
Phytochemical Study of Eryngium cymosum F. Delaroche and the Inhibitory Capacity of Its Main Compounds on Two Glucose-Producing Pathway Enzymes

 , ,
, , 
Abstract
:1. Introduction
2. Results
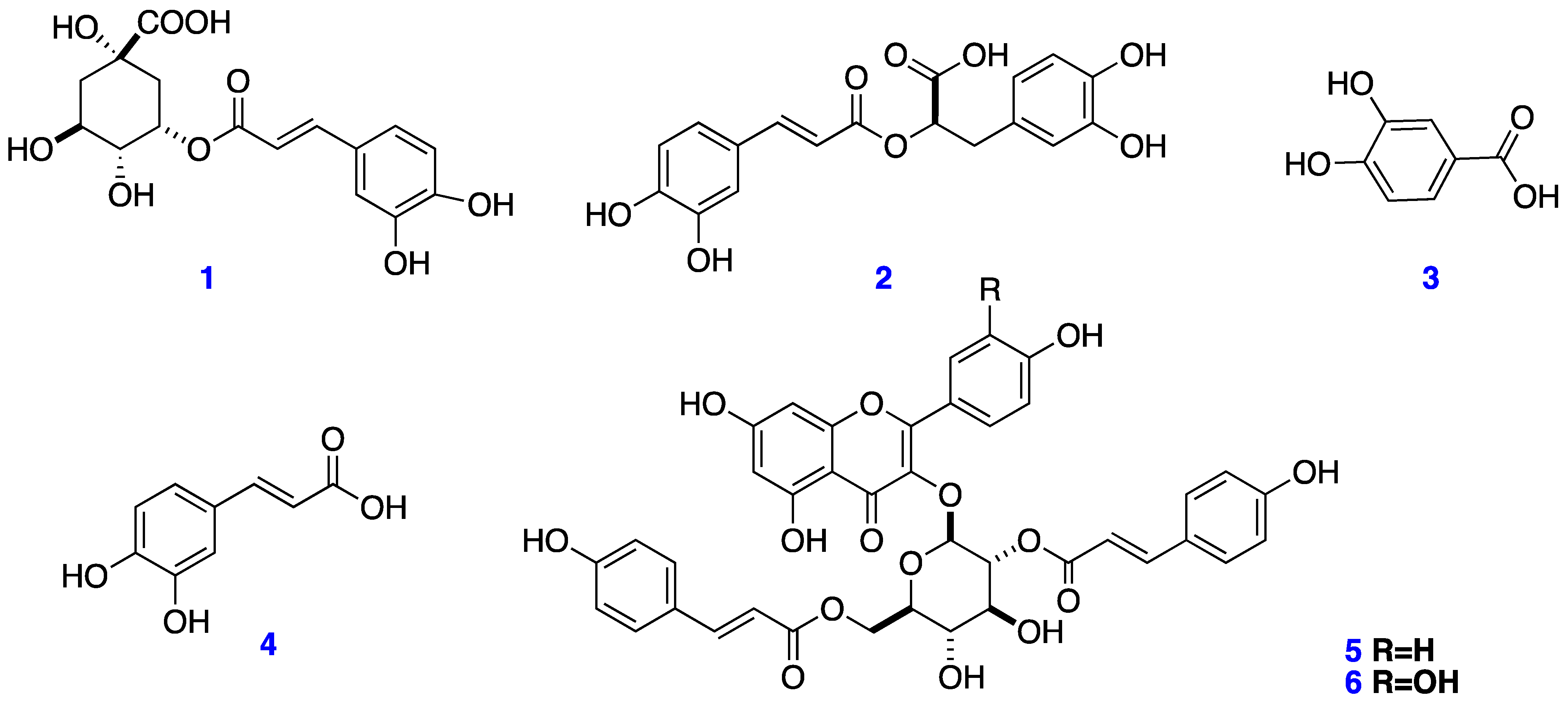
2.1. Extraction and Isolation
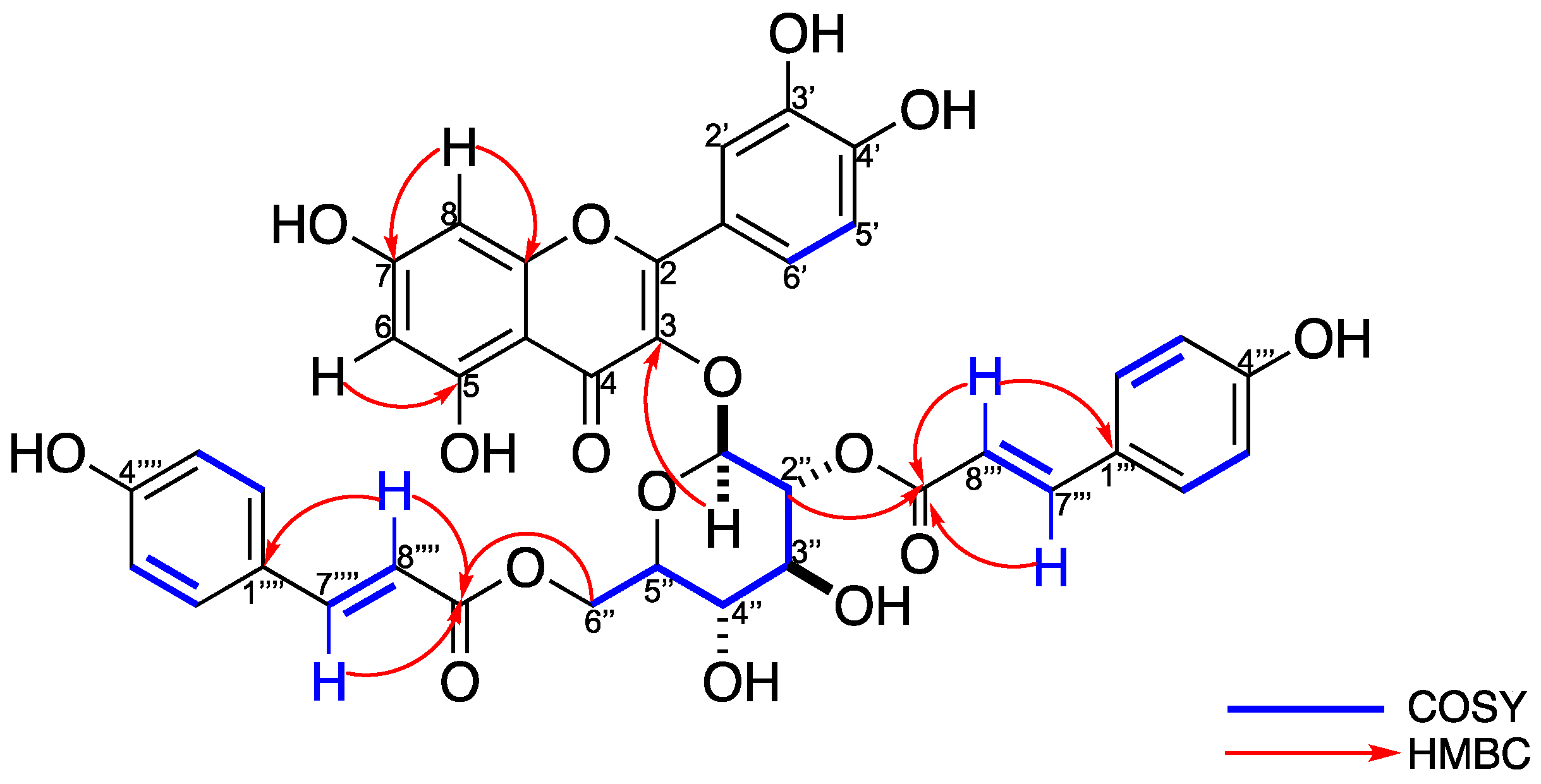
2.2. Structural Elucidation of Compound 6
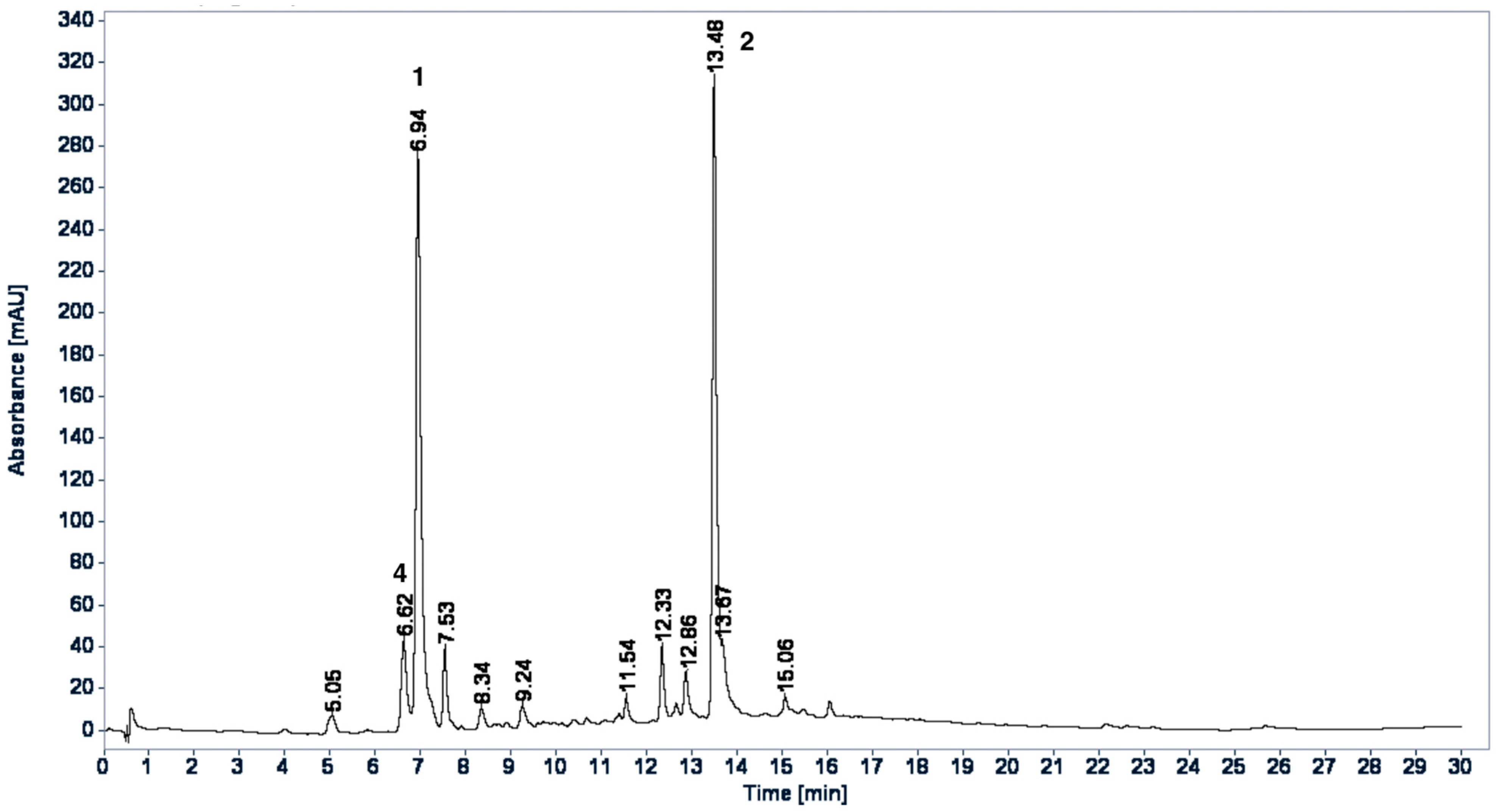
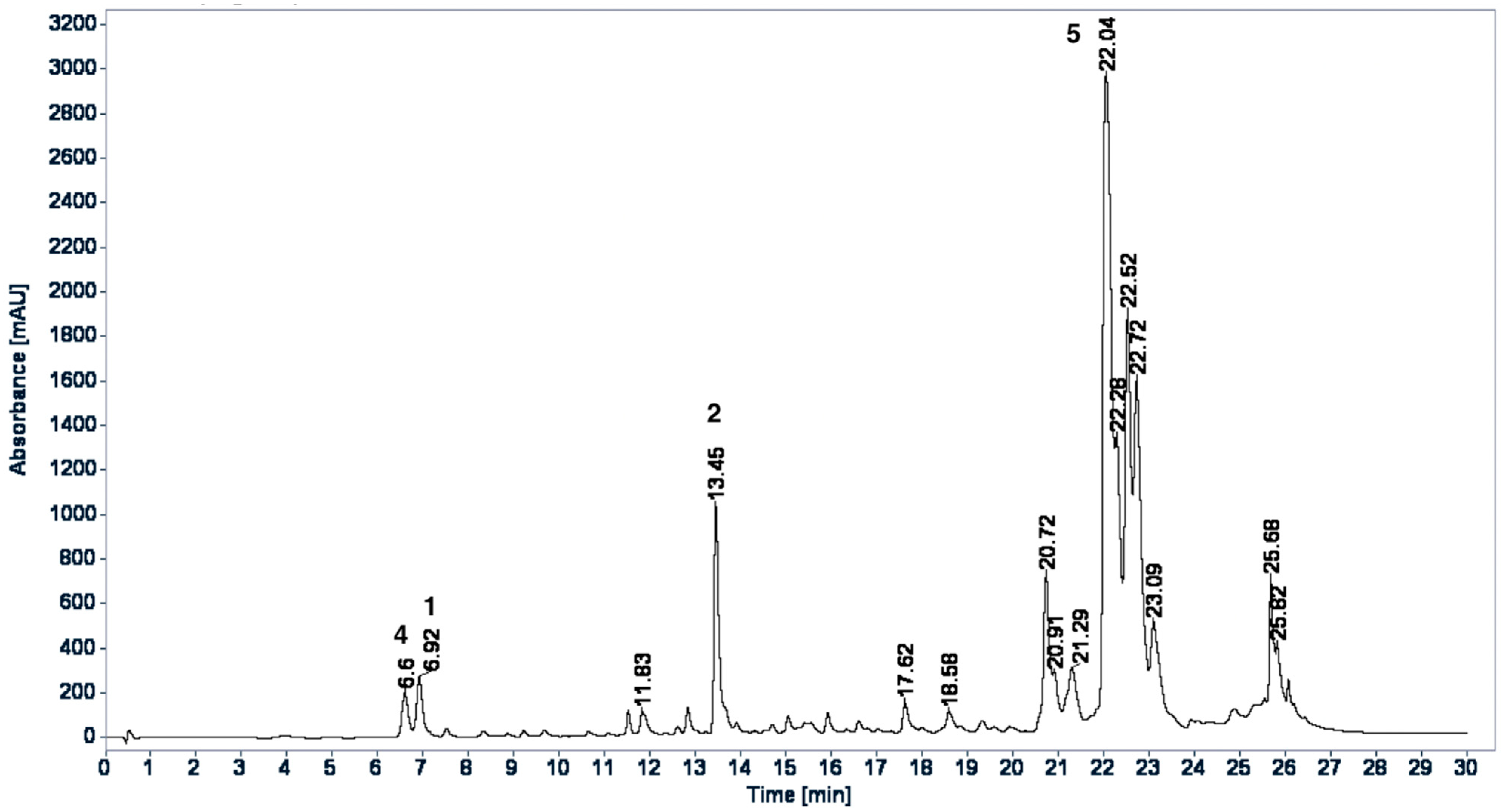
2.3. HPLC Profiles
2.4. In Vitro Inhibitory Assays
