
From Host Defense to Metabolic Signatures: Unveiling the Role of γδ T Cells in Bacterial Infections

Department of Dermatology, Johns Hopkins University School of Medicine, Baltimore, MD 21231, USA
*
Author to whom correspondence should be addressed.
Biomolecules 2024, 14(2), 225; https://doi.org/10.3390/biom14020225
Submission received: 30 December 2023
/
Revised: 12 February 2024
/
Accepted: 12 February 2024
/
Published: 15 February 2024
(This article belongs to the Special Issue Novel Insights into Gamma Delta T Cells (γδ T Cells): Receptors, Molecular Targets, and Therapeutic Potentials)
{kind=link}
{kind=link}
Abstract
:The growth of antibiotic-resistant bacterial infections necessitates focusing on host-derived immunotherapies. γδ T cells are an unconventional T cell subset, making up a relatively small portion of healthy circulating lymphocytes but a substantially increased proportion in mucosal and epithelial tissues. γδ T cells are activated and expanded in response to bacterial infection, having the capability to produce proinflammatory cytokines to recruit neutrophils and clear infection. They also play a significant role in dampening immune response to control inflammation and protecting the host against secondary challenge, making them promising targets when developing immunotherapy. Importantly, γδ T cells have differential metabolic states influencing their cytokine profile and subsequent inflammatory capacity. Though these differential metabolic states have not been well studied or reviewed in the context of bacterial infection, they are critical in understanding the mechanistic underpinnings of the host’s innate immune response. Therefore, this review will focus on the context-specific host defense conferred by γδ T cells during infection with Staphylococcus aureus, Streptococcus pneumoniae, Listeria monocytogenes, and Mycobacterium tuberculosis.
1. Introduction
γδ T cells are an unconventional T cell subset, making up about 1–5% of circulating lymphocytes in most healthy animal species [1]. However, in healthy mucosal or epithelial tissue, these cells can comprise up to 50% of the T cell population [2]. Furthermore, γδ T cell population increases dramatically in response to invading pathogens [3]. Along with Mucosal associated invariant T cells (MAIT), Natural Killer T cells (NKT), and Intraepithelial lymphocytes (IEL), and other unconventional T cell subsets, γδ T cells are non-MHC restricted, can recognize a host of diverse self- and foreign molecules, and have both innate and adaptive immune cell features [4].
γδ T cells are classified by their TCRγ and TCRδ chain usage. In humans, this is primarily divided into Vδ1+ and Vδ2+ subsets, which arise during fetal development. Vδ1+ subsets typically associate with the Vγ1+ group, including Vγ2,3,4,5, and 8 [5], and are found in the mucosal epithelium [6,7]. Vδ2+ subsets typically only associate with the Vγ2+ group, including Vγ9 [5], and form the largest γδ T cell population in the peripheral blood [6,7]. The functions of human Vγ9Vδ2+ T cells are well studied, including cytokine production, killing of infected target cells, regulation of monocyte and Dendritic cell (DC) differentiation and maturation, and antigen presentation [8,9,10,11].
In mice, however, γδ T cells are classified primarily via their Vγ chain expression and can be resident to specific tissues with non-redundant functions [12]. The first γδ T cell population to arise in the thymus are Vγ5Vδ1 T cells, which migrate to the epidermis and become dendritic epidermal T cells (DETCs), a population not found in humans [13]. DETC progenitors are found between embryonic days 14–16 [14]. In the dermis, however, γδ T cells are primarily Vγ6+, arising at around embryonic day 5, but found obviously on day 3 in mice [12]. These cells are bona fide resident dermal cells and are essential in neonatal skin immunity [12]. Vγ4+ cells increase in number through development [12]. Vγ4+ γδ T cells typically make IFNγ cytokine, while Vγ6+ γδ T cells typically make IL-17 and IL-22 [15]. Thymic signals regulate these cells’ subsequent effector function and critical role during early infection stages [16].
γδ T cells may also have memory functions, although it is unclear if these functions are entirely analogous to αβ T cell memory functions [17,18,19,20]. Though not fully elucidated, human Vγ9Vδ2 T cells respond to (E)-4-Hydroxy-3-methyl-but-2-enyl pyrophosphate (HMB-PP), an intermediate in the non-mevalonate (MEP) pathway of isoprenoid synthesis in some pathogenic bacterial species [8,9,21,22]; isopentenyl phosphate (IPP) an intermediate in the mevalonate pathway of isoprenoid synthesis [21]; and superantigens [23]. Other subsets of γδ T cells respond to antigens such as CMV virus and MHC-related molecules [24].
γδ T cells are thus important in understanding the host response to bacterial infection. Part of the host immune response includes metabolic reprogramming, coined as immunometabolism, driving subsequent cytokine, chemokine, and interferon responses [25]. This review aims to elucidate the immunometabolic role of γδ T cells during Staphylococcus aureus, Streptococcus pneumoniae, Listeria monocytogenes, and M. tuberculosis infections, focusing on their contextual roles in pro- and anti-inflammatory signaling.
2. Immunometabolism of γδ T Cells during Bacterial Infection
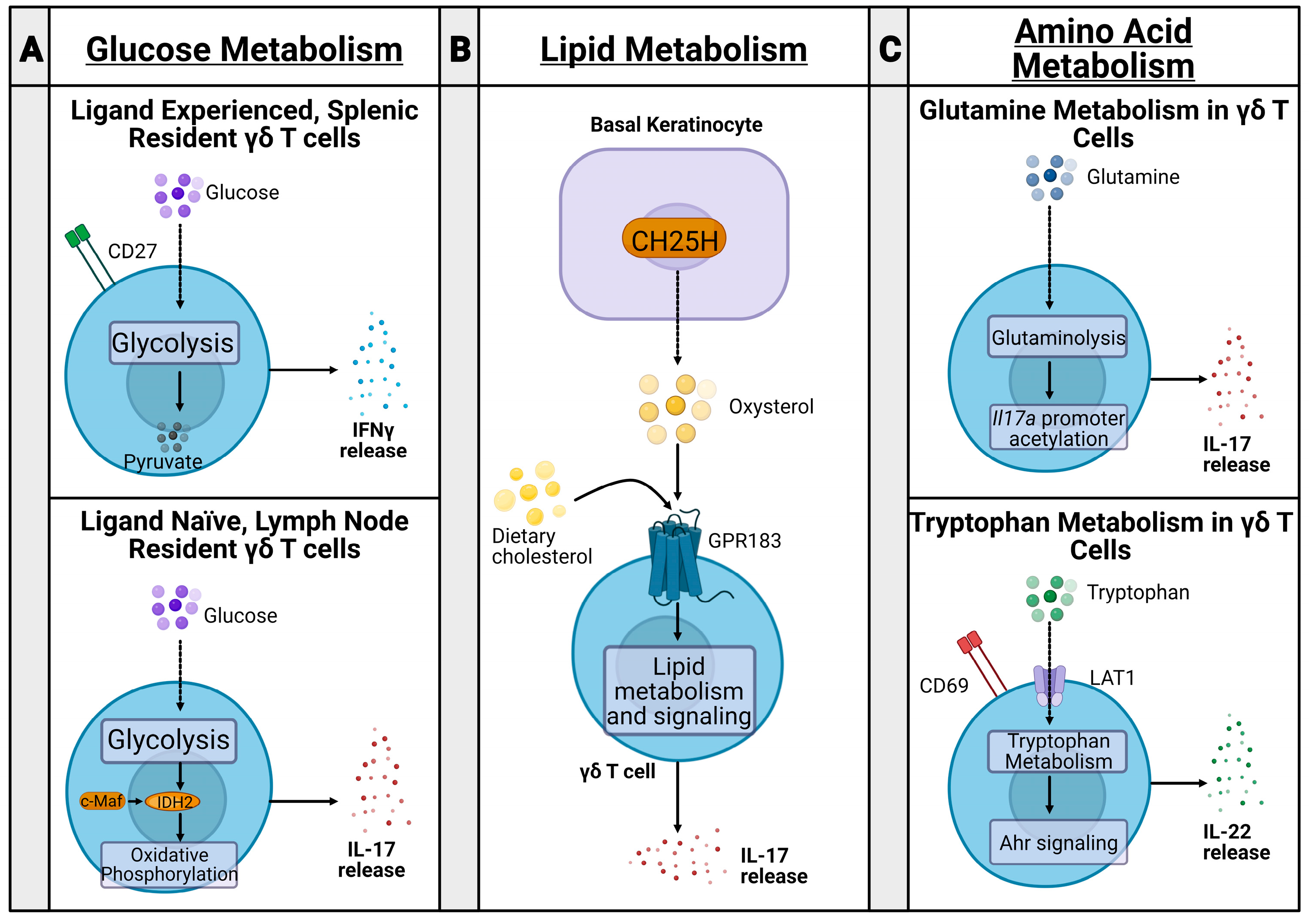
Murine γδ T cells can be functionally divided into two subsets via the expression of CD27 and their subsequent metabolic states [26,27] (Figure 1A). CD27+ γδ T cells are generally splenic-resident, ligand-experienced, and secrete IFNγ [26,27]. Alternatively, CD27− γδ T cells are found in the lymph nodes (LNs), are ligand naive, and secrete IL-17 [26,27]. The transcription factor c-Maf is an essential regulator of IL-17 producing γδ T cells by increasing chromatin availability of genes involved in producing Th17 cytokines, Rorc, Il17a, and Blk, and downregulating TCF1, which limits IL-17 producing γδ T cell differentiation [28]. These two cell sets have differential metabolic states established during thymic development [29]. IFNγ producing cells are dependent on glycolysis, while IL-17-producing cells rely on oxidative phosphorylation [29], controlled by c-Maf regulation of rate-limiting enzyme isocitrate dehydrogenase 2 (IDH2) [30].
While glucose metabolic pathways may mark differential γδ T cell subsets by cytokine production, lipid metabolic changes may impact the severity of inflammation in inflammatory disease (Figure 1B). In a psoriasis model, dietary cholesterol metabolites, such as oxysterol, regulate IL-17+ γδ T cell development and trafficking to the skin, with a higher fat diet in mice being associated with more severe psoriasis [31]. Similarly, IL-17+ γδ T cells had increased lipid metabolism and storage in murine breast adenocarcinoma, colon adenocarcinoma, and melanoma models [29].
Amino acid metabolism in T cells has been well studied, particularly in the context of cancer; however, recent studies aim to relate amino acid metabolism in γδ T cells to changes in inflammation severity in psoriasis and polymicrobial sepsis (Figure 1C). For example, glutamine metabolism was upregulated in IL-17+ γδ T cells during psoriasis, promoting acetylation of the Il17a promoter and increasing IL-17 production [32,33]. Additionally, CD69 interacted with the heterodimeric LAT1-CD98 amino acid transporter to upregulate tryptophan uptake during psoriasis, leading to mTORC- and AhR-mediated IL-22 production and increased inflammation [33]. Furthermore, glutamine metabolism is also implicated in mouse models of polymicrobial sepsis. Mice treated with glutamine administration had increased γδ T cell populations, decreased inflammatory lesions, and reduced lung damage compared with mice treated with saline [34]. While these metabolic states were established and studied in both an inflammatory and tumor microenvironment, these metabolic changes have not yet been fully elucidated during bacterial infection. To this end, our lab is currently studying immunometabolic states of immune cells, including conventional and unconventional T cell types, including γδ T cell and MAIT cell populations, during S. aureus infection.
