
In-Depth Genome Characterization and Pan-Genome Analysis of Strain KMM 296, a Producer of Highly Active Alkaline Phosphatase; Proposal for the Reclassification of Cobetia litoralis and Cobetia pacifica as the Later Heterotypic Synonyms of Cobetia amphilecti and Cobetia marina, and Emended Description of the Species Cobetia amphilecti and Cobetia marina
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Method
2.1. Strain Cultivation
2.2. Morphological, Biochemical, and Physiological Characterization
2.3. Whole-Cell Fatty Acid, Polar Lipid, and Respiratory Quinone Composition
2.4. The 16S rRNA Gene Sequencing and DNA–DNA Hybridization
2.5. Whole-Genome Shotgun Sequencing and Phylogenetic Analysis
3. Results and Discussion
3.1. Morphological, Biochemical, and Physiological Characterization
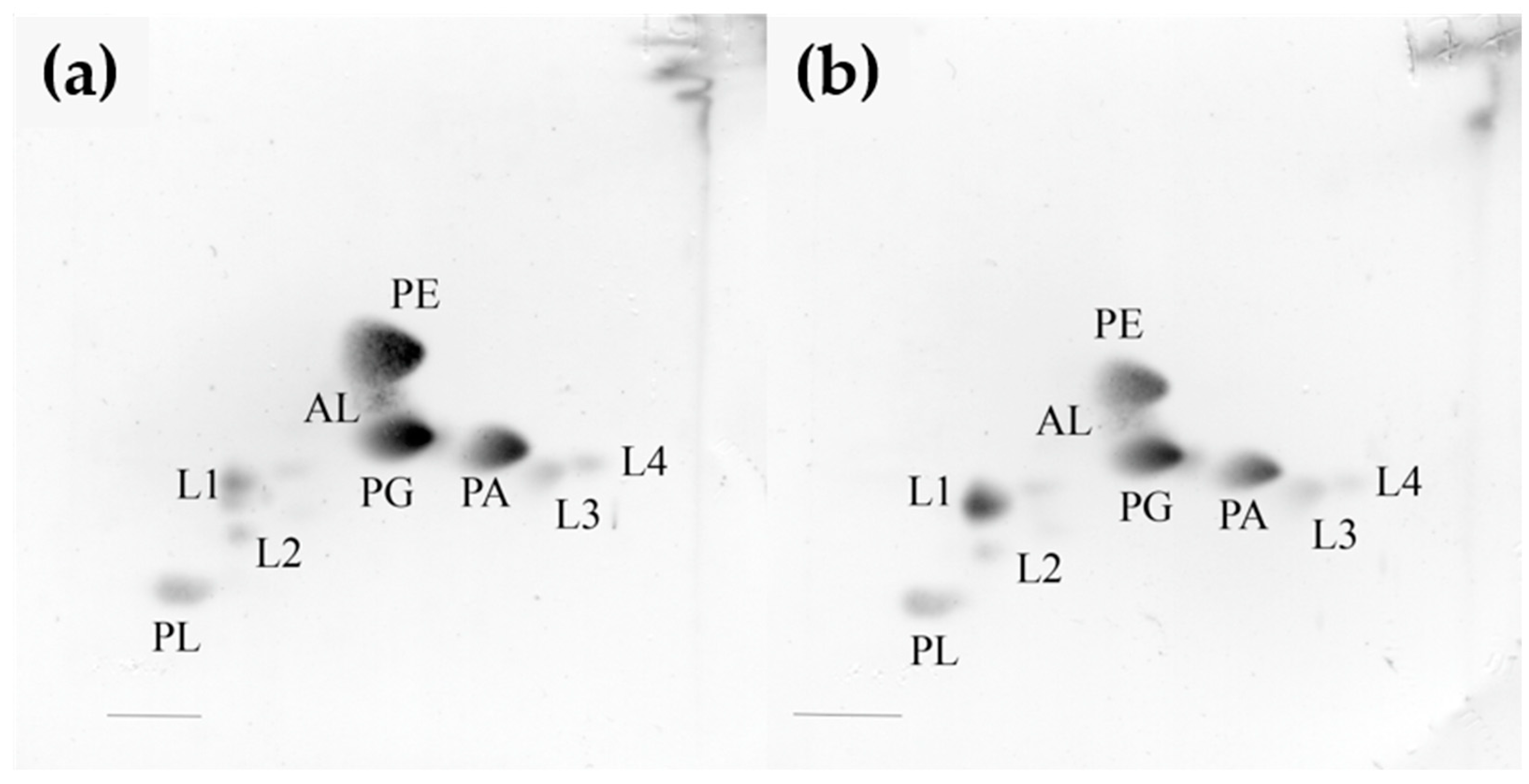
3.2. Whole-Cell Fatty Acid, Polar Lipid, and Respiratory Quinone Composition in the Strains KMM 296 and C. amphilecti NRIC 0815T
3.3. The 16S rRNA Gene Phylogenetic Analysis of the Strains KMM 296, C. marina LMG 2217T, and C. amphilecti NRIC 0815T
3.4. GC Comparison between Strains KMM 296 and C. amphilecti NRIC 0815T
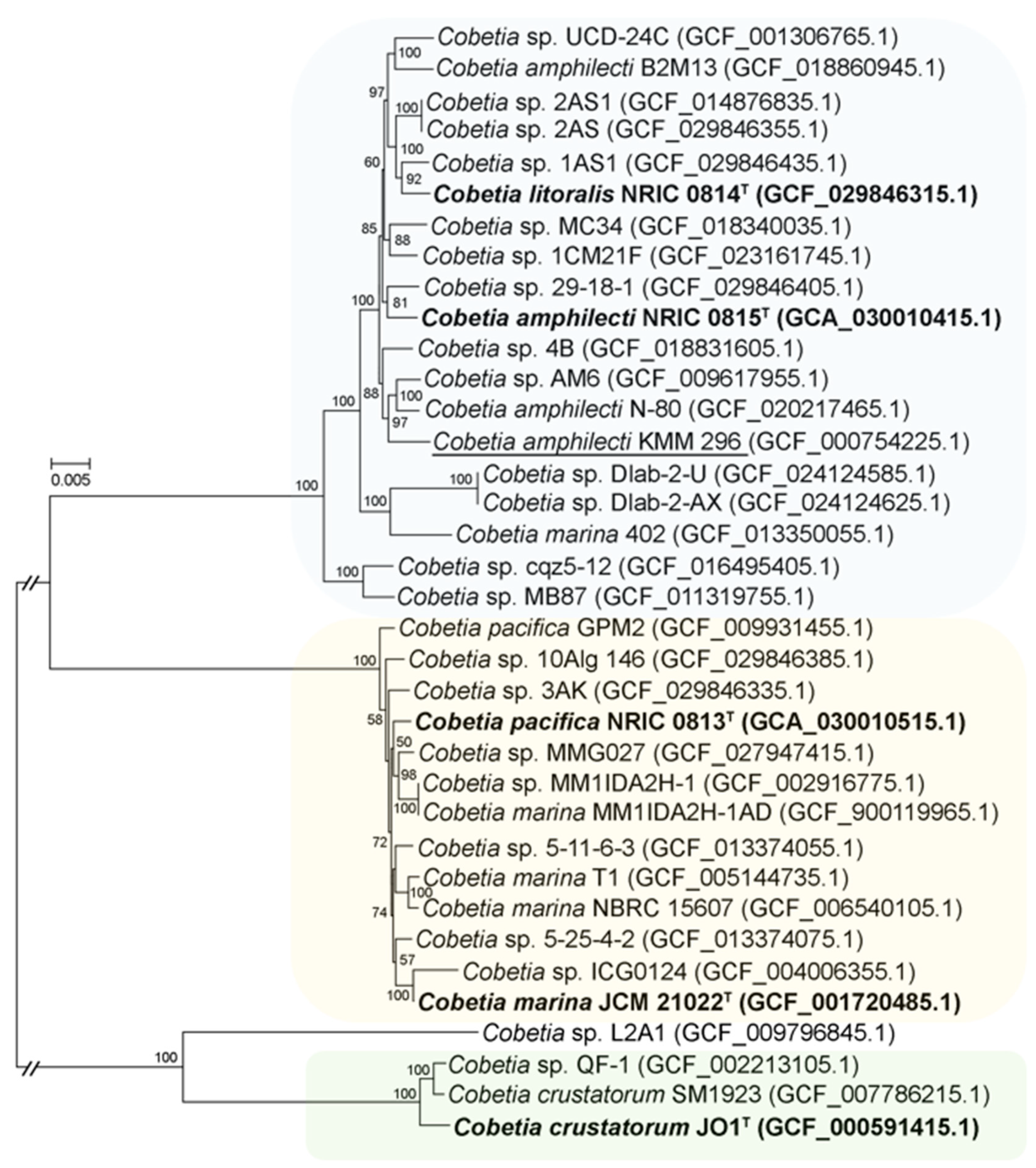
3.5. Whole-Genome-Based Phylogeny and Analysis of Cobetia Strains
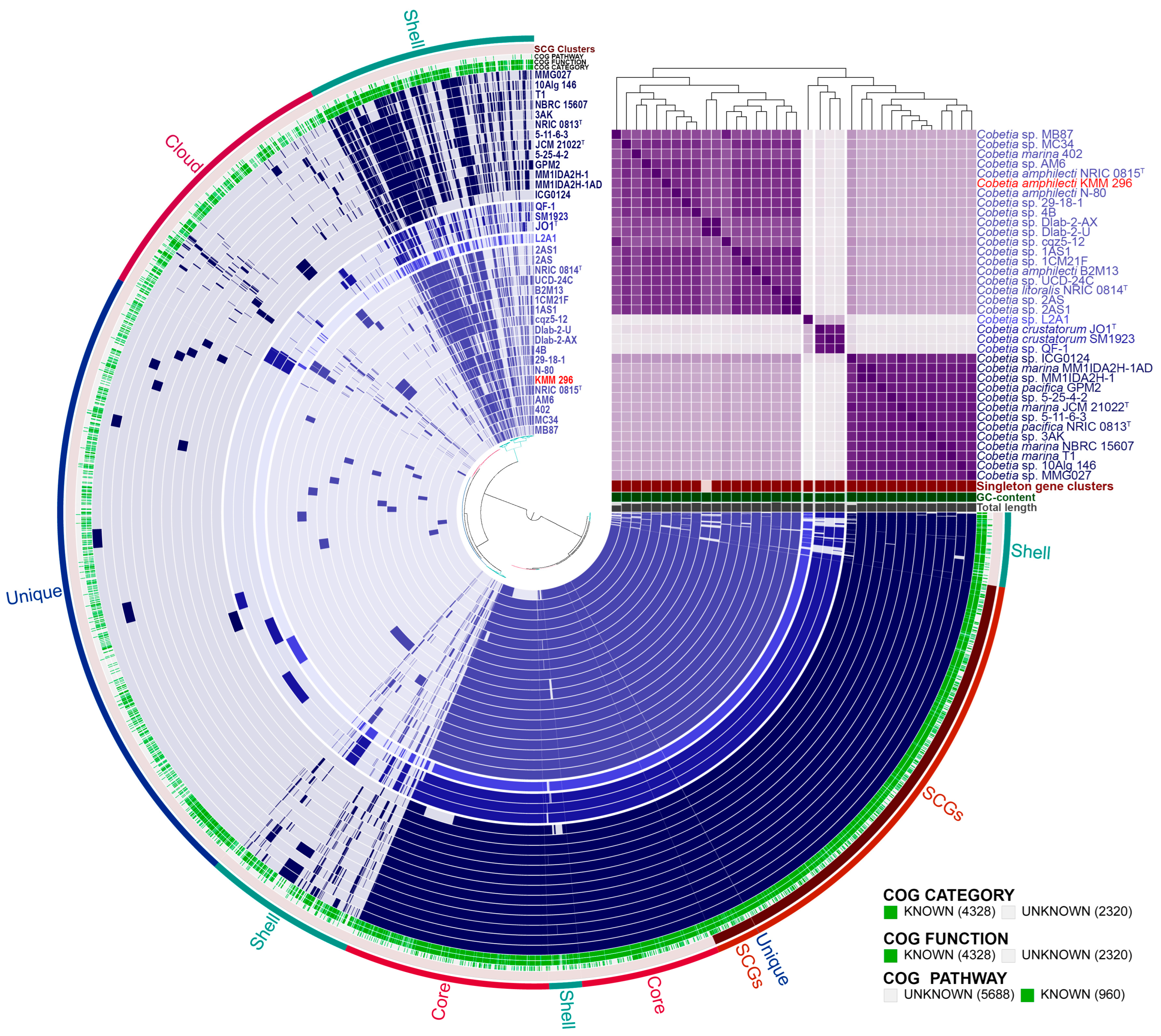
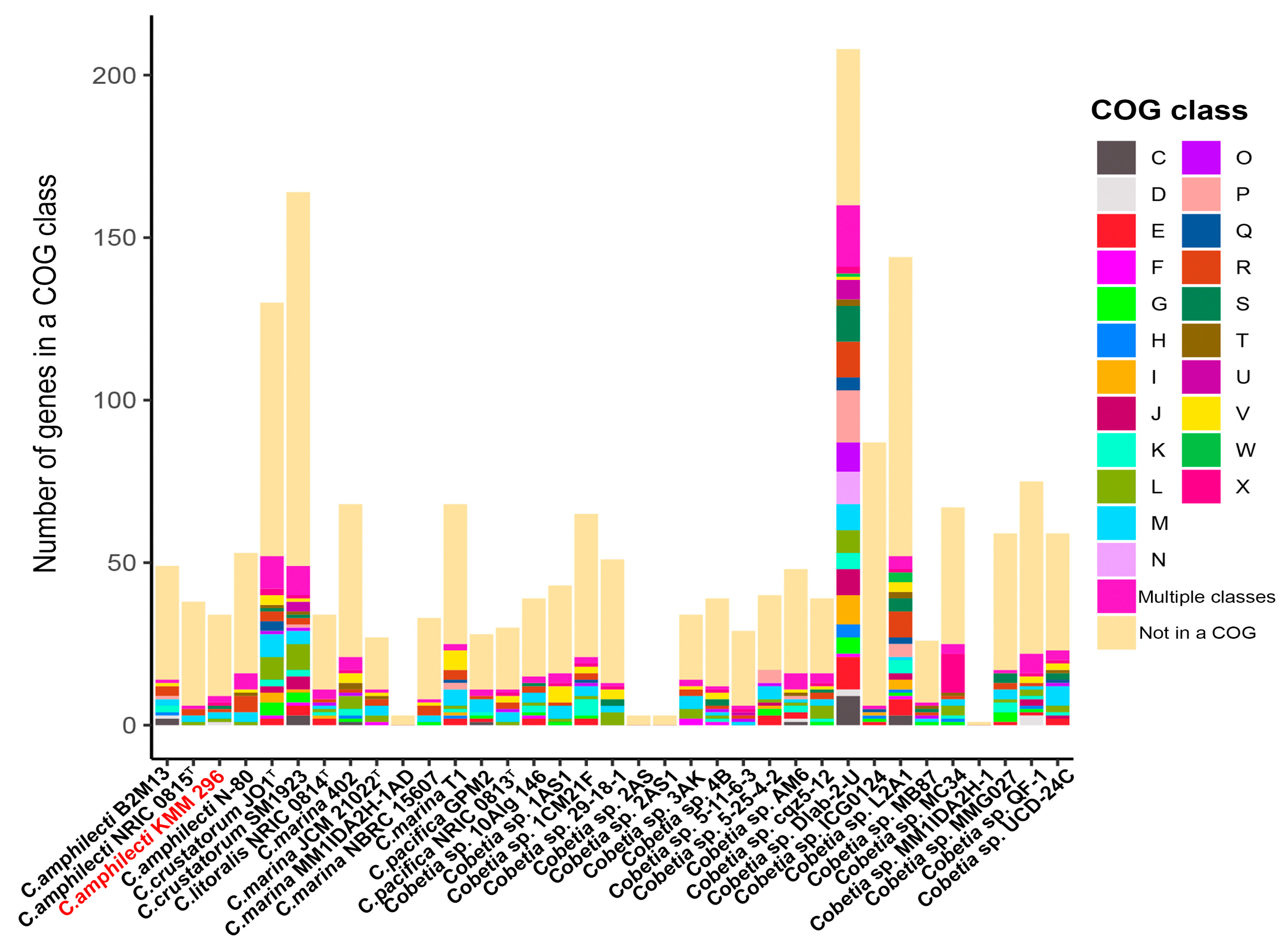
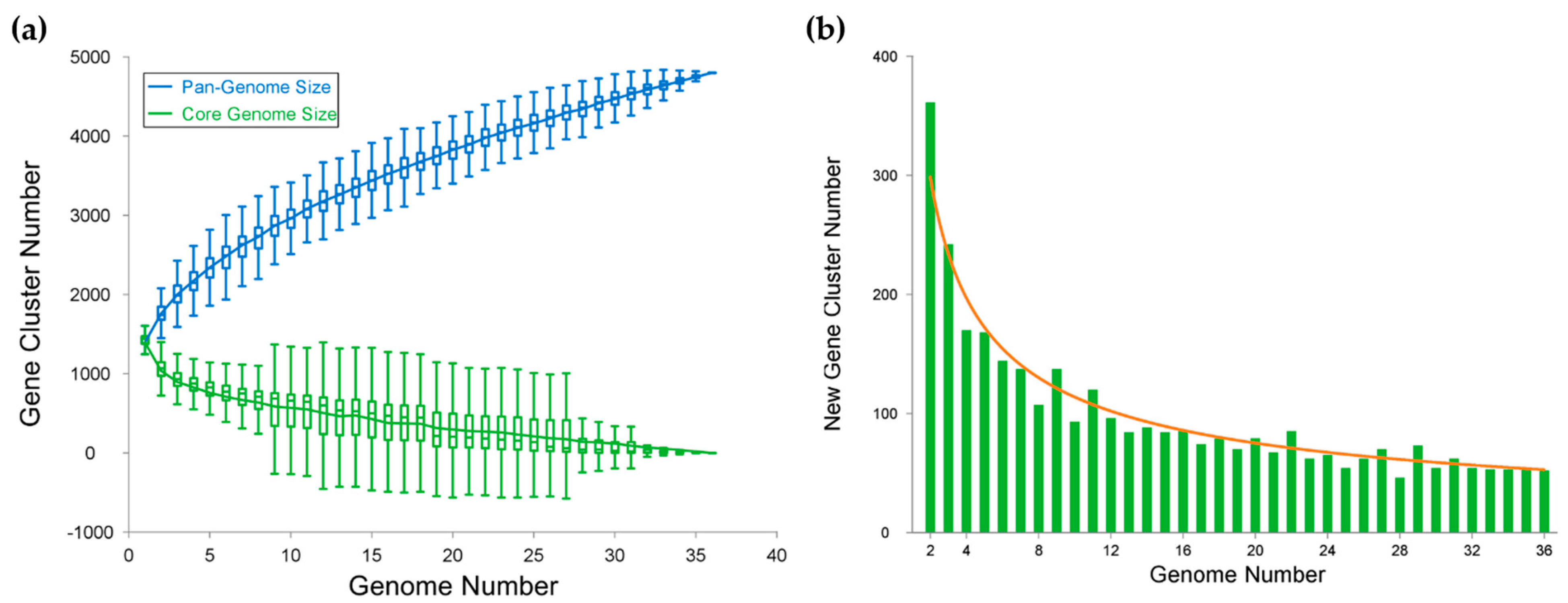
3.6. Pan-Genome-Based Phylogeny and Analysis of Cobetia Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arahal, D.R.; Castillo, A.M.; Ludwig, W.; Schleifer, K.H.; Ventosa, A. Proposal of Cobetia marina gen. nov., comb. nov., within the family Halomonadaceae, to include the species Halomonas marina. Syst. Appl. Microbiol. 2002, 25, 207–211. [Google Scholar] [CrossRef]
- Kim, M.S.; Roh, S.W.; Bae, J.W. Cobetia crustatorum sp. nov., a novel slightly halophilic bacterium isolated from traditional fermented seafood in Korea. Int. J. Syst. Evol. Microbiol. 2010, 60, 620–626. [Google Scholar] [CrossRef]
