
Conditional Seed Dormancy Helps Silene hicesiae Brullo & Signor. Overcome Stressful Mediterranean Summer Conditions

 ,
,
Abstract
:1. Introduction
2. Results
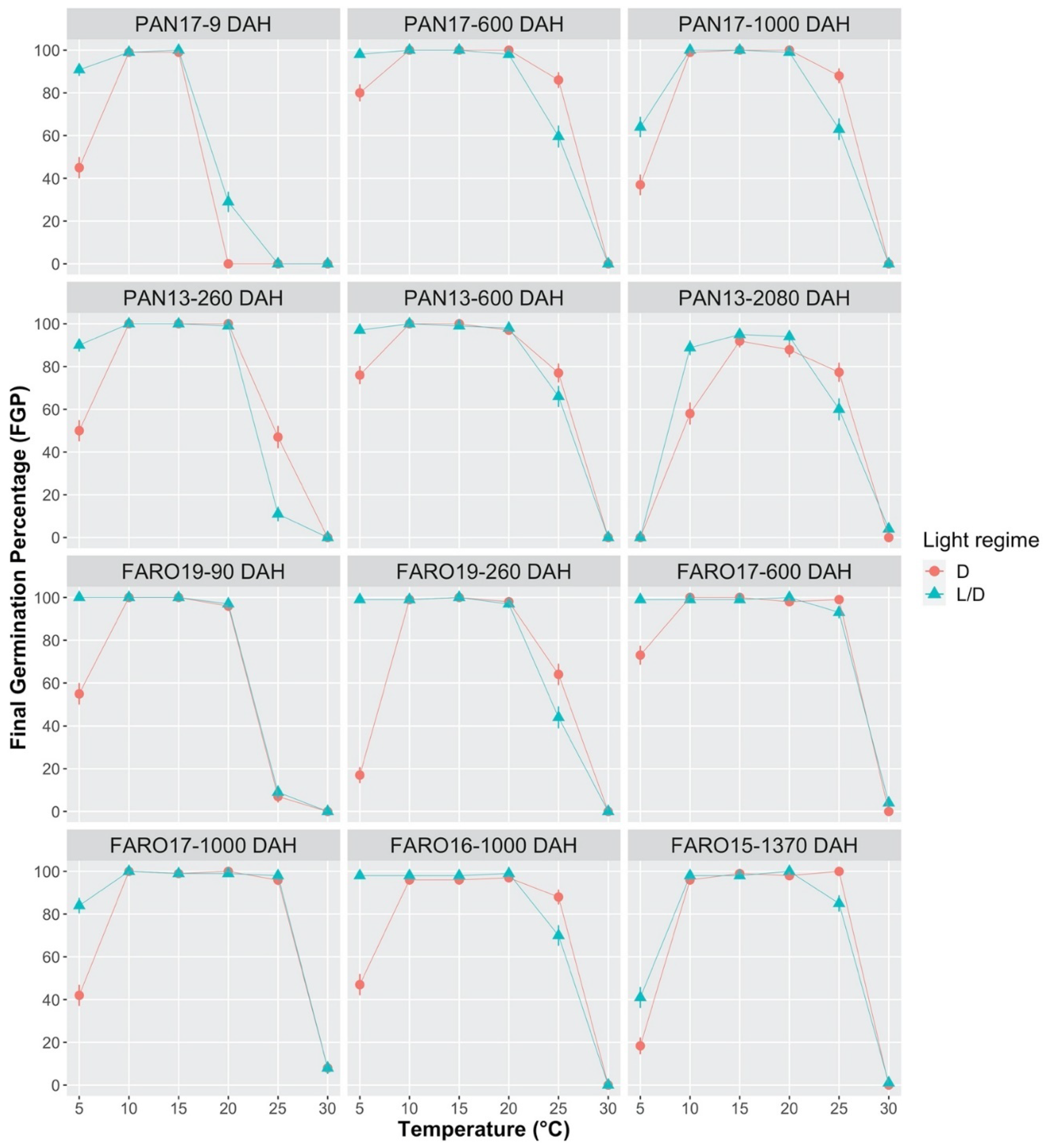
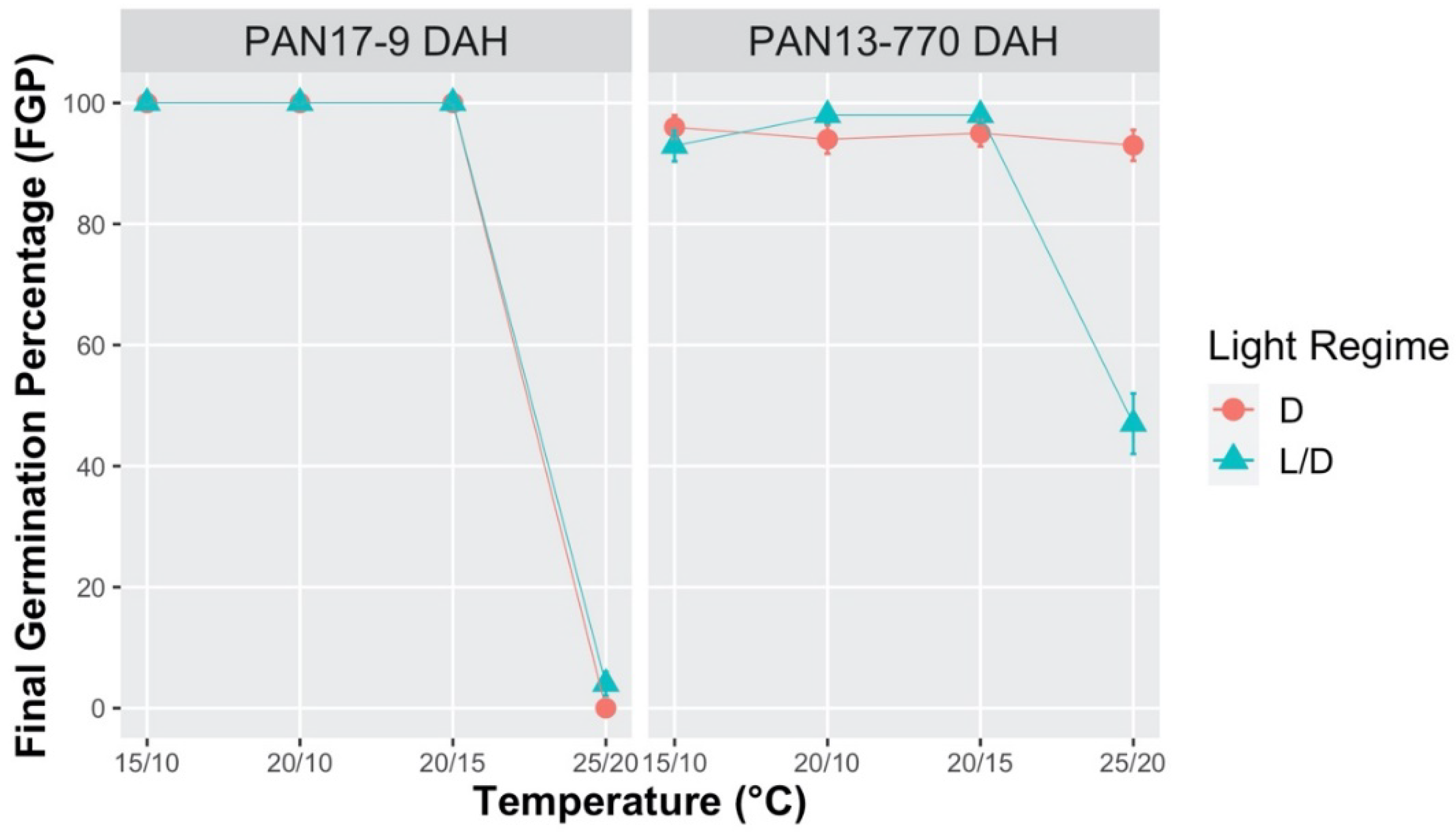
2.1. Effect of Temperature and Light on Germination Process
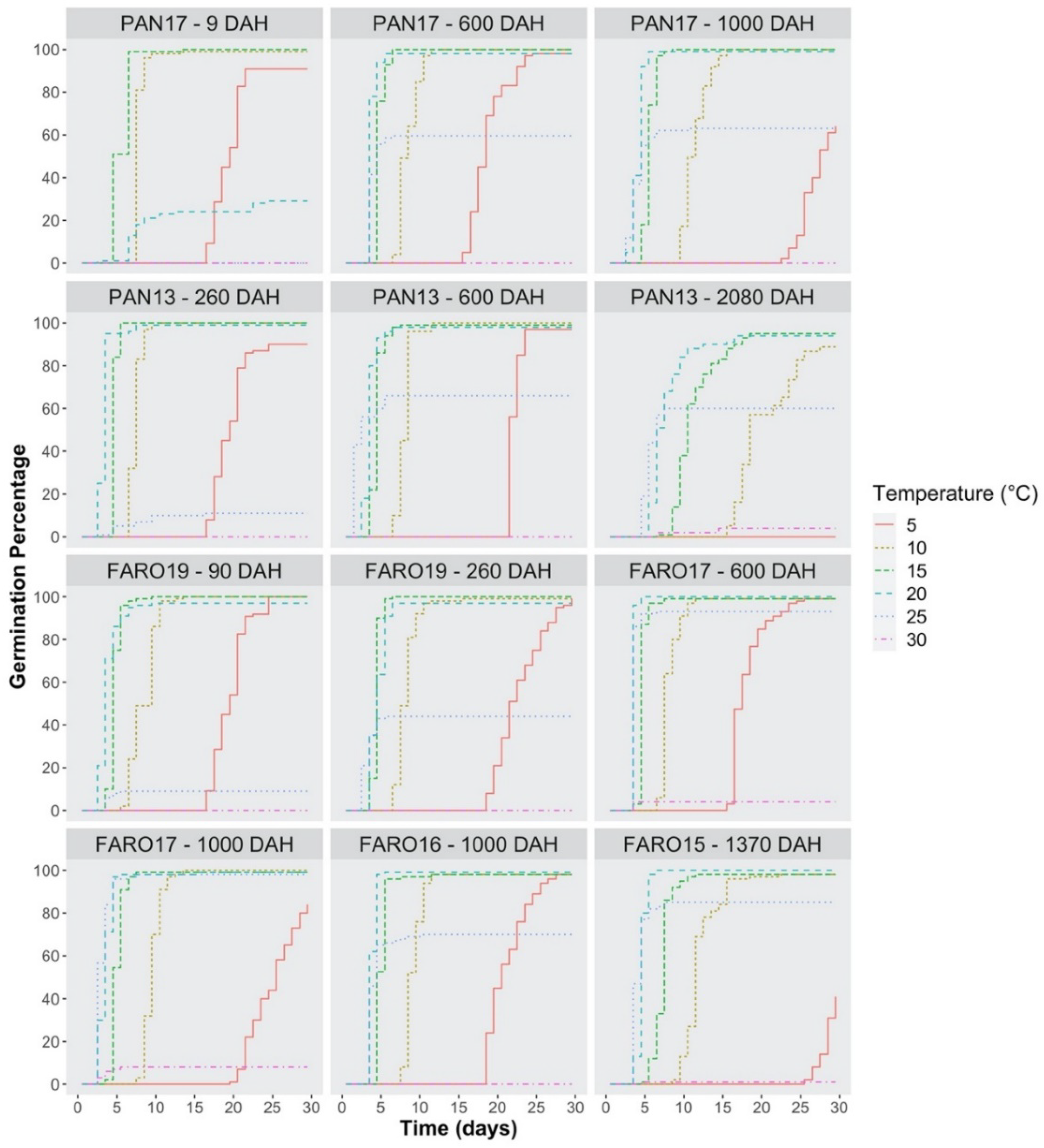
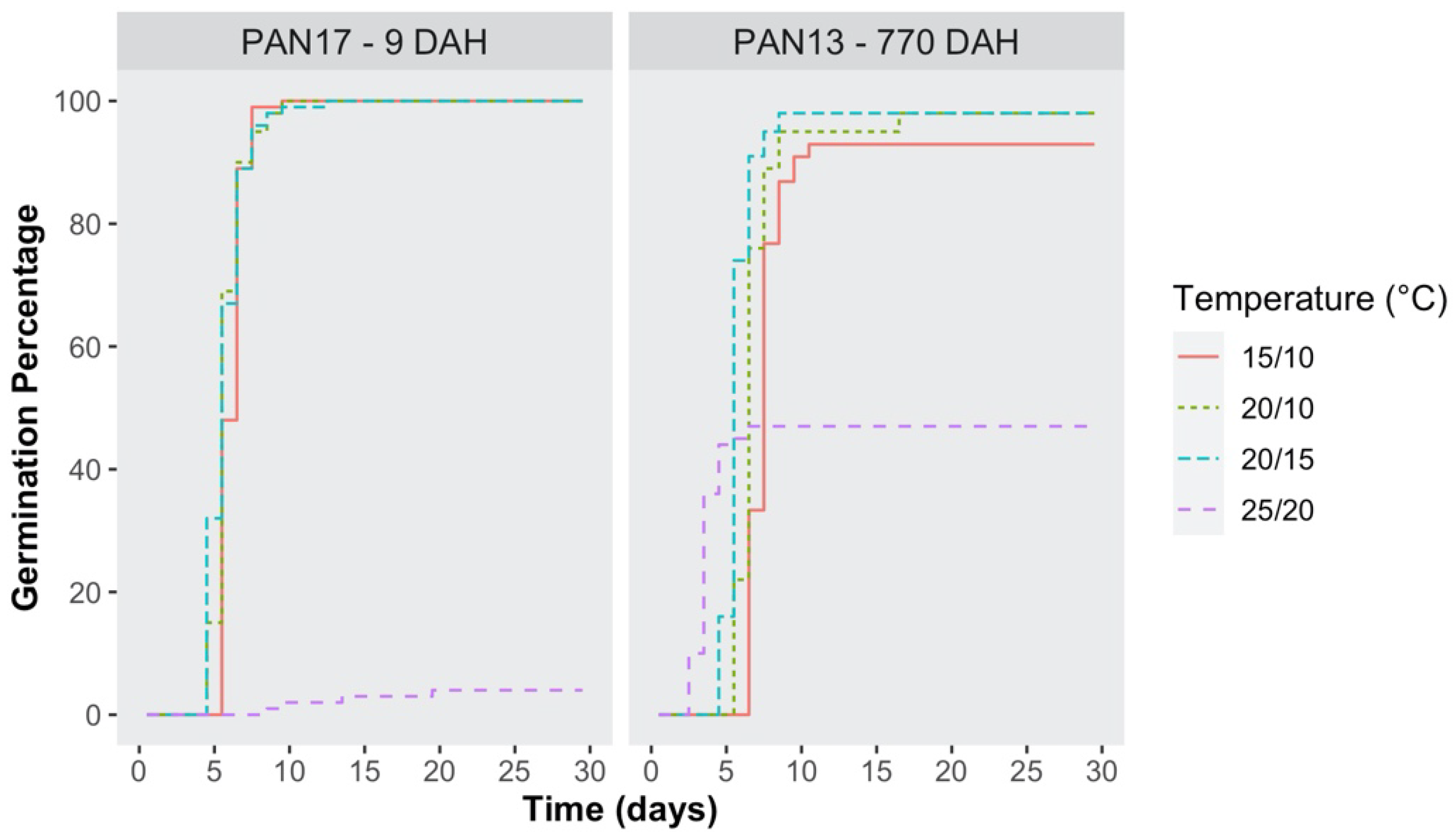
2.2. Germination Time-Course
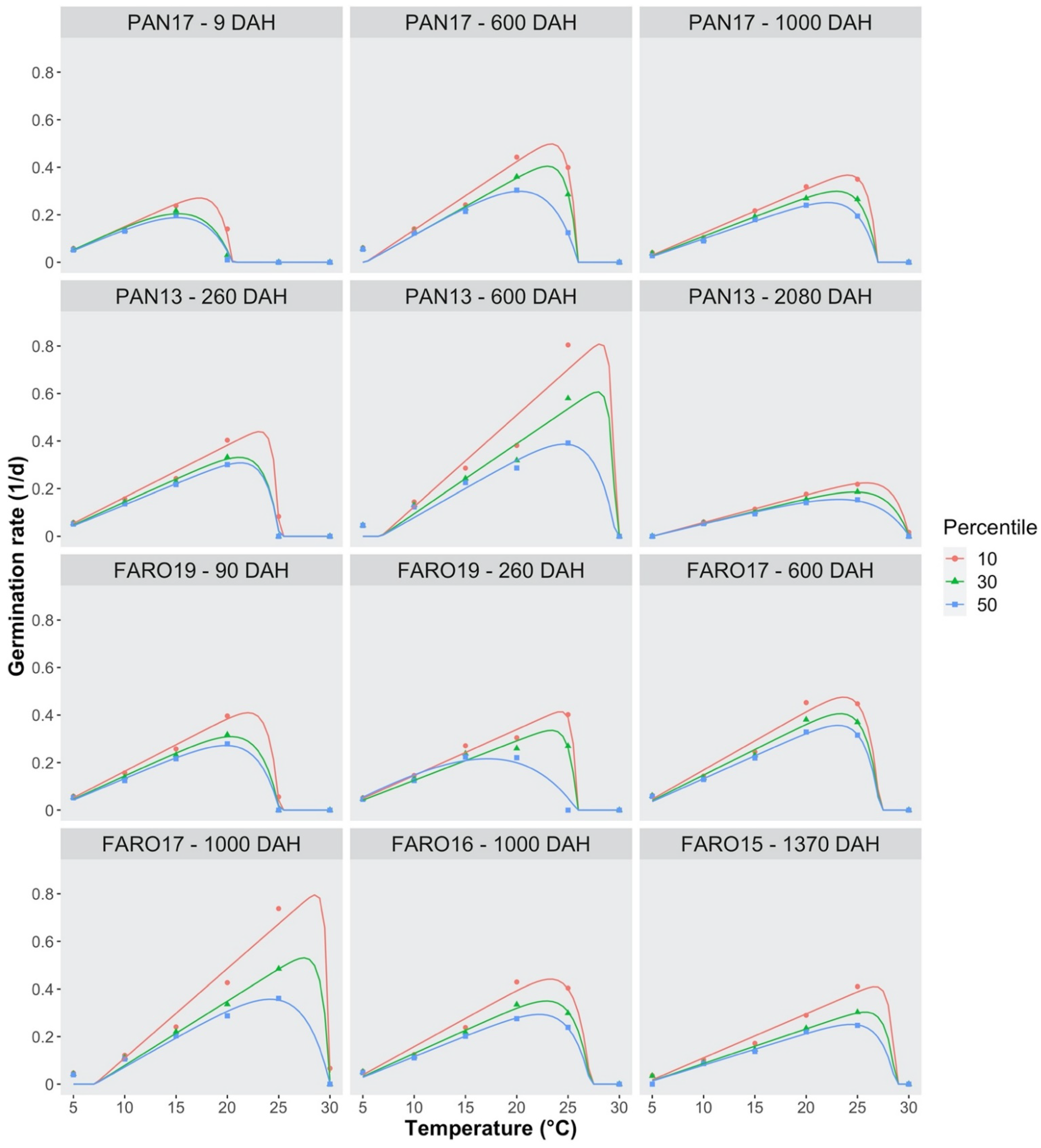
2.3. Germination Rate, Threshold Temperatures and Thermal Requirements
3. Discussion
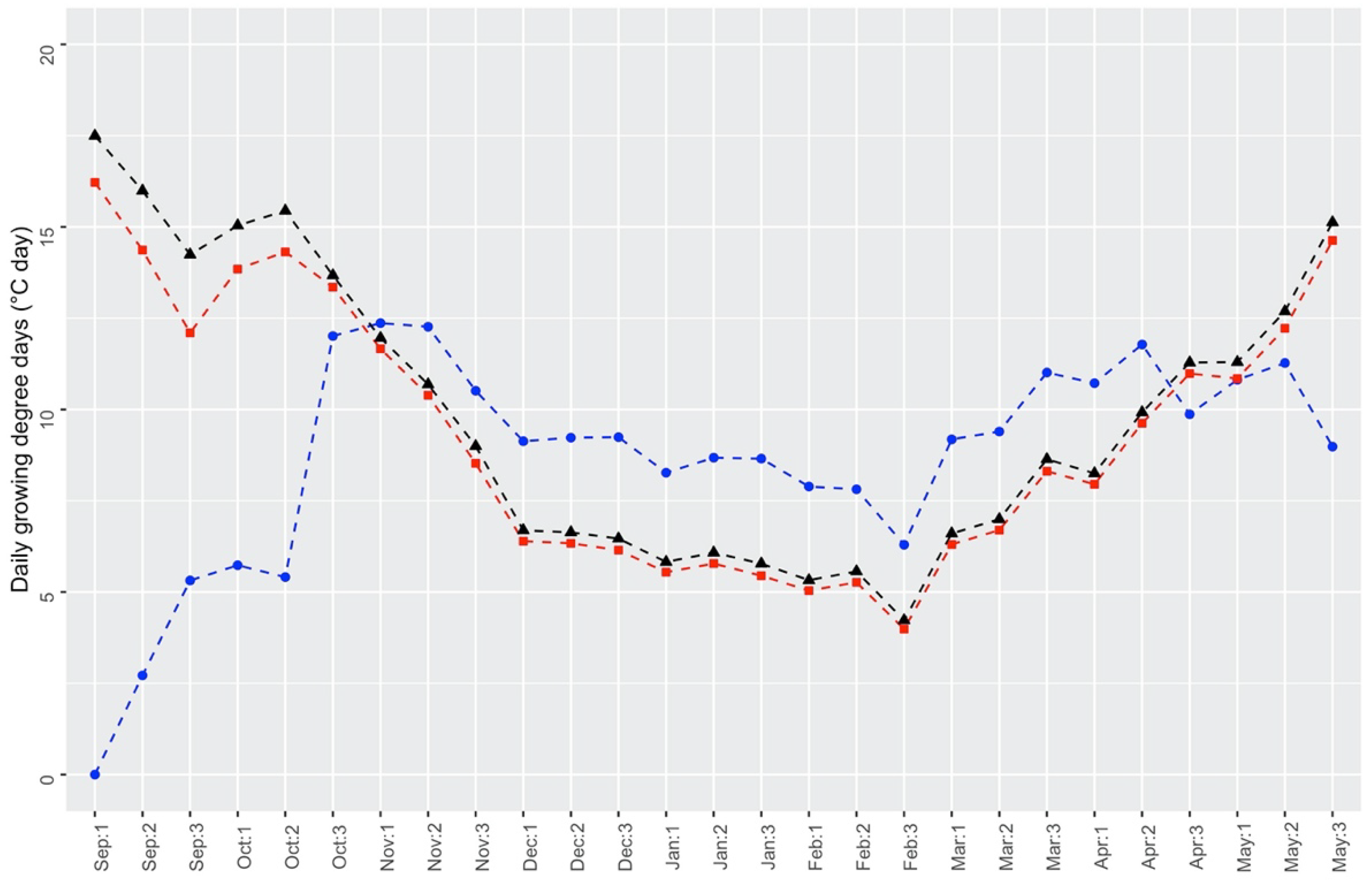
4. Materials and Methods
4.1. Seed Lot Details
4.2. Seed Germination Behavior Assessment
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganatsas, P.; Tsakaldimi, M.; Damianidis, C.; Stefanaki, A.; Kalapothareas, T.; Karydopoulos, T.; Papapavlou, K. Regeneration ecology of the rare plant species Verbascum dingleri: Implications for species conservation. Sustainability 2019, 11, 3305. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, M.; Dayrell, R.L.C.; Wardil, M.V.; Damasceno, A.; Fernandes, T.; Castilho, A.; Santos, F.M.G.; Silveira, F.A.O. Seed functional traits provide support for ecological restoration and ex situ conservation in the threatened Amazon ironstone outcrop flora. Front. Plant Sci. 2020, 11, 599496. [Google Scholar] [CrossRef]