3. Response of Human Vγ9Vδ2 T Cells to Phosphoantigens
Human Vγ9Vδ2 T cells have remarkably rapid responses to phosphoantigens. IPP, an intermediate in the mevalonate pathway of isoprenoid synthesis, is a stimulator of human Vγ9Vδ2 T cells, though less potent than HMB-PP [24]. Zoledronate (ZOL) and pamidronate (PAM) are aminobisphosphonate drugs and stimulators of human Vγ9Vδ2 T cells [35]. They work by inhibiting farnesyl diphosphate synthase, an enzyme that metabolizes IPP, which leads to an accumulation of IPP, and subsequent activation of Vγ9Vδ2 T cells [35]. The microbial side of γδ T cell immunometabolism largely centers around HMB-PP, one of the most potent stimulators of human Vγ9Vδ2 T cells. HMB-PP is a microbial prenyl phosphate metabolite recognized as a pathogen-associated molecular pattern (PAMP) [35]. To be stimulated by HMB-PP, γδ T cells require butyrophilin membrane proteins [36]. Phosphoantigens like HMB-PP interact with and bind to the B30.2 and juxtamembrane domains within the intracellular domain of BTN3A to modulate phopshoantigen sensing [35].
Furthermore, in Vγ9Vδ2 T cells, HMB-PP alone is not sufficient to trigger proliferation; IL-2, IL-17, IL-22, and IL-23 have all been implicated as co-stimulatory cytokines, with IL-23 co-stimulation seen during memory response challenge [36]. This expansion of Vδ2+ cells was found to be a marker for HMB–PP-producing bacterial infection, including E. coli, K. pneumoniae, P. aeruginosa, C. striatum, and L. monocytogenes. It produced TNFα and IFNγ [8,22,37].
HMB-PP stimulated γδ T cells are notably expanded during acute bacterial peritonitis in kidney disease patients [22]. In these patients, HMB-PP stimulated γδ T cells underwent crosstalk with mesothelial cells and peritoneal fibroblasts to induce the proinflammatory response of CCL2, CXCL8, CXCL10, and IL-6 production [22]. These γδ T cells promote short-term neutrophil survival and activation and respond further to neutrophil-phagocytosed bacteria through crosstalk with monocytes [8]. Cell-cell interactions are necessary for such crosstalk, and induce an APC-like phenotype in monocytes, characterized by upregulation of CD40, CD86, and HLA-DR, causing a subsequent inflammatory positive feedback loop [9]. Neutrophils stimulated by Vδ2+ γδ T cells also obtained a unique APC-like phenotype, not found in other circulating neutrophils, again expressing CD40, CD64, CD86, HLA-DR, CD54, and HLA-ABC [38].
Interestingly, exposure to TNFα or IFNγ derived from HMB-PP stimulated γδ T cell supernatant induced a morphological change in primary omentum-derived mesothelial cells, causing them to become fibroblastic, coupled with an upregulation of the mesenchymal marker fibronectin [22]. This cellular remodeling may impact the integrity of the peritoneal membrane and could explain increased peritoneal dialysis technique failure rates in patients with bacterial peritonitis [22], making γδ T cell activation by HMB-PP a salient target for therapeutics. Taken in sum, evidence from initial studies marks γδ T cell immunometabolism as a promising field of study in better understanding the microbe and subsequent host response to bacterial invasion.
4. The Site-Specific γδ T Cell Response to S. aureus Infection
S. aureus is a gram-positive bacterium that is highly pathogenic and is the major cause of skin and soft tissue infection (SSTI), infective endocarditis, bone and joint infection, medical device-related infection, and bacteremia. [39]. Mainly, community-associated methicillin-resistant S. aureus (CA-MRSA) infection is a looming threat, claiming over 20,000 lives in the US annually [40]. Furthermore, vaccine efforts against S. aureus to date have been unsuccessful [41,42]. As a result, alternative approaches to combat the threat have become essential, and immunotherapies against S. aureus infection have emerged in recent years [43,44].
There are various non-pore-forming toxins, pore-forming toxins, and bacterial components from S. aureus that are antigenic to the host. γδ T cells have been reportedly involved in early immune responses to S. aureus infection. γδ T cells respond to staphylococcal superantigens, including staphylococcal enterotoxin A (SEA) and toxic shock syndrome toxin 1 (TSST-1) [45]. In conventional T cells, staphylococcal superantigens bind outside the MHCII-TCR antigen presenting complex, leading to rapid T cell expansion and inflammation [46].
In human adults, Vγ2+ T cell response and proliferation to SEA in specific requires APC presentation, particularly on MHC II [23,47]. SEA recognition requires the N terminal third of the toxin for partial γδ T cell activity and the N terminal two-thirds of the protein for optimal activity, most critically needing amino acid residues 20–27 [23]. Subsequent reactivity to SEA was also dependent on and specific to the Vγ9 region of the TCR, which may explain why Vγ2+ cells are largely extrathymic [47].
The γδ T cell cytokine response to superantigens primarily involves IFNγ and IL-17A. IFNγ production by γδ T cells in response to SEA proceeds through an IL-12-dependent pathway and helps induce the expansion of memory-like CD45RO+ Vγ9Vδ2 T cells [41,45]. This finding was echoed in bovine WC1+ γδ T cells [48]. On the other hand, SEA-induced IL-17A production by γδ T cells has a unique role in lung host response. In a lung immunity model, Kumar and colleagues found that γδ T cells were the primary source of proinflammatory cytokine IL-17A after the SEA challenge [49].
Interestingly, however, after the SEA challenge, there was no decrease in neutrophil and monocyte recruitment in TCRδ(−/−) mice, and αβ T cells were found to be responsible for neutrophil and monocyte recruitment to the infection site [49]. Neutrophil recruitment is critical for clearance of S. aureus: individuals with impaired neutrophil function or decreased neutrophil count display increased infection susceptibility [50]. Since IL-17A is known to play a critical role in neutrophil recruitment [51], the researchers hypothesized that IL-17A production was being compensated by CD3 + CD8− αβ T cells in TCRδ(−/−) mice [49]. In a follow-up study, IL-17 producing γδ T cells in the lung were termed lung granular γδ T cells due to their increased side scatter measured during flow cytometry analysis [52]. Lung granular γδ T cells were activated by inflammasome-derived IL-1β and IL-2 to produce IL-17 in a JAK/STAT-dependent manner, marking a novel mechanism by which γδ T cells respond to superantigen exposure [52]. Interestingly, this finding contrasts the widely accepted paradigm of IL-2 inhibition of Th17 differentiation [53], though the exact function of γδ T cell-derived IL-17 in this context has not yet been elucidated.
Unlike SEA stimulation, TSST-1 stimulation of γδ T cells upregulated secretion of proinflammatory cytokines IFNγ, TNFα, and IL-2 and suppressed anti-inflammatory IL-10 production [54]. This response was specific to TSST-1 stimulation and was not demonstrated in other staphylococcal toxins [54].
While staphylococcal superantigens were shown to activate the inflammatory response of γδ T cells, S. aureus α toxin was contrastingly found to delay IL-17+ γδ T cell recruitment to the infection site, slowing neutrophil recruitment and worsening infection in a mouse dermonecrosis model [55]. These studies indicate a critical role for γδ T cells in activating or suppressing host immune responses against S. aureus in response to toxins the bacterium produces. The following section of the review will explore current literature elucidating the host γδ T cell response to S. aureus in different infection sites and contexts.
4.1. Cutaneous Infection
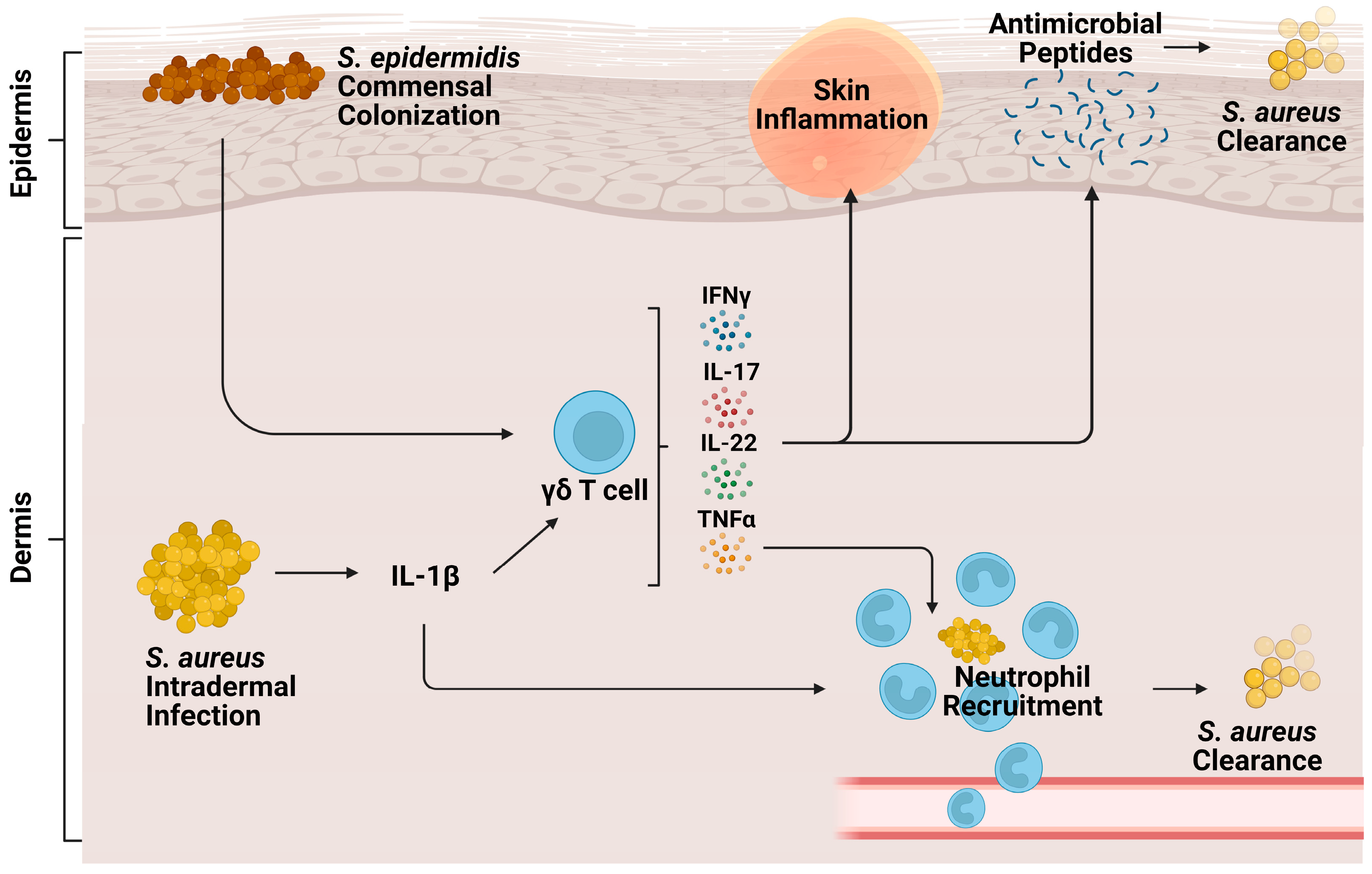
Cutaneous γδ T cells are important in early immune defense against S. aureus skin infection [56] (Figure 2). In a murine in vivo study of S. aureus skin infection, epidermal Vγ5+ γδ T cells induced by IL-1R and IL-23 signaling were protective against worsened infection; γδ T cell deficient mice had more significant lesion sizes, increased bacterial burdens, and lessened neutrophil recruitment and activity compared to both αβ T cell deficient mice and wild type (WT) mice [57]. This protective role is mediated by IL-17 production. γδ T cells comprise most of the IL-17-producing cells in S. aureus-infected skin [57,58]. Treatment of γδ T cell-deficient mice with a single dose of IL-17 abrogated the detrimental effects of γδ T cell deficiency [57]. IL-17+ γδ T and Th17 cells also play a compensatory role, promoting neutrophil recruitment in IL-1β deficient mice [59].
