
Allium pallasii and A. caricifolium—Surprisingly Diverse Old Steppe Species, Showing a Clear Geographical Barrier in the Area of Lake Zaysan

 ,
,
Abstract
:1. Introduction
2. Results
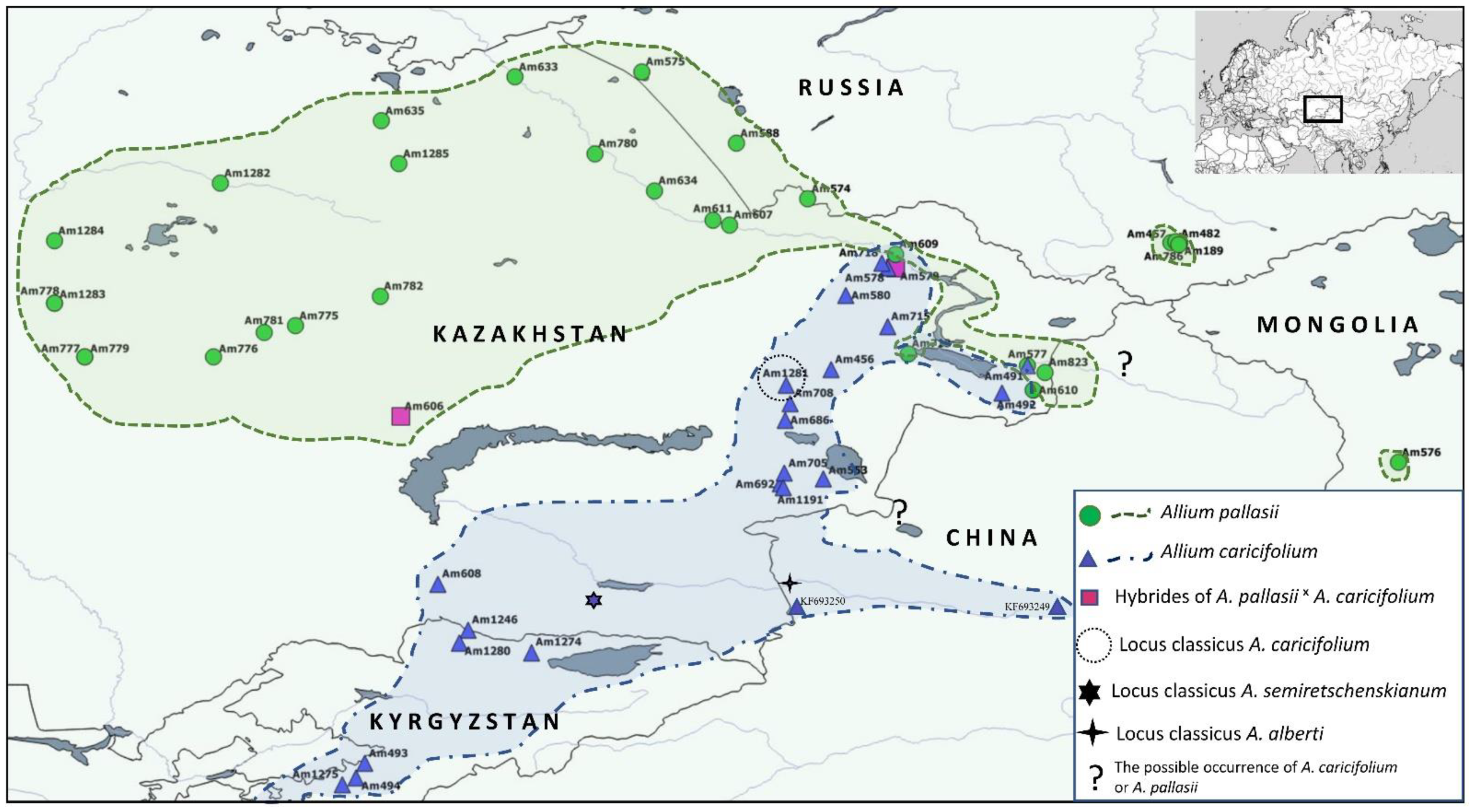
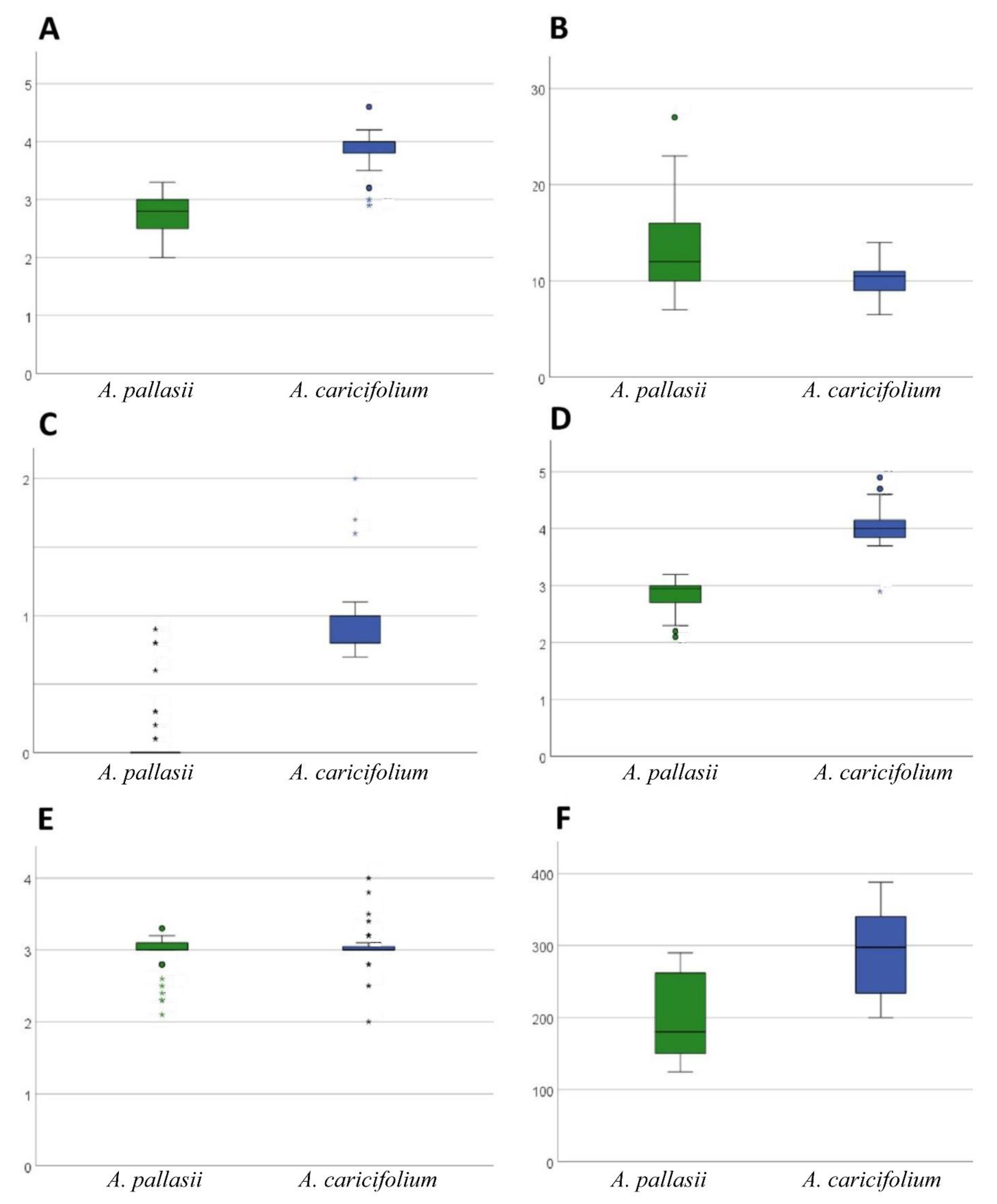
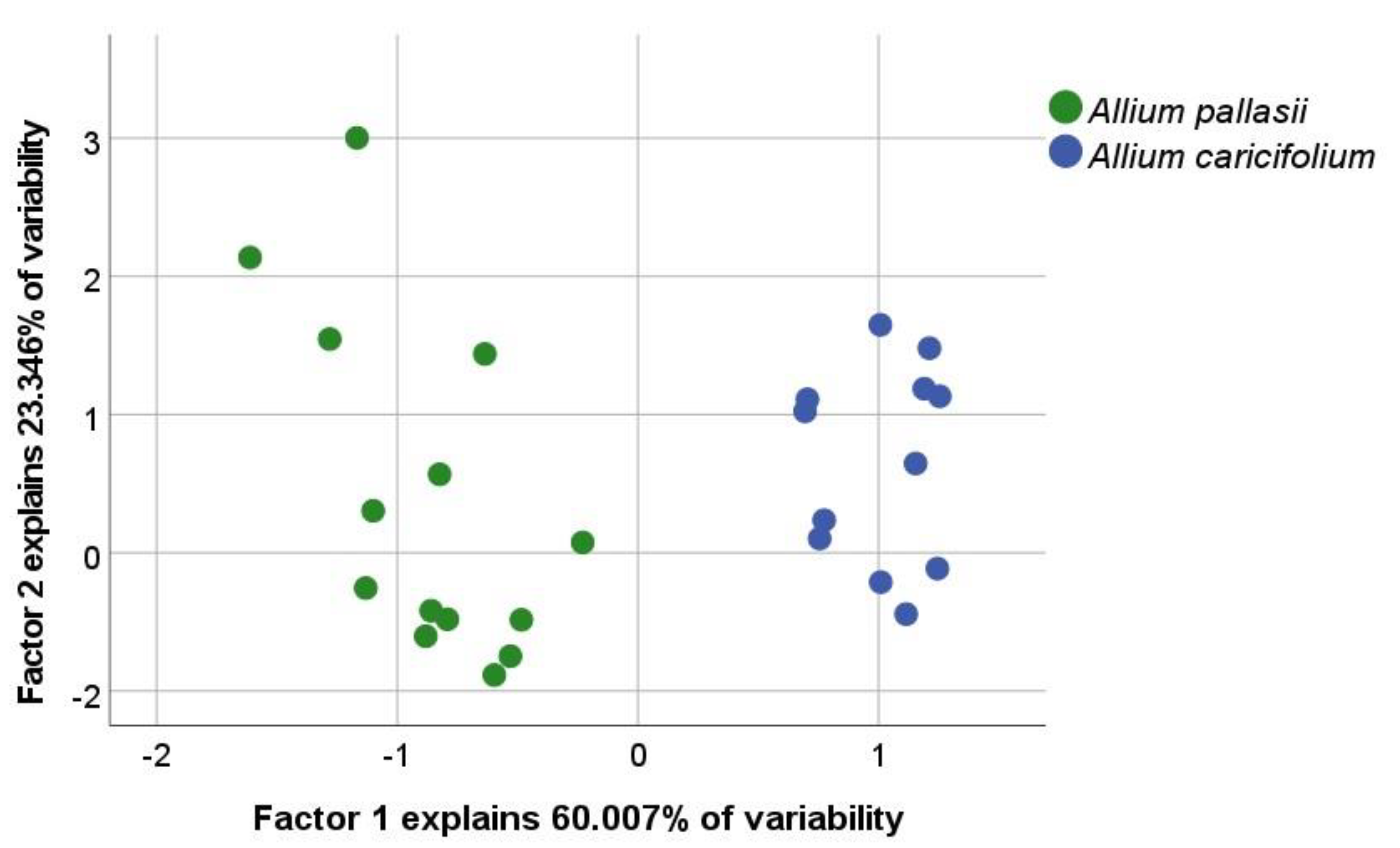
2.1. Morphology and Distribution
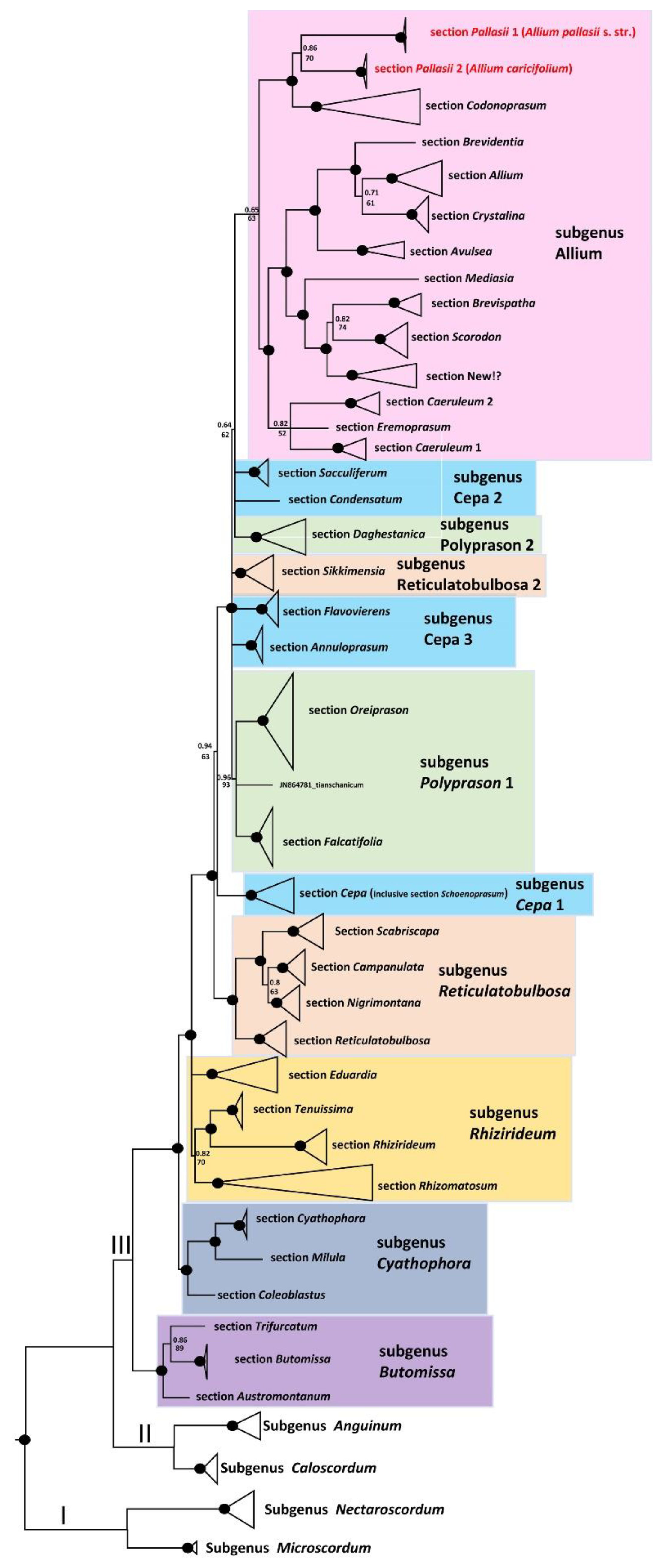
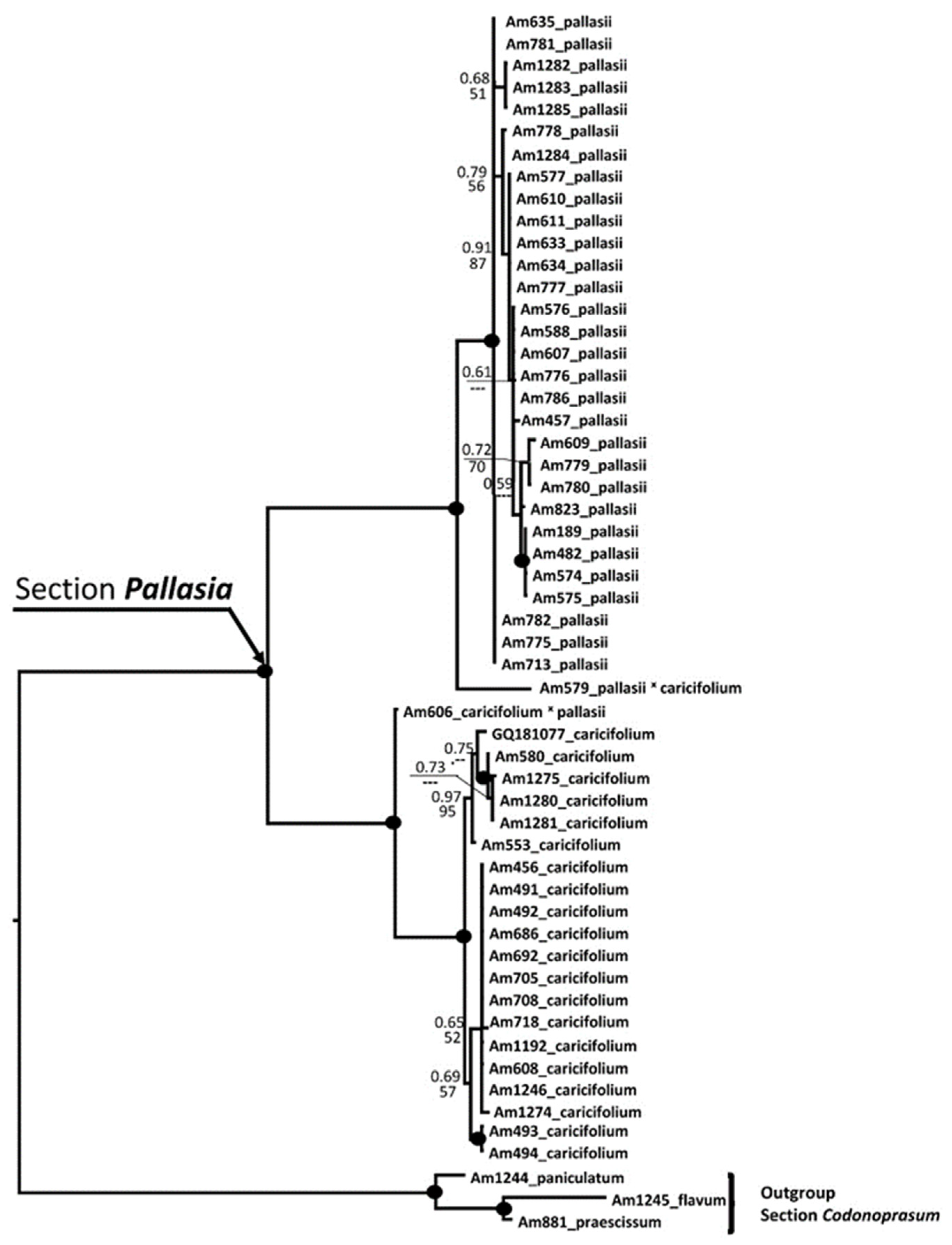
2.2. Phylogenetic Analyses
2.2.1. Position of A. pallasii s.l. in the Third Evolution Line
2.2.2. Phylogeny of Allium pallasii s.l.
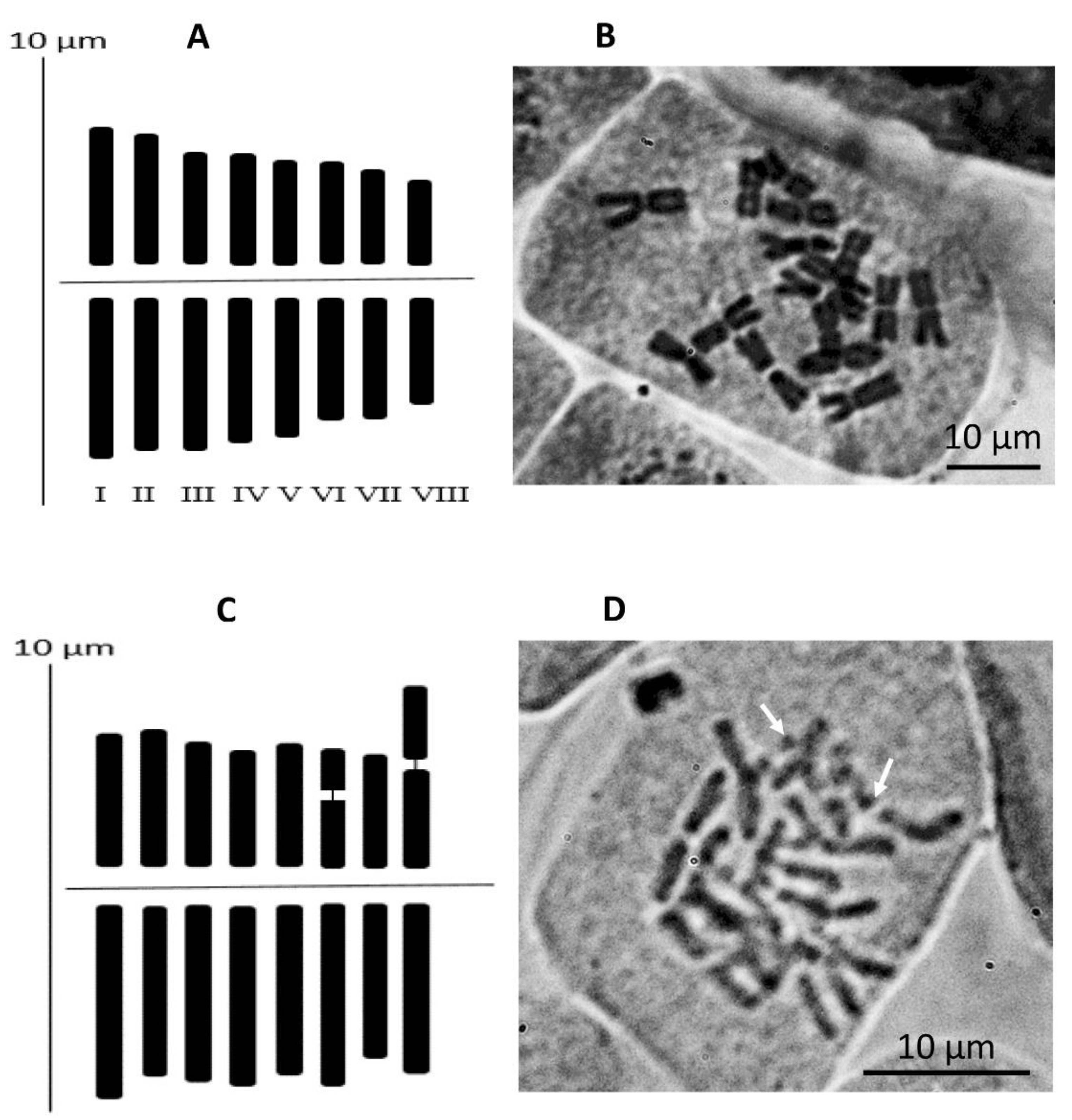
2.3. Cytology, Flowcytometry
2.4. Nomenclatural Remarks
3. Discussion
4. Materials and Methods
4.1. Morphological and Distribution Analyses
4.2. Taxon Sampling
4.3. DNA Extraction, Amplification and Sequencing
4.4. Phylogenetic Analyses
4.5. Cytology, Flowcytometry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Art Name | Coordinates | Country | Voucher | rITS | trnQ-rps16 | rpl32trnL |
|---|---|---|---|---|---|---|---|
| Am189 | Allium pallasii | 50°14′15″ N 87°57′34″ E | RU | OSBU: 18243 | OM891893 | ||
| Am457 | Allium pallasii | 50°14′14″ N 87°48′40″ E | RU | OSBU: 22275 | OM891894 | OM983335 | OM983382 |
| Am482 | Allium pallasii | 50°14′38″ N 87°53′48″ E | RU | OSBU: 23174 | OM891895 | ||
| Am574 | Allium pallasii | 51°2′49″ N 81°2′16″ E | KZ | NS: 0012843 | OM891896 | ||
| Am575 | Allium pallasii | 53°25′0″ N 77°57′0″ E | RU | NS: 0012841 | OM891897 | OM983336 | OM983383 |
| Am576 | Allium pallasii | 45°58′30″ N 92°14′59″ E | MN | ALTB: Smirnov, 19 June 1999 | OM891898 | OM983337 | OM983384 |
| Am577 | Allium pallasii | 47°55′60″ N 85°7′60″ E | KZ | ALTB: Smirnov, Antonjuk, 29 May 2000 | OM891899 | OM983338 | OM983385 |
