
Investigation of the Copper Requirements of the Metallophyte Liverworts Cephaloziella nicholsonii Douin and C. massalongoi (Spruce) Müll.Frib

Abstract
:1. Introduction
2. Results
2.1. Soil Analysis
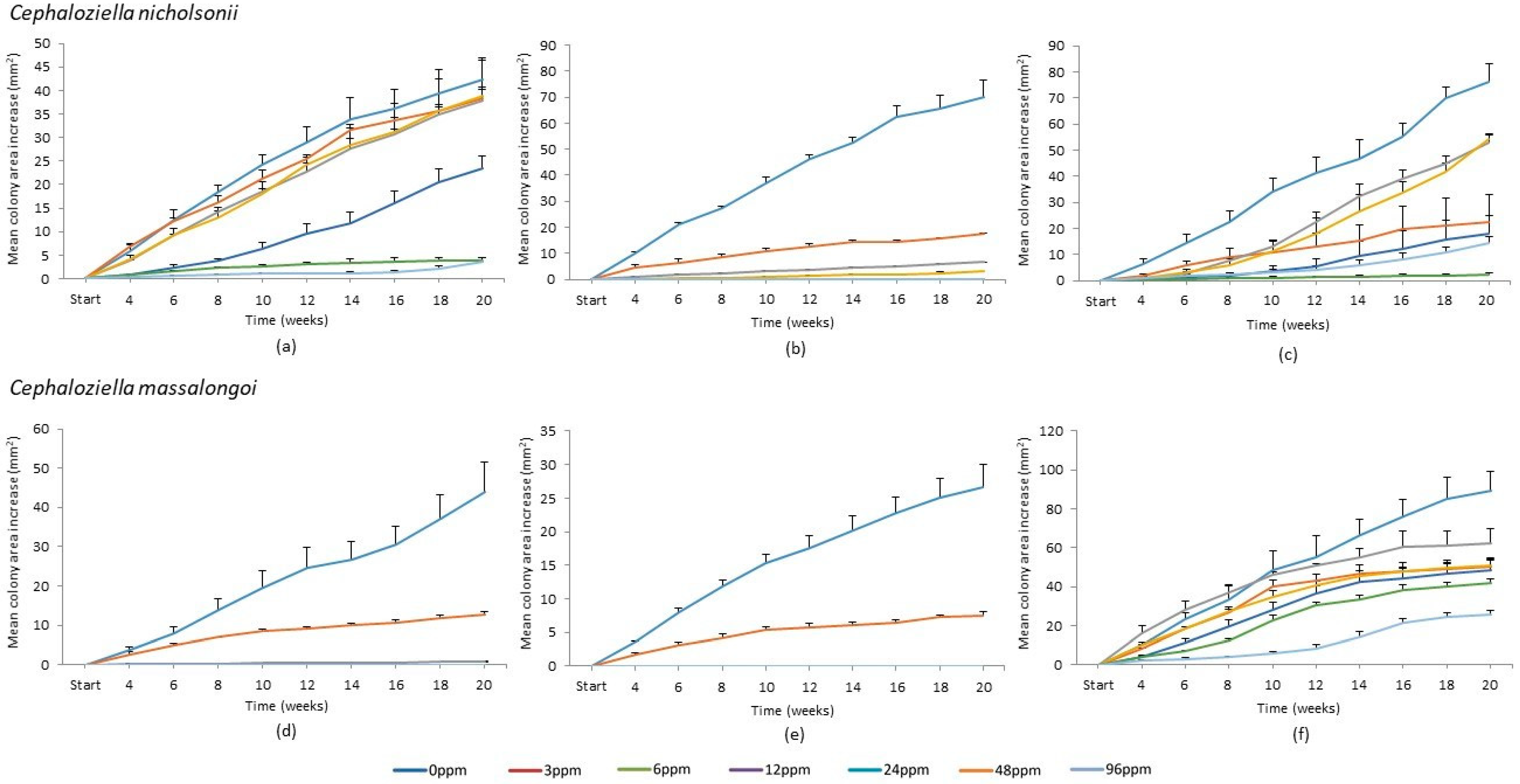
2.2. Growth through Course of Experiment
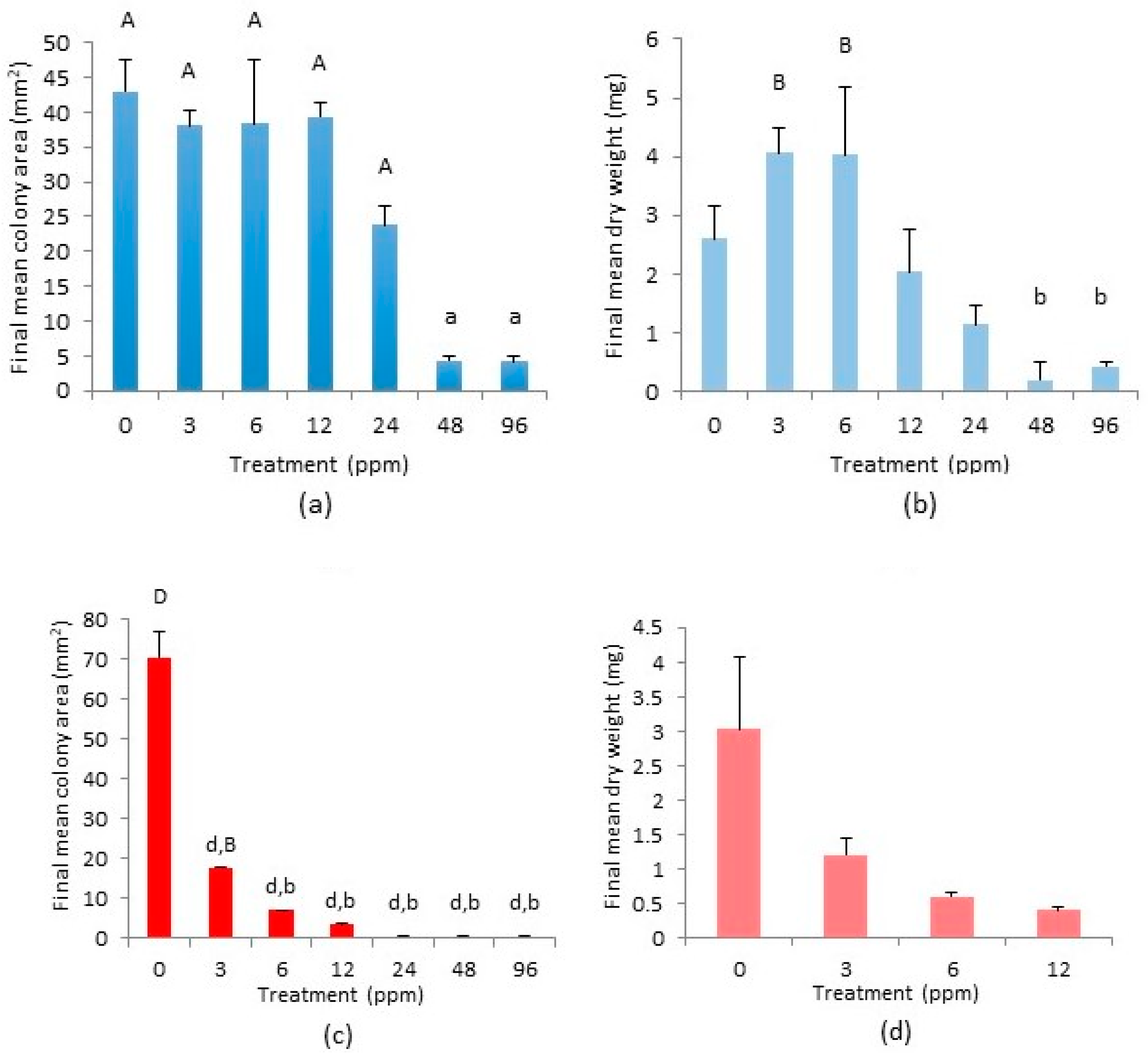
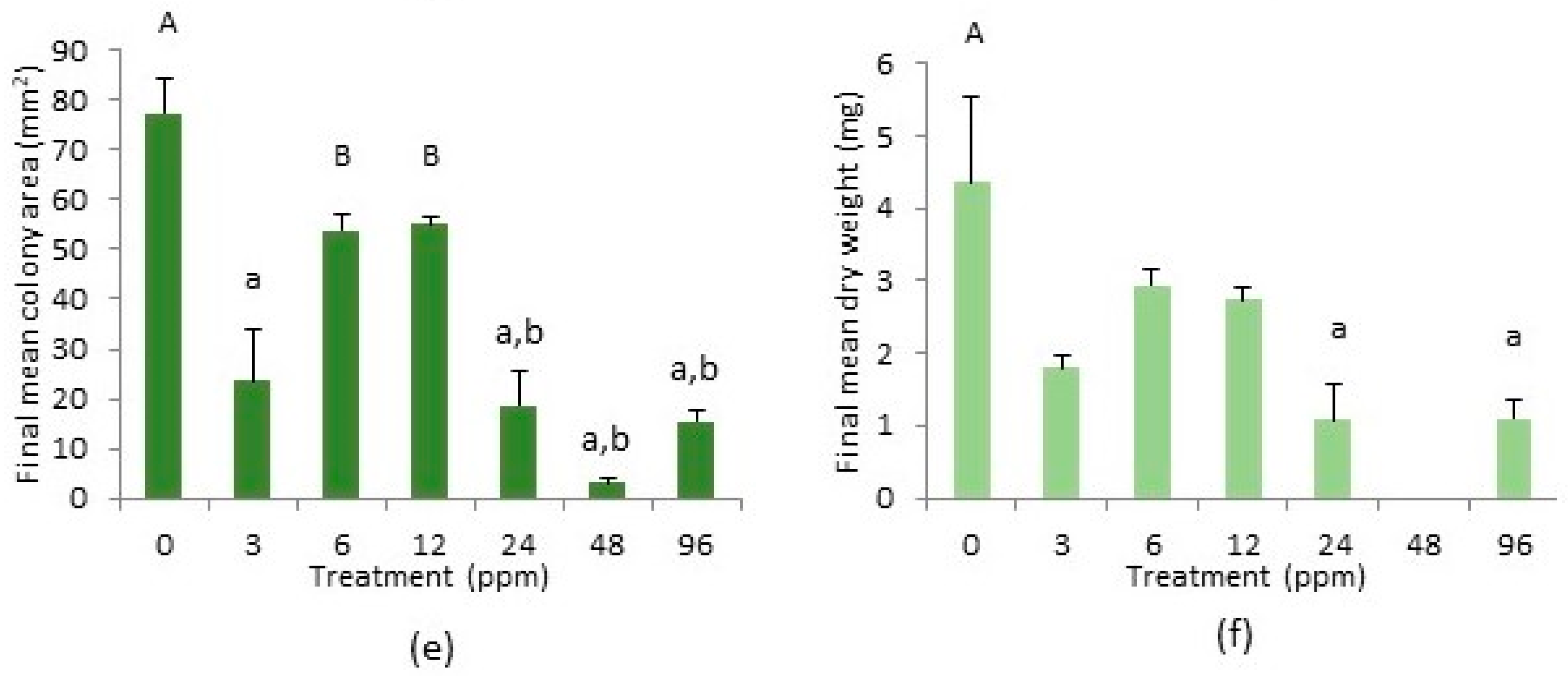
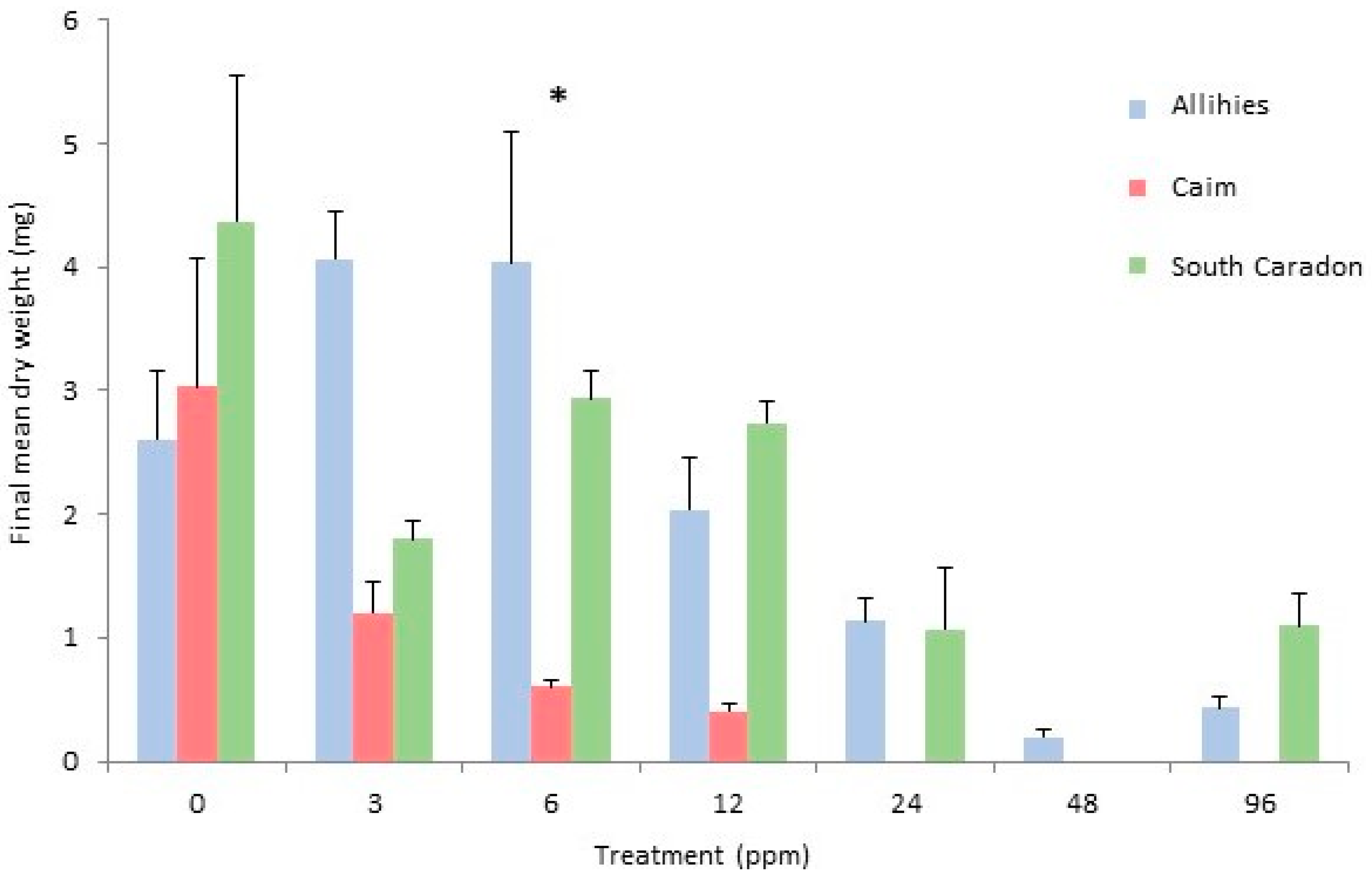
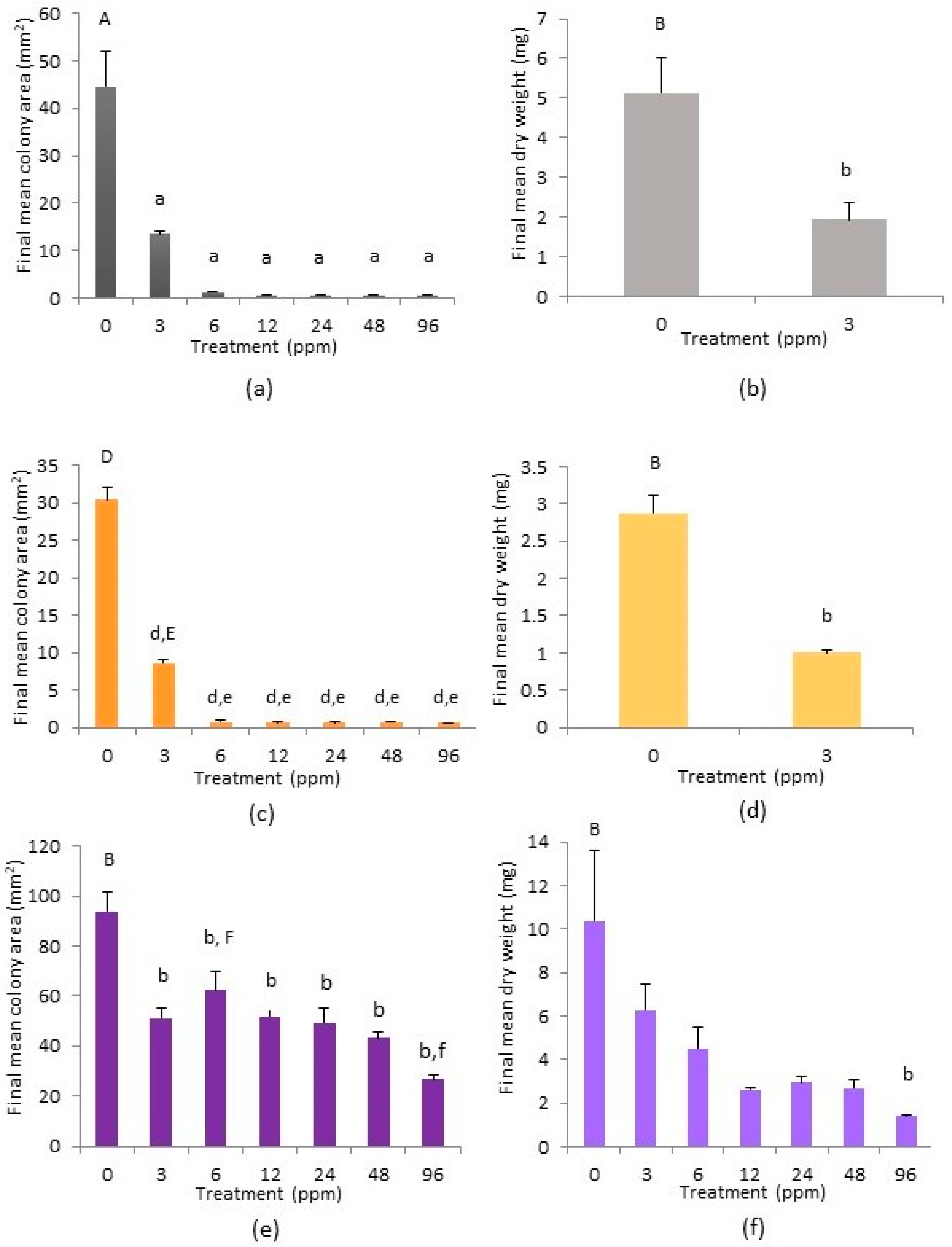
2.3. Cephaloziella nicholsonii
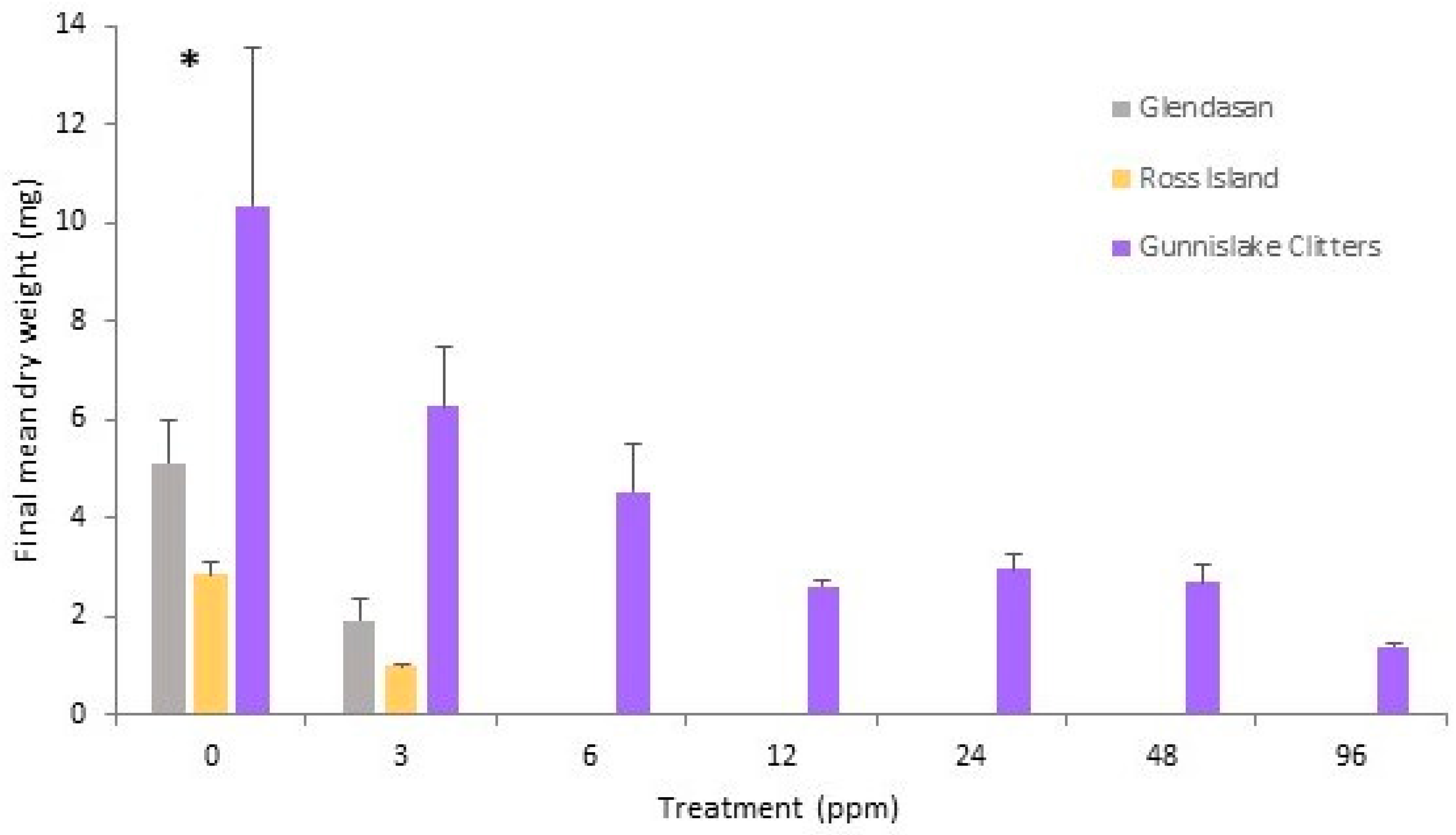
2.4. Cephaloziella massalongoi
2.5. Tolerance Indices of C. nicholsonii and C. massalongoi
3. Discussion
4. Materials and Methods
4.1. Sampling and Initiation into Culture
4.2. Growth Media for Experimental Investigation
4.3. Experimental Procedure