A study from our research group supported the importance of IL-17 producing γδ T cell populations during S. aureus skin infection, finding that these cells were being trafficked from the LNs [60]. T cell receptor (TCR) RNA sequencing revealed clonotypic expansion of Vγ6+ Vδ4+ T cells but not Vγ5+ γδ T cells in the skin and LNs [60]. This expansion was specific to S. aureus infection; however, during P. aeruginosa infection, both TRGV6 and V5 were expanded [60]. The trafficked Vγ6 + Vδ4+ T cells enhanced the neutrophilic response during secondary S. aureus infection in IL-1β deficient mice [61]. These cells produced not only IL-17 but also other proinflammatory cytokines like TNFα, IL-22, and IFNγ [60]. IL-17A and IL-17F produced by LN-trafficked γδ T cells have compensatory roles to one another in the skin, as they do during mucocutaneous S. aureus infection [62]. Both cytokines had to be neutralized to note any differences in mouse lesion size or bacterial burden after cutaneous S. aureus infection [60]. While IL-17 plays a critical role in γδ T cell signaling in cutaneous infection, it is important to note that this is not the case in all infection contexts. For example, response to early IL-1R signaling, but surprisingly not IL-17 or TNFα signaling, in γδ T cells promotes host survival and monocyte recruitment to the spleen in an S. aureus-induced bacteremia model [63].
γδ T cells also have memory-like function during skin inflammation and S. aureus infection [61,64]. In an imiquimod model of inflammation, IL-17 producing Vγ4+ γδ T cells leave the LNs and are trafficked to the skin, with previously sensitized mice showing more significant γδ T cell-induced inflammation proliferation and IL-17 production, indicating a memory-like role [64]. In the context of S. aureus infection, our colleagues found that while the primary challenge of IL-1β deficient mice led to decreased bacterial clearance, greater lesions, and impaired neutrophil abscess formation, these functions were restored upon secondary infection by LN draining γδ T cells through TLR2/MyD88 signaling to produce IFNγ and TNF [61]. Overall, inflammatory cytokine production in the skin by LN-draining γδ T cells seems to confer protective immunity against primary and secondary S. aureus infections.
Along with inherent host protective responses to S. aureus infection, commensal bacteria may protect the host against S. aureus infection by driving host antimicrobial peptide production by γδ T cells [65]. One study finds that S. epidermidis commensal colonization of the skin induces γδ T cells to upregulate perforin-2 expression, a cytolysin constitutively expressed by γδ T cells to form pores in bacterial membranes [66], along with upregulating other cytotoxicity markers against MRSA infection [67]. As such, S. epidermidis co-colonization with S. aureus on the skin led to an increased anti-S. aureus effect [67].
Lactobacilli have also been identified as commensal bacteria that may modulate host immune protection (Figure 2). One study found that in vitro co-colonization of human PBMCs with S. aureus and Lactobacillus strains dampened IFNγ secretion in γδ T cells, MAIT cells, and NK cells stimulated by S. aureus cell free supernatants [68]. However, recent other studies utilize Lactobacillus recombinant strains as vaccine models against S. aureus due to the commensal bacteria’s ability to induce a robust immune response [69,70]. Thus, it is evident that during S. aureus infection, lactobacilli have differential effects in vitro versus in vivo and must be studied contextually.
4.2. Pneumonia
In the lung, the major subsets of γδ T cells are Vγ1+ and Vγ4+ and accumulate after S. aureus-induced pneumonia [71]. While murine γδ T cells are involved in decreasing the bacterial burden and increasing neutrophil infiltration, with corresponding increases in keratinocyte-derived chemokine (KC), MIP2, GM-CSF, IL-6, and TNFα, acute lung damage was decreased in γδ T cell deficient mice, likely due to tamped inflammation from reduced neutrophil recruitment [71]. An early burst of γδ T cell-produced IL-17 was also implicated in increased lung damage post-pneumonia [71], indicating a possibly detrimental role for γδ T cells in this model.
Interestingly, nociceptor sensory neurons in the lung may have an immunosuppressive role on pulmonary γδ T cell function during MRSA infection. Selective ablation of TRPV1+, an ion channel expressed on nociceptors mediating airway allergic pathways, increased absolute Vγ1+ γδ T cell population, increased survival, and increased bacterial clearance [72]. This neuroimmunological finding is important as it marks sensory neurons as targets to protect against S. aureus pneumonia.
Cell death pathways, including the necroptotic pathway, may also suppress the host immune response against S. aureus-induced pneumonia by targeting IL-17 signaling by lung γδ T cells [73]. α toxin from S. aureus is known to activate the NLRP3 inflammasome in vitro [74]. NLRC4, an NLR family protein involved in inflammasome assembly, is upregulated during S. aureus-induced pneumonia in myeloid and non-myeloid cells [73]. Furthermore, α toxin from S. aureus induces necroptosis leading to increased IL-18 and IL-1β production and suppressed γδ T cell recruitment, dampening the IL-17 response, leading to decreased neutrophil recruitment and inflammation [73]. However, necroptotic suppression of IL-17 response is site-specific; in a surgical site S. aureus infection model, IL-17 producing γδ T cells at the wound site relied by NLRP3/IL-1β signaling for IL-17A production. It is interesting to note that these observed during infection with SH1000 S. aureus strain, but not PS80 [37].
Therefore, the pneumonia model indicates the necessity of balance in the γδ T cell response; too much IL-17 signaling may lead to excessive inflammation and tissue damage, while too little may hinder bacterial clearance.
4.3. Peritonitis
Similar to cutaneous and pulmonary infection, γδ T cells are the primary source of IL-1β-dependent IL-17 in a primary challenge during a recurrent peritonitis murine model of S. aureus infection [18]. Some mice in this study were also found to have a biphasic wave of IL-17 production, with one peak at 3 h and the second at 72 h post-infection, with Vγ4+ γδ T cells at 72 h being primed for later infection and IL-1β independent IL-17 production [18], suggesting a memory function for γδ T cells during acute peritonitis as well. In the kidney, chronic systemic S. aureus infection induced the expansion of a population of kidney-resident γδ T cells that constitutively express CD69 and provide protection against S. aureus [15]. Thus, in mice, S. aureus infection seems to expand both resident and memory γδ T cells.
In human peritoneal dialysis (PD) patients, on the other hand, it is unclear whether S. aureus-induced peritonitis expands γδ T cells or not. In one study, peritoneal Vδ2+ γδ T cells were reduced during acute peritonitis [37]. However, this was not specific to S. aureus-induced peritonitis and was used partially as an immune fingerprint to classify gram-positive acute peritonitis [37]. On the other hand, other studies found Vδ2+ γδ T cell recruitment and response to S. aureus-induced peritonitis in PD patients, possibly due to superantigen recognition [22,23].
5. γδ T Cell Response during S. pneumoniae Infection
S. pneumoniae infection is the leading cause of community-acquired pneumonia, meningitis, and bacteremia, inflicting a heavy disease burden [75]. Despite vaccination against S. pneumoniae, multidrug-resistant bacterial infections run rampant, causing 19,336 hospitalizations in the US annually and millions of dollars in medical and other costs [76]. The immune response to S. pneumoniae infection utilizes neutrophil recruitment, subsequent accumulation in the alveolar spaces, and killing through both phagocytosis and, more critically, degranulation [77,78]. Proinflammatory cytokine synthesis and release induced by Vα14+ NKT cells, another unconventional T cell subset, has been shown to promote rapid neutrophil recruitment to the infection site [79]. Alveolar macrophages function to control the resolution of inflammation [80]. γδ T cells have also been critically implicated during innate immune response to S. pneumoniae in the lung.
S. pneumoniae infection stimulated human peripheral Vγ9Vδ2 T cells in an in vitro study [81]. In mice, however, Vγ4+ γδ T cells accumulated in the lungs after pulmonary infection [78] due to their lung-homing capability [82]. TCR Vγ4(−/−) mice had increased bacterial burden, increased mortality, decreased neutrophil accumulation, and decreased TNFα and MIP2 production early after S. pneumoniae infection, indicating a role for γδ T cells during initial innate immunity [78]. In the lung, Vγ4+ γδ T cells produce IL-17 in response to infection [82]. IL-17 production was stimulated by IL-23 and IL-1β, further recruiting neutrophils [83]. Neutrophils, via NLRP3 inflammasome activation, were in turn the greatest source of IL-1β in the interstitial area of the lung, activated by alveolar macrophage-derived TNFα and pneumolysin, a pore-forming virulence toxin [83].
The lung γδ T cell population is regulated to prevent excess inflammation. IL-17-producing γδ T cells are regulated by S. pneumonia-activated regulatory T cells with upregulated TNFR2; lack of TNFR2 caused a dysregulated IL-17 response and increased lung damage due to excessive inflammation [84]. The lung’s inflammatory state may also regulate γδ T cell responses [85]. In one study, mice deficient in SOD3, the main enzyme involved in clearing damaging ROS, had increased phagosomal ROS levels [85]. This led to early neutrophil apoptosis in S. pneumoniae infection, thereby hampering the subsequent pro-inflammatory γδ T cell response [85]. As such, the above data suggest that γδ T cells have a regulated proinflammatory role during early S. pneumoniae clearance in the lung.
γδ T cells have also been found to play an important part in inflammation resolution during S. pneumoniae infection. In a murine model, the lung γδ T cell population increased by 30 fold, 7–10 days post-challenge, when no more bacterial burden was detectable [80], indicating that these cells were not involved in bacterial clearance. Instead, lung γδ T cells were cytotoxic towards both naive and challenged host-derived inflammatory alveolar macrophages and pulmonary dendritic cells, thus aiding in dampening inflammation [80], similar to S. aureus-induced pneumonia [71]. γδ T cells are also involved in non-bacterial inflammation resolution in the lungs, interacting with M2 macrophages to promote clearance of apoptotic cells during ozone-induced pulmonary inflammation [86]. In summary, early γδ T cell response to S. pneumoniae promotes neutrophil recruitment and bacterial clearance, while an end-stage response assists more during the resolution phase of infection.
6. γδ T Cell Response during L. monocytogenes Infection
L. monocytogenes is one of the deadliest food-borne pathogens and can cause listeriosis, which results in a 90% hospitalization rate [87]. Vγ9Vδ2+ T cells have a site-specific proliferative response in the liver and spleen to secreted ligands from the bacterium, indicating a possible role for γδ T cells during immunity [88,89]. HMB-PP is a particularly potent stimulator of human Vγ9Vδ2+ T cells. L. monocytogenes undergoes both the mevalonate and non-mevalonate pathways for isoprenoid biosynthesis, producing both IPP and HMB-PP. However, HMB-PP affects a greater response in γδ T cell expansion [90,91]. This is evidenced by the finding that in vitro, human Vγ9Vδ2+ T cells were more bioactive when co-cultured with an L. monocytogenes mutant strain with a deficiency in the Lytb (HMB-PP reductase) enzyme leading to HMB-PP overproduction [90]. HMB-PP is also important in developing Vγ9Vδ2 T cells with a memory phenotype, which was demonstrated in a macaque model using an attenuated L. monocytogenes immunization [91].