| Am579 | Allium pallasii x A. caricifolium | 49°45′59″ N 82°38′9″ E | KZ | ALTB: Smirnov, Antonyuk, 20 May 2000 | OM891900 | OM983339 | OM983386 |
| Am588 | Allium pallasii | 52°4′48″ N 79°42′35″ E | RU | ALTB: Smirnov, 22 July 13 | OM891901 | OM983340 | OM983387 |
| Am607 | Allium pallasii | 50°33′40″ N 79°35′14″ E | KZ | OSBU: 23346 | OM891902 | OM983341 | OM983388 |
| Am609 | Allium pallasii | 49°54′58″ N 82°40′21″ E | KZ | TK: Prokopjev, 13 June 1982 | OM891903 | OM983342 | OM983389 |
| Am610 | Allium pallasii | 47°28′57″ N 85°14′25″ E | KZ | TK: Schischkin, 3 June 1914 | OM891904 | ||
| Am611 | Allium pallasii | 50°38′47″ N 79°16′5″ E | KZ | TK: Prokopjev et al., 1 June 1975 | OM891905 | ||
| Am633 | Allium pallasii | 53°19′0″ N 75°34′60″ E | KZ | NS: 0015875 | OM891906 | OM983343 | OM983390 |
| Am634 | Allium pallasii | 51°12′0″ N 78°10′60″ E | KZ | NS: 0015873 | OM891907 | OM983344 | OM983391 |
| Am635 | Allium pallasii | 52°29′55″ N 73°5′38″ E | KZ | NS: 0015872 | OM891908 | OM983345 | OM983392 |
| Am713 | Allium pallasii | 48°9′52″ N 82°54′23″ E | KZ | OSBU: 24384 | OM891909 | OM983346 | OM983393 |
| Am775 | Allium pallasii | 48°40′45″ N 71°29′20″ E | KZ | OSBU: 24800 | OM891910 | OM983348 | OM983394 |
| Am776 | Allium pallasii | 48°5′47″ N 69°57′58″ E | KZ | OSBU: 24841 | OM891911 | OM983349 | OM983395 |
| Am777 | Allium pallasii | 48°6′4″ N 67°34′3″ E | KZ | OSBU: 24890 | OM891912 | OM983347 | OM983396 |
| Am778 | Allium pallasii | 49°6′41″ N 66°59′48″ E | KZ | OSBU: 24875 | OM891913 | OM983350 | OM983397 |
| Am779 | Allium pallasii | 48°6′4″ N 67°34′3″ E | KZ | OSBU: 24898 | OM891914 | OM983351 | OM983398 |
| Am780 | Allium pallasii | 51°53′29″ N 77°3′51″ E | KZ | OSBU: 25000 | OM891915 | OM983352 | OM983399 |
