
Morphological and Molecular Characterization of Some Alternaria Species Isolated from Tomato Fruits Concerning Mycotoxin Production and Polyketide Synthase Genes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Tomato Samples
2.2. Isolation and Morphological Description of Alternaria
2.3. DNA Extraction of Alternaria Species
2.4. Polymerase Chain Reaction, Amplification of 5.8S rDNA, and Gene Sequencing
2.5. Amplification of pksH and pksJ Genes
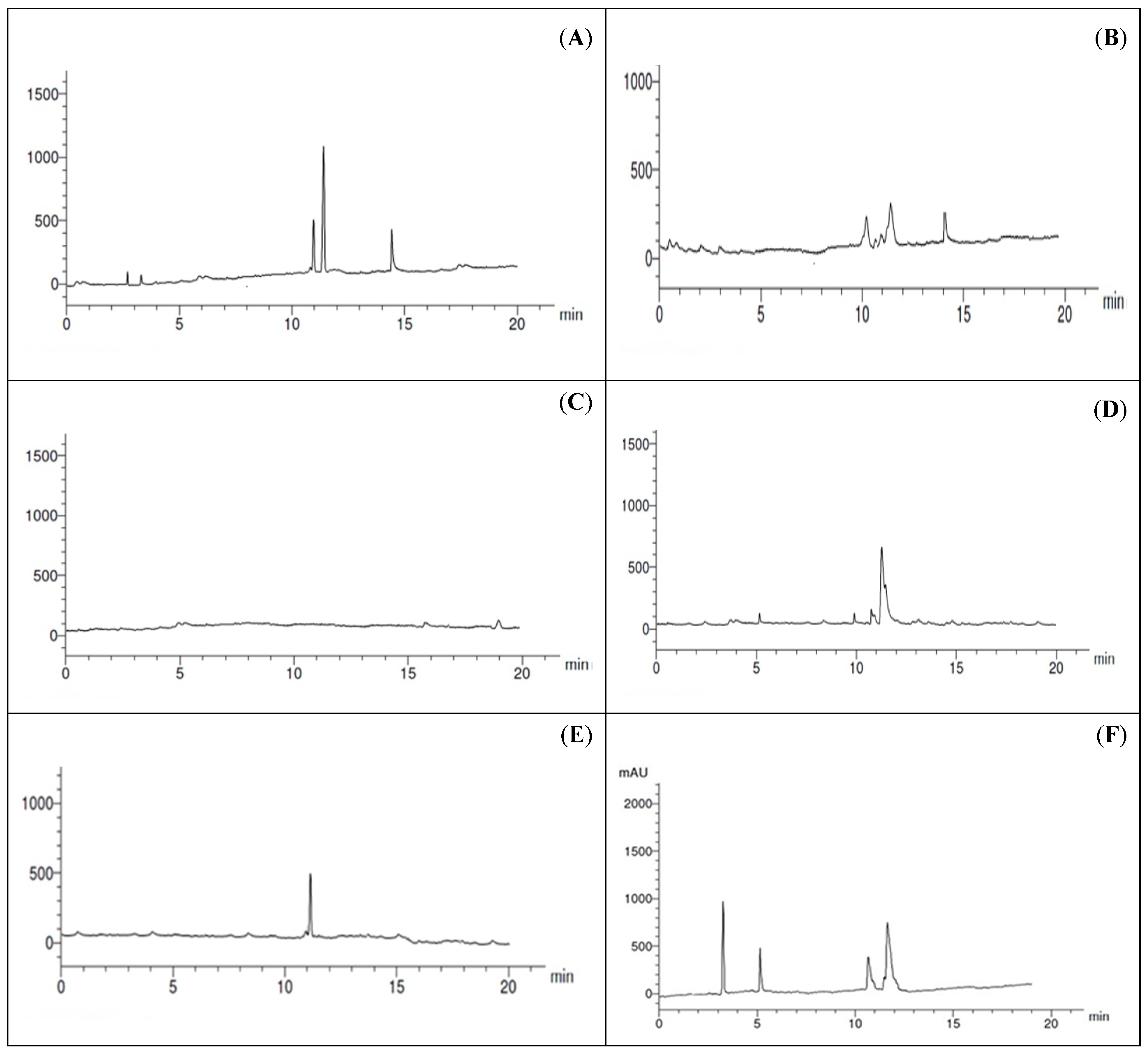
2.6. Toxin Production and Purification for HPLC Analysis
2.7. Chromatographic Analysis
3. Results
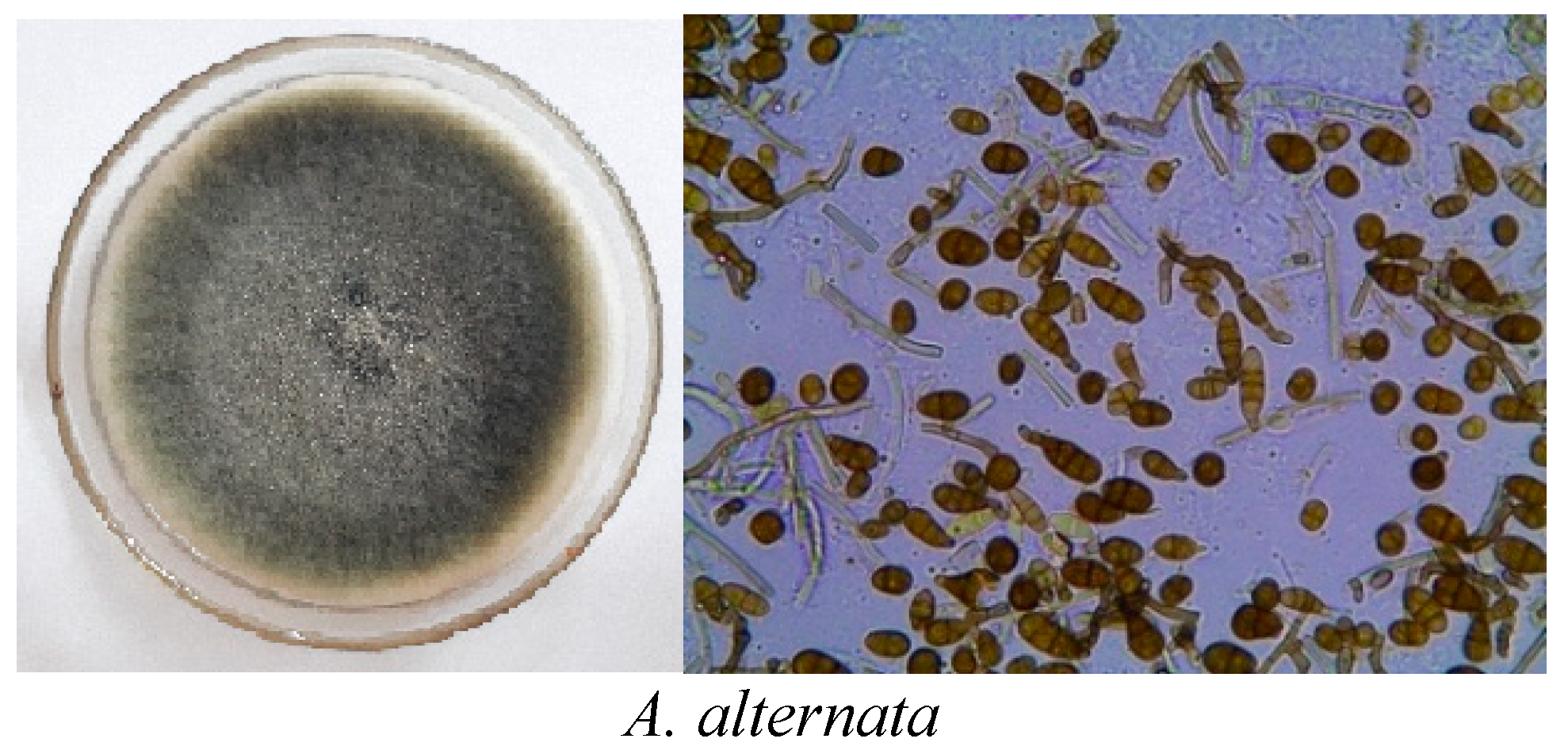
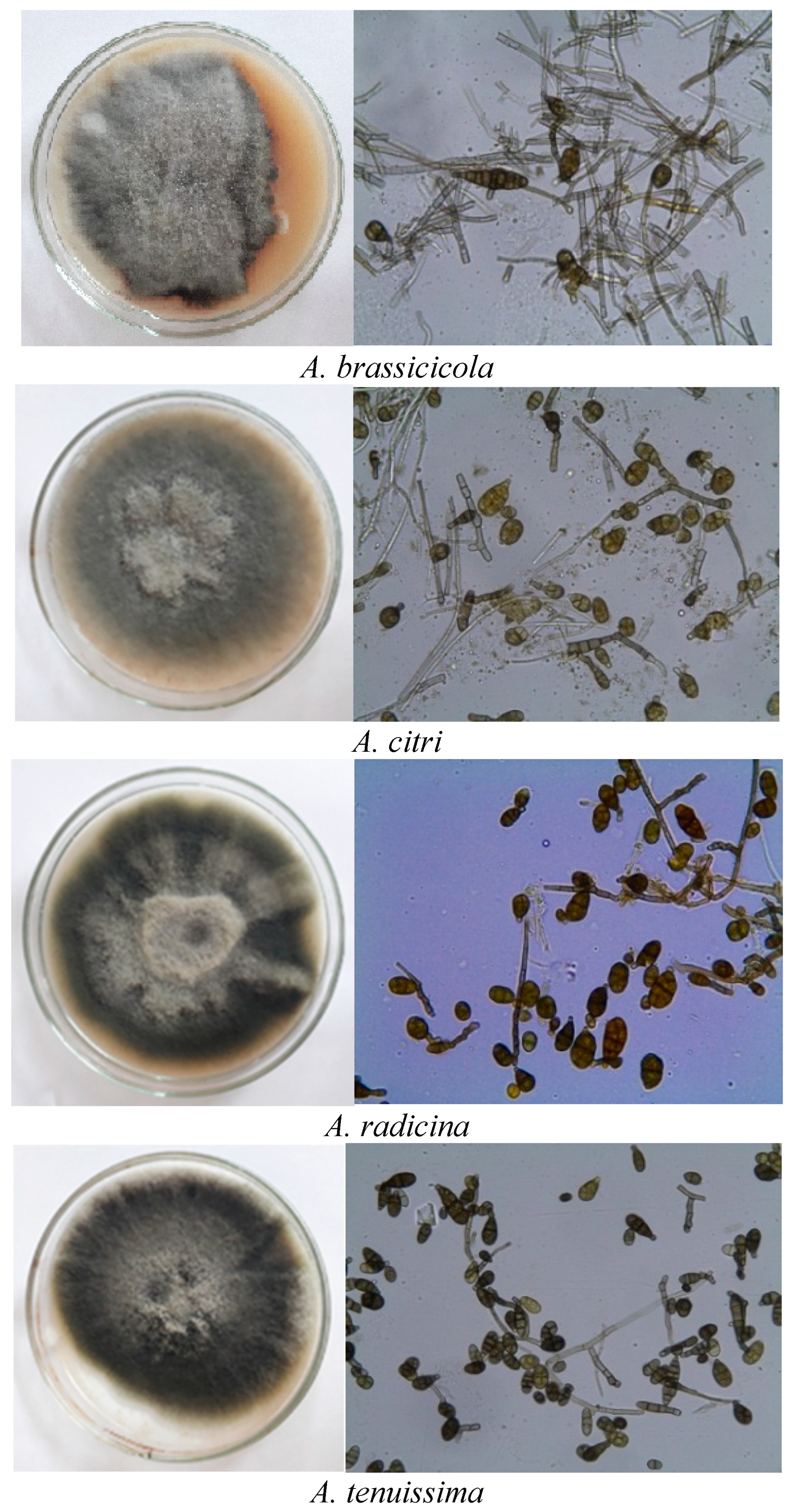
3.1. Morphological Description of Alternaria Species
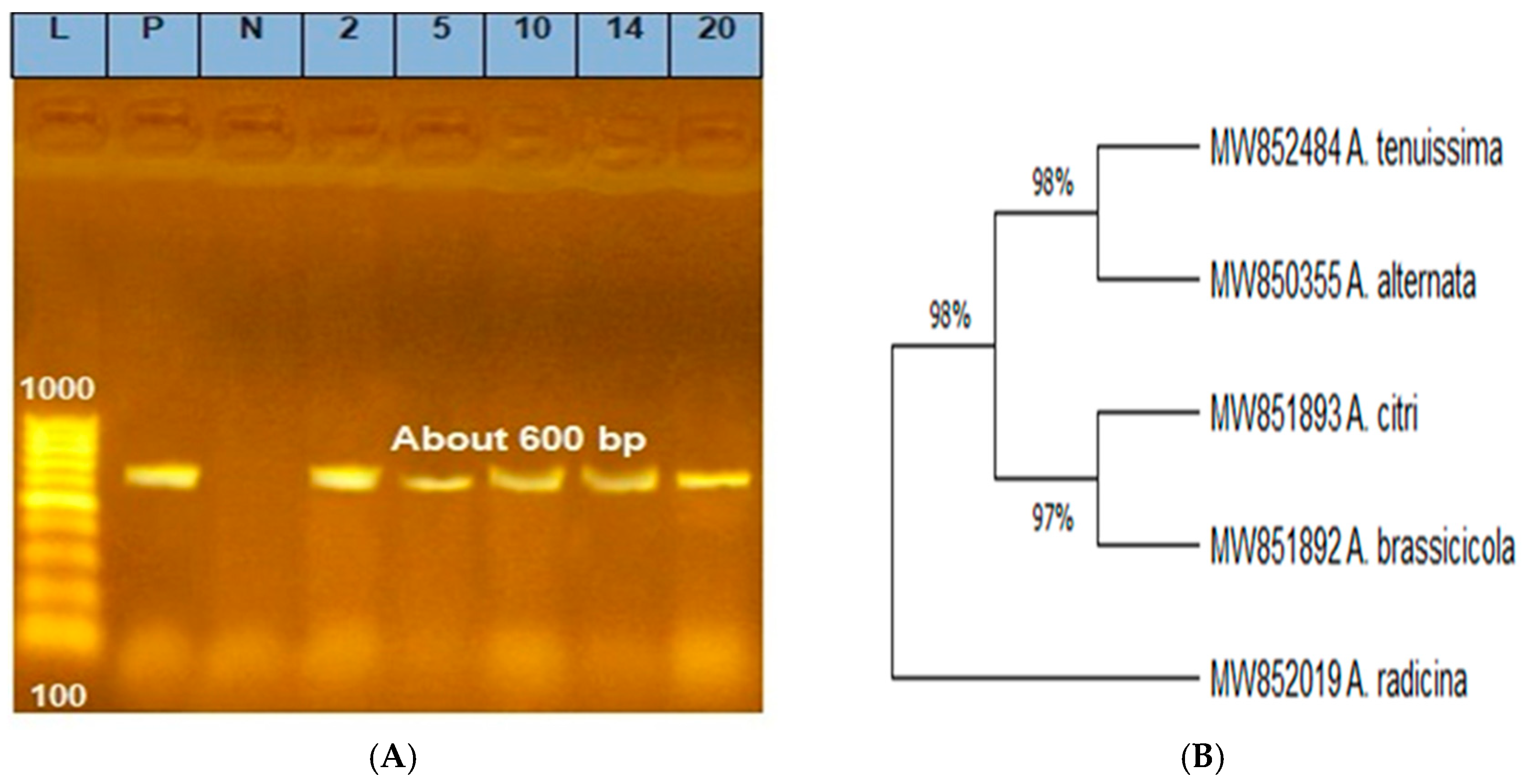
3.2. Molecular Characterization of Alternaria Species Using ITS rDNA Gene Sequencing
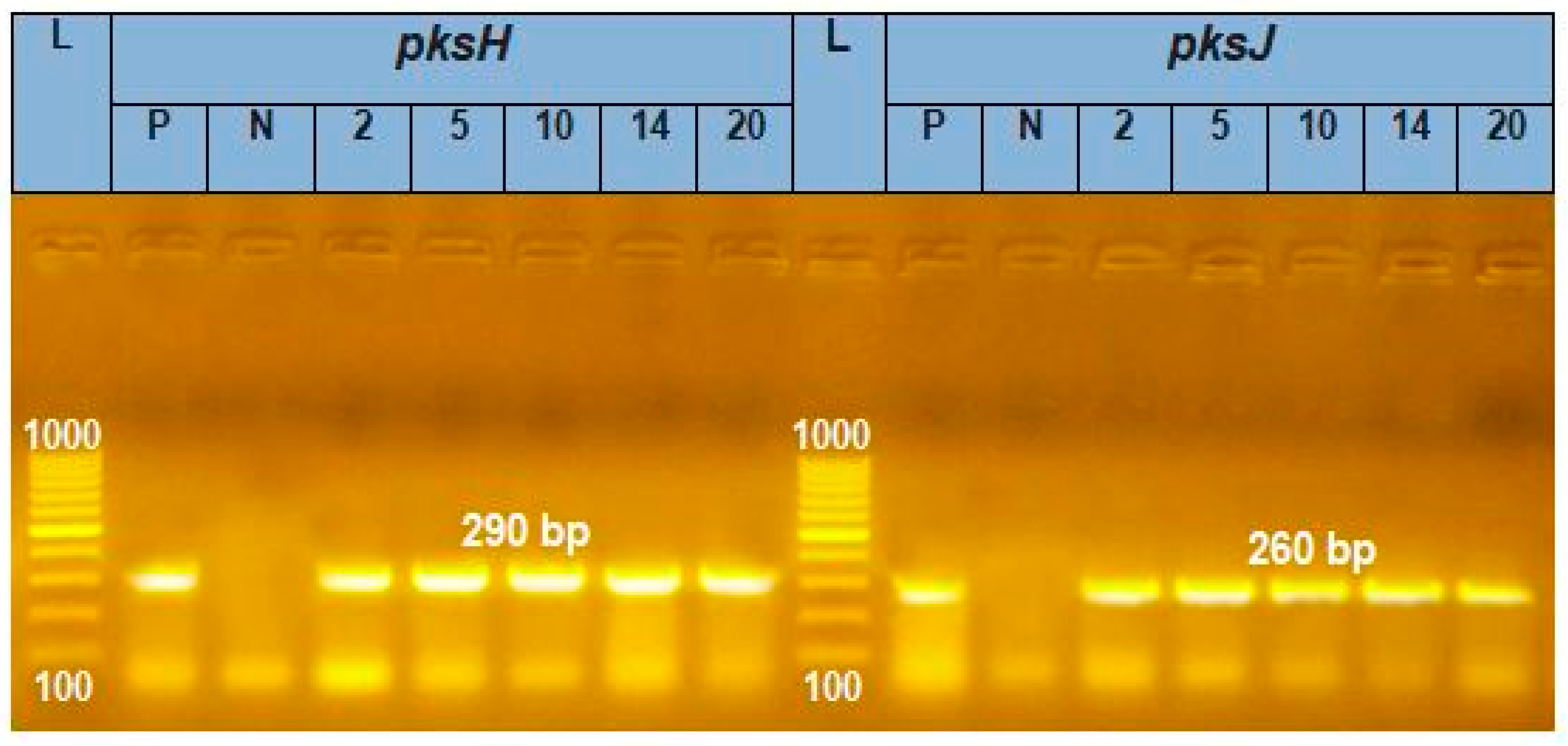
3.3. Amplification of pksH and pksJ Genes
3.4. Toxin Production and Purification for HPLC Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria toxins: Potential virulence factors and genes related to pathogenesis. Front. Microbiol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pinto, V.E.F.; Patriarca, A. Alternaria species, and their associated mycotoxins. In Mycotoxigenic Fungi: Methods in Molecular Biology; Moretti, A., Susca, A., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1542, Chapter 2; pp. 13–32. [Google Scholar]