γδ T cell activation seems to have a protective role against L. monocytogenes infection, though the exact γδ subset involved is unclear. In a murine intraperitoneal infection model, administration of an anti-Vγ1 antibody led to increased bacterial burden in the spleen and liver, indicating a role for Vγ1Vδ6+ T cells during early infection in mice [92]. In contrast, another study found that, despite an overall increase in γδ T cell population after L. monocytogenes infection, there was a relative decrease in the frequency of Vγ1+ γδ T cells in the liver, while Vγ6+ T cells increased in frequency [93]. In another inflammation study, Vγ6+ γδ T cells also seemed to have a proportionally increased response compared to other γδ T cell subsets [94].
Intravenous injection of L. monocytogenes into a lateral tail vein led to γδ T cell activation, which, though non-essential for survival, helped control the inflammatory response in the liver by promoting macrophage influx via MCP-1 production and reducing neutrophil influx [95]. αβ T cells, as opposed to γδ T cells, are critical in survival and protection against necrotic hepatitis [95]. During early infection, IL-17-producing γδ T cells are vital in cytotoxic T cell recruitment. They act on dendritic cells to upregulate MHC I, increasing the production of IL-12, IL-6, and IL-1β, thus leading to the cross-priming and proliferation of CD8+ T cells [59]. In the murine liver, these early IL-17-producing cells are activated by IL-23 from macrophages or dendritic cells, express Vγ4 or Vγ6, and are involved not only in neutrophil recruitment and host protection but also in negatively regulating granuloma formation [59]. Vγ6Vδ1 T cells, in specific, play a protective role during infection and produce IFNγ along with IL-17 [96]. On the other hand, Vγ4+ γδ T cells are protective against CD8+ T cell-mediated liver injury in an IL-10-dependent fashion by controlling TNFα production and dysregulation [97]. IL-10 production in this context occurs in response to signaling from activated splenic macrophages and L. monocytogenes-elicited CD8+ T cells.
γδ T cells have also been shown to have an immunomodulatory response during L. monocytogenes infection. Infection in TCRδ deficient mice led to an accumulation of activated macrophages and neutrophils in the infected mice’s peritoneum, leading to overwhelming hepatic necrosis [98], indicating a suppressive role for γδ T cells. Macrophages had decreased apoptosis in TCRδ deficient mice, indicating that γδ T cells are cytotoxic towards macrophages via either the TNFα or the Fas pathway towards the end of L. monocytogenes infection [98]. The binding between γδ T cells and macrophages is TCR-mediated [99].
During L. monocytogenes infection, γδ T cells also have memory functions. In a mouse oral inoculation model that closely replicates human enteric L. monocytogenes infection, mesenteric lymph node-derived Vγ4Vδ1 T cells produced both IL-17A and IFNγ and were identified as a unique CD27−CD44hi memory subset that persisted for at least 5 months and launched a robust and rapid proliferative response to secondary challenge [19]. Though it is unclear what causes this memory-like expansion, the secondary memory response was infection route dependent; intravenous infection of mice did not elicit a similar γδ T cell expansion and response [19]. A subsequent study found that Vγ4+ γδ T cells were bona fide resident memory T cells, expressing gut-homing genes like integrin b7, CCR2, CCR5, CXCR3, and CXCR6 [20]. Though these cells had restricted motility, they rapidly formed clusters with neutrophils and monocytes in an IL-17-dependent manner at bacterial replication foci to quickly clear infection after a secondary challenge [20]. A similar γδ T cell population was found in an imiquimod sensitization model of psoriasis, in which they produced elevated levels of IL-17, upregulated IL-1R1 expression, and increasingly responded to IL-1β stimulation [64]. Interestingly, memory Vγ4+ γδ T cells were resistant to cellular senescence [100]. Their adaptive-like response to food-borne L. monocytogenes infection increased with age, making these memory T cells an essential target for antibacterial therapies in elderly patients [100]. Thus, γδ T cells have multi-faceted functions, including early protection, inflammatory resolution, and memory functions, during L. monocytogenes infection.
7. γδ T Cell Response during M. tuberculosis Infection
Tuberculosis is the leading cause of death worldwide from a single infectious agent, primarily due to latent tuberculosis infection [101]. In M. tuberculosis-infected lung cells, γδ T cells produce IL-17, dependent upon IL-23 production from infected dendritic cells in mice, and IL-2 co-stimulation and HMB-PP stimulation in primates [36,102,103]. Post expansion, IL-17 producing γδ T cells upregulate expression of ICAM-1 and LFA-1 in macrophages [104,105]. ICAM-1 and LFA-1 are involved in forming granulomas in the lung, thereby helping to sequester and kill the mycobacteria [104,105]. γδ T cell expansion also led to IL-12 production, enhancing the Th1 cytokine response of Ag-specific CD4 and CD8 T cells [36,103]. Macrophages have been implicated in the chemotaxis of γδ T cells to the infection site via secretion of IP-10, inducing changes in chemokine expression pattern [106]; after being trafficked to the pulmonary compartment, Vγ9Vδ2 T cells can differentiate into IFNγ-, perforin-, and granulysin-producing cells, thus reducing bacterial burden and increasing immune resistance against M. tuberculosis [36,103].
In vitro, HMB-PP-stimulated expansion and effector function are also driven by Th17 cytokines, particularly IL-23 [107]. In one study, IL-2 facilitated IL-23 mediated expansion of HMB-PP stimulated Vγ9Vδ2 T cells from human patients with latent tuberculosis infection [108]. However, tuberculosis infection also selectively impaired IL-23 stimulated expansion during human latent tuberculosis by inhibiting JAK-STAT signaling, leading to loss of effector function and cytokine production [108]. This indicates a mechanism by which M. tuberculosis can suppress infection clearance. Specifically, miRNAs hsa-miR-337-3p and hsa-miR-125b-5p were found to be upregulated during tuberculosis infection, inhibiting STAT3 expression and IL-23 stimulated γδ T cell expansion; silencing of these miRNAs restored expansion [108]. M. tuberculosis infection may also site-specifically impair Vγ9Vδ2 T cell activation in response to antigen recognition. In human tuberculosis patients, γδ T cells isolated from bronchoalveolar lavage had downmodulation of CD3ε, as compared to γδ T cells from the peripheral blood [109]. Therefore, the M. tuberculosis model represents an arms race between the bacteria and the corresponding innate immune response.
8. Future Clinical Role for γδ T Cells
Given the immunoprotective role of γδ T cells, several new therapies have emerged involving stimulation of γδ T cell expansion. γδ T cell-related therapeutics have to date primarily focused on anti-cancer therapeutics. Initially, bisphosphonate-based drugs, such as PAM, ZOL, and IL-2, were found to stimulate Vγ9Vδ2 T cells but were not clinically effective in cancer models [110]. However, due to MHC independence and lessened graft vs. host disease risk, γδ T cells have been identified as a potential target for allogeneic T cell transfer immunotherapy in many cancer states [110,111]. In fact, an ongoing clinical trial is using an expanded γδ T cell infusion intending to increase host defense in leukemia and myelodysplastic patients while reducing the risk of graft vs. host disease seen in conventional bone marrow transplants [110], (Clinical trial ID: NCT03533816). Additionally, induced pluripotent stem cell-derived CAR-modified γδ T cells seem to be effective against tumors in a xenograft mouse model, and there are several commercial entities now developing CAR γδ T cell therapies [110]. A recent murine study found that genetically modified γδ T cells that expressed NY-ESO-1 (a cancer-testis antigen widely expressed in a number of cancers) specific αβTCR are effective in killing NY-ESO-1 expressing tumors [112]. Interestingly, these αβ T cell-transduced γδ T cells seemed to undergo oxidative phosphorylation at a higher rate, with IFNγ production being dependent on ATP production [112]. Mitochondria have been demonstrated to play an important role in sustained CD8 T cell cytotoxicity [113], which may apply to these αβ T cell transduced γδ T cells as well. Though a further examination of the metabolic state of γδ T cells in therapeutic models is required, it could further our understanding of γδ T cell-mediated cytotoxicity, as demonstrated in cancer models.
Apart from cancer models, vaccine development in nonhuman primate tuberculosis models is currently ongoing and focuses on stimulating Vγ9Vδ2 T cells via phosphoantigen delivery. ZOL, in particular, is known to expand γδ T cells, and initial studies focused on adequately promoting γδ T cells to use during immunization. In vitro, ZOL coupled with IL-2 (ZOL/IL-2) stimulation was found to expand IFNγ and TNFα producing γδ T cells [114]. Direct ZOL/IL-2 administration led to an in vivo expansion of Vγ9Vδ2 T cells during multidrug-resistant M. tuberculosis infection [36,114]. These cells then traveled to the pulmonary compartment, producing IFNγ, perforin, and TNFα, thereby decreasing bacterial burden and disease state [36,114]. An initial proof of concept study then found that adoptive transfer of ZOL/IL-2-expanded Vγ9Vδ2 T cells in macaques infected with M. tuberculosis had reduced bacterial burden in the lung and reduced bacterial dissemination [115]. In another study, a mutant HMB-PP-producing L. monocytogenes vector was used to selectively immunize Vγ9Vδ2 T cells, which conferred protection against M. tuberculosis in macaques [116]. Vaccine development efforts against M. tuberculosis are a promising yet ongoing field of research.
There is also increasing evidence for bisphosphonate-based drugs as an immunotherapy in other bacterial infection states. ZOL activates circulating γδ T cells via monocyte accumulation of IPP and its stereoisomer DMAPP [35,117]. In one study, human septic PBMCs treated with ZOL led to monocyte activation, marked by upregulated HLA-DR, CD40, and CD64, allowing for downstream activation of Vγ9Vδ4 γδ T cells [118]. PAM, a known safe human Vγ9Vδ2 T cell-specific aminobisphosphonate drug [119], may also be a promising therapeutic agent. A chimeric SCID mouse model was used to elucidate the role of memory Vγ9Vδ2 T cells in antibacterial resistance, finding that treatment with PAM rendered PBMCs 100x more efficient in clearing both S. aureus and E. coli infection [120].
9. Conclusions
In summary, this review outlines the critical role of γδ T cells during bacterial infection, focusing on the microenvironment and promising advances in studying immune metabolism. γδ T cells are part in all stages of infection—early defense, inflammatory resolution, and defense during reinfection—making these unconventional T lymphocytes a potential target for therapeutics against hard-to-treat bacterial infection. As such, further research must focus on utilizing γδ T cells in immunotherapies to provide lasting protection against the threat of antibiotic-resistant microbes.
Author Contributions
Conceptualization, M.P.A.; writing—original draft preparation, N.N.; writing—review and editing, N.N. and M.P.A. visualization, N.N. and M.P.A.; supervision, M.P.A.; funding acquisition, M.P.A. All authors have read and agreed to the published version of the manuscript.