- Romanenko, L.A.; Tanaka, N.; Svetashev, V.I.; Falsen, E. Description of Cobetia amphilecti sp. nov., Cobetia litoralis sp. nov. and Cobetia pacifica sp. nov., classification of Halomonas halodurans as a later heterotrophic synonym of Cobetia marina and emended descriptions of the genus Cobetia and Cobetia marina. Int. J. Syst. Evol. Microbiol. 2013, 63, 288–297. [Google Scholar] [CrossRef]
- Ibacache-Quiroga, C.; Canales, C.; Charifeh, M.; Dinamarca, M.A. Genome sequence of Cobetia sp. strain MM1IDA2H-1, a hydrocarbon-degrading and biosurfactant-producing marine bacterium. Genome Announc. 2017, 5, e00132-17. [Google Scholar] [CrossRef]
- Liu, W.; Cong, B.; Lin, J.; Zhao, L.; Liu, S. Complete genome sequencing and comparison of two nitrogen-metabolizing bacteria isolated from Antarctic deep-sea sediment. BMC Genom. 2022, 23, 713. [Google Scholar] [CrossRef]
- Cheng, W.; Yan, X.; Xiao, J.; Chen, Y.; Chen, M.; Jin, J.; Bai, Y.; Wang, Q.; Liao, Z.; Chen, Q. Isolation, identification, and whole genome sequence analysis of the alginate-degrading bacterium Cobetia sp. cqz5-12. Sci. Rep. 2020, 10, 10920. [Google Scholar] [CrossRef]
- Christensen, M.; Jablonski, P.; Altermark, B.; Irgum, K.; Hansen, H. High natural PHA production from acetate in Cobetia sp. MC34 and Cobetia marina DSM 4741T and in silico analyses of the genus specific PhaC2 polymerase variant. Microb. Cell Fact. 2021, 20, 225. [Google Scholar] [CrossRef] [PubMed]
- Moriya, H.; Takita, Y.; Matsumoto, A.; Yamahata, Y.; Nishimukai, M.; Miyazaki, M.; Shimoi, H.; Kawai, S.J.; Yamada, M. Cobetia sp. bacteria, which are capable of utilizing alginate or waste Laminaria sp. for poly(3-hydroxybutyrate) synthesis, isolated from a marine environment. Front. Bioeng. Biotechnol. 2020, 8, 974. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Mikhailov, V.V.; Plisova, E.J.; Balabanova, L.A.; Svetashev, V.V.; Vysockyi, M.V.; Stepanenko, V.I.; Rasskazov, V.A. Characterization of the marine bacterium Deleya marina producing highly active alkaline phosphatase and associated with the mussel Crenomytilus grayanus. Russ. J. Mar. Biol. 1994, 20, 340–545. [Google Scholar]
