
Interactive Effects of Flooding Duration and Sediment Texture on the Growth and Adaptation of Three Plant Species in the Poyang Lake Wetland
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
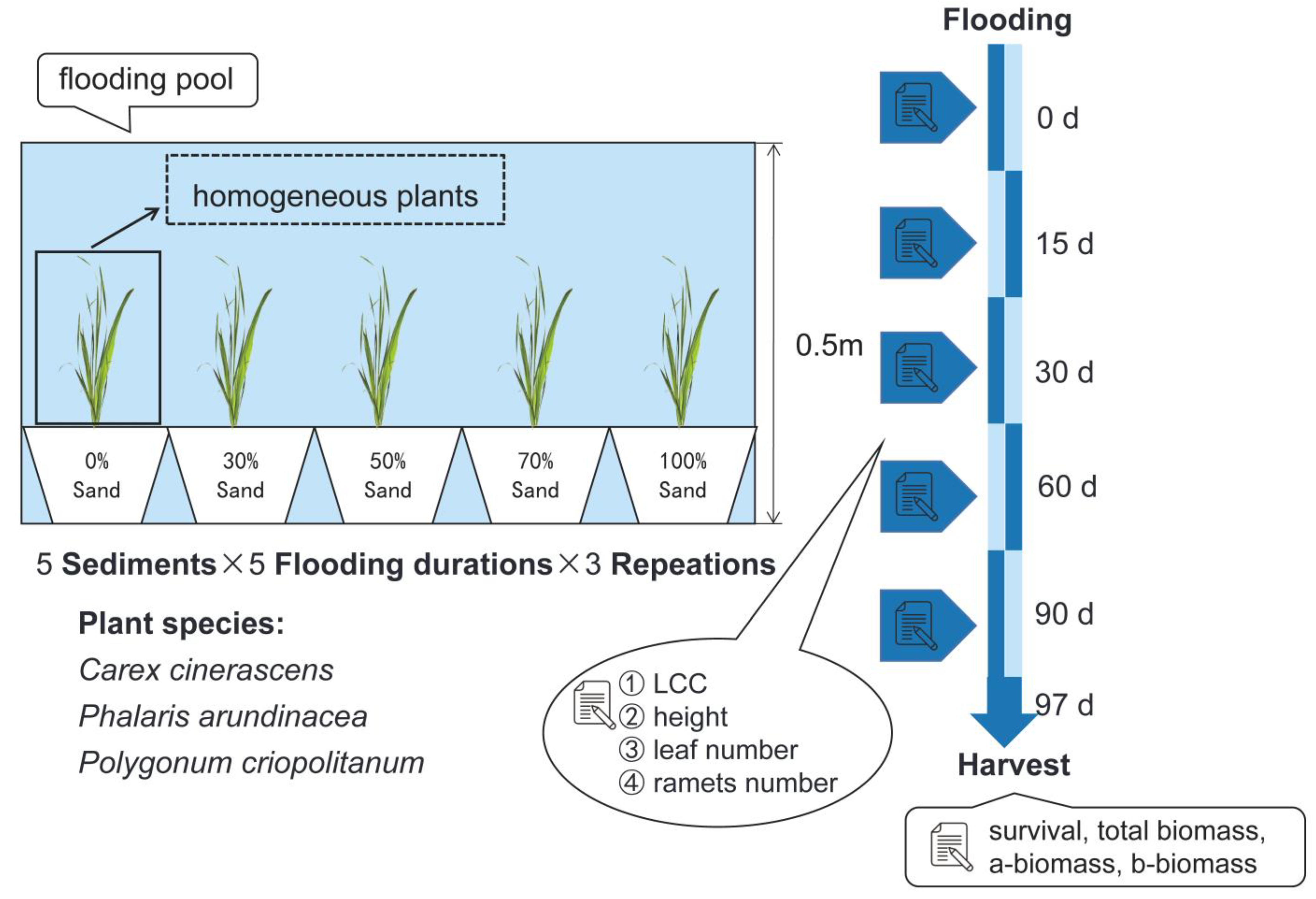
2.2. Experimental Design
2.3. Experimental Measurements
2.4. Data Analysis
3. Results
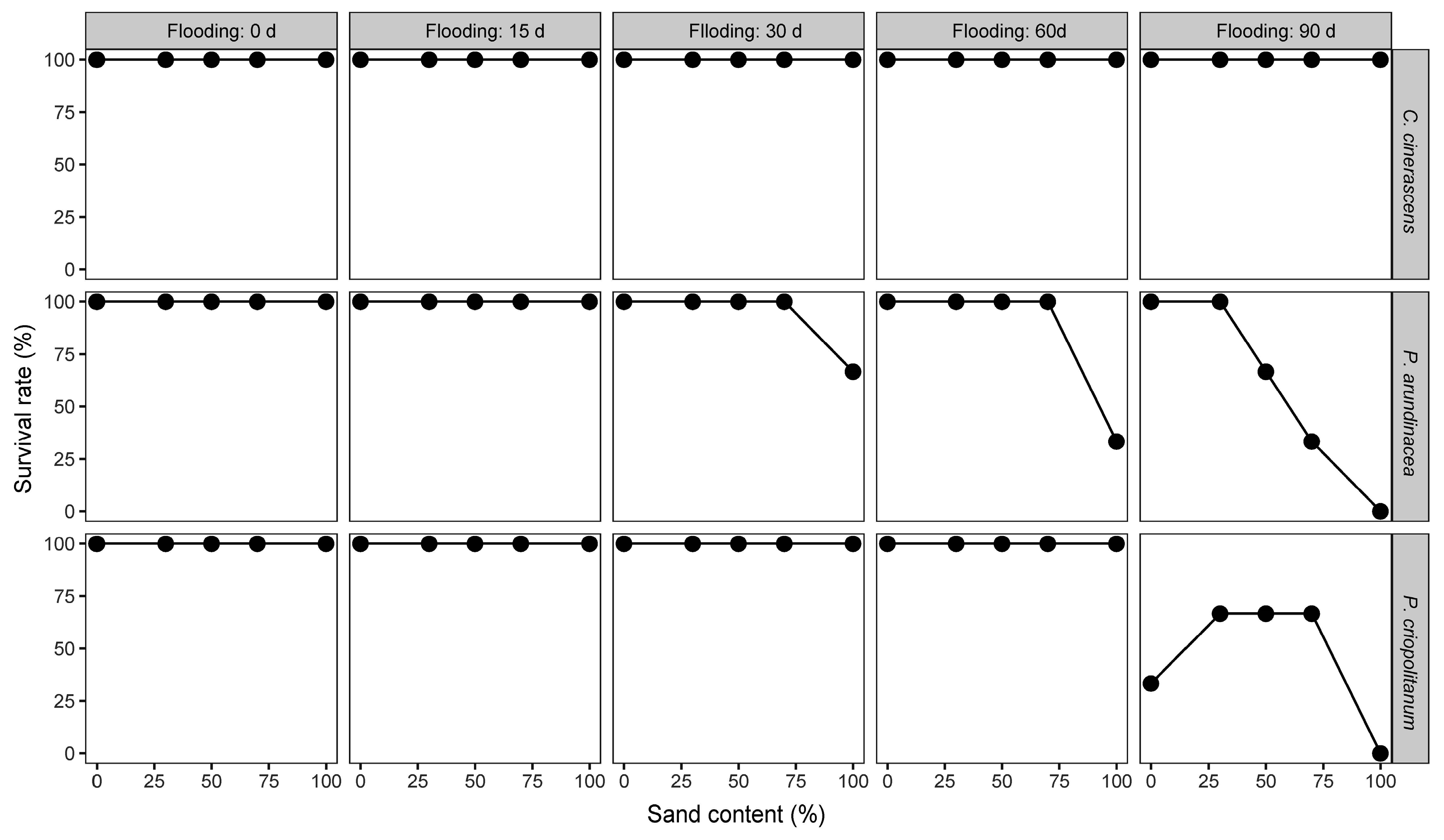
3.1. Effects of Flooding Duration and Sediment Texture on the Survival of Hygrophilous Plants
3.2. Effects of Flooding Duration and Sediment Texture on the Functional Traits of Hygrophilous Plants
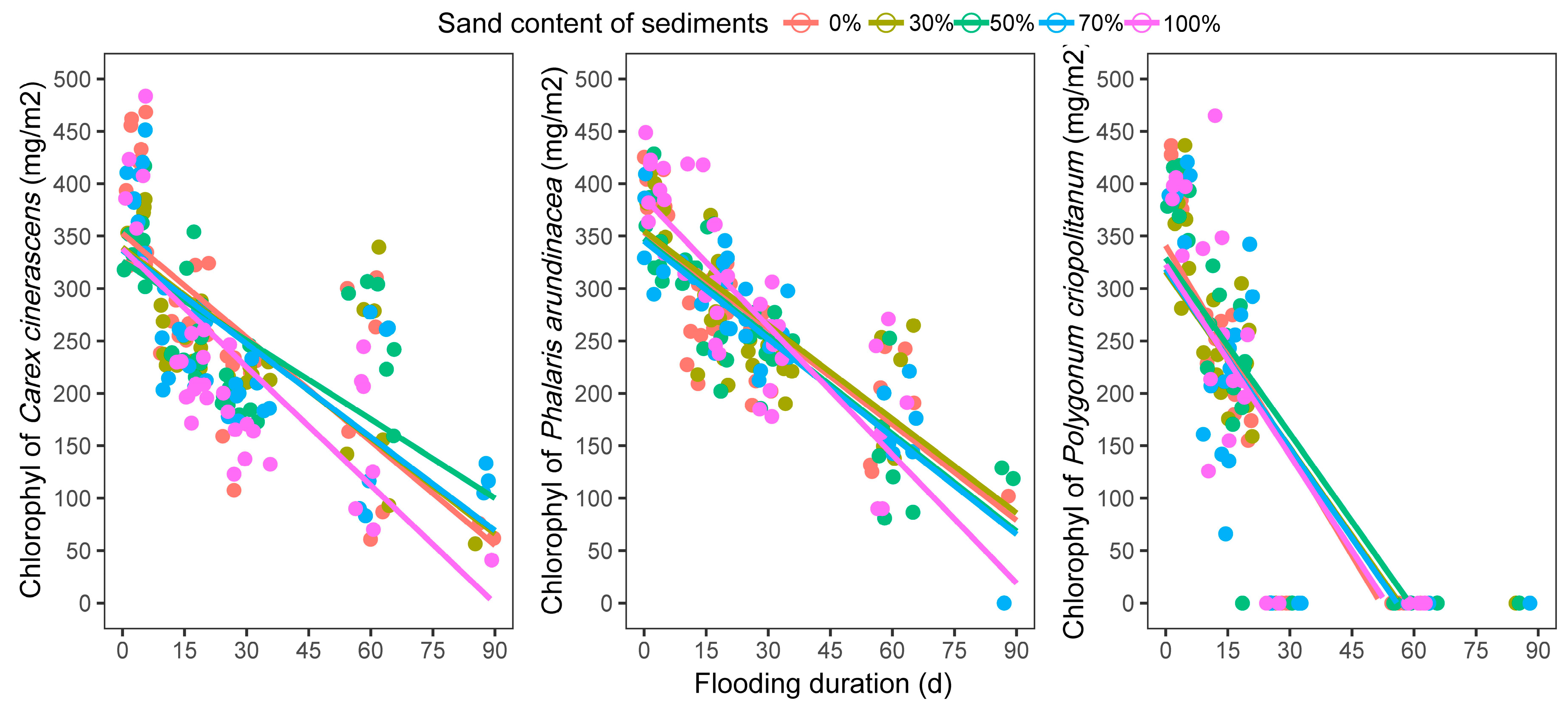
3.2.1. Leaf Chlorophyll Content
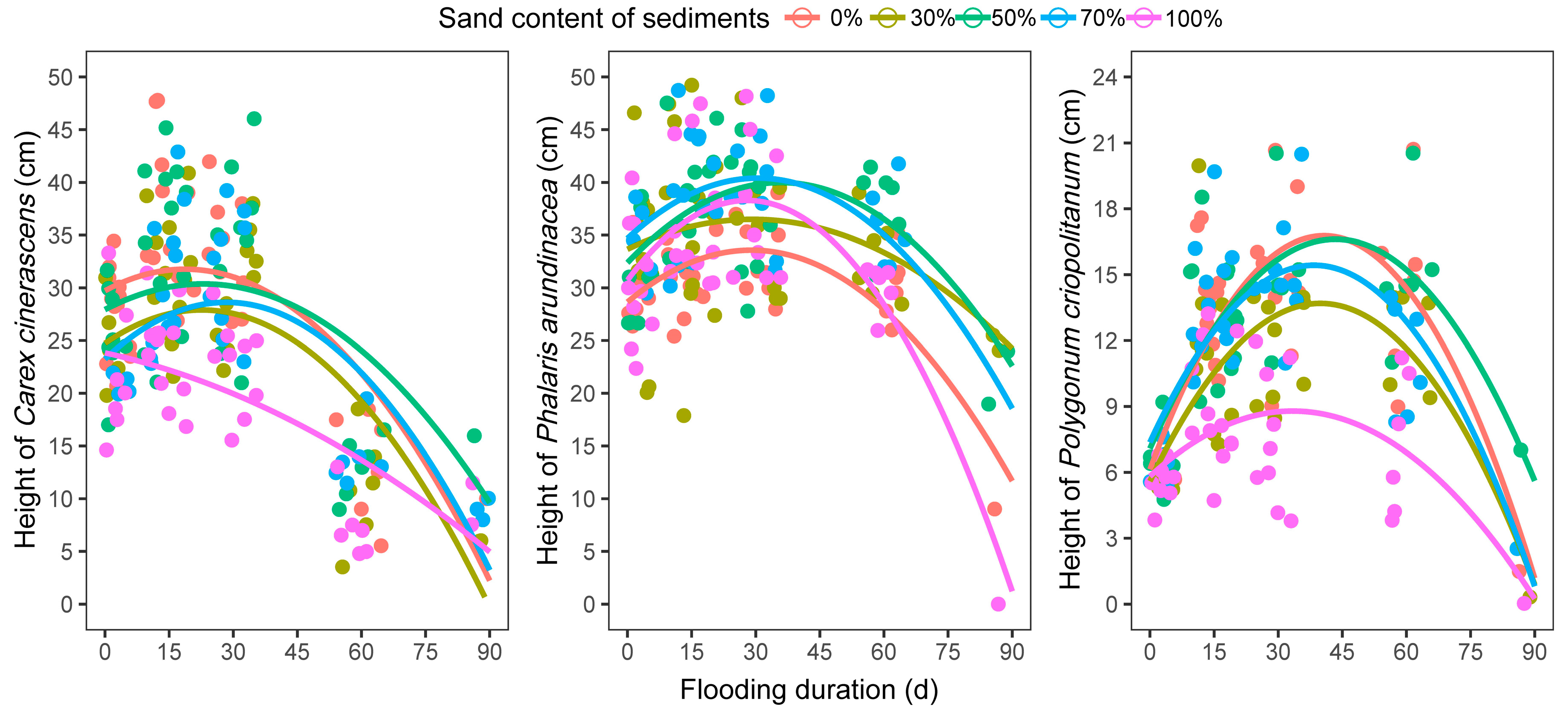
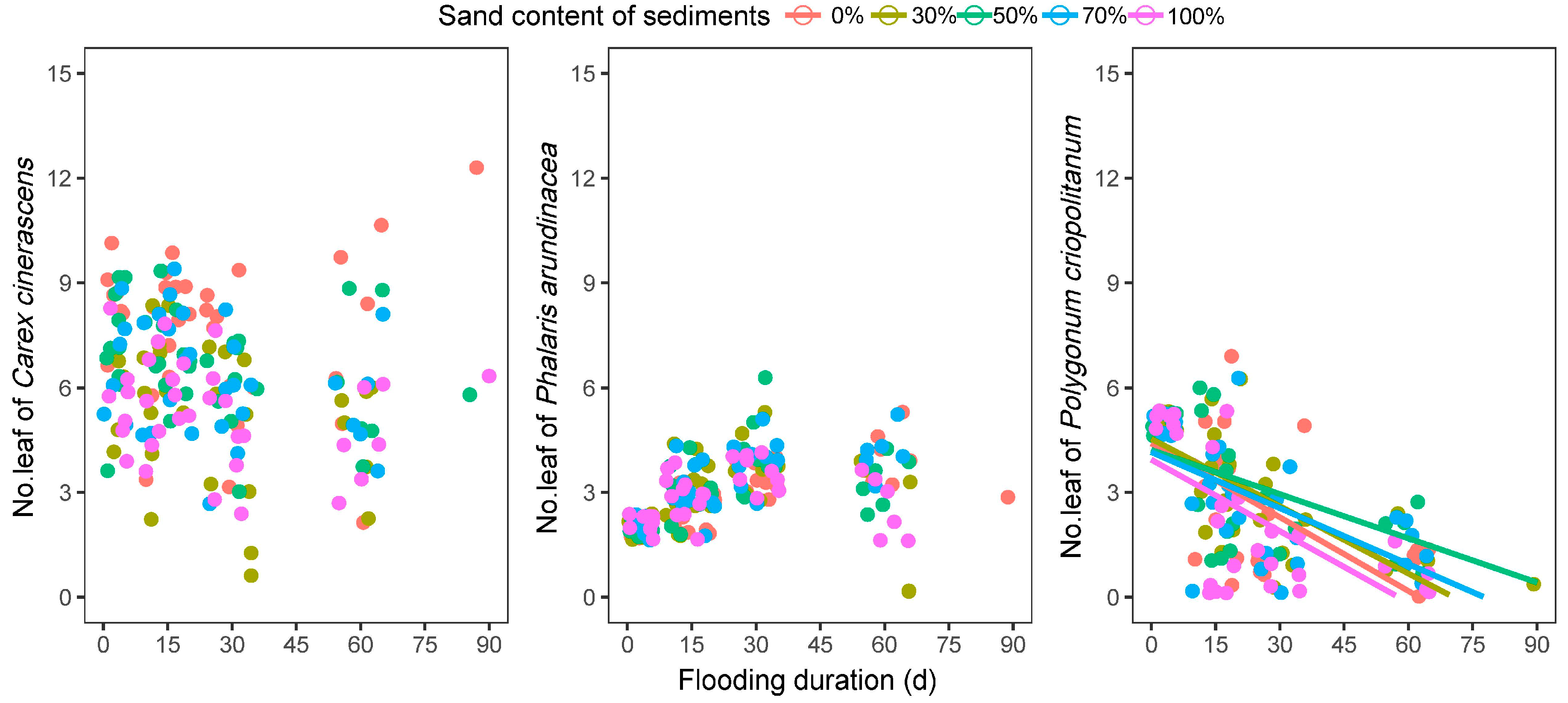
3.2.2. Plant Height and Number of Leaves
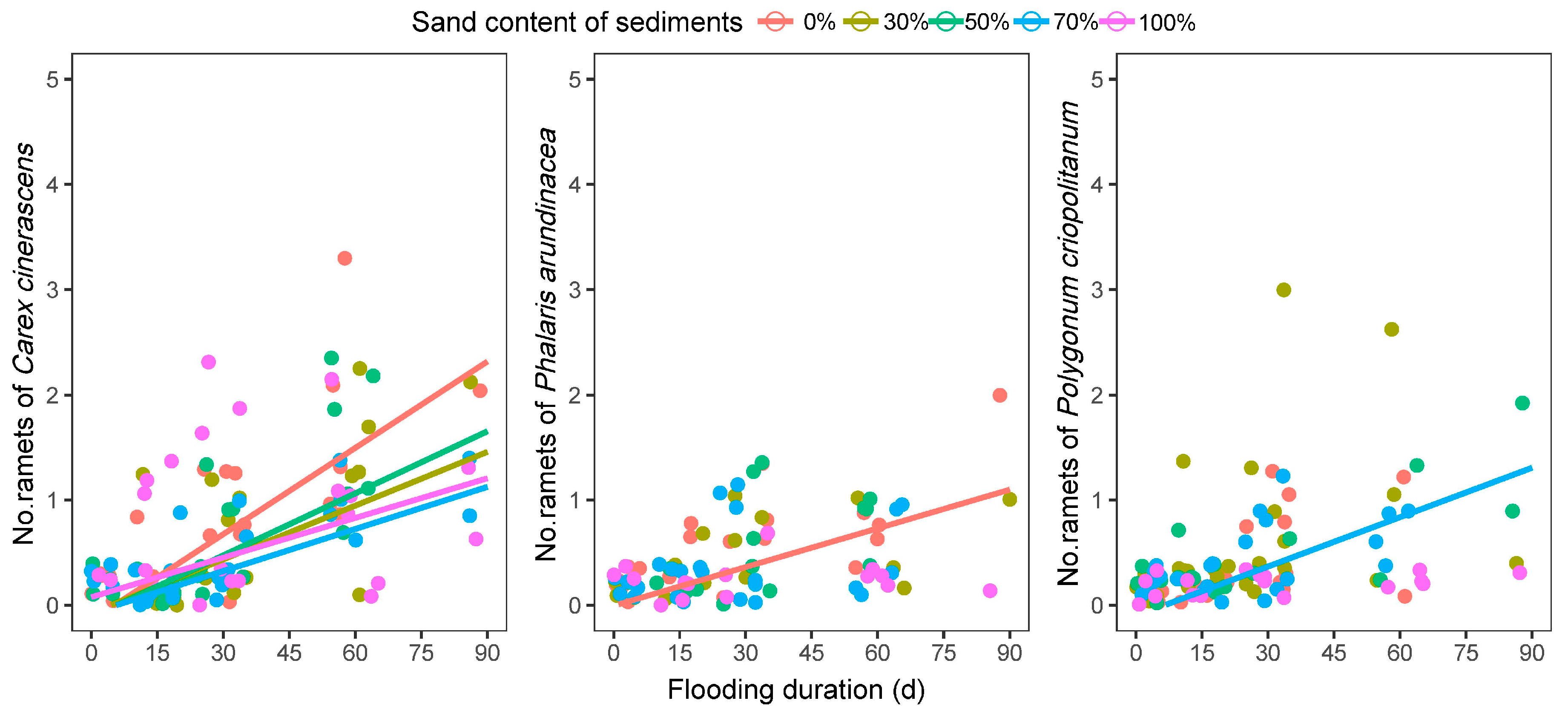
3.2.3. Number of Ramets
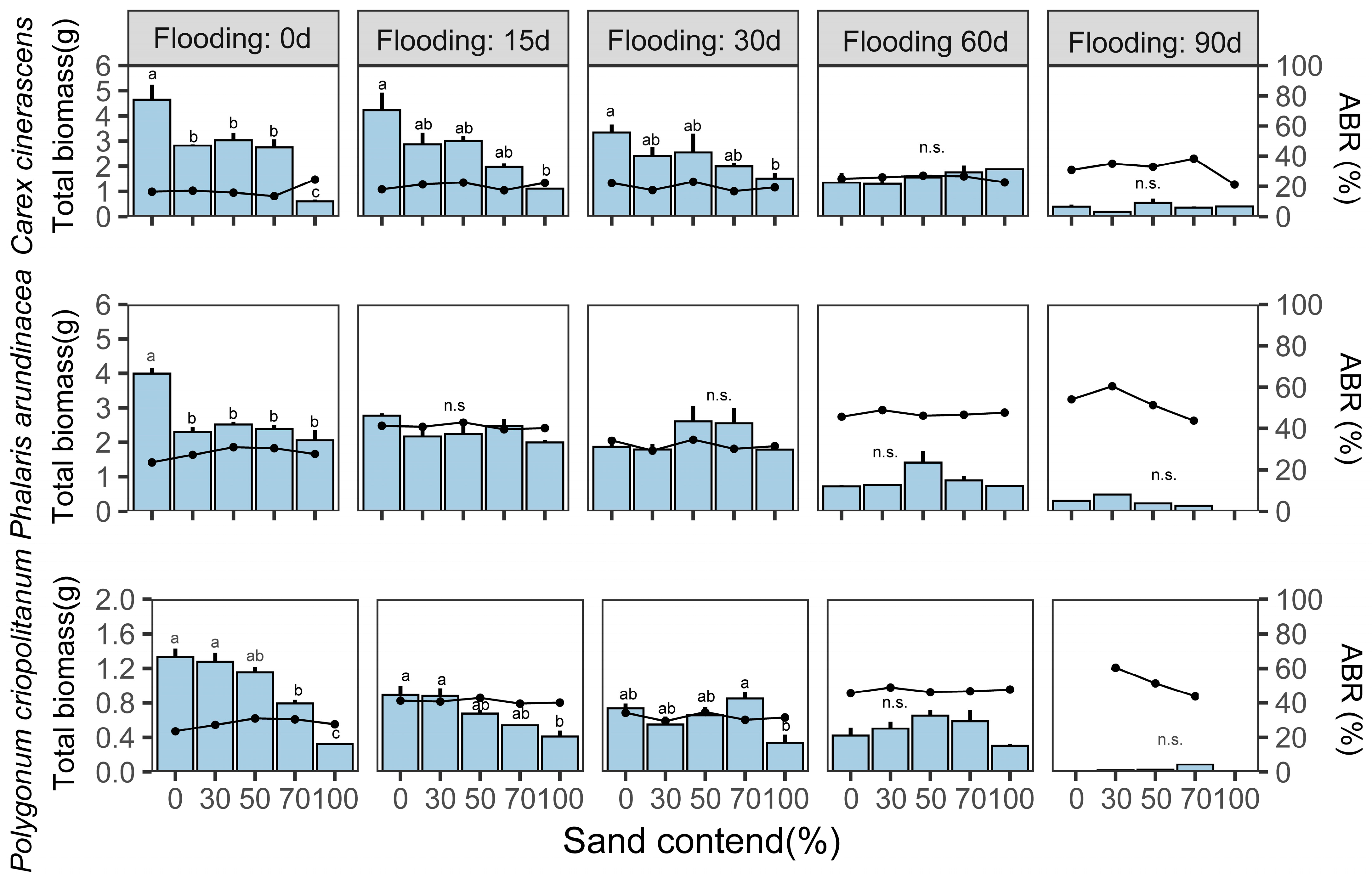
3.3. Effects of Flooding Duration and Sediment Texture on the Biomass of Three Hygrophilous Plants
4. Discussion
4.1. The Texture of Sediments Plays a Crucial Role in Shaping Functional Traits of Hygrophilous Plants Respond to Flooding
4.2. Flooding Duration and Sediment Texture Interact to Affect Wetland Plant Biomass Accumulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jinks, K.I.; Rasheed, M.A.; Brown, C.J.; Olds, A.D.; Schlacher, T.A.; Sheaves, M.; York, P.H.; Connolly, R. Saltmarsh grass supports fishery food webs in subtropical Australian estuaries. Estuar. Coast. Shelf Sci. 2020, 238, 106719. [Google Scholar] [CrossRef]