- Walter, G.M.; Catara, S.; Bridle, J.R.; Cristaudo, A. Population variation in early development can determine ecological resilience in response to environmental change. New Phytol. 2020, 226, 1312–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schemske, D.W.; Husband, B.C.; Ruckelshaus, M.H.; Goodwillie, C.; Parker, I.M.; Bishop, J.G. Evaluating approaches to the conservation of rare and endangered plants. Ecology 1994, 75, 584–606. [Google Scholar] [CrossRef] [Green Version]
- Cristaudo, A.; Catara, S.; Mingo, A.; Restuccia, A.; Onofri, A. Temperature and storage time strongly affect the germination success of perennial Euphorbia species in Mediterranean regions. Ecol. Evol. 2019, 9, 10984–10999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, J.L. Population Biology of Plants; Academic Press: New York, NY, USA, 1977; ISBN 0123258529. [Google Scholar]
- Probert, R.J. The role of temperature in the regulation of seed dormancy and germination. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 261–292. ISBN 0851994326. [Google Scholar]
- Gresta, F.; Cristaudo, A.; Onofri, A.; Restuccia, A.; Avola, G. Germination response of four pasture species to temperature, light, and post-harvest period. Plant Biosyst. 2010, 144, 849–856. [Google Scholar] [CrossRef]
- Thanos, C.A.; Georghiou, K.; Skarou, F. Glaucium flavum seed germination: An ecophysiological approach. Ann. Bot. 1989, 63, 121–130. [Google Scholar] [CrossRef]
