Salt Tolerance in Machilus faberi: Elucidating Growth and Physiological Adaptations to Saline Environments


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Salt Stress Treatment Using Sodium Chloride
2.3. Morphological Measurements
2.4. Chlorophyll Determination
2.5. Determination of Pn and Gs
2.6. Determination of Water Content, Relative Conductivity, Malondialdehyde Content, and Root Activity of Plant Leaves
2.7. The Activities of SOD, POD, and APX Were Determined
2.8. The Contents of Pro, SS, and SP Were Determined
2.9. Statistical Analysis
3. Results
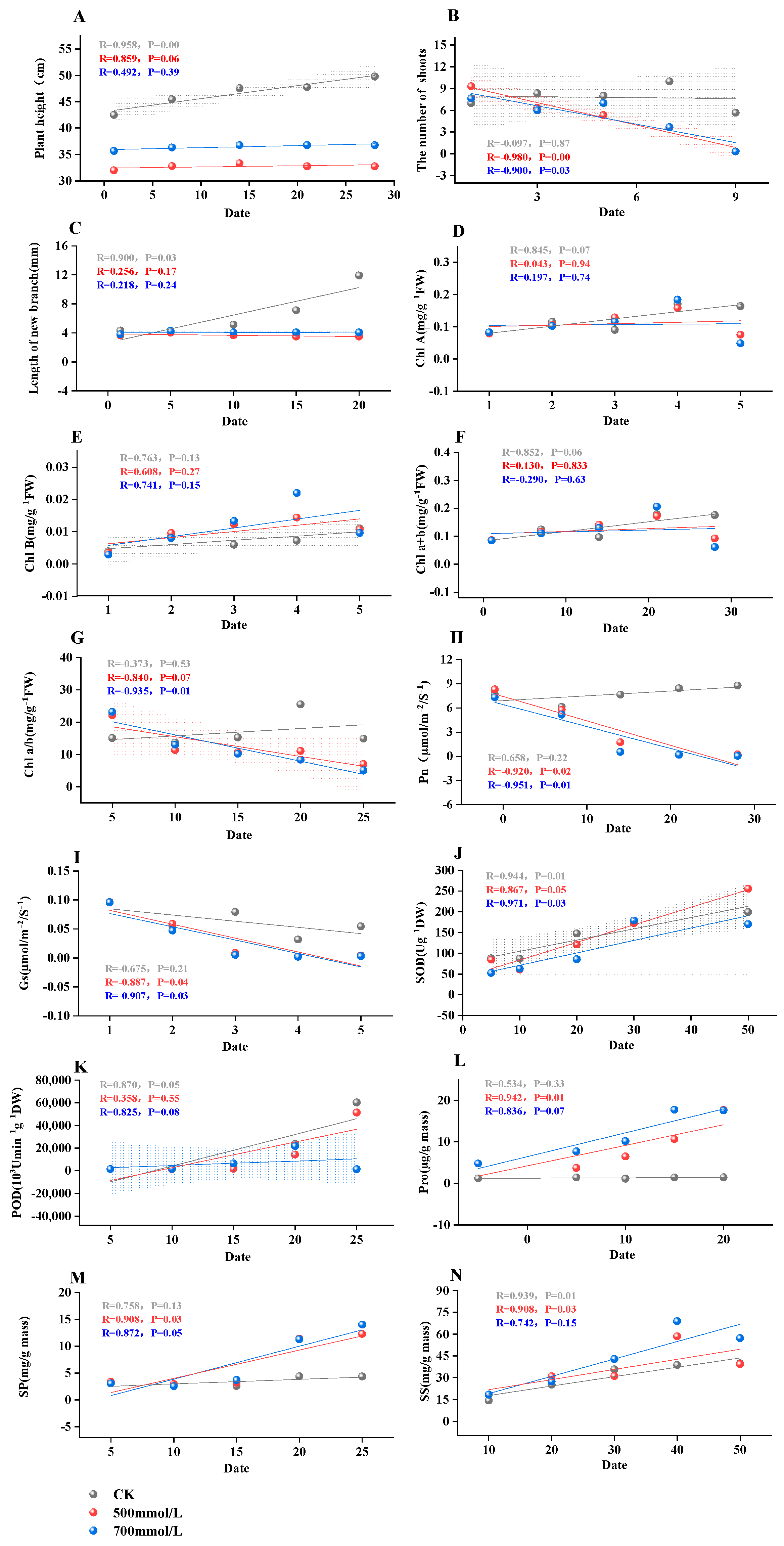
3.1. Observation of Leaf Phenotype and Change in Growth Index
3.2. Changes in Chlorophyll Content in Leaves of M. faberi
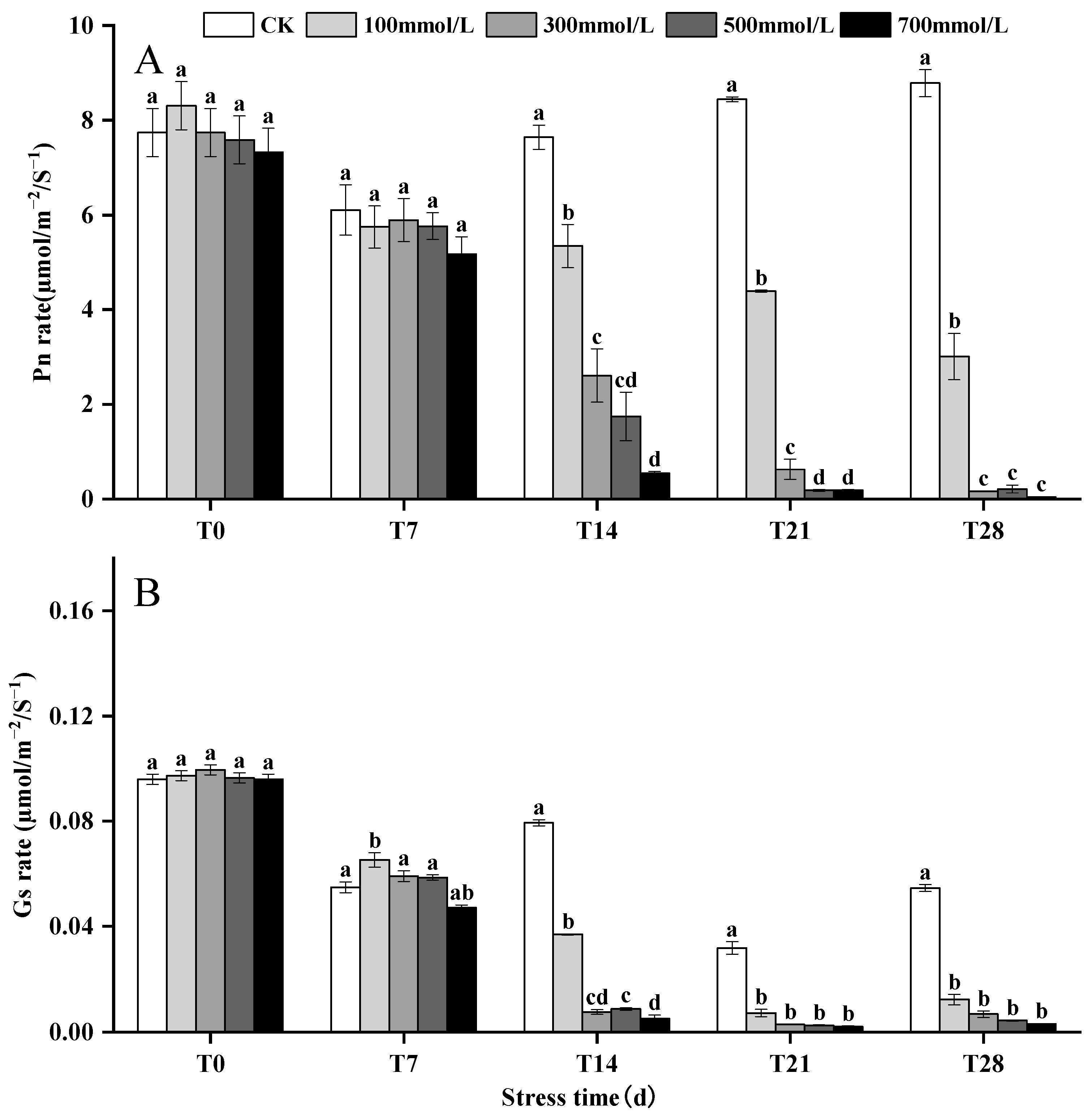
3.3. Effects of Salt Stress on Pn and Gs
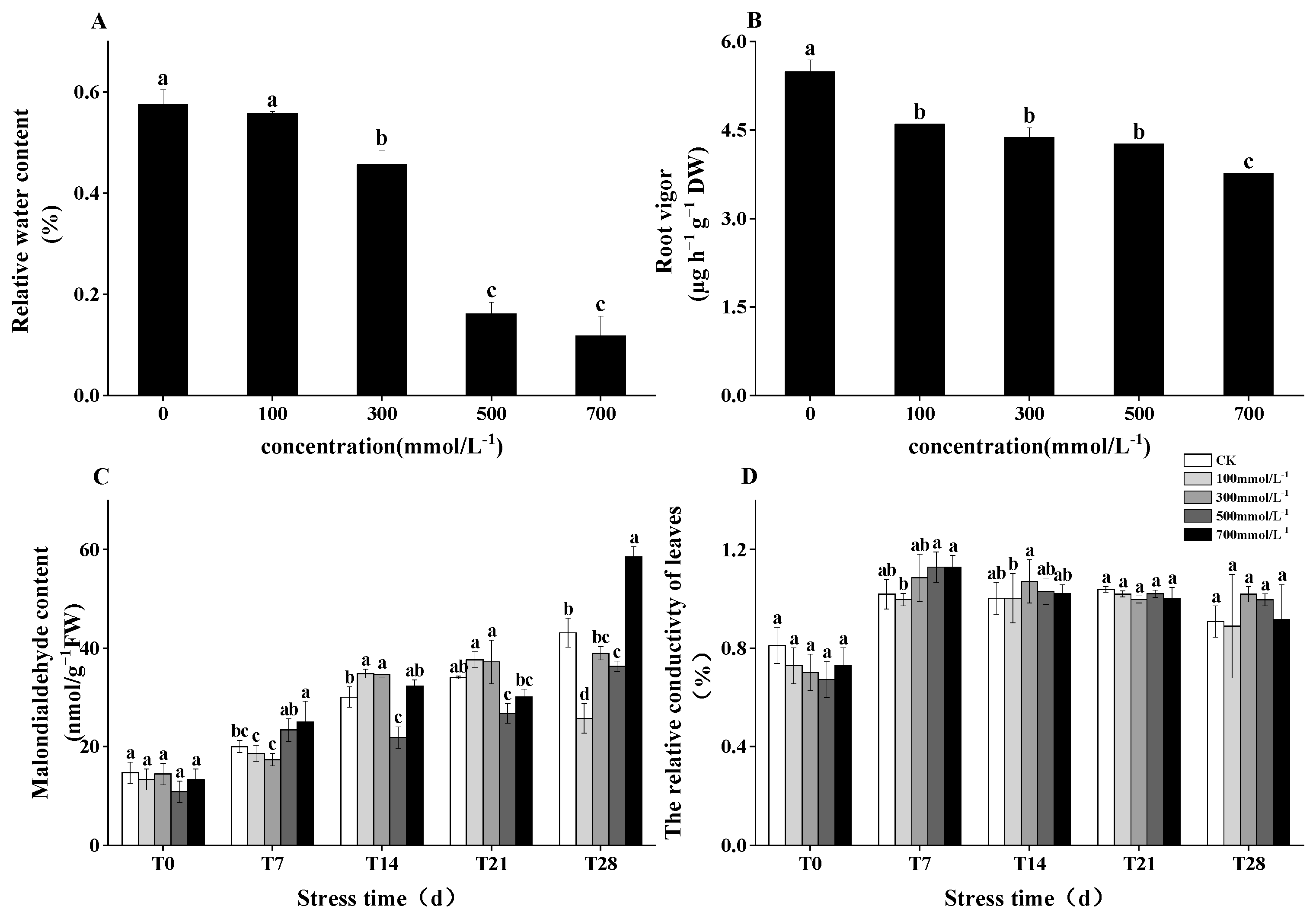
3.4. Root Activity, MDA Content, and Electrical Conductivity in Response to Salt Stress
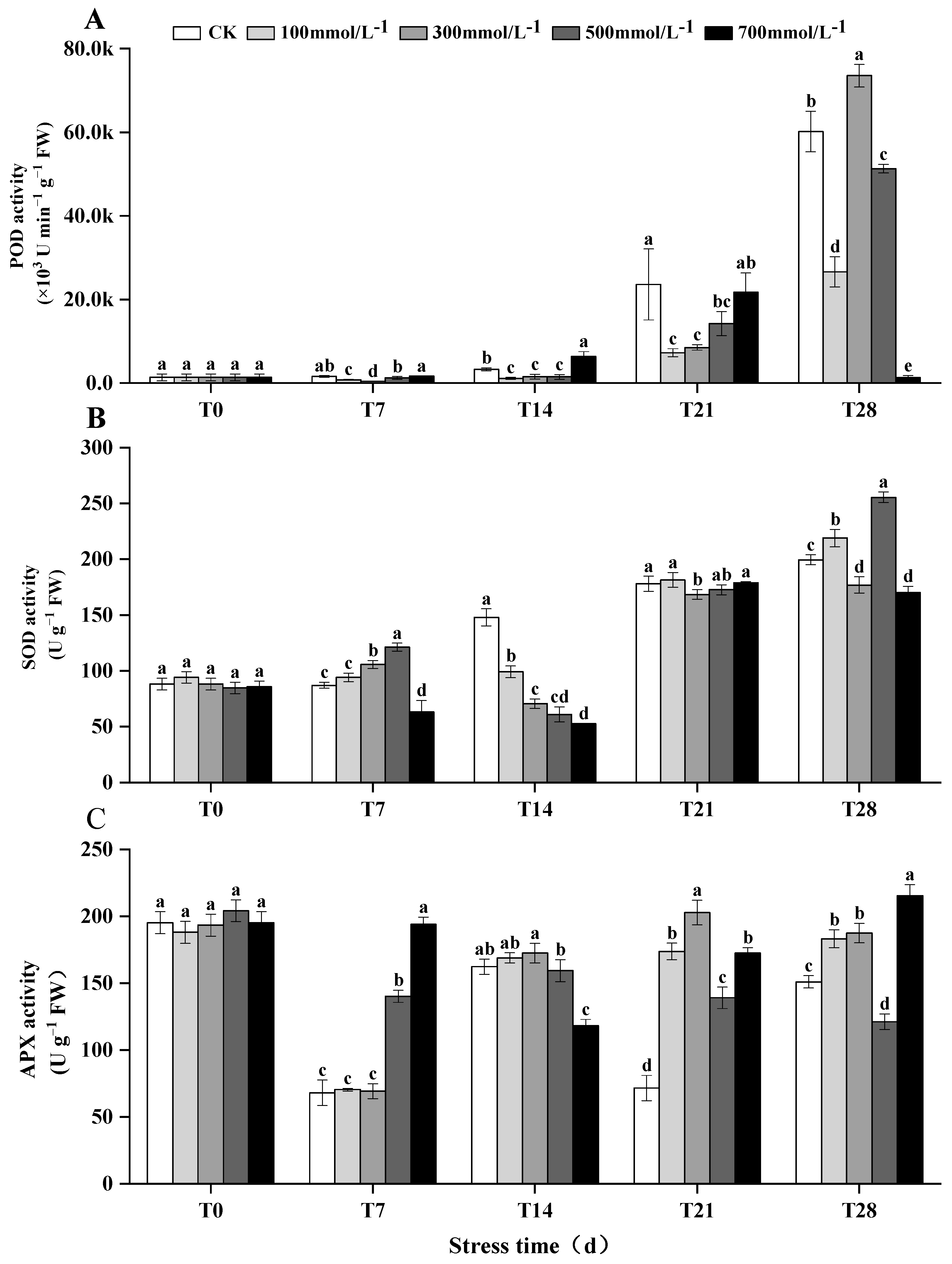
3.5. Enzymatic Responses in M. faberi Leaves under Salt Stress
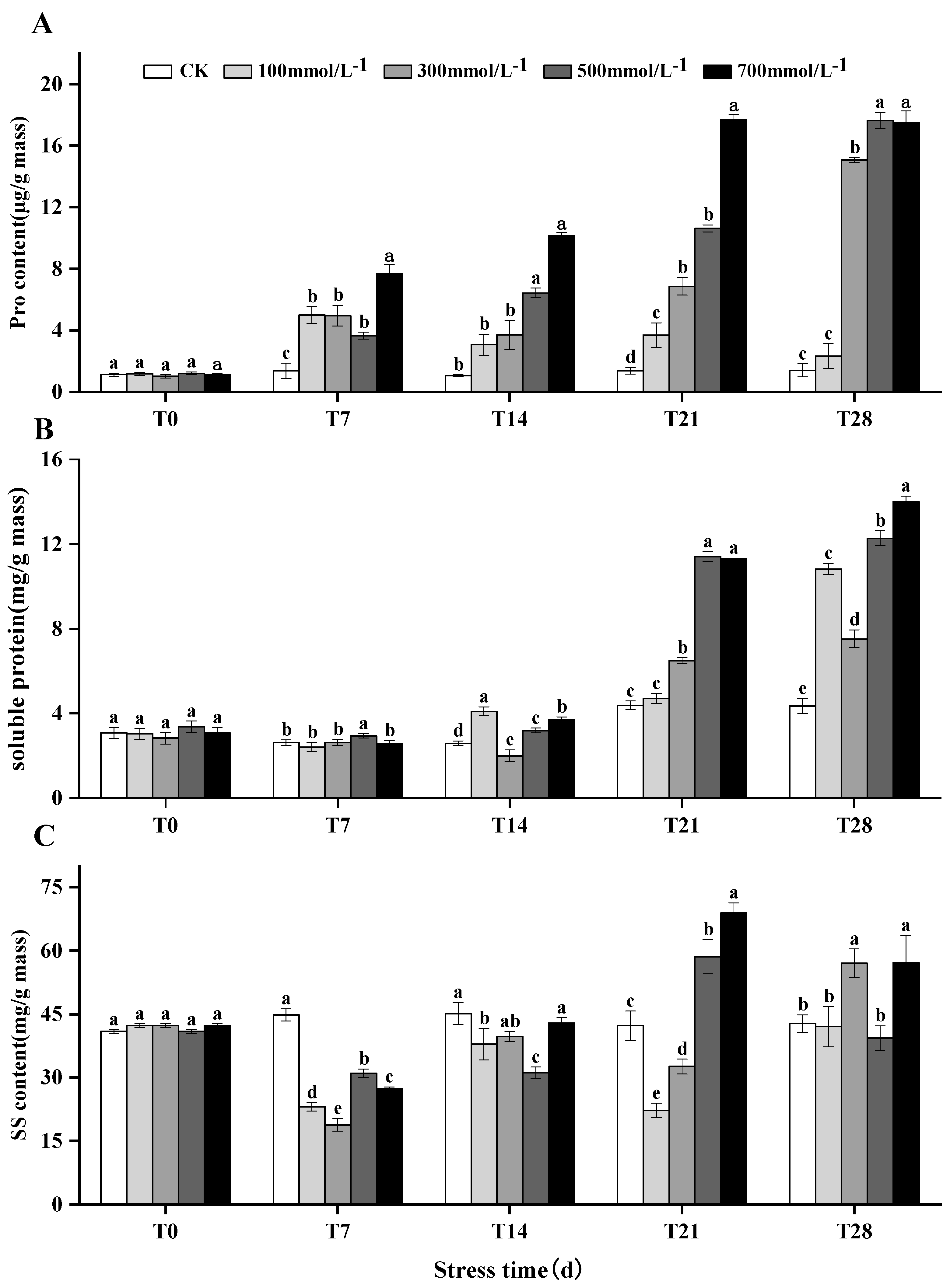
3.6. Accumulation of Soluble Proteins, Proline, and Soluble Sugars in M. faberi under Salt Stress
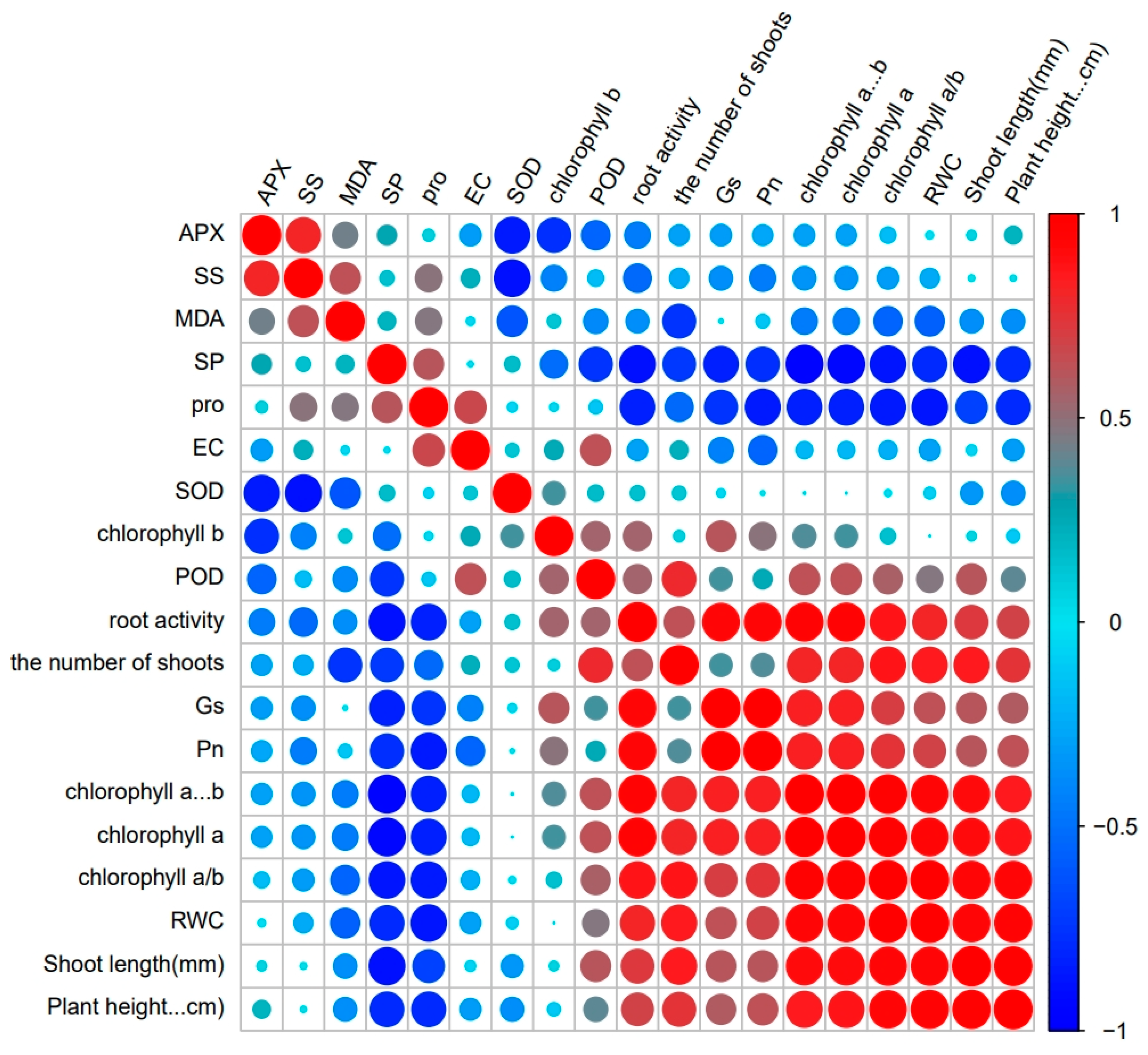
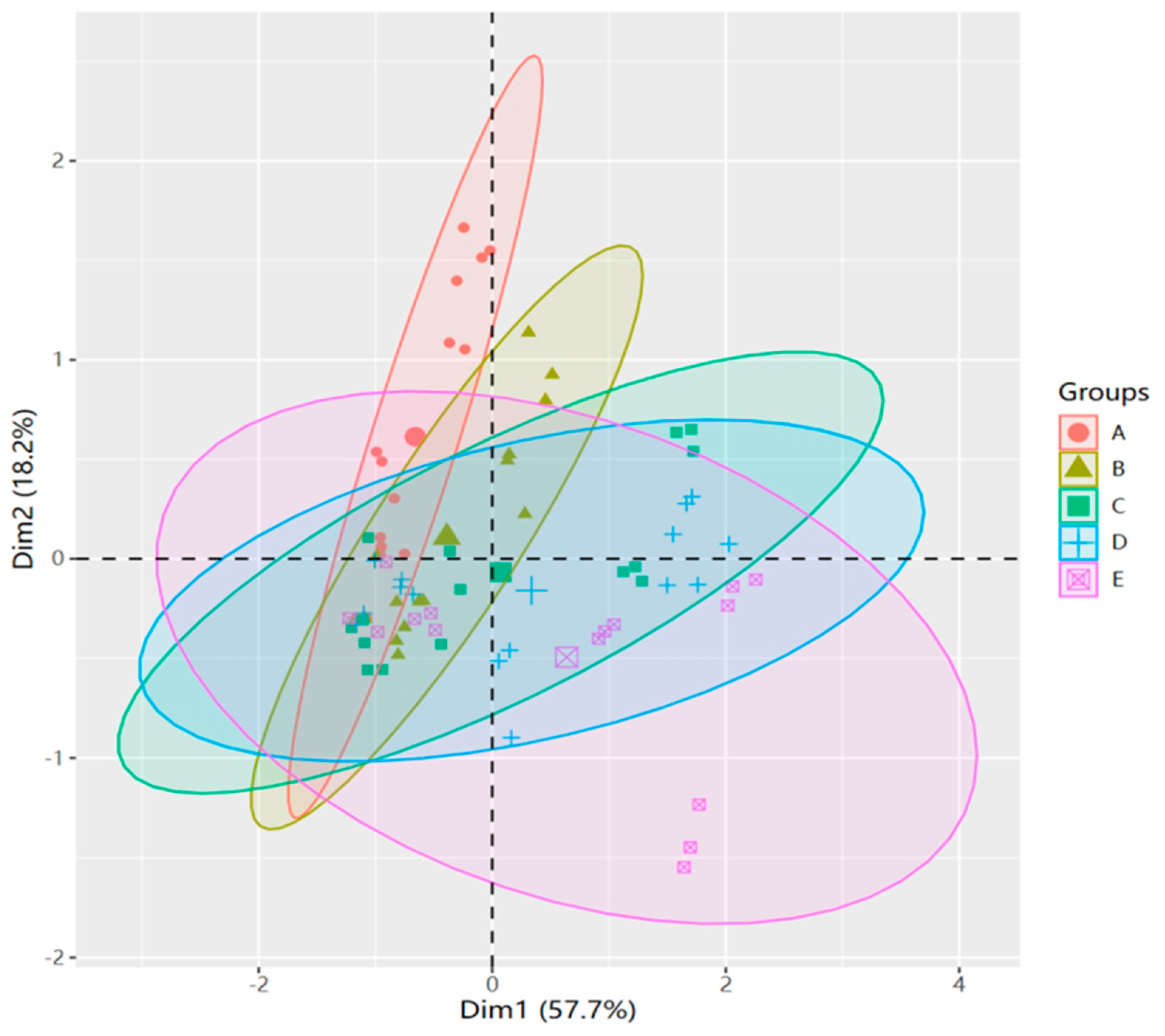
3.7. Principal Component Analysis, Correlation, and Regression Insights in M. faberi under Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Leonardi, C.; Bie, Z. Role of grafting in vegetable crops grown under saline conditions. Sci. Hortic. 2010, 127, 147–155. [Google Scholar] [CrossRef]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Othman, Y.A.; Hani, M.B.; Ayad, J.Y.; St Hilaire, R. Salinity level influenced morpho-physiology and nutrient uptake of young citrus rootstocks. Heliyon 2023, 9, e13336. [Google Scholar] [CrossRef]
- Zhang, S.-H.; Xu, X.-F.; Sun, Y.-M.; Zhang, J.-L.; Li, C.-Z. Influence of drought and salt stress on the growth of young. New For. 2018, 17, 336–347. [Google Scholar]