| Am781 | Allium pallasii | 48°33′43″ N 70°54′16″ E | KZ | OSBU: 24835 | OM891916 | OM983353 | OM983400 |
| Am782 | Allium pallasii | 49°14′1″ N 73°3′58″ E | KZ | OSBU: 24770 | OM891917 | OM983354 | OM983401 |
| Am786 | Allium pallasii | 50°11′54″ N 87°56′31″ E | RU | OSBU: 23556 | OM891918 | OM983355 | OM983402 |
| Am823 | Allium pallasii | 47°48′46″ N 85°27′40″ E | KZ | ALTB: Starikov. 21 May 1990 | OM891919 | OM983356 | OM983403 |
| Am1282 | Allium pallasii | 51°20′11″ N 70°5′25″ E | KZ | OSBU: 24956 | OM891920 | OM983357 | OM983404 |
| Am1283 | Allium pallasii | 49°6′41″ N 66°59′48″ E | KZ | OSBU: 24765 | OM891921 | OM983358 | OM983405 |
| Am1284 | Allium pallasii | 50°15′49″ N 66°59′32″ E | KZ | OSBU:24931 | OM891922 | OM983359 | OM983406 |
| Am1285 | Allium pallasii | 51°41′58″ N 73°25′27″ E | KZ | OSBU: 28157 | OM891923 | OM983360 | OM983407 |
| Am456 | Allium caricifolium | 47°52′6″ N 81°28′34″ E | KZ | ALTB: Kechaikin, 2 June 12 | OM891924 | OM983361 | OM983408 |
| Am491 | Allium caricifolium | 47°55′60″ N 85°7′60″ E | KZ | OSBU: 22298 | OM891925 | OM983362 | OM983409 |
| Am492 | Allium caricifolium | 47°25′53″ N 84°39′11″ E | KZ | OSBU: 22290 | OM891926 | OM983410 | |
| Am493 | Allium caricifolium | 40°31′51″ N 72°47′11″ E | KG | GAT: 0017895 | OM891927 | ||
| Am494 | Allium caricifolium | 40°16′23″ N 72°37′30″ E | KG | GAT: 0017900 | OM891928 | ||
| Am553 | Allium caricifolium | 45°50′5″ N 81°19′56″ E | KZ | TK: Goloskokov, 18 June 1959 | OM891929 | OM983363 | OM983411 |
| Am578 | Allium caricifolium | 49°45′59″ N 82°32′9″ E | ALTB: Kechaikin, Tjutjunic, 1 June 2012 | OM983364 | |||