Funding
MPA is funded by Research Career Development Award from Dermatology Foundation, USA (Funding number: 140819-22043024).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carding, S.R.; Egan, P.J. γδ T Cells: Functional Plasticity and Heterogeneity. Nat. Rev. Immunol. 2002, 2, 336–345. [Google Scholar] [CrossRef]
- Kabelitz, D.; Wesch, D.; Hinz, T. Gamma Delta T Cells, Their T Cell Receptor Usage and Role in Human Diseases. In Springer Seminars in Immunopathology; Springer International: Berlin, Germany, 1999; Volume 21, pp. 55–75. [Google Scholar]
- Brown, A.F.; Murphy, A.G.; Lalor, S.J.; Leech, J.M.; O’Keeffe, K.M.; Mac Aogáin, M.; O’Halloran, D.P.; Lacey, K.A.; Tavakol, M.; Hearnden, C.H.; et al. Memory Th1 Cells Are Protective in Invasive Staphylococcus aureus Infection. PLoS Pathog. 2015, 11, e1005226. [Google Scholar] [CrossRef]
- Mayassi, T.; Barreiro, L.B.; Rossjohn, J.; Jabri, B. A Multilayered Immune System through the Lens of Unconventional T Cells. Nature 2021, 595, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Wesch, D. Analysis of the TCR Vgamma Repertoire in Healthy Donors and HIV-1-Infected Individuals. Int. Immunol. 1998, 10, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Rosenthal, C.J.; Penn, N.E.; Dunn, Z.S.; Zhou, Y.; Yang, L. Human γδ T Cell Subsets and Their Clinical Applications for Cancer Immunotherapy. Cancers 2022, 14, 3005. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lin, L.; Xiao, Z.; Li, M.; Wu, X.; Li, W.; Li, X.; Zhao, Q.; Wu, Y.; Zhang, H.; et al. Protective Role of γδ T Cells in Different Pathogen Infections and Its Potential Clinical Application. J. Immunol. Res. 2018, 2018, 5081634. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.S.; Lin, C.-Y.; Roberts, G.W.; Heuston, S.; Brown, A.C.; Chess, J.A.; Toleman, M.A.; Gahan, C.G.M.; Hill, C.; Parish, T.; et al. Human Neutrophil Clearance of Bacterial Pathogens Triggers Antimicrobial γδ T Cell Responses in Early Infection. PLoS Pathog. 2011, 7, e1002040. [Google Scholar] [CrossRef]
- Eberl, M.; Roberts, G.W.; Meuter, S.; Williams, J.D.; Topley, N.; Moser, B. A Rapid Crosstalk of Human γδ T Cells and Monocytes Drives the Acute Inflammation in Bacterial Infections. PLoS Pathog. 2009, 5, e1000308. [Google Scholar] [CrossRef]
- Leslie, D.S.; Vincent, M.S.; Spada, F.M.; Das, H.; Sugita, M.; Morita, C.T.; Brenner, M.B. CD1-Mediated γ/δ T Cell Maturation of Dendritic Cells. J. Exp. Med. 2002, 196, 1575–1584. [Google Scholar] [CrossRef]
- Skeen, M.J.; Freeman, M.M.; Ziegler, H.K. Changes in Peritoneal Myeloid Populations and Their Proinflammatory Cytokine Expression during Infection with Listeria monocytogenes Are Altered in the Absence of Gamma/Delta T Cells. J. Leukoc. Biol. 2004, 76, 104–115. [Google Scholar] [CrossRef]
- Cai, Y.; Xue, F.; Fleming, C.; Yang, J.; Ding, C.; Ma, Y.; Liu, M.; Zhang, H.; Zheng, J.; Xiong, N.; et al. Differential Developmental Requirement and Peripheral Regulation for Dermal Vγ4 and Vγ6T17 Cells in Health and Inflammation. Nat. Commun. 2014, 5, 3986. [Google Scholar] [CrossRef]
- Vermijlen, D.; Prinz, I. Ontogeny of Innate T Lymphocytes—Some Innate Lymphocytes Are More Innate than Others. Front. Immunol. 2014, 5, 486. [Google Scholar] [CrossRef]
- Sutoh, Y.; Mohamed, R.H.; Kasahara, M. Origin and Evolution of Dendritic Epidermal T Cells. Front. Immunol. 2018, 9, 1059. [Google Scholar] [CrossRef]
- Bertram, T.; Reimers, D.; Lory, N.C.; Schmidt, C.; Schmid, J.C.; Heinig, L.; Bradtke, P.; Rattay, G.; Zielinski, S.; Hellmig, M.; et al. Kidney-Resident Innate-like Memory γδ T Cells Control Chronic Staphylococcus aureus Infection of Mice. Proc. Natl. Acad. Sci. USA 2023, 120, e2210490120. [Google Scholar] [CrossRef]
- Fiala, G.J.; Gomes, A.Q.; Silva-Santos, B. From Thymus to Periphery: Molecular Basis of Effector γδ-T Cell Differentiation. Immunol. Rev. 2020, 298, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, Y.; Lau, Y.-L.; Tu, W. γδ-T Cells: An Unpolished Sword in Human Anti-Infection Immunity. Cell. Mol. Immunol. 2013, 10, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.G.; O’Keeffe, K.M.; Lalor, S.J.; Maher, B.M.; Mills, K.H.G.; McLoughlin, R.M. Staphylococcus aureus Infection of Mice Expands a Population of Memory γδ T Cells That Are Protective against Subsequent Infection. J. Immunol. 2014, 192, 3697–3708. [Google Scholar] [CrossRef]
- Sheridan, B.S.; Romagnoli, P.A.; Pham, Q.-M.; Fu, H.-H.; Alonzo, F.; Schubert, W.-D.; Freitag, N.E.; Lefrançois, L. γδ T Cells Exhibit Multifunctional and Protective Memory in Intestinal Tissues. Immunity 2013, 39, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, P.A.; Sheridan, B.S.; Pham, Q.-M.; Lefrançois, L.; Khanna, K.M. IL-17A-Producing Resident Memory γδ T Cells Orchestrate the Innate Immune Response to Secondary Oral Listeria monocytogenes Infection. Proc. Natl. Acad. Sci. USA 2016, 113, 8502–8507. [Google Scholar] [CrossRef] [PubMed]
- Morita, C.T.; Jin, C.; Sarikonda, G.; Wang, H. Nonpeptide Antigens, Presentation Mechanisms, and Immunological Memory of Human Vγ2Vδ2 T Cells: Discriminating Friend from Foe through the Recognition of Prenyl Pyrophosphate Antigens. Immunol. Rev. 2007, 215, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, A.R.; Kift-Morgan, A.; Lopez-Anton, M.; Friberg, I.M.; Zhang, J.; Brook, A.C.; Roberts, G.W.; Donovan, K.L.; Colmont, C.S.; Toleman, M.A.; et al. Unconventional Human T Cells Accumulate at the Site of Infection in Response to Microbial Ligands and Induce Local Tissue Remodeling. J. Immunol. 2016, 197, 2195–2207. [Google Scholar] [CrossRef]
- Morita, C.T.; Li, H.; Lamphear, J.G.; Rich, R.R.; Fraser, J.D.; Mariuzza, R.A.; Lee, H.K. Superantigen Recognition by γδ T Cells. Immunity 2001, 14, 331–344. [Google Scholar] [CrossRef]
- Vermijlen, D.; Gatti, D.; Kouzeli, A.; Rus, T.; Eberl, M. γδ T Cell Responses: How Many Ligands Will It Take till We Know? In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2018; Volume 84, pp. 75–86. [Google Scholar] [CrossRef]
- Rosenberg, G.; Riquelme, S.; Prince, A.; Avraham, R. Immunometabolic Crosstalk during Bacterial Infection. Nat. Microbiol. 2022, 7, 497–507. [Google Scholar] [CrossRef]
- Jensen, K.D.C.; Su, X.; Shin, S.; Li, L.; Youssef, S.; Yamasaki, S.; Steinman, L.; Saito, T.; Locksley, R.M.; Davis, M.M.; et al. Thymic Selection Determines γδ T Cell Effector Fate: Antigen-Naive Cells Make Interleukin-17 and Antigen-Experienced Cells Make Interferon γ. Immunity 2008, 29, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Ribot, J.C.; deBarros, A.; Pang, D.J.; Neves, J.F.; Peperzak, V.; Roberts, S.J.; Girardi, M.; Borst, J.; Hayday, A.C.; Pennington, D.J.; et al. CD27 Is a Thymic Determinant of the Balance between Interferon-γ- and Interleukin 17–Producing γδ T Cell Subsets. Nat. Immunol. 2009, 10, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Zuberbuehler, M.K.; Parker, M.E.; Wheaton, J.D.; Espinosa, J.R.; Salzler, H.R.; Park, E.; Ciofani, M. The Transcription Factor C-Maf Is Essential for the Commitment of IL-17-Producing γδ T Cells. Nat. Immunol. 2019, 20, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Lopes, N.; McIntyre, C.; Martin, S.; Raverdeau, M.; Sumaria, N.; Kohlgruber, A.C.; Fiala, G.J.; Agudelo, L.Z.; Dyck, L.; Kane, H.; et al. Distinct Metabolic Programs Established in the Thymus Control Effector Functions of γδ T Cell Subsets in Tumor Microenvironments. Nat. Immunol. 2021, 22, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cai, Y.; Hu, X.; Ding, C.; He, L.; Zhang, X.; Chen, F.; Yan, J. Differential Metabolic Requirement Governed by Transcription Factor C-Maf Dictates Innate γδT17 Effector Functionality in Mice and Humans. Sci. Adv. 2022, 8, eabm9120. [Google Scholar] [CrossRef] [PubMed]
- Frascoli, M.; Ferraj, E.; Miu, B.; Malin, J.; Spidale, N.A.; Cowan, J.; Shissler, S.C.; Brink, R.; Xu, Y.; Cyster, J.G.; et al. Skin gδ T Cell Inflammatory Responses Are Hardwired in the Thymus by Oxysterol Sensing via GPR183 and Calibrated by Dietary Cholesterol. Immunity 2023, 56, 562–575.e6. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Cao, G.; Sun, G.; Zhu, L.; Tian, Y.; Song, Y.; Guo, C.; Wang, X.; Zhong, J.; Zhou, W.; et al. GLS1-Mediated Glutaminolysis Unbridled by MALT1 Protease Promotes Psoriasis Pathogenesis. J. Clin. Investig. 2020, 130, 5180–5196. [Google Scholar] [CrossRef]
- Meng, Z.; Cao, G.; Yang, Q.; Yang, H.; Hao, J.; Yin, Z. Metabolic Control of γδ T Cell Function. Infect. Microbes Dis. 2021, 3, 142–148. [Google Scholar] [CrossRef]
- Hu, Y.-M.; Yeh, C.-L.; Pai, M.-H.; Lee, W.-Y.; Yeh, S.-L. Glutamine Administration Modulates Lung γδ T Lymphocyte Expression in Mice with Polymicrobial Sepsis. Shock 2014, 41, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, T.; Fichtner, A.S.; Karunakaran, M.M. An Update on the Molecular Basis of Phosphoantigen Recognition by Vγ9Vδ2 T Cells. Cells 2020, 9, 1433. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W. Protective Immune Responses of Major Vγ2Vδ2 T-Cell Subset in M. tuberculosis Infection. Curr. Opin. Immunol. 2016, 42, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Roberts, G.W.; Kift-Morgan, A.; Donovan, K.L.; Topley, N.; Eberl, M. Pathogen-Specific Local Immune Fingerprints Diagnose Bacterial Infection in Peritoneal Dialysis Patients. J. Am. Soc. Nephrol. 2013, 24, 2002–2009. [Google Scholar] [CrossRef]
