
Integrative Taxonomy of Nitraria (Nitrariaceae), Description of the New Enigmatic Species and Key to All Currently Known Species
 ,
,
Abstract
:1. Introduction
2. Results and Discussion
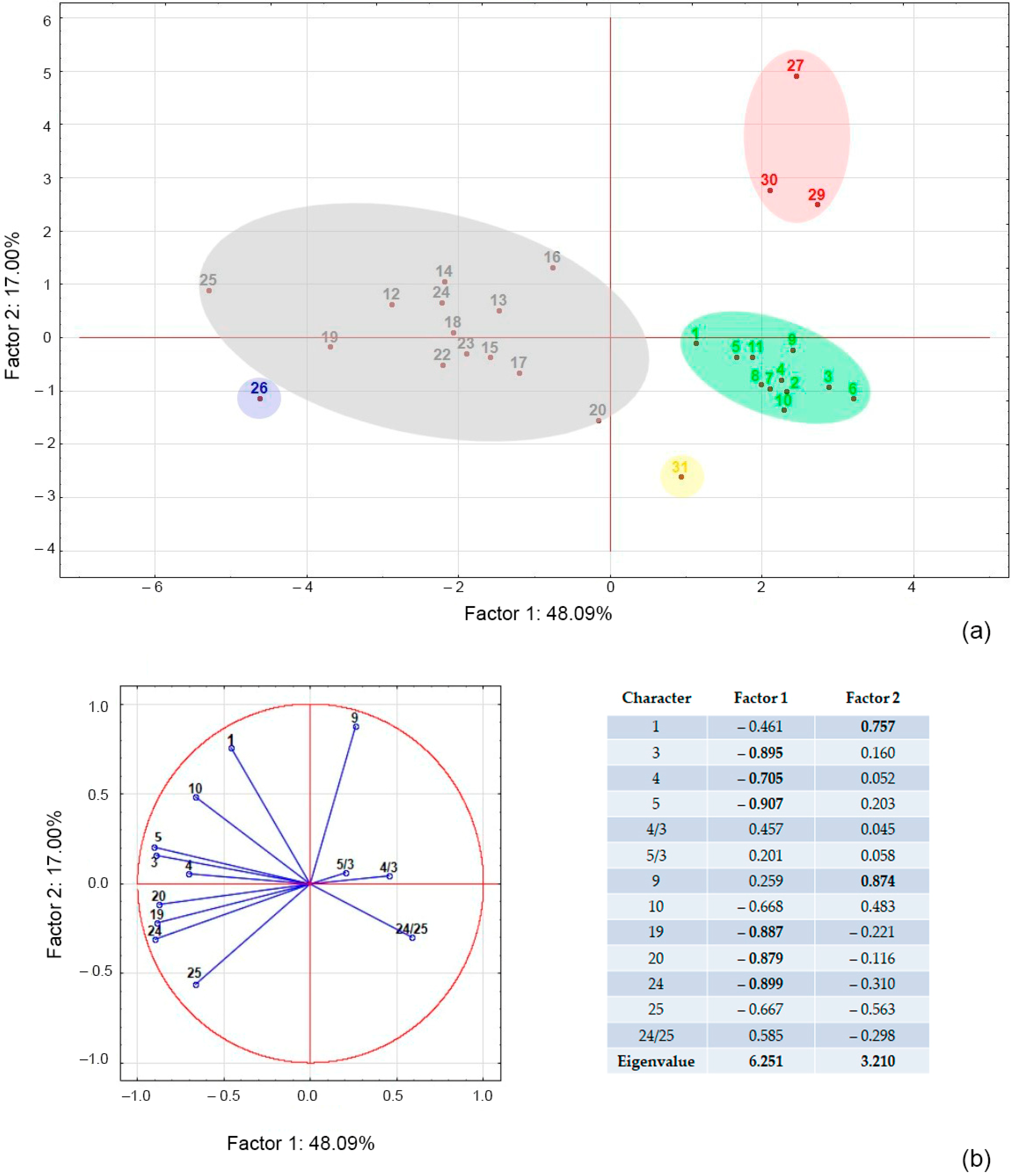
2.1. Morphological Analysis
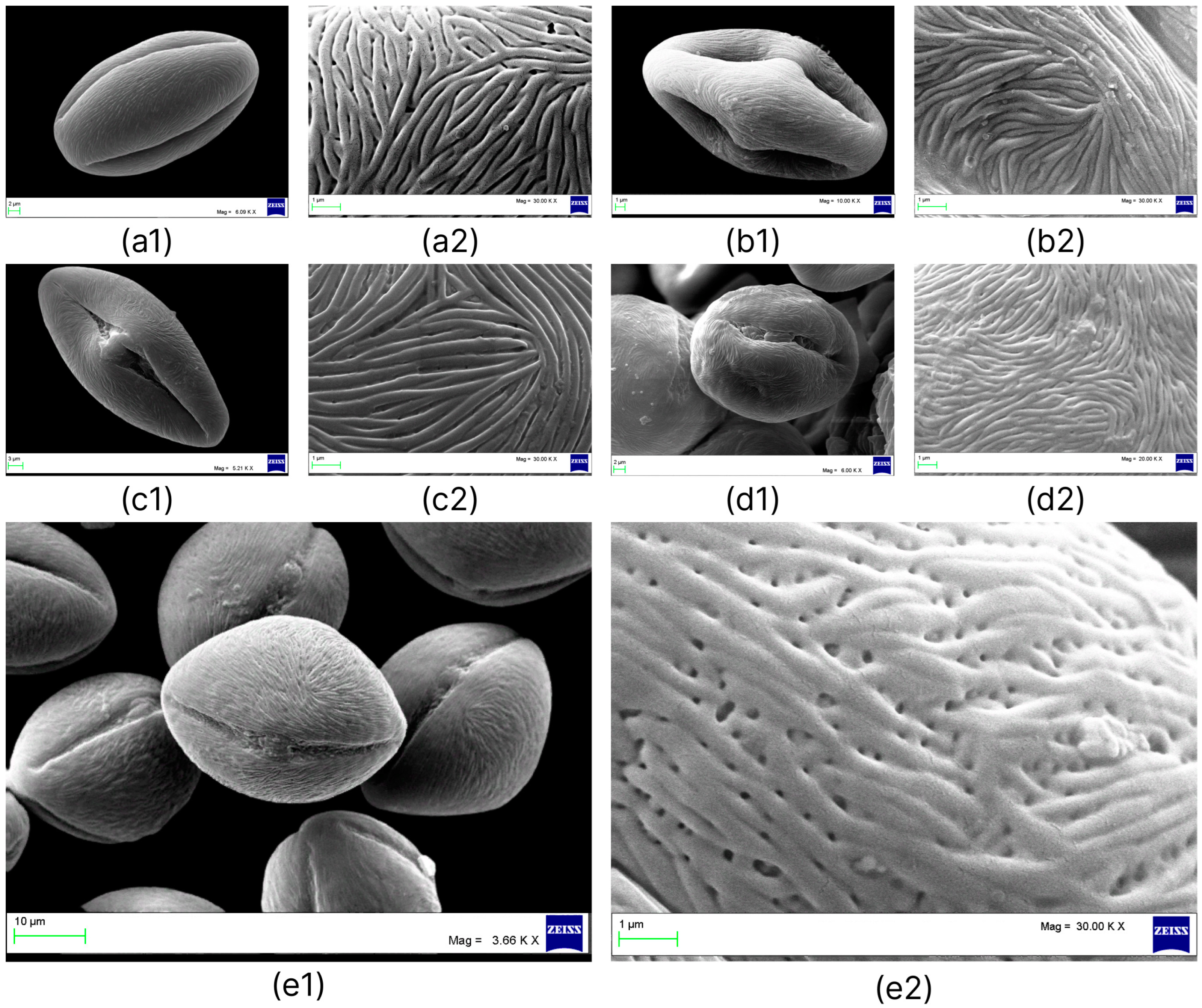
2.2. Palynological Analysis
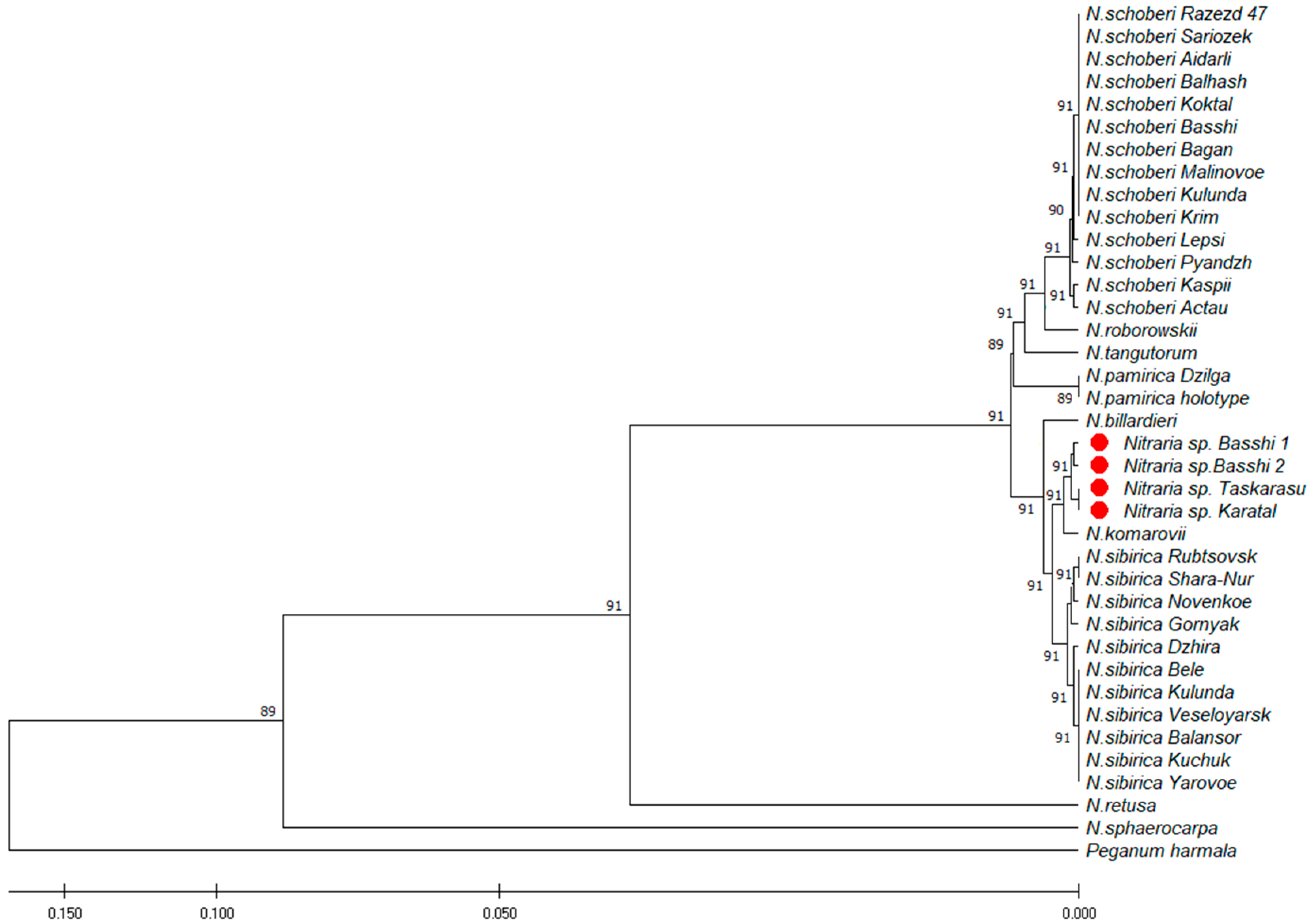
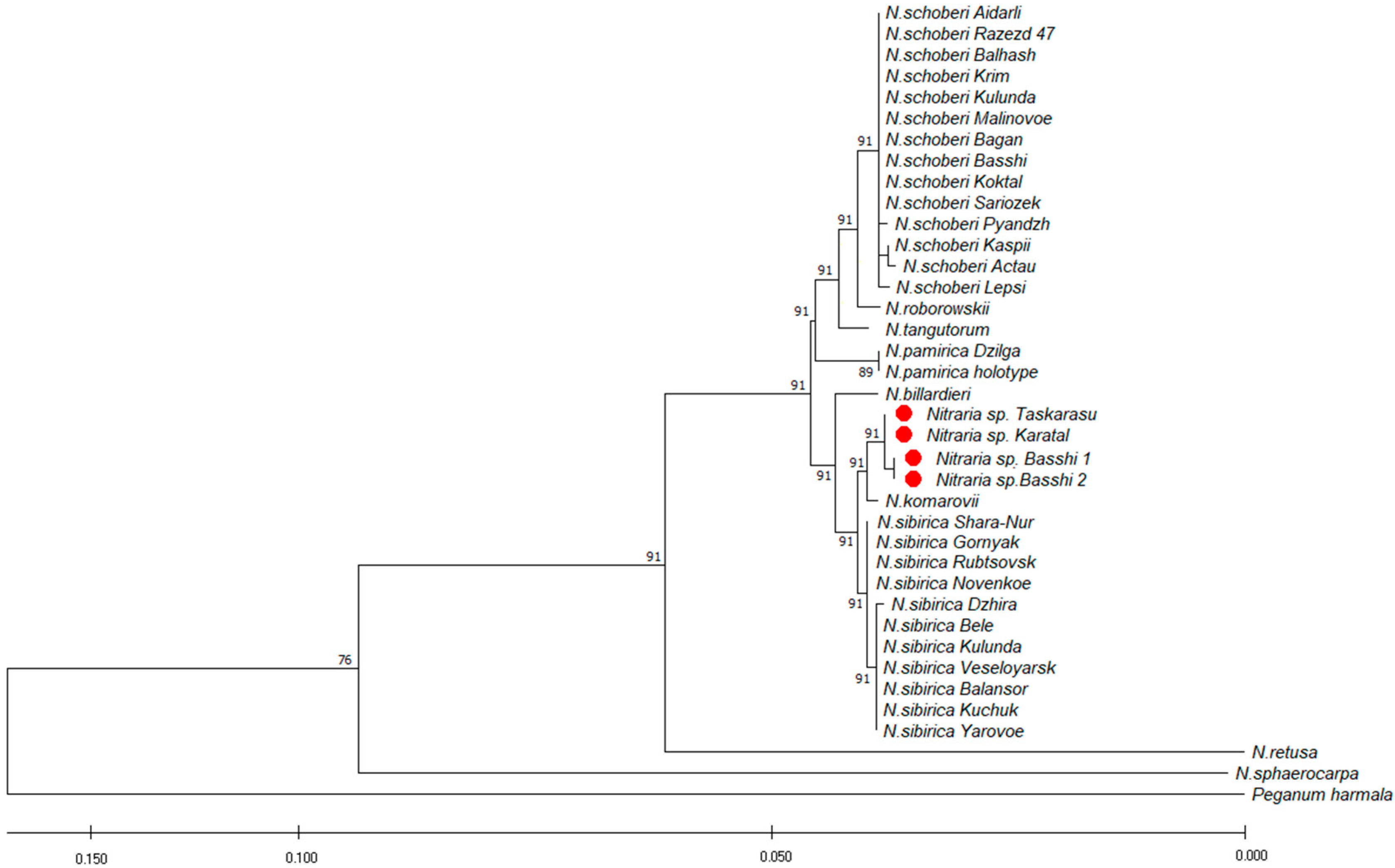
2.3. Molecular Analysis
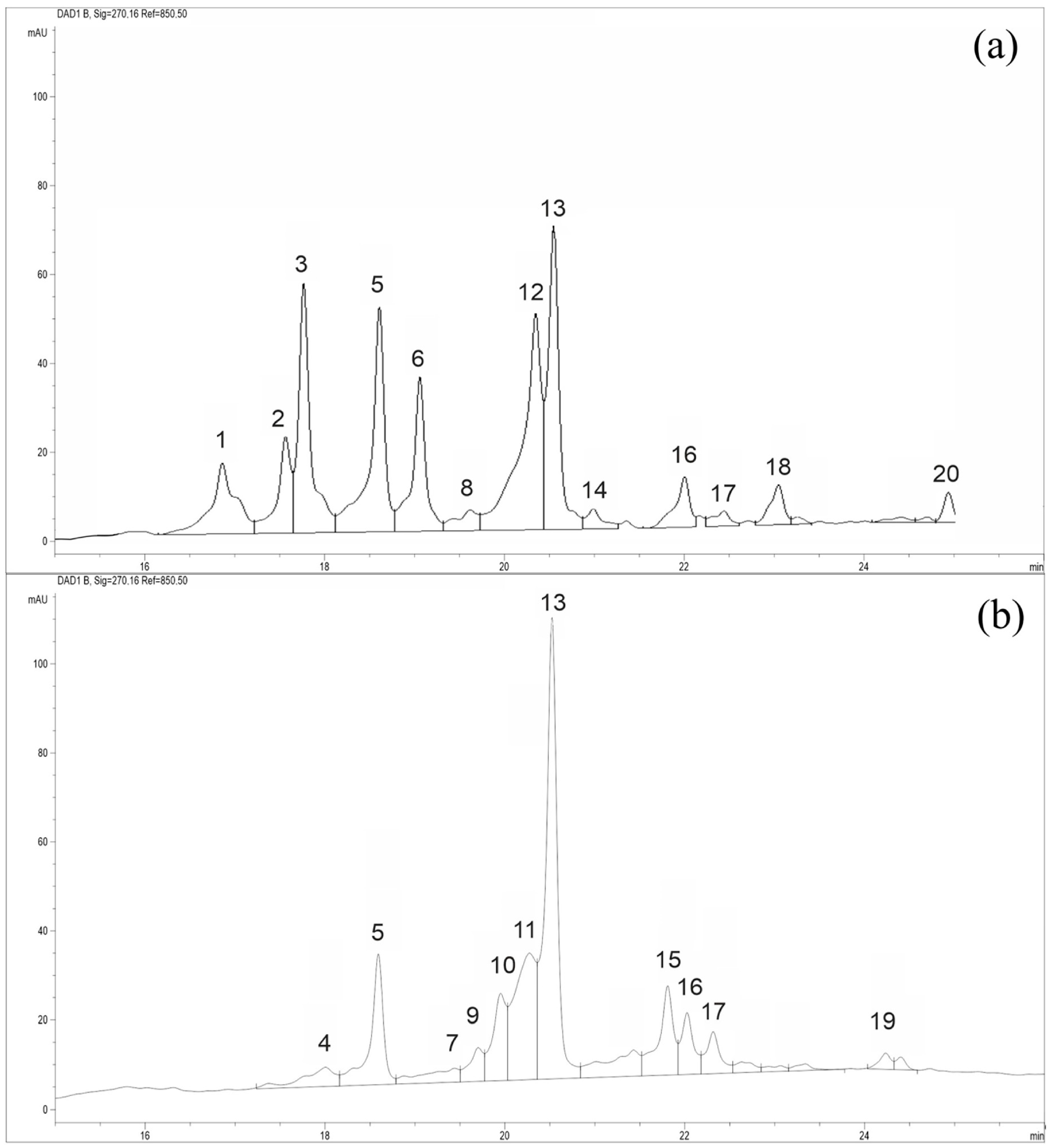
2.4. HPLC-MS Analysis
2.5. Taxonomy
2.5.1. Description
2.5.2. Affinity
2.5.3. Phenology
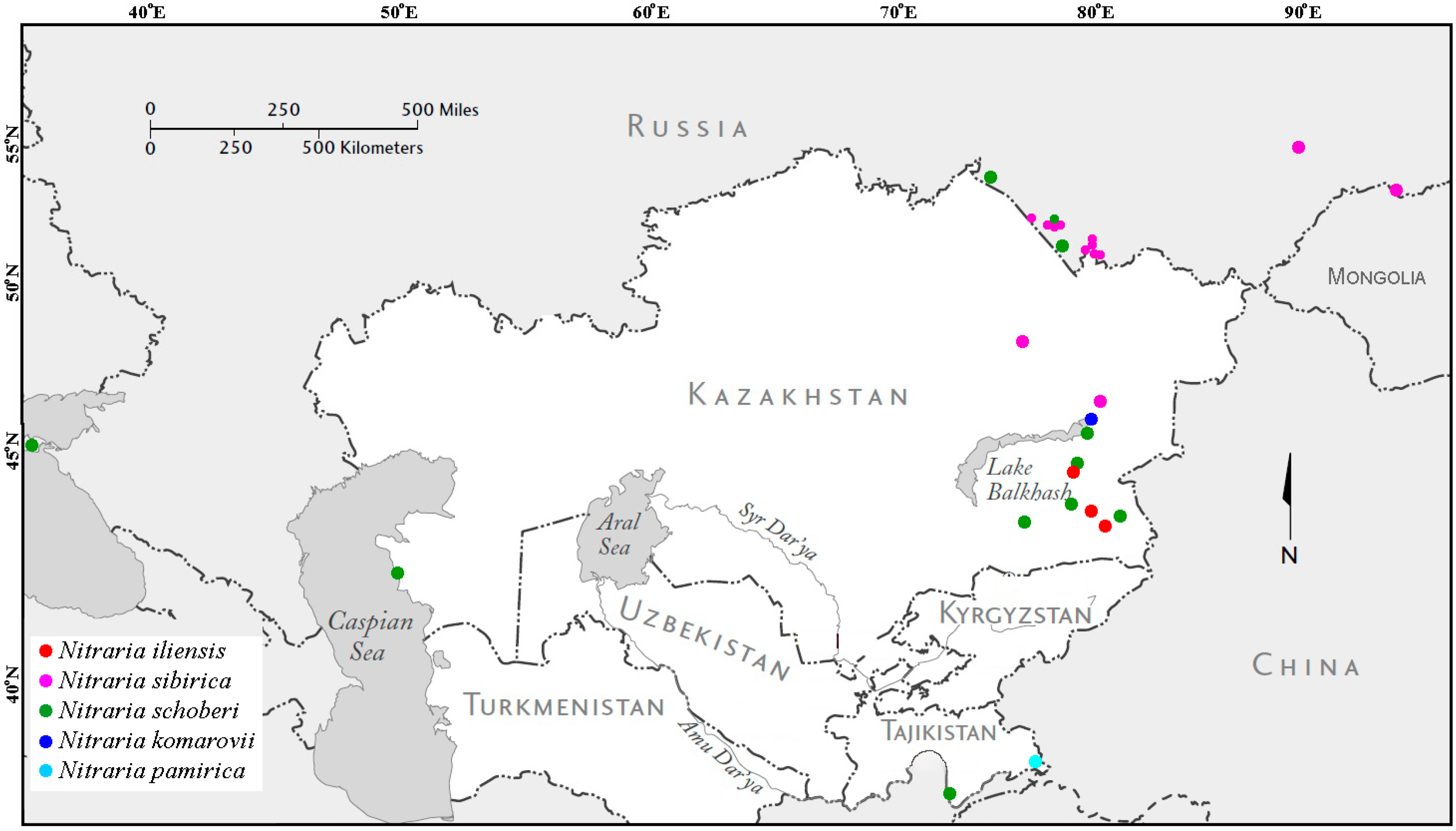
2.5.4. Distribution
2.5.5. Habitat and Ecology
2.5.6. Etymology
2.5.7. Additional Specimens Examined
2.5.8. Notes
2.5.9. Key to Nitraria Species
| 1. Drupe juicy, oval or spherical; stone ovoid or conical.............................................................2 |
| - Drupe dry, swollen, spherical up to 1 cm in diameter, with a membranous swollen pubescent shell; narrow stone, almost fusiform, 0.8 × 0.2–0.3 cm. Thorny bush, prostrate, up to 20–40 cm high, with curved branches, pillow-shaped. Branches are grayish-white in color. Leaves narrow, linear-lanceolate, 20–40 × 1–3 mm. Inflorescence shoot short, up to 5–10 cm. Flowers white, small, up to 0.5 cm....................................................N. sphaerocarpa Maxim |
| 2. Leaves obovate, oblanceolate or linear, entire above, very rarely 1–2-toothed; stipules membranous, deciduous....3 |
| - Leaves spatulate or cuneiform, entire above or 3–5 crenate-toothed; stipules present........N. retusa (Forsk.) Aschers |
| 3. Leaves entire, blunt at the apex, rarely pointed...................................................................4 |
| - Leaves are the largest of all species of the genus, 25–46 × 6–9 mm, either entire or 1–2-toothed at the apex (on vegetative shoots); drupe large, round, juicy, pale pink to maroon in color, stone 10–12 mm long, 3.5–4.5 mm wide, ovoid pointed to apex, barely pointed or almost blunt; bush 1–1.5 m high................................N. roborowskii Kom. |
| 4. Bush above 0.5 m, spreading-branching..........................................................................5 |
| - Bush 0.1–0.3 m high, spreading, dense; leaves green, matte, oblanceolate with a pointed apex; number of flowers in the inflorescence 9–15; petals white, oblong-oval with short claws; drupe cherry, 7–8 × 4–5 mm, stone oblong-conical, 5–6 × 2–2.5 mm.............................................................................N. pamirica L.I. Vassiljeva |
| 5. Drupe maroon to black in color, oval or spherical.................................................................7 |
| - Drupe yellow, orange, pale to bright red, oval, confined to Central Asia............................................6 |
| - Bush grows in Australia, leaves narrow at the apex, slightly pointed, stone conically ovoid.........N. billardierei DC. |
| 6. Stems ascending, leaves greenish-yellow, narrow, long, linear spatulate, 25–28 mm long, 2–3.5 mm wide; petals yellowish-white, ovate with short claws; number of flowers per inflorescence 17–30; drupe oval, 8–12 × 7–11 mm; stone conically ovoid with a recurved apex, 8.5–11 mm long, 4.5–6 mm wide; drupe sap light pink.......................................................................................................N. komarovii Iljin & Lava ex Bobrov |
