
Chickpea (Cicer arietinum L.) Biology and Biotechnology: From Domestication to Biofortification and Biopharming
 , ,
, ,
Abstract
:1. Introduction
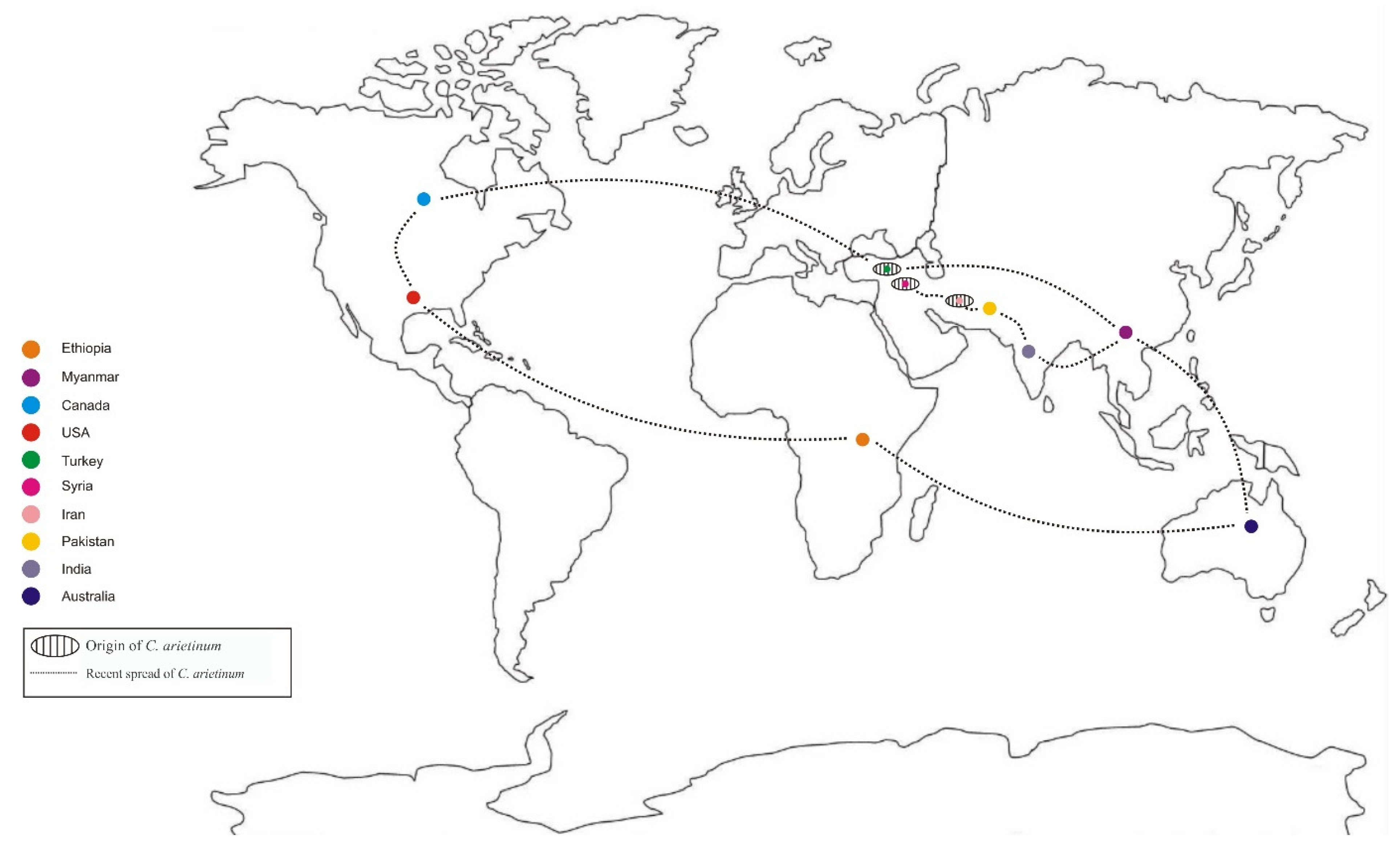
1.1. Origin and Distribution
1.2. Morphology
1.3. Nutritional Profile
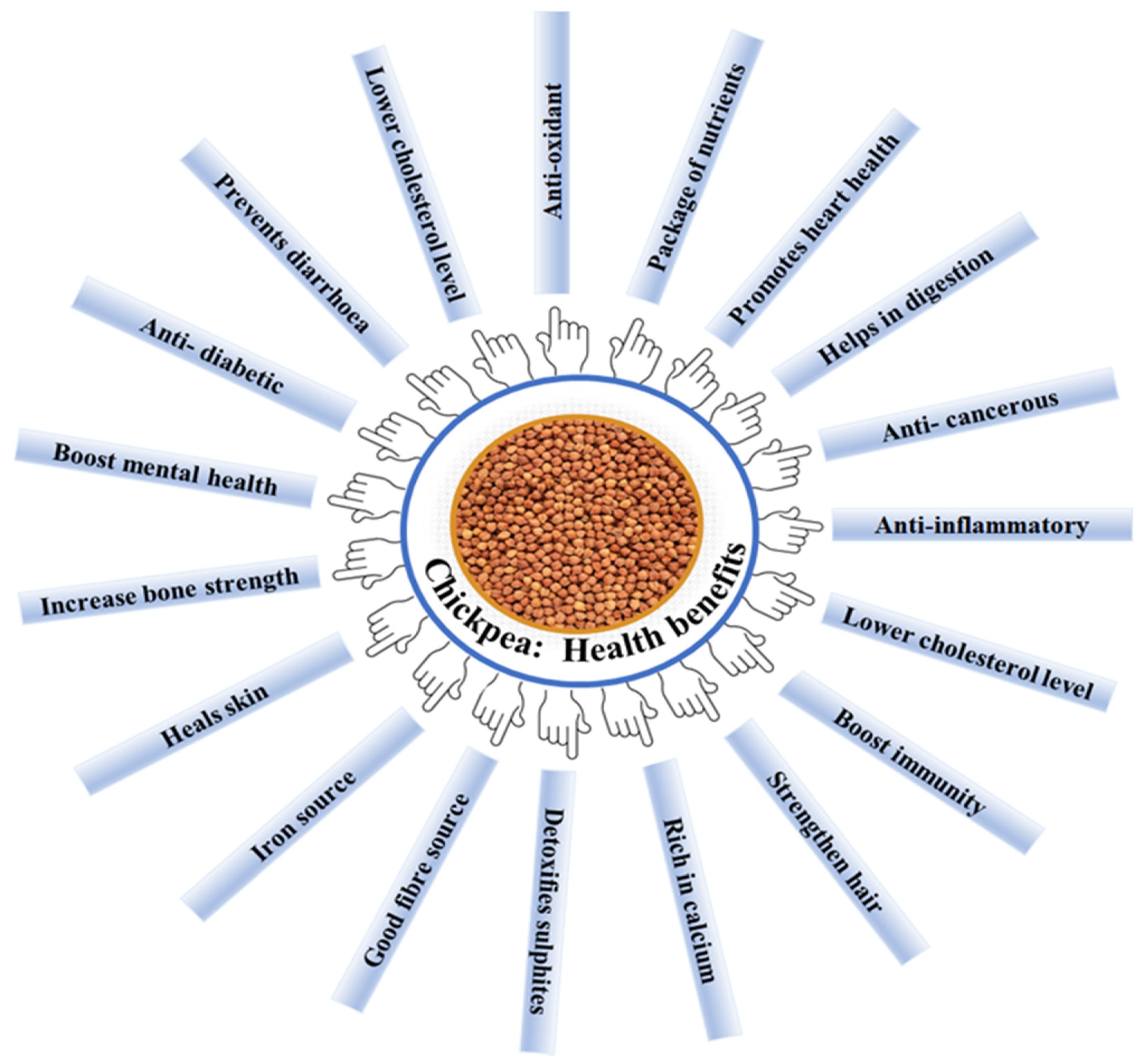
1.4. Pharmacological Characteristics
1.5. Genomic Analysis
1.6. Mapping Populations
1.7. Molecular Markers
1.8. Genome Mapping
1.9. QTL Analysis
1.10. Marker-Assisted Breeding (MAB)
2. Abiotic and Biotic Constraints to Chickpea Production
Crop Improvement through Transformation Regime
3. Biofortification
3.1. Foliar Method
3.2. Microbial Treatment
4. Constraints in the Development of Transgenic Chickpea
5. Industrial Application of Chickpea
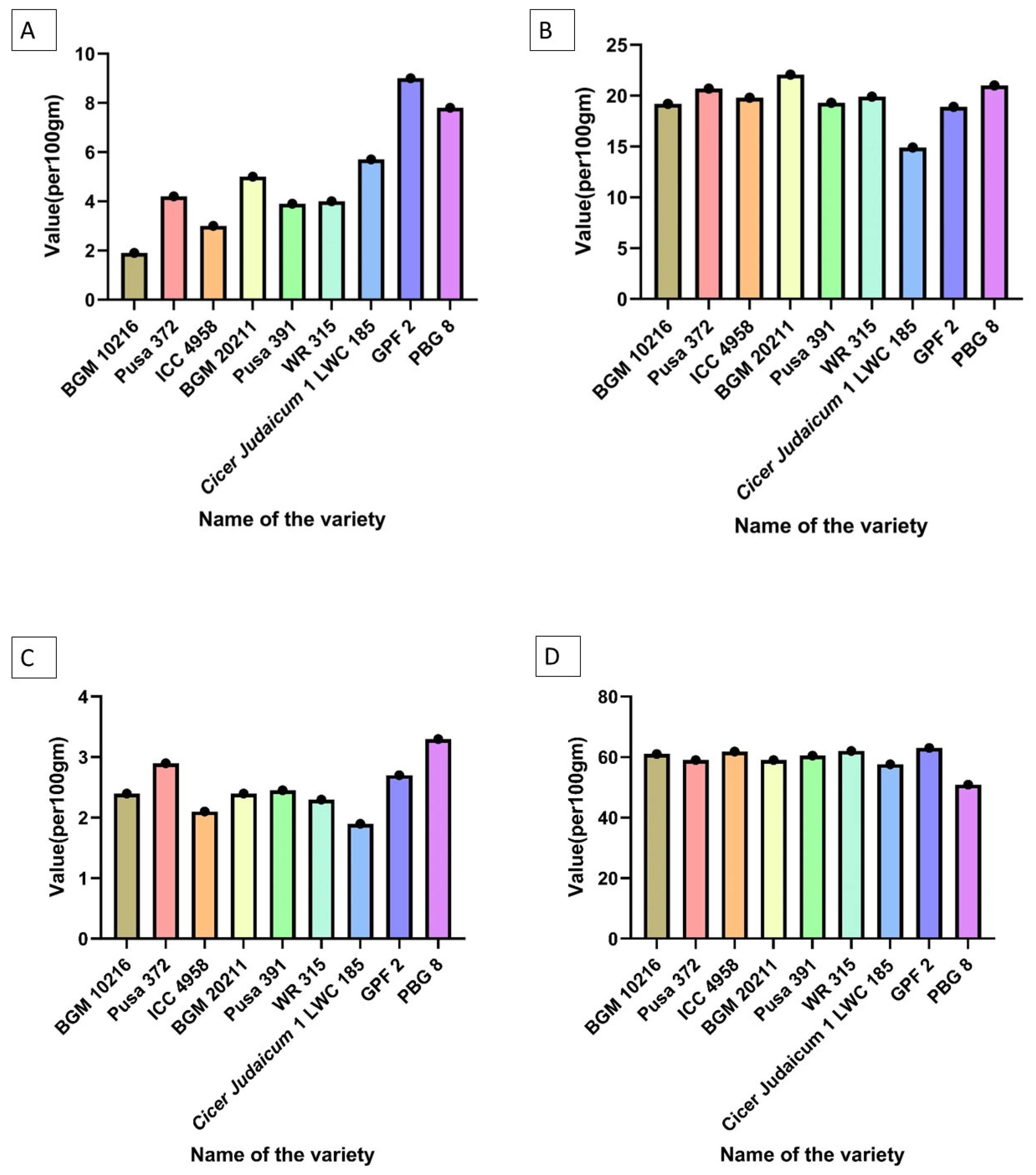
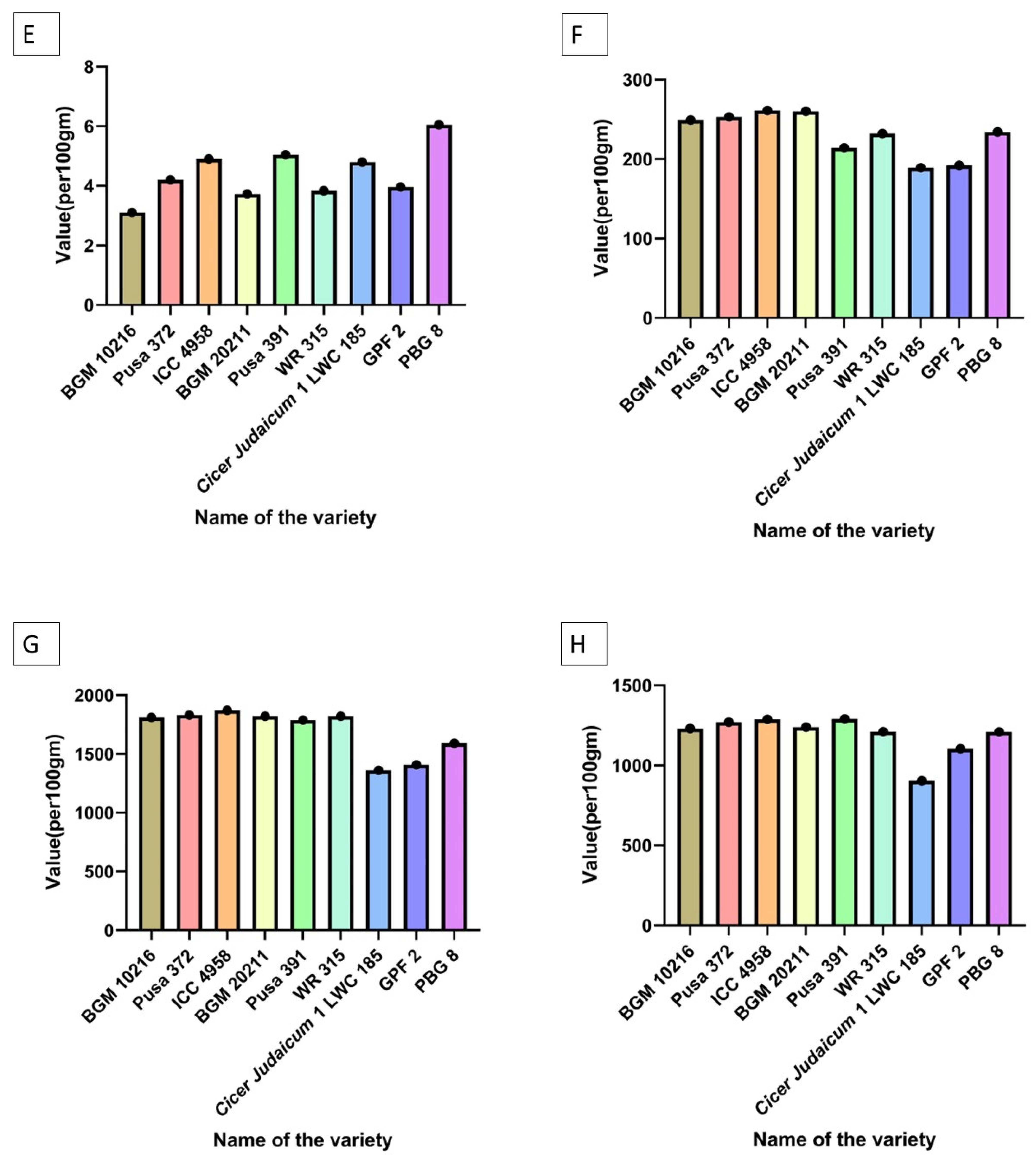
6. Characterization of Chickpea Varieties
7. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaur, P.M.; Tripathi, S.; Gowda, C.L.; Ranga Rao, G.; Sharma, H.; Pande, S.; Sharma, M. Chickpea Seed Production Manual; International Crops Research Institute for the Semi-Arid Tropics (ICRISAT): Patancheru, Andhra Pradesh, India, 2010. [Google Scholar]
- Rasheed, A.; Gill, R.A.; Hassan, M.U.; Mahmood, A.; Qari, S.; Zaman, Q.U.; Ilyas, M.; Aamer, M.; Batool, M.; Li, H. A critical review: Recent advancements in the use of CRISPR/Cas9 technology to enhance crops and alleviate global food crises. Curr. Issues Mol. Biol. 2021, 43, 1950–1976. [Google Scholar] [CrossRef] [PubMed]
- Grasso, N.; Lynch, N.L.; Arendt, E.K.; O’Mahony, J.A. Chickpea protein ingredients: A review of composition, functionality, and applications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Ladizinsky, G.; Adler, A. The origin of chickpea Cicer arietinum L. Euphytica 1976, 25, 211–217. [Google Scholar] [CrossRef]
- Acharjee, S.; Sarmah, B.K. Biotechnologically generating ‘super chickpea’ for food and nutritional security. Plant Sci. 2013, 207, 108–116. [Google Scholar] [CrossRef]
- Roorkiwal, M.; Bharadwaj, C.; Barmukh, R.; Dixit, G.P.; Thudi, M.; Gaur, P.M.; Chaturvedi, S.K.; Fikre, A.; Hamwieh, A.; Kumar, S. Integrating genomics for chickpea improvement: Achievements and opportunities. Theor. Appl. Genet. 2020, 133, 1703–1720. [Google Scholar] [CrossRef] [Green Version]
- Benkadri, S.; Salvador, A.; Zidoune, M.N.; Sanz, T. Gluten-free biscuits based on composite rice—Chickpea flour and xanthan gum. Food Sci. Technol. Int. 2018, 24, 607–616. [Google Scholar] [CrossRef]