- Todd, M.J.; Muneepeerakul, R.; Pumo, D.; Azaele, S.; Miralles-Wilhelm, F.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydrological drivers of wetland vegetation community distribution within Everglades National Park, Florida. Adv. Water Resour. 2010, 33, 1279–1289. [Google Scholar] [CrossRef]
- Kopec, D.; Michalska-Hejduk, D.; Krogulec, E. The relationship between vegetation and groundwater levels as an indicator of spontaneous wetland restoration. Ecol. Eng. 2013, 57, 242–251. [Google Scholar] [CrossRef]
- Gathumbi, S.M.; Bohlen, P.J.; Graetz, D.A. Nutrient enrichment of wetland vegetation and sediments in subtropical pastures. Soil Sci. Soc. Am. J. 2005, 69, 539–548. [Google Scholar] [CrossRef]
- Zhang, Q.J.; Wang, Z.S.; Xia, S.X.; Zhang, G.S.; Li, S.X.; Yu, D.K.; Yu, X.B. Hydrologic-induced concentrated soil nutrients and improved plant growth increased carbon storage in a floodplain wetland over wet-dry alternating zones. Sci. Total Environ. 2022, 822, 153512. [Google Scholar] [CrossRef]
- Reents, S.; Mueller, P.; Tang, H.; Jensen, K.; Nolte, S. Plant genotype determines biomass response to flooding frequency in tidal wetlands. Biogeosciences 2021, 18, 403–411. [Google Scholar] [CrossRef]
- Zhou, D.Y.; Ni, Y.H.; Yu, X.N.; Lin, K.X.; Du, N.; Liu, L.L.; Guo, X.; Guo, W.H. Trait-based adaptability of Phragmites australis to the effects of soil water and salinity in the Yellow River Delta. Ecol. Evol. 2021, 11, 11352–11361. [Google Scholar] [CrossRef]