- Rychlik, M.; Zappa, G.; Añorga, L.; Belc, N.; Castanheira, I.; Donard, O.F.X.; Kourimská, L.; Ogrinc, N.; Ocké, M.C.; Presser, K.; et al. Ensuring food integrity by metrology and FAIR data principles. Front. Chem. 2018, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, M.A.; Voigt, K. Phylogenetic and enzymatic variability of Alternaria species isolated from various substrates in Qena governorate of Upper Egypt. Arch. Phytopathol. Plant Prot. 2019, 52, 530–541. [Google Scholar] [CrossRef]
- Lee, H.B.; Patriarca, A.; Magan, N. Alternaria in food: Ecophysiology, mycotoxin production, and toxicology. Mycobiology 2015, 43, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, Y.; Taheri, P.; Mamarabadi, M. Identification of Alternaria spp. associated with tomato early blight in Iran and investigating some of their virulence factors. J. Plant Pathol. 2019, 101, 647–659. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes. Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, J.P.; Kushwaha, G.D.; Shukla, D.N. Black point disease of wheat and its implications on seed quality. Crop Res. 2014, 47, 21–23. [Google Scholar]
- Al-Nadabi, H.H.; Maharachchikumbura, S.S.N.; Agrama, H.; Al-Azri, M.; Nasehi, A.; Al-Sadi, A.M. Molecular characterization, and pathogenicity of Alternaria species on wheat and date palms in Oman. Eur. J. Plant Pathol. 2018, 152, 577–588. [Google Scholar] [CrossRef]
- Masiello, M.; Somma, S.; Susca, A.; Ghionna, V.; Logrieco, A.F.; Franzoni, M.; Ravaglia, S.; Meca, G.; Moretti, A. Molecular identification and mycotoxin production by Alternaria species occurring on Durum wheat, showing black point symptoms. Toxins 2020, 12, 275. [Google Scholar] [CrossRef] [Green Version]