| Am580 | Allium caricifolium | 49°15′0″ N 81°45′0″ E | KZ | ALTB: Smirnov, Antonjuk, 18 May 2000 | OM891930 | OM983365 | OM983412 |
| Am606 | Allium caricifolium x A.pallasii | 46°59′40″ N 73°26′60″ E | KZ | TK: Pavlov, 28 May 1951 | OM891931 | OM983366 | OM983413 |
| Am608 | Allium caricifolium | 43°51′55″ N 74°8′19″ E | KZ | TK: Pavlov, 11 May 1951 | OM891932 | OM983367 | OM983414 |
| Am686 | Allium caricifolium | 46°55′29″ N 80°36′52″ E | KZ | OSBU: 24335 | OM891933 | OM983368 | OM983415 |
| Am692 | Allium caricifolium | 45°43′55″ N 80°31′33″ E | KZ | OSBU: 24340 | OM891934 | OM983369 | OM983416 |
| Am705 | Allium caricifolium | 45°55′58″ N 80°36′31″ E | KZ | OSBU: 24368 | OM891935 | OM983370 | OM983417 |
| Am708 | Allium caricifolium | 47°14′20″ N 80°43′6″ E | KZ | OSBU: 24370 | OM891936 | OM983371 | OM983418 |
| Am718 | Allium caricifolium | 49°51′16″ N 82°25′26″ E | KZ | OSBU: 24390 | OM891937 | OM983372 | OM983419 |
| Am1192 | Allium caricifolium | 45°39′37″ N 80°34′56″ E | KZ | OSBU: 24354 | OM891938 | OM983373 | OM983420 |
| Am1246 | Allium caricifolium | 43°0′56″ N 74°42′19″ E | KG | OSN:2021-0746-W | OM891939 | OM983374 | OM983421 |
| Am1274 | Allium caricifolium | 42°36′6″ N 75°53′39″ E | KG | GAT: 3060090 | OM891940 | OM983375 | OM983422 |
| Am1275 | Allium caricifolium | 40°8′29″ N 72°21′47″ E | KG | GAT: 2543277 | OM891941 | OM983376 | OM983423 |
| Am1280 | Allium caricifolium | 42°47′0″ N 74°32′53″ E | KG | FRU: Usulbaev A.K., 20 May 2019 | OM891942 | OM983377 | OM983424 |
| Am1281 | Allium caricifolium | 47°34′42″ N 80°37′47″ E | KZ | OSBU: 24328 | OM891943 | OM983378 | OM983425 |
| Am826 | Allium caesium | 48°3′22″ N 68°28′33″ E | KZ | OSBU: 24961 | OM891944 | OM983426 | |
| Am683 | Allium caeruleum | 47°24′53″ N 80°34′56″ E | KZ | OSBU: 24332 | OM891945 | ||