- Garg, A.K.; Kim, J.-K.; Owens, T.G.; Ranwala, A.P. Trehalose accumulation in rice plants confers hightolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 10, 99. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Litalien, A.; Zeeb, B. Curing the earth: A review of anthropogenic soil salinization and plant-based strategies for sustainable mitigation. Sci. Total Environ. 2020, 698, 134235. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, F.; Zhang, H.; Ren, S. Chemical constituents and antifungal activities of essential oil from leaves of Phoebe faberi. Guihaia 2000, 20, 181–184. [Google Scholar]
- Qunsheng, Z. Transplanting technology of Machilus faberi Hemsley tree. Flower Plant Penjing 2011, 7, 43–46, 43–44,46. [Google Scholar]
- Jinjing, C. Study on Transplanting Cultivation Techniques of Greening Seedlings of Machilus bambusoides. Anhui Agric. Sci. Bull. 2013, 19, 110–112. [Google Scholar]
- Xiangjun, C. Study on Forestland Selection and Fertilization Experiment of Machilus faberi Hemsley; Hunan Forestry Science & Technology: Changsha, China, 1998; pp. 27–31. [Google Scholar]
- Karimi, S.; Karami, H.; Vahdati, K.; Mokhtassi-Bidgoli, A. Antioxidative responses to short-term salinity stress induce drought tolerance in walnut. Sci. Hortic. 2020, 267, 109322. [Google Scholar] [CrossRef]
- Yue, D. Study on extraction method of chlorophyll from rape pods. Crop Res. 2023, 37, 10–13. [Google Scholar]
- Chen, A.K.; Han, R.H.; Li, D.Y.; Lin, L.L.; Luo, H.X.; Tang, S.J. A Comparison of Two Methods for Electrical Conductivity about Plant leaves. J. Guangdong Univ. Educ. 2010, 30, 88–91. [Google Scholar]
- Zhang, Q.; Wu, X.; Zheng, J.; Sun, M. Progress of Researches on Methods FOR Determination of Malondialdenhyde in Biological Sampies. Phys. Test. Chem. Anal. (Part B Chem. Anal.) 2016, 52, 979–985. [Google Scholar]