- Bessadat, N.; Berruyer, R.; Hamon, B.; Bataille-Simoneau, N.; Benichou, S.; Kihal, M.; Henni, D.E.; Simoneau, P. Alternaria species associated with early blight epidemics on tomato and other Solanaceae crops in northwestern Algeria. Eur. J. Plant Pathol. 2017, 148, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, A.; Bahramikia, S. Molecular identification and genetic variation of Alternaria species isolated from tomatoes using ITS1 sequencing and inter simple sequence repeat methods. Curr. Med. Mycol. 2019, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aung, S.L.L.; Liu, H.F.; Pei, D.F.; Lu, B.B.; Oo, M.M.; Deng, J.X. Morphology and molecular characterization of a fungus from the Alternaria alternata species complex causing black spots on Pyrus sinkiangensis (Koerle pear). Mycobiology 2020, 48, 233–239. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Dietary exposure to Alternaria toxins in the European population. EFSA J. 2016, 14, e04654. [Google Scholar]
- Rychlik, A.M. Potential health hazards due to the occurrence of the mycotoxin tenuazonic acid in infant food. Eur. Food. Res. Technol. 2013, 236, 491–497. [Google Scholar]
- Meena, M.; Samal, S. Alternaria host-specific (HSTs) toxins: An overview of chemical characterization, target sites, regulation, and their toxic effects. Toxicol. Rep. 2019, 6, 445–458. [Google Scholar] [CrossRef]
- Saha, D.; Fetzner, R.; Burkhardt, B.; Podlech, J.; Metzler, M.; Dang, H.; Lawrence, C.; Fischer, R. Identification of a polyketide synthase required for alternariol (AOH) and alternariol-9-methyl ether (AME) formation in Alternaria alternata. PLoS ONE 2012, 7, e40564. [Google Scholar] [CrossRef]
