Really Onychocellids? Revisions and New Findings Increase the Astonishing Bryozoan Diversity of the Mediterranean Sea



Abstract
:1. Introduction
2. Material and Methods
3. Systematic Account
3.1. Family Onychocellidae Jullien, 1882
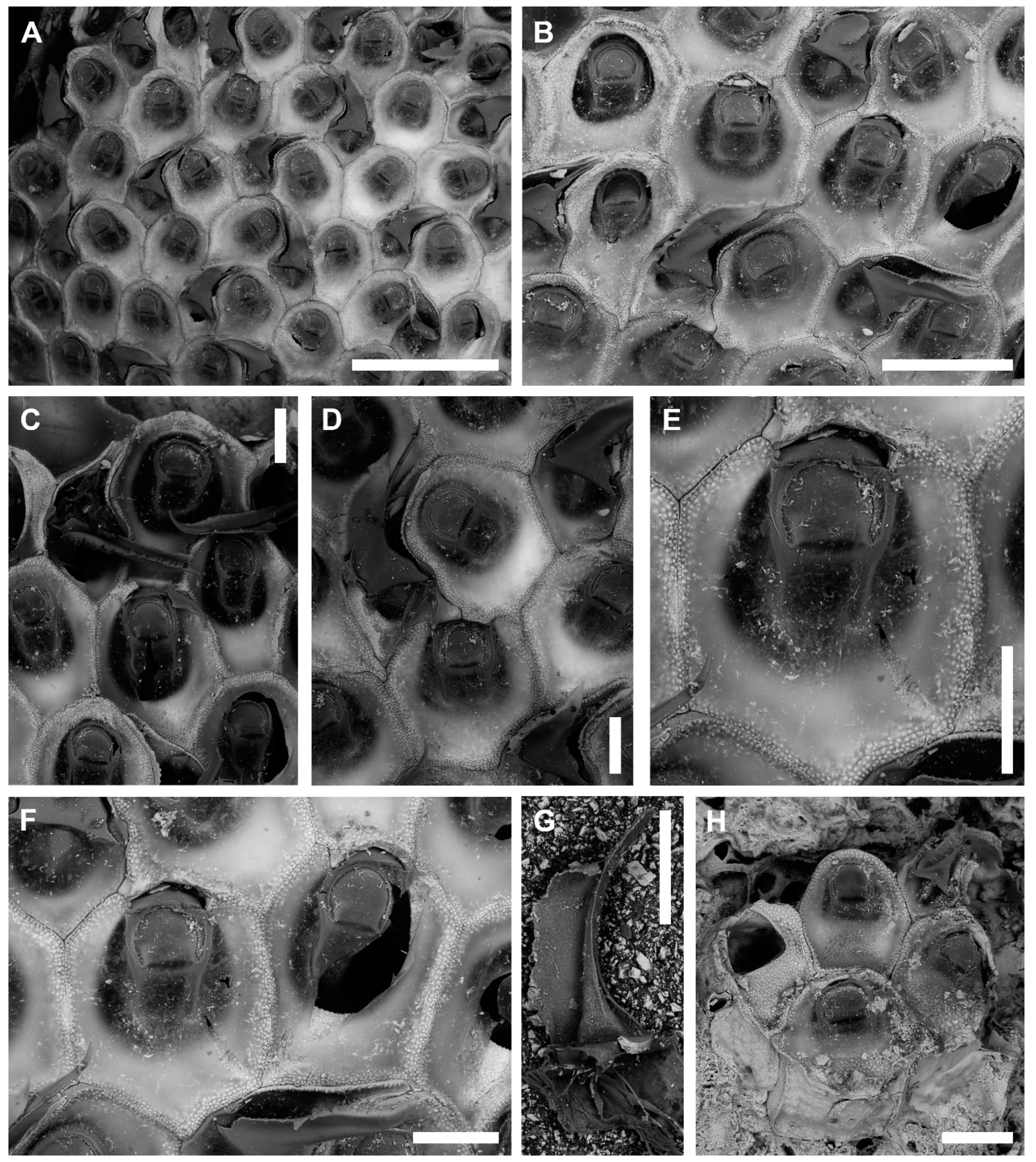
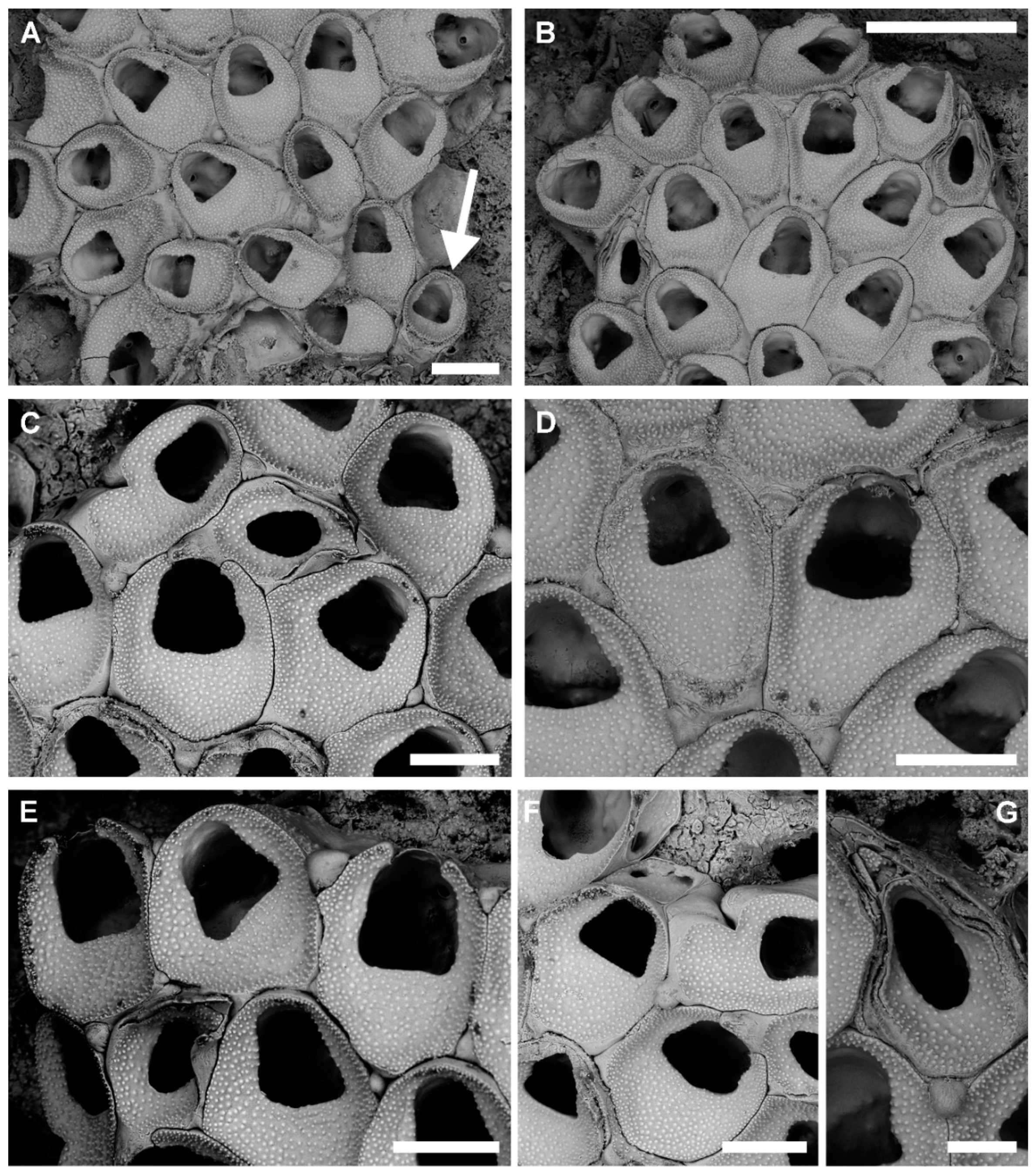
3.1.1. Onychocella marioni Jullien, 1882
Synonymies
Material Examined
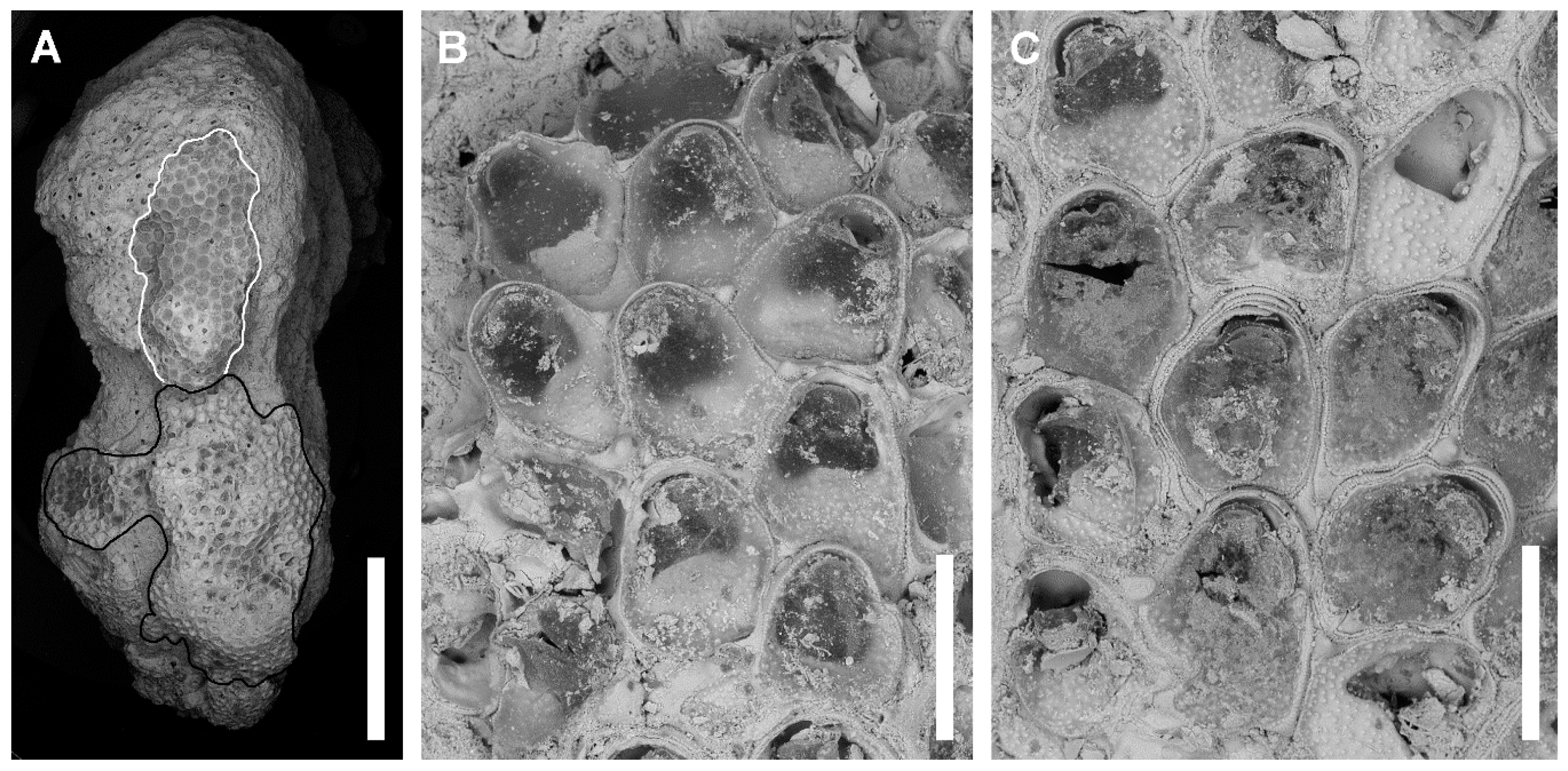
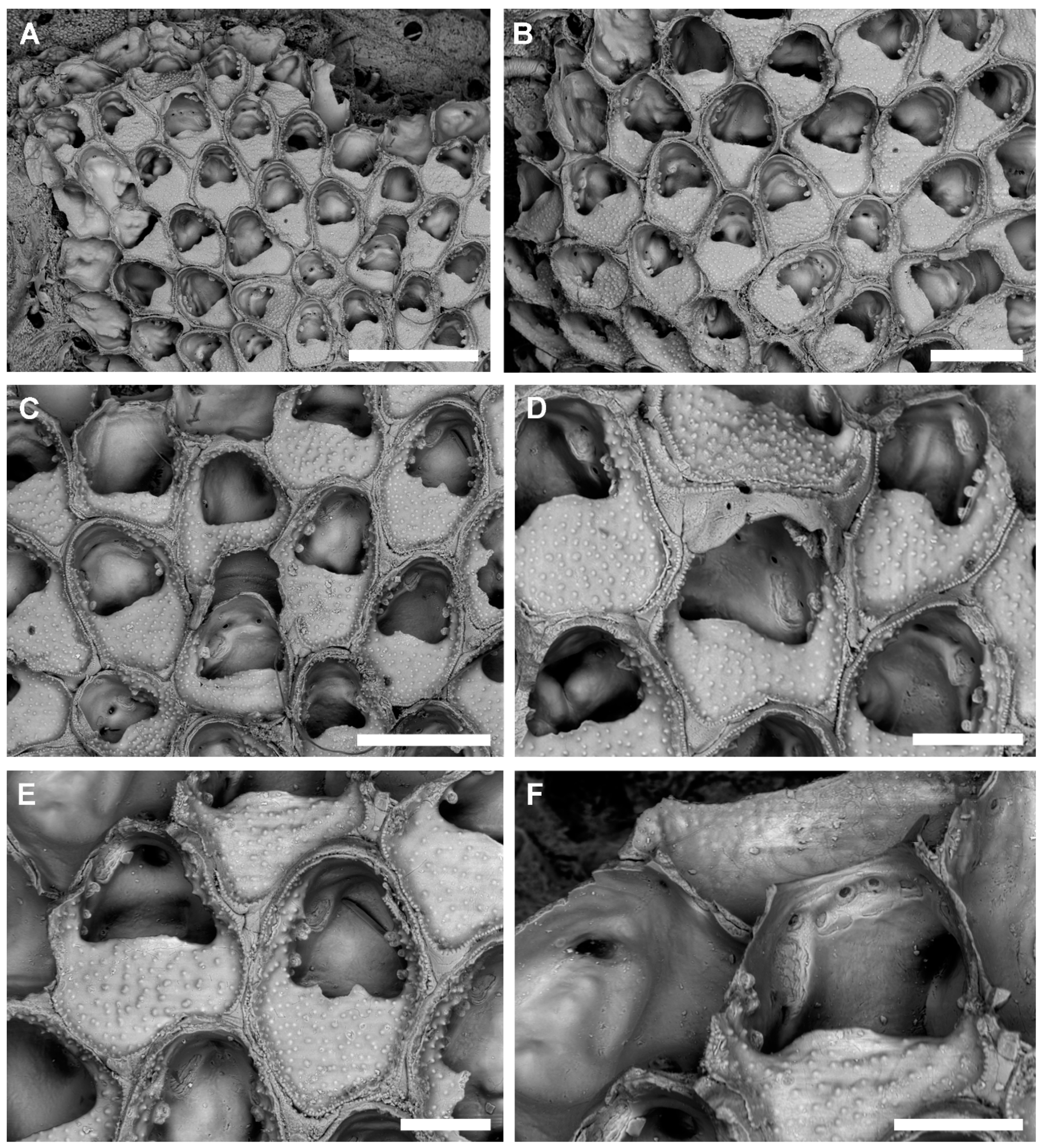
Description
Remarks
Distribution
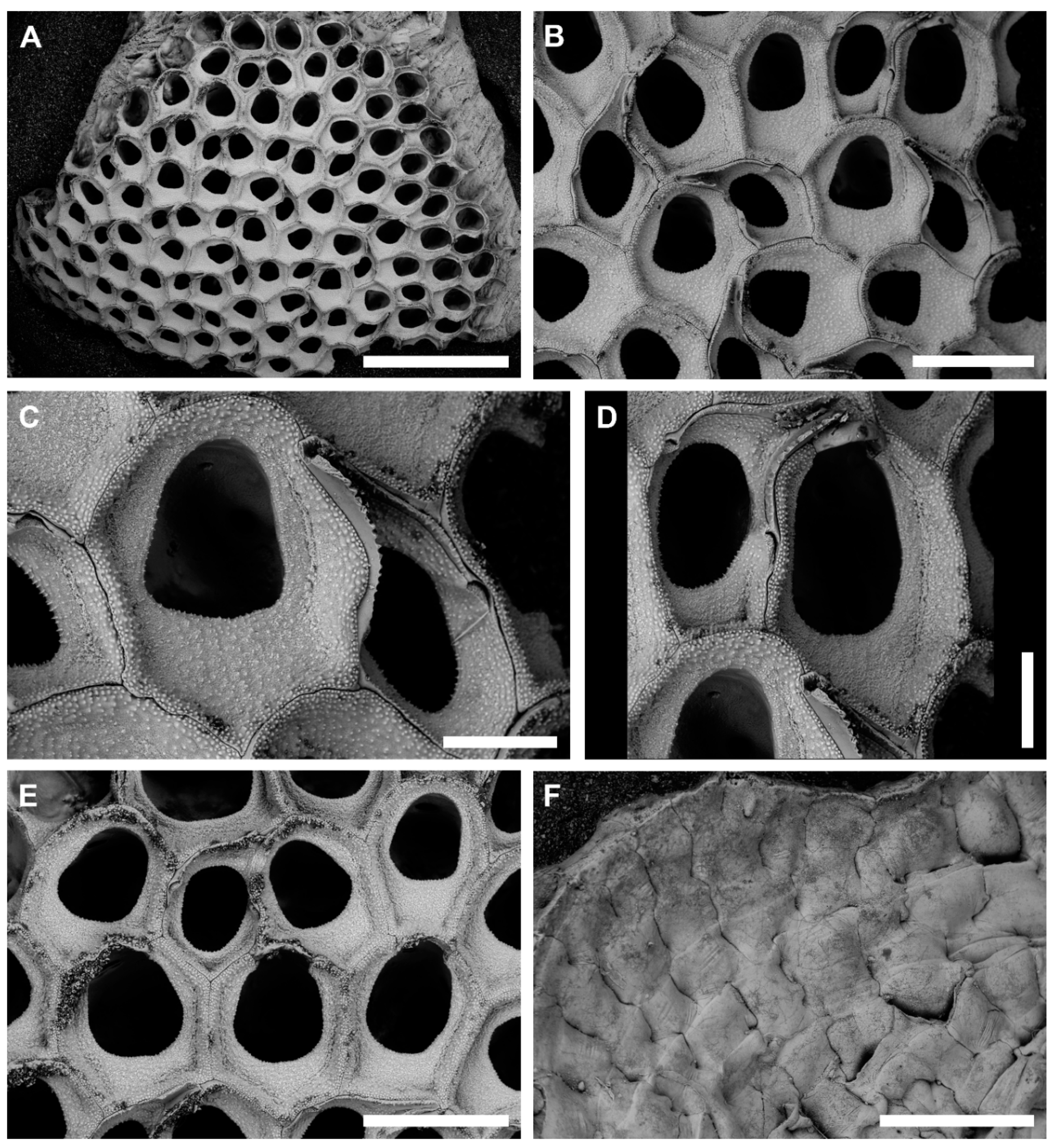
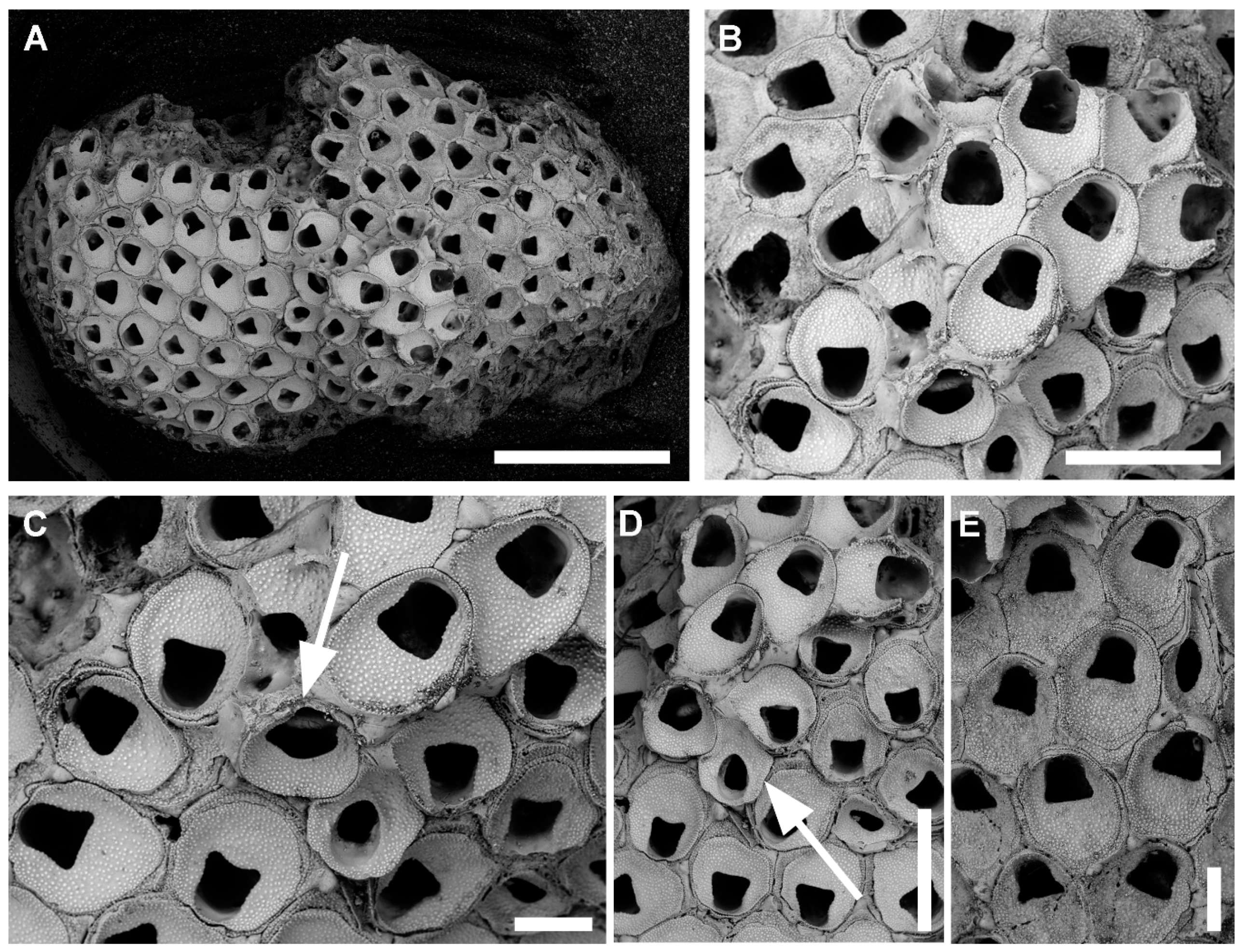
3.1.2. Onychocella vibraculifera Neviani, 1895
Synonymies
Material Examined
Description
Remarks
Distribution
3.2. Family Microporidae Gray, 1848
3.2.1. Genus Bryobifallax gen. nov.
Diagnosis
Remarks
Etymology
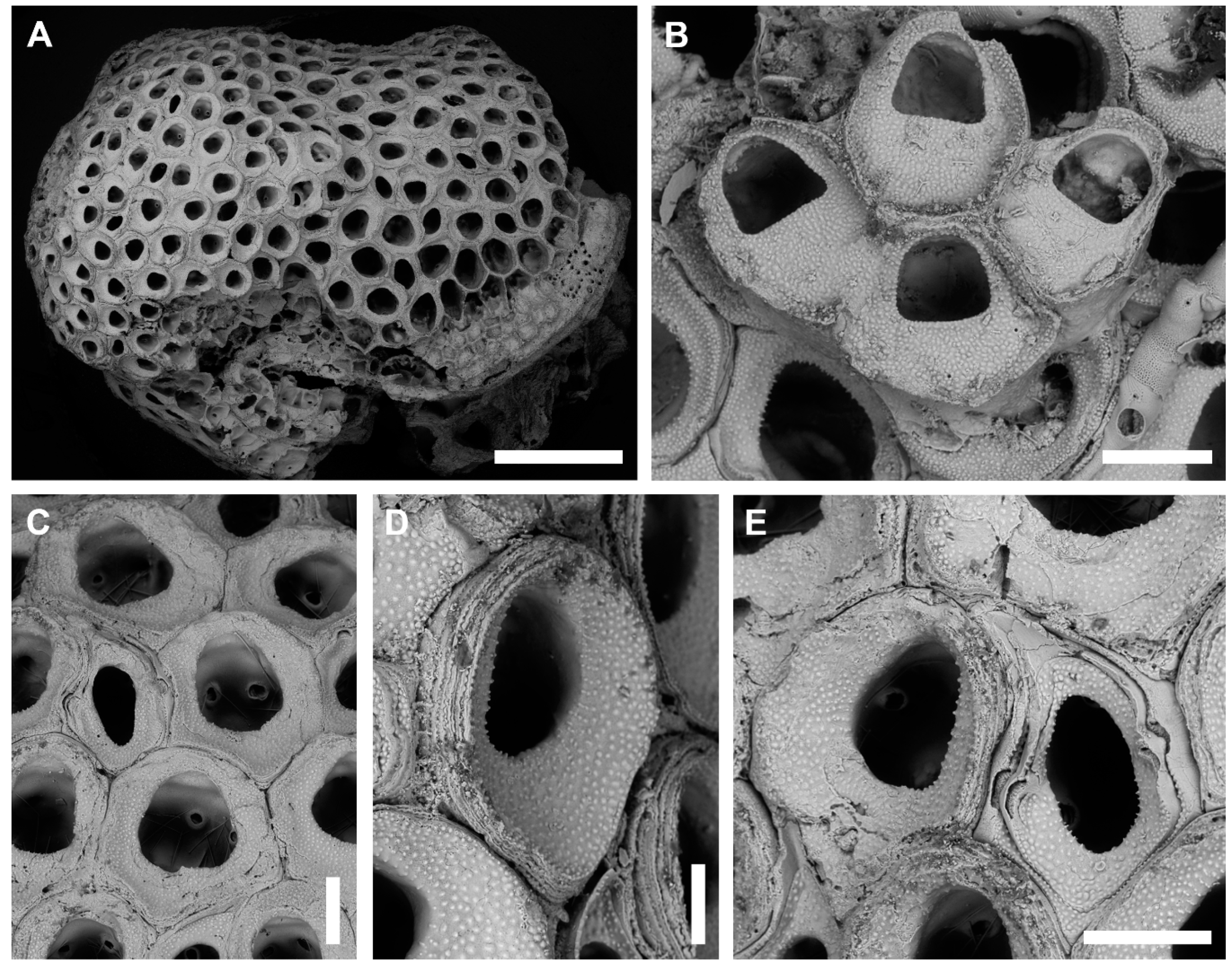
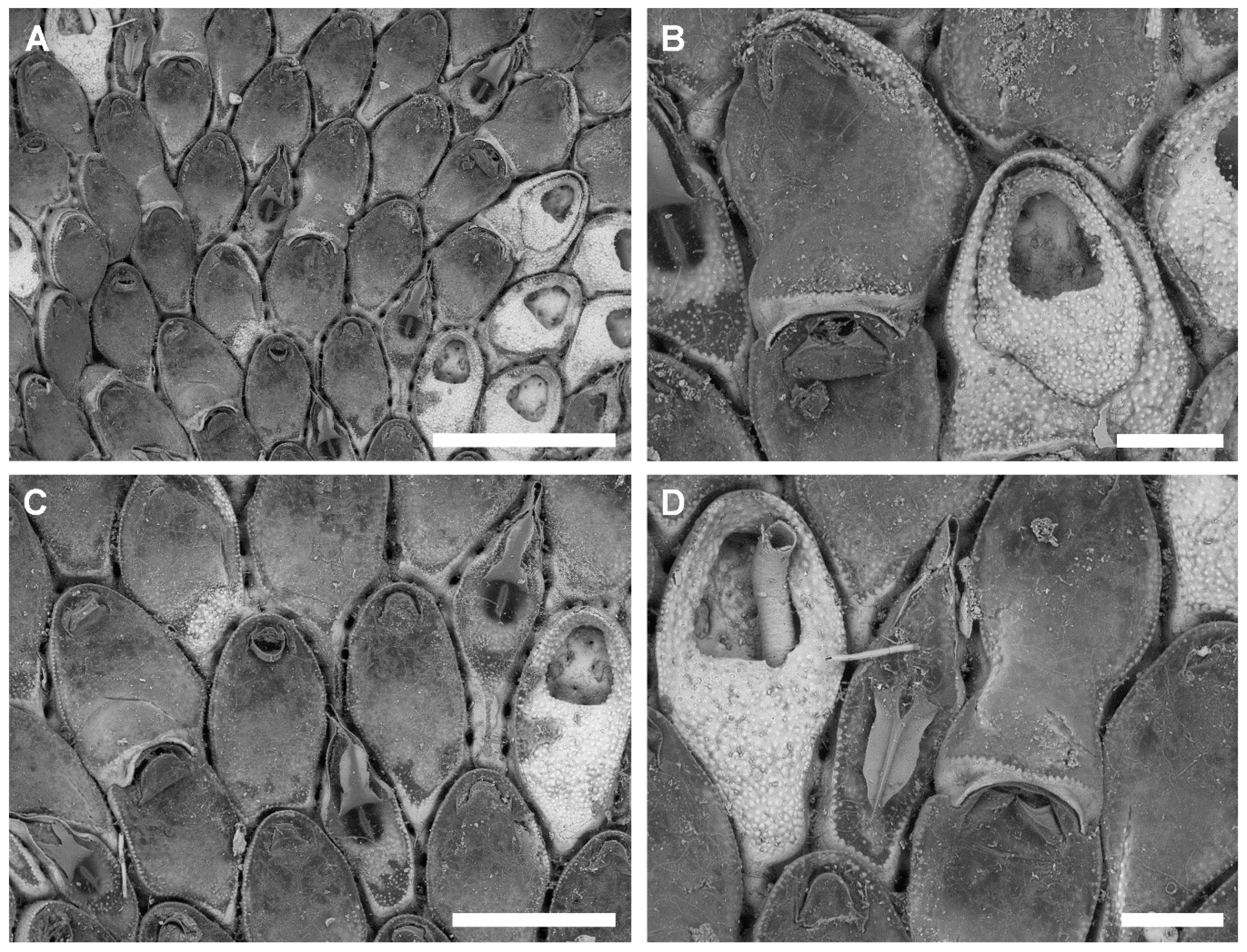
3.2.2. Bryobifallax disjuncta (Canu & Bassler, 1930) comb. nov.
Synonymies
Material Examined
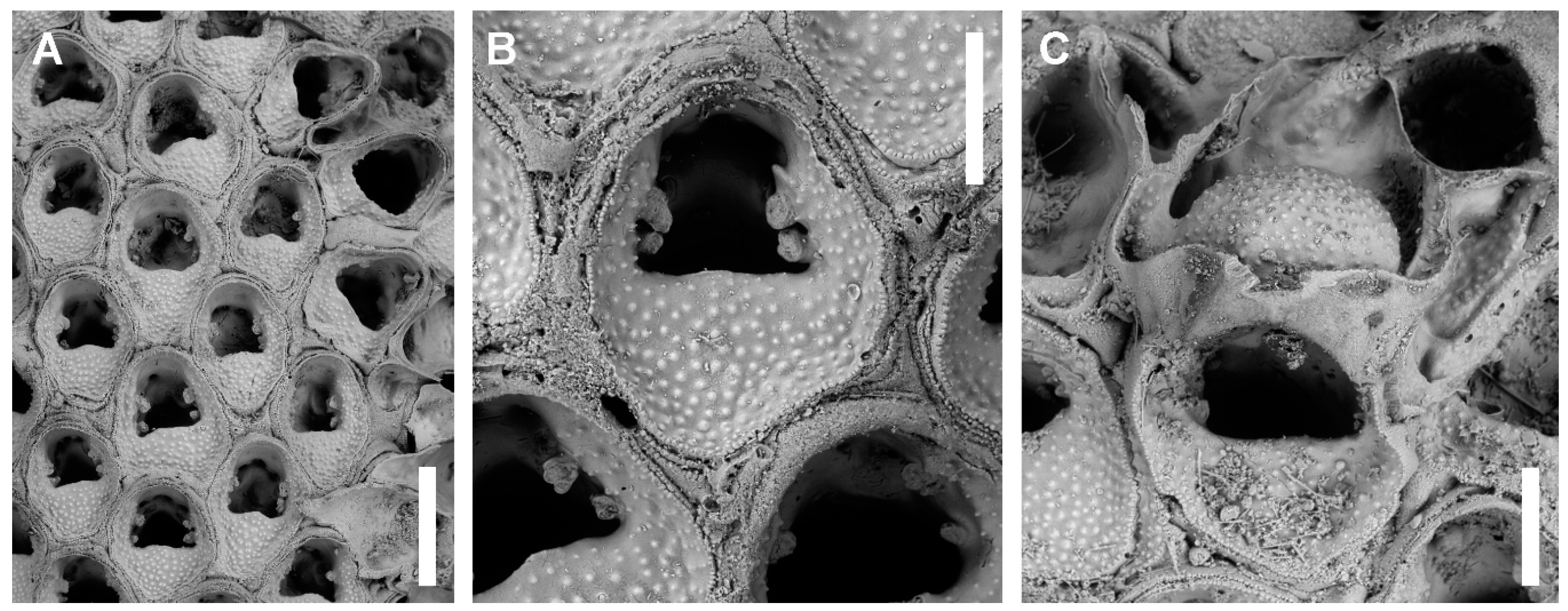
Description
Remarks
Distribution
3.3. Family Calescharidae Cook & Bock, 2001 [62]
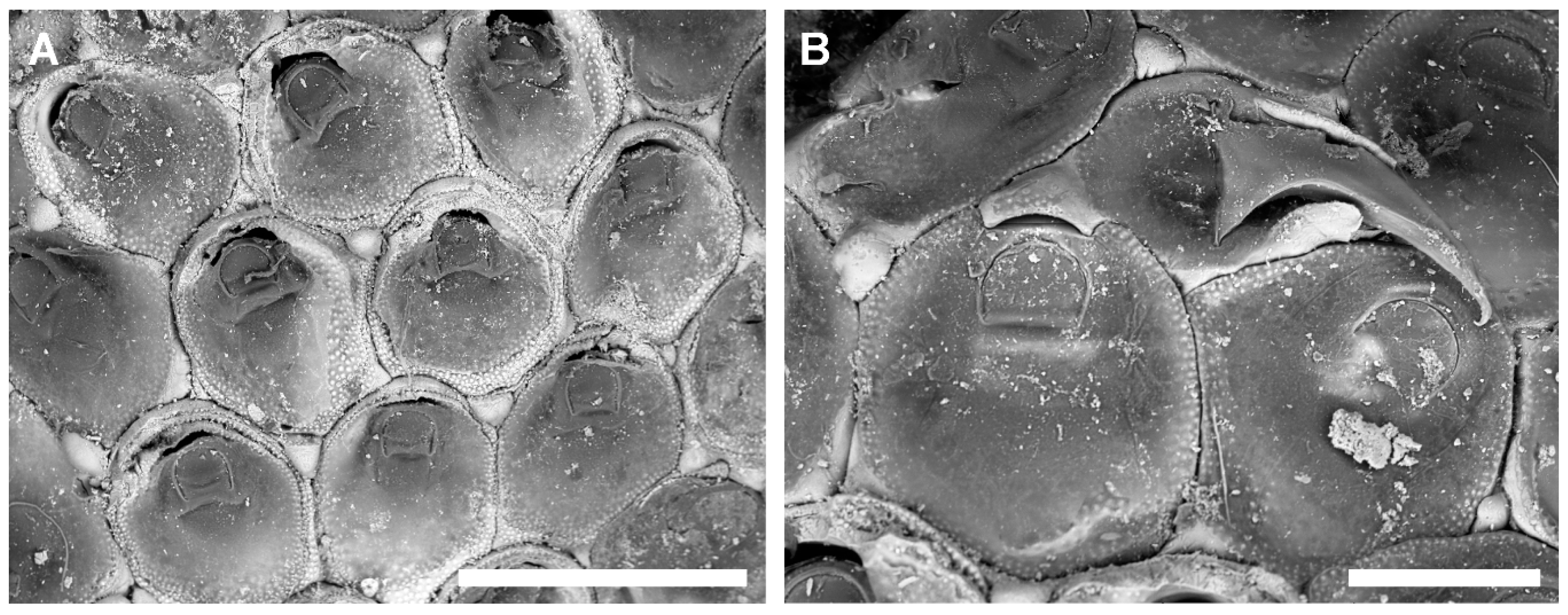
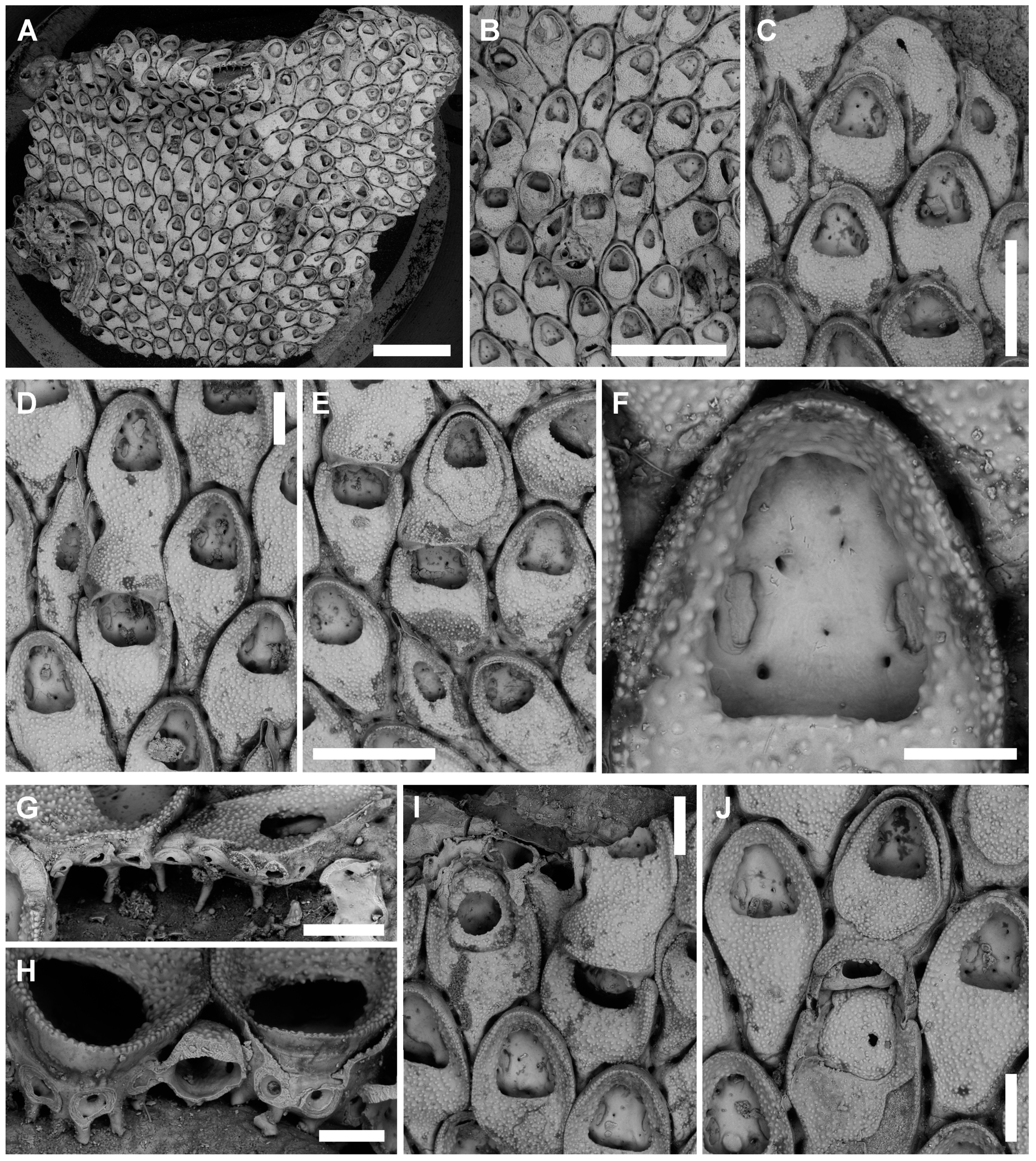
3.3.1. Tretosina arculifera (Canu & Bassler, 1927) comb. nov.
Synonymies
Material Examined
Description
Remarks
Distribution