- Kim, S.; Park, H.J.; Lee, C.W.; Kim, N.Y.; Hwang, J.E.; Lee, B.D.; Bin Park, H.; An, J.E.; Baek, J. Endangered plant species under differing anthropogenic interventions: How to preserve Pterygopleurum neurophyllum in Wondong wetland? Peerj 2022, 10, e14050. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, X.; Zhang, Q.W.; Zhang, Y.L.; Liu, D.; Cai, A.D.; Zhang, X. Assessing wetland sustainability by modeling water table dynamics under climate change. J. Clean. Prod. 2020, 263, 121293. [Google Scholar] [CrossRef]
- Junk, W.J.; da Cunha, N.; Thomaz, S.M.; Agostinho, A.A.; Ferreira, F.A.; de Souza, E.E.; Stevaux, J.C.; da Silva, J.C.B.; Rocha, P.C.; Kawakita, K. Macrohabitat classification of wetlands as a powerful tool for management and protection: The example of the Parana River floodplain, Brazil. Ecohydrol. Hydrobiol. 2021, 21, 411–424. [Google Scholar] [CrossRef]
- Byun, C.; Nam, J.M.; Kim, J.G. Effects of flooding regime on wetland plant growth and species dominance in a mesocosm experiment. Plant Ecol. 2017, 218, 517–527. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- De Battisti, D.; Fowler, M.S.; Jenkins, S.R.; Skov, M.W.; Bouma, T.J.; Neyland, P.J.; Griffin, J.N. Multiple trait dimensions mediate stress gradient effects on plant biomass allocation, with implications for coastal ecosystem services. J. Ecol. 2020, 108, 1227–1240. [Google Scholar] [CrossRef]
- Hough-Snee, N.; Nackley, L.L.; Kim, S.H.; Ewing, K. Does plant performance under stress explain divergent life history strategies? The effects of flooding and nutrient stress on two wetland sedges. Aquat. Bot. 2015, 120, 151–159. [Google Scholar] [CrossRef]