- Zhu, X.; Liang, M.; Ma, Y. A Review Report on the Experiments for the Determination of Root Activiyy by TTC Method. Guangdong Chem. Ind. 2020, 47, 211–212. [Google Scholar]
- Han, Y.; Fan, S.; Zhang, Q.; Wang, Y. Effect of heat stress on the MDA, proline and soluble sugar content in leaf lettuce seedlings. Agric. Sci. 2013, 4, 112–115. [Google Scholar] [CrossRef]
- Yue, S.-Y.; Zhou, R.-R.; Nan, T.-G.; Huang, L.-Q.; Yuan, Y. Comparison of major chemical components in Puerariae Thomsonii Radix and Puerariae Lobatae Radix. China J. Chin. Mater. Med. 2022, 47, 2689–2697. [Google Scholar] [CrossRef]
- Ribeiro, H.; Duque, L.; Sousa, R.; Abreu, I. Ozone effects on soluble protein content of Acer negundo, Quercus robur and Platanus spp. pollen. Aerobiologia 2013, 29, 443–447. [Google Scholar] [CrossRef]
- Bayomy, H.M.; Alamri, E.S.; Alharbi, B.M.; Foudah, S.H.; Genaidy, E.A.; Atteya, A.K. Response of Moringa oleifera trees to salinity stress conditions in Tabuk region, Kingdom of Saudi Arabia. Saudi J. Biol. Sci. 2023, 30, 103810. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (Oryza sativa L.) varieties. Biomed. Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [PubMed]
- Fatima, N.; Akram, M.; Shahid, M.; Abbas, G.; Hussain, M.; Nafees, M.; Wasaya, A.; Tahir, M.; Amjad, M. Germination, growth and ions uptake of moringa (Moringa oleifera L.) grown under saline condition. J. Plant Nutr. 2018, 41, 1555–1565. [Google Scholar] [CrossRef]
- Farooq, F.; Rashid, N.; Ibrar, D.; Hasnain, Z.; Ullah, R.; Nawaz, M.; Irshad, S.; Basra, S.M.A.; Alwahibi, M.S.; Elshikh, M.S.; et al. Impact of varying levels of soil salinity on emergence, growth and biochemical attributes of four Moringa oleifera landraces. PLoS ONE 2022, 17, e0263978. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef]