| Am473 | Allium delicatulum | 47°52′59″ N 81°28′34″ E | KZ | ALTB: Kechaikin, 2 June 2012 | OM891946 | ||
| Am573 | Allium delicatulum | 51°44′13″ N 94°28′26″ E | RU | NS: 0014638 | OM891947 | ||
| Am712 | Allium delicatulum | 47°55′39″ N 82°4′10″ E | KZ | OSBU: 24380 | OM891948 | OM983427 | |
| Am749 | Allium zaissanicum | 48°49′11″ N 83°46′12″ E | KZ | OSBU: 23940 | OM891949 | ||
| Am1243 | Allium moschatum | 46°47′22″ N 17°18′24″ E | HUN | OSN: 2019-0872-W | OM891950 | OM983429 | |
| Am1245 | Allium flavum | 48°54′14″ N 21°57′56″ E | SVK | OSN:2004-0826-W | OM891951 | OM983379 | OM983428 |
| Am1239 | Allium vineale | 48°17′16″ N 16°50′36″ E | A | OSBU: 17147 | OM891952 | ||
| Am1244 | Allium paniculatum | 49°55′26″ N 42°25′22″ E | RU | OSN: 2018-1203-W | OM891953 | OM983380 | OM983430 |
| Am881 | Allium praescisum | 52°29′48″ N 61°58′0″ E | KZ | OSBU: 25829 | OM891954 | OM983381 | OM983431 |
| Am630 | Allium amphibolum | 49°17′14″ N 87°43′4″ E | RU | OSBU: 23610 | OM983432 | ||
| Am875 | Allium lineare | 49°6′56″ N 72°40′59″ E | KZ | OSBU: 24823 | OM983433 | ||
| Am848 | Allium strictum | 49°55′58″ N 14°8′31″ E | CZ | FRT 2015/62 | OM983434 | ||
| Am1286 | Allium scabriscapum | 36°2′12″ N 51°12′10″ E | IR | GAT 19116 | OM983435 | ||
| Am1240 | Allium kunthianum | 42°39′52″ N 44°37′0″ E | GEO | OSN: 2020-0819-W | OM983436 | ||
| Am1241 | Allium oleraceum | 52°21′22″ N 14°10′12″ E | DE | OSN: 2016-0568-W | OM983437 |
References
- Govaerts, R.; Kington, S.; Friesen, N.; Fritsch, R.; Snijman, D.A.; Marcucci, R.; Silverstone-Sopkin, P.A.; Brullo, S. 2005–2020. World Checklist of Amaryllidaceae. Available online: http://apps.kew.org/wcsp/ (accessed on 20 June 2020).