| 7. Drupe black, oval or spherical; stone smaller than 7 mm..........................................................8 |
| - Drupe oval, maroon to black in color, 7–10 × 6–9 mm; stone ovoid, obtuse, 7–10 mm long, 4.5–6.5 mm wide; bush 0.7–1.5 m high, spreading-branching; stems arcuate, with large spines; leaves dark green, shiny, oblong-spatulate, 20–26 mm long, 3–6 mm wide; flowers yellowish-white in color; petals ovoid or diamond-shaped with short claws; number of flowers per inflorescence 11–28; fruit sap pale reddish................................................N. schoberi L. |
| - Drupe oval, burgundy to dark purple in color, 8 × 6 mm; stone ovoid, obtuse, 6 × 3 mm; leaves oblong-lanceolate, 24–25 x 4.5–6 mm; bush 1–2 m high (up to 3–4)...................................................N. tangutorum Bobrov |
| 8. Leaves pubescent or slightly pubescent...................................................9 |
| - Leaves silvery from dense pubescence, calyx densely hairy, petals and ovaries hairy on the outside; leaf 10–15 × 4–6 mm..........................................................................N. praevisa Bobrov |
| 9. Bush 0.6–1.8 m high, dense, with small spines; leaves oblanceolate, elongated, green, 14–16 × 2–3 mm; number of flowers per inflorescence 40–80 (90); petals white, ovate or rhombic with short claws; drupe black, oval or spherical, 4–6 × 4–4.5 mm; stone narrowly ovoid with a narrow pointed apex; 3.5–5 mm long, 2–2.5 mm wide; fruit sap green-black.....................................................................................N. iliensis Banaev&Tomoshevich |
| - Bush 0.2–0.8 m high, dense, with tightly arranged small spines and oblanceolate bluish-green leaves, 10–13 mm long, 2–3 mm wide; number of flowers per inflorescence 20–48; flowers white, pale-purple in buds; petals white, pointed-elliptical with narrow claws; drupe black, 4–9 × 4–8 mm; stone ovoid-acuminate, 3.6–6 mm long and 2.5–3.5 mm wide; fruit sap dark blue..............................................................................N. sibirica Pall. |
3. Materials and Methods
3.1. Plant Material
3.2. Morphological Analysis
3.3. Palynological Analysis
3.4. Molecular Analysis
3.5. HPLC-MS Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engler, A. Zygophyllaceae, Rutaceae, Simaroubaceae, Burseraceae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, A., Prantl, K., Eds.; Engelmann: Leipzig, Germany, 1931; Volume 19a, pp. 144–184, 187–456. [Google Scholar]
- Bachelier, J.B.; Endress, P.K.; Ronse de Craene, L. Comparative floral structure and development of Nitrariaceae (Sapindales) and systematic implications. In Flowers on the Tree of Life; Wantntorp, L., Ronse de Craene, L., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 181–217. [Google Scholar]
- Woutersen, A.; Jardine, P.E.; Bogotá-Angel, R.G.; Zhang, H.; Silvestro, D.; Antonelli, A.; Gogna, E.; Erkens, R.H.J.; Gosling, W.D.; Dupont-Nivet, G.; et al. A novel approach to study the morphology and chemistry of pollen in a phylogenetic context, applied to the halophytic taxon Nitraria L. (Nitrariaceae). PeerJ 2018, 6, e5055. [Google Scholar] [CrossRef] [Green Version]
- Tomoshevich, M.; Banaev, E.; Khozyaykina, S.; Erst, A. Pollen morphology of some species from genus Nitraria. Plants 2022, 11, 2359. [Google Scholar] [CrossRef] [PubMed]