- Ghogdi, E.A.; lzadi-Darbandi, A.; Borzouei, A. Effects of salinity on some physiological traits in wheat (Triticum aestivum L.) cultivars. Indian J. Sci. Technol. 2012, 5, 1901–1906. [Google Scholar] [CrossRef]
- Dos Santos Araujo, G.; de Oliveira Paula-Marinho, S.; de Paiva Pinheiro, S.K.; de Castro Miguel, E.; de Sousa Lopes, L.; Camelo Marques, E.; de Carvalho, H.H.; Gomes-Filho, E. H2O2 priming promotes salt tolerance in maize by protecting chloroplasts ultrastructure and primary metabolites modulation. Plant Sci. 2021, 303, 110774. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Bi, Y.; Wang, L.; Tang, L.; Yu, X.; Ohtani, M.; Demura, T.; Zhuge, Q. Overexpression of PtSOS2 Enhances Salt Tolerance in Transgenic Poplars. Plant Mol. Biol. Rep. 2014, 32, 185–197. [Google Scholar] [CrossRef]
- Zhang, L.X.; Chang, Q.S.; Hou, X.G.; Liu, W.; Li, X.P.; Gao, Y.H.; Zhang, X.-L.; Ding, S.-Y.; Xiao, R.-X.; Zhang, Y.; et al. Effects of NaCl stress on antioxidant capacity and photosynthetic characteristics of Prunella vulgaris seedlings. Acta Prataculturae Sin. 2017, 26, 167–175. [Google Scholar] [CrossRef]
- Lu, W.; Wei, G.; Zhou, B.; Liu, J.; Zhang, S.; Guo, J. A comparative analysis of photosynthetic function and reactive oxygen species metabolism responses in two hibiscus cultivars under saline conditions. Plant Physiol. Biochem. 2022, 184, 87–97. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, S.; Wei, M.; Gong, B.; Shi, Q. Effect of Different Rootstocks on the Salt Stress Tolerance in Watermelon Seedlings. Hortic. Plant J. 2018, 4, 239–249. [Google Scholar] [CrossRef]
- Penella, C.; Landi, M.; Guidi, L.; Nebauer, S.G.; Pellegrini, E.; San Bautista, A.; Remorini, D.; Nali, C.; Lopez-Galarza, S.; Calatayud, A. Salt-tolerant rootstock increases yield of pepper under salinity through maintenance of photosynthetic performance and sinks strength. J. Plant Physiol. 2016, 193, 1–11. [Google Scholar] [CrossRef]