- Kushwaha, P.; Srivastava, R.; Pandiyan, K.; Singh, A.; Chakdar, H.; Kashyap, P.L.; Bhardwaj, A.K.; Murugan, K.; Karthikeyan, N.; Bagul, S.Y. Enhancement in plant growth and zinc biofortification of chickpea (Cicer arietinum L.) by Bacillus altitudinis. J. Soil Sci. Plant Nutr. 2021, 21, 922–935. [Google Scholar] [CrossRef]
- Biradar, S.S.; Sridevi, O.; Salimath, P. Genetic enhancement of chickpea for pod borer resistance through expression of CryIAc protein. Karnataka J. Agric. Sci. 2009, 22, 467–470. [Google Scholar]
- Mehrotra, M.; Singh, A.K.; Sanyal, I.; Altosaar, I.; Amla, D. Pyramiding of modified cry1Ab and cry1Ac genes of Bacillus thuringiensis in transgenic chickpea (Cicer arietinum L.) for improved resistance to pod borer insect Helicoverpa armigera. Euphytica 2011, 182, 87–102. [Google Scholar] [CrossRef]
- Summerfield, R.; Roberts, E. Photo-thermal regulation of flowering in pea, lentil, faba bean and chickpea. In World Crops: Cool Season Food Legumes; Springer: Cham, Switzerland, 1988; pp. 911–922. [Google Scholar]
- Yadav, S.; Longnecker, N.; Dusunceli, F.; Bejiga, G.; Yadav, M.; Rizvi, A.; Manohar, M.; Reddy, A.; Xaxiao, Z.; Chen, W. Uses, consumption and utilization. In Chickpea Breeding and Management; CAB International: Oxfordshire, UK, 2007; pp. 101–142. [Google Scholar]
- Jain, M.; Misra, G.; Patel, R.K.; Priya, P.; Jhanwar, S.; Khan, A.W.; Shah, N.; Singh, V.K.; Garg, R.; Jeena, G. A draft genome sequence of the pulse crop chickpea (Cicer arietinum L.). Plant J. 2013, 74, 715–729. [Google Scholar] [CrossRef]
- López-Bellido, F.J.; López-Bellido, R.J.; Khalil, S.K.; López-Bellido, L. Effect of planting date on winter kabuli chickpea growth and yield under rainfed Mediterranean conditions. Agron. J. 2008, 100, 957–964. [Google Scholar] [CrossRef]
- De Santis, M.A.; Rinaldi, M.; Menga, V.; Codianni, P.; Giuzio, L.; Fares, C.; Flagella, Z. Influence of organic and conventional farming on grain yield and protein composition of chickpea genotypes. Agronomy 2021, 11, 191. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, N. Studies on functional, thermal and pasting properties of flours from different chickpea (Cicer arietinum L.) cultivars. Food Chem. 2005, 91, 403–411. [Google Scholar] [CrossRef]
- Singh, M.; Bisht, I.S.; Dutta, M. Broadening the Genetic Base of Grain Legumes; Springer: Cham, Switzerland, 2014. [Google Scholar]
- Heuzé, V.; Tran, G.; Boudon, A.; Bastianelli, D.; Lebas, F. Chickpea (Cicer Arietinum). 2015. Available online: https://www.feedipedia.org/node/319 (accessed on 9 December 2016).