4.4. Soil Sampling and Analysis
4.5. Data Handling and Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Širka, P.; Kubešová, S.; Mišíková, K. Bryophytes of spoil heaps rich in toxic metals in Central Slovakia. Thaiszia J. Bot. 2018, 28, 59–77. [Google Scholar]
- Holyoak, D.T.; Lockhart, N. Survey of bryophytes and metallophyte vegetation of metalliferous mine spoil in Ireland. J. Min. Herit. Trust. Irel. 2011, 11, 3–16. [Google Scholar]
- Callaghan, D.A.; Bowyer, H.M. Chemical characteristics of the soil occupied by Cephaloziella massalongi and C. nicholsonii in Cornwall (UK). J. Bryol. 2011, 33, 170–173. [Google Scholar] [CrossRef]
- European Commission. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. In Official Journal of the European Union–Legislation; European Commission: Luxembourg, 1992; Volume 206, pp. 7–50. [Google Scholar]
- Rodwell, J.S.; Morgan, V.; Jefferson, R.G.; Moss, D. The European Context of British Lowland Grassland; JNCC Report No. 394; Joint Nature Conservation Committee: Peterborough, UK, 2007. [Google Scholar]
- Doyle, G.J. Minuartio-Thlaspietum alpestris (Violetea calaminariae) in Ireland. J. Life Sci. R. Dublin Soc. 1982, 3, 147–164. [Google Scholar]
- Baumbach, H. Metallophytes and metallicolous vegetation: Evolutionary aspects, taxonomic changes and conservational status in Central Europe. In Perspectives on Nature Conservation–Patterns, Pressures and Prospects; Tiefenbacher, J.P., Ed.; InTech: Rijeka, Croatia, 2002; pp. 93–118. [Google Scholar]
- Baker, A.J.M.; Ernst, W.H.O.; Van Der Ent, A.; Malaisse, F.; Ginocchio, R. Metallophytes: The unique biological resource, its ecology and conservational status in Europe, central Africa and Latin America. In Ecology of Industrial Pollution; Batty, L.C., Hallberg, K.B., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 7–40. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publications: Oxford, UK; London, UK, 1989. [Google Scholar]
- Alloway, B.J. Heavy Metals in Soils; Blackie: Glasgow, Scotland, 1990. [Google Scholar]
- Stanley, G.; Gallagher, V.; Ní Mhairtín, F.; Brogan, J.; Lally, P.; Doyle, E.; Farrell, L. Historic Mine Sites–Inventory and Risk Classification Volume 1; Environmental Protection Agency and the Geological Society of Ireland: Dublin, Ireland, 2009.