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Imperato, F. Historia Naturale; Presso Combi, & la Noù: Venetia, Italy, 1599. [Google Scholar]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, B.R.F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosso, A.; Di Martino, E. Bryozoan diversity in the Mediterranean Sea: An up-date. Med. Mar. Sci. 2016, 17, 567–607. [Google Scholar] [CrossRef]
- Rosso, A.; Gerovasileiou, V.; Sanfilippo, R.; Guido, A. Bryozoans assemblages from two submarine caves in the Aegean Sea (Eastern Mediterranean). Mar. Biodivers. 2019, 49, 707–726. [Google Scholar] [CrossRef]
- Rosso, A.; Gerovasileiou, V.; Sanfilippo, R.; Guido, A. Undisclosed bryodiversity of submarine caves of the Aegean Sea (Eastern Mediterranean). In Proceedings of the 2nd Mediterranean Symposium on the Conservation of Dark Habitats, Antalya, Turkey, 16 January 2019; RAC/SPA: Antalya, Turkey, 2019; pp. 47–52. [Google Scholar]
- Rosso, A.; Di Martino, E.; Gerovasileiou, V. Revision of the genus Setosella (Bryozoa: Cheilostomata) with description of new species from deep-waters and submarine caves of the Mediterranean. Zootaxa 2020, 4728, 401–442. [Google Scholar] [CrossRef] [PubMed]
- Schwaha, T.; Bernhard, J.M.; Edgcomb, V.P.; Todaro, M.A. Aethozooides uraniae, a new deep-sea genus and species of solitary bryozoan from the Mediterranean Sea, with a revision of the Aethozoidae. Mar. Biodiv. 2019, 49, 1843–1856. [Google Scholar] [CrossRef] [Green Version]