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyimu, X.G.; Bozlar, M.A.; Wulamujiang, A. Pharmacology properties of Cicer arietinum L. Int. J. ChemTech Res. 2020, 13, 251–256. [Google Scholar] [CrossRef]
- Faridy, J.-C.M.; Stephanie, C.-G.M.; Gabriela, M.-M.O.; Cristian, J.-M. Biological activities of chickpea in human health (Cicer arietinum L.). A review. Plant Foods Hum. Nutr. 2020, 75, 142–153. [Google Scholar] [CrossRef]
- Guillon, F.; Champ, M.-J. Carbohydrate fractions of legumes: Uses in human nutrition and potential for health. Br. J. Nutr. 2002, 88, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Barmukh, R.; Soren, K.R.; Madugula, P.; Gangwar, P.; Shanmugavadivel, P.; Bharadwaj, C.; Konda, A.K.; Chaturvedi, S.K.; Bhandari, A.; Rajain, K. Construction of a high-density genetic map and QTL analysis for yield, yield components and agronomic traits in chickpea (Cicer arietinum L.). PLoS ONE 2021, 16, e0251669. [Google Scholar] [CrossRef]
- Gaur, P.M.; Jukanti, A.K.; Varshney, R.K. Impact of genomic technologies on chickpea breeding strategies. Agronomy 2012, 2, 199–221. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C.; Nayyar, H.; Palakurthi, R.; Jha, R.; Valluri, V.; Bajaj, P.; Chitikineni, A.; Singh, N.P.; Varshney, R.K.; Thudi, M. Major QTLs and potential candidate genes for heat stress tolerance identified in chickpea (Cicer arietinum L.). Front. Plant Sci. 2021, 12, 655103. [Google Scholar] [CrossRef]
- Varshney, R.K.; Roorkiwal, M.; Sun, S.; Bajaj, P.; Chitikineni, A.; Thudi, M.; Singh, N.P.; Du, X.; Upadhyaya, H.D.; Khan, A.W. A chickpea genetic variation map based on the sequencing of 3366 genomes. Nature 2021, 599, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, H.D.; Thudi, M.; Dronavalli, N.; Gujaria, N.; Singh, S.; Sharma, S.; Varshney, R.K. Genomic tools and germplasm diversity for chickpea improvement. Plant Genet. Resour. 2011, 9, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, C.; Sachdeva, S.; Singh, R.K.; Patil, B.; Roorkiwal, M.; Chaturvedi, S.; Varshney, R. Chickpea Genomics. In Biotechnologies of Crop Improvement; Springer: Cham, Switzerland, 2018; Volume 3, pp. 289–316. [Google Scholar]
- Nguyen, D.T.; Hayes, J.E.; Atieno, J.; Li, Y.; Baumann, U.; Pattison, A.; Bramley, H.; Hobson, K.; Roorkiwal, M.; Varshney, R.K. The genetics of vigour-related traits in chickpea (Cicer arietinum L.): Insights from genomic data. Theor. Appl. Genet. 2022, 135, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Gaur, P.M.; Thudi, M.; Samineni, S.; Varshney, R.K. Advances in chickpea genomics. In Legumes in the Omic Era; Springer: Cham, Switzerland, 2014; pp. 73–94. [Google Scholar]
- Thudi, M.; Bohra, A.; Nayak, S.; Varghese, N.; Shah, T.; Penmetsa, R.; Thirunavukkarasu, N.; Gudipati, S.; Gaur, P.; Kulwal, P. Novel SSR Markers from BAC-End Sequences, DArT Arrays and a Comprehensive Genetic Map with 1291 Marker Loci for Chickpea (Cicer arietinum L.). PLoS ONE 2011, 6, e27275. [Google Scholar] [CrossRef] [Green Version]
- Madrid, E.; Seoane, P.; Claros, M.; Barro, F.; Rubio, J.; Gil, J.; Millán, T. Genetic and physical mapping of the QTLAR3 controlling blight resistance in chickpea (Cicer arietinum L). Euphytica 2014, 198, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C. Current advances in chickpea genomics: Applications and future perspectives. Plant Cell Rep. 2018, 37, 947–965. [Google Scholar] [CrossRef]
- Lichtenzveig, J.; Scheuring, C.; Dodge, J.; Abbo, S.; Zhang, H.-B. Construction of BAC and BIBAC libraries and their applications for generation of SSR markers for genome analysis of chickpea, Cicer arietinum L. Theor. Appl. Genet. 2005, 110, 492–510. [Google Scholar] [CrossRef]
- Coram, T.E.; Pang, E.C. Isolation and analysis of candidate Ascochyta blight defence genes in chickpea. Part I. Generation and analysis of an expressed sequence tag (EST) library. Physiol. Mol. Plant Pathol. 2005, 66, 192–200. [Google Scholar] [CrossRef]
- Hiremath, P.J.; Farmer, A.; Cannon, S.B.; Woodward, J.; Kudapa, H.; Tuteja, R.; Kumar, A.; BhanuPrakash, A.; Mulaosmanovic, B.; Gujaria, N. Large-scale transcriptome analysis in chickpea (Cicer arietinum L.), an orphan legume crop of the semi-arid tropics of Asia and Africa. Plant Biotechnol. J. 2011, 9, 922–931. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.N.; Zhu, H.; Varghese, N.; Datta, S.; Choi, H.-K.; Horres, R.; Jüngling, R.; Singh, J.; Kavi Kishor, P.; Sivaramakrishnan, S. Integration of novel SSR and gene-based SNP marker loci in the chickpea genetic map and establishment of new anchor points with Medicago truncatula genome. Theor. Appl. Genet. 2010, 120, 1415–1441. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G.; Cannon, S.; Baek, J.; Rosen, B.D.; Tar’an, B. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katna, G.; Nitesh, S.; Sharma, K.D. Conventional cytogenetic manipulations. In Chickpea: Crop Wild Relatives for Enhancing Genetic Gains; Elsevier: Amsterdam, The Netherlands, 2020; pp. 63–93. [Google Scholar]
- Gujaria, N.; Kumar, A.; Dauthal, P.; Dubey, A.; Hiremath, P.; Bhanu Prakash, A.; Farmer, A.; Bhide, M.; Shah, T.; Gaur, P.M. Development and use of genic molecular markers (GMMs) for construction of a transcript map of chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2011, 122, 1577–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srungarapu, R.; Mahendrakar, M.D.; Mohammad, L.A.; Chand, U.; Jagarlamudi, V.R.; Kondamudi, K.P.; Kudapa, H.; Samineni, S. Genome-Wide Association Analysis Reveals Trait-Linked Markers for Grain Nutrient and Agronomic Traits in Diverse Set of Chickpea Germplasm. Cells 2022, 11, 2457. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Alsamman, A.M.; Jighly, A.; Mubarak, M.H.; Al-Shamaa, K.; Istanbuli, T.; Momtaz, O.A.; El Allali, A.; Hamwieh, A. Genome-wide association analysis of chickpea germplasms differing for salinity tolerance based on DArTseq markers. PLoS ONE 2021, 16, e0260709. [Google Scholar] [CrossRef]
- Gaur, P.; Slinkard, A. Genetic control and linkage relations of additional isozyme markers in chick-pea. Theor. Appl. Genet. 1990, 80, 648–656. [Google Scholar] [CrossRef]