- Haque, A.; Ali, G.; Macrae, M.; Badiou, P.; Lobb, D. Hydroclimatic influences and physiographic controls on phosphorus dynamics in prairie pothole wetlands. Sci. Total Environ. 2018, 645, 1410–1424. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, I.; D’Odorico, P.; Laio, F.; Ridolfi, L.; Tamea, S. Challenges in humid land ecohydrology: Interactions of water table and unsaturated zone with climate, soil, and vegetation. Water Resour. Res. 2007, 43, W0930. [Google Scholar] [CrossRef]
- Sinha, R.; Gupta, A.; Mishra, K.; Tripathi, S.; Nepal, S.; Wahid, S.M.; Swarnkar, S. Basin-scale hydrology and sediment dynamics of the Kosi river in the Himalayan foreland. J. Hydrol. 2019, 570, 156–166. [Google Scholar] [CrossRef]
- Feng, W.J.; Santonja, M.; Bragazza, L.; Buttler, A. Shift in plant-soil interactions along a lakeshore hydrological gradient. Sci. Total Environ. 2020, 742, 140254. [Google Scholar] [CrossRef]
- Reza, S.K.; Nayak, D.C.; Chattopadhyay, T.; Mukhopadhyay, S.; Singh, S.K.; Srinivasan, R. Spatial distribution of soil physical properties of alluvial soils: A geostatistical approach. Arch. Agron. Soil Sci. 2016, 62, 972–981. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, X.; Shen, C.; Meng, Z. Potamogeton crispus physiological characteristics on sediment physical properties change response. Ecol. Environ. Sci. 2012, 21, 1311–1314. [Google Scholar]
- Brotherton, S.J.; Joyce, C.B.; Berg, M.J.; Awcock, G.J. Immediate and lag effects of hydrological change on floodplain grassland plants. Plant Ecol. 2019, 220, 345–359. [Google Scholar] [CrossRef]