- Ivanova, E.P.; Christen, R.; Sawabe, T.; Alexeeva, Y.V.; Lysenko, A.M.; Chelomin, V.P.; Mikhailov, V.V. Presence of ecophysiologically diverse populations within Cobetia marina strains isolated from marine invertebrate, algae and the environments. Microbes Environ. 2005, 20, 200–207. [Google Scholar] [CrossRef]
- Balabanova, L.A.; Golotin, V.A.; Kovalchuk, S.N.; Babii, A.V.; Shevchenko, L.S.; Son, O.M.; Kosovsky, G.Y.; Rasskazov, V.A. The Genome of the marine bacterium Cobetia marina KMM 296 isolated from the mussel Crenomytilus grayanus (Dunker, 1853). Russ. J. Mar. Biol. 2016, 42, 106–109. [Google Scholar] [CrossRef]
- Plisova, E.Y.; Balabanova, L.A.; Ivanova, E.P.; Kozhemyako, V.B.; Mikhailov, V.V.; Agafonova, E.V.; Rasskazov, V.A. A highly active alkaline phosphatase from the marine bacterium Cobetia. Mar. Biotechnol. 2005, 7, 173–178. [Google Scholar] [CrossRef]
- Golotin, V.; Balabanova, L.; Likhatskaya, G.; Rasskazov, V. Recombinant production and characterization of a highly active alkaline phosphatase from marine bacterium Cobetia marina. Mar. Biotechnol. 2015, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Balabanova, L.; Podvolotskaya, A.; Slepchenko, L.; Eliseikina, M.; Noskova, Y.; Nedashkovskaya, O.; Son, O.; Tekutyeva, L.; Rasskazov, V. Nucleolytic enzymes from the marine bacterium Cobetia amphilecti KMM 296 with antibiofilm activity and biopreservative effect on meat products. Food Control 2017, 78, 270–278. [Google Scholar] [CrossRef]
- Buinovskaya, N.S.; Bakholdina, S.I.; Balabanova, L.A. Dephosphorylation of lipopolysaccharides by alkaline phosphatase from marine bacterium. Vestn. FEB RAS 2018, 6, 80–81. [Google Scholar]