- Winter, P.; Benko-Iseppon, A.-M.; Hüttel, B.; Ratnaparkhe, M.; Tullu, A.; Sonnante, G.; Pfaff, T.; Tekeoglu, M.; Santra, D.; Sant, V. A linkage map of the chickpea (Cicer arietinum L.) genome based on recombinant inbred lines from a C. arietinum × C. reticulatum cross: Localization of resistance genes for fusarium wilt races 4 and 5. Theor. Appl. Genet. 2000, 101, 1155–1163. [Google Scholar] [CrossRef]
- Simon, C.; Muehibauer, F. Construction of a chickpea linkage map and its comparison with maps of pea and lentil. J. Hered. 1997, 88, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Anuradha, C.; Gaur, P.M.; Pande, S.; Gali, K.K.; Ganesh, M.; Kumar, J.; Varshney, R.K. Mapping QTL for resistance to botrytis grey mould in chickpea. Euphytica 2011, 182, 1–9. [Google Scholar] [CrossRef]
- Garg, V.; Kaushik, P. Influence of short-term irrigation of textile mill wastewater on the growth of chickpea cultivars. Chem. Ecol. 2006, 22, 193–200. [Google Scholar] [CrossRef]
- Millan, T.; Winter, P.; Jüngling, R.; Gil, J.; Rubio, J.; Cho, S.; Cobos, M.; Iruela, M.; Rajesh, P.; Tekeoglu, M. A consensus genetic map of chickpea (Cicer arietinum L.) based on 10 mapping populations. Euphytica 2010, 175, 175–189. [Google Scholar] [CrossRef]
- Rajesh, P.; Coyne, C.; Meksem, K.; Sharma, K.; Gupta, V.; Muehlbauer, F. Construction of a HindIII bacterial artificial chromosome library and its use in identification of clones associated with disease resistance in chickpea. Theor. Appl. Genet. 2004, 108, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Scheuring, C.F.; Zhang, M.; Dong, J.J.; Zhang, Y.; Huang, J.J.; Lee, M.-K.; Abbo, S.; Sherman, A.; Shtienberg, D. A BAC/BIBAC-based physical map of chickpea, Cicer arietinum L. BMC Genom. 2010, 11, 501. [Google Scholar] [CrossRef] [Green Version]
- Mazumdar, D.; Saha, S.P.; Ghosh, S. Isolation, screening and application of a potent PGPR for enhancing growth of Chickpea as affected by nitrogen level. Int. J. Veg. Sci. 2020, 26, 333–350. [Google Scholar] [CrossRef]
- Tekeoglu, M.; Santra, D.K.; Kaiser, W.J.; Muehlbauer, F.J. Ascochyta blight resistance inheritance in three chickpea recombinant inbred line populations. Crop Sci. 2000, 40, 1251–1256. [Google Scholar] [CrossRef]
- Rakshit, S.; Winter, P.; Tekeoglu, M.; Juarez Muñoz, J.; Pfaff, T.; Benko-Iseppon, A.-M.; Muehlbauer, F.; Kahl, G. DAF marker tightly linked to a major locus for Ascochyta blight resistance in chickpea (Cicer arietinum L.). Euphytica 2003, 132, 23–30. [Google Scholar] [CrossRef]
- Flandez-Galvez, H.; Ford, R.; Pang, E.; Taylor, P. An intraspecific linkage map of the chickpea (Cicer arietinum L.) genome based on sequence tagged microsatellite site and resistance gene analog markers. Theor. Appl. Genet. 2003, 106, 1447–1456. [Google Scholar] [CrossRef]
- Udupa, S.M.; Baum, M. Genetic dissection of pathotype-specific resistance to ascochyta blight disease in chickpea (Cicer arietinum L.) using microsatellite markers. Theor. Appl. Genet. 2003, 106, 1196–1202. [Google Scholar] [CrossRef]
- Collard, B.C.; Pang, E.C.; Ades, P.K.; Taylor, P.W. Preliminary investigation of QTLs associated with seedling resistance to ascochyta blight from Cicer echinospermum, a wild relative of chickpea. Theor. Appl. Genet. 2003, 107, 719–729. [Google Scholar] [CrossRef]
- Cho, S.; Chen, W.; Muehlbauer, F.J. Pathotype-specific genetic factors in chickpea (Cicer arietinum L.) for quantitative resistance to ascochyta blight. Theor. Appl. Genet. 2004, 109, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Millan, T.; Rubio, J.; Iruela, M.; Daly, K.; Cubero, J.; Gil, J. Markers associated with Ascochyta blight resistance in chickpea and their potential in marker-assisted selection. Field Crops Res. 2003, 84, 373–384. [Google Scholar] [CrossRef]
- Iruela, M.; Rubio, J.; Cubero, J.; Gil, J.; Millan, T. Phylogenetic analysis in the genus Cicer and cultivated chickpea using RAPD and ISSR markers. Theor. Appl. Genet. 2002, 104, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, A.; Bhatia, D.; Singh, I.; Thudi, M.; Singh, G.; Bindra, S.; Vij, S.; Gill, B.; Bharadwaj, C.; Singh, S. Identification of stable heat tolerance QTLs using inter-specific recombinant inbred line population derived from GPF 2 and ILWC 292. PLoS ONE 2021, 16, e0254957. [Google Scholar] [CrossRef] [PubMed]
- Sudheesh, S.; Kahrood, H.V.; Braich, S.; Dron, N.; Hobson, K.; Cogan, N.O.; Kaur, S. Application of genomics approaches for the improvement in ascochyta blight resistance in chickpea. Agronomy 2021, 11, 1937. [Google Scholar] [CrossRef]
- Sharma, K.D.; Muehlbauer, F.J. Fusarium wilt of chickpea: Physiological specialization, genetics of resistance and resistance gene tagging. Euphytica 2007, 157, 1–14. [Google Scholar] [CrossRef]
- Varshney, R.K.; Thudi, M.; Nayak, S.N.; Gaur, P.M.; Kashiwagi, J.; Krishnamurthy, L.; Jaganathan, D.; Koppolu, J.; Bohra, A.; Tripathi, S. Genetic dissection of drought tolerance in chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2014, 127, 445–462. [Google Scholar] [CrossRef]
- Razzaq, M.K.; Akhter, M.; Ahmad, R.M.; Cheema, K.L.; Hina, A.; Karikari, B.; Raza, G.; Xing, G.; Gai, J.; Khurshid, M. CRISPR-Cas9 based stress tolerance: New hope for abiotic stress tolerance in chickpea (Cicer arietinum). Mol. Biol. Rep. 2022, 49, 8977–8985. [Google Scholar] [CrossRef]
- Amiri, S.R.; Deihimfard, R.; Eyni-Nargeseh, H. Toward dormant seeding of rainfed chickpea as an adaptation strategy to sustain productivity in response to changing climate. Field Crops Res. 2020, 247, 107674. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, V.; Dubey, A.K.; Narayan, S.; Sawant, S.V.; Pande, V.; Shirke, P.A.; Sanyal, I. CAMTA transcription factor enhances salinity and drought tolerance in chickpea (Cicer arietinum L.). Plant Cell Tissue Organ Cult. (PCTOC) 2022, 148, 319–330. [Google Scholar]
- Acharjee, S.; Sarmah, B.K.; Kumar, P.A.; Olsen, K.; Mahon, R.; Moar, W.J.; Moore, A.; Higgins, T. Transgenic chickpeas (Cicer arietinum L.) expressing a sequence-modified cry2Aa gene. Plant Sci. 2010, 178, 333–339. [Google Scholar] [CrossRef]
- Pandey, A.; Yadav, R.; Kumar, S.; Kumar, A.; Shukla, P.; Yadav, A.; Sanyal, I. Expression of the entomotoxic Cocculus hirsutus trypsin inhibitor (ChTI) gene in transgenic chickpea enhances its underlying resistance against the infestation of Helicoverpa armigera and Spodoptera litura. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 146, 41–56. [Google Scholar] [CrossRef]