- Komarov, V.L. Introduction to the floras of China and Mongolia. Tr. S.-Peterb. Bot. Sada 1908, 19, 1–179. (In Russian) [Google Scholar]
- Chafoor, A. Zygophyllaceae. In Flora of West Pakistan; Nasir, E., Ali, S.I., Eds.; Department of Botany, University of Karachi: Karachi, Pakistan, 1974; Volume 76, pp. 1–35. [Google Scholar]
- Pallas, P.S. Journey through Different Provinces of the Russian State. Part Three. Half First; Imperial Academy of Sciences: St. Petersburg, Russia, 1788; 655p. [Google Scholar]
- Trautvetter, R.E. Observations on plants by Dre. G. Radde in 1870 collected in Turcomania and Transcaucasia, and also in some other places. Acta Horti Petropolitani 1871, 1, 13–34. [Google Scholar]
- Il’in, M.M. Nitraria and the origin of desert flora. Priroda 1944, 5–6, 116–118. (In Russian) [Google Scholar]
- Kovtonyuk, N.K.; Tomoshevich, M.A.; Banaev, E.V. Typification of the name Nitraria komarovii (Nitrariaceae). Bot. Pac. 2019, 8, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Petrov, M.P. Systematics and geography of Nitraria L. in Asia. In History of Flora and Vegetation of Eurasia; Nauka: Leningrad, Russia, 1972; pp. 156–181. (In Russian) [Google Scholar]
- Grubov, V.I. Guide for Identification of Vascular Plants of Mongolia; Nauka: Leningrad, Russia, 1982; pp. 175–177. (In Russian) [Google Scholar]
- Su, Z.; Lu, W.; Zhang, M. Phylogeographical patterns of two closely related desert shrubs, Nitraria roborowskii and N. sphaerocarpa (Nitrariaceae), from arid north-western China. Bot. J. Linn. Soc. 2016, 108, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Bobrov, E.G. On the origin of flora of the deserts of the Old World in conjunction with the review of Nitraria genus. Bot. Zh. 1965, 50, 1053–1057. (In Russian) [Google Scholar]
- Bobrov, E.G. About Asian species of the genus Nitraria L. Sov. Bot. 1946, 14, 19–30. (In Russian) [Google Scholar]
- Vasil’eva, L.I. A new species of the genus Nitraria L. from the Pamirs. Nov. Sist. Vyssh. Rast. 1974, 11, 341–344. [Google Scholar]
- Pan, X.-Y.; Wei, X.-P.; Yu, Q.-S.; Chen, J.-K.; Wang, G.-X. Polyploidy: Classification, evolution and applied perspective of the genus Nitraria. Chin. Bull. Bot. 2003, 20, 632–638. [Google Scholar]
- El-Kheir, Z.A.A.; El-Monayeri, M.O.; EL-Raof, D.A. Evaluation of genetic diversity between some Nitraria retusa populations using molecular markers. Al-Azhar Bull. Sci. 2006, 17, 47–66. [Google Scholar] [CrossRef] [Green Version]
- Temirbayeva, K.; Zhang, M.L. Molecular phylogenetic and biogeographical analysis of Nitraria based on nuclear and chloroplast DNA sequences. Plant Syst. Evol. 2015, 30, 1897–1906. [Google Scholar] [CrossRef]
- Zhang, Z.-L.; Song, M.-F.; Guan, Y.-H.; Li, H.-T.; Niu, Y.-F.; Zhang, L.-X.; Ma, X.-J. DNA barcoding in medicinal plants: Testing the potential of a proposed barcoding marker for identification of Uncaria species from China. Biochem. Syst. Ecol. 2015, 60, 8–14. [Google Scholar] [CrossRef]
- Banaev, E.V.; Tomoshevich, M.A.; Yamtyrov, M.B. On variation of metric and qualitative characters of Nitraria L. species in the context of ecological-climatic conditions of habitats in Siberia. Contemp. Probl. Ecol. 2017, 10, 664–673. [Google Scholar] [CrossRef]