- Thanos, C.A.; Kadis, C.C.; Skarou, F. Ecophysiology of germination in the aromatic plants thyme, savory and oregano (Labiatae). Seed Sci. Res. 1995, 5, 161–170. [Google Scholar] [CrossRef]
- Tlig, T.; Gorai, M.; Neffati, M. Germination responses of Diplotaxis harra to temperature and salinity. Flora 2008, 203, 421–428. [Google Scholar] [CrossRef]
- Bradford, K.J. Threshold models applied to seed germination ecology. New Phytol. 2005, 165, 338–341. [Google Scholar] [CrossRef]
- Wang, W.Q.; Cheng, H.Y.; Song, S.Q. Development of a threshold model to predict germination of Populus tomentosa seeds after harvest and storage under ambient condition. PLoS ONE 2013, 8, e62868. [Google Scholar] [CrossRef] [Green Version]
- Cristaudo, A.; Gresta, F.; Catara, S.; Mingo, A. Assessment of daily heat pulse regimes on the germination of six Amaranthus species. Weed Res. 2014, 54, 366–376. [Google Scholar] [CrossRef]
- Thanos, C.A.; Georghiou, K.; Douma, D.J.; Marangaki, C.J. Photoinhibition of seed germination in Mediterranean maritime plants. Ann. Bot. 1991, 68, 469–475. [Google Scholar] [CrossRef]
- Thanos, C.A.; Georghiou, K.; Delipetrou, P. Photoinhibition of seed germination in the maritime plant Matthiola tricuspidata. Ann. Bot. 1994, 73, 639–644. [Google Scholar] [CrossRef]
- Catara, S.; Cristaudo, A.; Gualtieri, A.; Galesi, R.; Impelluso, C.; Onofri, A. Threshold temperatures for seed germination in nine species of Verbascum (Scrophulariaceae). Seed Sci. Res. 2016, 26, 30–46. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier Academic Press: London, UK, 2014; ISBN 9780124166776. [Google Scholar]