- Kaiser, W.; Schaad, N.; Mink, G.; Hampton, R. Workshop: Seed pathogens of food legumes. In World Crops: Cool Season Food Legumes; Springer: Cham, Switzerland, 1988; pp. 515–517. [Google Scholar]
- Das, A.; Datta, S.; Thakur, S.; Shukla, A.; Ansari, J.; Sujayanand, G.; Chaturvedi, S.K.; Kumar, P.; Singh, N. Expression of a chimeric gene encoding insecticidal crystal protein Cry1Aabc of Bacillus thuringiensis in chickpea (Cicer arietinum L.) confers resistance to gram pod borer (Helicoverpa armigera Hubner.). Front. Plant Sci. 2017, 8, 1423. [Google Scholar] [CrossRef] [Green Version]
- Ignacimuthu, S.; Prakash, S. Agrobacterium-mediated transformation of chickpea with α-amylase inhibitor gene for insect resistance. J. Biosci. 2006, 31, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Basu, D.; Das, S.; Ramkrishnan, N.A.; Mukherjee, P.; Nayak, P.; Sen, S.K. Expression of cryIA (c) gene of Bacillus thuringiensis in transgenic chickpea plants inhibits development of pod-borer (Heliothis armigera) larvae. Transgenic Res. 1997, 6, 177–185. [Google Scholar] [CrossRef]
- Sanyal, I.; Singh, A.K.; Kaushik, M.; Amla, D.V. Agrobacterium-mediated transformation of chickpea (Cicer arietinum L.) with Bacillus thuringiensis cry1Ac gene for resistance against pod borer insect Helicoverpa armigera. Plant Sci. 2005, 168, 1135–1146. [Google Scholar] [CrossRef]
- Indurker, S.; Misra, H.S.; Eapen, S. Genetic transformation of chickpea (Cicer arietinum L.) with insecticidal crystal protein gene using particle gun bombardment. Plant Cell Rep. 2007, 26, 755–763. [Google Scholar] [CrossRef]
- Chakraborti, D.; Sarkar, A.; Mondal, H.A.; Das, S. Tissue specific expression of potent insecticidal, Allium sativum leaf agglutinin (ASAL) in important pulse crop, chickpea (Cicer arietinum L.) to resist the phloem feeding Aphis craccivora. Transgenic Res. 2009, 18, 529–544. [Google Scholar] [CrossRef]
- Indurker, S.; Misra, H.S.; Eapen, S. Agrobacterium-mediated transformation in chickpea (Cicer arietinum L.) with an insecticidal protein gene: Optimization of different factors. Physiol. Mol. Biol. Plants 2010, 16, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, J.; Sen, S.; Ghosh, P.; Sengupta, A.; Basu, D.; Das, S. Homologous promoter derived constitutive and chloroplast targeted expression of synthetic cry1Ac in transgenic chickpea confers resistance against Helicoverpa armigera. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 125, 521–535. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, M.; Molla, K.A.; Karmakar, S.; Datta, K.; Datta, S.K. Development of pod borer-resistant transgenic chickpea using a pod-specific and a constitutive promoter-driven fused cry1Ab/Ac gene. Theor. Appl. Genet. 2014, 127, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Sawardekar, S.; Katageri, I.; Salimath, P.; Kumar, P.; Kelkar, V. Standardization of in-vitro genetic transformation technique in chickpea (Cicer arietinum L.) for pod-borer resistance. Adv. Agric. Res. Technol. J 2017, 1, 198–205. [Google Scholar]
- Bhatnagar-Mathur, P.; Vadez, V.; Jyostna Devi, M.; Lavanya, M.; Vani, G.; Sharma, K.K. Genetic engineering of chickpea (Cicer arietinum L.) with the P5CSF129A gene for osmoregulation with implications on drought tolerance. Mol. Breed. 2009, 23, 591–606. [Google Scholar] [CrossRef]
- Kiran Kumar Ghanti, S.; Sujata, K.; Kumar, V.; Nataraja Karba, N.; Srinath Rao, M.; Kavi Kishor, P. Heterologous expression of P5CS gene in chickpea enhances salt tolerance without affecting yield. Biol. Plant. 2011, 55, 634–640. [Google Scholar] [CrossRef]
- Anbazhagan, K.; Bhatnagar-Mathur, P.; Sharma, K.K.; Baddam, R.; Kishor, P.K.; Vadez, V. Changes in timing of water uptake and phenology favours yield gain in terminal water stressed chickpea AtDREB1A transgenics. Funct. Plant Biol. 2014, 42, 84–94. [Google Scholar] [CrossRef]
- Mubina, N.; Hoque, M.; Sarker, R. In vitro Regeneration and Over Expression of Pea DNA Helicase 45 (PDH45) Gene into the Local Cultivars of Chickpea (Cicer arietinum L.) through Agrobacterium-mediated Genetic Transformation. Plant Tissue Cult. Biotechnol. 2018, 28, 125–140. [Google Scholar] [CrossRef]
- Hajyzadeh, M.; Turktas, M.; Khawar, K.M.; Unver, T. miR408 overexpression causes increased drought tolerance in chickpea. Gene 2015, 555, 186–193. [Google Scholar] [CrossRef]
- Das, A.; Basu, P.S.; Kumar, M.; Ansari, J.; Shukla, A.; Thakur, S.; Singh, P.; Datta, S.; Chaturvedi, S.K.; Sheshshayee, M. Transgenic chickpea (Cicer arietinum L.) harbouring AtDREB1a are physiologically better adapted to water deficit. BMC Plant Biol. 2021, 21, 39. [Google Scholar] [CrossRef]
- Lande, N.V.; Barua, P.; Gayen, D.; Wardhan, V.; Jeevaraj, T.; Kumar, S.; Chakraborty, S.; Chakraborty, N. Dehydration-responsive chickpea chloroplast protein, CaPDZ1, confers dehydration tolerance by improving photosynthesis. Physiol. Plant. 2022, 174, e13613. [Google Scholar] [CrossRef]
- Tabe, L.; Wirtz, M.; Molvig, L.; Droux, M.; Hell, R. Overexpression of serine acetlytransferase produced large increases in O-acetylserine and free cysteine in developing seeds of a grain legume. J. Exp. Bot. 2010, 61, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Romeis, J.; Sharma, H.C.; Sharma, K.K.; Das, S.; Sarmah, B.K. The potential of transgenic chickpeas for pest control and possible effects on non-target arthropods. Crop Prot. 2004, 23, 923–938. [Google Scholar] [CrossRef] [Green Version]
- Lingappa, S.; Hegde, R. Exploitation of biocontrol potential in the management of insect pests of pulse crops. In Biocontrol Potential and Its Exploitation in Sustainable Agriculture; Springer: Cham, Switzerland, 2001; pp. 321–344. [Google Scholar]