- Soliman, S.; Hall, R.C. Forensic issues in medical evaluation: Competency and end-of-life issues. Adv. Psychosom. Med. 2015, 34, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, G.; Shen, W.; Li, X.; Qi, J.; Xu, J.; Tao, A.; Liu, X. Effect of Salt Stress on the Growth and Antioxidant Enzyme Activity of Kenaf Seedlings. PLant Sci. J. 2011, 29, 493–501. [Google Scholar] [CrossRef]
- Wasim, M.A.; Naz, N.; Zehra, S. Anatomical characteristic, ionic contents and nutritional potential of Buffel grass (Cenchrus ciliaris L.) under high salinity. S. Afr. J. Bot. 2022, 144, 471–479. [Google Scholar] [CrossRef]
- Heyno, E.; Mary, V.; Schopfer, P.; Krieger-Liszkay, A. Oxygen activation at the plasma membrane: Relation between superoxide and hydroxyl radical production by isolated membranes. Planta 2011, 234, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 2027–2038. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Gan, X.; Luo, H.; Zhang, Y.; Zhang, W. Effects of Exogenous Nitric Oxide on Growth, Antioxidant System and Photosynthetic Characterics in Seedling of Cotton Cultivar Under Chilling Lnjury Stress. Sci. Agric. Sin. 2012, 45, 3058–3067. [Google Scholar] [CrossRef]
- Thornalley, P.J. The glyoxalase system new developments towards functional characterization of a metabolic pathway fundamental to biological lifetrue. Biochem. J. 1990, 269, 1–11. [Google Scholar] [CrossRef]
- Guo, Q.; Wu, Y.; Lin, Y.; Zheng, H. Effect of NaCl stress on growth and antioxidant systems of Pogostemon cablin. China J. Chin. Mater. Med. 2009, 34, 530–534. [Google Scholar]
- Mubushar, M.; El-Hendawy, S.; Tahir, M.U.; Alotaibi, M.; Mohammed, N.; Refay, Y.; Tola, E. Assessing the Suitability of Multivariate Analysis for Stress Tolerance Indices, Biomass, and Grain Yield for Detecting Salt Tolerance in Advanced Spring Wheat Lines Irrigated with Saline Water under Field Conditions. Agronomy 2022, 12, 3084. [Google Scholar] [CrossRef]