- Prieto, K.R.; de Medeiros, L.S.; Isidoro, M.M.; Toffano, L.; da Silva, M.F.G.F.; Fernandes, J.B.; Vieira, P.C.; Forim, M.R.; Rodrigues-Filho, E.; Stuart, R.M.; et al. Rapid detection of ACTG- and AK-Toxins in Alternaria alternata by LC-ESI-MS/MS analysis and antifungal properties of Citrus compounds. J. Braz. Chem. Soc. 2016, 27, 1493–1505. [Google Scholar]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An overview. J. Food Qual. 2017, 10, 1569748. [Google Scholar]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef]
- Montemurro, N.; Visconti, A. Alternaria metabolites, chemical and biological data. In Alternaria: Biology, Plant Diseases, and Metabolites; Chelkowski, J., Visconti, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; pp. 449–557. [Google Scholar]
- Hickert, S.; Krug, I.; Cramer, B.; Humpf, H.U. Detection and quantitative analysis of the non-cytotoxicallo-Tenuazonic acid in tomato products by stable isotope dilution HPLC-MS/MS. J. Agric. Food Chem. 2015, 63, 10879–10884. [Google Scholar] [CrossRef] [PubMed]
- Garganese, F.; Schena, L.; Siciliano, I.; Prigigallo, M.I.; Spadaro, D.; De Grassi, A.; Ippolito, A.; Sanzani, S.M. Characterization of Citrus-associated Alternaria species in Mediterranean areas. PLoS ONE 2016, 16, e0163255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motta, S.D.; Soares, L.M.V. A method for the determination of two Alternaria toxins, alternariol, and alternariol monomethyl ether, in tomato products. Braz. J. Microbiol. 2000, 31, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.M. Analysis of agricultural commodities and foods for Alternaria mycotoxins. J. AOAC Int. 2001, 84, 1809–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitenbach, M.; Simon-Nobbe, B. The allergens of Cladosporium herbarum and Alternaria alternata. Chem. Immunol. 2002, 81, 48–72. [Google Scholar] [PubMed] [Green Version]
