ecol. 2005, 327, 36–46. [Google Scholar] [CrossRef]
- Wulff, J. Regeneration of sponges in ecological context: Is regeneration an integral part of life history and morphological strategies? Integr. Comp. Biol. 2010, 50, 494–505. [Google Scholar] [CrossRef]
- Alcolado, P.M. Nuevas especies de esponjas encontradas en Cuba. Poeyana 1984, 271, 1–22. [Google Scholar]
- Wulff, J.L. Parrotfish predation on cryptic sponges of Caribbean coral reefs. Mar. Biol. 1997, 129, 41–52. [Google Scholar] [CrossRef]
- Wulff, J.L. Sponge predators may determine differences in sponge fauna between two sets of mangrove cays, Belize barrier reef. Atoll Res. Bull. 2000, 477, 251–263. [Google Scholar] [CrossRef]
- De Goeij, J.M.; Van Oevelen, D.; Vermeij, M.J.; Osinga, R.; Middelburg, J.J.; De Goeij, A.F.; Admiraal, W. Surviving in a marine desert: The sponge loop retains resources within coral reefs. Science 2013, 342, 108–110. [Google Scholar] [CrossRef]
- Barthel, D.; Wolfrath, B. Tissue sloughing in the spongeHalichondriapanicea: A fouling organism prevents being fouled. Oecologia 1989, 78, 357–360. [Google Scholar] [CrossRef]
- Hansen, I.V.; Weeks, J.M.; Depledge, M.H. Accumulation of copper, zinc, cadmium and chromium by the marine spongeHalichondriapanicea Pallas and the implications for biomonitoring. Mar. Pollut. Bull. 1995, 31, 133–138. [Google Scholar] [CrossRef]
- Langenbruch, P.F.; Scalera-Liaci, L. Structure of Choanocyte Chambers in Haplosclerid Sponges; Smithsonian Institution Press: Washington, DC, USA; Woods Hole, MA, USA, 1985; pp. 245–251. [Google Scholar]
- Haeckel, E. XXVII. On the Calcispongiae, their position in the animal kingdom, and their relation to the theory of descendence. J. Nat. Hist. 1873, 11, 241–262. [Google Scholar]
- Reiswig, H.M. The aquiferous systems of three marine Demospongiae. J. Morphol. 1975, 145, 493–502. [Google Scholar] [CrossRef]
- Fry, W.G. The sponge as a population: A biometric approach. Symp. Zool. Soc. Lond. 1970, 25, 135–162. [Google Scholar]
- Fry, W.G. Taxonomy, the individual and the sponge. In Biology and Systematics of Colonial Organisms: Proceedings of an International Symposium Held at the University of Durham; Published for the Systematics Association; Academic Press: London, UK, 1979; Volume 11, pp. 39–80. [Google Scholar]
- Riisgård, H.U.; Larsen, P.S. Filtration rates and scaling in demosponges. J. Mar. Sci. Eng. 2022, 10, 643. [Google Scholar] [CrossRef]
- Larsen, P.S.; Riisgård, H.U. The sponge pump. J. Theor. Biol. 1994, 168, 53–63. [Google Scholar] [CrossRef]
- Weissenfels, N. The filtration apparatus for food collection in freshwater sponges (Porifera, Spongillidae). Zoomorphology 1992, 112, 51–55. [Google Scholar] [CrossRef]
- Mah, J.L.; Christensen-Dalsgaard, K.K.; Leys, S.P. Choanoflagellate and choanocyte collar-flagellar systems and the assumption of homology. Evol. Dev. 2014, 16, 25–37. [Google Scholar] [CrossRef]
- Suarez, P.A.; Leys, S.P. The sponge pump as a morphological character in the fossil record. Paleobiology 2022, 48, 446–461. [Google Scholar] [CrossRef]
- Evans, C.W. The ultrastructure of larvae from the marine spongeHalichondriamoorei Bergquist (Porifera, Demospongiae). Cah. Biol. Mar. 1997, 18, 427–433. [Google Scholar]