- Achilleos, K.; Jimenez, C.; Berning, B.; Petrou, A. Bryozoan diversity of Cyprus (eastern Mediterranean Sea): First results from census surveys (2011–2018). Med. Mar. Sci. 2020, 21, 228–237. [Google Scholar] [CrossRef]
- Jullien, J. Note sur une nouvelle division des Bryozoaires Cheilostomiens. Bull. Soc. Zool. Fr. 1882, 6, 271–285. [Google Scholar]
- Taylor, P.D.; Martha, S.O.; Gordon, D.P. Synopsis of ‘onychocellid’ cheilostome bryozoan genera. J. Nat. Hist. 2018, 52, 1657–1721. [Google Scholar] [CrossRef]
- Gautier, Y.V. Recherches écologiques sur les Bryozoaires Chilostomes en Méditerranée occidentale. Rec. Trav. Stat. Mar. Endoume 1962, 38, 1–434. [Google Scholar]
- Prenant, M.; Bobin, G. Bryozoaires, deuxieme partie. Chilostomes Anasca; Fédération Française des sociétés de sciences naturelles: Paris, France, 1966; pp. 1–647. [Google Scholar]
- Zabala, M.; Maluquer, P. Illustrated keys for the classification of Mediterranean Bryozoa. Treb. Mus, Zool. Barcelona 1988, 4, 1–294. [Google Scholar]
- Harmelin, J.-G. Bryozoaires récoltés au cours de la campagne du Jean Charcot en Méditerranée orientale (Août-Septembre 1967). I. Dragages. Bull. Mus. nat. Hist. nat. Sér. 2 1969, 40, 1179–1208. [Google Scholar]
- Hayward, P.J. Studies on the cheilostome bryozoan fauna of the Aegean island of Chios. J. Nat. Hist. 1974, 8, 369–402. [Google Scholar] [CrossRef]
- Chimenz-Gusso, C.; Nicoletti, L.; Bondanese, C. Briozoi. Biol. Mar. Medit. 2014, 20 (Suppl. 1), 1–336. [Google Scholar]