- Yang, Y.; Wandler, A.; Postlethwait, J.; Guillemin, K. Dynamic evolution of the LPS-detoxifying enzyme intestinal alkaline phosphatase in Zebrafish and other vertebrates. Front. Immunol. 2012, 3, 314. [Google Scholar] [CrossRef]
- Lalles, J.P. Intestinal alkaline phosphatase: Multiple biological roles in maintenance of intestinal homeostasis and modulation by diet. Nutr. Rev. 2010, 68, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Hamarneh, S.R.; Mohamed, M.M.; Economopoulos, K.P.; Morrison, S.A.; Phupitakphol, T.; Tantillo, T.J.; Gul, S.S.; Gharedaghi, M.H.; Tao, Q.; Kaliannan, K.; et al. A novel approach to maintain gut mucosal integrity using an oral enzyme supplement. Ann. Surg. 2014, 260, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Kühn, F.; Adiliaghdam, F.; Cavallaro, P.M.; Hamarneh, S.R.; Tsurumi, A.; Hoda, R.S.; Munoz, A.R.; Dhole, Y.; Ramirez, J.M.; Liu, E.; et al. Intestinal alkaline phosphatase targets the gut barrier to prevent aging. JCI Insight 2020, 26, e134049. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, P.; Murray, R.G.E.; Wood, W.A.; Krieg, N.R. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994; p. 224. [Google Scholar]
- Lemos, M.L.; Toranzo, A.E.; Barja, J.L. Modified medium for oxidation-fermentation test in the identification of marine bacteria. Appl. Environ. Microbiol. 1985, 40, 1541–1543. [Google Scholar] [CrossRef]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids; Tech. Note 101; MIDI, Inc.: Newark, DE, USA, 1990. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Shah, H.M. Fatty acid, menaquinone and polar lipid composition of Rothia dentosacariosa. Arch. Microbiol. 1984, 137, 247–249. [Google Scholar] [CrossRef]