- Harris, A.K. Cell motility and the problem of anatomical homeostasis. J. Cell. Sci. 1987, 8, 121–140. [Google Scholar] [CrossRef]
- Gaino, E.; Burlando, B. Sponge cell motility: A model system for the study of morphogenetic processes. Ital. J. Zool. 1990, 57, 109–118. [Google Scholar] [CrossRef]
- Bond, C. Continuous cell movements rearrange anatomical structures in intact sponges. J. Exp. Zool. 1992, 263, 284–302. [Google Scholar] [CrossRef]
- Lavrov, A.I.; Kosevich, I.A. Sponge cell reaggregation: Cellular structure and morphogenetic potencies of multicellular aggregates. J. Exp. Zool. A. Ecol. Genet. Physiol. 2016, 325, 158–177. [Google Scholar] [CrossRef]
- Saito, Y. Self and nonself recognition in a marine sponge,Halichondriajaponica (Demospongiae). Zool. Sci. 2013, 30, 651–657. [Google Scholar] [CrossRef]
- Storr, J.F. Field observations of sponge reactions as related to their ecology. In Aspects of Sponge Biology; Harrison, F.W., Cowden, R.R., Eds.; Academic Press Inc.: New York, NY, USA, 1976; pp. 277–282. [Google Scholar]
- Reiswig, H.M. In situ pumping activities of tropical Demospongiae. Mar. Biol. 1971, 9, 38–50. [Google Scholar] [CrossRef]
- Nickel, M. Kinetics and rhythm of body contractions in the sponge Tethya wilhelma (Porifera: Demospongiae). J. Exp. Biol. 2004, 207, 4515–4524. [Google Scholar] [CrossRef]
- Nickel, M.; Scheer, C.; Hammel, J.U.; Herzen, J.; Beckmann, F. The contractile sponge epithelium sensu lato—Body contraction of the demosponge Tethya wilhelma is mediated by the pinacoderm. J. Exp. Biol. 2011, 214, 1692–1698. [Google Scholar] [CrossRef]
- Elliott, G.R.; Leys, S.P. Coordinated contractions effectively expel water from the aquiferous system of a freshwater sponge. J. Exp. Biol. 2007, 210, 3736–3748. [Google Scholar] [CrossRef]
- Kumala, L.; Canfield, D.E. Contraction dynamics and respiration of small single-osculum explants of the demospongeHalichondriapanicea. Front. Mar. Sci. 2018, 5, 410. [Google Scholar] [CrossRef]
- Kumala, L.; Larsen, M.; Glud, R.N.; Canfield, D.E. Spatial and temporal anoxia in single-osculumHalichondriapanicea demosponge explants studied with planar optodes. Mar. Biol. 2021, 168, 173. [Google Scholar] [CrossRef]
- Leys, S.P. Elements of a ‘nervous system’ in sponges. J. Exp. Biol. 2015, 218, 581–591. [Google Scholar] [CrossRef]
- Gerrodette, T.; Flechsig, A.O. Sediment-induced reduction in the pumping rate of the tropical sponge Verongia lacunosa. Mar. Biol. 1979, 55, 103–110. [Google Scholar] [CrossRef]
- Bell, J.J.; McGrath, E.; Biggerstaff, A.; Bates, T.; Bennett, H.; Marlow, J.; Shaffer, M. Sediment impacts on marine sponges. Mar. Pollut. Bull. 2015, 94, 5–13. [Google Scholar] [CrossRef]
- Nickel, M. Evolutionary emergence of synaptic nervous systems: What can we learn from the non-synaptic, nerveless Porifera? Invertebr. Biol. 2010, 129, 1–16. [Google Scholar] [CrossRef]
- Jones, W.C. Is there a nervous system in sponges? Biol. Rev. 1962, 37, 1–47. [Google Scholar] [CrossRef]