- Gerovasileiou, V.; Rosso, A. Marine bryozoa of Greece: An annotated checklist. Biodiv. Data J. 2016, 4, e10672. [Google Scholar] [CrossRef] [Green Version]
- Harmelin, J.-G.; Bitar, G.; Zibrowius, H. High xenodiversity versus low native diversity in the south-eastern Mediterranean: Bryozoans from the coastal zone of Lebanon. Med. Mar. Sci. 2016, 17, 417–439. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Trygonis, V.; Sini, M.; Koutsoubas, D.; Voultsiadou, E. Three-dimensional mapping of marine caves using a handheld echosounder. Mar. Ecol. Prog. Ser. 2013, 486, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Voultsiadou, E. Sponge diversity gradients in marine caves of the eastern Mediterranean. J. Mar. Biol. Assoc. UK 2016, 96, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Dimitriadis, C.; Arvanitidis, C.; Voultsiadou, E. Taxonomic and functional surrogates of sessile benthic diversity in Mediterranean marine caves. PLoS ONE 2017, 12, e0183707. [Google Scholar] [CrossRef] [Green Version]
- Sanfilippo, R.; Rosso, A.; Guido, A.; Gerovasileiou, V. Serpulid communities from two marine caves in the Aegean Sea, eastern Mediterranean. J. Mar. Biol. Assoc. UK 2017, 97, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Rosso, A.; Di Martino, E.; Sanfilippo, R.; Di Martino, V. Bryozoan communities and thanatocoenoses from submarine caves in the plemmirio marine protected area (SE Sicily). In Bryozoan Studies 