
Comparative Study of Plastomes in Solanum tuberosum with Different Cytoplasm Types
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Structural Characteristics of Plastid Genomes of Solanum tuberosum Acessions
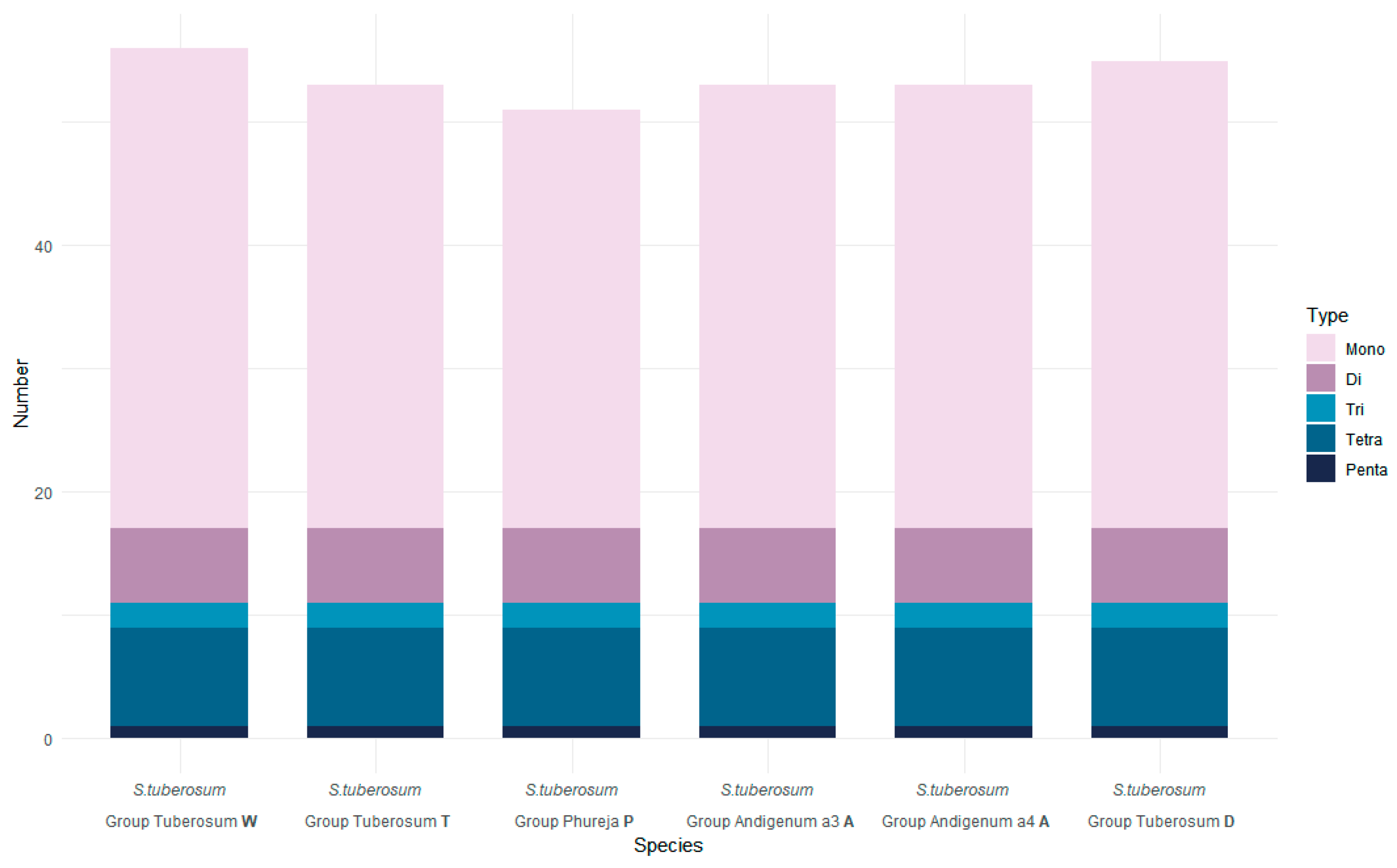
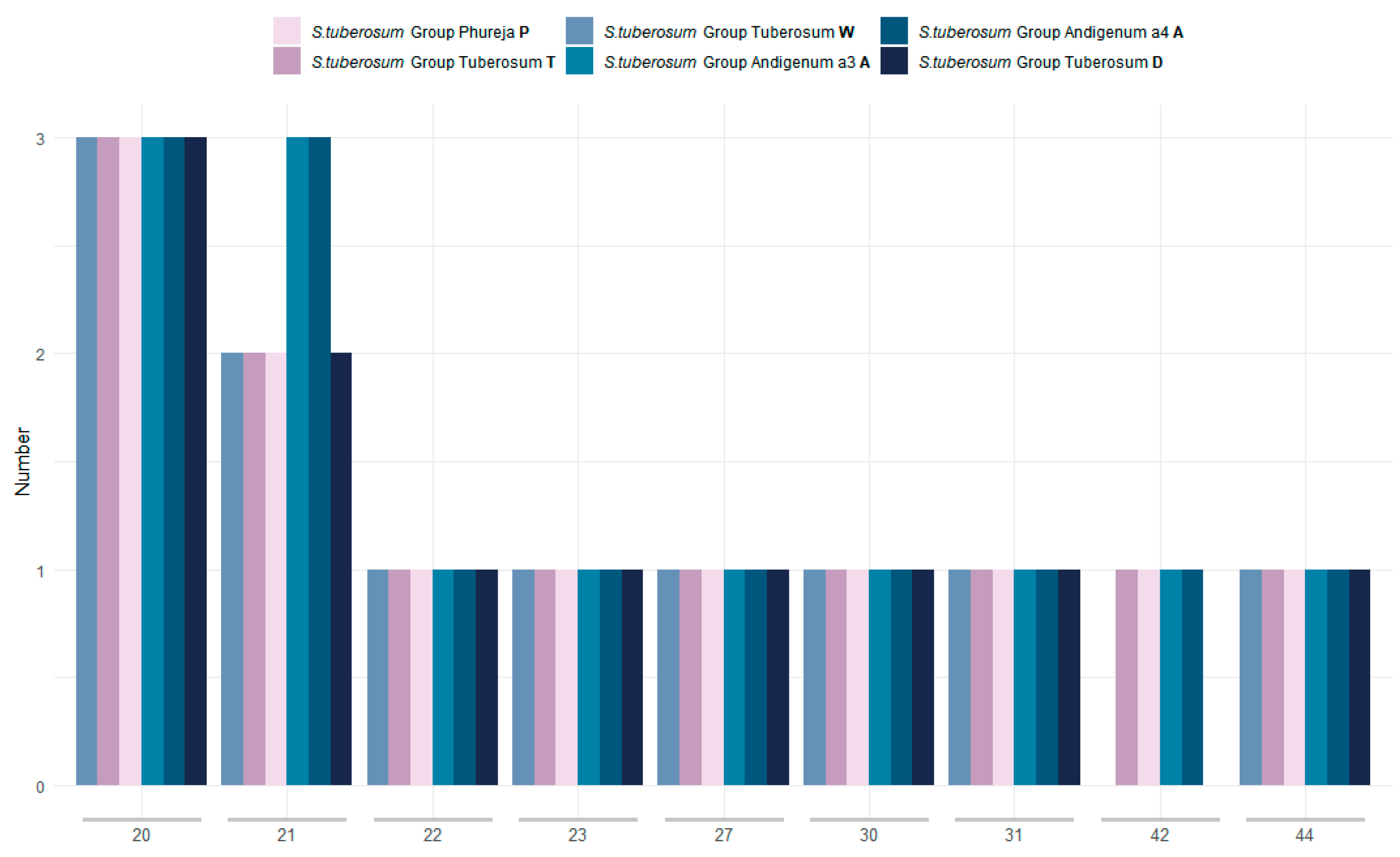
2.2. Repetitive Elements and SSR-Analysis
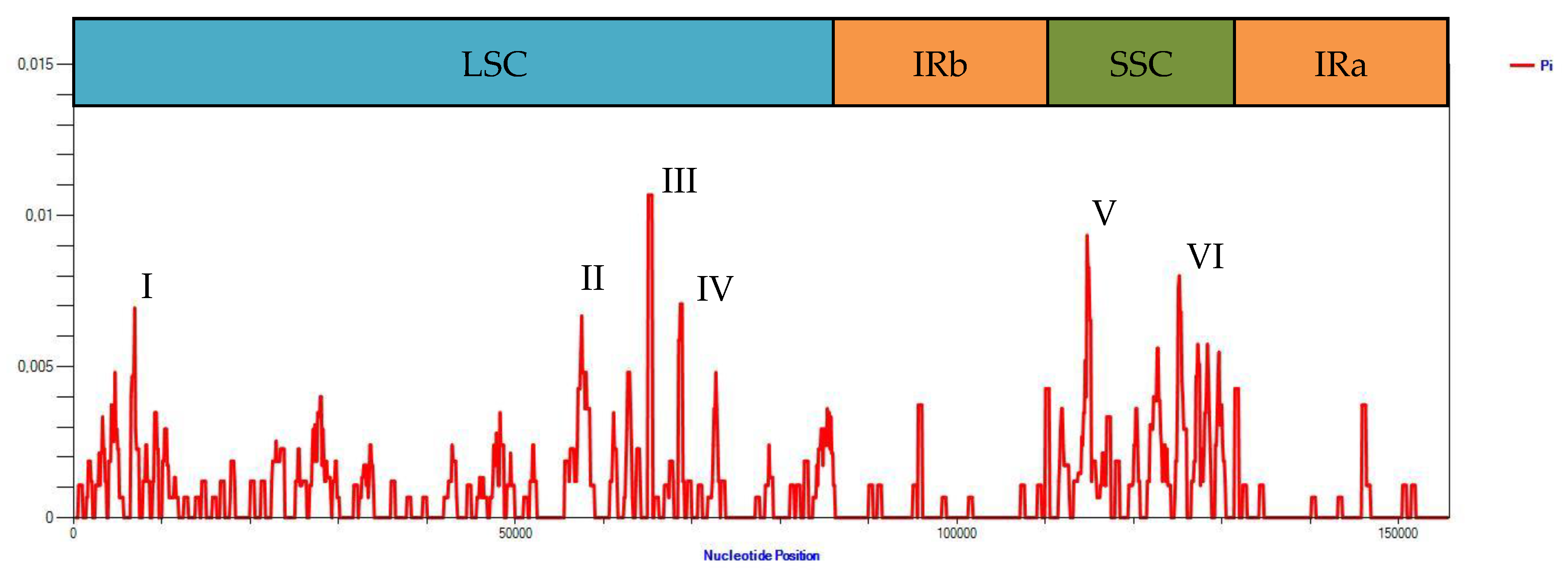
2.3. Comparison and Nucleotide Diversity Analysis of Assembled Genomes
2.4. Localization of Molecular Markers for Identification of Cytoplasm Types (A, T, SAC, D, and S) in Potato Chloroplast Genomes
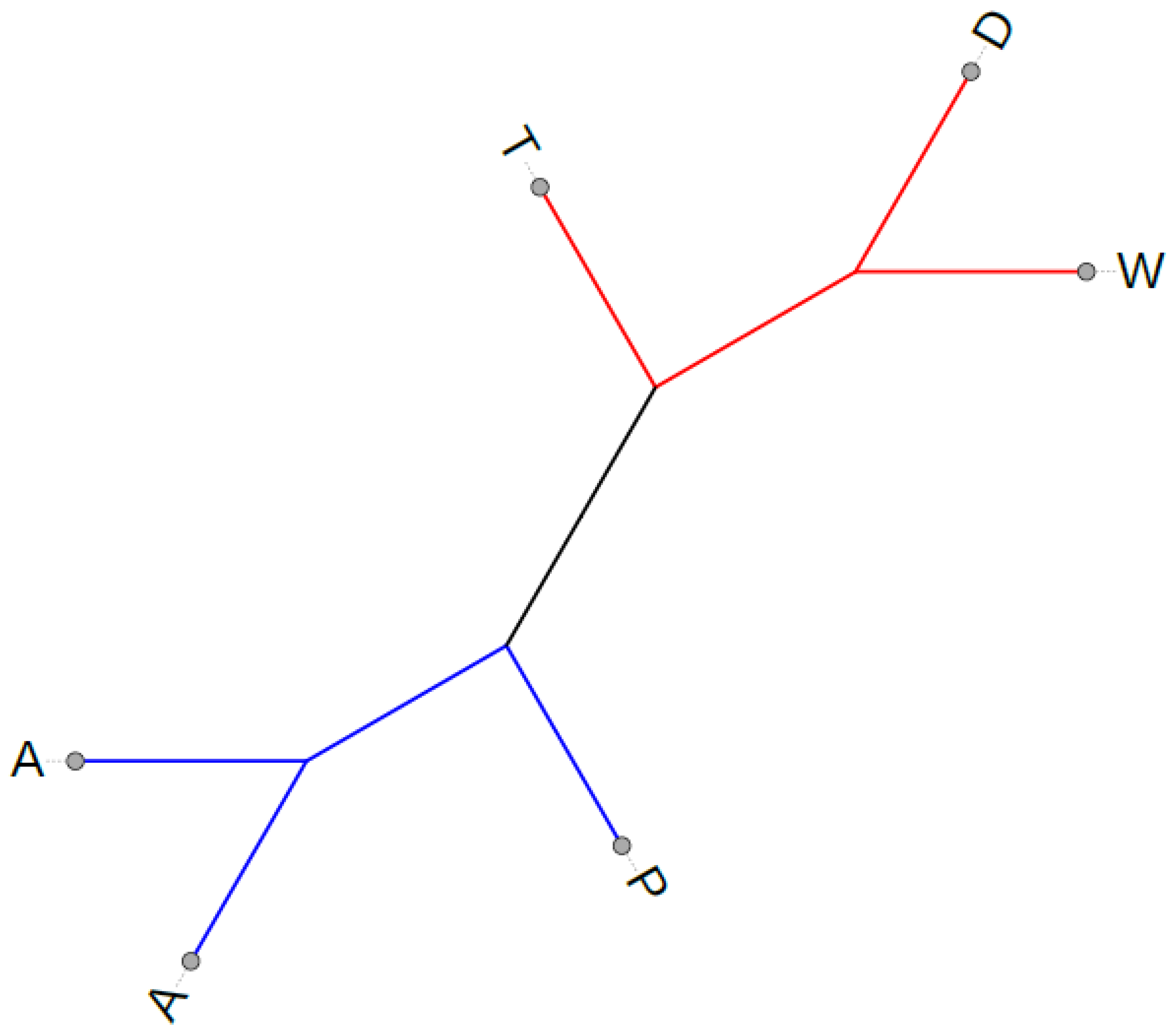
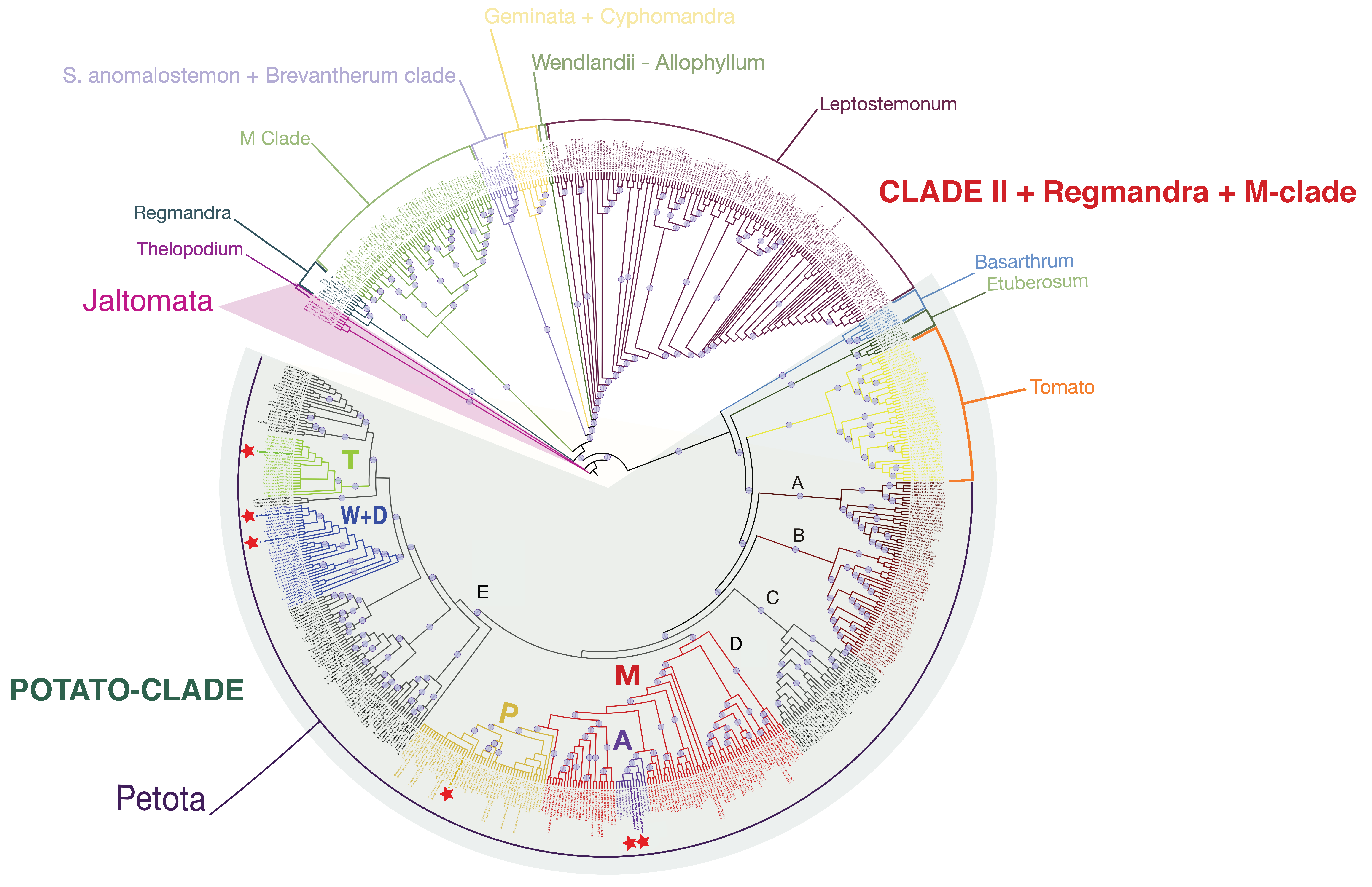
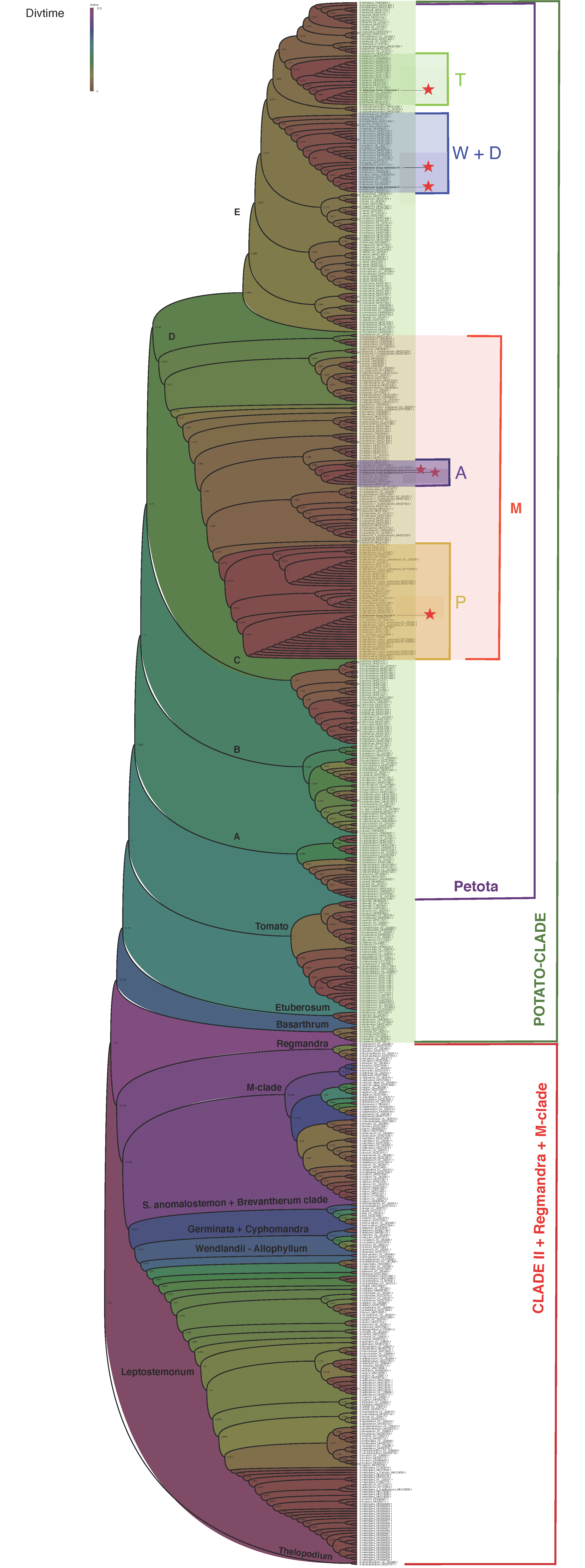
2.5. Potato Chloroplast Genome Phylogeny and Molecular Dating Analyses
2.6. Polymorphism of Protein Sequences Encoded by Potato Chloroplast Genomes
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. DNA Extraction and Sequencing
4.3. Chloroplast Genome Assembly and Annotation
4.4. Simple Sequence Repeats Analysis
4.5. Analysis of Repetitive Elements
4.6. Genome Comparison and Nucleotide Diversity Analysis
4.7. Phylogenetic Reconstruction
4.8. Molecular Dating Analyses
4.9. Adaptive Evolution Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosaka, K.; Sanetomo, R. Development of a rapid identification method for potato cytoplasm and its use for evaluating Japanese collections. Theor. Appl. Genet. 2012, 125, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Lindhout, P.; Meijer, D.A.; Schotte, T.; Hutten, R.C.B.; Visser, R.G.F.; van Eck, H. Towards F1 Hybrid Seed Potato Breeding. Eur. Potato J. 2011, 54, 301–312. [Google Scholar] [CrossRef]
- Jansky, S.H.; Charkowski, A.O.; Douches, D.S.; Gusmini, G. Reinventing Potato as a Diploid Inbred Line–Based Crop. Crop Sci. 2016, 56, 1412–1422. [Google Scholar] [CrossRef]
- Ames, M.; Spooner, D.M. DNA from herbarium specimens settles a controversy about origins of the European potato. Am. J. Bot. 2008, 95, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Simakov, E.A.; Anisimov, B.V.; Sklyarova, N.P.; Yashina, I.M.; Elansky, S.N. Potato Varieties Grown in Russia; NP Russian University of Modern Additional Education for Youth: Moscow, Russian, 2005; p. 112. [Google Scholar]