- Ludeman, D.A.; Farrar, N.; Riesgo, A.; Paps, J.; Leys, S.P. Evolutionary origins of sensation in metazoans: Functional evidence for a new sensory organ in sponges. BMC Evol. Biol. 2014, 14, 3. [Google Scholar] [CrossRef]
- Leys, S.P.; Nichols, S.A.; Adams, E.D. Epithelia and integration in sponges. Integr. Comp. Biol. 2009, 49, 167–177. [Google Scholar] [CrossRef]
- Bagby, R.M. The fine structure of myocytes in the sponges Microciona prolifera (Ellis and Solander) and Tedania ignis (Duchassaing and Michelotti). J. Morphol. 1966, 118, 167–181. [Google Scholar] [CrossRef]
- Hammel, J.U.; Nickel, M. A new flow-regulating cell type in the demosponge Tethya wilhelma—Functional cellular anatomy of a leuconoid canal system. PLoS ONE 2014, 9, e113153. [Google Scholar] [CrossRef]
- Kanzawa, N.; Takano-Ohmuro, H.; Maruyama, K. Isolation and characterization of sea sponge myosin. Zool. Sci. 1995, 12, 765–769. [Google Scholar] [CrossRef]
- De Ceccatty, M.P. Coordination in sponges. The foundations of integration. Am. Zool. 1974, 14, 895–903. [Google Scholar] [CrossRef]
- Perovic, S.; Krasko, A.; Prokic, I.; Müller, I.M.; Müller, W.E. Origin of neuronal-like receptors in Metazoa: Cloning of a metabotropic glutamate/GABA-like receptor from the marine sponge Geodia cydonium. Cell Tissue Res. 1999, 296, 395–404. [Google Scholar] [CrossRef]
- Lieberkühn, N. Über Bewegungserscheinungen der Zellen; Band IX, N.G., Ed.; Elwert’sche Universitätsbuchhandlung: Marburg/Leipzig, Germany, 1870; pp. 9–22. [Google Scholar]
- Galtsoff, P.S. The amoeboid movement of dissociated sponge cells. Biol. Bull. 1923, 45, 153–161. [Google Scholar] [CrossRef]
- Loewenstein, W.R. On the genesis of cellular communication. Devel. Biol. 1967, 15, 503–520. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of research articles on Halichondria Fleming (1828) based on genus- and species-level (cf. [33]) according to Google Scholar (Web of Science) along with the main Web of Science research categories (accessed on 6 July 2022).
Table 1.
Number of research articles on Halichondria Fleming (1828) based on genus- and species-level (cf. [33]) according to Google Scholar (Web of Science) along with the main Web of Science research categories (accessed on 6 July 2022).
| Species | No. Articles | (%) | Web of Science Categories (%) |
|---|---|---|---|
| Halichondria panicea | 4040 (229) | 36.4 | Marine Freshwater Biology (43.5), Ecology (19.2), Oceanography (16.2) |
| Halichondria okadai | 2980 (96) | 26.8 | Organic Chemistry (39.6), Pharmacology Pharmacy (16.7), Biochemistry Molecular Biology (14.5) |
| Halichondria sp./spp. | 1390 (73) | 12.5 | Organic Chemistry (34.3), Medicinal Chemistry (20.6), Pharmacology Pharmacy (20.6) |
| Genus Halichondria | 723 (14) | 6.5 | Organic Chemistry (21.4), Pharmacology Pharmacy (21.4), Biochemistry Molecular Biology (14.3) |
| Halichondria bowerbanki | 447 (10) | 4.0 | Ecology (50.0), Marine Freshwater Biology (40.0), Zoology (30.0) |
| Halichondria japonica | 260 (20) | 2.3 | Biochemistry Molecular Biology (30.0), Organic Chemistry (20.0), Fisheries (15.0) |
| Halichondria cylindrata | 173 (10) | 1.6 | Organic Chemistry (70.0), Medicinal Chemistry (30.0), Biochemistry Molecular Biology (10.0) |