2010, Proceedings of the 15th IBA Conference, Kiel, Germany, 16−18 May 2010; Ernst, A., Schäfer, P., Scholz, J., Eds.; Lecture Notes in Earth System Sciences; Springer: Berlin/Heidelberg, Germany, 2013; Volume 143, pp. 251–269. [Google Scholar]
- Canu, F. Révision des Bryozoaires du Crétacé figurés par d’Orbigny. II, Cheilostomata. Bull. Soc. Géol. Fr. 1900, 28, 334–463. [Google Scholar]
- Canu, F.; Bassler, R.S. Bryozoaires marins de Tunisie. Ann. Stat. Océanogr. Salammbô. 1930, 5, 1–91. [Google Scholar]
- Rosso, A. Popolamenti e tanatocenosi a Briozoi di fondi mobili circalitorali del Golfo di Noto (Sicilia SE). Nat. Sicil. 1996, 20, 189–225. [Google Scholar]
- Di Geronimo, I.; Di Geronimo, R.; Improta, S.; Rosso, A.; Sanfilippo, R. Preliminary observation on a columnar coralline build-up from off SE Sicily. Biol. Mar. Medit. 2001, 8, 229–237. [Google Scholar]
- Rosso, A.; Sanfilippo, R.; Taddei Ruggiero, E.; Di Martino, E. Faunas and ecological groups of Serpuloidea, Bryozoa and Brachiopoda from submarine caves in Sicily (Mediterranean Sea). Boll. Soc. Paleont. Ital. 2013, 52, 167–176. [Google Scholar]
- Berning, B. The cheilostome bryozoan fauna from the Late Miocene of Niebla (Guadalquivir Basin, SW Spain): Environmental and biogeographic implications. Mitt. Geol. Paläont. Inst. Univ. Hambg. 2006, 90, 7–156. [Google Scholar]
- Unsal, I. Bryozaires marins de Turquie. Istambul Üniv. Fen Fakül Mecmuasi Ser. B 1975, 40, 37–54. [Google Scholar]