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
4.3.1. Chlorogenic Acid (1)
4.3.2. Rosmarinic Acid (2)
4.3.3. Protocatechuic Acid (3)
4.3.4. Caffeic Acid (4)
4.3.5. Kaempferol-3-O-(2,6-di-O-trans-ρ-coumaryl)-β-d-glucopyranoside (5)
4.3.6. Quercetin-3-O-(2,6-di-O-trans-ρ-coumaryl)-β-d-glucopyranoside (6)
4.4. HPLC Analysis
4.5. In Vitro Assays
4.5.1. Inhibition Assay of Glucose-6-phosphatase (G6Pase)
4.5.2. Inhibition Assay of Fructose-1,6-bisphosphatase (FBPase)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erdem, S.A.; Nabavi, S.F.; Orhan, I.E.; Daglia, M.; Izadi, M.; Nabavi, S.M. Blessings in disguise: A review of phytochemical composition and antimicrobial activity of plants belonging to the genus Eryngium. DARU J. Pharm. Sci. 2015, 23, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P. Phytochemical Constituents and Pharmacological Activities of Eryngium L. (Apiaceae). Pharm. Crop. 2012, 3, 99–120. [Google Scholar] [CrossRef]
- García-Ruiz, I. Contribución al conocimiento del género Eryngium (Apiaceae) en el estado de Michoacán, México. Acta Bot. Mex. 2013, 103, 65–118. [Google Scholar] [CrossRef] [Green Version]
- Cho, N.; Kirigia, J.; Ogurstova, K.; Reja, A. IDF Diabetes Atlas (Internet); International Diabetes Federation: Brussels, Belgium, 2021; ISBN 9782930229874. [Google Scholar]
- World Health Organization (WHO). WHO Traditional Medicine Strategy 2014–2023; World Health Organization: Geneva, Switzerland, 2013; pp. 1–76.