- Babiceanu, M.C.; Howard, B.A.; Rumore, A.C.; Kita, H.; Lawrence, C.B. Analysis of global gene expression changes in human bronchial epithelial cells exposed to spores of the allergenic fungus, Alternaria alternata. Front. Microbiol. 2013, 4, 196. [Google Scholar] [CrossRef] [Green Version]
- Hayes, T.; Rumore, A.; Howard, B.; He, X.; Luo, M.; Wuenschmann, S.; Chapman, M.; Kale, S.; Li, L.; Kita, H.; et al. Innate immunity induced by the major allergen alt a 1 from the fungus Alternaria is dependent upon toll-like receptors 2/4 in human lung epithelial cells. Front. Immunol. 2018, 9, 1507. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.; Turner, G.; Bennett, J.W. Fungal secondary metabolism-from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Crawford, J.M.; Townsend, C.A. New insights into the formation of fungal aromatic polyketides. Nat. Rev. Microbiol. 2010, 8, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Gulder, T.A.; Freeman, M.F.; Piel, J. The catalytic diversity of multimodular polyketide synthases: Natural product biosynthesis beyond textbook assembly rules. In Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–53. [Google Scholar]
- Cox, R. Polyketides, proteins and genes in fungi: Programmed nanomachines begin to reveal their secrets. Org. Biomol. Chem. 2007, 5, 2010–2026. [Google Scholar] [CrossRef]
- Zghair, F.S.; Mohamed, B.T.; Neda, S.M. Molecular assay of polyketide synthase gene of Alternariol (AOH) produce by Alternaria alternata. Int. J. Innov. Appl. Stud. 2014, 9, 1124–1127. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Domsch, K.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd ed.; IHW-Verlag: Eching, Germany, 2007; p. 672. [Google Scholar]