- Komagata, K.; Suzuki, K.I. Lipid and cell wall analysis in bacterial systematics. Methods Microbiol. 1987, 19, 161–207. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Volume 1. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Noskova, Y.; Seitkalieva, A.; Nedashkovskaya, O.; Shevchenko, L.; Tekutyeva, L.; Son, O.; Balabanova, L. Are the closely related Cobetia strains of different species? Molecules 2021, 26, 690. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- De Ley, J.; Cattoir, H.; Reynaerts, A. The quantitative measurement of DNA hybridization from renaturation rates. Eur. J. Biochem. 1970, 12, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Eren, A.M.; Esen, O.C.; Quince, C.; Vineis, J.H.; Morrison, H.G.; Sogin, M.L.; Delmont, T.O. Anvi’o: An advanced analysis and visualization platformfor ‘omics data. PeerJ 2015, 3, e1319. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Prepr. 2016, 4, e1900v1. [Google Scholar]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Riley, D.; Cattuto, C.; Medini, D. Comparative genomics: The bacterial pan-genome. Curr. Opin. Microbiol. 2008, 11, 472–477. [Google Scholar] [CrossRef]
- Tang, X.; Xu, K.; Han, X.; Mo, Z.; Mao, Y. Complete genome of Cobetia marina JCM 21022T and phylogenomic analysis of the family Halomonadaceae. J. Ocean. Limnol. 2018, 36, 528–536. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Accession ID | Genome Size, bp | G+C (mol%) | Contigs | Completeness (%) | Contamination (%) | Isolation Source |
|---|---|---|---|---|---|---|---|
| Cobetia sp. UCD-24C | GCF_001306765.1 | 4,229,986 | 62.5 | 51 | 98.9 | 1.76 | Seagrass sediment |
| C. amphilecti B2M13 | GCF_018860945.1 | 4,289,324 | 62.5 | 58 | 95.88 | 0.91 | Artificial alginate particle |
| Cobetia sp. 2AS1 | GCF_014876835.1 | 4,248,424 | 62.5 | 49 | 95.96 | 0.9 | Coastal sediment |
| Cobetia sp. 2AS | GCF_029846355.1 | 4,247,060 | 62.5 | 39 | 99.15 | 1.3 | |
| Cobetia sp. 1AS1 | GCF_029846435.1 | 4,235,090 | 62.5 | 43 | 99.27 | 1.71 | Coastal seawater |
| C. litoralis NRIC 0814T | GCF_029846315.1 | 4,621,254 | 62.5 | 51 | 99.15 | 2.93 | Sandy sediment |
| Cobetia sp. MC34 | GCF_018340035.1 | 4,022,416 | 62.5 | 175 | 95.47 | 0.87 | Fish-landing facility |