- Espinoza-Hernández, F.; Andrade-Cetto, A.; Escandón-Rivera, S.; Mata-Torres, G.; Mata, R. Contribution of fasting and postprandial glucose-lowering mechanisms to the acute hypoglycemic effect of traditionally used Eryngium cymosum F. Delaroche. J. Ethnopharmacol. 2021, 279, 114339. [Google Scholar] [CrossRef] [PubMed]
- Romo-Pérez, A.; Escandón-Rivera, S.M.S.M.; Andrade-Cetto, A. Chronic hypoglycemic effect and phytochemical composition of Smilax moranensis roots. Rev. Bras. Farmacogn. 2019, 29, 246–253. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Rosmarinic acid derivatives from Salvia officinalis. Phytochemistry 1999, 51, 91–94. [Google Scholar] [CrossRef]
- da Silva, L.A.L.; Faqueti, L.G.; Reginatto, F.H.; dos Santos, A.D.C.; Barison, A.; Biavatti, M.W. Phytochemical analysis of Vernonanthura tweedieana and a validated UPLC-PDA method for the quantification of eriodictyol. Braz. J. Pharmacogn. 2015, 25, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.U.; Hashmi, M.A.; Tehseen, Y.; Khan, A.; Khan, S.S.; Iqbal, J.; Perveen, S.; Khan, S.; Farooq, U.; Ahmad, V.U. Antidiabetic Flavonol Glycosides from Eryngium caeruleum. Rec. Nat. Prod. 2017, 11, 229–234. [Google Scholar]
- Le Claire, E.; Schwaiger, S.; Banaigs, B.; Stuppner, H.; Gafner, F. Distribution of a new rosmarinic acid derivative in Eryngium alpinum L. and other Apiaceae. J. Agric. Food Chem. 2005, 53, 4367–4372. [Google Scholar] [CrossRef]
- Kikowska, M.; Budzianowski, J.; Krawczyk, A.; Thiem, B. Accumulation of rosmarinic, chlorogenic and caffeic acids in in vitro cultures of Eryngium planum L. Acta Physiol. Plant. 2012, 34, 2425–2433. [Google Scholar] [CrossRef] [Green Version]
- de la Luz Cádiz-Gurrea, M.; Fernández-Arroyo, S.; Joven, J.; Segura-Carretero, A. Comprehensive characterization by UHPLC-ESI-Q-TOF-MS from an Eryngium bourgatii extract and their antioxidant and anti-inflammatory activities. Food Res. Int. 2013, 50, 197–204. [Google Scholar] [CrossRef]
- Singh, S.; Singh, D.R.; Banu, S.; Salim, K.M. Determination of bioactives and antioxidant activity in eryngium foetidum L.: A traditional culinary and medicinal herb. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2013, 83, 453–460. [Google Scholar] [CrossRef]
- Vukic, M.D.; Vukovic, N.L.; Djelic, G.T.; Obradovic, A.; Kacaniova, M.M.; Markovic, S.; Popović, S.; Baskić, D. Phytochemical analysis, antioxidant, antibacterial and cytotoxic activity of different plant organs of Eryngium serbicum L. Ind. Crops Prod. 2018, 115, 88–97. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S.; Ownby, S.; Wang, P.; Yuan, W.; Zhang, W.; Scott Beasley, R. Phenolic compounds and rare polyhydroxylated triterpenoid saponins from Eryngium yuccifolium. Phytochemistry 2008, 69, 2070–2080. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A Review on Protocatechuic Acid and Its Pharmacological Potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.; Simmonds, M.S.J. Rosmarinic acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef]