- Reuss, A.E. Die fossilen Polyparien des Wiener Tertiärbeckens. Haid. Naturwis. Abhand. 1848, 2, 1–109. [Google Scholar]
- Zágoršek, K. Bryozoa from the Langhian (Miocene) of the Czech Republic. Part I: Geology of the studied sections, systematic description of the orders Cyclostomata, Ctenostomata and “Anascan” Cheilostomata (Suborders Malacostega Levinsen, 1902 and Flustrina Smitt, 1868). Acta Mus. Nat. Prague Ser. B Hist. Nat. 2010, 66, 3–136. [Google Scholar]
- Cook, P.L. Polyzoa from West Africa. 1. Notes on the Steganoporellidae, Thalamoporellidae, and Onychocellidae. Result. Sci. Camp. Calypso 1964, 41, 43–78. [Google Scholar]
- Cook, P.L. Bryozoa from Ghana: A preliminary survey. Ann. Mus. Afr. Cent. Sci. Zool. 1985, 238, 1–315. [Google Scholar]
- Gautier, Y.V. Résultats scientifiques des campagnes de la Calypso. Ann. Inst. Oceanogr. 1956, 32, 189–225. [Google Scholar]
- Zabala, M. Fauna dels Briozous dels Països Catalans. Inst. Est. Catalans Arx. Sec. Ciénc. Barc. 1986, 84, 1–836. [Google Scholar]
- Harmelin, J.-G. Bryozoan-dominated assemblages in Mediterranean cryptic environments. In Bryozoa: Ordovician to Recent; Nielsen, C., Larwood, G.P., Eds.; Olsen & Olsen: Fredensborg, Denmark, 1985; pp. 135–143. [Google Scholar]
- WoRMS Editorial Board. World Register of Marine Species. 2020. Available online: http://www.marinespecies.org at VLIZ (accessed on 3 September 2020).