| Cobetia sp. 1CM21F | GCF_023161745.1 | 4,261,659 | 62.5 | 24 | 95.88 | 0.49 | Sea cave |
| Cobetia sp. 29-18-1 | GCF_029846405.1 | 4,117,019 | 62.5 | 70 | 99.35 | 1.48 | Esperiopsis digitata |
| C. amphilecti NRIC 0815T | GCA_030010415.1 | 4,171,304 | 62.5 | 112 | 98.54 | 1.34 | Internal tissue, Amphilectus digitatus |
| Cobetia sp. 4B | GCF_018831605.1 | 4,325,922 | 62.5 | 3 | 99.41 | 1.33 | Current Humbolt system, Heterostera chilensis |
| Cobetia sp. AM6 | GCF_009617955.1 | 4,229,996 | 62.5 | 1 | 98.74 | 1.36 | Japan: Tokyo |
| C. amphilecti N-80 | GCF_020217465.1 | 4,160,095 | 62.5 | 1 | 98.74 | 1.28 | Marine sediment |
| C. amphilecti KMM 296 | GCF_000754225.1 | 3,965,007 | 62.5 | 97 | 98.29 | 1.28 | Crenomytilus grayanus |
| Cobetia sp. Dlab-2-U | GCF_024124585.1 | 4,144,083 | 62.5 | 137 | 96.63 | 1.3 | Coral surface mucus layer and tissue Diploria labyrinthiformis |
| Cobetia sp. Dlab-2-AX | GCF_024124625.1 | 4,001,795 | 62.5 | 20 | 96.29 | 1.3 | |
| C. marina 402 | GCF_013350055.1 | 3,978,956 | 62.5 | 132 | 95.93 | 0.6 | Seawater, aquarium |
| Cobetia sp. cqz5-12 | GCF_016495405.1 | 4,209,007 | 62.5 | 1 | 99.45 | 1.71 | Brown algae Sargassum fusiforme |
| Cobetia sp. MB87 | GCF_011319755.1 | 3,101,384 | 62.5 | 12 | 81.26 | 1.58 | Sea cucumber gut |
| C. pacifica GPM2 | GCF_009931455.1 | 4,195,186 | 62.5 | 1 | 99.59 | 1.33 | Neopyropia tenera |
| Cobetia sp. 10Alg 146 | GCF_029846385.1 | 4,095,141 | 62.5 | 33 | 99.44 | 1.68 | Ahnfeltia tobuchiensis |
| Cobetia sp. 3AK | GCF_029846335.1 | 4,073,243 | 62.5 | 58 | 99.44 | 1.71 | Coastal seawater |
| C. pacifica NRIC 0813T | GCA_030010515.1 | 4,066,371 | 62.5 | 42 | 99.44 | 1.3 | Sandy sediment |
| Cobetia sp. MMG027 | GCF_027947415.1 | 4,168,882 | 62.5 | 47 | 99.24 | 1.71 | - |
| Cobetia sp. MM1IDA2H-1 | GCF_002916775.1 | 4,151,052 | 62 | 88 | 99.65 | 1.33 | Eulittoral intertidal pond at sea level |
| C. marina MM1IDA2H-1AD | GCF_900119965.1 | 4,155,178 | 62 | 105 | 99.65 | 1.33 | - |