- Davey, M.S.; Morgan, M.P.; Liuzzi, A.R.; Tyler, C.J.; Khan, M.W.A.; Szakmany, T.; Hall, J.E.; Moser, B.; Eberl, M. Microbe-Specific Unconventional T Cells Induce Human Neutrophil Differentiation into Antigen Cross-Presenting Cells. J. Immunol. 2014, 193, 3704–3716. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Kourtis, A.P.; Hatfield, K.; Baggs, J.; Mu, Y.; See, I.; Epson, E.; Nadle, J.; Kainer, M.A.; Dumyati, G.; Petit, S.; et al. Vital Signs: Epidemiology and Recent Trends in Methicillin-Resistant and in Methicillin-Susceptible Staphylococcus aureus Bloodstream Infections—United States. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 214–219. [Google Scholar] [CrossRef]
- Cooper, A.J.R.; Lalor, S.J.; McLoughlin, R.M. Activation of Human Vδ2+ γδ T Cells by Staphylococcus aureus Promotes Enhanced Anti-Staphylococcal Adaptive Immunity. J. Immunol. 2020, 205, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Clegg, J.; Soldaini, E.; McLoughlin, R.M.; Rittenhouse, S.; Bagnoli, F.; Phogat, S. Staphylococcus aureus Vaccine Research and Development: The Past, Present and Future, Including Novel Therapeutic Strategies. Front. Immunol. 2021, 12, 705360. [Google Scholar] [CrossRef] [PubMed]
- Alphonse, M.P.; Rubens, J.H.; Ortines, R.V.; Orlando, N.A.; Patel, A.M.; Dikeman, D.; Wang, Y.; Vuong, I.; Joyce, D.P.; Zhang, J.; et al. Pan-Caspase Inhibition as a Potential Host-Directed Immunotherapy against MRSA and Other Bacterial Skin Infections. Sci. Transl. Med. 2021, 13, eabe9887. [Google Scholar] [CrossRef]
- Cahill, E.; Oladipo, O.O.; Dikeman, D.; Prifti, D.; Mento, S.J.; Miller, L.S.; Alphonse, M.P. An Oral Caspase Inhibitor as Monotherapy or with Antibiotics Eradicates MRSA Skin Infections in Mice. Drug Dev. Res. 2023, 84, 1567–1571. [Google Scholar] [CrossRef] [PubMed]
- Mata Forsberg, M.; Arasa, C.; Van Zwol, W.; Uzunçayır, S.; Schönbichler, A.; Regenthal, P.; Schelin, J.; Lindkvist-Petersson, K.; Björkander, S.; Sverremark-Ekström, E. Activation of Human γδ T Cells and NK Cells by Staphylococcal Enterotoxins Requires Both Monocytes and Conventional T Cells. J. Leukoc. Biol. 2022, 111, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal Enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef] [PubMed]
- Rust, C.J.J.; Verreck, F.; Vietor, H.; Koning, F. Specific Recognition of Staphylococcal Enterotoxin A by Human T Cells Bearing Receptors with the Vγ9 Region. Nature 1990, 346, 572–574. [Google Scholar] [CrossRef]
- Fikri, Y.; Denis, O.; Pastoret, P.-P.; Nyabenda, J. Purified Bovine WC1+ γδ T Lymphocytes Are Activated by Staphylococcal Enterotoxins and Toxic Shock Syndrome Toxin-1 Superantigens: Proliferation Response, TCR Vγ Profile and Cytokines Expression. Immunol. Lett. 2001, 77, 87–95. [Google Scholar] [CrossRef]
- Kumar, S.; Colpitts, S.L.; Ménoret, A.; Budelsky, A.L.; Lefrancois, L.; Vella, A.T. Rapid αβ T-Cell Responses Orchestrate Innate Immunity in Response to Staphylococcal Enterotoxin A. Mucosal Immunol. 2013, 6, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.S.; Cho, J.S. Immunity against Staphylococcus aureus Cutaneous Infections. Nat. Rev. Immunol. 2011, 11, 505–518. [Google Scholar] [CrossRef]
- Zenobia, C.; Hajishengallis, G. Basic Biology and Role of Interleukin-17 in Immunity and Inflammation. Periodontology 2000 2015, 69, 142–159. [Google Scholar] [CrossRef]
- Ménoret, A.; Buturla, J.A.; Xu, M.M.; Svedova, J.; Kumar, S.; Rathinam, V.A.K.; Vella, A.T. T Cell-Directed IL-17 Production by Lung Granular γδ T Cells Is Coordinated by a Novel IL-2 and IL-1β Circuit. Mucosal Immunol. 2018, 11, 1398–1407. [Google Scholar] [CrossRef]
- Laurence, A.; Tato, C.M.; Davidson, T.S.; Kanno, Y.; Chen, Z.; Yao, Z.; Blank, R.B.; Meylan, F.; Siegel, R.; Hennighausen, L.; et al. Interleukin-2 Signaling via STAT5 Constrains T Helper 17 Cell Generation. Immunity 2007, 26, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Kalyan, S.; Chow, A.W. Human Peripheral γδ T Cells Potentiate the Early Proinflammatory Cytokine Response to Staphylococcal Toxic Shock Syndrome Toxin–1. J. Infect. Dis. 2004, 189, 1892–1896. [Google Scholar] [CrossRef] [PubMed]
- Tkaczyk, C.; Hamilton, M.M.; Datta, V.; Yang, X.P.; Hilliard, J.J.; Stephens, G.L.; Sadowska, A.; Hua, L.; O’Day, T.; Suzich, J.; et al. Staphylococcus aureus Alpha Toxin Suppresses Effective Innate and Adaptive Immune Responses in a Murine Dermonecrosis Model. PLoS ONE 2013, 8, e75103. [Google Scholar] [CrossRef] [PubMed]
- Mölne, L.; Corthay, A.; Holmdahl, R.; Tarkowski, A. Role of Gamma/Delta T Cell Receptor-Expressing Lymphocytes in Cutaneous Infection Caused by Staphylococcus aureus. Clin. Exp. Immunol. 2003, 132, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Pietras, E.M.; Garcia, N.C.; Ramos, R.I.; Farzam, D.M.; Monroe, H.R.; Magorien, J.E.; Blauvelt, A.; Kolls, J.K.; Cheung, A.L.; et al. IL-17 Is Essential for Host Defense against Cutaneous Staphylococcus aureus Infection in Mice. J. Clin. Investig. 2010, 120, 1762–1773. [Google Scholar] [CrossRef] [PubMed]
- Myles, I.A.; Fontecilla, N.M.; Valdez, P.A.; Vithayathil, P.J.; Naik, S.; Belkaid, Y.; Ouyang, W.; Datta, S.K. Signaling via the IL-20 Receptor Inhibits Cutaneous Production of IL-1β and IL-17A to Promote Infection with Methicillin-Resistant Staphylococcus aureus. Nat. Immunol. 2013, 14, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Hamada, S.; Umemura, M.; Shiono, T.; Tanaka, K.; Yahagi, A.; Begum, M.D.; Oshiro, K.; Okamoto, Y.; Watanabe, H.; Kawakami, K.; et al. IL-17A Produced by Gammadelta T Cells Plays a Critical Role in Innate Immunity against Listeria monocytogenes Infection in the Liver. J. Immunol. 2008, 181, 3456–3463. [Google Scholar] [CrossRef] [PubMed]
- Marchitto, M.C.; Dillen, C.A.; Liu, H.; Miller, R.J.; Archer, N.K.; Ortines, R.V.; Alphonse, M.P.; Marusina, A.I.; Merleev, A.A.; Wang, Y.; et al. Clonal Vγ6 + Vδ4 + T Cells Promote IL-17–Mediated Immunity against Staphylococcus aureus Skin Infection. Proc. Natl. Acad. Sci. USA 2019, 116, 10917–10926. [Google Scholar] [CrossRef]
- Dillen, C.A.; Pinsker, B.L.; Marusina, A.I.; Merleev, A.A.; Farber, O.N.; Liu, H.; Archer, N.K.; Lee, D.B.; Wang, Y.; Ortines, R.V.; et al. Clonally Expanded γδ T Cells Protect against Staphylococcus aureus Skin Reinfection. J. Clin. Investig. 2018, 128, 1026–1042. [Google Scholar] [CrossRef]
- Ishigame, H.; Kakuta, S.; Nagai, T.; Kadoki, M.; Nambu, A.; Komiyama, Y.; Fujikado, N.; Tanahashi, Y.; Akitsu, A.; Kotaki, H.; et al. Differential Roles of Interleukin-17A and -17F in Host Defense against Mucoepithelial Bacterial Infection and Allergic Responses. Immunity 2009, 30, 108–119. [Google Scholar] [CrossRef]
- Wang, Y.; Ahmadi, M.Z.; Dikeman, D.A.; Youn, C.; Archer, N.K. γδ T Cell-Intrinsic IL-1R Promotes Survival during Staphylococcus aureus Bacteremia. Front. Immunol. 2023, 14, 1171934. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valle, F.; Gray, E.E.; Cyster, J.G. Inflammation Induces Dermal Vγ4+ γδT17 Memory-like Cells That Travel to Distant Skin and Accelerate Secondary IL-17-Driven Responses. Proc. Natl. Acad. Sci. USA 2015, 112, 8046–8051. [Google Scholar] [CrossRef]
- Christensen, G.J.M.; Brüggemann, H. Bacterial Skin Commensals and Their Role as Host Guardians. Benef. Microbes 2014, 5, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Strbo, N.; Pastar, I.; Romero, L.; Chen, V.; Vujanac, M.; Sawaya, A.P.; Jozic, I.; Ferreira, A.D.F.; Wong, L.L.; Head, C.; et al. Single Cell Analyses Reveal Specific Distribution of Anti-Bacterial Molecule Perforin-2 in Human Skin and Its Modulation by Wounding and Staphylococcus aureus Infection. Exp. Dermatol. 2019, 28, 225–232. [Google Scholar] [CrossRef]
- Pastar, I.; O’Neill, K.; Padula, L.; Head, C.R.; Burgess, J.L.; Chen, V.; Garcia, D.; Stojadinovic, O.; Hower, S.; Plano, G.V.; et al. Staphylococcus Epidermidis Boosts Innate Immune Response by Activation of Gamma Delta T Cells and Induction of Perforin-2 in Human Skin. Front. Immunol. 2020, 11, 550946. [Google Scholar] [CrossRef]
- Johansson, M.A.; Björkander, S.; Mata Forsberg, M.; Qazi, K.R.; Salvany Celades, M.; Bittmann, J.; Eberl, M.; Sverremark-Ekström, E. Probiotic Lactobacilli Modulate Staphylococcus aureus-Induced Activation of Conventional and Unconventional T Cells and NK Cells. Front. Immunol. 2016, 7, 273. [Google Scholar] [CrossRef]
- Pan, N.; Liu, B.; Bao, X.; Zhang, H.; Sheng, S.; Liang, Y.; Pan, H.; Wang, X. Oral Delivery of Novel Recombinant Lactobacillus Elicit High Protection against Staphylococcus aureus Pulmonary and Skin Infections. Vaccines 2021, 9, 984. [Google Scholar] [CrossRef]