- Stearn, W.T. European species of Allium and allied genera of Alliaceae: A synonymy enumeration. Ann. Mus. Goulandris 1978, 4, 83–198. [Google Scholar]
- Stearn, W.T. How many species of Allium are known? Kew Mag. 1992, 9, 180–182. [Google Scholar] [CrossRef]
- Fritsch, R.M. and Friesen, N. Evolution, domestication, and taxonomy. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 5–30. [Google Scholar]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Friesen, N. Chapter 1. Genus Allium: Evolution, Classification, and Domestication. In Edible Alliums: Modern Biology, Production and Uses; Rabinowitch, H.D., Brian, T., Eds.; CABI Publishing: Wallingford, UK, 2022; in press. [Google Scholar]
- Friesen, N.; Fritsch, R.M.; Blattner, F.R. Phylogeny and new intrageneric classification of Allium L. (Alliaceae) based on nuclear rDNA ITS sequences. Aliso 2006, 22, 372–395. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Driscoll, H.E.; Specht, C.D. A molecular phylogeny of the wild onions (Allium; Alliaceae) with a focus on the western North American center of diversity. Mol. Phylogenet. Evol. 2008, 47, 1157–1172. [Google Scholar] [CrossRef]
- Li, Q.Q.; Zhou, S.D.; He, X.J.; Yu, Y.; Zhang, Y.C.; Wei, X.Q. Phylogeny and biogeography of Allium (Amaryllidaceae: Allieae) based on nuclear ribosomal internal transcribed spacer and chloroplast rps16 sequences, focusing on the inclusion of species endemic to China. Ann. Bot. 2010, 106, 709–733. [Google Scholar] [CrossRef]
- Wheeler, E.J.; Mashayekhi, S.; McNeal, D.W.; Columbus, J.T.; Pires, J.C. Molecular systematics of Allium subgenus Amerallium (Amaryllidaceae) in North America. Am. J. Bot. 2013, 100, 701–711. [Google Scholar] [CrossRef]
- Hauenschild, F.; Favre, A.; Schnitzler, J.; Michalak, I.; Freiberg, M.; Muellner-Riehl, A.N. Spatio–temporal evolution of Allium L. in the Qinghai–Tibet–Plateau region: Immigration and in situ radiation. Plant Divers. 2017, 39, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.F.; Yu, H.X.; Price, M.; Xie, C.; Deng, Y.-Q.; Chen, J.P.; Yu, Y.; Zhou, S.D.; He, X.J. Phylogeny of Chinese Allium species in Section Daghestanica and adaptive evolution of Allium (Amaryllidaceae, Allioideae) species revealed by the chloroplast complete genome. Front. Plant Sci. 2019, 10, 460. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.F.; Tan, J.B.; Yu, Y.; Gui, L.J.; Su, D.M.; Zhou, S.D.; He, X.J. Insights into phylogeny, age and evolution of Allium (Amaryllidaceae) based on the whole plastome sequences. Ann. Bot. 2020, 125, 1039–1055. [Google Scholar] [CrossRef]
- Costa, L.; Jimenez, H.; Carvalho, R.; Carvalho-Sobrinho, J.; Escobar, I.; Souza, G. Divide to Conquer: Evolutionary History of Allioideae Tribes (Amaryllidaceae) Is Linked to Distinct Trends of Karyotype Evolution. Front. Plant Sci. 2020, 11, 320. [Google Scholar] [CrossRef]
- Li, Q.Q.; Zhou, S.D.; Huang, D.Q.; He, X.J.; Wei, X.Q. Molecular phylogeny, divergence time estimates and historical biogeography within one of the world’s largest monocot genera. AoB Plants 2016, 8, plw041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, N.; Smirnov, S.; Herden, T.; Oyuntsetseg, B.; Shmakov, A.; Hurka, H. Allium species of section Rhizomatosa, early members of the Central Asian steppe vegetation. Flora 2020, 263, 151536. [Google Scholar] [CrossRef]
- Murray, A. Observationes botanica. In Novi Commentarii Societatis Regiae Scientiarum Gottingensis; Joann Christian Dieterich: Göttingen, Germany, 1775; Volume 6, pp. 23–40. (In Latin) [Google Scholar]
- Friesen, N. Rody Allium i Calloscordum Herbert. In Flora Sibiri. Tom 4 Araceae—Orchidaceae; Malyshev, L.I., Peshkova, G.A., Eds.; Nauka—Sibirskoe Otdelenie: Novosibirsk, Russia, 1987; pp. 55–97, 177–195. (In Russian) [Google Scholar]
- Friesen, N. The Genus Allium L. in the Flora of Mongolia. Feddes Repert. 1995, 106, 59–81. [Google Scholar] [CrossRef]
- Baasanmunkh, S.; Oyuntsetseg, B.; Oyundary, C.; Oyundelger, K.; Urgamal, M.; Darikhand, D.; Soninkhishig, N.; Kechaykin, A.A.; Shmakov, A.I.; Erst, A.S.; et al. The vascular plant diversity of Dzungarian Gobi in western Mongolia, with an annotated checklist. Phytotaxa 2021, 501, 1–55. [Google Scholar] [CrossRef]
- Vvedensky, A.I. Luk—Allium L. In Flora SSSR. Tom 4; Komarov, V.L., Ed.; Izd. Akad. Nauk SSSR: Leningrad, Russia, 1935; pp. 112–280. (In Russian) [Google Scholar]
- Pavlov, N.V.; Polyakov, P.P. Genus Allium L. In Flora Kazakhstana; Pavlov, N.V., Ed.; Publishing House of the Academy of Sciences of the KazSSR: Almaty, Kazakhstan, 1958; Volume 2, pp. 134–193. (In Russian) [Google Scholar]
- Vakhtina, L.I.; Kudryashova, G.L. Cytotaxonomic studies of some Allium species in Kazakhstan. Bot. Zhurn. 1977, 62, 539–547. [Google Scholar]
- Zakirova, R.O.; Nafanailova, I.I. Chromosome numbers in some species of the Kazakhstan flora. Bot. Zhurn. 1988, 73, 1493–1494. [Google Scholar]
- He, X.J.; Xu, J.M. Karyotypes of 4 species in Allium sect. Haplostemon Boiss. from China. Acta Phytotax. Sin. 2001, 39, 423–432. [Google Scholar]
- POWO. Available online: https://powo.science.kew.org/results?q=Allium%20pallasii (accessed on 1 April 2022).
- Xie, F.M.; Xie, D.F.; Xie, C.; Yu, Y.; Zhou, S.D.; He, X.J. Adaptation Evolution and Phylogenetic Analyses of Species in Chinese Allium Section Pallasia and Related Species Based on Complete Chloroplast Genome Sequences. BioMed Res. Int. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Friesen, N.; Smirnov, S.V.; Leweke, M.; Seregin, A.P.; Fritsch, R.M. Taxonomy and Phylogeny of Allium section Decipientia (Amaryllidaceae): Morphological characters do not reflect the evolutionary history verified by molecular markers. Bot. J. Linn. Soc. 2021, 197, 190–228. [Google Scholar] [CrossRef]
- Linnaeus, C. Species Plantarum; Laurentii Salvii: Stockholm, Sweden, 1753; Volume 1. (In Latin) [Google Scholar]
- Linnaeus, C. Systema Naturae. Editio Decima, Reformata; Laurentius Salvius: Stockholm, Sweden, 1759; Volume 2. (In Latin) [Google Scholar]
- Reichenbach, H.G.L. Iconographia Botanica Seu Plantae Criticae; Apud Fridericum Hofmeister: Leipzig, Germany, 1827; Volume 5. (In Latin) [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Tzanoudakis, D. Karyotypes of ten taxa of Allium section Scorodon from Greece. Caryologia 1983, 36, 259–284. [Google Scholar] [CrossRef] [Green Version]
- Don, G. A monograph of the genus Allium. Mem. Wernerian Nat. Hist. Soc. 1832, 6, 1–102. [Google Scholar]