- Sanetomo, R.; Gebhardt, C. Cytoplasmic genome types of European potatoes and their effects on complex agronomic traits. BMC Plant Biol. 2015, 15, 162. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, K.; Sanetomo, R.; Tamiya, S.; Asano, K.; Shimosaka, E. Creation of long-day adapted tetraploid potatoes from crosses of Solanum tarijense females with Andigena males. Euphytica 2018, 214, 191–220. [Google Scholar] [CrossRef]
- Gavrilenko, T.A.; Klimenko, N.S.; Alpatieva, N.V.; Kostina, L.I.; Lebedeva, V.A.; Evdokimova, Z.Z.; Apalikova, O.V.; Novikova, L.Y.; Antonova, O.Y. Cytoplasmic genetic diversity of potato varieties bred in Russia and FSU countries. Vavilov J. Genet. Breed. 2019, 23, 753–764. [Google Scholar] [CrossRef]
- Vanishree, G.; Patil, V.U.; Kaur, R.P. Cytoplasmic Types of Indian Potato Cultivars and Their Effect on Important Agronomic Traits. Agric. Res. 2022, 11, 390–397. [Google Scholar] [CrossRef]
- Sood, S.; Kumar, A.; Singh, B.; Sundaresha, S.; Bhardwaj, V. Cytoplasmic genome of Indian potato varieties and breeding lines vis a vis prospects in potato breeding. Heliyon 2021, 7, e06365. [Google Scholar] [CrossRef]
- Chimote, V.P.; Chakrabarti, S.K.; Pattanayak, D.; Pandey, S.K.; Naik, P.S. Molecular analysis of cytoplasm type in Indian potato varieties. Euphytica 2008, 162, 69–80. [Google Scholar] [CrossRef]
- Mori, K.; Mukojima, N.; Nakao, T.; Tamiya, S.; Sakamoto, Y.; Sohbaru, N. Germplasm release: Saikai 35, a male and female fertile breeding line carrying Solanum phureja-derived cytoplasm and potato cyst nematode resistance (H1) and Potato virus Y resistance (Rychc ) genes. Am. J. Potato Res. 2012, 89, 63–72. [Google Scholar] [CrossRef]
- Glendinning, D.R. Potato introductions and breeding up to the early 20th century. New Phytol. 1983, 94, 479–505. [Google Scholar] [CrossRef]
- Sanford, J.C.; Hanneman Jr, R.E. Reciprocal differences in the photoperiod reaction of hybrid populations in Solanum tuberosum. Am Potato J. 1979, 56, 531–540. [Google Scholar] [CrossRef]
- Hoopes, R.W.; Plaisted, R.L.; Cubillos, A.G. Yield and fertility of reciprocal-cross Tuberosum-Andigena hybrids. Am. Potato J. 1980, 57, 275–284. [Google Scholar] [CrossRef]
- Sanford, J.C.; Hanneman Jr, R.E. Large yield differences between reciprocal families of Solanum tuberosum. Euphytica 1982, 31, 1–12. [Google Scholar] [CrossRef]