- Rooney, C.P.; Zhao, F.-J.; McGrath, S.P. Soil factors controlling the expression of copper toxicity to plants in a wide range of European soils. Environ. Toxicol. Chem. 2006, 25, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.A.; Han, T.; Ahn, S.K.; Kang, H.; Cho, M.R.; Lee, S.C.; Im, K.H. Effects of heavy metals on plant growths and pigment contents in Arabidopsis thaliana. Plant Pathol. J. 2012, 28, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Holyoak, D.T. Bryophytes and Metallophytes Vegetation on Metalliferous Mine-Waste in Ireland: Report to National Parks and Wildlife Service of a Survey in 2008; National Parks and Wildlife Service: Dublin, Ireland, 2008.
- Lockhart, N. Natural Heritage Areas (NHAs) for Calaminarian Grassland: Selection Criteria. In Irish Wildlife Manuals, No. 130; National Parks and Wildlife Service, Department of Housing, Local Government and Heritage: Dublin, Ireland, 2022. [Google Scholar]
- Hodd, R.L.; Hodgetts, N.G. Results of a survey to monitor the EU Annex I habitat Calaminarian grassland, 2018. In Irish Wildlife Manuals, No. 105; National Parks and Wildlife Service, Department of Culture, Heritage and the Gaeltacht: Dublin, Ireland, 2018. [Google Scholar]
- Lockhart, N.; Hodgetts, N.G.; Holyoak, D.T. Rare and Threatened Bryophytes of Ireland; National Museums: Holywood, UK, 2012. [Google Scholar]
- Stanković, J.D.; Sabovljević, A.D.; Sabovljević, M.S. Bryophytes and heavy metals: A review. Acta Bot. Croat. 2018, 77, 109–118. [Google Scholar] [CrossRef]
- Callaghan, D.A.; Samson, L. Population status and ecology of the globally threatened moss Ditrichum plumbicola Crundw. on the Isle of Man. J. Bryol. 2022, 45, 30–37. [Google Scholar] [CrossRef]
- Sassmann, S.; Wernitznig, S.; Lichtscheidl, I.K.; Lang, I. Comparing copper resistance in two bryophytes: Mielichhoferia elongata Hornsch. versus Physcomitrella patens Hedw. Protoplasma 2010, 246, 119–123. [Google Scholar] [CrossRef]
- Hartman, E.L. The ecology of the “copper moss” Mielichhoferia mielichhoferi in Colorado. Bryologist 1969, 72, 56–59. [Google Scholar] [CrossRef]
- Callaghan, D.A. Survival of the critically endangered Ditrichum cornubicum and dependence on conservation management intervention. J. Bryol. 2021, 43, 175–180. [Google Scholar] [CrossRef]
- Callaghan, D.A. Status, conservation and ecology of the exceptionally rare metallophyte Cornish path-moss (Ditrichum cornubicum Paton). J. Bryol. 2018, 40, 358–370. [Google Scholar] [CrossRef]
- Campbell, C.; Kelly, D.L.; Smyth, N.; Lockhart, N.; Holyoak, D.T. Genetic variation in the Red-Listed moss Ditrichum cornubicum Paton (Ditrichaceae) and implications for its conservation. J. Bryol. 2017, 39, 141–151. [Google Scholar] [CrossRef]
- Campbell, C. Conservation of Selected Legally Protected and Red Listed Bryophyte Species in Ireland. Ph.D. Thesis, University of Dublin (Trinity College), Dublin, Ireland, 2013. [Google Scholar]
- Boquete, M.T.; Lang, I.; Weidinger, M.; Richards, C.L.; Alonso, C. Patterns and mechanisms of heavy metal accumulation and tolerance in two terrestrial moss species with contrasting habitat specialization. Environ. Exp. Bot. 2021, 182, 104336. [Google Scholar] [CrossRef]
- Shaw, A.J. Evolution of heavy metal tolerance in bryophytes II. An ecological and experimental investigation of the “copper moss” Scopelophila cataractae (Pottiaceae). Am. J. of Bot. 1987, 74, 813–821. [Google Scholar] [CrossRef]
- Blockeel, T.L.; Bosanquet, S.D.S.; Hill, M.O.; Preston, C.D. Atlas of British and Irish Bryophytes; Pisces Publications: Newbury, UK, 2014. [Google Scholar]
- Hill, M.O.; Preston, C.D. The geographical relationships of British and Irish bryophytes. J. Bryol. 1998, 20, 127–226. [Google Scholar] [CrossRef]
- Hodgetts, N.G. Checklist and country status of European bryophytes–towards a new Red List for Europe. In Irish Wildlife Manuals, No. 84; National Parks and Wildlife Service, Department of Arts, Heritage and the Gaeltacht: Dublin, Ireland, 2015. [Google Scholar]
- Söderström, L.; Urmi, E.; Váňa, J. Distribution of Hepaticae and Anthocerotae in Europe and Macaronesia–update. Cryptogam. Bryol. 2007, 28, 229–350. [Google Scholar]
- Yamada, K.; Iwatsuki, Z. Catalogue of the hepatics of Japan. J. Hattori. Bot. Lab. 2006, 99, 1–106. [Google Scholar]
- Damsholt, K. Illustrated Flora of Nordic Liverworts and Hornworts; Nordic Bryological Society: Lund, Sweden, 2002. [Google Scholar]
- Schuster, R.M. The Hepaticae and Anthrocerotae of North America. Vol. IV; Columbia University Press: New York, NY, USA, 1980. [Google Scholar]
- Ohashi, H. Flora of Eastern Himalaya. Third Report; The University Museum, The University of Tokyo Bulletin 8; The University of Tokyo Press: Tokyo, Japan, 1975. [Google Scholar]