- Amoah, S.K.S.; Sandjo, L.P.; Kratz, J.M.; Biavatti, M.W. Rosmarinic Acid-Pharmaceutical and Clinical Aspects. Planta Med. 2016, 82, 388–406. [Google Scholar] [CrossRef] [Green Version]
- Jayanthy, G.; Subramanian, S. Rosmarinic acid, a polyphenol, ameliorates hyperglycemia by regulating the key enzymes of carbohydrate metabolism in high fat diet-STZ induced experimental diabetes mellitus. Biomed. Prev. Nutr. 2014, 4, 431–437. [Google Scholar] [CrossRef]
- Azevedo, M.F.; Lima, C.F.; Fernandes-Ferreira, M.; Almeida, M.J.; Wilson, J.M.; Pereira-Wilson, C. Rosmarinic acid, major phenolic constituent of Greek sage herbal tea, modulates rat intestinal SGLT1 levels with effects on blood glucose. Mol. Nutr. Food Res. 2011, 55, S15–S25. [Google Scholar] [CrossRef] [Green Version]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic acids and the acyl-quinic acids: Discovery, biosynthesis, bioavailability and bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef] [Green Version]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: A review. Evid.-Based Complement. Altern. Med. 2013, 2013, 801457. [Google Scholar] [CrossRef] [PubMed]
- Harini, R.; Pugalendi, K.V. Antihyperglycemic effect of protocatechuic acid on streptozotocin-diabetic rats. J. Basic Clin. Physiol. Pharmacol. 2010, 21, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Agunloye, O.M.; Adefegha, S.A.; Akinyemi, A.J.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 165–170. [Google Scholar] [CrossRef]
- Un, J.J.; Lee, M.K.; Yong, B.P.; Jeon, S.M.; Choi, M.S. Antihyperglycemic and antioxidant properties of caffeic acid in db/db mice. J. Pharmacol. Exp. Ther. 2006, 318, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Noriega-Cisneros, R.; Ortiz-Ávila, O.; Esquivel-Gutiérrez, E.; Clemente-Guerrero, M.; Manzo-Avalos, S.; Salgado-Garciglia, R.; Cortés-Rojo, C.; Boldogh, I.; Saavedra-Molina, A. Hypolipidemic Activity of Eryngium carlinae on Streptozotocin-Induced Diabetic Rats. Biochem. Res. Int. 2012, 2012, 603501. [Google Scholar] [CrossRef] [Green Version]
- Jaghabir, M. Hypoglycemic effects of Eryngium creticum. Arch. Pharm. Res. 1991, 14, 295–297. [Google Scholar] [CrossRef] [Green Version]
- Kasabri, V.; AbuDahab, R.; Afifi, F.; Naffa, R.; Majdalawi, L. Modulation of pancreatic MIN6 insulin secretion and proliferation and extrapancreatic glucose absorption with Achillea santolina, Eryngium creticum and Pistacia atlantica extracts: In vitro evaluation. J. Exp. Integr. Med. 2012, 2, 245. [Google Scholar] [CrossRef] [Green Version]