- Hilali, A.; Lauer, F.I.; Veilleux, R.E. Reciprocal differences between hybrids of Solanum tuberosum Groups Tuberosum (haploid) and Phureja. Euphytica 1987, 36, 631–639. [Google Scholar] [CrossRef]
- Maris, B. Analysis of an incomplete diallel cross among three ssp. Tuberosum varieties and seven long-day adapted ssp. andigena clones of the potato (Solanum tuberosum L.). Euphytica 1989, 41, 163–182. [Google Scholar] [CrossRef]
- Grun, P.; Ochoa, C.; Capage, D. Evolution of cytoplasmic factors in tetraploid cultivated potatoes (Solanaceae). Am. J. Bot. 1977, 64, 412–420. [Google Scholar] [CrossRef]
- Grun, P. Cytoplasmic sterilities that separate the group tuberosum cultivated potato from its putative tetraploid ancestor. Evolution 1973, 27, 633–643. [Google Scholar] [CrossRef]
- Sanetomo, R.; Ono, S.; Hosaka, K. Characterization of cross- ability in the crosses between Solanum demissum and S. tuberosum, and the F1 and BC1 progenies. Am. J. Potato Res. 2011, 88, 500–510. [Google Scholar] [CrossRef]
- Ross, H.; Hunnius, W. Potato Breeding—Problems and Perspectives. In Fortschritte der Pflanzenzüchtung; V. P. Parey: Berlin, German, 1986; p. 132. [Google Scholar]
- Plaisted, R.L.; Hoopes, R.W. The past record and future prospects for the use of exotic potato germplasm. Am. Potato J. 1989, 66, 603–627. [Google Scholar] [CrossRef]
- Dionne, L.A. Cytoplasmic sterility in derivatives of Solanum demissum. Am. Potato J. 1961, 38, 117–120. [Google Scholar] [CrossRef]
- Song, Y.S.; Schwarzfischer, A. Development of STS markers for selection of extreme resistance (Rysto) to PVY and maternal pedigree analysis of extremely resistant cultivars. Am. J. Potato Res. 2008, 85, 159–170. [Google Scholar] [CrossRef]
- Lössl, A.; Götz, M.; Braun, A.; Wenzel, G. Molecular markers for cytoplasm in potato: Male sterility and contribution of different plastid-mitochondrial configurations to starch production. Euphytica 2000, 116, 221–230. [Google Scholar] [CrossRef]
- Hosaka, K. Distribution of the 241 bp deletion of chloroplast DNA in wild potato species. Am. J. Potato Res. 2002, 79, 119–123. [Google Scholar] [CrossRef]
- Hosaka, K.; Hanneman Jr, R.E. The origin of the cultivated tetraploid potato based on chloroplast DNA. Theor. Appl. Genet. 1988, 76, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Kawagoe, Y.; Kikuta, Y. Chloroplast DNA evolution in potato (Solanum tuberosum L.). Theor. Appl. Genet. 1991, 81, 13–20. [Google Scholar] [CrossRef]