- Luo, W.; Song, F.; Xie, Y. Trade-off between tolerance to drought and tolerance to flooding in three wetland plants. Wetlands 2008, 28, 866–873. [Google Scholar] [CrossRef]
- Luo, F.L.; Jiang, X.X.; Li, H.L.; Yu, F.H. Does hydrological fluctuation alter impacts of species richness on biomass in wetland plant communities? J. Plant Ecol. 2016, 9, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zheng, L.D.; Pan, X.; Li, W.; Kang, X.M.; Li, J.; Ning, Y.; Zhang, M.X.; Cui, L.J. Hydrological Conditions Affect the Interspecific Interaction between Two Emergent Wetland Species. Front. Plant Sci. 2018, 8, 2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.P.; Xia, J.B.; Zhao, X.M.; Zhuge, Y.P. Soil moisture ecological characteristics of typical shrub and grass vegetation on Shell Island in the Yellow River Delta, China. Geoderma 2019, 348, 45–53. [Google Scholar] [CrossRef]
- Hu, Z.; Ge, G.; Liu, C.; Chen, F.; Li, S. Structure of Poyang Lake wetland plants ecosystem and influence of lake water level for the structure. Resour. Environ. Yangtze Basin 2010, 19, 597–605. [Google Scholar]
- Dai, X.; Wan, R.R.; Yang, G.S.; Wang, X.L.; Xu, L.G.; Li, Y.Y.; Li, B. Impact of seasonal water-level fluctuations on autumn vegetation in Poyang Lake wetland, China. Front. Earth Sci. 2019, 13, 398–409. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.Z.; Liu, Y.; Guo, H.C.; Chen, G.; Fu, Z.T.; Fu, Y.; Ge, G. Effects of Sediment types on the distribution and diversity of plant communities in the Poyang Lake wetlands. Diversity 2022, 14, 491. [Google Scholar] [CrossRef]