- Lawo, N.; Mahon, R.; Milner, R.; Sarmah, B.; Higgins, T.; Romeis, J. Effectiveness of Bacillus thuringiensis-transgenic chickpeas and the entomopathogenic fungus Metarhizium anisopliae in controlling Helicoverpa armigera (Lepidoptera: Noctuidae). Appl. Environ. Microbiol. 2008, 74, 4381–4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbazhagan, K.; Bhatnagar-Mathur, P.; Vadez, V.; Dumbala, S.R.; Kishor, P.; Sharma, K.K. DREB1A overexpression in transgenic chickpea alters key traits influencing plant water budget across water regimes. Plant Cell Rep. 2015, 34, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Pathak, G.C.; Gupta, B.; Pandey, N. Improving reproductive efficiency of chickpea by foliar application of zinc. Braz. J. Plant Physiol. 2012, 24, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Laus, M.F.; Vales, L.D.M.F.; Costa, T.M.B.; Almeida, S.S. Early postnatal protein-calorie malnutrition and cognition: A review of human and animal studies. Int. J. Environ. Res. Public Health 2011, 8, 590–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cynober, L.; Bier, D.M.; Stover, P.; Kadowaki, M.; Morris, S.M., Jr.; Elango, R.; Smriga, M. Proposals for upper limits of safe intake for methionine, histidine, and lysine in healthy humans. J. Nutr. 2020, 150, 2606S–2608S. [Google Scholar] [CrossRef]
- Pellegrino, E.; Bedini, S. Enhancing ecosystem services in sustainable agriculture: Biofertilization and biofortification of chickpea (Cicer arietinum L.) by arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2014, 68, 429–439. [Google Scholar] [CrossRef]
- Ullah, A.; Farooq, M.; Hussain, M. Improving the productivity, profitability and grain quality of kabuli chickpea with co-application of zinc and endophyte bacteria Enterobacter sp. MN17. Arch. Agron. Soil Sci. 2020, 66, 897–912. [Google Scholar] [CrossRef]
- Shivay, Y.S.; Prasad, R.; Pal, M. Effects of source and method of zinc application on yield, zinc biofortification of grain, and Zn uptake and use efficiency in chickpea (Cicer arietinum L.). Commun. Soil Sci. Plant Anal. 2015, 46, 2191–2200. [Google Scholar] [CrossRef]
- Ullah, A.; Farooq, M.; Nadeem, F.; Rehman, A.; Hussain, M.; Nawaz, A.; Naveed, M. Zinc application in combination with zinc solubilizing Enterobacter sp. MN17 improved productivity, profitability, zinc efficiency, and quality of desi chickpea. J. Soil Sci. Plant Nutr. 2020, 20, 2133–2144. [Google Scholar] [CrossRef]
- Pal, V.; Singh, G.; Dhaliwal, S. Agronomic biofortification of chickpea with zinc and iron through application of zinc and urea. Commun. Soil Sci. Plant Anal. 2019, 50, 1864–1877. [Google Scholar] [CrossRef]
- Ullah, A.; Farooq, M.; Nadeem, F.; Rehman, A.; Nawaz, A.; Naveed, M.; Wakeel, A.; Hussain, M. Zinc seed treatments improve productivity, quality and grain biofortification of desi and kabuli chickpea (Cicer arietinum). Crop Pasture Sci. 2020, 71, 668–678. [Google Scholar] [CrossRef]
- Poblaciones, M.J.; Rodrigo, S.; Santamaria, O.; Chen, Y.; McGrath, S.P. Selenium accumulation and speciation in biofortified chickpea (Cicer arietinum L.) under Mediterranean conditions. J. Sci. Food Agric. 2014, 94, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Rathod, S.; Channakeshava, S.; Basavaraja, B.; Shashidhara, K. Effect of soil and foliar application of zinc and Boron on growth, yield and micro nutrient uptake of Chickpea. J. Pharmacogn. Phytochem. 2020, 9, 3356–3360. [Google Scholar]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Verma, V.; Behera, S.K.; Singh, P.; Alotaibi, S.S.; Gaber, A.; Hossain, A. Comparative efficiency of mineral, chelated and nano forms of zinc and iron for improvement of zinc and iron in chickpea (Cicer arietinum L.) through biofortification. Agronomy 2021, 11, 2436. [Google Scholar] [CrossRef]
- Saira, K.; Asghar, H.N.; Akhtar, M.J.; Ana, A.; Zahir, Z.A. Biofortification of iron in chickpea by plant growth promoting rhizobacteria. Pak. J. Bot. 2015, 47, 1191–1194. [Google Scholar]
- Verma, J.; Yadav, J.; Tiwari, K.N. Application of Rhizobium sp. BHURC01 and plant growth promoting rhizobacteria on nodulation, plant biomass and yields of chickpea (Cicer arietinum L.). Int. J. Agric. Res. 2010, 5, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Mehboob, N.; Yasir, T.A.; Ul-Allah, S.; Nawaz, A.; Ahmad, N.; Hussain, M. Interactive Effect of Boron Application Methods and Boron-Tolerant Bacteria (Bacillus sp. MN54) Improves Nodulation, Grain Yield, Profitability and Biofortification of kabuli Chickpea Grown Under Irrigated and Rainfed Conditions. J. Soil Sci. Plant Nutr. 2022, 1–18. [Google Scholar] [CrossRef]
- Batool, S.; Asghar, H.N.; Shehzad, M.A.; Yasin, S.; Sohaib, M.; Nawaz, F.; Akhtar, G.; Mubeen, K.; Zahir, Z.A.; Uzair, M. Zinc-solubilizing bacteria-mediated enzymatic and physiological regulations confer zinc biofortification in chickpea (Cicer arietinum L.). J. Soil Sci. Plant Nutr. 2021, 21, 2456–2471. [Google Scholar] [CrossRef]
- Yasmin, R.; Hussain, S.; Rasool, M.H.; Siddique, M.H.; Muzammil, S. Isolation, characterization of Zn solubilizing bacterium (Pseudomonas protegens RY2) and its contribution in growth of chickpea (Cicer arietinum L) as deciphered by improved growth parameters and Zn content. Dose Response 2021, 19, 15593258211036791. [Google Scholar] [CrossRef]
- Hussain, M.; Mehboob, N.; Naveed, M.; Shehzadi, K.; Yasir, T.A. Optimizing boron seed coating level and boron-tolerant bacteria for improving yield and biofortification of chickpea. J. Soil Sci. Plant Nutr. 2020, 20, 2471–2478. [Google Scholar] [CrossRef]
- Yadav, R.; Mehrotra, M.; Singh, A.K.; Niranjan, A.; Singh, R.; Sanyal, I.; Lehri, A.; Pande, V.; Amla, D. Improvement in Agrobacterium-mediated transformation of chickpea (Cicer arietinum L.) by the inhibition of polyphenolics released during wounding of cotyledonary node explants. Protoplasma 2017, 254, 253–269. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrients | Nutrient Value Per 100 g |
|---|---|
| Calories | 378–396 |
| Protein (g) | 18.77–24 |
| Fat (g) | 4.1–6.04 |
| Carbohydrate (g) | 39.56–54.2 |
| Fiber (g) | 7.4–12.22 |
| Ash (g) | 3.4 |
| Minerals | |
| Ca (mg) | 57–160 |
| P (mg) | 250–310 |
| Fe (mg) | 4.0–12.3 |
| Na (mg) | 24 |
| K (mg) | 700–718 |
| Zn (mg) | 2.76–4.1 |
| Mg (mg) | 79–138 |
| Vitamins | |
| β-carotene (μg) | 67 |
| Thiamine (mg) | 0.45–0.5 |
| Riboflavin (mg) | 0.2–0.26 |
| Niacin (mg) | 1.54–2 |
| Tocopherol (mg) | 11.2–12.9 |
| Folic acid (mg) | 206–290 |
| Pantothenic acid (mg) | 1–2 |
| Pyridoxine (mg) | 0.3–0.38 |
| Amino acids | |
| Lysine (g) | 6.6–7.2 |
| Methionine (g) | 1.2–1.4 |
| Cysteine (g) | 0–1 |
| Arginine (g) | 8–8.8 |
| Glycine (g) | 3.5–4 |
| Histidine (g) | 2.3–2.5 |
| Isoleucine (g) | 3.5–4.4 |
| Leucine (g) | 7.1–7.6 |
| Phenylalanine (g) | 5.5–6.6 |
| Tyrosine (g) | 3–3.3 |
| Threonine (g) | 3.4–3.5 |