- Bryan, G.J.; McNicoll, J.; Ramsay, G.; Meyer, R.C.; De Jong, W.S. Polymorphic simple sequence repeat markers in chloroplast genomes of Solanaceous plants. Theor. Appl. Genet. 1999, 99, 859–867. [Google Scholar] [CrossRef]
- Provan, J.; Powell, W.; Dewar, H.; Bryan, G.; Machray, G.C.; Waugh, R. An extreme cytoplasmic bottleneck in the modern European cultivated potato (Solanum tuberosum) is not reflected in decreased levels of nuclear diversity. Proc. R. Soc. Lond. 1999, 266, 633–639. [Google Scholar] [CrossRef]
- Sanetomo, R.; Hosaka, K. A maternally inherited DNA marker, descended from Solanum demissum (2n = 6x = 72) to S. tuberosum (2n = 4x = 48). Breed. Sci. 2011, 61, 426–434. [Google Scholar] [CrossRef]
- Chung, H.J.; Jung, J.D.; Park, H.W.; Kim, J.H.; Cha, H.W.; Min, S.R.; Jeong, W.-J.; Liu, J.R. The complete chloroplast genome sequences of Solanum tuberosum and comparative analysis with Solanaceae species identified the presence of a 241- bp deletion in cultivated potato chloroplast DNA sequence. Plant Cell Rep. 2006, 25, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Ruess, H.; Liang, Q.; Colleoni, C.; Spooner, D.M. Analyses of 202 plastid genomes elucidate the phylogeny of Solanum section Petota. Sci. Rep. 2019, 9, 4454. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, E.; Hilgenhof, R.; Orejuela, A.; McDonnell, A.; Sablok, G.; Aubriot, X.; Giacomin, L.; Gouvêa, Y.; Bragionis, T.; Stehmann, J.R.; et al. Phylogenomic discordance suggests polytomies along the backbone of the large genus Solanum. Am. J. Bot. 2022, 109, 580–601. [Google Scholar] [CrossRef] [PubMed]
- Särkinen, T.; Bohs, L.; Olmstead, R.; Knapp, S. A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): A dated 1000-tip tree. BMC Evol. Biol. 2013, 13, 214. [Google Scholar] [CrossRef]
- Knight, S.; Andersson, I.; Brindn, C.-I. Crystallographic Analysis of Ribulose 1,5-Bisphosphate Carboxylase from Spinach at 2.4 A Resolution Subunit Interactions and Active Site. J. Mol. Biol. 1990, 215, 113–160. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 4th ed.; WH Freeman and Company: New York, NY, USA, 2005; p. 1120. [Google Scholar]