- Gao, J.H.; Jia, J.; Kettner, A.J.; Xing, F.; Wang, Y.P.; Xu, X.N.; Yang, Y.; Zou, X.Q.; Gao, S.; Qi, S.; et al. Changes in water and sediment exchange between the Changjiang River and Poyang Lake under natural and anthropogenic conditions, China. Sci. Total Environ. 2014, 481, 542–553. [Google Scholar] [CrossRef]
- Qi, S.; Xiong, M.; Liao, F.; Liu, G.; Zheng, H. Impacts of human activities on sediments budget in Poyang Lake. Sci. Geogr. Sin. 2016, 36, 888–894. [Google Scholar]
- Wu, C.; Tian, B.; Gao, P.; Mu, X.; Zhao, G.; Yin, D. Characteristics and driving factors of water area change of Poyang Lake during dry season in recent 40 years. J. Soil Water Conserv. 2021, 35, 177. [Google Scholar]
- Dai, Z.J.; Mei, X.F.; Darby, S.E.; Lou, Y.Y.; Li, W.H. Fluvial sediment transfer in the Changjiang (Yangtze) river-estuary depositional system. J. Hydrol. 2018, 566, 719–734. [Google Scholar] [CrossRef] [Green Version]
- De Wilde, M.; Puijalon, S.; Bornette, G. Sediment type rules the response of aquatic plant communities to dewatering in wetlands. J. Veg. Sci. 2017, 28, 172–183. [Google Scholar] [CrossRef]
- Chen, F.Q.; Li, Y.; Qie, G.W.; Xu, W.-N. The morphological responses and endurance of Polygonum hydropiper to flooding stress. Wuhan Zhiwuxue Yanjiu 2008, 26, 142–146. [Google Scholar]
- Barman, U.; Choudhury, R.D. Soil texture classification using multi class support vector machine. Inf. Process. Agric. 2020, 7, 318–332. [Google Scholar] [CrossRef]
- Li, W.; Wang, X.; He, L.; Liu, Y.; Ge, G. The responses of growth and vegetative reproduction of wetland plants in Poyang Lake to duration of submergence. Acta Ecol. Sin. 2018, 38, 8176–8183. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: https://www.R-project.org/ (accessed on 8 April 2021).