- Banaev, E.V.; Khozyaykina, S.A.; Dorogina, O.V.; Tomoshevich, M.A. Genetic diversity of Nitraria sibirica Pall. (Nitrariaceae) in the populations of Siberia and Kazakhstan based on the polymorphism of intermicrosatellite markers. Contemp. Probl. Ecol. 2022, 15, 400–408. [Google Scholar] [CrossRef]
- Fan, W.Y.; Zhou, C.J.; Wang, Y.C. Confirmation of natural hybrids between two species of Nitraria in Inner Mongolia based on molecular evidence. Acta Bot. Boreal-Occident. Sin. 2011, 31, 913–919. [Google Scholar]
- Baitenov, M.S. Flora of Kazakhstan, 1st ed.; Gylym: Almaty, Kazakhstan, 1999; 400p. [Google Scholar]
- Baitenov, M.S. Flora of Kazakhstan, 2nd ed.; Gylym: Almaty, Kazakhstan, 2001; 280p. [Google Scholar]
- Ovchinnikov, P.N. Flora of the Tajik SSR, 6th ed.; Nauka: Leningrad, Russia, 1981; 725p. [Google Scholar]
- Krasnoborov, I.M.; Lomonosova, M.N.; Shaulo, D.N.; Kutsev, M.G.; Krasnikov, A.A.; Shmakov, A.I.; German, D.A.; Smirnov, S.V.; Doronkin, V.M.; Durnikin, D.A.; et al. Key to Plants of the Altai Territory; Publishing House of the Siberian Branch of the Russian Academy of Sciences, Branch “Geo”: Novosibirsk, Russia, 2003; 492p. [Google Scholar]
- Krasnoborov, I.M.; Lomonosova, M.N.; Shaulo, D.N.; Vibe, E.I.; Zhirova, O.S.; Korolyuk, E.A.; Krasnikov, A.A.; Snytko, O.N.; Tupitsyna, N.N. Key to Plants of the Novosibirsk Region; Nauka SO RAN: Novosibirsk, Russia, 2000; 492p. [Google Scholar]
- Shaulo, D.N.; Krasnoborov, I.M.; Lomonosova, M.N.; Krasnikov, A.A.; Artemov, I.A.; Baikov, K.S. Key to Plants of the Tyva Republic; Publishing House of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2007; 706p. [Google Scholar]
- Pavlov, N.V. Flora of Kazakhstan, 6th ed.; Publishing House of the Academy of Sciences of the Kazakh SSR: Alma-Ata, Kazakhstan, 1963; pp. 52–53. [Google Scholar]
- Tomoshevich, M.A.; Banaev, E.V.; Ak-Lama, T.A. Nitraria komarovii Iljin & Lava ex Bobrov (Nitrariaceae), a new record for the flora of Kazakhstan. Check List 2019, 15, 891–897. [Google Scholar] [CrossRef]
- Polyakova, T.A.; Banaev, E.V.; Tomoshevich, M.A. Identification of species in the genus Nitraria L. (Nitrariaceae) based on nucleotide variability of nuclear ribosomal DNA. Vavilovskii Zh. Genet. Sel. 2020, 24, 481–488. [Google Scholar] [CrossRef]
- Banaev, E.V.; Poliakova, T.A.; Tomoshevich, M.A.; Ak-Lama, T.A. Using the ITS2 transcribed spacer of nuclear ribosomal DNA to identify species of the genus Nitraria. In Proceedings of the International Conferences Plant Diversity: Status, Trends, Conservation Concept, Novosibirsk, Russia, 30 September–3 October 2020; Volume 24, p. 00009. [Google Scholar] [CrossRef]
- Voronkova, M.S.; Banaev, E.V.; Tomoshevich, M.A.; Ak-Lama, T.A. Possibilities of using the HPLC method in the taxonomy of the genus Nitraria (Nitrariaceae). In Proceedings of the International Conferences Plant Diversity: Status, Trends, Conservation Concept, Novosibirsk, Russia, 30 September–3 October 2020; Volume 24, p. 00096. [Google Scholar] [CrossRef]
- Banaev, E.V.; Tomoshevich, M.A.; Ak-Lama, T.A. Morphological variability of Nitraria species in Central and Southern Kazakhstan. Contemp. Probl. Ecol. 2021, 14, 255–268. [Google Scholar] [CrossRef]
- Peshkova, G.A. Family Nitrariaceae. In Flora of Siberia; Nauka: Novosibirsk, Russia, 1996; Volume 10, pp. 34–35. (In Russian) [Google Scholar]
- Koropachinskii, I.Y. Tree Flora of Siberia; Botanica Pacifica: Novosibirsk, Russia, 2016. (In Russian) [Google Scholar]