- Olmstead, R.; Santiago-Valentin, E.; Bohs, L.; Migid, A.A. Molecular phylogeny of Solanaceae. TAXON 2008, 57, 1159–1181. [Google Scholar] [CrossRef]
- Jansky, S.H.; Roble, J.; Spooner, D.M. Solanum clarum and S. morelliforme as Novel Model Species for Studies of Epiphytism. Front. Plant Sci. 2016, 7, 231. [Google Scholar] [CrossRef]
- Simon, R.; Fuentes, A.F.; Spooner, D.M. Biogeographic implications of the striking discovery of a 4,000 kilometer disjunct population of the wild potato Solanum morelliforme in South America. Syst. Bot. 2011, 36, 1062–1067. [Google Scholar] [CrossRef]
- Sukhotu, T.; Kamijima, O.; Hosaka, K. Nuclear and chloroplast DNA differentiation in Andean potatoes. Genome 2004, 47, 46–56. [Google Scholar] [CrossRef]
- Hosaka, K.; Sanetomo, R. Comparative differentiation in mitochondrial and chloroplast DNA among cultivated potatoes and closely related wild species. Genes Genet. Syst. 2009, 84, 371–378. [Google Scholar] [CrossRef]
- Spooner, D.M.; Hetterscheid, W.L.A. Origins, evolution, and group classification of cultivated potatoes. In Darwin’s Harvest: New Approaches to the Origins, Evolution, and Conservation of Crops; Motley, T.J., Zerega, N., Cross, H., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 285–307. [Google Scholar]
- Spooner, D.M.; McLean, K.; Ramsay, G.; Waugh, R.; Bryan, G.J. A single domestication for potato based on multilocus AFLP genotyping. Proc. Natl. Acad. Sci. USA 2005, 102, 14694–14699. [Google Scholar] [CrossRef] [PubMed]
- Hardigan, M.A.; Parker, F.; Newtow, L.; Buell, C.R. Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proc. Natl. Acad. Sci. USA 2017, 114, E9999–E10008. [Google Scholar] [CrossRef] [PubMed]
- Spooner, D.; Glenn, D. Species Limits of Solanum berthaultii Hawkes and S. tarijense Hawkes and the Implications for Species Boundaries in Solanum Sect.Petota. Taxon 2007, 56, 987–999. [Google Scholar] [CrossRef]
- Hosaka, K. T-type chloroplast DNA in Solanum tuberosum L. ssp. tuberosum was conferred from some populations of S. tarijense Hawkes. Am. J. Potato Res. 2003, 80, 21–32. [Google Scholar] [CrossRef]