- Rosso, A.; Sanfilippo, R. The contribution of bryozoans and serpuloideans to coralligenous concretions from SE Sicily. In Proceedings of the First Symposium on the Coralligenous and Other Calcareous Bio-concretions of the Mediterranean Sea, Tabarka, Tunisia, 15–16 January 2009; UNEP-MAP-RAC/SPA: Tabarka, Tunisia, 2009; pp. 123–128. [Google Scholar]
- Di Geronimo, I.; Di Geronimo, R.; Rosso, A.; Sanfilippo, R. Structural and taphonomic analysis of a columnar build-up from Sicily shelf. Géobios. Mem. Spec. 2002, 24, 86–95. [Google Scholar] [CrossRef]
- Di Geronimo, I.; Giacobbe, S.; Rosso, A.; Sanfilippo, R. Popolamenti e tanatocenosi del Banco Apollo (Ustica, Mar Tirreno meridionale). Boll. Mus. Reg. Sc. Nat. Torino 1990, spec, 697–729. [Google Scholar]
- Rosso, A. Valutazione della biodiversità in Mediterraneo: L’esempio dei popolamenti a briozoi della Biocenosi del Detritico Costiero. Biol. Mar. Medit. 1996, 3, 58–65. [Google Scholar]
- Balduzzi, A.; Rosso, A. Briozoi. In Grotte Marine: Cinquant’anni di Ricerca in Italia; Cicogna, F., Bianchi, N.C., Ferrari, G., Forti, P., Eds.; CLEM-ONLUS, Ministero dell’Ambiente e della Tutela del Territorio: Massa Lubrense, Napoli, Italy, 2003; pp. 195–202. [Google Scholar]
- Harmelin, J.-G. Le sous-ordre des Tubuliporina (Bryozoaires Cyclostomes) en Méditerranée. Écologie et systématique. Mém. Inst. Océanogr. 1976, 10, 1–326. [Google Scholar]
- Harmelin, J.-G. Biodiversité des habitats cryptiques marins du parc national de Port-Cros (Méditerranée, France). Assemblages de bryozoaires d’une grotte sous-marine et des faces inférieures de pierres. Sci. Rep. Port-Cros Natl. Park Fr. 2003, 19, 101–116. [Google Scholar]
- Harmelin, J.G. Patterns in the distribution of bryozoans in the Mediterranean marine caves. Stygologia 1986, 2, 10–25. [Google Scholar]
- Gili, J.M.; Olivella, I.; Zabala, M.; Ros, J. Primera contribución al conocimiento del poblamiento de las cuevas submarinas del litoral catalán. In Actas del Ier Simposio Ibérico de Estudio del Bentos Marino; Niell, F.X., Ros, J.D., Eds.; Universidad del País Vasco: Bilbao, Spain, 1982; pp. 818–836. [Google Scholar]
- Morri, C.; Bianchi, C.N.; Cocito, S.; Peirano, A.; De Biase, A.M.; Aliani, S.; Pansini, M.; Boyer, M.; Ferdeghini, F.; Pestarino, M.; et al. Biodiversity of marine sessile epifauna at an Aegean island subject to hydrothermal activity: Milos, eastern Mediterranean Sea. Mar. Biol. 1999, 135, 729–739. [Google Scholar] [CrossRef]
- Koçak, F.; Balduzzi, A.; Benli, H.A. Epiphytic bryozoan community of Posidonia oceanica (L.) Delile meadow in the northern Cyprus (Eastern Mediterranean). Indian J. Mar. Sci. 2002, 31, 235–238. [Google Scholar]
- Koçak, F.; Aydin Önen, S. Checklist of Bryozoa on the coasts of Turkey. Turkish J. Zool. 2014, 38, 880–891. [Google Scholar] [CrossRef]
- Neviani, A. Briozoi fossili della Farnesina e Monte Mario presso Roma. Palaeontogr. Ital. 1985, 1, 79–100. [Google Scholar]
- Rosso, A.; Chimenz Gusso, C.; Balduzzi, A. Bryozoa. Checklist della Flora e della Fauna dei Mari Italiani (Parte II). Biol. Mar. Medit. 2010, 17, 589–615. [Google Scholar]
- Winston, J.E.; Vieira, L.M. Systematics of intertidal incrusting bryozoans from southeastern Brazil. Zootaxa 2013, 3710, 101–146. [Google Scholar] [CrossRef] [PubMed]
- Di Geronimo, I.; La Perna, R.; Rosso, A.; Sanfilippo, R. Popolamento e tanatocenosi bentonica della Grotta dell’Accademia (Ustica, Mar Tirreno meridionale). Nat. Sicil. 1993, 17, 45–63. [Google Scholar]
- Di Geronimo, I.; Allegri, L.; Improta, S.; La Perna, R.; Rosso, A.; Sanfilippo, R. Spatial and Temporal aspects of Recent benthic thanatocoenoses in a Mediterranean Infralittoral cave. Riv. It. Paleont. Strat. 1997, 103, 15–28. [Google Scholar]
- Harmer, S.F. The Polyzoa of the Siboga Expedition, 2. Cheilostomata Anasca. Siboga Exped. Rep. 1926, 28, 183–501. [Google Scholar]
- Bock, P.E. Bryozoa Home Page. Available online: http://www.bryozoa.net (accessed on 31 August 2020).
- Ostrovsky, A.N. Evolution of sexual reproduction in marine invertebrates. In Example of Gymnolaemate Bryozoans; Springer: Dordrecht, The Netherlands, 2013; pp. 1–356. [Google Scholar]
- Hayward, P.J. Antarctic Cheilostomatous Bryozoa; Oxford University Press: Oxford, UK, 1995; pp. 1–355. [Google Scholar]
- Moyano, G.H.I. Flustrapora magellanica nov. gen. nov. sp., (Bryozoa, Cheilostomata, Anasca). Bol. Soc. Biol. Concepción 1970, 42, 59–65. [Google Scholar]
- Souto, J.; Reverter-Gil, O.; Ostrovsky, A.N. New species of Bryozoa from Madeira associated with rhodoliths. Zootaxa 2014, 3795, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Cook, P.L.; Bock, P.E. Calescharidae, a new family for the Tertiary to Recent genera Caleschara MacGillivray and Tretosina Canu & Bassler (Bryozoa: Cheilostomata). Inv. Taxon. 2001, 15, 527–550. [Google Scholar]
- Canu, F.; Bassler, R.S. Bryozoaires des Iles Hawaii. Bull. Soc. Sci. Seine Oise 1927, 7, 1–66. [Google Scholar]
- Canu, F.; Bassler, R.S. A synopsis of American Early Tertiary cheilostome Bryozoa. Bull. U.S. Natl. Mus. 1917, 96, 1–87. [Google Scholar] [CrossRef]
- Canu, F.; Bassler, R.S. North American early tertiary bryozoa. Bull. U.S. Natl. Mus. 1920, 106, 1–879. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.D.; McKinney, F.K. Cretaceous Bryozoa from the cCmpanian and Maastrichtian of the Atlantic and gulf coastal plains, United States. Scr. Geol. 2006, 132, 1–346. [Google Scholar]
- Tilbrook, K.J. Cheilostomatous Bryozoa from the Solomon Islands. Stud. Biodivers. St. Barbara Mus. Nat. Hist. Monogr. 4 2006, 3, 1–386. [Google Scholar]
- Gordon, D.P. The marine fauna of New Zealand: Bryozoa Ceilostomata from the Kermadek Ridge. N. Zeal. Oceanogr. Inst. 1984, 91, 1–198. [Google Scholar]
- Gordon, D.P.; Taylor, P.D. Late Paleocene to earliest Eocene bryozoans from Chatham Islands, New Zealand. Bull. Nat. Hist. Mus. Lond. (Geol.) 1999, 55, 1–45. [Google Scholar]
- Meunier, A.; Pergens, E. Les Bryozoaires du Systéme Montien (Éocène inférieur); Louvain, Belgium, 1886; Volume 1, pp. 1–15. Available online: https://books.google.com.hk/books/about/Les_bryozoaires_du_syst%C3%A8me_montien_%C3%A9oc.html?id=F7e_GwAACAAJ&redir_esc=y (accessed on 3 September 2020).