- Zhang, S.-H.; Xu, X.-F.; Sun, Y.-M.; Zhang, J.-L.; Li, C.-Z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef]
- Gao, G.; Tester, M.A.; Julkowska, M.M. The Use of High-Throughput Phenotyping for Assessment of Heat Stress-Induced Changes in Arabidopsis. Plant Phenomics 2020, 2020, 3723916. [Google Scholar] [CrossRef]
- Al-Farsi, S.M.; Nawaz, A.; Anees ur, R.; Nadaf, S.K.; Al-Sadi, A.M.; Siddique, K.H.M.; Farooq, M. Effects, tolerance mechanisms and management of salt stress in lucerne (Medicago sativa). Crop Pasture Sci. 2020, 71, 411–428. [Google Scholar] [CrossRef]
- Liu, X.; Su, S. Growth and Physiological Response of Viola tricolor L. to NaCl and NaHCO3 Stress. Plants 2023, 12, 178. [Google Scholar] [CrossRef]
- Meng, S.; Xu, T.; Zhu, X.; Di, J.; Zhu, Y.; Yang, X.; Zou, S.; Yang, X.; Qin, C.; Yan, W. Diversity Analysis of Soybean Landraces Collected from Jiangsu Province Using Phenotypic Traits. J. Plant Genet. Resour. 2023, 24, 419–436. [Google Scholar] [CrossRef]
- Liang, F.; Huang, Q.W.; Yu, Y.P.; Liang, H.; Huang, Q.Y.; Liu, X.M.; Chen, Q.Y.; Tan, X.H. Physiological response of endangered semi-mangrove Barringtonia racemosa to salt stress and its correlation analysis. J. Cent. South Univ. For. Technol. 2019, 39, 12–18. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphologic Characteristics | Days | T0 | T7 | T14 | T21 | T28 |
|---|---|---|---|---|---|---|
| Concentration (mmol−1/L) | ||||||
| Number of new shoots (pcs) | CK | 7.00 ± 1.73a | 8.33 ± 1.89a | 8.00 ± 1.46ab | 10.00 ± 1.66a | 5.67 ± 1.04a |
| 100 | 8.67 ± 1.53a | 12.33 ± 1.11a | 10.33 ± 1.21a | 7.33 ± 1.13a | 6.00 ± 1.00a | |
| 300 | 13.00 ± 1.00a | 11.33 ± 1.53a | 10.33 ± 1.16a | 8.00 ± 1.36 | 7.00 ± 1.73a | |
| 500 | 9.33 ± 1.53a | 6.33 ± 1.31a | 5.33 ± 1.15ab | 3.67 ± 1.52a | 3.67 ± 1.52a | |
| 700 | 7.67 ± 1.08a | 6.00 ± 1.00a | 1.00 ± 1.00b | 0.33 ± 0.58a | 0.33 ± 0.58a | |
| Length of new branch (mm) | CK | 4.32 ± 0.59a | 4.23 ± 0.66a | 5.13 ± 0.95a | 7.10 ± 1.95a | 16.94 ± 5.56a |