- Gavrilenko, T.; Antonova, O.; Shuvalova, A.; Krylova, E.; Alpatyeva, N.; Spooner, D.M.; Novikova, L. Genetic diversity and origin of cultivated potatoes based on plastid microsatellite polymorphism T. Genet. Resour. Crop Evol. 2013, 60, 1997–2015. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Spooner, D.M.; Salas, A.R.; Guarino, L.; de la Cruz, J. Atlas of wild potatoes. In Systematic and Ecogeographic Studies in Crop Genepools 8; IPGRI: Rome, Italy, 2002; p. 130. [Google Scholar]
- Hosaka, K. Evolutionary pathway of T-type chloroplast DNA in potato. Am. J. Potato Res. 2004, 81, 153–158. [Google Scholar] [CrossRef]
- Potato Pedigree Database. Available online: https://www.plantbreeding.wur.nl/PotatoPedigree/pedigree_imagemap.php?id=17289 (accessed on 31 October 2023).
- Spreitzer, R.J.; Salvucci, M.E. Rubisco: Structure, regulatory interactions, and possibilities for a better enzyme. Annu. Rev. Plant Biol. 2002, 53, 449–475. [Google Scholar] [CrossRef]
- Sen, L.; Gao, L.; Fares, M. Molecular evolution of rbcL in three gymnosperm families: Identifying adaptive and coevolutionary patterns. Biol. Direct 2011, 6, 29. [Google Scholar] [CrossRef]
- Kapralov, M.V.; Filatov, D.A. Molecular Adaptation during Adaptive Radiation in the Hawaiian Endemic Genus Schiedea. PLoS ONE 2006, 1, e8. [Google Scholar] [CrossRef]
- Andrews, S. Fast QC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 6 January 2023).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. Multi QC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Jiang, H.; Lei, R.; Ding, S.-W.; Zhu, S. Skewer: A fast and accurate adapter trimmer for next-generation sequencing paired-end reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Zheng, S.; Poczai, P.; Hyvönen, J.; Tang, J.; Amiryousefi, A. Chloroplot: An online program for the versatile plotting of organelle genomes. Front. Genet. 2020, 11, 1123. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L. GMATA: An Integrated Software Package for Genome-Scale SSR Mining, Marker Development and Viewing. Front. Plant Sci. 2016, 7, 1350. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; p. 268. ISBN 978-3-319-24277-4. [Google Scholar]
- Kaupp, J. nord: Arctic Ice Studio’s Nord and Group of Seven Inspired Colour Palettes for ggplot2. R Package Version 1.0.0. 2019. Available online: https://CRAN.R-project.org/package=nord (accessed on 31 October 2023).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2023. Available online: https://www.R-project.org (accessed on 31 October 2023).