- Zhang, M.L.; Temirbayeva, K.; Sanderson, S.; Chen, X. Young dispersal of xerophil Nitraria lineages in intercontinental disjunctions of the Old World. Sci. Rep. 2015, 5, 13840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneyer, V.S.; Rodionov, A.V. Plant DNA barcodes. Uspekhi Sovrem. Biol. Adv. Curr. Biol. 2018, 138, 531–538. (In Russian) [Google Scholar] [CrossRef]
- Lu, L.; Li, X.; Hao, Z.; Yang, L.; Zhang, J.; Peng, Y.; Xu, H.; Lu, Y.; Zhang, J.; Shi, J.; et al. Phylogenetic studies and comparative chloroplast genome analyses elucidate the basal position of halophyte Nitraria sibirica (Nitrariaceae) in the Sapindales. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2018, 29, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Hu, N.; Suo, Y.; Chi, X.; Wang, H. The complete chloroplast genome sequences of two species from Nitraria. Mitochondrial DNA Part B 2019, 4, 1229–1230. [Google Scholar] [CrossRef] [Green Version]
- Paica, I.C.; Banciu, C.; Maria, G.M.; Vladimirescu, M.; Manole, A. Genetic diversity in marginal populations of Nitraria schoberi L. from Romania. Diversity 2022, 14, 882. [Google Scholar] [CrossRef]
- Du, Q.; Xin, H.; Peng, C. Pharmacology and phytochemistry of the Nitraria genus (Review). Mol. Med. Rep. 2015, 11, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Saleh, N.A.M.; El-Hadidi, M.H. An approach to the chemosystematics of the Zygophyllaceae. Biochem. Syst. Ecol. 1977, 5, 121–128. [Google Scholar] [CrossRef]
- Shen, M.Y.; Zuanazzi, J.A.; Kan, C.; Quirion, J.C.; Husson, H.P.; Bick, I.R.C. Alkaloids from Nitraria billardieri. Nat. Prod. Lett. 1995, 6, 119–125. [Google Scholar] [CrossRef]
- Sameh, R.H.; Salwa, A.K.; Mohamed, E.T.; Nabiel, A.M.; Saleh, N. Chemosystematic studies of Nitraria retusa and selected taxa of Zygophyllaceae in Egypt. Plant Syst. Evol. 2009, 277, 251–264. [Google Scholar] [CrossRef]
- Tulyaganov, T.S.; Allaberdiev, F.K. Alkaloids from plants of the Nitraria genus. Structure of sibiridine. Chem. Nat. Compd. 2003, 39, 292–293. [Google Scholar] [CrossRef]
- Tulyaganov, T.S.; Nazarov, O.M.; Levkovich, M.G.; Abdullaev, N.D. Alkaloids of the Nitraria genus. Komavine and acetylkomavine. Chem. Nat. Compd. 2001, 37, 61–64. [Google Scholar] [CrossRef]
- Turghun, C.; Bakri, M.; Zou, G.A.; Bobakulov, K.M.; Aisa, H.A. Phenolic compounds from leaves of Nitraria sibirica. Chem. Nat. Compd. 2018, 54, 987–989. [Google Scholar] [CrossRef]
- Duan, J.A.; Zhou, R.H.; Zhao, S.X. The chemical constituents of flavonoids and phenolic acid compounds of leaves from Nitraria tangutorum Bor. J. Plant Resour. Environ. 1999, 8, 6–9. [Google Scholar]
- Barbhan, N.; Miladi, I.; Ali, S.I.; Poupon, E.; Mohamed, A.A.; Beniddir, M.A. Chemical constituents of Nitraria retusa grown in Egypt. Chem. Nat. Compd. 2017, 53, 994–996. [Google Scholar] [CrossRef]
- Banaev, E.V.; Voronkova, M.S.; Vysochina, G.I.; Tomoshevich, M.A. Population structure and differentiation of the Siberian representatives of the genus Nitraria L. (Nitrariaceae) based on the composition and content of phenolic compounds in leaves. Contemp. Probl. Ecol. 2015, 8, 735–742. [Google Scholar] [CrossRef]
- Voronkova, M.S.; Banaev, E.V.; Tomoshevich, M.A. Comparative study of the composition and content of phenolic compounds of leaves of plants of the genus Nitraria (Nitrariaceae). Khim. Rast. Syr’ya 2017, 4, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Akhani, H. Notes on the flora of Iran: 1. Asparagus (Asparagaceae) and Nitraria (Zygophyllaceae). Edinb. J. Bot. 2002, 59, 295–302. [Google Scholar] [CrossRef]