- Meinunger, L.; Schröder, W. Verbreitungsatlas der Moose Deutschlands. Band 1; Dürhammer, O., Ed.; Regensburgische Botanische Gesellschaft: Regensburg, Germany, 2007. [Google Scholar]
- Holyoak, D.T.; Lockhart, N. Notes on some rare and newly recorded bryophytes of metalliferous mine sites in Ireland. J. Bryol. 2009, 31, 267–271. [Google Scholar] [CrossRef]
- Paton, J.A. The Liverwort Flora of the British Isles; Harley Books: Essex, UK, 1999. [Google Scholar]
- Castell, C.P. New vice-county records. Hepaticae. Trans. Brit. Bryol. Soc. 1957, 3, 322–326. [Google Scholar]
- Callaghan, D.A. Notes on Cephaloziella massalongi and C. nicholsonii in Snowdonia. Field Bryol. 2011, 105, 2–9. [Google Scholar]
- Hébrard, J.-P.; Pierrot, R.B.; Quétu, G.; Rogeon, M.A. Contribution à la bryoflore de la Haute Cerdagne et du Capcir. Bull. De La Société Bot. Du Cent.-Ouest 1988, 19, 401–414. [Google Scholar]
- Pierrot, L.; Pierrot, R.B. Cephaloziella massalongi (Spr.) K. Mull. dans les Pyrénées. Le Monde Des Plantes 1971, 15, 370–371. [Google Scholar]
- Hodgetts, N.; Cálix, M.; Englefield, E.; Fettes, N.; García Criado, M.; Patin, L.; Nieto, A.; Bergamini, A.; Bisang, I.; Baisheva, E.; et al. A Miniature World in Decline: European Red List of Mosses, Liverworts and Hornworts; IUCN: Brussels, Belgium, 2019. [Google Scholar]
- Callaghan, D.A. A new IUCN Red List of the bryophytes of Britain. J. Bryol. 2023, 44, 271–389. [Google Scholar] [CrossRef]
- Godfrey, M. Some chromosome counts for the genus Cephaloziella. Field Bryol. 2011, 105, 10–11. [Google Scholar]
- Paton, J.A. Cephaloziella nicholsonii Douin and Schiffn. distinguished from C. massalongi (Spruce) K. Müll. J. Bryol. 1984, 13, 1–8. [Google Scholar] [CrossRef]
- Rouen, S. Substrate Metal Contamination in Relation to Rare Bryophyte Species Distribution at Former Metalliferous Mining Locations in Cornwall; Unpublished Research Project Report as part of BSc (Hons) degree; University of Hertfordshire: Hertfordshire, UK, 2000. [Google Scholar]
- Church, J.M.; Hodgetts, N.G.; Preston, C.D.; Stewart, N.F. British Red Data Books: Mosses and Liverworts; Joint Nature Conservation Council: Peterborough, UK, 2001. [Google Scholar]
- De Agostini, A.; Cogoni, A.; Cortis, P.; Vacca, A.; Becerril, J.M.; Hernández, A.; Esteban, R. Heavy metal tolerance strategies in metallicolous and non-metallicolous populations of mosses: Insights of γ+β-tocopherol regulatory role. Environ. Exp. Bot. 2022, 194, 104738. [Google Scholar] [CrossRef]
- Rowntree, J.K. Development of novel methods for the initiation of in vitro bryophyte cultures for conservation. Plant Cell Tissue Organ Cult. 2006, 87, 191–201. [Google Scholar] [CrossRef]
- Stanković, J.D.; Janković, S.; Lang, I.; Vujičić, M.M.; Sabovljević, M.S.; Sabovljević, A.D. The toxic metal stress in two mosses of different growth forms under axenic and controlled conditions. Bot. Serbica 2021, 45, 31–47. [Google Scholar] [CrossRef]
- Sabovljević, A.; Vujičić, M.; Stanković, J.; Sabovljević, M. Effects of zinc and copper on development and survival of the moss Atrichum undulatum in controlled conditions. Bot. Serbica 2018, 42, 181–184. [Google Scholar]
- Vukojević, V.; Sabovljević, A.; Sabovljević, M. Effect of ferri(III)citrate and potassium hexacyanoferrate(III) on growth of the moss Bryum argenteum Hedw. (Bryaceae) in vitro. Arch. Biol. Sci. 2014, 56, 75–78. [Google Scholar] [CrossRef]
- Blockeel, T.L.; Bell, N.E.; Hill, M.O.; Hodgetts, N.G.; Long, D.G.; Pilkington, S.L.; Rothero, G.P. A new checklist of the bryophytes of Britain and Ireland, 2020. J. Bryol. 2021, 43, 1–51. [Google Scholar] [CrossRef]
- Evans, D.E.; Coleman, J.O.D.; Kearns, A. Plant Cell Culture; BIOS Scientific Publishers Taylor & Francis group: Oxford, UK, 2019. [Google Scholar]
- Brown, D.H. Mineral nutrition. In Bryophyte Ecology; Smith, A.J.E., Ed.; Chapman & Hall: London, UK, 1982. [Google Scholar]
- Wilkins, P. Observations on ecology of Mielichoferia elongata and other ‘copper mosses’ in the British Isles. Bryologist 1977, 80, 175–181. [Google Scholar] [CrossRef]
- Schatz, A. Speculations on the ecology and photosynthesis of the ‘copper mosses’. Bryologist 1955, 58, 113–120. [Google Scholar] [CrossRef]
- Coombes, A.J.; Lepp, N.W. The effect of copper and zinc on the growth of Marchantia polymorpha and Funaria hygrometrica. Bryologist 1974, 77, 447–452. [Google Scholar] [CrossRef]
- Yagura, R.; Imanishi, J.; Shibata, S. Effects of copper ions on the growth and photosynthetic activity of Scopelophila cataractae. Lindbergia 2019, 2019, 1–7. [Google Scholar] [CrossRef]