- Rozas, J.; Ferrer-Mata, A.; Librado, P.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Entfellner, D.J.-B.; Wilkinson, E.; Correia, D.; Davila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Peterson, D.; Tamura, K. MEGA-CC: Computing core of molecular evolutionary genetics analysis program for automated and iterative data analysis. Bioinformatics 2012, 28, 2685–2686. [Google Scholar] [CrossRef]
- Tamura, K.; Battistuzzi, F.U.; Billing-Ross, P.; Murillo, O.; Filipski, A.; Kumar, S. Estimating divergence times in large molecular phylogenies. Proc. Natl. Acad. Sci. USA 2012, 109, 19333–19338. [Google Scholar] [CrossRef]
- Tamura, K.; Tao, Q.; Kumar, S. Theoretical Foundation of the RelTime Method for Estimating Divergence Times from Variable Evolutionary Rates. Mol. Biol. Evol. 2018, 35, 1770–1782. [Google Scholar] [CrossRef]
- FigTree v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 31 October 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | Type of Cytoplasm | Genbank Accession Number | Read Pairs Number | Average Genome Coverage | Sequence Length, bp | |||
|---|---|---|---|---|---|---|---|---|
| IR | LSC | SSC | Total | |||||
| S. tuberosum Group Tuberosum cv. Nakra | W | OR632697 | 3,302,337 | 784× | 25,592 | 85,991 | 18,374 | 155,549 |
| S. tuberosum Group Tuberosum cv. Vitelotte | T | OR632698 | 7,906,999 | 761× | 25,593 | 85,737 | 18,373 | 155,296 |
| S. tuberosum Group Tuberosum cv. Barin | D | OR632702 | 8,790,748 | 697× | 25,593 | 86,003 | 18,373 | 155,562 |
| S. tuberosum Group Phureja | P | OR632699 | 5,036,719 | 804× | 25,593 | 85,930 | 18,376 | 155,492 |
| S. tuberosum Group Andigenum a3 | A | OR632700 | 1,887,454 | 531× | 25,593 | 85,968 | 18,364 | 155,518 |
| S. tuberosum Group Andigenum a4 | A | OR632701 | 5,751,620 | 778× | 25,593 | 85,967 | 18,364 | 155,517 |
| Marker | Primer Sequence | References | Position in Chloroplast Genome of S. tuberosum cv. Barin with D Type of Cytoplasm |
|---|---|---|---|
| T | GGAGGGGTTTTTCTTGGTTG | [27] | 52,459–52,478 |
| AAGTTTACTCACGGCAATCG | 52,882–52,901 | ||
| S | GGTTCGAATCCTTCCGTC | [31] | 7282–7299 |
| GATTCTTTCGCATCTCGATTC | GATTCTTTCGC/TATCTCGATTC 7126–7146 | ||
| SAC | TTGGAGTTGTTGCGAATGAG | [1] | 63,681–63,700 |
| GTTCCCTAGCCACGATTCTG | 63,973–63,992 | ||
| D | CGGGAGGTGGTGTACTTTCT | [32] | not found |
| ACGGCTGACTGTGTGTTTGA | not found | ||
| A | AACTTTTTGAACTCTATTCCTTAATTG | [1] | 115,410–115,436 |
| ACGCTTCATTAGCCCATACC | 116,577–116,596 |
| Function | Gene | Length, bp | Substitutions Number | Alignment Position | Cytoplasm Type | A/T Genomes Substitution | (A + P)/(T + W + D) Genomes Substitution | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P | A | D | W | T | |||||||
| ATP synthase | atpB | 498 | 1 | 99 | S | S | R | R | R | S/R | S/R |
| Cytochrome c biogenesis protein | ccsA | 313 | 1 | 199 | K | K | Q | Q | K | ||
| Maturase K | matK | 509 | 3 | 54 | F | F | L | L | L | F/L | F/L |
| 191 | L | L | W | W | L | ||||||
| 333 | D | D | N | N | D | ||||||
| NADH dehydrogenase | ndhA | 363 | 1 | 31 | I | I | V | V | V | I/V | I/V |
| ndhB | 510 | 1 | 70 | M | M | M | M | L | M/L | ||
| ndhF | 739 | 3 | 466 | G | G | G | G | D | G/D | ||
| 649 | L | L | I | L | L | ||||||
| 677 | L | L | F | F | L | ||||||
| ndhG | 176 | 1 | 44 | V | V | I | I | I | V/I | V/I | |
| Photosystem II | psbC | 461 | 1 | 425 | L | L | F | F | F | L/F | L/F |
| Rubisco large subunit | rbcL | 477 | 7 | 117 | F | F | L | L | L | F/L | F/L |
| 142 | T | T | V | V | V | T/V | T/V | ||||
| 230 | T | T | A | A | A | T/A | T/A | ||||
| 262 | V | V | T | T | T | V/T | V/T | ||||
| 309 | I | I | M | M | M | I/M | I/M | ||||
| 439 | Q | Q | R | R | R | Q/R | Q/R | ||||
| 449 | C | C | A | A | A | C/A | C/A | ||||
| DNA dependent RNA polymerase | rpoB | 1070 | 2 | 31 | V | V | L | L | L | V/L | V/L |
| 587 | I | I | V | V | I | ||||||
| rpoC1 | 688 | 1 | 668 | V | V | A | A | A | V/A | V/A | |
| rpoC2 | 1392 | 1 | 1052 | F | F | V | V | V | F/V | F/V | |
| Large subunit of ribosome | rpl14 | 122 | 1 | 57 | I | I | V | V | V | I/V | I/V |
| rpl20 | 128 | 1 | 65 | I | I | L | L | I | |||
| Small subunit of ribosome | rps11 | 138 | 1 | 77 | A | A | V | V | A | ||