- Kasabri, V.; Afifi, F.U.; Hamdan, I. Evaluation of the acute antihyperglycemic effects of four selected indigenous plants from Jordan used in traditional medicine. Pharm. Biol. 2011, 49, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Dalar, A.; Türker, M.; Zabaras, D.; Konczak, I. Phenolic Composition, Antioxidant and Enzyme Inhibitory Activities of Eryngium bornmuelleri leaf. Plant Foods Hum. Nutr. 2014, 69, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Malik, T.; Pandey, D.K.; Roy, P.; Okram, A. Evaluation of phytochemicals, antioxidant, antibacterial and antidiabetic potential of alpinia galanga and eryngium foetidum plants of Manipur (India). Pharmacogn. J. 2016, 8, 459–464. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 6 | ||
|---|---|---|
| Position | δH, Multiplicity, (J in Hz) | δC |
| 2 | - | 158.8 |
| 3 | - | 134.6 |
| 4 | - | 179.2 |
| 5 | - | 163.0 |
| 6 | 6.09, d, J = 2.0 Hz | 99.8 |
| 7 | 165.7 | |
| 8 | 6.27, d, J = 2.1 Hz | 94.6 |
| 9 | - | 158.3 |
| 10 | - | 105.7 |
| 1′ | - | 123.1 |
| 2′ | 7.56, d, J = 2.0 Hz | 117.1 |
| 3′ | - | 146.0 |
| 4′ | - | 149.7 |
| 5′ | 6.85, d, J = 9.0 Hz | 116.0 |
| 6′ | 7.55, dd, J = 2.1, 8.6 Hz | 123.5 |
| 1″ | 5.63, d, J = 8.0 Hz | 100.4 |
| 2″ | 5.13, dd, J = 9.5, 8.0 Hz | 75.7 |
| 3″ | 3.68, t, J = 9.2 Hz | 76.2 |
| 4″ | 3.46, m | 72.0 |
| 5″ | 3.56, ddd, J = 9.3, 6.9, 2.3 Hz | 76.0 |
| 6″ | 4.38, dd, J = 11.7, 2.2 Hz, H-6a 4.22, dd, J = 11.8, 6.8 Hz, H6b | 64.2 |
| 1‴ | - | 127.1 |
| 2‴, 6‴ | 7.50, d, J = 8.7 Hz | 131.2 |
| 3‴, 5‴ | 6.83, d, J = 8.6 Hz | 116.8 |
| 4‴ | - | 161.3 |
| 7‴ | 7.73, d, J = 15.9 Hz | 147.1 |
| 8‴ | 6.44, d, J = 15.9 Hz | 115.2 |
| 9‴ | - | 168.6 |
| 1⁗ | - | 127.3 |
| 2⁗, 6⁗ | 7.33, d, J = 8.6 Hz | 131.3 |
| 3⁗, 5⁗ | 6.83, d, J = 8.6, Hz | 116.8 |
| 4⁗ | 161.2 | |
| 7⁗ | 7.42, d, J = 15.9 Hz | 146.6 |
| 8⁗ | 6.10, d, J = 15.9 Hz | 114.7 |
| 9⁗ | 168.8 |
| Compound | G6Pase IC50 (μg/mL) | FBPase IC50 (μg/mL) |
|---|---|---|
| Caffeic acid (4) | 719.8 ± 192.9 | - |
| kaempferol-3-O-(2,6-di-O-trans-ρ-coumaryl)-β-d-glucopyranoside (5) | 27.7 ± 1.9 | 52.5 ± 6.4 |
| Chlorogenic acid * | 179.7 ± 27.5 | - |
| Adenosine 5′-monophosphate + | - | 16.9 ± 2.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romo-Pérez, A.; Escandón-Rivera, S.M.; Miranda, L.D.; Andrade-Cetto, A. Phytochemical Study of Eryngium cymosum F. Delaroche and the Inhibitory Capacity of Its Main Compounds on Two Glucose-Producing Pathway Enzymes. Plants 2022, 11, 992. https://doi.org/10.3390/plants11070992
Romo-Pérez A, Escandón-Rivera SM, Miranda LD, Andrade-Cetto A. Phytochemical Study of Eryngium cymosum F. Delaroche and the Inhibitory Capacity of Its Main Compounds on Two Glucose-Producing Pathway Enzymes. Plants. 2022; 11(7):992. https://doi.org/10.3390/plants11070992
Chicago/Turabian StyleRomo-Pérez, Adriana, Sonia Marlen Escandón-Rivera, Luis D. Miranda, and Adolfo Andrade-Cetto. 2022. "Phytochemical Study of Eryngium cymosum F. Delaroche and the Inhibitory Capacity of Its Main Compounds on Two Glucose-Producing Pathway Enzymes" Plants 11, no. 7: 992. https://doi.org/10.3390/plants11070992