- Pan, N.; Liu, Y.; Zhang, H.; Xu, Y.; Bao, X.; Sheng, S.; Liang, Y.; Liu, B.; Lyu, Y.; Li, H.; et al. Oral Vaccination with Engineered Probiotic Limosilactobacillus reuteri Has Protective Effects against Localized and Systemic Staphylococcus aureus Infection. Microbiol. Spectr. 2023, 11, e0367322. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Liu, T.; Zhou, W.-Y.; Zhuang, Y.; Peng, L.; Zhang, J.; Yin, Z.-N.; Mao, X.; Guo, G.; Shi, Y.; et al. Role of Gamma-Delta T Cells in Host Response against Staphylococcus aureus-Induced Pneumonia. BMC Immunol. 2012, 13, 38. [Google Scholar] [CrossRef]
- Baral, P.; Umans, B.D.; Li, L.; Wallrapp, A.; Bist, M.; Kirschbaum, T.; Wei, Y.; Zhou, Y.; Kuchroo, V.K.; Burkett, P.R.; et al. Nociceptor Sensory Neurons Suppress Neutrophil and γδ T Cell Responses in Bacterial Lung Infections and Lethal Pneumonia. Nat. Med. 2018, 24, 417–426. [Google Scholar] [CrossRef]
- Paudel, S.; Ghimire, L.; Jin, L.; Baral, P.; Cai, S.; Jeyaseelan, S. NLRC4 Suppresses IL-17A-Mediated Neutrophil-Dependent Host Defense through Upregulation of IL-18 and Induction of Necroptosis during Gram-Positive Pneumonia. Mucosal Immunol. 2019, 12, 247–257. [Google Scholar] [CrossRef]
- Craven, R.R.; Gao, X.; Allen, I.C.; Gris, D.; Wardenburg, J.B.; McElvania-TeKippe, E.; Ting, J.P.; Duncan, J.A. Staphylococcus aureus α-Hemolysin Activates the NLRP3-Inflammasome in Human and Mouse Monocytic Cells. PLoS ONE 2009, 4, e7446. [Google Scholar] [CrossRef]
- Kim, L.; McGee, L.; Tomczyk, S.; Beall, B. Biological and Epidemiological Features of Antibiotic-Resistant Streptococcus pneumoniae in Pre- and Post-Conjugate Vaccine Eras: A United States Perspective. Clin. Microbiol. Rev. 2016, 29, 525–552. [Google Scholar] [CrossRef]
- Reynolds, C.A.; Finkelstein, J.A.; Ray, G.T.; Moore, M.R.; Huang, S.S. Attributable Healthcare Utilization and Cost of Pneumonia Due to Drug-Resistant Streptococcus pneumonia: A Cost Analysis. Antimicrob. Resist. Infect. Control 2014, 3, 16. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Kimmey, J.M. Neutrophil Recruitment in Pneumococcal Pneumonia. Front. Cell. Infect. Microbiol. 2022, 12, 894644. [Google Scholar] [CrossRef] [PubMed]
- Nakasone, C.; Yamamoto, N.; Nakamatsu, M.; Kinjo, T.; Miyagi, K.; Uezu, K.; Nakamura, K.; Higa, F.; Ishikawa, H.; O’Brien, R.L.; et al. Accumulation of Gamma/Delta T Cells in the Lungs and Their Roles in Neutrophil-Mediated Host Defense against Pneumococcal Infection. Microbes Infect. 2007, 9, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Yamamoto, N.; Kinjo, Y.; Miyagi, K.; Nakasone, C.; Uezu, K.; Kinjo, T.; Nakayama, T.; Taniguchi, M.; Saito, A. Critical Role of Vα14+ Natural Killer T Cells in the Innate Phase of Host Protection against Streptococcus pneumoniae Infection. Eur. J. Immunol. 2003, 33, 3322–3330. [Google Scholar] [CrossRef] [PubMed]
- Kirby, A.; Newton, D.; Carding, S.; Kaye, P. Pulmonary Dendritic Cells and Alveolar Macrophages Are Regulated by γδ T Cells during the Resolution of S. pneumoniae-induced Inflammation. J. Pathol. 2007, 212, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.; Kabelitz, D. Preferential Activation of Peripheral Blood Vγ9+γ/δ T Cells by Group A, B and C but Not Group D or F Streptococci. Clin. Exp. Immunol. 2008, 89, 301–304. [Google Scholar] [CrossRef] [PubMed]
- De Souza Costa, M.F.; Bastos Trigo De Negreiros, C.; Ugarte Bornstein, V.; Hemmi Valente, R.; Mengel, J.; Henriques, M.D.G.; Farias Benjamim, C.; Penido, C. Murine IL-17+ Vγ4 T Lymphocytes Accumulate in the Lungs and Play a Protective Role during Severe Sepsis. BMC Immunol. 2015, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Hassane, M.; Demon, D.; Soulard, D.; Fontaine, J.; Keller, L.E.; Patin, E.C.; Porte, R.; Prinz, I.; Ryffel, B.; Kadioglu, A.; et al. Neutrophilic NLRP3 Inflammasome-Dependent IL-1β Secretion Regulates the γδT17 Cell Response in Respiratory Bacterial Infections. Mucosal Immunol. 2017, 10, 1056–1068. [Google Scholar] [CrossRef]
- Xu, R.; Jacques, L.C.; Khandaker, S.; Beentjes, D.; Leon-Rios, M.; Wei, X.; French, N.; Neill, D.R.; Kadioglu, A. TNFR2+ Regulatory T Cells Protect against Bacteremic Pneumococcal Pneumonia by Suppressing IL-17A-Producing γδ T Cells in the Lung. Cell Rep. 2023, 42, 112054. [Google Scholar] [CrossRef]
- Anthony, D.; Papanicolaou, A.; Wang, H.; Seow, H.J.; To, E.E.; Yatmaz, S.; Anderson, G.P.; Wijburg, O.; Selemidis, S.; Vlahos, R.; et al. Excessive Reactive Oxygen Species Inhibit IL-17A+ Γδ T Cells and Innate Cellular Responses to Bacterial Lung Infection. Antioxid. Redox Signal. 2020, 32, 943–956. [Google Scholar] [CrossRef]
- Mathews, J.A.; Kasahara, D.I.; Ribeiro, L.; Wurmbrand, A.P.; Ninin, F.M.C.; Shore, S.A. γδ T Cells Are Required for M2 Macrophage Polarization and Resolution of Ozone-Induced Pulmonary Inflammation in Mice. PLoS ONE 2015, 10, e0131236. [Google Scholar] [CrossRef] [PubMed]
- Matle, I.; Mbatha, K.R.; Madoroba, E. A Review of Listeria Monocytogenes from Meat and Meat Products: Epidemiology, Virulence Factors, Antimicrobial Resistance and Diagnosis. Onderstepoort J. Vet. Res. 2020, 87, e1–e20. [Google Scholar] [CrossRef] [PubMed]
- Belles, C.; Kuhl, A.K.; Donoghue, A.J.; Sano, Y.; O’Brien, R.L.; Born, W.; Bottomly, K.; Carding, S.R. Bias in the Gamma Delta T Cell Response to Listeria monocytogenes. V Delta 6.3+ Cells Are a Major Component of the Gamma Delta T Cell Response to Listeria monocytogenes. J. Immunol. 1996, 156, 4280–4289. [Google Scholar] [CrossRef] [PubMed]
- Munk, M.E.; Elser, C.; Kaufmann, S.H. Human Gamma/Delta T-Cell Response to Listeria monocytogenes Protein Components In Vitro. Immunology 1996, 87, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Gahan, C.G.M.; Kollas, A.-K.; Hintz, M.; Hill, C.; Jomaa, H.; Eberl, M. The Interplay between Classical and Alternative Isoprenoid Biosynthesis Controls γδ T Cell Bioactivity of Listeria monocytogenes. FEBS Lett. 2004, 561, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Frencher, J.T.; Shen, H.; Yan, L.; Wilson, J.O.; Freitag, N.E.; Rizzo, A.N.; Chen, C.Y.; Chen, Z.W. HMBPP-Deficient Listeria Mutant Immunization Alters Pulmonary/Systemic Responses, Effector Functions, and Memory Polarization of Vγ2Vδ2 T Cells. J. Leukoc. Biol. 2014, 96, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Nakamura; Matsuzaki; Nomoto. The Protective Role of T-cell Receptor Vγ1+ T Cells in Primary Infection with Listeria monocytogenes. Immunology 1999, 96, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Roark, C.E.; Vollmer, M.K.; Campbell, P.A.; Born, W.K.; O’Brien, R.L. Response of a Gamma Delta+ T Cell Receptor Invariant Subset during Bacterial Infection. J. Immunol. 1996, 156, 2214–2220. [Google Scholar] [CrossRef]
- Roark, C.L.; Aydintug, M.K.; Lewis, J.; Yin, X.; Lahn, M.; Hahn, Y.-S.; Born, W.K.; Tigelaar, R.E.; O’Brien, R.L. Subset-Specific, Uniform Activation among Vγ6/Vδ1+ γδ T Cells Elicited by Inflammation. J. Leukoc. Biol. 2004, 75, 68–75. [Google Scholar] [CrossRef] [PubMed]
- DiTirro, J.; Rhoades, E.R.; Roberts, A.D.; Burke, J.M.; Mukasa, A.; Cooper, A.M.; Frank, A.A.; Born, W.K.; Orme, I.M. Disruption of the Cellular Inflammatory Response to Listeria monocytogenes Infection in Mice with Disruptions in Targeted Genes. Infect. Immun. 1998, 66, 2284–2289. [Google Scholar] [CrossRef] [PubMed]
- Hamada, S.; Umemura, M.; Shiono, T.; Hara, H.; Kishihara, K.; Tanaka, K.; Mayuzumi, H.; Ohta, T.; Matsuzaki, G. Importance of Murine Vdelta1gammadelta T Cells Expressing Interferon-Gamma and Interleukin-17A in Innate Protection against Listeria monocytogenes Infection. Immunology 2008, 125, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, K.A.; Andrew, E.M.; Newton, D.J.; Tramonti, D.; Carding, S.R. A Subset of IL-10-Producing Gammadelta T Cells Protect the Liver from Listeria-Elicited, CD8(+) T Cell-Mediated Injury. Eur. J. Immunol. 2008, 38, 2274–2283. [Google Scholar] [CrossRef]
- Egan, P.J.; Carding, S.R. Downmodulation of the Inflammatory Response to Bacterial Infection by Gammadelta T Cells Cytotoxic for Activated Macrophages. J. Exp. Med. 2000, 191, 2145–2158. [Google Scholar] [CrossRef]
- Dalton, J.E.; Pearson, J.; Scott, P.; Carding, S.R. The Interaction of Gamma Delta T Cells with Activated Macrophages Is a Property of the V Gamma 1 Subset. J. Immunol. 2003, 171, 6488–6494. [Google Scholar] [CrossRef]
- Khairallah, C.; Chu, T.H.; Qiu, Z.; Imperato, J.N.; Yang, D.; Sheridan, B.S. The Accumulation of Vγ4 T Cells with Aging Is Associated with an Increased Adaptive Vγ4 T Cell Response after Food-borne Listeria monocytogenes Infection of Mice. Immun. Ageing 2022, 19, 19. [Google Scholar] [CrossRef]
- Ragonnet, R.; Trauer, J.M.; Geard, N.; Scott, N.; McBryde, E.S. Profiling Mycobacterium Tuberculosis Transmission and the Resulting Disease Burden in the Five Highest Tuberculosis Burden Countries. BMC Med. 2019, 17, 208. [Google Scholar] [CrossRef]
- Lockhart, E.; Green, A.M.; Flynn, J.L. IL-17 Production Is Dominated by γδ T Cells Rather than CD4 T Cells during Mycobacterium tuberculosis Infection. J. Immunol. 2006, 177, 4662–4669. [Google Scholar] [CrossRef]