| Tryptophan (g) | 0–0.9 |
| Valine (g) | 3.9–4.6 |
| Alanine (g) | 3.7–4.1 |
| Aspartic acid (g) | 10–11 |
| Glutamic acid (g) | 16–17 |
| Proline (g) | 4–4.3 |
| Serine (g) | 4.8–5.2 |
| QTL | Marker | Reference(s) |
|---|---|---|
| Ascochyta blightresistance | ||
| 1 | GAA47 | [54] |
| 2 | TA72, GA2 | |
| ar2 | TA72 | [55] |
| TA146 | ||
| I | STMS11, GA2, GAA47, TR20 4 | |
| 1 | TS12b | [56] |
| 2/3 | TA3a/TA3b | |
| 4/5/6 | TA30/TA146/TR20 | |
| QTL-2 | TA3a | |
| TA146 | ||
| QTL-2/QTL-3 | TA72 | |
| GA2 | ||
| TA3a/TA3b | ||
| ar1 | GA16 2 | [57] |
| ar2a | GA16 | |
| ar2b | TA130, TA72, TS72 | |
| ar1b | TA37, TA200 | |
| ar2a | GA24, GAA47 | [58] |
| ar2b | TA130 | |
| TA72 | ||
| TS72 | ||
| ar19 | TR19 | |
| GA16 | ||
| QTL-1 | GAA47 | [54,56,59] |
| TS12b | ||
| STMS28 | ||
| STMS11 | ||
| GA2 | ||
| TS12b | ||
| TR20 | ||
| QTL-3 | TS45 | [57,60,61] |
| TA3b | ||
| TA194 | ||
| TS82 | ||
| TR58 | ||
| ar1a | GA16 | [57,59] |
| GA20 | ||
| ar1b | TA37 | [57] |
| TA200 | ||
| ar2a | GA16 | [59] |
| GA24 | ||
| GAA47 | ||
| ar1 | GA16 | |
| ar1a | GA20, GA16 | |
| qab-4.1, qab-4.2LG7, qab-7.1 | qab-4.1: CNC_021163.1.32280291, CNC_021163.1.37933917 qab-4.2: CNC_021163.1.23799836 CNC_021163.1.24184658 qab-7.1: CNC_021166.1.34330294 CNC_021166.1.34330283 | [62] |
| QTL1 | Ca_Ce_18445 [Ca_Ce_18577 & Ca_Ce_18594] Ca_Ce_18656 | [63] |
| ar2 | SC/OPK13603 4 SC/OPM02935 TA72, TA146 | [60,61] |
| Fusarium wiltresistance | ||
| Foc-0/foc-0 | TR59 | [64] |
| foc-1 | TA59 | |
| TA96 | ||
| TA27 | ||
| foc-2 | TA96 | |
| TA27 | ||
| TR19 | ||
| Foc-3/foc-3 | TA96 | |
| TA27 | ||
| TR59 | ||
| foc-4 | TA59 | |
| TA96 | ||
| TA27 | ||
| TR19 | ||
| TA194 | ||
| Foc-5/foc-5 | TA27 | |
| TA59 | ||
| TA96 | ||
| TA110 | ||
| TA59 | ||
| TA53 | ||
| TA103 | ||
| TS82 | ||
| TR58 | ||
| Foc 1 & 3 | GA 16 | [65] |
| TAA60 | ||
| TA194 | ||
| TS82 | ||
| TA110 | ||
| TR19 | ||
| Gene Transferred | Source of Gene | Transformation Method | Explant | Trait Introduced | Expression Level | Reference(s) |
|---|---|---|---|---|---|---|
| Biotic stress | ||||||
| cryIAc+ nptII+ CaMV35S | Bacillus thuringiensis | A. tumefaciens | Embryo axis | H. armigera resistance | Inhibits the development of Heliothis armigera larvae | [74] |
| cry1Ac + nptII +CaMV35S | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | Cry1Ac protein showed 80–85% protection with high mortality rate i.e., >80% | [75] |
| αAI1+ nptII+ CaMV3S | Phaseolus—vulgaris | A. tumefaciens | Embryogeni c axis | Bruchids resistance | Larval growth reduction | [73] |
| cry1Ac + nptII+ CaMV35S | Bacillus thuringiensis | Particle gun bombardment | Embryonal axis, Epicotyl and stem | Protection from H. armigera and S. litura | Higher mortality of Heliothis armigera and Spodoptera litura larvae | [76] |
| ASAL+ nptII+ CaMV35S + rolC | Allium sativum leaf agglutinin | A. tumefaciens | Single cotyledon with half embryo | Aphis craccivora resistance | Increase in mortality rate upto 42% | [77] |
| CryIAc+ nptII | Bacillus thuringiensis | A. tumefaciens | - | H. armigera resistance | Mortality of >60% for H. armigera | [9] |
| cry2Aa+ nptII + ats1A | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | Showed higher toxicity to the insect. | [69] |
| cryIAc+ nptII+ uidA+ CaMV35S | Bacillus thuringiensis | A. tumefaciens | Embryonic axis, epicotyl and stem explants | H. armigera resistance | Tolerance to infection by H. armigera | [78] |
| cry1Ac+ nptII+ uidA + rbcS+ CaMV35S | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | High level protection against pod borer | [79] |
| cry1Ab and cry1Ac+ CaMV35S or Pcec+ nptII | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | Showed higher mortality of the insect (95%). | [10] |
| cry1Ab/Ac+ actin1+ msg | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | Showed higher toxicity to the Pod borer. | [80] |
| cryIIAa+ nptII + CaMV35S | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | Showed higher toxicity to the insect. | [81] |
| cry1Aabc+ nptII | Bacillus thuringiensis | A. tumefaciens | Cotyledon nodes | H. armigera resistance | Highly effective against pod borer | [72] |
| ChTI+ nptII + CaMV35S+ nos | Cocculus hirsutus | A. tumefaciens | Cotyledon nodes | Protection from H. armigera and S. litura | Showed mortality rate of 60–80% | [70] |
| Abiotic stress | ||||||
| P5CSF129A+ nptII + uidA+ CaMV 35S | Vigna aconitifolia | A. tumefaciens | Axillary meristem | Drought tolerance | Enhanced proline overcame the adverse effects of drought stress | [82] |
| P5CS+ hpt + CaMV35S | Vigna aconitifolia | A. tumefaciens | Cotyledon node | Salt tolerance | Proline overproduction alleviated salt stress | [83] |
| AtDREB1A + rd29A promoter | Arabidopsis thaliana | A. tumefaciens | Axillary meristem | Transpiration efficiency under drought stress | Increased transpiration efficiency | [84] |
| PDH45+ hpt+ CaMV 35S | Pea DNA Helicase 45 | A. tumefaciens | Zygotic embryo, decapitated embryo and decapitated embryo with single cotyledon disc | Salt tolerance | Alleviated salt stress | [85] |
| miR408 (over expression) | Arabidopsis thaliana | Terrestrial plants | Mature embryo | Drought tolerance | Increased drought tolerance | [86] |
| AtDREB1a+ rd29a promoter | Arabidopsis thaliana | A. tumefaciens | Cotyledon with half embryo axis | Drought tolerance | Enhanced drought tolerance | [87] |
| CAMTA (over expression) | Gossypium herbaceum | A. tumefaciens | Cotyledon nodes | Salinity and drought stress | Enhanced activities of antioxidant enzymes under drought and salinity | [68] |
| CaPDZ1 (Over expression) | Cicer arietinum | A. tumefaciens | Single cotyledon with embryo | Dehydration tolerance | Conferred dehydration tolerance by improving photosynthesis | [88] |
| Nutritional enhancement | ||||||
| SSA+ CaMV 35S+ uidA+ pea vicilin gene | Sunflower seed albumin gene (Brassica napus) | A. tumefaciens | Embryo axis | Increased methionine content | Increased methionine content in normal soil state | [69,89] |
| Treatment | Source | Trait Transferred | Expression Level | Reference(s) |
|---|---|---|---|---|
| Foliar application of Zn | ZnSO4.7H2O (33% Zn) | Zn biofortification | Increased Zn content in seeds | [102] |
| Foliar application of Zn | 0.1% ZnSO4 foliar spray | Efficiency of chickpea | Increased Zn content in seeds | [94] |