- Simmons, E.G. Alternaria: An Identification Manual; CBS Biodiversity Series; CBS: Utrercht, The Netherlands, 2007; Volume 6, pp. 1–775. [Google Scholar]
- Basim, H.; Basim, E.; Baki, D.; Abdulai, M.; Öztürk, N.; Balkic, R. Identification, and characterization of Alternaria alternata (Fr.) Keissler causing Ceratonia blight disease of carob (Ceratonia siliqua L.) in Turkey. Eur. J. Plant Pathol. 2018, 151, 73–86. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.R.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Tarini, N.M.A.; Wahid, M.H.; Ibrahim, F.; Yasmon, A.; Djauzi, S. Development of multiplex-PCR assay for rapid detection of Candida spp. Med. J. Indones. 2010, 19, 83–87. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Saleem, A.; Al-Johani, M. Mycobiota and chromatographic analyses of aflatoxin contamination of Aspergillus species isolated from poultry feed. J. Food Saf. 2018, 38, e12560. [Google Scholar] [CrossRef]
- Saleem, A. Mycobiota and molecular detection of Aspergillus flavus and A. parasiticus aflatoxin contamination of Strawberry (Fragaria ananassa Duch.) fruits. Arch. Phytopathol. Plant Prot. 2018, 50, 982–996. [Google Scholar] [CrossRef]
- Habib, W.; Masiello, M.; El Ghorayeb, R.; Gerges, E.; Susca, A.; Meca, G.; Quiles, J.M.; Logrieco, A.F.; Moretti, A. Mycotoxin profile and phylogeny of pathogenic Alternaria species isolated from symptomatic tomato plants in Lebanon. Toxins 2021, 13, 513. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Species | Average Conidial Length (µm) ± SD | Average Conidial Width (µm) ± SD | L/W Ratio ± SD | Average of Transversa Septa ± SD | Average of Longitudinal Septa ± SD | Average Transversa/Longitudinal Septa ± SD |