- Chen, C.Y.; Yao, S.; Huang, D.; Wei, H.; Sicard, H.; Zeng, G.; Jomaa, H.; Larsen, M.H.; Jacobs, W.R.; Wang, R.; et al. Phosphoantigen/IL2 Expansion and Differentiation of Vγ2Vδ2 T Cells Increase Resistance to Tuberculosis in Nonhuman Primates. PLoS Pathog. 2013, 9, e1003501. [Google Scholar] [CrossRef] [PubMed]
- Okamoto Yoshida, Y.; Umemura, M.; Yahagi, A.; O’Brien, R.L.; Ikuta, K.; Kishihara, K.; Hara, H.; Nakae, S.; Iwakura, Y.; Matsuzaki, G. Essential Role of IL-17A in the Formation of a Mycobacterial Infection-Induced Granuloma in the Lung. J. Immunol. 2010, 184, 4414–4422. [Google Scholar] [CrossRef]
- Peng, M.Y.; Wang, Z.H.; Yao, C.Y.; Jiang, L.N.; Jin, Q.L.; Wang, J.; Li, B.Q. Interleukin 17-Producing Gamma Delta T Cells Increased in Patients with Active Pulmonary Tuberculosis. Cell. Mol. Immunol. 2008, 5, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, E.; Biswas, P.; Vettoretto, K.; Ferrarini, M.; Uguccioni, M.; Piali, L.; Leone, B.E.; Moser, B.; Rugarli, C.; Pardi, R. Macrophages Exposed to Mycobacterium tuberculosis Release Chemokines Able to Recruit Selected Leucocyte Subpopulations: Focus on Gammadelta Cells. Immunology 2003, 108, 365–374. [Google Scholar] [CrossRef]
- Shen, H.; Wang, Y.; Chen, C.Y.; Frencher, J.; Huang, D.; Yang, E.; Ryan-Payseur, B.; Chen, Z.W. Th17-Related Cytokines Contribute to Recall-like Expansion/Effector Function of HMBPP-Specific Vγ2Vδ2 T Cells after Mycobacterium tuberculosis Infection or Vaccination. Eur. J. Immunol. 2015, 45, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Gu, J.; Xiao, H.; Liang, S.; Yang, E.; Yang, R.; Huang, D.; Chen, C.; Wang, F.; Shen, L.; et al. Selective Destruction of Interleukin 23-Induced Expansion of a Major Antigen-Specific γδ T-Cell Subset in Patients With Tuberculosis. J. Infect. Dis. 2017, 215, 420–430. [Google Scholar] [CrossRef]
- El Daker, S.; Sacchi, A.; Montesano, C.; Altieri, A.M.; Galluccio, G.; Colizzi, V.; Martini, F.; Martino, A. An Abnormal Phenotype of Lung Vγ9Vδ2 T Cells Impairs Their Responsiveness in Tuberculosis Patients. Cell. Immunol. 2013, 282, 106–112. [Google Scholar] [CrossRef]
- Saura-Esteller, J.; de Jong, M.; King, L.A.; Ensing, E.; Winograd, B.; de Gruijl, T.D.; Parren, P.W.H.I.; van der Vliet, H.J. Gamma Delta T-Cell Based Cancer Immunotherapy: Past-Present-Future. Front. Immunol. 2022, 13, 915837. [Google Scholar] [CrossRef]
- Kobayashi, H.; Tanaka, Y.; Shimmura, H.; Minato, N.; Tanabe, K. Complete Remission of Lung Metastasis Following Adoptive Immunotherapy Using Activated Autologous Gammadelta T-Cells in a Patient with Renal Cell Carcinoma. Anticancer Res. 2010, 30, 575–579. [Google Scholar] [PubMed]
- Ishihara, M.; Miwa, H.; Fujiwara, H.; Akahori, Y.; Kato, T.; Tanaka, Y.; Tawara, I.; Shiku, H. Aβ-T Cell Receptor Transduction Gives Superior Mitochondrial Function to γδ -T Cells with Promising Persistence. iScience 2023, 26, 107802. [Google Scholar] [CrossRef] [PubMed]
- Lisci, M.; Barton, P.R.; Randzavola, L.O.; Ma, C.Y.; Marchingo, J.M.; Cantrell, D.A.; Paupe, V.; Prudent, J.; Stinchcombe, J.C.; Griffiths, G.M. Mitochondrial Translation Is Required for Sustained Killing by Cytotoxic T Cells. Science 2021, 374, eabe9977. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yang, E.; Guo, M.; Yang, R.; Huang, G.; Peng, Y.; Sha, W.; Wang, F.; Shen, L. Adjunctive Zoledronate + IL-2 Administrations Enhance Anti-Tuberculosis Vγ2Vδ2 T-Effector Populations, and Improve Treatment Outcome of Multidrug-Resistant Tuberculosis1. Emerg. Microbes Infect. 2022, 11, 1790–1805. [Google Scholar] [CrossRef] [PubMed]
- Qaqish, A.; Huang, D.; Chen, C.Y.; Zhang, Z.; Wang, R.; Li, S.; Yang, E.; Lu, Y.; Larsen, M.H.; Jacobs, W.R.; et al. Adoptive Transfer of Phosphoantigen-Specific Γδ T Cell Subset Attenuates Mycobacterium tuberculosis Infection in Nonhuman Primates. J. Immunol. 2017, 198, 4753–4763. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Frencher, J.; Huang, D.; Wang, W.; Yang, E.; Chen, C.Y.; Zhang, Z.; Wang, R.; Qaqish, A.; Larsen, M.H.; et al. Immunization of Vγ2Vδ2 T Cells Programs Sustained Effector Memory Responses That Control Tuberculosis in Nonhuman Primates. Proc. Natl. Acad. Sci. USA 2019, 116, 6371–6378. [Google Scholar] [CrossRef]
- Roelofs, A.J.; Jauhiainen, M.; Mönkkönen, H.; Rogers, M.J.; Mönkkönen, J.; Thompson, K. Peripheral Blood Monocytes Are Responsible for Gammadelta T Cell Activation Induced by Zoledronic Acid through Accumulation of IPP/DMAPP. Br. J. Haematol. 2009, 144, 245–250. [Google Scholar] [CrossRef]
- Raffray, L.; Burton, R.J.; Baker, S.E.; Morgan, M.P.; Eberl, M. Zoledronate Rescues Immunosuppressed Monocytes in Sepsis Patients. Immunology 2020, 159, 88–95. [Google Scholar] [CrossRef]
- Zacharin, M.; O’Sullivan, M. Intravenous Pamidronate Treatment of Polyostotic Fibrous Dysplasia Associated with the McCune Albright Syndrome. J. Pediatr. 2000, 137, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kamath, A.; Das, H.; Li, L.; Bukowski, J.F. Antibacterial Effect of Human V Gamma 2V Delta 2 T Cells in Vivo. J. Clin. Investig. 2001, 108, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Immune metabolism of γδ T cells. (A) CD27+ γδ T cells are ligand-experienced and splenic resident. They undergo glycolysis after thymic development and release IFNγ. CD27− γδ T cells, however, are ligand naive and lymph node resident. They undergo oxidative phosphorylation, governed by c-Maf regulation of rate-limiting enzyme isocitrate dehydrogenase 2 (IDH2) and Th17 cytokine-producing genes Rorc, Il17a, and Blk. (B) In inflammatory psoriasis, basal keratinocytes produce oxysterols through the enzyme cholesterol-25-hydroxylase (CH25H). The oxysterols are ligands for the G-protein-coupled receptor GPR183 on γδ T cells. Subsequent lipid metabolism and signaling leads to IL-17 release and inflammation. In a mouse model, dietary cholesterol contributes to this signaling cascade, worsening psoriasis. (C) During psoriasis, γδ T cells undergo glutaminolysis, promoting subsequent acetylation of the Il17a promoter and releasing IL-17, increasing inflammation. CD69 and LAT1 expression on γδ T cells promote tryptophan uptake and metabolism in psoriasis. The resulting Ahr signaling cascade leads to IL-22 release and increased inflammation.
Figure 1.
Immune metabolism of γδ T cells. (A) CD27+ γδ T cells are ligand-experienced and splenic resident. They undergo glycolysis after thymic development and release IFNγ. CD27− γδ T cells, however, are ligand naive and lymph node resident. They undergo oxidative phosphorylation, governed by c-Maf regulation of rate-limiting enzyme isocitrate dehydrogenase 2 (IDH2) and Th17 cytokine-producing genes Rorc, Il17a, and Blk. (B) In inflammatory psoriasis, basal keratinocytes produce oxysterols through the enzyme cholesterol-25-hydroxylase (CH25H). The oxysterols are ligands for the G-protein-coupled receptor GPR183 on γδ T cells. Subsequent lipid metabolism and signaling leads to IL-17 release and inflammation. In a mouse model, dietary cholesterol contributes to this signaling cascade, worsening psoriasis. (C) During psoriasis, γδ T cells undergo glutaminolysis, promoting subsequent acetylation of the Il17a promoter and releasing IL-17, increasing inflammation. CD69 and LAT1 expression on γδ T cells promote tryptophan uptake and metabolism in psoriasis. The resulting Ahr signaling cascade leads to IL-22 release and increased inflammation.

Figure 2.
Role of γδ T cells during cutaneous S. aureus infection. Intradermal S. aureus infection and subsequent IL-1β signaling stimulate cutaneous γδ T cells to produce proinflammatory cytokines IFNy, TNFα, IL-22, and IL-17. These cytokines contribute to neutrophil recruitment, production of antimicrobial peptides, skin inflammation, and eventual bacterial clearance. Commensal S. epidermidis, skin colonization, stimulates γδ T cells to produce perforin-2, a pore-forming cytolysin that helps clear S. aureus infection.
Figure 2.
Role of γδ T cells during cutaneous S. aureus infection. Intradermal S. aureus infection and subsequent IL-1β signaling stimulate cutaneous γδ T cells to produce proinflammatory cytokines IFNy, TNFα, IL-22, and IL-17. These cytokines contribute to neutrophil recruitment, production of antimicrobial peptides, skin inflammation, and eventual bacterial clearance. Commensal S. epidermidis, skin colonization, stimulates γδ T cells to produce perforin-2, a pore-forming cytolysin that helps clear S. aureus infection.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nanda, N.; Alphonse, M.P. From Host Defense to Metabolic Signatures: Unveiling the Role of γδ T Cells in Bacterial Infections. Biomolecules 2024, 14, 225. https://doi.org/10.3390/biom14020225
AMA Style
Nanda N, Alphonse MP. From Host Defense to Metabolic Signatures: Unveiling the Role of γδ T Cells in Bacterial Infections. Biomolecules. 2024; 14(2):225. https://doi.org/10.3390/biom14020225
Chicago/Turabian StyleNanda, Namya, and Martin P. Alphonse. 2024. "From Host Defense to Metabolic Signatures: Unveiling the Role of γδ T Cells in Bacterial Infections" Biomolecules 14, no. 2: 225. https://doi.org/10.3390/biom14020225
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.