- El-Hadidi, M.N. Zygophyllaceae. In Flora Iranica; Rechinger, K.H., Ed.; Akademische Druck-u.-Verlagsanstalt: Graz, Austria, 1972; Volume 98, pp. 1–32. [Google Scholar]
- Erdtman, G. Pollen and Plant Taxonomy Angiosperms; Hafner Publ. Co.: New York, NY, USA, 1986; 553p. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Hessen, M.; Halbritter, H.; Zetter, R.; Weber, M.; Buchner, R.; Frosch-Radivo, A.; Ulrich, S. Pollen Terminology an Illustrated Handbook; SpringerWein: New York, NY, USA, 2009; 266p. [Google Scholar]
- Halbritter, H.; Ulrich, S.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Frosch-Radivo, A. Illustrated Pollen Terminology; Springer International Publishing AG part of Springer Nature: Cham, Switzerland, 2018; p. 483. [Google Scholar] [CrossRef] [Green Version]
- Erdtman, G. Pollen Morphology and Plant Taxonomy–Angiosperms: An Introduction to Palynology; Almqvist & Wiksell: Stockholm, Sweden; Chronica Botanica: Waltham, MA, USA, 1952; Volume 1, 539p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 12–15. [Google Scholar]
- Potter, D.; Still, S.M.; Grebenc, T.; Ballian, D.; Božič, G.; Franjiæ, J.; Kraigher, H. Phylogenetic relationships in tribe Spiraea (Rosaceae) inferred from nucleotide sequence data. Pl. Syst. Evol. 2007, 266, 105–118. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Character | Character | N. schoberi | N. sibirica | N. komarovii | N. pamirica | N. iliensis |
|---|---|---|---|---|---|---|
| 1 | Height of bush (m) | 0.7–1.5 | 0.2–0.8 | 0.5–1 | 0.1–0.3 | 0.6–1.8 |
| 2 | Habit | Spreading-branching | Spreading-branching, dense | Branching, graceful | Spreading, dense | Spreading-branching, dense |
| 3 | Leaf length (mm) | 20–26 | 10–13 | 25–28 | 13–17 | 14–16 |
| 4 | Leaf width (mm) | 3–6 | 2–4 | 2–3,5 | 2–3 | 2–3 |
| 5 | Distance from the base to the widest point of the leaf blade (mm) | 15–16 | 8–9 | 22 | 10 | 10–12 |
| 6 | Leaf color | Dark-green, shiny | Glaucescent-green | Pale green | Green, matte | Green |
| 7 | Leaf shape | Oblanceolate | Oblanceolate | Narrow, linear-spathulate | Oblanceolate, with a pointed apex | Oblanceolate |
| 8 | Flowers | Yellowish-white | White (buds pale-violet) | Yellowish-white | White | White |
| 9 | Number of flowers per inflorescence | 11–28 | 20–48 | 17–30 | 9–15 | 40–80 |
| 10 | Flower spacing (mm) | 0.4–0.8 | 0.2–0.3 | 0.3–0.7 | 0.2–0.3 | 0.3–0.5 |
| 11 | Petal shape | Ovate or rhombic; claws short | Acuminate-elliptica; claws narrow | Ovate; claws short | Ovate; claws short | Ovate or rhombic; claws short |
| 12 | Petal length (mm) | 3–4.7 | 2.6–4 | 1.7–3.9 | 2.1–2.3 | 3.5 |
| 13 | Petal width (mm) | 2–3.6 | 1–2.8 | 1–2.9 | 0.7–0.9 | 2.6 |
| 14 | Anther length (mm) | 1.2–1.4 | 0.6–0.9 | 1.3 | - | 0.8 |
| 15 | Anther width (mm) | 0.5–0.7 | 0.3–0.5 | 0.7 | - | 0.5–0.6 |
| 16 | Pestle length (mm) | 2.02–4.5 | 1.35–3.19 | 2.01–3.59 | - | 2.05 |
| 17 | Pestle width (mm) | 1.38–2.23 | 0.97–1.41 | 1.09–1.82 | - | 1.22 |
| 18 | Fruit color | Dark-red to black | Black | Yellow, orange, or pale to bright red | Cherry red | Black |
| 19 | Fruit length (mm) | 7–10 | 4–9 | 8–12 | 7–8 | 4–6 |
| 20 | Fruit width (mm) | 6–11 | 4–8 | 7–11 | 4–5 | 4–4.5 |