|---|---|---|---|---|---|---|---|
| 1 | A. alternata | 22.6 ± 7.0 | 11.4 ± 1.6 | 2.0 ± 0.5 | 3.1 ± 1.7 | 2.6 ± 0.8 | 1.2 ± 1.1 |
| 2 | 28.2 ± 6.74 | 10.4 ± 1.78 | 2.7 ± 0.87 | 3.3 ± 1.1 | 1.4 ± 0.66 | 2.4 ± 0.95 | |
| 3 | A. citri | 29.4 ± 11.18 | 15.9 ± 3.07 | 1.9 ± 0.73 | 4.3 ± 1.25 | 2.5 ± 0.97 | 1.7 ± 0.98 |
| 4 | 27.1 ± 5.11 | 10.5 ± 1.70 | 2.6 ± 0.80 | 3.2 ± 1.01 | 2.4 ± 0.92 | 1.3 ± 0.99 | |
| 5 | 41.4 ± 11.88 | 11.4 ± 2.64 | 3.6 ± 1.21 | 5.9 ± 1.45 | 1.4 ± 0.52 | 4.2 ± 1.86 | |
| 6 | 40.3 ± 10.11 | 11.0 ± 2.50 | 3.7 ± 1.22 | 5.6 ± 1.11 | 2.0 ± 0.88 | 2.8 ± 0.76 | |
| 7 | A. radicina | 24.4 ± 8.84 | 10.6 ± 3.53 | 2.3 ± 1.56 | 3.0 ± 1.49 | 1.6 ± 0.84 | 1.9 ± 1.53 |
| 8 | 34.8 ± 5.66 | 15.2 ± 1.72 | 2.3 ± 0.51 | 3.8 ± 1.40 | 2.5 ± 0.85 | 1.5 ± 1.32 | |
| 9 | 33.2 ± 4.5 | 15.3 ± 1.60 | 2.2 ± 0.40 | 3.8 ± 1.20 | 2.0 ± 0.98 | 1.9 ± 1.44 | |
| 10 | A. brassicicola | 30.6 ± 14.61 | 15.3 ± 1.43 | 2.0 ± 1.02 | 6.0 ± 2.31 | 3.2 ± 0.79 | 1.9 ± 0.72 |
| 11 | 32.8 ± 2.35 | 16.2 ± 1.68 | 2.0 ± 0.40 | 5.4 ± 0.70 | 2.2 ± 0.42 | 2.5 ± 1.07 | |
| 12 | A. alternata | 31.4 ± 5.53 | 14.3 ± 2.88 | 2.2 ± 0.91 | 3.8 ± 1.14 | 1.7 ± 0.82 | 2.2 ± 1.38 |
| 13 | 21.5 ± 1.33 | 10.1 ± 3.34 | 2.1 ± 0.39 | 3.7 ± 1.37 | 1.3 ± 0.66 | 2.8 ± 0.67 | |
| 14 | A. radicina | 31.4 ± 5.28 | 15.5 ± 2.03 | 2.0 ± 0.48 | 2.0 ± 0.82 | 3.1 ± 0.57 | 0.7 ± 0.28 |
| 15 | 28.5 ± 7.61 | 15.8 ± 2.33 | 1.8 ± 0.48 | 3.1 ± 1.20 | 1.8 ± 1.14 | 1.7 ± 0.88 | |