| 100 | 3.60 ± 0.17a | 3.50 ± 0.51a | 3.53 ± 1.25a | 4.25 ± 0.50b | 12.98 ± 5.11a | |
| 300 | 3.58 ± 1.40a | 7.39 ± 4.34a | 3.82 ± 1.49a | 3.81 ± 0.28b | 16.39 ± 7.45b | |
| 500 | 3.65 ± 0.94a | 3.97 ± 0.19a | 3.66 ± 0.92a | 3.49 ± 1.18b | 3.49 ± 1.18b | |
| 700 | 3.79 ± 0.53a | 4.26 ± 1.34a | 4.05 ± 0.99a | 4.05 ± 0.99b | 4.05 ± 0.99a | |
| Plant height (cm) | CK | 42.51 ± 6.21a | 45.47 ± 5.91a | 47.56 ± 6.10a | 47.77 ± 5.91a | 49.77 ± 3.36a |
| 100 | 41.00 ± 8.77a | 42.98 ± 8.501a | 44.94 ± 8.71a | 44.66 ± 7.64ab | 48.13 ± 8.78ab | |
| 300 | 39.97 ± 10.61a | 43.00 ± 11.01a | 44.11 ± 12.49a | 43.37 ± 12.77ab | 47.57 ± 10.37ab | |
| 500 | 31.98 ± 3.94a | 32.80 ± 4.10a | 33.34 ± 4.20a | 32.78 ± 3.61b | 32.78 ± 3.61c | |
| 700 | 35.67 ± 3.11a | 36.31 ± 2.50a | 36.79 ± 2.84a | 36.79 ± 2.84ab | 36.79 ± 2.84b |
| Chl Species | Days | T0 | T7 | T14 | T21 | T28 |
|---|---|---|---|---|---|---|
| Concentration (mmol−1/L) | ||||||
| CK | 0.081 ± 0.002a | 0.115 ± 0.005a | 0.090 ± 0.007c | 0.171 ± 0.027b | 0.164 ± 0.017a | |
| 100 | 0.082 ± 0.002a | 0.098 ± 0.007b | 0.113 ± 0.002b | 0.207 ± 0.009a | 0.119 ± 0.001b | |
| Chl a (mg/g) | 300 | 0.102 ± 0.013a | 0.096 ± 0.001b | 0.126 ± 0.006a | 0.129 ± 0.015c | 0.117 ± 0.003b |
| 500 | 0.158 ± 0.022a | 0.106 ± 0.010ab | 0.129 ± 0.004a | 0.158 ± 0.018c | 0.075 ± 0.005c | |
| 700 | 0.157 ± 0.005a | 0.102 ± 0.005b | 0.116 ± 0.004b | 0.184 ± 0.004ab | 0.049 ± 0.003d | |
| CK | 0.004 ± 0.001a | 0.009 ± 0.001a | 0.006 ± 0.001c | 0.007 ± 0.003d | 0.011 ± 0.001a | |
| 100 | 0.007 ± 0.003a | 0.008 ± 0.002a | 0.031 ± 0.002a | 0.014 ± 0.004c | 0.009 ± 0.001a | |
| Chl b (mg/g) | 300 | 0.006 ± 0.002a | 0.008 ± 0.003a | 0.004 ± 0.001c | 0.051 ± 0.006a | 0.010 ± 0.001a |
| 500 | 0.008 ± 0.003a | 0.010 ± 0.002a | 0.012 ± 0.002c | 0.014 ± 0.003c | 0.011 ± 0.001a | |