| Foliar application of Se | Sodium selenate and Sodium selenite at four rates (0, 10, 20, 40 g ha−1) | Se biofortification | Selenomethionine was found in high concentrations in chickpea grains (>70%). | [103] |
| Foliar application of Zn-EDTA | Zn-EDTA three sprays (V + F + G) | Zinc Biofortification | Enrichment of seed with Zn | [99] |
| Foliar application of Zn + urea | ZnSO4 @ 0.5% + @ 2% urea | Biofortification of chickpea with Zn and Fe | Enrichment of seed with Zn and Fe | [101] |
| Foliar application of Zn and Fe | Zn @ 0.5% + Fe @ 0.1% | Biofortification of chickpea with Zn and Fe | Enrichment of chickpeas with Fe and Zn | [104] |
| Foliar application of ZnO NPs + Fe2O3 NPs | 0.5% ZnO NPs + 0.5% Fe2O3 | Biofortification of chickpea with Zn and Fe | Enrichment of chickpeas with Zn and Fe | [105] |
| Treatment | PGPB | Trait Transferred | Expression Level | Reference(s) |
|---|---|---|---|---|
| Zn + PGPB | Enterobacter sp. MN17 | Zn biofortification | Enhanced Zn content in seed | [98,100] |
| Fe + PGPR (plant growth promoting rhizobacteria) | Bacillus cereus UW 85, Azotobacter vinelandi MAC 259, Pseudomonas, Bacillus megaterium, E. coli | Fe biofortification | Enhanced Fe content and 81–75% increase in productivity | [106] |
| Rhizobium sps. BHURC01 + PGPR + Pseudomonas fluorescens | Azotobacter chroococcum, Bacillus megaterium | Plant biomass and yield | Inhibited the phytopathogenic fungi leading to suppression of plant disease, Promotion of plant growth and nodule formation. | [107] |
| Boron coated seed + PGPB | Bacillus sp. MN54 | Boron efficiency | Increased B content, nodulation and yield | [108] |
| Zinc-solubilizing bacteria | ZnSB13 | Zinc biofortifcation in chickpea | Increased Zn content in seeds | [109] |
| Zinc-solubilizing bacteria | B. altitudinis (BT3 and CT8) | Zinc biofortifcation in chickpea | Improved Zn uptake by 3.9–6.0%. | [8] |
| Zinc-solubilizing bacteria | Pseudomonas protegens (RY2, MF351762) | Zinc biofortifcation in chickpea | Enhanced Zn in soil | [110] |
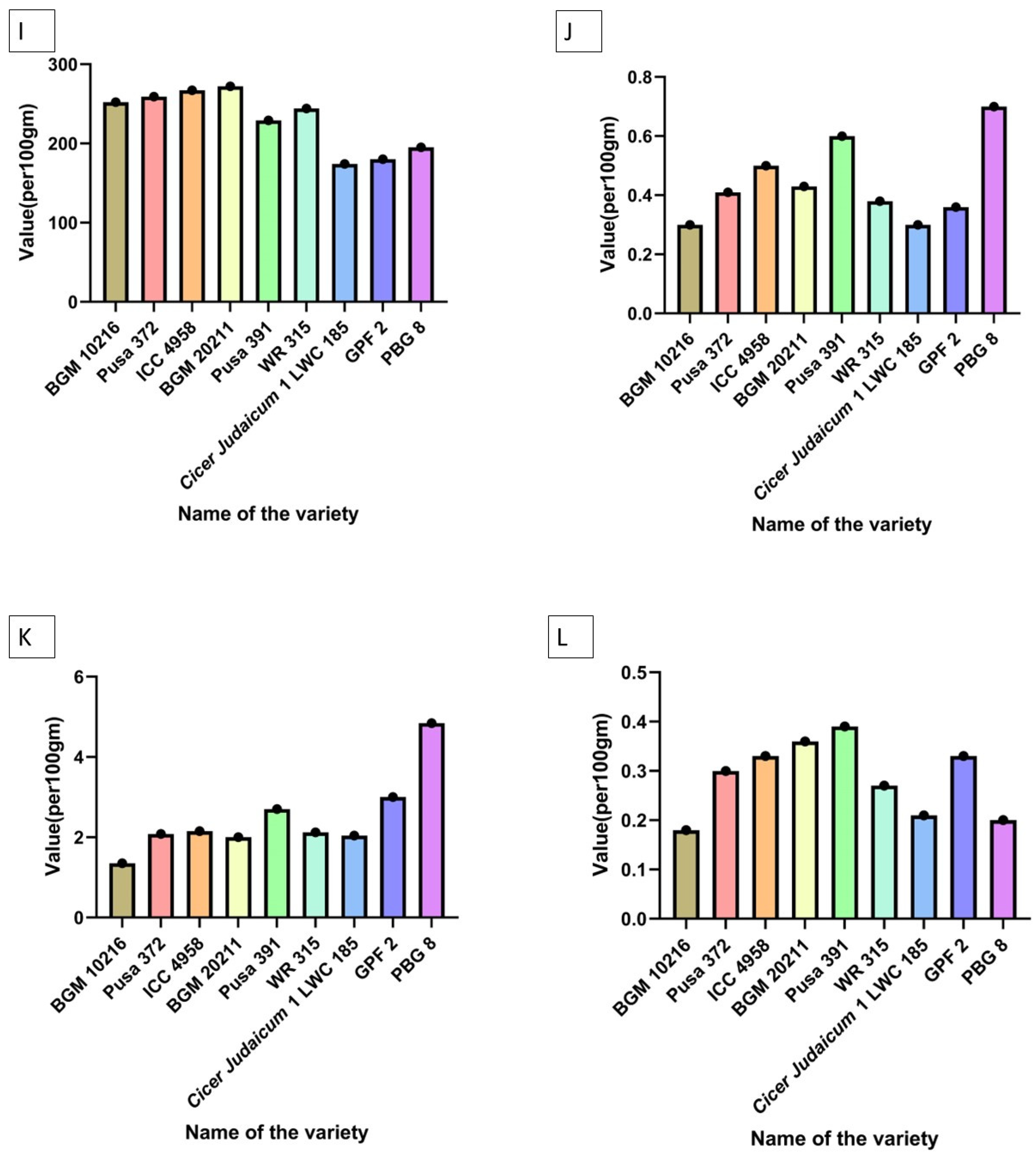
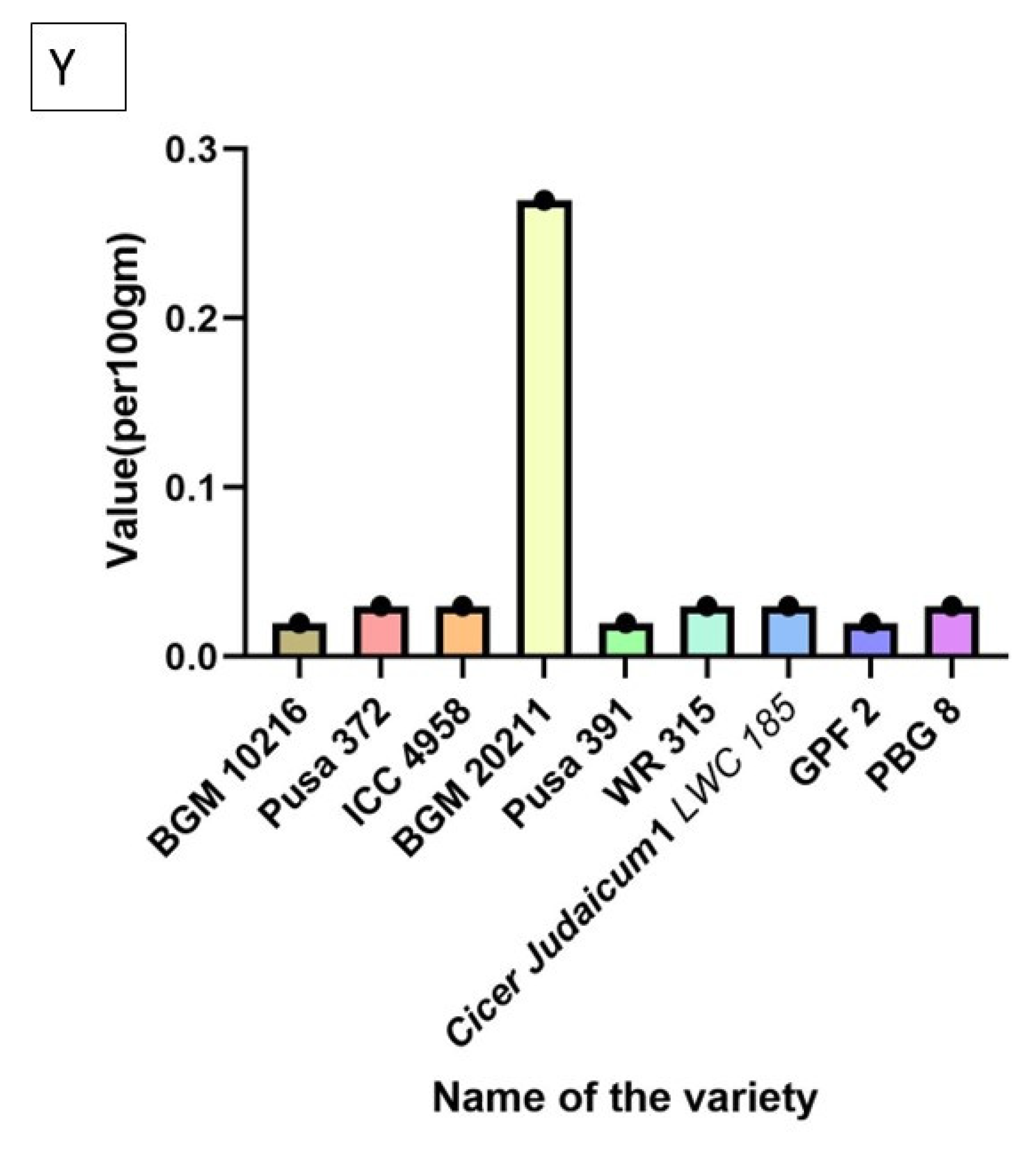
| Parameter Tested | Trend Observed |
|---|---|
| Crude fiber | GPF 2 > PBG 8 > Cicer judaicum 1 LWC 185 > BGM 20211 > Pusa 372 > WR 315 > Pusa 391 > ICC 4958 > BGM 10216 |
| Crude protein | BGM 20211 > PBG 8 > Pusa 372 > WR 315 > ICC 4958 > Pusa 391 > BGM 10216 > GPF 2 > Cicer judaicum 1 LWC 185 |
| Total ash | PBG 8 > Pusa 372 > GPF 2 > Pusa 391 > BGM 20211 = BGM 10216 > WR 315 > ICC 4958 > Cicer judaicum 1 LWC 185 |
| Carbohydrates | GPF 2 > WR 315 > ICC 4958 > BGM 10216 > Pusa 391 > BGM 20211 = Pusa 372 > Cicer judaicum 1 LWC 185 > PBG 8 |
| Crude lipid—fat | PBG 8 > Pusa 391 > ICC 4958 > Cicer judaicum 1 LWC 185 > Pusa 372 > GPF 2 > WR 315 > BGM 20211 > BGM 10216 |
| Methionine | ICC 4958 > BGM 20211 > Pusa 372 > BGM 10216 > PBG 8 > WR 315 > Pusa 391 > GPF 2 > Cicer judaicum 1 LWC 185 |
| Arginine | ICC 4958 > Pusa 372 > BGM 20211 = WR 315 > BGM 10216 > Pusa 391 > PBG 8 > GPF 2 > Cicer judaicum 1 LWC 185 |
| Lysine | Pusa 391 > ICC 4958 > Pusa 372 > BGM 20211 > BGM 10216 > WR 315 > PBG 8 > GPF 2 > Cicer judaicum 1 LWC 185 |
| Cysteine | BGM 20211 > ICC 4958 > Pusa 372 > BGM 10216 > WR 315 > Pusa 391 > PBG 8 > GPF 2 > Cicer judaicum 1 LWC 185 |
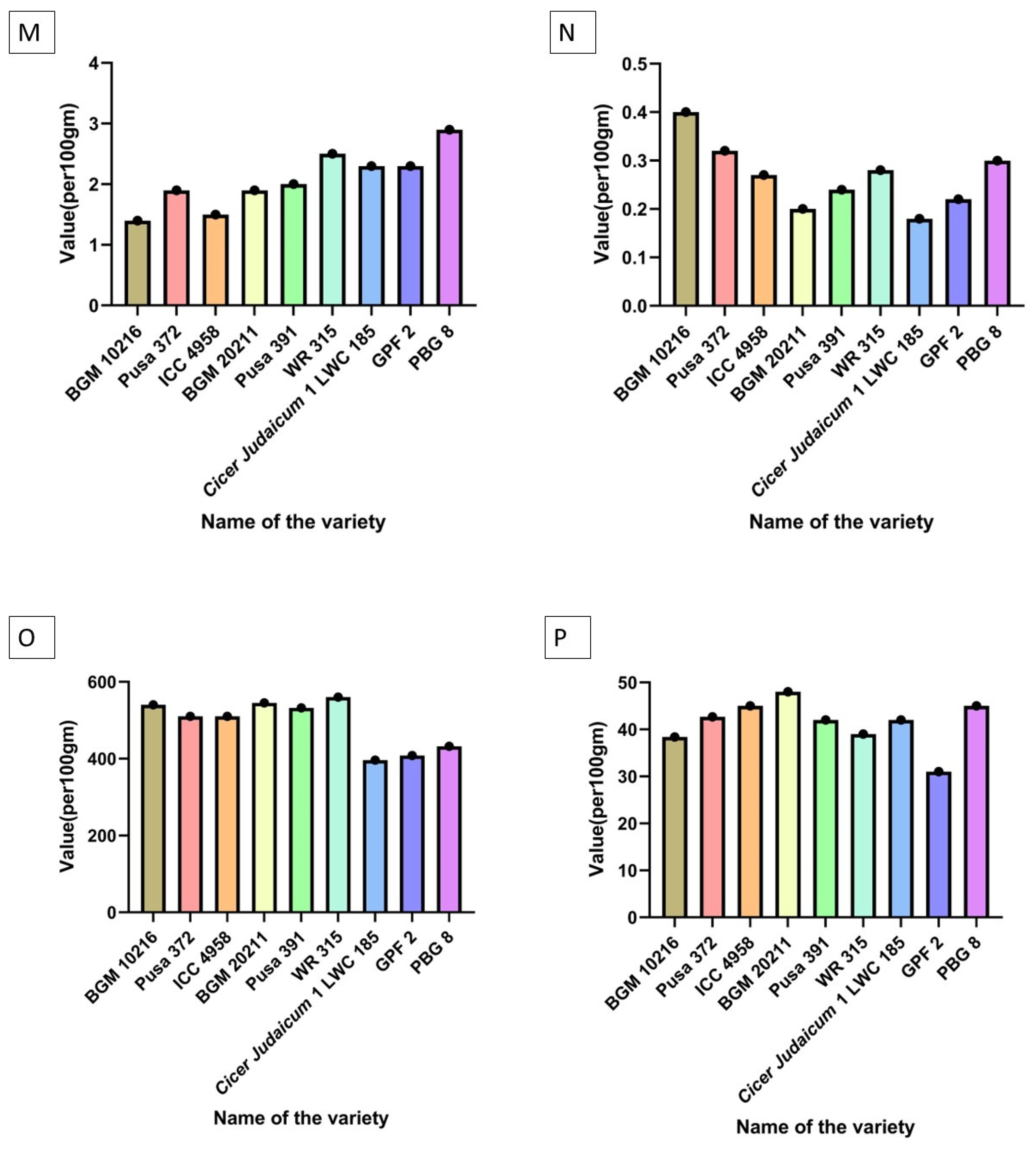
| Saturated fatty acids | PBG 8 > Pusa 391 > ICC 4958 > BGM 20211 > Pusa 372 > WR 315 > GPF 2 > BGM 10216 = Cicer judaicum 1 LWC 185 |
| Polyunsaturated fatty acids | PBG 8 > GPF 2 > Pusa 391 > ICC 4958 > WR 315 > Pusa 372 > Cicer judaicum 1 LWC 185 > BGM 20211 > BGM 10216 |
| Riboflavin | Pusa 391 > BGM 20211 > GPF 2 = ICC 4958 > Pusa 372 > WR 315 > Cicer judaicum 1 LWC 185 > PBG 8 > BGM 10216 |
| Niacin | PBG 8 > WR 315 > Cicer judaicum 1 LWC 185 = GPF 2 > Pusa 391 > BGM 20211 = Pusa 372 > ICC 4958 > BGM 10216 |
| Thiamin | BGM 10216 > Pusa 372 > PBG 8 > WR 315 > ICC 4958 > Pusa 391 > GPF 2 > BGM 20211 > Cicer judaicum 1 LWC 185 |
| Folate | WR 315 > BGM 20211 > BGM 10216 > Pusa 391 > Pusa 372 = ICC 4958 > PBG 8 > GPF 2 > Cicer judaicum 1 LWC 185 |
| B—Carotene (vitamin A) | BGM 20211 > ICC 4958 = PBG 8 > Pusa 372 > Pusa 391 = Cicer judaicum 1 LWC 185 > WR 315 > BGM 10216 > GPF 2 |
| Phenolics | PBG 8 > WR 315 > Pusa 391 > GPF 2 > BGM 10216 > Cicer judaicum 1 LWC 185 > ICC 4958 > BGM 20211 > Pusa 372 |
| Flavanoids | GPF 2 = BGM 10216 > Pusa 391 > ICC 4958 = PBG 8 > WR 315 > Pusa 372 = Cicer judaicum 1 LWC 185 > BGM 20211 |
| Omega 6: Omega 3 | Pusa 391 > BGM 20211 > BGM 10216 > ICC 4958 = WR 315 = PBG 8 > GPF 2 > Pusa 372 > Cicer judaicum 1 LWC 185 |
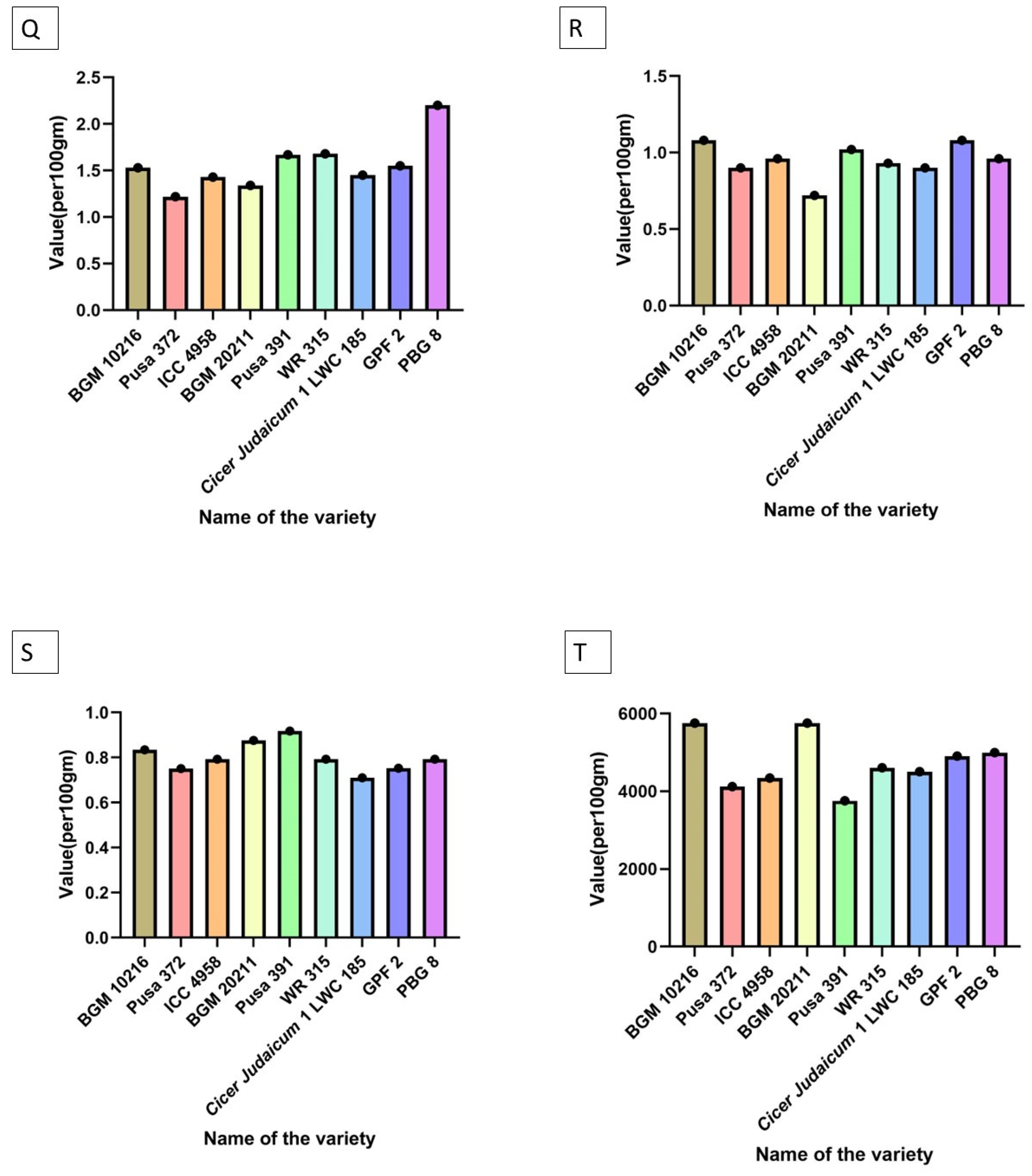
| Antioxidant activity | BGM 10216 = BGM 20211 > PBG 8 > GPF 2 > WR 315 > Cicer judaicum 1 LWC 185 > ICC 4958 > Pusa 372 > Pusa 391 |
| Lectin and hemaglutination assay | PBG 8 > GPF 2 > WR 315 > Pusa 391 > ICC 4958 > Cicer judaicum 1 LWC 185 > BGM 20211 > BGM 10216 > Pusa 372 |
| Uric acid | BGM 20211 > ICC 4958 > BGM 10216 = PBG 8 > GPF 2 > Pusa 372 > Pusa 391 > WR 315 > Cicer judaicum 1 LWC 185 |
| Energy value | ICC 4958 > Pusa 391 = GPF 2 > WR 315 > BGM 20211 > Pusa 372 > BGM 10216 > PBG 8 > Cicer judaicum 1 LWC 185 |
| Protein bioavailability | BGM 20211 > BGM 10216 > Pusa 391 > PBG 8 > WR 315 > GPF 2 > ICC 4958 > Cicer judaicum 1 LWC 185 > Pusa 372 |
| Phytic acid | Pusa 372 = PBG 8 > BGM 20211 = WR 315 > Cicer judaicum 1 LWC 185 = ICC 4958 > GPF 2 > BGM 10216 > Pusa 391 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koul, B.; Sharma, K.; Sehgal, V.; Yadav, D.; Mishra, M.; Bharadwaj, C. Chickpea (Cicer arietinum L.) Biology and Biotechnology: From Domestication to Biofortification and Biopharming. Plants 2022, 11, 2926. https://doi.org/10.3390/plants11212926
Koul B, Sharma K, Sehgal V, Yadav D, Mishra M, Bharadwaj C. Chickpea (Cicer arietinum L.) Biology and Biotechnology: From Domestication to Biofortification and Biopharming. Plants. 2022; 11(21):2926. https://doi.org/10.3390/plants11212926
Chicago/Turabian StyleKoul, Bhupendra, Komal Sharma, Vrinda Sehgal, Dhananjay Yadav, Meerambika Mishra, and Chellapilla Bharadwaj. 2022. "Chickpea (Cicer arietinum L.) Biology and Biotechnology: From Domestication to Biofortification and Biopharming" Plants 11, no. 21: 2926. https://doi.org/10.3390/plants11212926