- Papafilippaki, A.; Gasparatos, D.; Haidouti, C.; Stavroulakis, G. Total and bioavailable forms of Cu, Zn, Pb and Cr in agricultural soils: A study from the hydrological basin of Keritis, Chania, Greece. Glob. NEST J. 2007, 9, 201–206. [Google Scholar]
- Alva, A.K.; Huang, B.; Paramasivam, S. Soil pH affects copper fractionation and phytotoxicity. Soil Sci. Soc. Am. J. 2000, 64, 955–962. [Google Scholar] [CrossRef]
- Shaw, A.J. Heavy Metal Tolerance in Plants: Evolutionary Aspects; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Shaw, A.J.; Antonovics, J.; Anderson, L.E. Inter- and intraspecific variation of mosses in tolerance to copper and zinc. Evolution 1987, 41, 1312–1325. [Google Scholar]
- Brown, D.H.; House, K.L. Evidence of a copper-tolerant ecotype of the hepatic Solenostoma crenulatum. Ann. Bot. 1977, 42, 1383–1392. [Google Scholar] [CrossRef]
- Briggs, D. Population differentiation in Marchantia polymorpha L. in various lead pollution levels. Nature 1972, 238, 166–167. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.J.; Beer, S.C.; Lutz, J. Potential for the evolution of heavy metal tolerance in Bryum argenteum, a moss. I. Variation within and among populations. Bryologist 1989, 92, 73–80. [Google Scholar] [CrossRef]
- Shaw, A.J.; Jules, E.S.; Beer, S.C. Effects of metals on growth, morphology and reproduction of Ceratodon purpureus. Bryologist 1991, 94, 270–277. [Google Scholar] [CrossRef]
- Shaw, A.J. Genetic variation for tolerance to copper and zinc within and among populations of the moss Funaria hygrometrica. New Phytol. 1988, 109, 211–222. [Google Scholar] [CrossRef]
- Shaw, A.J. Adaptation to metals in widespread and endemic plants. Environ. Health Perspect. 1994, 102, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.P.G.; Bradshaw, A.D. Heavy metal tolerance in populations of Agrostis tenuis Sibth. and other grasses. New Phytol. 1965, 64, 131–143. [Google Scholar] [CrossRef]
- Shaw, A.J. Genetic and environmental effects on morphology and asexual reproduction in the moss Bryum bicolor. Bryologist 1990, 93, 1–6. [Google Scholar] [CrossRef]
- Duckett, J.G.; Burch, J.; Fletcher, P.W.; Matcham, H.W.; Read, D.J.; Russell, A.J.; Pressel, S. In vitro cultivation of bryophytes: A review of practicalities, problems, progress and promise. J. Bryol. 2004, 26, 3–20. [Google Scholar]
- Pressel, S.; Ligrone, R.; Duckett, J.G.; Davis, E.C. A novel ascomycetous endophytic association in the rhizoids of the leafy liverwort family, Schistochilaceae (Jungermanniidae, Hepaticopsida). Am. J. Bot. 2008, 95, 531–541. [Google Scholar] [CrossRef]
- Pressel, S.; Bidartondo, M.I.; Ligrone, R.; Duckett, J.G. Fungal symbioses in bryophytes: New insights in the Twenty First Century. Phytotaxa 2010, 9, 238–253. [Google Scholar] [CrossRef]
- Duckett, J.G.; Pressel, S.; Ligrone, R. Cornish bryophytes in the Atlantic Arc: Cell biology, culturing, conservation and climate change. In Botanical Links in the Atlantic Arc, Proceedings of the an Anglo-Hiberno-French Meeting Arranged by the Botanical Society of the British Isles, Camborne, Cornwal, UK, 12–18 May 2003; Leach, S.J., Page, C.N., Peytoureau, Y., Sanford, M.N., Eds.; Botanical Society of the British Isles: Durham, UK, 2006. [Google Scholar]
- Bogdanović, M.; Sabovljević, M.; Sabovljević, A.; Grubišic, D. The influence of gypsiferous substrata on bryophyte growth: Are there obligatory gypsophilous bryophytes? Bot. Serbica 2009, 33, 75–82. [Google Scholar]
- Schuster, R.M. The Hepatiae and Anthocerotae of North America East of the Hundreth Meridian; Columbia University Press: New York, NY, USA, 1992; Volume 6. [Google Scholar]
- Bell, D.; Long, D.; Hollingsworth, P. The Use of DNA Barcoding to Address Major Taxonomic Problems for Rare British Bryophytes; Final Revised Report; Royal Botanic Garden: Edinburgh, Scotland, 2013. [Google Scholar]
- Sawicki, J.; Krawczyk, K.; Ślipiko, M.; Szczecińska, M. Sequencing of Organellar Genomes of Nowellia curvifolia (Cephaloziaceae Jungermanniales) Revealed the Smallest Plastome with Complete Gene Set and High Intraspecific Variation Suggesting Cryptic Speciation. Diversity 2021, 13, 81. [Google Scholar] [CrossRef]
- Myszczyński, K.; Bączkiewicz, A.; Buczkowska, K.; Ślipiko, M.; Szczecińska, M.; Sawicki, J. The extraordinary variation of the organellar genomes of the Aneura pinguis revealed advanced cryptic speciation of the early land plants. Sci. Rep. 2017, 7, 9804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachowiak, W.; Bączkiewicz, A.; Chudzińska, E.; Buczkowska, K. Cryptic speciation in liverworts–A case study in the Aneura pinguis complex. Bot. J. Linn. Soc. 2007, 155, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Batty, L.C. The potential importance of mine sites for biodiversity. Mine Water Environ. 2005, 24, 101–103. [Google Scholar] [CrossRef]