| 21 | Weight of 100 fruits (g) | 270–600 | 100–270 | 380–490 | 80–90 | 80–130 |
| 22 | Fruit shape | Oval | Globate or oval | Oval | Oval | Globate or oval |
| 23 | Fruit sap | Pale reddish | Dark-blue | Pale pink | Pale pink | Black–green |
| 24 | Stone length (mm) | 7–10 | 3,6–6 | 8,6–11 | 5–6 | 3.5–5 |
| 25 | Stone width (mm) | 4.5–6.5 | 2.5–3.5 | 4.7–6 | 2.0–2.5 | 2–2.6 |
| 26 | Stone shape | Ovate, obtuse | Ovate, pointed | Conic-ovate with a pointed apex | Oblong-conical, | Narrow ovoid with a narrow pointed apex |
| No. | Taxon | Specimen | Locality | Date | Herbarium, Specimen Number |
|---|---|---|---|---|---|
| 1 | N. sibirica | Noven’koe | Russia, Altai Krai, Loktevsky District, vicinity of Noven’koe village | 31 May 2011 | NSK3001284 |
| 2 | N. sibirica | Veseloyarsk | Russia, Altai Krai, Rubtsovsky District, vicinity of Veseloyarsk village | 31 May 2011 | NSK3001286 |
| 3 | N. sibirica | Kulunda | Russia, Altai Krai, Slavgorodskiy District, on the shore of Lake Kulundinskoe | 2 June 2011 | NSK3001276 |
| 4 | N. sibirica | Bele | Russia, Republic of Tyva, Tandinsky kozhuun, northern shore of Lake Bele | 22 July 2011 | NSK3001016 |
| 5 | N. sibirica | Balansor | Russia, Altai Krai, Uglovskiy District, on the shore of Lake Balansor | 1 June 2011 | NSK3001280 |
| 6 | N. sibirica | Dzhira | Russia, Altai Krai, Kulundinsky District, eastern shore of Lake Dzhira | 2 June 2011 | NSK3001283 |
| 7 | N. sibirica | Gornyak | Russia, Altai Krai, Loktevsky District, vicinity of Gornyak village | 31 May 2011 | NSK3001289 |
| 8 | N. sibirica | Kuchuk | Russia, Altai Krai, Blagoveshchensky District, vicinity of Nizhny Kuchuk village | 2 June 2011 | NSK3001274 |
| 9 | N. sibirica | Yarovoe | Russia, Altai Krai, Tabunsky District, southern shore of Bolshoye Yarovoe Lake | 3 June 2011 | NSK3001282 |
| 10 | N. sibirica | Shara-Nur | Russia, Tyva Republic, Tes-Khemsky kozhuun, the shore of Lake Shara-Nur | 28 July 2011 | NSK3000991 |
| 11 | N. sibirica | Rubtsovsk | Russia, Altai Krai, vicinity of Rubtsovsk city | 15 June 2013 | NSK3001785 |
| 12 | N. schoberi | Balhash | Republic of Kazakhstan, Almaty Region, on the shore of Lake Balkhash, sandy desert | 31 May 2016 | NSK3000948 |
| 13 | N. schoberi | Sariozek | Republic of Kazakhstan, Almaty Region, 30 km north of Sariozek village | 25 May 2016 | NSK3000947 |
| 14 | N. schoberi | Basshi | Republic of Kazakhstan, Almaty Region, vicinity of Basshi village | 25 May 2016 | NSK3000982 |
| 15 | N. schoberi | Aidarli | Republic of Kazakhstan, Almaty Region, Zhambylskii District, 17 km south of Aidarli village | 21 August 2017 | NSK3000958 |
| 16 | N. schoberi | Koktal | Republic of Kazakhstan, Almaty Region, vicinity of Koktal village | 30 July 2013 | NSK3000999 |
| 17 | N. schoberi | Lepsi | Republic of Kazakhstan, Almaty region, on the bank of the Lepsi River in outskirts of Lepsi village | 30 May 2016 | NSK3000981 |
| 18 | N. schoberi | Raz’ezd_47 | Republic of Kazakhstan, Almaty region, vicinity of the village of Molaly (railroad junction No. 47) | 15 August 2017 | NSK3000944 |
| 19 | N. schoberi | Bagan | Russia, Novosibirskaya Oblast’, southwest of the village Grushevka, on the terrace of Lake Bol’shoy Bagan | 4 June 2011 | NSK3000973 |
| 20 | N. schoberi | Kaspii | Republic of Kazakhstan, Mangistauskaya Oblast’, vicinity of Aktau city, on sandy mound | 12 June 2012 | NSK3000979 |