- Laanaya, F.; St-Hilaire, A.; Gloaguen, E. Water temperature modelling: Comparison between the generalized additive model, logistic, residuals regression and linear regression models. Hydrol. Sci. J.-J. Des Sci. Hydrol. 2017, 62, 1078–1093. [Google Scholar] [CrossRef]
- Wu, L.W.; Zhang, Y.; Guo, X.; Ning, D.L.; Zhou, X.S.; Feng, J.J.; Yuan, M.M.; Liu, S.; Guo, J.J.; Gao, Z.P.; et al. Reduction of microbial diversity in grassland soil is driven by long-term climate warming. Nat. Microbiol. 2022, 7, 1054–1062. [Google Scholar] [CrossRef]
- Heminway, A.W.; Wilcox, D.A. Response of Typha to phosphorus, hydrology, and land use in Lake Ontario coastal wetlands and a companion greenhouse study. Wetl. Ecol. Manag. 2022, 30, 1167–1180. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Ghosh, J.K. AIC, BIC and recent advances in model selection. In Philosophy of Statistics; Bandyopadhyay, P.S., Forster, M.R., Eds.; North-Holland: Amsterdam, The Netherlands, 2011; Volume 7, pp. 583–605. [Google Scholar]
- Mendonça, F.; Lucena, M.; Quirino, R.; Govêia, C.; Guimaraes, G. Risk factors for postoperative hypothermia in the post-anesthetic care unit: A prospective prognostic pilot study. Braz. J. Anesthesiol. (Engl. Ed.) 2018, 69, 122–130. [Google Scholar] [CrossRef]
- Richard, M.H. HH: Statistical analysis and data display: Heiberger and Holland. Available online: https://CRAN.R-project.org/package=HH (accessed on 30 May 2022).