- Cristaudo, A.; Gresta, F.; Restuccia, A.; Catara, S.; Onofri, A. Germinative response of redroot pigweed (Amaranthus retroflexus L.) to environmental conditions: Is there a seasonal pattern? Plant Biosyst. 2016, 150, 583–591. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, C.C.; Baskin, J.M. A graphical method for identifying the six types of non-deep physiological dormancy in seeds. Plant Biol. 2017, 19, 673–682. [Google Scholar] [CrossRef]
- Andersson, L.; Milberg, P. Variation in seed dormancy among mother plants, populations and years of seed collection. Seed Sci. Res. 1998, 8, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Gutterman, Y. Maternal effects on seeds during development. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 59–84. ISBN 0851994326. [Google Scholar]
- Donohue, K. Completing the cycle: Maternal effects as the missing link in plant life histories. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1059–1074. [Google Scholar] [CrossRef] [Green Version]
- Godefroid, S.; Le Pajolec, S.; Van Rossum, F. Pre-translocation considerations in rare plant reintroductions: Implications for designing protocols. Plant Ecol. 2016, 217, 169–182. [Google Scholar] [CrossRef]
- Iralu, V.; Barbhuyan, H.S.A.; Upadhaya, K. Ecology of seed germination in threatened trees: A review. Energy Ecol. Environ. 2019, 4, 189–210. [Google Scholar] [CrossRef]
- Saatkamp, A.; Cochrane, A.; Commander, L.; Guja, L.; Jimenez-Alfaro, B.; Larson, J.; Nicotra, A.; Poschlod, P.; Silveira, F.A.O.; Cross, A.; et al. A research agenda for seed-trait functional ecology. New Phytol. 2019, 221, 1764–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basey, A.C.; Fant, J.B.; Kramer, A.T. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Nativ. Plants J. 2015, 16, 37–52. [Google Scholar] [CrossRef]
- Volis, S. Conservation-oriented restoration—A two for one method to restore both threatened species and their habitats. Plant Divers. 2019, 41, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Volis, S. Complementarities of two intermediate conservation approaches. Plant Divers. 2017, 39, 379–382. [Google Scholar] [CrossRef]
- Ensslin, A.; Godefroid, S. How the cultivation of wild plants in botanic gardens can change their genetic and phenotypic status and what this means for their conservation value. Sibbaldia 2019, 17, 51–70. [Google Scholar] [CrossRef]
- Pasta, S.; Perez-Graber, A.; Fazan, L.; de Montmollin, B. (Eds.) The Top 50 Mediterranean Island Plants UPDATE 2017; IUCN/SSC/Mediterranean Plant Specialist Group: Neuchâtel, Switzerland, 2017; 141p, Available online: https://top50.iucn-mpsg.org/book (accessed on 20 April 2021).