| Cobetia sp. 5-11-6-3 | GCF_013374055.1 | 4,120,053 | 62.5 | 40 | 99.44 | 1.3 | Seaweed |
| C. marina T1 | GCF_005144735.1 | 4,177,239 | 62 | 21 | 99.44 | 1.28 | Saltwater |
| C. marina NBRC 15607 | GCF_006540105.1 | 4,184,377 | 62.5 | 113 | 99.24 | 1.68 | - |
| Cobetia sp. 5-25-4-2 | GCF_013374075.1 | 4,118,344 | 62.5 | 41 | 99.44 | 1.3 | Seaweed |
| Cobetia sp. ICG0124 | GCF_004006355.1 | 3,345,506 | 63 | 1 | 90.85 | 2.1 | Marine waters |
| C. marina JCM 21022T | GCF_001720485.1 | 4,176,400 | 62.5 | 1 | 99.51 | 1.51 | Littoral water |
| Cobetia sp. L2A1 | GCF_009796845.1 | 4,118,938 | 57.5 | 1 | 98.9 | 1.28 | Arctic Ocean beach |
| Cobetia sp. QF-1 | GCF_002213105.1 | 4,084,184 | 57.5 | 31 | 99.04 | 0.93 | Seawater |
| C. crustatorum SM1923 | GCF_007786215.1 | 4,215,468 | 57.5 | 163 | 98.83 | 0.82 | Surface seawater |
| C. crustatorum JO1T | GCF_000591415.1 | 4,049,952 | 57.5 | 138 | 90.05 | 0.52 | Fermented shrimp |
| Characteristic | KMM 296 | C. amphilecti NRIC 0815T |
|---|---|---|
| Source and site of isolation | Mollusk C. grayanus, the Sea of Japan, Pacific Ocean | Sponge A. digitatus, the Gulf of Alaska, Pacific Ocean |
| Temperature range for growth (°C) | 4–42 | 4–42 |
| Salinity range for growth (% NaCl) | 0.5–19 | 0–20 |
| Nitrate reduction | - | + |
| Hydrolysis of: | ||
| Tween 80 | - | + |
| DNA | - | + |
| Acid production from: | ||
| D-Fructose, D-lactose | - | + |
| L-Arabinose, D-melibiose, L-rhamnose | + | - |
| Assimilation of: | ||
| Amygdalin, maltose, sodium malonate, glycogen, capric acid, valeric acid, 3-hydroxybutiric acid, L-proline | + | - |
| N-acetylglucosamine, L-serine | - | + |
| Enzyme activities: | ||
| Valine arylamidase, cysteine arylamidase | - | + |
| Trypsin | + | - |
| DNA G+C content (mol%) | 62.5 | 62.5 |
| Fatty Acid * | KMM 296 | C. amphilecti NRIC 0815T |
|---|---|---|
| C10:0 | 3.2 | 4.3 |
| C12:0 | 11.0 | 8.7 |
| C16:0 | 21.6 | 21.9 |