- Kabala, C.; Singh, B.R. Fractionation and mobility of copper, lead and zinc in soil profiles in the vicinity of a copper smelter. J. Environ. Qual. 2001, 30, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Andreu, V.; Gimeno-García, E. Evolution of heavy metals in marsh areas under rice farming. Environ. Pollut. 1999, 104, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Klekowski, E.J., Jr. Reproductive Biology of the Pteridophyta. III. A study of the Blechnaceae. Bot. J. Linn. Soc. 1969, 62, 361–377. [Google Scholar] [CrossRef]
- Abramoff, M.D.; Magelhaes, P.J.; Ram, S.J. Image processing with ImageJ. Biophonetics Int. 2004, 11, 36–42. [Google Scholar]
- Robertson, G.P.; Coleman, D.C.; Bledsoe, C.S.; Sollins, P. Standard Soil Methods for Long-Term Ecological Research; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Shapiro, S.S.; Wilks, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Population | Grid Ref. | Bedrock | Metal(s) Mined | Yrs Worked | pH | Cu | Pb | Zn |
|---|---|---|---|---|---|---|---|---|---|
| Cephaloziella nicholsonii | Caim, Co. Waterford | S885408 | Metamorphic | Cu & Pb | 1815–1855 | 5.83 | 2513 | 37,004 | 1495 |
| Mountain Mine, Allihies, Co. Cork | V589456 | Sandstone | Cu | 1813–1918 | 5.81 | 9877 | 129 | 162 | |
| South Caradon Mine, Cornwall | SX264698 | Granite | Cu | 1837–1885 | ND | ND | ND | ND | |
| Cephaloziella massalongoi | Ross Island, Co. Kerry | V945880 | Limestone | Cu & Pb | 1804–1810 | 6.28 | 3815 | 9911 | 12,647 |
| Glendasan, Co. Wicklow | T098981 | Granite, Schist | Pb | 1807–1900 | 7.03 | 478 | 30,522 | 14,502 | |
| Gunnislake Clitters, Cornwall | SX422720 | Granite | Cu, Sn, Ar & W | 1820–1919 | ND | ND | ND | ND | |
| Bunmahon, Co. Waterford | X443986 | Slate | Cu | 1730–1907 | 5.80 | 12,528 | 34 | 81 | |
| Mountain Mine, Allihies, Co. Cork | V590458 | Sandstone | Cu | 1813–1918 | 5.99 | 6019 | 52 | 32 |
| Source | Degrees of Freedom | Sum of Squares | Mean Square | F-Ratio | p |
|---|---|---|---|---|---|
| Constant | 1 | 162.563 | 162.563 | 239.34 | ≤0.0001 |
| Population | 2 | 23.2821 | 11.641 | 17.139 | ≤0.0001 |
| Treatment | 6 | 78.5549 | 13.0925 | 19.276 | ≤0.0001 |
| Population × Treatment | 12 | 26.9134 | 2.24278 | 3.3021 | 0.0020 |
| Error | 42 | 28.5267 | 0.679206 | ||
| Total | 62 | 157.277 |
| Source | Degrees of Freedom | Sum of Squares | Mean Square | F-Ratio | p |
|---|---|---|---|---|---|
| Constant | 1 | 244.864 | 244.864 | 122.65 | ≤0.0001 |
| Population | 2 | 215.456 | 35.9094 | 17.986 | ≤0.0001 |
| Treatment | 6 | 188.919 | 94.4595 | 47.312 | ≤0.0001 |
| Population × Treatment | 12 | 41.3471 | 3.44559 | 1.7258 | 0.0954 |
| Error | 42 | 83.8533 | 1.99651 | ||
| Total | 62 | 529.576 |
| Treatment | Cephaloziella nicholsonii | Cephaloziella massalongoi | p |
|---|---|---|---|
| 3 ppm | 0.79 (0.19) | 0.44 (0.06) | 0.26 (ns) |
| 6 ppm | 0.81 (0.23) | 0.15 (0.08) | 0.007 ** |
| 12 ppm | 0.51 (0.11) | 0.08 (0.04) | 0.005 ** |
| 24 ppm | 0.22 (0.07) | 0.10 (0.05) | 0.09 (ns) |
| 48 ppm | 0.03 (0.01) | 0.09 (0.04) | 0.67 (ns) |
| 96 ppm | 0.14 (0.04) | 0.05 (0.02) | 0.09 (ns) |
| Chemical Name | Chemical Formula | mg Litre−1 | |
|---|---|---|---|
| Macroelements | Ammonium nitrate 1 | NH4NO3 | 125.0 |
| Potassium dihydrogen phosphate 1 | KH2PO4 | 500.0 | |
| Magnesium sulphate 1 | MgSO4 | 120.0 | |
| Calcium chloride 1 | CaCl2 | 20.0 | |
| Microelements | Sequestrene 330 1 | C14H18FeN3Na2O10·× H2O | 14.0 |
| Boric Acid 1 | H3BO3 | 11.4 | |
| Zinc sulphate 2 | ZnSO4·7H2O | 8.82 | |
| Copper sulphate 2 | CuSO4·5H2O | 1.57 | |
| Manganese sulphate 2 | MnSO4·4H2O | 0.0223 | |
| Sodium molybdate 2 | Na2MoO4·2H2O | 0.0250 | |
| Cobalt chloride 2 | CoCl2·6H2O | 0.0025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, C.; Kelly, D.L.; Smyth, N.; Lockhart, N.; Holyoak, D.T.; Long, D. Investigation of the Copper Requirements of the Metallophyte Liverworts Cephaloziella nicholsonii Douin and C. massalongoi (Spruce) Müll.Frib. Plants 2023, 12, 2265. https://doi.org/10.3390/plants12122265
Campbell C, Kelly DL, Smyth N, Lockhart N, Holyoak DT, Long D. Investigation of the Copper Requirements of the Metallophyte Liverworts Cephaloziella nicholsonii Douin and C. massalongoi (Spruce) Müll.Frib. Plants. 2023; 12(12):2265. https://doi.org/10.3390/plants12122265
Chicago/Turabian StyleCampbell, Christina, Daniel L. Kelly, Noeleen Smyth, Neil Lockhart, David T. Holyoak, and David Long. 2023. "Investigation of the Copper Requirements of the Metallophyte Liverworts Cephaloziella nicholsonii Douin and C. massalongoi (Spruce) Müll.Frib" Plants 12, no. 12: 2265. https://doi.org/10.3390/plants12122265