- Murru, V.; Santo, A.; Piazza, C.; Hugot, L.; Bacchetta, G. Seed germination, salt-stress tolerance, and the effect of nitrate on three Tyrrhenian coastal species of the Silene mollissima aggregate (Caryophyllaceae). Botany 2015, 93, 881–892. [Google Scholar] [CrossRef]
- Murru, V.; Santo, A.; Gallo, M.; Cardona, C.; Boi, M.; Bacchetta, G. Comparative germination ecology and seedling growth of two Ibero-Levantine endemic species belonging to the Silene mollissima aggregate (Caryophyllaceae). Flora 2017, 227, 10–17. [Google Scholar] [CrossRef]
- Brullo, S.; Signorello, P. Silene hicesiae, a new species from Aeolian Islands. Willdenowia 1984, 14, 141–144. [Google Scholar] [CrossRef]
- Jeanmonod, D. Révision de la section Siphonomorpha Otth du genre Silene L. (Caryophyllaceae) en Méditerranée occidentale II: Le groupe de S. mollissima. Candollea 1984, 39, 195–259. [Google Scholar]
- Murru, V.; Grillo, O.; Santo, A.; Ucchesu, M.; Piazza, C.; Gaio, A.; Carta, A.; Bacchetta, G. Seed morpho-colorimetric analysis on some Tyrrhenian species of the Silene mollissima aggregate (Caryophyllaceae). Flora 2019, 258, 151445. [Google Scholar] [CrossRef]
- Royal Botanic Gardens Kew. Seed Information Database (SID). Version 7.1 (May 2008). Available online: https://data.kew.org/sid/SidServlet?ID=46635&Num=813#Germination (accessed on 19 March 2021).
- Aldhous, J.R. Nursery Practice; Forestry Commission: London, UK, 1972; Volume 43, 184p.