| 16 | 24.8 ± 3.39 | 13.1 ± 2.54 | 1.9 ± 0.32 | 3.1 ± 0.34 | 1.2 ± 0.32 | 2.7 ± 0.60 | |
| 17 | 29.0 ± 8.61 | 13.7 ± 1.94 | 2.1 ± 0.95 | 2.6 ± 0.97 | 1.3 ± 0.48 | 2.0 ± 0.84 | |
| 18 | 34.1 ± 4.30 | 11.3 ± 2.35 | 3.0 ± 1.22 | 4.1 ± 1.55 | 1.5 ± 0.54 | 2.7 ± 0.63 | |
| 19 | A. tenuissima | 28.7 ± 2.98 | 12.4 ± 1.20 | 2.3 ± 0.33 | 3.2 ± 0.42 | 1.5 ± 0.53 | 2.1 ± 1.07 |
| 20 | 36.0 ± 6.20 | 11.0 ± 0.93 | 3.3 ± 0.71 | 4.6 ± 1.65 | 1.5 ± 0.71 | 3.1 ± 1.41 |
| Mycotoxins | RT | A. alternata | A. brassicicola | A. citri | A. radicina | A. tenuissima |
|---|---|---|---|---|---|---|
| Altenuene | 10 | ND | ND | 1.35 | ND | ND |
| Alternariol | 11 | 5.12 | ND | ND | 0.89 | 4.56 |
| Alternariol methyl ether | 11.5 | 9.85 | ND | 5.12 | 6.65 | ND |
| Tenuazonic acid | 14.8 | 4.12 | ND | 1.51 | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleem, A.; El-Shahir, A.A. Morphological and Molecular Characterization of Some Alternaria Species Isolated from Tomato Fruits Concerning Mycotoxin Production and Polyketide Synthase Genes. Plants 2022, 11, 1168. https://doi.org/10.3390/plants11091168
Saleem A, El-Shahir AA. Morphological and Molecular Characterization of Some Alternaria Species Isolated from Tomato Fruits Concerning Mycotoxin Production and Polyketide Synthase Genes. Plants. 2022; 11(9):1168. https://doi.org/10.3390/plants11091168
Chicago/Turabian StyleSaleem, Abdelrahman, and Amany A. El-Shahir. 2022. "Morphological and Molecular Characterization of Some Alternaria Species Isolated from Tomato Fruits Concerning Mycotoxin Production and Polyketide Synthase Genes" Plants 11, no. 9: 1168. https://doi.org/10.3390/plants11091168