- Ledebour, C.F. Icones Plantarum Novarum Vel Imperfecte Cognitarum Floram Rossicam, Imprimis Altaicam Illustrantes, Centuria II; I. Deubner: Riga, Latvia, 1833; p. 16. (In Latin) [Google Scholar]
- Ledebour, C.F. Flora Rossica Sive, Enumeratio Plantarum in Totius Imperii Rossici Provinciis Europaeis, Asiaticis et Americanis Hucusque Observatarum; Sumptibus Librariae E. Schweizerbart: Stuttgart, Germany, 1853; Volume 4. (In Latin) [Google Scholar]
- Regel, E. Allii Species Asiae Centralis in Asia Media a Turcomania Desertisque Araliensibus et Caspicis Usque ad Mongoliam Crescents. Acta Horti Petropolitani 1887, 10, 278–362. (In Latin) [Google Scholar]
- Karelin, G.S.; Kirilow, I.P. Enumeratio Plantarum in Desertis Songoriae Orientalis et in Jugo Summarum Alpium Alatau Anno 1841 Collectarum. Bull. Soc. Imp. Nat. Moscou 1842, 15, 503–542. (In Latin) [Google Scholar]
- Gubanov, I.A. Catalogue of Authentic Specimens of Vascular Plants of the Moscow State University Herbarium (MW); Gerbarii im. DP Syreishchikov: Moscow, Russia, 2002; p. 213. [Google Scholar]
- Regel, E. Descriptiones Plantarum Novarum Et Minus Cognitarum, Fasciculus VI. Acta Horti Petropolitani 1878, 5, 575–646. (In Latin) [Google Scholar]
- Regel, E. Alliorum Adhuc Cognitorum Monographia. Acta Horti Petropolitani 1875, 3, 1–266. (In Latin) [Google Scholar]
- Regel, A. Reisen in Central Asien, 1876–1879. In Dr. A. Petermann’s Mittheilungen aus Justus Perthes’ Geographischer Anstalt; Justus Perthes: Gotha, Germany, 1879. (In German) [Google Scholar]
- Egorova, T.V. Genus Allium. In Plantae Asian Centralis; Grubov, V.I., Egorova, T.V., Eds.; Nauka: Saint Petersburg, Russia, 1977; Volume 7, pp. 17–67. [Google Scholar]
- Khasanov, F. Amaryllidaceae in Flora of Uzbekistan; Sennikov, A.N., Ed.; Navro’z Publishers: Tashkent, Uzbekistan, 2017; Volume 1, p. 173. [Google Scholar]
- Zagolova, V.G. To the taxonomy of the section Rhizirideum of the genus Allium in Kazakhstan. Bot. Mater. Gerbarija Inst. Bot. Akad. Nauk. KazSSR 1974, 8, 16–19. (In Russian) [Google Scholar]
- Khasanov, F.O.; Karimov, F.; Tirkasheva, B. Taxonomic revision and Lectotypification of Allium sect. Coerulea (Omelch.) F.O.Khass. Stapfia 2013, 99, 208–234. [Google Scholar]
- Akhmetiev, M.A.; Zaporozhets, N.I. Paleogene events in central Eurasia: Their role in the flora and vegetation cover evolution, migration of phytochore boundaries, and climate changes. Stratigr. Geol. Correl. 2014, 22, 312–335. [Google Scholar] [CrossRef]
- Popova, S.; Utescher, T.; Gromyko, D.V.; Bruch, A.A.; Henrot, A.-J.; Mosbrugger, V. Cenozoic vegetation gradients in the mid- and higher latitudes of Central Eurasia and climatic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 467, 69–82. [Google Scholar] [CrossRef]
- Akhmetyev, M.A.; Dodonov, A.E.; Sotnikova, M.V.; Spasskaya, I.I.; Kremenetzky, K.V.; Klimanov, V.A. Kazakhstan and central Asia—Plains and foothills. In Climate and Environment Changes during the Last 65 Million Years; Cenozoic: From Paleocene to Holocene; Velichko, A.A., Nechaev, V.P., Eds.; GEOS: Moscow, Russia, 1999; pp. 165–191. (In Russian) [Google Scholar]
- Popova, S.; Utescher, T.; Averyanova, A.; Tarasevich, V.; Tropina, P.; Xing, Y. Early Miocene flora of central Kazakhstan (Turgai Plateau) and its paleoenvironmental implications. Plant Divers. 2019, 41, 183–197. [Google Scholar]
- Dorfman, B.F. Zaysan—the Only Surviving Cretaceous Lake—May be Lost. Procedia Environ. Sci. 2011, 10, 1376–1382. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Carter, A.; Dong, J.; Bao, Z.; An, Y.; Guo, Z. Mesozoic–Tertiary exhumation history of the Altai Mountains, northern Xinjiang, China: New constraints from apatite fission-track data. Tectonophysics 2006, 412, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zheng, D.; Zhang, H.; Pang, J.; Wang, W.; Wang, Y.; Wu, Y.; He, H.; Stuart, F.M.; Xu, S. Plio-Pleistocene establishment of Irtysh River in Junggar, northwest China: Implications for Siberian-Arctic river system evolution and resulting climate impact. Geophys. Res. Lett. 2021, 48, e2021GL093217. [Google Scholar] [CrossRef]
- Kremenetsky, K.V.; Tarasov, P.E.; Cherkinsky, A.E. Holocene history of the Kazakhstan “island” pine forests. Bot. Zhurn. 1994, 94, 13–29. (In Russian) [Google Scholar]
- Rudaya, N.; Krivonogov, S.; Slowinski, M.; Cao, X.; Zhilich, S. Postglacial history of the Steppe Altai: Climate, fire and plant diversity. Quat. Sci. Rev. 2020, 249, 106616. [Google Scholar]
- Butvilovsky, V.V. Paleogeography of the Last Glaciation and the Holocene of Altai: An Event-Catastrophic Model/v.v. Butvilovsky; Publishing House Tomsk University: Tomsk, Russia, 1993; p. 252. (In Russian) [Google Scholar]
- Rudoy, A.N. Glacier-Dammed Lakes and geological work of glacial super floods in the Late Pleistocene, Southern Siberia, Altai Mountains. Quat. Int. 2002, 87, 119–140. [Google Scholar] [CrossRef]
- Seregin, A.; Anackov, G.; Friesen, N. Molecular and morphological revision of the Allium saxatile group (Amaryllidaceae): Geographical isolation as the driving force of underestimated speciation. Bot. J. Linn. Soc. 2015, 178, 67–101. [Google Scholar] [CrossRef] [Green Version]
- Friesen, N.; Vesselova, P.; Osmonaly, B.; Sitpaeva, G.; Luferov, A.; Shmakov, A. Allium toksanbaicum (Amaryllidaceae), a new species from Southeast Kazachstan. Phytotaxa 2021, 494, 251–267. [Google Scholar] [CrossRef]
- Seidl, A.; Tremetsberger, K.; Planzelt, S.; Blattner, F.R.; Neuffer, B.; Friesen, N.; Hurka, H.; Shmakov, A.; Oyuntsetseg, B.; Zerdoner Calasan, A.; et al. The phylogeographic history of Krascheninnikovia reflecs the development of the dry steppes and semy-deserts in Eurasia. Sci. Rep. 2021, 11, 6645. [Google Scholar] [CrossRef]
- Volkova, P.; Herden, T.; Friesen, N. Genetic variation in Goniolimon speciosum (Plumbaginaceae) reveals a complex history of steppe vegetation. Bot. J. Linn. Soc. 2017, 184, 113–121. [Google Scholar]
- Hurka, H.; Friesen, N.; Bernhardt, K.-G.; Neuffer, B.; Smirnov, S.V.; Shmakov, A.I.; Blattner, F.R. The Eurasian steppe belt: Status quo, origin and evolutionary history. Turczaninowia 2019, 22, 5–71. [Google Scholar]
- Török, P.; Neuffer, B.; Heilmeier, H.; Bernhardt, K.-G.; Wesche, K. Climate, landscape history and management drive Eurasian steppe biodiversity. Flora 2020, 271, 151685. [Google Scholar] [CrossRef]
- Seregin, A.P. (Ed.) Moscow Digital Herbarium: Electronic Resource. Moscow State University: Moscow, Russia. Available online: https://plant.depo.msu.ru/ (accessed on 1 April 2022).