| 21 | N. schoberi | Actau | Republic of Kazakhstan, Mangistauskaya Oblast’, vicinity of Aktau city | 12 June 2012 | NSK3000978 |
| 22 | N. schoberi | Krim | Crimea, on the sandy coast of the Black Sea in Dvuyakornaya Bay | 16 September 2013 | NSK3000960 |
| 23 | N. schoberi | Pyandzh | Republic of Tajikistan, Gorno-Badakhshan Autonomous Region, on the sandy bank of the Pyandzh River | 8 August 2014 | NSK3000994 |
| 24 | N. schoberi | Kulunda | Russia, Altai Krai, Slavgorodskiy District, on the shore of Lake Kulundinskoe | 2 June 2011 | NSK3000975 |
| 25 | N. schoberi | Malinovoe | Russia, Altai Krai, Mikhailovskiy District, on the shore of Lake Malinovoe | 1 June 2011 | NSK3000971 |
| 26 | N. komarovii | Balhash | Republic of Kazakhstan, Almaty Region, on the shore of Lake Balhash, sandy desert | 31 May 2016 | NSK3000920 |
| 27 | N. iliensis | Basshi 1 | Republic of Kazakhstan, Almaty Region, vicinity of Basshi village | 25 May 2016 | NSK3001499 |
| 28 | N. iliensis | Basshi 2 | Republic of Kazakhstan, Almaty Region, vicinity of Basshi village | 30 July 2013 | NSK3001277 |
| 29 | N. iliensis | Taskarasu | Republic of Kazakhstan, Almaty Region, vicinity of Taskarasu village | 26 May 2016 | NSK3001244 |
| 30 | N. iliensis | Karatal | Republic of Kazakhstan, Almaty Region, Karatalskii District, vicinity of Ushtobe city, on the terrace of the Karatal River | 29 May 2016 | NSK3000922 |
| 31 | N. pamirica | Shaimak | Republic of Tajikistan, Gorno-Badakhshan Autonomous Region, Eastern Pamir, on the cliff of the Djilga River | 10 August 2014 | NSK3001238 |
| Taxa | Polar Axis (P) | Equatorial Axis (E) | P/E | Shape | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Range | X ± Sx | CV, % | Range | X ± Sx | CV, % | Range | X ± Sx | CV, % | ||
| N. sibirica | 36.11–43.33 | 40.24 ± 0.34 | 4.3 | 17.07–23.50 | 20.48 ± 0.30 | 7.5 | 1.73–2.28 | 1.97 ± 0.02 | 5.8 | prolate |
| N. komarovii | 23.38–29.50 | 27.15 ± 0.30 | 5.6 | 14.23–18.91 | 16.42 ± 0.30 | 9.1 | 1.44–1.89 | 1.66 ± 0.02 | 8.0 | prolate |
| N. schoberi | 38.19–49.90 | 42.84 ± 0.62 | 7.2 | 22.28–32.59 | 27.84 ± 0.65 | 11.0 | 1.24–2.05 | 1.57 ± 0.05 | 16.5 | prolate |
| N. pamirica | 30.13–36.07 | 32.29 ± 0.32 | 4.9 | 24.87–30.62 | 26.50 ± 0.28 | 5.3 | 1.18–1.27 | 1.22 ± 0.005 | 2.0 | subprolate |
| N. iliensis | 31.41–36.00 | 33.82 ± 0.32 | 4.8 | 21.10–27.08 | 25.81 ± 0.50 | 9.7 | 1.10–1.51 | 1.32 ± 0.02 | 6.7 | subprolate or prolate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banaev, E.V.; Tomoshevich, M.A.; Khozyaykina, S.A.; Erst, A.A.; Erst, A.S. Integrative Taxonomy of Nitraria (Nitrariaceae), Description of the New Enigmatic Species and Key to All Currently Known Species. Plants 2023, 12, 593. https://doi.org/10.3390/plants12030593
Banaev EV, Tomoshevich MA, Khozyaykina SA, Erst AA, Erst AS. Integrative Taxonomy of Nitraria (Nitrariaceae), Description of the New Enigmatic Species and Key to All Currently Known Species. Plants. 2023; 12(3):593. https://doi.org/10.3390/plants12030593
Chicago/Turabian StyleBanaev, Evgeny V., Maria A. Tomoshevich, Sofia A. Khozyaykina, Anna A. Erst, and Andrey S. Erst. 2023. "Integrative Taxonomy of Nitraria (Nitrariaceae), Description of the New Enigmatic Species and Key to All Currently Known Species" Plants 12, no. 3: 593. https://doi.org/10.3390/plants12030593