| rps15 | 87 | 2 | 13 | E | E | K | K | K | E/K | E/K | |
| 80 | E | E | D | D | D | E/D | E/D | ||||
| rps3 | 218 | 2 | 103 | L | L | F | F | F | L/F | L/F | |
| 217 | E | E | A | A | E | ||||||
| rps4 | 201 | 1 | 39 | V | G | G | G | G | |||
| Proteins of unknown function | ycf1 | 1887 | 10 | 360 | F | F | L | L | L | F/L | F/L |
| 487 | T | T | K | K | K | T/K | T/K | ||||
| 581 | K | K | E | E | E | K/E | K/E | ||||
| 972 | F | F | L | L | L | F/L | F/L | ||||
| 1008 | I | I | L | L | L | I/L | I/L | ||||
| 1116 | I | I | K | K | K | I/K | I/K | ||||
| 1258 | D | D | N | N | D | ||||||
| 1319 | Q | Q | Q | Q | K | Q/K | |||||
| 1321 | K | K | Q | Q | K | ||||||
| 1345 | S | L | S | S | S | L/S | |||||
| ycf2 | 2278 | 1 | 606 | Y | Y | S | S | Y | |||
| 29 | 25 | ||||||||||
| Gene | Ka | Ks | Ka/Ks |
|---|---|---|---|
| atpB | 0.0005 | 0.0009 | 0.55556 |
| ccsA | 0.0007 | 0.0025 | 0.28 |
| matK | 0.0014 | 0.0015 | 0.93333 |
| ndhA | 0.0007 | 0.0044 | 0.15909 |
| ndhB | 0.0003 | 0 | NA |
| ndhF | 0.0007 | 0.0029 | 0.24138 |
| ndhG | 0.0015 | 0.0047 | 0.31915 |
| psbC | 0.0006 | 0 | NA |
| rbcL | 0.0056 | 0.001 | 5.6 |
| rpl15 | 0.0022 | 0.0038 | 0.57895 |
| rpl20 | 0.0018 | 0 | NA |
| rpoB | 0.0005 | 0.0023 | 0.21739 |
| rpoC1 | 0.0004 | 0.0019 | 0.21053 |
| rpoC2 | 0.0002 | 0.001 | 0.2 |
| rps3 | 0.0022 | 0.0064 | 0.34375 |
| rps4 | 0.0007 | 0 | NA |
| rps11 | 0.0018 | 0 | NA |
| rps15 | 0.006 | 0.0093 | 0.64516 |
| ycf1 | 0.0016 | 0.0042 | 0.38095 |
| ycf2 | 0.0001 | 0.0004 | 0.25 |
| Codon No | Amino Acid Changes | Type of Changes a | Location of Residue b | |
|---|---|---|---|---|
| M + A + P | T + W + D | |||
| 117 | F | L | HR => HN | -helix C |
| 142 | T | V | UP => HN | -helix D |
| 230 | T | A | UP => HN | -helix 2 |
| 262 | V | T | HN => UP | between helix 3 и-strand 4 |
| 309 | I | M | HN => HN | -strand F |
| 439 | Q | R | UP => UB | -helix G |
| 449 | C | A | UP => HN | -helix G |
| GenBank No | Lengh, bp | Haplotype | Sample |
|---|---|---|---|
| DQ231562 | 155312 | h1 | Solanum tuberosum L. cv. Desiree |
| MH021416 | 155296 | h2 | Solanum berthaultii voucher PI 498105 |
| MT511702 | 155296 | h2 | Solanum tuberosum chloroplast clone 7506-01 (2n) |
| MW307947 | 155296 | h2 | Solanum tuberosum cultivar Atlantic |
| MZ030720 | 155296 | h2 | Solanum tuberosum cultivar Atlantic |
| NC_008096 | 155296 | h2 | Solanum tuberosum cv. Desiree |
| OR632698 | 155296 | h2 | Solanum tuberosum cv. Vitelotte (this study) |
| MT511703 | 155296 | h3 | Solanum tuberosum chloroplast clone 08675-21 (2n) |
| MT511708 | 155296 | h3 | Solanum tuberosum chloroplast clone DW84-1457 (2n) |
| MT511709 | 155296 | h3 | Solanum tuberosum chloroplast clone H412-1 (2n) |
| MW307946 | 155296 | h3 | Solanum tuberosum cultivar Yanshu No. 4 |
| MW307948 | 155296 | h3 | Solanum tuberosum cultivar Favorita |
| MW307949 | 155296 | h3 | Solanum tuberosum cultivar Shepody |
| MZ030723 | 155296 | h3 | Solanum tuberosum cultivar Colomba |
| MZ030724 | 155296 | h3 | Solanum tuberosum cultivar Spunta |
| KM489056 | 155296 | h4 | Solanum tuberosum |
| MH021575 | 155297 | h5 | Solanum tarijense |
| MH021576 | 155295 | h6 | Solanum tarijense |
| OM638071 | 155295 | h6 | Solanum tarijense |
| MH021573 | 155299 | h7 | Solanum tarijense |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goryunova, S.; Sivolapova, A.; Polivanova, O.; Sotnikova, E.; Meleshin, A.; Gaitova, N.; Egorova, A.; Semenov, A.; Gins, E.; Koroleva, A.; et al. Comparative Study of Plastomes in Solanum tuberosum with Different Cytoplasm Types. Plants 2023, 12, 3995. https://doi.org/10.3390/plants12233995
Goryunova S, Sivolapova A, Polivanova O, Sotnikova E, Meleshin A, Gaitova N, Egorova A, Semenov A, Gins E, Koroleva A, et al. Comparative Study of Plastomes in Solanum tuberosum with Different Cytoplasm Types. Plants. 2023; 12(23):3995. https://doi.org/10.3390/plants12233995
Chicago/Turabian StyleGoryunova, Svetlana, Anastasia Sivolapova, Oksana Polivanova, Evgeniia Sotnikova, Alexey Meleshin, Natalia Gaitova, Anna Egorova, Anatoly Semenov, Ekaterina Gins, Alina Koroleva, and et al. 2023. "Comparative Study of Plastomes in Solanum tuberosum with Different Cytoplasm Types" Plants 12, no. 23: 3995. https://doi.org/10.3390/plants12233995