| C17:0 cyclo | 5.2 | 9.4 |
| C16:1 ω7c | 38.2 | 32.5 |
| C18:1 ω7c | 14.9 | 12.7 |
| C19:1 ω6c | 1.6 | tr |
| C12:0 3-OH | 15.8 | 20.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedashkovskaya, O.; Balabanova, L.; Otstavnykh, N.; Zhukova, N.; Detkova, E.; Seitkalieva, A.; Bystritskaya, E.; Noskova, Y.; Tekutyeva, L.; Isaeva, M. In-Depth Genome Characterization and Pan-Genome Analysis of Strain KMM 296, a Producer of Highly Active Alkaline Phosphatase; Proposal for the Reclassification of Cobetia litoralis and Cobetia pacifica as the Later Heterotypic Synonyms of Cobetia amphilecti and Cobetia marina, and Emended Description of the Species Cobetia amphilecti and Cobetia marina. Biomolecules 2024, 14, 196. https://doi.org/10.3390/biom14020196
Nedashkovskaya O, Balabanova L, Otstavnykh N, Zhukova N, Detkova E, Seitkalieva A, Bystritskaya E, Noskova Y, Tekutyeva L, Isaeva M. In-Depth Genome Characterization and Pan-Genome Analysis of Strain KMM 296, a Producer of Highly Active Alkaline Phosphatase; Proposal for the Reclassification of Cobetia litoralis and Cobetia pacifica as the Later Heterotypic Synonyms of Cobetia amphilecti and Cobetia marina, and Emended Description of the Species Cobetia amphilecti and Cobetia marina. Biomolecules. 2024; 14(2):196. https://doi.org/10.3390/biom14020196
Chicago/Turabian StyleNedashkovskaya, Olga, Larissa Balabanova, Nadezhda Otstavnykh, Natalia Zhukova, Ekaterina Detkova, Aleksandra Seitkalieva, Evgenia Bystritskaya, Yulia Noskova, Liudmila Tekutyeva, and Marina Isaeva. 2024. "In-Depth Genome Characterization and Pan-Genome Analysis of Strain KMM 296, a Producer of Highly Active Alkaline Phosphatase; Proposal for the Reclassification of Cobetia litoralis and Cobetia pacifica as the Later Heterotypic Synonyms of Cobetia amphilecti and Cobetia marina, and Emended Description of the Species Cobetia amphilecti and Cobetia marina" Biomolecules 14, no. 2: 196. https://doi.org/10.3390/biom14020196