| 700 | 0.008 ± 0.003a | 0.008 ± 0.001a | 0.013 ± 0.006c | 0.022 ± 0.001b | 0.010 ± 0.001a | |
| CK | 0.085 ± 0.00a | 0.124 ± 0.004a | 0.096 ± 0.008c | 0.178 ± 0.030b | 0.175 ± 0.019a | |
| 100 | 0.0892 ± 0.00a | 0.107 ± 0.009b | 0.144 ± 0.004a | 0.222 ± 0.013a | 0.129 ± 0.001b | |
| Chl a + b (mg/g) | 300 | 0.108 ± 0.001a | 0.103 ± 0.004b | 0.129 ± 0.006b | 0.181 ± 0.021b | 0.127 ± 0.005b |
| 500 | 0.166 ± 0.02a | 0.116 ± 0.011ab | 0.141 ± 0.006a | 0.172 ± 0.020b | 0.086 ± 0.006c | |
| 700 | 0.165 ± 0.007a | 0.110 ± 0.004b | 0.129 ± 0.007b | 0.206 ± 0.004ab | 0.058 ± 0.004d | |
| CK | 22.180 ± 0.616a | 13.682 ± 0.931a | 15.294 ± 0.195b | 25.533 ± 0.368a | 14.932 ± 0.468a | |
| 100 | 13.655 ± 5.779a | 12.244 ± 0.976a | 3.654 ± 0.172c | 15.184 ± 0.183b | 13.009 ± 0.904b | |
| Chl a/b (mg/g) | 300 | 16.318 ± 2.242a | 13.552 ± 0.686a | 36.357 ± 0.750a | 2.511 ± 0.015c | 11.905 ± 0.015b |
| 500 | 23.080 ± 9.908a | 11.348 ± 0.895a | 10.649 ± 0.531b | 11.081 ± 0.733b | 7.017 ± 0.198c | |
| 700 | 23.726 ± 10.786a | 13.016 ± 0.374a | 10.221 ± 0.800b | 8.387 ± 0.449ab | 5.105 ± 0.420d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, Q.; Liu, Y.; Wei, H.; Jiang, L.; Wu, E.; Lin, L.; Yang, Q.; Yu, X.; Yan, L.; Li, Y. Salt Tolerance in Machilus faberi: Elucidating Growth and Physiological Adaptations to Saline Environments. Biology 2024, 13, 75. https://doi.org/10.3390/biology13020075
Mo Q, Liu Y, Wei H, Jiang L, Wu E, Lin L, Yang Q, Yu X, Yan L, Li Y. Salt Tolerance in Machilus faberi: Elucidating Growth and Physiological Adaptations to Saline Environments. Biology. 2024; 13(2):75. https://doi.org/10.3390/biology13020075
Chicago/Turabian StyleMo, Qiong, Yang Liu, Haohui Wei, Liyuan Jiang, En Wu, Ling Lin, Qihong Yang, Xiaoying Yu, Lihong Yan, and Yanlin Li. 2024. "Salt Tolerance in Machilus faberi: Elucidating Growth and Physiological Adaptations to Saline Environments" Biology 13, no. 2: 75. https://doi.org/10.3390/biology13020075