- Kovtonyuk, N.K.; Han, I.; Gatilova, E.; Friesen, N. Genus Allium in CSBS Digital Herbarium. BIO Web Conf. 2020, 24, 00042. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 15 January 2022).
- Blattner, F.R. Direct amplification of the entire ITS region from poorly preserved plant material using recombinant PCR. Biotechnology 1999, 27, 1180–1185. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Shaw, J.E.B.; Lickey, E.E.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X window interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.-L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2002.
- Kluge, A.G.; Farris, J.S. Quantitative phyletics and the evolution of anurans. Syst. Zool. 1969, 18, 1–32. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, R.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnov, Y.A. Accelerated method of study of somatic chromosomes of fruit plants. Tsitologiya 1968, 10, 1601–1602. (In Russian) [Google Scholar]
- Altinordu, F.; Peruzzi, L.; Yu, Y.; He, X. A tool for the analysis of chromosomes: KaryoType. Taxon 2016, 65, 586–592. [Google Scholar] [CrossRef]
- Pfosser, A.; Amon, A.; Lelley, T.; Heberle-Bors, E. Evaluation of sensitivity of flow cytometry in detecting aneuploidy in wheat using disomic and ditelosomic wheat-rye addition lines. Cytometry 1985, 21, 387–393. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef] [Green Version]







| Character | A. pallasii | A. caricifolium |
|---|---|---|
| Scape length | 194.7 ± 55.3 (125–290) | 275 ± 63 (200–388) |
| Pedicel length | 12.9 ± 3.7 (7.5–19) | 10.4 ± 1.6 (6.5–14) |
| Tepals length | 2.9 ± 0.1 (2.1–3.3) | 3 ± 0.2 (2.0–4.0) |
| The ratio of tepals length/pedicels length | 1.1 | 1.2 |
| Length of filaments outside the tepals | 0.1 ± 0.1 (0) | 1 ± 0.1 (0.7–1.1) |
| Filaments length in total | 2.9 ± 0.1 (2.6–3) | 4 ± 0.1 (3.8–4.3) |
| The ratio filaments length outside the tepals/total length | 1.02 | 1.25 |
| Carpel length | 2.8 ± 0.1 (2–3.3) | 3.9 ± 0.1 (2.9–4.6) |
| Flowers per inflorescence | 36.8 ± 17.5 (22–75) | 52.8 ± 12.9 (37–66) |
| Pair No. | TAL (µm) | RL % | LA (µm) | SA (µm) | Sat | CI % | Type |
|---|---|---|---|---|---|---|---|
| 1 | 6.6 ± 0.8 | 15.1 ± 0.2 | 3.5 ± 0.4 | 3.0 ± 0.5 | 0 | 46.3 ± 1.4 | m |
| 2 | 6.2 ± 0.7 | 14.3 ± 0.2 | 3.4 ± 0.3 | 2.9 ± 0.4 | 0 | 46.1 ± 2.0 | m |
| 3 | 5.9 ± 0.7 | 13.6 ± 0.2 | 3.4 ± 0.4 | 2.6 ± 0.4 | 0 | 43.4 ± 1.1 | m |
| 4 | 5.6 ± 0.9 | 12.9 ± 0.4 | 3.2 ± 0.5 | 2.4 ± 0.4 | 0 | 43.2 ± 1.1 | m |
| 5 | 5.4 ± 0.7 | 12.3 ± 0.1 | 3.1 ± 0.3 | 2.3 ± 0.5 | 0 | 42.8 ± 2.6 | m |
| 6 | 5.0 ± 0.6 | 11.5 ± 0.3 | 2.8 ± 0.2 | 2.2 ± 0.4 | 0 | 44.5 ± 2.4 | m |
| 7 | 4.7 ± 0.6 | 10.8 ± 0.1 | 2.7 ± 0.4 | 2.1 ± 0.3 | 0 | 43.8 ± 1.5 | m |
| 8 | 4.2 ± 0.6 | 9.6 ± 0.1 | 2.3 ± 0.2 | 1.9 ± 0.4 | 0 | 44.4 ± 2.6 | m |
| Pair No. | TAL (µM) | RL % | LA (µM) | SA (µM) | Sat | CI % | Type |
|---|---|---|---|---|---|---|---|
| 1 | 7.2 ± 1.4 | 13.9 ± 2.5 | 4.3 ± 0.7 | 2.9 ± 0.9 | 0 | 40.7 ± 8.7 | m |
| 2 | 6.7 ± 1.2 | 13.1 ± 2.1 | 3.7 ± 0.6 | 3.0 ± 1.1 | 0 | 44.6 ± 11.4 | m |
| 3 | 6.6 ± 1.1 | 12.9 ± 1.9 | 3.9 ± 0.7 | 2.8 ± 0.5 | 0 | 41.6 ± 4.0 | m |
| 4 | 6.5 ± 0.3 | 12.5 ± 1.1 | 3.9 ± 0.3 | 2.5 ± 0.3 | 0 | 39.1 ± 4.5 | m |
| 5 | 6.4 ± 0.5 | 12.4 ± 0.9 | 3.7 ± 0.3 | 2.7 ± 0.5 | 0 | 41.9 ± 5.9 | m |
| 6 | 6.4 ± 0.8 | 12.4 ± 1.2 | 3.7 ± 0.4 | 1.8 ± 0.8 | 1 | 27.4 ± 8.1 | m |
| 7 | 5.9 ± 1.3 | 11.4 ± 2.0 | 3.4 ± 0.7 | 2.5 ± 0.6 | 0 | 42.4 ± 5.6 | m |
| 8 | 5.9 ± 1.0 | 11.4 ± 2.5 | 3.7 ± 0.7 | 2.2 ± 0.6 | 2 | 36.7 ± 7.6 | m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friesen, N.; Grützmacher, L.; Skaptsov, M.; Vesselova, P.; Dorofeyev, V.; Luferov, A.N.; Turdumatova, N.; Lazkov, G.; Smirnov, S.V.; Shmakov, A.I.; et al. Allium pallasii and A. caricifolium—Surprisingly Diverse Old Steppe Species, Showing a Clear Geographical Barrier in the Area of Lake Zaysan. Plants 2022, 11, 1465. https://doi.org/10.3390/plants11111465
Friesen N, Grützmacher L, Skaptsov M, Vesselova P, Dorofeyev V, Luferov AN, Turdumatova N, Lazkov G, Smirnov SV, Shmakov AI, et al. Allium pallasii and A. caricifolium—Surprisingly Diverse Old Steppe Species, Showing a Clear Geographical Barrier in the Area of Lake Zaysan. Plants. 2022; 11(11):1465. https://doi.org/10.3390/plants11111465
Chicago/Turabian StyleFriesen, Nikolai, Lisa Grützmacher, Mikhail Skaptsov, Polina Vesselova, Vladimir Dorofeyev, Alexander N. Luferov, Nazgul Turdumatova, Georgii Lazkov, Sergei V. Smirnov, Alexander I. Shmakov, and et al. 2022. "Allium pallasii and A. caricifolium—Surprisingly Diverse Old Steppe Species, Showing a Clear Geographical Barrier in the Area of Lake Zaysan" Plants 11, no. 11: 1465. https://doi.org/10.3390/plants11111465