- Schütz, W.; Milberg, P.; Lamont, B.B. Seed dormancy, after-ripening and light requirements of four annual Asteraceae in south-western Australia. Ann. Bot. 2002, 90, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Baskin, J.M.; Baskin, C.C. Germination responses of buried seeds of Capsella bursa-pastoris exposed to seasonal temperature changes. Weed Res. 1989, 29, 205–212. [Google Scholar] [CrossRef]
- Thompson, P.A. Germination of species of Caryophyllaceae in relation to their geographical distribution in Europe. Ann. Bot. 1970, 34, 427–449. [Google Scholar] [CrossRef]
- Doussi, M.A.; Thanos, C.A. Ecophysiology of seed germination in Mediterranean geophytes. 1. Muscari spp. Seed Sci. Res. 2002, 12, 193–201. [Google Scholar] [CrossRef]
- Perez-Garcia, F.; Hornero, J.; Gonzalez-Benito, M.E. Interpopulation variation in seed germination of five Mediterranean Labiatae shrubby species. Isr. J. Plant Sci. 2003, 51, 117–124. [Google Scholar] [CrossRef]
- Zani, D.; Müller, J.V. Climatic control of germination in the genus Silene L. Plant Ecol. Divers. 2017, 10, 431–442. [Google Scholar] [CrossRef]
- Picciau, R.; Pritchard, H.; Mattana, E.; Bacchetta, G. Thermal thresholds for seed germination in Mediterranean species are higher in mountain compared with lowland areas. Seed Sci. Res. 2019, 29, 44–54. [Google Scholar] [CrossRef]
- Cogoni, D.; Mattana, E.; Fenu, G.; Bacchetta, G. From seed to seedling: A critical transitional stage for the Mediterranean psammophilous species Dianthus morisianus (Caryophyllaceae). Plant Biosyst. 2012, 146, 910–917. [Google Scholar] [CrossRef]
- De Vitis, M.; Seal, C.E.; Ulian, T.; Pritchard, H.W.; Magrini, S.; Fabrini, G.; Mattana, E. Rapid adaptation of seed germination requirements of the threatened Mediterranean species Malcolmia littorea (Brassicaceae) and implications for its reintroduction. S. Afr. J. Bot. 2014, 94, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Copete, M.Á.; Herranz, J.M.; Ferrandis, P. Seed germination ecology of the endemic Iberian winter annuals Iberis pectinata and Ziziphora aragonensis. Seed Sci. Res. 2009, 19, 155–169. [Google Scholar] [CrossRef]
- Carta, A.; Hanson, S.; Müller, J.V. Plant regeneration from seeds responds to phylogenetic relatedness and local adaptation in Mediterranean Romulea (Iridaceae) species. Ecol. Evol. 2016, 6, 4166–4178. [Google Scholar] [CrossRef]
- Visser, M.; Beaugendre, A. Conditional dormancy of Stipa lagascae (Poaceae) bulk-harvested on seed increase plots in South Tunisia: A reassessment and a surprise. Plant Ecol. Evol. 2019, 152, 450–459. [Google Scholar] [CrossRef]
- Walck, J.; Baskin, J.; Baskin, C.; Hidayati, S. Defining transient and persistent seed banks in species with pronounced seasonal dormancy and germination patterns. Seed Sci. Res. 2005, 15, 189–196. [Google Scholar] [CrossRef]
- Ozinga, W.A.; Römermann, C.; Bekker, R.M.; Prinzing, A.; Tamis, W.L.M.; Schaminée, J.H.J.; Hennekens, S.M.; Thompson, K.; Poschlod, P.; Kleyer, M.; et al. Dispersal failure contributes to plant losses in NW Europe. Ecol. Lett. 2009, 12, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Mattana, E.; Daws, M.I.; Bacchetta, G. Comparative germination ecology of the endemic Centranthus amazonum (Valerianaceae) and its widespread congener Centranthus ruber. Plant Spec. Biol. 2010, 25, 165–172. [Google Scholar] [CrossRef]
- Peng, D.; Sun, L.; Pritchard, H.W.; Yang, J.; Sun, H.; Li, Z. Species distribution modelling and seed germination of four threatened snow lotus (Saussurea), and their implication for conservation. Glob. Ecol. Conserv. 2019, 17, e00565. [Google Scholar] [CrossRef]
- Meyer, S.E.; Kitchen, S.G. Life-history variation in Blue Flax (Linum perenne, Linaceae)—Seed-germination phenology. Am. J. Bot. 1994, 81, 528–535. [Google Scholar] [CrossRef]
- Rojas-Aréchiga, M.; Casas, A.; Vázquez-Yanes, C. Seed germination of wild and cultivated Stenocereus stellatus (Cactaceae) from the Tehuacán-Cuicatlán Valley, Central México. J. Arid Environ. 2001, 49, 279–287. [Google Scholar] [CrossRef]
- Wilson, S.B.; Mecca, L.K. Seed production and germination of eight cultivars and the wild type of Ruellia tweediana: A potentially invasive ornamental. J. Environ. Hortic. 2003, 21, 137–143. [Google Scholar] [CrossRef]
- Maass, B.L. Changes in seed morphology, dormancy and germination from wild to cultivated hyacinth bean germplasm (Lablab purpureus: Papilionoideae). Genet. Resour. Crop Evol. 2006, 53, 1127–1135. [Google Scholar] [CrossRef]
- Wang, M.; Li, W.; Fang, C.; Xu, F.; Liu, Y.; Wang, Z.; Yang, R.; Zhang, M.; Liu, S.; Lu, S.; et al. Parallel selection on a dormancy gene during domestication of crops from multiple families. Nat. Genet. 2018, 50, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Ensslin, A.; Sandner, T.M.; Matthies, D. Consequences of ex situ cultivation of plants: Genetic diversity, fitness and adaptation of the monocarpic Cynoglossum officinale L. in botanic gardens. Biol. Conserv. 2011, 144, 272–278. [Google Scholar] [CrossRef]
- Schröder, R.; Prasse, R. Cultivation and hybridization alter the germination behavior of native plants used in revegetation and restoration. Restor. Ecol. 2013, 21, 793–800. [Google Scholar] [CrossRef]
- Ensslin, A.; Van de Vyver, A.; Vanderborght, T.; Godefroid, S. Ex situ cultivation entails high risk of seed dormancy loss on short-lived wild plant species. J. Appl. Ecol. 2018, 55, 1145–1154. [Google Scholar] [CrossRef]
- Therneau, T.M. A Package for Survival Analysis in R. R Package Version 3.1-12. 2020. Available online: https://CRAN.R-project.org/package=survival (accessed on 11 November 2020).
- Onofri, A.; Gresta, F.; Tei, F. A new method for the analysis of germination and emergence data of weed species. Weed Res. 2010, 50, 187–198. [Google Scholar] [CrossRef]
- Gresta, F.; Avola, G.; Onofri, A.; Anastasi, U.; Cristaudo, A. When Does Hard Coat Impose Dormancy in Legume Seeds? Lotus and Scorpiurus Case Study. Crop Sci. 2011, 51, 1739–1747. [Google Scholar] [CrossRef]
- Bradford, K.J. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Rete Stazioni Meteo Linea Meteo 2020. Available online: http://www.lineameteo.it/stazioni.php?id=678 (accessed on 24 March 2020).
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.0.2; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 5 February 2021).