- John Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, S.; Zhang, X.; Mao, L. glmm.hp: An R package for computing individual effect of predictors in generalized linear mixed models. J. Plant Ecol. 2022, 15, 1302–1307. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing AG: Houston, TX, USA, 2016. [Google Scholar]
- An, Y.; Gao, Y.; Tong, S.Z.; Liu, B. Morphological andphysiological traits related to the response and adaption of Bolboschoenus planiculmis seedlings grown under salt-alkaline stress conditions. Front. Plant Sci. 2021, 12, 567782. [Google Scholar] [CrossRef] [PubMed]
- Gentine, J.A.; Conard, W.M.; O’Reilly, K.E.; Cooper, M.J.; Fiorino, G.E.; Harrison, A.M.; Hein, M.; Moerke, A.H.; Iii, C.R.R.; Uzarski, D.G.; et al. Environmental predictors of phytoplankton chlorophyll-a in Great Lakes coastal wetlands. J. Great Lakes Res. 2022, 48, 927–934. [Google Scholar] [CrossRef]
- Robertson, A.I.; Bacon, P.; Heagney, G. The responses of floodplain primary production to flood frequency and timing. J. Appl. Ecol. 2001, 38, 126–136. [Google Scholar] [CrossRef]
- Batistel, C.; Porsche, C.; Jurasinski, G.; Schubert, H. Responses of four peatland emergent macrophytes to salinity and short salinity pulses. Wetlands 2022, 42, 67. [Google Scholar] [CrossRef]
- Mentges, M.I.; Reichert, J.M.; Rodrigues, M.F.; Awe, G.O.; Mentges, L.R. Capacity and intensity soil aeration properties affected by granulometry, moisture, and structure in no-tillage soils. Geoderma 2016, 263, 47–59. [Google Scholar] [CrossRef]
- Mats’ela, K.; Olaleye, A.O.; Rathebe, K.; Rasekoele, M.; Ntlele, M.; Pheko, T.; Odunuga, B.O. Morphological properties and soil nutrient changes in selected properties in two contrasting wetlands in lesotho. Commun. Soil Sci. Plant Anal. 2015, 46, 2274–2294. [Google Scholar] [CrossRef]
- Wallington, C.A.; Scholes, M.C. Pans as possible pedological hotspots in an arid African savanna. Wetl. Ecol. Manag. 2021, 29, 519–530. [Google Scholar] [CrossRef]
- Xie, Y.H.; Ren, B.; Li, F. Increased nutrient supply facilitates acclimation to high-water level in the marsh plant Deyeuxia angustifolia: The response of root morphology. Aquat. Bot. 2009, 91, 1–5. [Google Scholar] [CrossRef]
- Kretz, L.; Koll, K.; Seele-Dilbat, C.; van der Plas, F.; Weigelt, A.; Wirth, C. Plant structural diversity alters sediment retention on and underneath herbaceous vegetation in a flume experiment. PLoS ONE 2021, 16, e0248320. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.S.; Liao, Y.L.; Xie, Y.H.; Li, F.; Deng, Z.M.; Hou, Z.Y.; Wu, C. Concurrent Effects of sediment accretion and nutrient availability on the clonal growth strategy of Carex brevicuspis—A wetland sedge that produces both spreading and clumping ramets. Front. Plant Sci. 2017, 8, 1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Chen, J.; Cheng, P. Response of Carex cinerascens to Waterlogging Stress. J. Hydroecology 2017, 38, 24–29. [Google Scholar]
- Billah, M.M.; Bhuiyan, M.K.A.; Islam, M.A.; Das, J.; Hoque, A. Salt marsh restoration: An overview of techniques and success indicators. Environ. Sci. Pollut. Res. 2022, 29, 15347–15363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.J.; Zhang, M.Y.; Tong, S.Z.; Qi, Q.; Wang, X.H.; Lu, X.G. Growth and physiological responses of Carex schmidtii to water-level fluctuation. Hydrobiologia 2020, 847, 967–981. [Google Scholar] [CrossRef]
- Luo, F.L.; Nagel, K.A.; Scharr, H.; Zeng, B.; Schurr, U.; Matsubara, S. Recovery dynamics of growth, photosynthesis and carbohydrate accumulation after de-submergence: A comparison between two wetland plants showing escape and quiescence strategies. Ann. Bot. 2011, 107, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.S.; Tang, H.R.; Chen, F.Y.; Lou, Y.J. Functional traits response to flooding depth and nitrogen supply in the helophyte Glyceria spiculosa (Gramineae). Aquat. Bot. 2021, 175, 103449. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H.; Al-Sodany, Y.M.; Haroun, S.A.; Jensen, K. A comparison of the functional traits of Phragmites australis in Lake Burullus (a ramsar site in Egypt): Young vs. old populations over the nutrient availability gradient. Ecol. Eng. 2021, 166, 106244. [Google Scholar] [CrossRef]
- Sabaj, V.; Conde, D.; Rodriguez-Gallego, L.; Kandus, P. Postharvest growth dynamic of Schoenoplectus californicus along fluvio-estuarine and flooding gradients. Wetl. Ecol. Manag. 2018, 26, 125–138. [Google Scholar] [CrossRef]
- McCoy, M.M.; Sloey, T.M.; Howard, R.J.; Hester, M.W. Response of tidal marsh vegetation to pulsed increases in flooding and nitrogen. Wetl. Ecol. Manag. 2020, 28, 119–135. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| y | Fixed Factor | Random Factor | |
|---|---|---|---|
| Sediments | Flooding | Species | |
| Total biomass | −0.50 *** | −0.93 *** | 0.47 *** |
| Above-ground biomass | −0.48 *** | −0.89 *** | 0.38 *** |
| Below-ground biomass | −0.46 *** | −0.86 *** | 0.45 *** |
| ABR | −0.03 | 0.40 *** | −0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, J.; Liu, Y.; He, L.; Yang, S.; Gong, H.; Xu, R.; Yao, X.; Ge, G. Interactive Effects of Flooding Duration and Sediment Texture on the Growth and Adaptation of Three Plant Species in the Poyang Lake Wetland. Biology 2023, 12, 944. https://doi.org/10.3390/biology12070944
Liu Y, Li J, Liu Y, He L, Yang S, Gong H, Xu R, Yao X, Ge G. Interactive Effects of Flooding Duration and Sediment Texture on the Growth and Adaptation of Three Plant Species in the Poyang Lake Wetland. Biology. 2023; 12(7):944. https://doi.org/10.3390/biology12070944
Chicago/Turabian StyleLiu, Ying, Jie Li, Yizhen Liu, Liang He, Shanshan Yang, Huiying Gong, Ruixin Xu, Xingzi Yao, and Gang Ge. 2023. "Interactive Effects of Flooding Duration and Sediment Texture on the Growth and Adaptation of Three Plant Species in the Poyang Lake Wetland" Biology 12, no. 7: 944. https://doi.org/10.3390/biology12070944