| Halichondria melanadocia | 169 (17) | 1.5 | Marine Freshwater Biology (52.9), Ecology (29.4), Anatomy Morphology (50.0) |
| Halichondria moorei | 108 (2) | 1.0 | Marine and Freshwater Biology (50.0), Multidisciplinary Sciences (50.0) |
| Halichondria sitiens | 89 (5) | 0.8 | Biodiversity Conservation (20.0), Biology (20.0), Ecology (20.0) |
| Halichondria oshoro | 82 (2) | 0.7 | Microbiology (100.0) |
| Halichondria magniconulosa | 67 (2) | 0.6 | Applied Chemistry (50.0), Medicinal Chemistry (50.0), Ecology (50.0) |
| Halichondria semitubulosa | 25 (1) | 0.2 | Zoology (100.0) |
| Halichondria cartilaginea | 19 (0) | 0.2 | - |
| Halichondria genitrix | 19 (0) | 0.2 | - |
| Halichondria albescens | 18 (0) | 0.2 | - |
| Halichondria lutea | 18 (3) | 0.2 | Biochemistry Molecular Biology (66.7), Ecology (66.7), Evolutionary Biology (66.7) |
| Halichondria coerulea | 14 (1) | 0.1 | Ecology (100.0), Marine Freshwater Biology (100.0), Oceanography (100.0) |
| Halichondria glabrata | 14 (2) | 0.1 | Anatomy and Morphology (50.0), Biology (50.0), Food Science Technology (50.0) |
| Halichondria diazae | 13 (0) | 0.1 | - |
| Halichondria cebimarensis | 12 (1) | 0.1 | Ecology (100.0), Marine Freshwater Biology (100.0) |
| Halichondria phakellioides | 12 (1) | 0.1 | Fisheries (100.0), Limnology (100.0), Marine Freshwater Biology (100.0) |
| Halichondria attenuata | 11 (2) | 0.1 | Marine Freshwater Biology (50.0), Zoology (50.0) |
| Halichondria contorta | 10 (1) | 0.1 | Zoology (100.0) |
| Halichondria topsenti | 10 (0) | 0.1 | - |
| Halichondria oblonga | 9 (0) | 0.1 | - |
| Halichondria aspera | 8 (0) | 0.1 | - |
| Halichondria cristata | 7 (0) | 0.1 | - |
| Halichondria agglomerans | 5 (0) | 0.0 | - |
| Halichondria flava | 5 (0) | 0.0 | - |
| Halichondria kelleri | 5 (0) | 0.0 | - |
| Halichondria migottea | 5 (0) | 0.0 | - |
| Halichondria osculum | 5 (1) | 0.0 | Medicinal Chemistry (100.0), Pharmacology Pharmacy (100.0) |
| Halichondria colossea | 4 (0) | 0.0 | - |
| Halichondria marianae | 4 (2) | 0.0 | Marine Freshwater Biology (50.0), Zoology (50.0) |
| Halichondria prostrata | 4 (0) | 0.0 | - |
| Halichondria tenebrica | 4 (0) | 0.0 | - |
| Halichondria capensis | 3 (0) | 0.0 | - |
| Halichondria convolvens | 3 (0) | 0.0 | - |
| Halichondria elenae | 3 (1) | 0.0 | Ecology (100.0), Marine Freshwater Biology (100.0) |
| Other species | 316 (36) | 2.1 | Cell biology (100.0), Zoology (100.0) |
| Total | 11,100 (532) | 100.0 | Marine and Freshwater Biology (27.3), Organic Chemistry (17.5), Ecology (12.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Goldstein, J.; Funch, P. A Review on Genus Halichondria (Demospongiae, Porifera). J. Mar. Sci. Eng. 2022, 10, 1312. https://doi.org/10.3390/jmse10091312
AMA Style
Goldstein J, Funch P. A Review on Genus Halichondria (Demospongiae, Porifera). Journal of Marine Science and Engineering. 2022; 10(9):1312. https://doi.org/10.3390/jmse10091312
Chicago/Turabian StyleGoldstein, Josephine, and Peter Funch. 2022. "A Review on Genus Halichondria (Demospongiae, Porifera)" Journal of Marine Science and Engineering 10, no. 9: 1312. https://doi.org/10.3390/jmse10091312
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.