- Pizza, R.; Espeland, E.; Etterson, J. Eight generations of native seed cultivation reduces plant fitness relative to the wild progenitor population. Evol. Appl. 2021, 14, 1816–1829. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seed Lot ID | Seed Age DAH | Parameters | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Tb (°C) | Tc (°C) | To (°C) | θT (°Cd) | ||||||
| 10th | 30th | 50th | 10th | 30th | 50th | ||||
| PAN17 | 9 | 2.35 (0.79) | 20.52 (0.17) | 17.3 (0.91) | 15.2 (1.44) | 15.2 (1.56) | 50.92 (5.24) | 50.32 (9.67) | 54.02 (12.2) |
| PAN17 | 600 | 5.27 (0.67) | 25.98 (0.38) | 23.4 (1.53) | 23.0 (1.82) | 20.3 (1.33) | 34.74 (2.33) | 41.46 (3.12) | 41.23 (5.62) |
| PAN17 | 1000 | 3.36 (0.70) | 26.99 (3.11) | 24.1 (9.30) | 23.0 (7.97) | 22.1 (7.77) | 54.10 (4.73) | 60.76 (5.03) | 67.03 (10.5) |
| PAN13 | 260 | 2.50 (0.58) | 25.08 (0.04) | 23.2 (2.57) | 21.2 (1.97) | 21.3 (4.93) | 45.77 (2.61) | 52.04 (3.99) | 56.66 (5.31) |
| PAN13 | 600 | 6.77 (0.36) | 29.67 (0.18) | 28.2 (9.97) | 27.9 (8.58) | 24.6 (2.53) | 25.99 (0.86) | 34.02 (1.23) | 40.87 (3.84) |
| PAN13 | 2080 | 4.97 (0.85) | 30.06 (0.09) | 25.9 (7.66) | 24.7 (2.23) | 23.2 (1.61) | 86.66 (12.3) | 94.92 (12.7) | 96.52 (16.0) |
| FARO19 | 90 | 2.66 (0.45) | 25.09 (0.03) | 21.8 (0.91) | 20.3 (0.77) | 19.9 (0.70) | 44.89 (2.08) | 51.03 (3.49) | 55.07 (4.32) |
| FARO19 | 260 | 2.48 (0.49) | 25.90 (0.21) | 24.3 (0.45) | 23.4 (0.64) | 17.2 (0.88) | 51.64 (2.36) | 60.10 (3.11) | 45.40 (5.84) |
| FARO17 | 600 | 3.13 (0.37) | 27.20 (0.80) | 23.7 (1.50) | 23.4 (1.59) | 23.1 (1.76) | 40.71 (1.63) | 46.56 (2.04) | 51.94 (2.50) |
| FARO17 | 1000 | 7.08 (0.30) | 30.02 (0.21) | 28.6 (0.34) | 27.5 (1.98) | 24.2 (0.67) | 26.61 (0.75) | 37.03 (1.18) | 40.49 (2.68) |
| FARO16 | 1000 | 3.36 (0.40) | 27.19 (0.74) | 23.3 (1.26) | 22.9 (1.68) | 22.2 (1.42) | 42.09 (1.82) | 51.38 (2.69) | 57.43 (3.55) |
| FARO15 | 1370 | 4.02 (0.46) | 28.74 (7.85) | 26.7 (11.1) | 25.8 (11.5) | 24.4 (10.8) | 54.05 (2.16) | 68.76 (3.68) | 74.80 (5.72) |
| Seed Source | Seed Lot ID | Collection Date | Seed Age DAH | Incubation Temperature (°C) | Light Conditions |
|---|---|---|---|---|---|
| Panarea (PAN) | PAN13 | July 2013 | 260 | 5, 10, 15, 20, 25, 30 | Light/Dark (12/12 h) Darkness (24 h) |
| 600 | |||||
| 770 | 15/10, 20/10, 20/15, 25/20 | ||||
| 2080 | 5, 10, 15, 20, 25, 30 | ||||
| PAN17 | July 2017 | 9 | 5, 10, 15, 20, 25, 30 15/10, 20/10, 20/15, 25/20 | ||
| 600 | 5, 10, 15, 20, 25, 30 | ||||
| 1000 | |||||
| Faro nursery (FARO) | FARO15 | July 2015 | 1370 | 5, 10, 15, 20, 25, 30 | Light/Dark (12/12 h) Darkness (24 h) |
| FARO16 | July 2016 | 1000 | |||
| FARO17 | July 2017 | 600 | |||
| 1000 | |||||
| FARO19 | July 2019 | 90 | |||
| 260 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carruggio, F.; Onofri, A.; Catara, S.; Impelluso, C.; Castrogiovanni, M.; Lo Cascio, P.; Cristaudo, A. Conditional Seed Dormancy Helps Silene hicesiae Brullo & Signor. Overcome Stressful Mediterranean Summer Conditions. Plants 2021, 10, 2130. https://doi.org/10.3390/plants10102130
Carruggio F, Onofri A, Catara S, Impelluso C, Castrogiovanni M, Lo Cascio P, Cristaudo A. Conditional Seed Dormancy Helps Silene hicesiae Brullo & Signor. Overcome Stressful Mediterranean Summer Conditions. Plants. 2021; 10(10):2130. https://doi.org/10.3390/plants10102130
Chicago/Turabian StyleCarruggio, Francesca, Andrea Onofri, Stefania Catara, Carmen Impelluso, Maria Castrogiovanni, Pietro Lo Cascio, and Antonia Cristaudo. 2021. "Conditional Seed Dormancy Helps Silene hicesiae Brullo & Signor. Overcome Stressful Mediterranean Summer Conditions" Plants 10, no. 10: 2130. https://doi.org/10.3390/plants10102130