- Cook, P.L.; Chimonides, P.J. Morphology and systematics of some rooted cheilostome Bryozoa. J. Nat. Hist. 1981, 15, 97134. [Google Scholar] [CrossRef]
- Winston, J.E. An annotate checklist of coral-associated bryozoans. Am. Mus. Novit. 1986, 2859, 1–39. [Google Scholar]
- Harmelin, J.-G. Diversity of bryozoans in a Mediterranean sublittoral cave with bathyal-like conditions: Role of dispersal processes and local factors. Mar. Ecol. Progr. Ser. 1997, 153, 139–152. [Google Scholar] [CrossRef]
- Harmelin, J.-G.; Vacelet, J.; Vasseur, P. Les grottes sous-marines obscures: Un milieu extrême et un remarquable biotope refuge. Téthys 1985, 11, 214–229. [Google Scholar]
- Brown, D.A. The Tertiary cheilostomatous Polyzoa of New Zealand; Trustees of the British Museum (Natural History): London, UK, 1952; pp. 1–405. [Google Scholar]
- Berning, B.; Tilbrook, K.J.; Rosso, A. Revision of the north-eastern Atlantic and Mediterranean species of the genera Herentia and Therenia (Bryozoa: Cheilostomata). J. Nat. Hist. 2008, 42, 1509–1547. [Google Scholar] [CrossRef]
- Harmelin, J.-G.; Bishop, J.D.D.; Madurell, T.; Souto, J.; Jones, M.E.S.; Zabala, M. The genus Collarina (Bryozoa, Cheilostomatida) in the NE Atlantic-Mediterranean region: Unexpected diversity. New species and reappraisal of C. balzaci and C. fayalensis. Zoosystema 2019, 41, 385–418. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cave | Fara Cave | Agios Vasilios Cave | |||||
|---|---|---|---|---|---|---|---|
| Cave depth (m) | 11–18 | 24–40 | |||||
| Sampling station | F3 | FC2 | F4 | VC1 | V1 | VC2 | V2 |
| Sample location inside caves | Walls | Ceiling | Walls | Ceiling | Walls | Ceiling | Walls |
| Distance from the entrance (m) | 5–10 | 15–20 | 20–30 | 5–10 | 5–10 | 15–20 | 15–20 |
| Depth (m) | 17 | 15 | 17 | 25 | 30 | 27 | 30 |
| Biocoenosis | SD | Trans | Dark | SD | SD | Dark | Dark |
| Dominant encrusters | Sc-Sp | Sr-B | Sr-Sp | Sc-Sp | Sp | Sc-Sp-Sr | Sp-Sr |
| Onychocella marioni Jullien, 1882 | 1 (1) | 17 (4) | 20 (10) | 6 (3) | 17 (12) | 12 (4) | 34 (14) |
| Onychocella vibraculifera Neviani, 1895 | 2 | 6 (1) | |||||
| Bryobifallax disjuncta (Canu & Bassler, 1930) | 4 (1) | 2 (1) | |||||
| Tretosina arculifera (Canu & Bassler, 1927) | 3 (2) | (1) | |||||
| Species | Onychocella marioni | Onychocella vibraculifera |
|---|---|---|
| Jullien, 1882 | Neviani, 1895 | |
| Zooid length | 457.67–633.84; 537.72 ± 49.26 (n = 20) | 372.33–477.72; 426.97 ± 27.57 (n = 20) |
| Zooid width | 376.88–527.94; 441.68 ± 37.86 (n = 20) | 233.27–388.75; 325.67 ± 34.09 (n = 20) |
| L/W | 1.22 | 1.31 |
| Opesial length | 226.63–312.37; 274.69 ± 24.60 (n = 20) | 127.28–216.33; 174.41 ± 25.89 (n = 20) |
| Opesial width | 201.57–281.54; 233.44 ± 23.35 (n = 20) | 134.48–191.81; 165.39 ± 18.76 (n = 20) |
| Ovicellate zooidal length | 574.89–685.52; 620.34 ± 37.82 (n = 8) | 430.80–472.63; 450.56 ± 16.65 (n = 5) |
| Ovicellate zooidal width | 415.24–607.05; 525.30 ± 60.95 (n = 8) | 314.49–352.17; 336.03 ± 13.78 (n = 5) |
| Ovicellate zooid opesial length | 382.62–422.59; 401.92 ± 15.14 (n = 8) | 150.42–231.93; 196.51 ± 37.55 (n = 5) |
| Ovicellate zooid opesial width | 207.64–345.15; 303.19 ± 45.51 (n = 8) | 167.23–205.84; 183.16 ± 15.95 (n = 5) |
| Ooecium length | 42.04–47.69; 44.87 ± 4.00 (n = 2) | 55.96–59.56; 58.31 ± 2.04 (n = 3) |
| Ooecium width | 150.32–205.11; 177.72 ± 38.74 (n = 2) | 110.52–189.21; 141.73 ± 41.79 (n = 3) |
| Avicularium length | 435.83–729.55; 566.61 ± 69.09 (n = 20) | 285.24–419.75; 363.63 ± 62.80 (n = 5) |
| Avicularium width | 244.06–393.10; 315.07 ± 39.96 (n = 20) | 184.00–239.27; 209.69 ± 21.78 (n = 5) |
| Avicularium opesial length | 214.57–366.37; 267.31 ± 40.66 (n = 20) | 147.95–173.69; 158.98 ± 11.78 (n = 5) |
| Avicularium opesial width | 132.58–224.46; 170.64 ± 25.16 (n = 20) | 63.71–94.01; 79.73 ± 12.39 (n = 5) |
| Ancestrula length | 326.13–353.02; 359.58 ± 19.01 (n = 2) | 213.23 |
| Ancestrula width | 399.70–426.12; 412.91 ± 18.68 (n = 2) | 191.32 |
| Ancestrula opesial length | 157.57 | 128.22 |
| Ancestrula opesial width | 168.82 | 117.66 |
| Tubercle diameter | Absent | 33.80–69.96; 54.35 ± 11.45 (n = 10) |
| Species | Bryobifallax disjuncta | Tretosina arculifera |
|---|---|---|
| (Canu & Bassler, 1930) comb. nov. | (Canu & Bassler, 1927) comb. nov. | |
| Zooid length | 481.32–701.91; 594.33 ± 64.00 (n = 20) | 510.23–776.94; 648.36 ± 70.24 (n = 20) |
| Zooid width | 278.80–406.40; 362.81 ± 28.65 (n = 20) | 233.67–485.80; 418.57 ± 44.90 (n = 20) |
| L/W | 1.64 | 1.55 |
| Opesial length | 184.53–257.25; 223.45 ± 21.83 (n = 20) | 218.04–333.56; 282.21 ± 30.55 (n = 20) |
| Opesial width | 180.28–233.48; 208.81 ± 14.72 (n = 20) | 212.20–304.17; 263.00 ± 30.16 (n = 20) |
| Ooecium length | 161.36–281.15; 223.06 ± 37.24 (n = 12) | 210.07–319.72; 266.71 ± 54.91 (n = 3) |
| Ooecium width | 307.71–353.94; 325.02 ± 16.81 (n = 12) | 407.75–444.96; 426.17 ± 18.61 (n = 3) |
| Avicularium length | 501.44–657.21; 574.23 ± 47.97 (n = 10) | Absent |
| Avicularium width | 211.52–256.26; 235.47 ± 14.44 (n = 10) | Absent |
| Avicularium opesial length | 120.92–197.04; 162.33 ± 22.47 (n = 10) | Absent |
| Avicularium opesial width | 89.26–113.40; 102.53 ± 7.87 (n = 10) | Absent |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosso, A.; Gerovasileiou, V.; Di Martino, E. Really Onychocellids? Revisions and New Findings Increase the Astonishing Bryozoan Diversity of the Mediterranean Sea. J. Mar. Sci. Eng. 2020, 8, 904. https://doi.org/10.3390/jmse8110904
Rosso A, Gerovasileiou V, Di Martino E. Really Onychocellids? Revisions and New Findings Increase the Astonishing Bryozoan Diversity of the Mediterranean Sea. Journal of Marine Science and Engineering. 2020; 8(11):904. https://doi.org/10.3390/jmse8110904
Chicago/Turabian StyleRosso, Antonietta, Vasilis Gerovasileiou, and Emanuela Di Martino. 2020. "Really Onychocellids? Revisions and New Findings Increase the Astonishing Bryozoan Diversity of the Mediterranean Sea" Journal of Marine Science and Engineering 8, no. 11: 904. https://doi.org/10.3390/jmse8110904